sperm competition and reproductive mode influence sperm dimensions and structure among snakes
TRANSCRIPT

ORIGINAL ARTICLE
doi:10.1111/j.1558-5646.2009.00739.x
SPERM COMPETITION AND REPRODUCTIVEMODE INFLUENCE SPERM DIMENSIONSAND STRUCTURE AMONG SNAKESMaximiliano Tourmente,1,2,3,4 Montserrat Gomendio,5 Eduardo R. S. Roldan,5,6 Laura C. Giojalas,2,3
and Margarita Chiaraviglio1
1Laboratorio de Biologıa del Comportamiento, Facultad de Ciencias Exactas, Fısicas y Naturales, Universidad Nacional de
Cordoba, Cordoba, Argentina2Centro de Biologıa Celular y Molecular, Facultad de Ciencias Exactas, Fısicas y Naturales, Universidad Nacional de
Cordoba, Cordoba, Argentina3Consejo Nacional de Investigaciones Cientıficas y Tecnologicas (CONICET), Argentina
4E-mail: [email protected] Ecology and Biology Group, Museo Nacional de Ciencias Naturales (CSIC), Madrid, Spain6Department of Veterinary Basic Sciences, The Royal Veterinary College, University of London, London, United Kingdom
Received July 11, 2008
Accepted May 4, 2009
The role of sperm competition in increasing sperm length is a controversial issue, because findings from different taxa seem
contradictory. We present a comparative study of 25 species of snakes with different levels of sperm competition to test whether
it influences the size and structure of different sperm components. We show that, as levels of sperm competition increase, so does
sperm length, and that this elongation is largely explained by increases in midpiece length. In snakes, the midpiece is comparatively
large and it contains structures, which in other taxa are present in the rest of the flagellum, suggesting that it may integrate some
of its functions. Thus, increases in sperm midpiece size would result in more energy as well as greater propulsion force. Sperm
competition also increases the area occupied by the fibrous sheath and outer dense fibers within the sperm midpiece, revealing
for the first time an effect upon structural elements within the sperm. Finally, differences in male–male encounter rates between
oviparous and viviparous species seem to lead to differences in levels of sperm competition. We conclude that the influence of
sperm competition upon different sperm components varies between taxa, because their structure and function is different.
KEY WORDS: Mating systems, reproductive strategies, selection-sexual, snakes, sperm competition, sperm structure.
An outstanding diversity of spermatozoa has evolved in differ-
ent species, particularly among internal fertilizers (Roldan et al.
1992; Anderson and Dixson 2002; Snook 2005; Gomendio and
Roldan 2008). The selective pressures, which may have favored
such variation, are likely to be related to differences in female
reproductive physiology and to levels of sperm competition be-
tween species. The sequence of events leading to fertilization
constitute such a complex process that much of sperm form and
function is influenced by the requirements of finding, interacting
with and fertilizing eggs, which vary between species (Gomendio
and Roldan 1993; Clark et al. 1999; Byrne et al. 2003; Pitnick
et al. 2003). In addition, the morphology of the female reproduc-
tive organs may influence sperm dimensions (Briskie et al. 1997;
Presgraves et al. 1999; Miller and Pitnick 2002; Minder et al.
2005; Anderson et al. 2006; Holman et al. 2008).
On the other hand, when sperm from rival males compete
to fertilize the ova (i.e., sperm competition) additional selec-
tive pressures influence sperm traits. In species in which sperm
competition is prevalent, males evolve larger testes to produce
more sperm. Thus, an increase in relative testes size has been
1C© 2009 The Author(s).Evolution

MAXIMILIANO TOURMENTE ET AL.
associated to high sperm competition levels in many taxa (mam-
mals: Gomendio et al. 1998, birds: Briskie and Montgomerie
1992, amphibians: Jennions and Passmore 1993; Byrne et al.
2002, fish: Stockley et al. 1997, butterflies: Cook and Gage 1995).
Additionally, males from species that experience intense sperm
competition have more viable sperm (Hunter and Birkhead 2002),
a greater proportion of sperm that are competent to fertilize and
sperm that are more responsive to the signals emitted by the ovum
(Gomendio et al. 2006). The competitiveness of an ejaculate is
also largely determined by sperm motility (birds: Birkhead et al.
1999) and sperm swimming velocity (fish: Gage et al. 2004). The
competitive advantage gained by faster swimming sperm is re-
lated either to their ability to outcompete other sperm in the race
to reach the ova (Gage et al. 2004) or to their ability to remain in
the sperm storage sites (Froman et al. 2002).
Theoretical models have predicted a trade-off between sperm
numbers and sperm size (Parker 1998), so the increase in sperm
numbers under sperm competition was believed to be achieved at
the expense of a reduction in sperm size. However, the finding
that polyandrous mammals have longer sperm than monandrous
species led to the suggestion that sperm competition selects for
an elongation of the flagellum presumably because it enhances
sperm swimming speed (Gomendio and Roldan 1991). Prelimi-
nary evidence linking sperm size and sperm swimming speed in a
few species of mammals (Gomendio and Roldan 1991), has now
received further support by a recent analysis on a larger set of
data that shows a strong relationship between the two after con-
trolling for phylogenetic effects (Gomendio and Roldan 2008).
Additionally, recent studies in birds (Lupold et al. 2009), and
fish (Fitzpatrick et al. 2009) have found a strong positive rela-
tionship between sperm velocity and sperm length, with sperm
length being associated to levels of sperm competition. Many
studies exploring this hypothesis have looked for an association
between sperm competition levels and sperm size on other taxa
but the results are not entirely consistent between studies. Most
studies have found an association between sperm size and sperm
competition levels (birds: Briskie and Montgomerie 1992; Briskie
et al. 1997; Johnson and Briskie 1999, passerines: Lupold et al.
2008, finches: Immler and Birkhead 2007, amphibians: Jennions
and Passmore 1993; Byrne et al. 2003, fish: Balshine et al. 2001;
Fitzpatrick et al. 2009, lepidopterans: Gage 1994; Cook and Gage
1995; Morrow and Gage 2000), but others have found no re-
lationship (bats: Hosken 1997, pheasants: Immler et al. 2007),
or an inverse relationship (warblers: Immler and Birkhead 2007,
fish: Stockley et al. 1997, Drosophila: Holman et al. 2008). In
groups with ameboid spermatozoa, sperm competition does favor
larger spermatozoa (mites: Radwan 1996, nematodes: LaMunyon
and Ward 1999), which crawl faster and displace smaller sperma-
tozoa from the spermathecae (LaMunyon and Ward 1998). The
causal link between sperm competition and sperm size has been
shown by experimental studies in which levels of sperm compe-
tition have been manipulated leading to changes in sperm size
(LaMunyon and Ward 2002).
It has also been suggested that in mammals sperm competi-
tion should favor an increase in the size of the midpiece to increase
the amount of energy that fuels motility (Anderson and Dixson
2002; Anderson et al. 2005). Few comparative studies have tested
this possibility, but evidence from studies at the intraspecific level
show that, in mammals, sperm that swim faster have elongated
heads and longer relative flagellum size, but shorter midpieces
(Malo et al. 2006). Conversely, in birds, sperm competition pro-
motes an elongation of the midpiece, which in turn shows a posi-
tive association with sperm velocity (Lupold et al. 2009).
One possible explanation for the contradictory results found
in studies examining the influence of sperm competition on sperm
dimensions, is that the different sperm components may differ in
structure and function between taxa. Thus, a study on animal
groups in which sperm morphology and structure departs from
the better-known groups is needed.
Snakes show more prolonged copulations and higher gonado-
somatic indexes in males than other close taxonomic groups (i.e.,
lizards) (Olsson and Madsen 1998), and females possess sperm
storage structures (Sever and Hamlett 2002). There is also consid-
erable variation between species in reproductive traits (see Zug
et al. 2001 for a partial analysis), ranging from viviparous species
in which females rely on “capital” energy (stored) for reproduc-
tion and reproduce bi- or even tri-annually, to oviparous species in
which females rely on “income” energy (obtained through current
feeding) and reproduce annually (Shine and Madsen 1997; Brown
and Shine 2002). Moreover, snakes’ mating systems exhibit a high
degree of variability, which depends on the temporal availability
and spatial predictability of the females, which influences sperm
competition levels ranging from mating aggregations in which
male/male competition is intense, to prolonged mate search sys-
tems with very low female/male and male/male encounter rates
(Duvall et al. 1992; Shine 2003). Here, we present a comparative
study of 25 species of snakes to test the hypothesis that sperm
competition influences sperm dimensions and structure, and that
reproductive mode (i.e., viviparity vs. oviparity) influences rela-
tive testes size and sperm dimensions.
Materials and MethodsSPECIMENS AND SPERM COLLECTION
Males of 25 species of snakes were obtained by two different
methods: animals captured in the wild and preserved specimens
from museum and private collections. Total body length (cm),
tail length (cm), snout–vent length (SVL) (cm), and body mass
(g) was recorded for each male. The sample size per species
varied depending on the availability of data (mean = 6.52 ± 0.67
2 EVOLUTION 2009

SPERM COMPETITION AND SPERM STRUCTURE AMONG SNAKES
individuals, range: 4–17); in 24 of the 25 species, at least five
males were included in the study sample.
Live (captured) animals were sacrificed by decapitation fol-
lowing anesthetic overdosage with ketamine hydrochloride and
xylazine. Testes were removed and its mass (g) was measured
with a precision scale (Ohaus TA302). Because Boa constrictor
occidentalis is considered a threatened species (Scrocchi et al.
2000) and has been included in Appendix I of CITES (CITES
2008), no males from this species were sacrificed. Instead, the
length and width of the testes were measured by ultrasound scan-
ning (Sonosite 180 plus portable ecographer, linear 5–10 Mhz
transducer, Sonosite Inc., Bothell, WA) and testes volume was cal-
culated using the equation for the volume of an ellipsoid (Mendez
and Villagran 1998). Then testes volume was converted to testes
mass using a linear regression calculated from the data for the
other species included in this study (testes mass = 0.0007 ×testes volume − 0.0293; P < 0.0001, R2 = 0.97). To obtain this
regression line, testicular width and length (mm) of all species
sampled (with the exception of B.c. occidentalis) were recorded
to the nearest 0.1 mm with calipers after dissection, and testicular
volume (mm3) was calculated.
Spermatozoa were obtained from the terminal portion of the
vas deferens for both living and preserved specimens following
dissection. In the case of B. c. occidentalis, semen extraction was
achieved by surgery: the specimens were anesthetized with intra-
muscular injections of ketamine hydrochloride (40 mg/kg) and
xylazine (0.6 mg/kg) (Carpenter et al. 2001; Mader 2005). Sub-
sequently, males were immobilized with adherent hypoallergenic
tape and a 5 cm incision was performed at approximately 10 cm
from the cloacae. The semen samples were extracted from the def-
erent tubes. To close the wound, a double suture was performed
at muscular and dermal levels. The animals were maintained in
observation for at least 15 days and then released to the wild. All
animals survived the procedure and none of them showed signs of
infection or behavioral alterations during the observation period.
All the samples obtained for living animals were collected in
a 1.5 mL plastic tube containing approximately 450 μl of phos-
phate buffer saline (PBS). Sperm concentration was estimated us-
ing a Neubauer chamber and the samples were diluted in PBS to a
1 × 106 cells/mL prior to observation. Sperm from these samples
were motile and were fixed for photography with formaldehyde
2%. In the case of preserved animals the vas deferens were gently
minced with a pair of scissors while immersed in PBS before
sperm collection.
SPERM MORPHOMETRY
The samples were examined at 400× magnification under a phase
contrast microscopy. Microphotographs of the samples were taken
using a Sony DSCW50 digital camera (Sony Corporation, Tokyo,
Japan) with a 6× zoom. The lengths (μm) of head (HL), midpiece
(MPL), and principal piece (PPL) and total sperm length (TSL)
of a minimum of 15 spermatozoa per sample were measured us-
ing software Image J version 1.38 (NIH, Bethesda, MD). Mean
trait values for each species were calculated from the means from
each individual of that species. Within the four species (Both-
rops diporus, Liophis poecilogyrus, Lystrophis dorbignyi, and
Philodryas patagoniensis) for which data from living and pre-
served specimens were used, no differences between them were
found on sperm length or the length of any of the sperm compo-
nents (Kruskal–Wallis n.s. in all comparisons).
SPERM ULTRASTRUCTURE
Sperm samples of living specimens of five species (Boa constric-
tor occidentalis, Bothrops alternatus, Bothrops diporus, Liophis
poecilogyrus, and Waglerophis merremii) were used for transmis-
sion electron microscopy analysis. Sperm samples were processed
as previously described in other studies (Tourmente et al. 2006,
2008). Briefly, the samples were washed twice with PBS and cen-
trifuged for 7 minutes at 700G. The supernatant was eliminated
and the resulting pellet was fixed during 3 h at room temperature
in a 3% glutaraldehyde solution in 0.1 M sodium cacodylate. The
samples were postfixed in 1% osmium tetroxide in cacodylate
buffer during 2 h at room temperature. The material was washed
with distilled water and dehydrated in a series of ascending ace-
tone concentrations (50%, 70%, 90%, and 100%), one intermedi-
ate step consisting of 50% acetone and 50% Araldite epoxy resin
and two final steps consisting of 100% resin. The samples were
then embedded in Araldite epoxy resin at 60 ◦C for 24–72 h. Ultra-
thin sections (approximately 60 nm) were mounted on 250 mesh
nickel grids, and stained with a saturated solution of uranyl in
etanol and lead citrate. Finally, the specimens were observed with
a Zeiss LEO 906E transmission electron microscope (Carl Zeiss
AG, Oberkochen, Germany) at different magnifications. The area
(μm2) of the fibrous sheath and the outer dense fibers in position
3 and 8 was measured using Image J version 1.38 (NIH).
DATA ANALYSIS
To evaluate the influence of relative testes mass on sperm di-
mensions, we used HL, MPL, PPL, and TSL for all species
as dependent variables and relative testes mass (i.e., an index
of sperm competition) as predictor. To accurately represent rel-
ative testes mass as a measure of sperm competition, we per-
formed multiple regression analyses including both testes mass
and body mass as predictors of sperm dimensions. Moreover, be-
cause our exploratory analyses appeared to indicate that reproduc-
tive mode could influence sperm competition in snakes, we added
this variable as a factor in our model. Because predictor variables
were related to each other (thus nonorthogonal), multiple regres-
sion analysis was performed using a sequential (Type I) sum of
squares, in which the predictor variables were added to the model
EVOLUTION 2009 3

MAXIMILIANO TOURMENTE ET AL.
in the following order: reproductive mode, body mass, testes
mass.
Additionally, we tested the association between relative testes
mass and ultrastructural measurements within the midpiece. This
analysis was performed via multiple regressions using the areas
of the fibrous sheath and outer dense fibers 3 and 8 as dependent
variables, with testes mass and body mass as predictors. We could
not include reproductive mode as a factor in this analysis because
of the low number of datapoints, therefore inferences about its
effects on sperm ultrastructure will not be made in this study.
Given that species data are not necessarily free of phyloge-
netic association, they may share character values as a result of a
common ancestry rather than independent evolution (Harvey and
Pagel 1991), and thus, would not provide independent datapoints
for comparative analysis (Felsenstein 1985). To control for this
phylogenetic inertia, we used a generalized least-squares (GLS)
approach in a phylogenetic framework (Freckleton et al. 2002).
This method estimates (using maximum likelihood) a phyloge-
netic scaling parameter lambda (λ), which represents the transfor-
mation that makes the data fit the Brownian motion evolutionary
model. If λ values are close to 0, the variables are likely to have
evolved independently of phylogeny, whereas λ values close to 1
indicate strong phylogenetic association of the variables. The GLS
method allows a variable degree of phylogenetic correction ac-
cording each tested model, and this constitutes an advantage when
different traits have different levels of phylogenetic association.
The estimation of λ values and GLS analyses were performed
using a code written by R. Freckleton for the statistical package
R version 2.7.2 (R Foundation for Statistical Computing, Vienna,
Austria) and the maximum likelihood value of λ was compared
against models with λ = 1 and λ = 0.
Because a complete phylogeny for all analyzed species was
not available, a phylogenetic reconstruction was used (Fig. 1). To
construct it, the phylogeny of the Neotropical Xenodontines based
in molecular characters (Vidal et al. 2000) provided most of the
tree; taxa not included in that tree were added from other studies:
an hemipenial morphology analysis for other Xenodontine snakes
(Zaher 1999), a morphologic phylogeny for pit vipers (Martins
et al. 2002), and two molecular phylogenies (Slowinski and Law-
son 2002; Vidal and Hedges 2002) for the placement of Boa, the
Elapidae, and the general relationships among the groups. Be-
cause the branch lengths were available only for a small fraction
of the species, and some phylogenetic relationships are uncertain,
equal branch lengths were used in the analysis.
The original data were log10-transformed to normalize the
distributions and all the analyses were conducted with R ver-
sion 2.7.2 and InfoStat version 2007p (Grupo Infostat, Facultad
de Ciencias Agropecuarias-Universidad Nacional de Cordoba,
Cordoba, Argentina) at α = 0.05. We avoided the use of Bon-
ferroni correction because it increases the chances of committing
Figure 1. Phylogenetic reconstruction for the species included in
the study. This reconstruction was based on Vidal et al. (2000),
Zaher (1999), Martins et al. (2002), Vidal and Hedges (2002), and
Slowinski and Lawson (2002).
type II errors (Nagakawa 2004). Alternatively, we calculated the
effect size r from F values (Rosenthal 1991; Rosenthal 1994;
Rosnow and Rosenthal 2003) obtained from the GLS model; ef-
fect sizes ≥ 0.5 were considered large (Cohen 1988). Finally, we
calculated the noncentral confidence limits (CLs) for r, which
indicate statistical significance if 0 is not contained within the
interval (Smithson 2003).
To be able to show values for relative testes size in the figures,
we used gonadosomatic index (GSI), which was calculated as
testes mass (g) divided by body mass (g), multiplied by 100
(Olsson and Madsen 1998). However, because this measure has
been criticized as an accurate index of sperm competition levels
due to allometric problems (Tomkins and Simmons 2002), we
have not used it in any of the statistical analyses.
ResultsSPERM MORPHOMETRY
The spermatozoa of all analyzed species followed the general mor-
phological pattern described for the ophidian (Oliver et al. 1996)
as they presented a filiform shape and were clearly divided in
two parts: head and flagellum, the latter containing two sections
4 EVOLUTION 2009
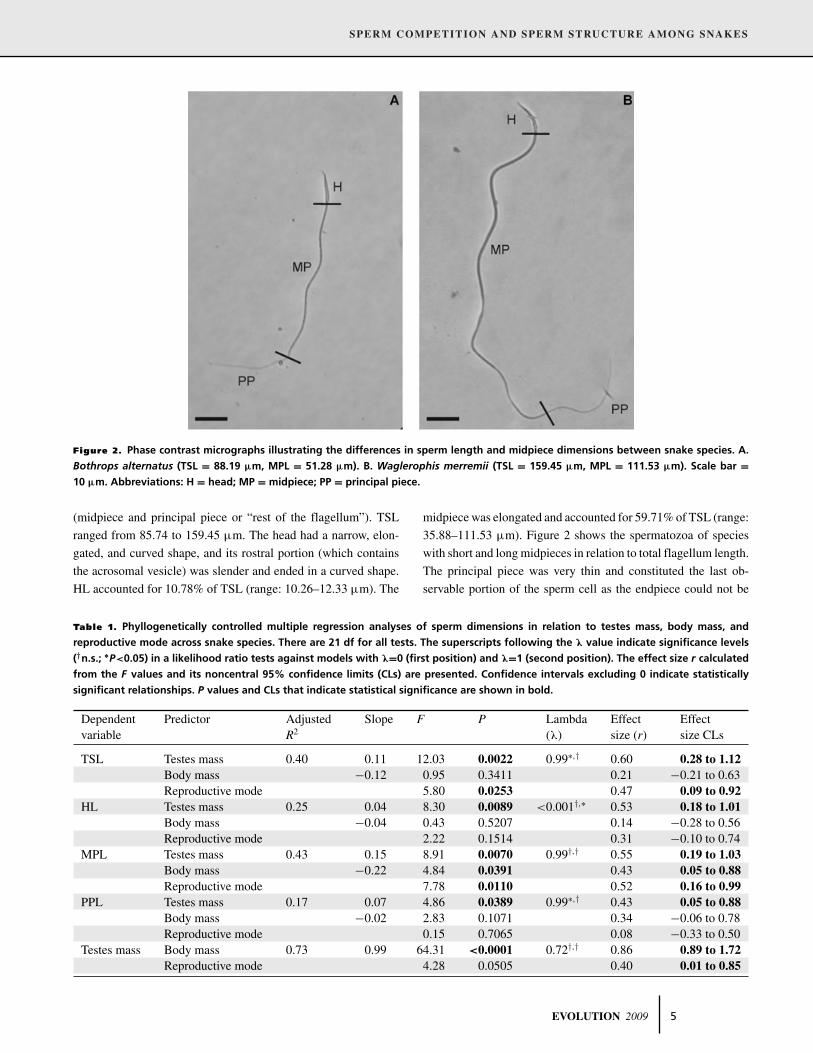
SPERM COMPETITION AND SPERM STRUCTURE AMONG SNAKES
Figure 2. Phase contrast micrographs illustrating the differences in sperm length and midpiece dimensions between snake species. A.
Bothrops alternatus (TSL = 88.19 μm, MPL = 51.28 μm). B. Waglerophis merremii (TSL = 159.45 μm, MPL = 111.53 μm). Scale bar =10 μm. Abbreviations: H = head; MP = midpiece; PP = principal piece.
(midpiece and principal piece or “rest of the flagellum”). TSL
ranged from 85.74 to 159.45 μm. The head had a narrow, elon-
gated, and curved shape, and its rostral portion (which contains
the acrosomal vesicle) was slender and ended in a curved shape.
HL accounted for 10.78% of TSL (range: 10.26–12.33 μm). The
Table 1. Phyllogenetically controlled multiple regression analyses of sperm dimensions in relation to testes mass, body mass, and
reproductive mode across snake species. There are 21 df for all tests. The superscripts following the λ value indicate significance levels
(†n.s.; ∗P<0.05) in a likelihood ratio tests against models with λ=0 (first position) and λ=1 (second position). The effect size r calculated
from the F values and its noncentral 95% confidence limits (CLs) are presented. Confidence intervals excluding 0 indicate statistically
significant relationships. P values and CLs that indicate statistical significance are shown in bold.
Dependent Predictor Adjusted Slope F P Lambda Effect Effectvariable R2 (λ) size (r) size CLs
TSL Testes mass 0.40 0.11 12.03 0.0022 0.99∗,† 0.60 0.28 to 1.12Body mass −0.12 0.95 0.3411 0.21 −0.21 to 0.63Reproductive mode 5.80 0.0253 0.47 0.09 to 0.92
HL Testes mass 0.25 0.04 8.30 0.0089 <0.001†,∗ 0.53 0.18 to 1.01Body mass −0.04 0.43 0.5207 0.14 −0.28 to 0.56Reproductive mode 2.22 0.1514 0.31 −0.10 to 0.74
MPL Testes mass 0.43 0.15 8.91 0.0070 0.99†,† 0.55 0.19 to 1.03Body mass −0.22 4.84 0.0391 0.43 0.05 to 0.88Reproductive mode 7.78 0.0110 0.52 0.16 to 0.99
PPL Testes mass 0.17 0.07 4.86 0.0389 0.99∗,† 0.43 0.05 to 0.88Body mass −0.02 2.83 0.1071 0.34 −0.06 to 0.78Reproductive mode 0.15 0.7065 0.08 −0.33 to 0.50
Testes mass Body mass 0.73 0.99 64.31 <0.0001 0.72†,† 0.86 0.89 to 1.72Reproductive mode 4.28 0.0505 0.40 0.01 to 0.85
midpiece was elongated and accounted for 59.71% of TSL (range:
35.88–111.53 μm). Figure 2 shows the spermatozoa of species
with short and long midpieces in relation to total flagellum length.
The principal piece was very thin and constituted the last ob-
servable portion of the sperm cell as the endpiece could not be
EVOLUTION 2009 5
MAXIMILIANO TOURMENTE ET AL.
distinguished. It accounted for 29.76% of TSL (range: 25.32–
43.52 μm). The midpiece showed extreme interspecies variability
(CV = 30.276) in comparison to head and principal piece (CV =4.595 and CV = 12.857, respectively).
ASSOCIATIONS BETWEEN SPERM MORPHOMETRY,
RELATIVE TESTES MASS, AND REPRODUCTIVE MODE
Both MPL and PPL showed a positive correlation with TSL after
accounting for phylogenetic effects (MPL: P < 0.0001, R2 =0.87; PPL: P = 0.0295, R2 = 0.19), which was stronger for MPL.
On the other hand, HL was not significantly associated with any
other sperm component; neither was midpiece related to PPL.
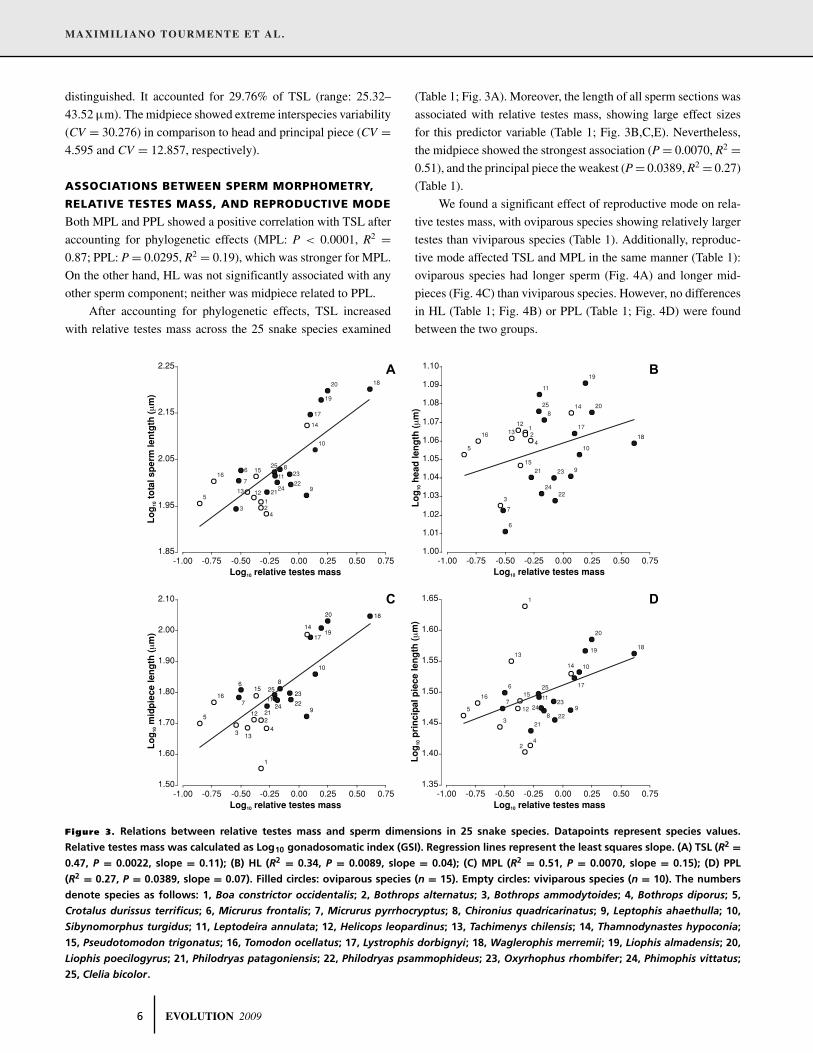
After accounting for phylogenetic effects, TSL increased
with relative testes mass across the 25 snake species examined
Figure 3. Relations between relative testes mass and sperm dimensions in 25 snake species. Datapoints represent species values.
Relative testes mass was calculated as Log10 gonadosomatic index (GSI). Regression lines represent the least squares slope. (A) TSL (R2 =0.47, P = 0.0022, slope = 0.11); (B) HL (R2 = 0.34, P = 0.0089, slope = 0.04); (C) MPL (R2 = 0.51, P = 0.0070, slope = 0.15); (D) PPL
(R2 = 0.27, P = 0.0389, slope = 0.07). Filled circles: oviparous species (n = 15). Empty circles: viviparous species (n = 10). The numbers
denote species as follows: 1, Boa constrictor occidentalis; 2, Bothrops alternatus; 3, Bothrops ammodytoides; 4, Bothrops diporus; 5,
Crotalus durissus terrificus; 6, Micrurus frontalis; 7, Micrurus pyrrhocryptus; 8, Chironius quadricarinatus; 9, Leptophis ahaethulla; 10,
Sibynomorphus turgidus; 11, Leptodeira annulata; 12, Helicops leopardinus; 13, Tachimenys chilensis; 14, Thamnodynastes hypoconia;
15, Pseudotomodon trigonatus; 16, Tomodon ocellatus; 17, Lystrophis dorbignyi; 18, Waglerophis merremii; 19, Liophis almadensis; 20,
Liophis poecilogyrus; 21, Philodryas patagoniensis; 22, Philodryas psammophideus; 23, Oxyrhophus rhombifer; 24, Phimophis vittatus;
25, Clelia bicolor.
(Table 1; Fig. 3A). Moreover, the length of all sperm sections was
associated with relative testes mass, showing large effect sizes
for this predictor variable (Table 1; Fig. 3B,C,E). Nevertheless,
the midpiece showed the strongest association (P = 0.0070, R2 =0.51), and the principal piece the weakest (P = 0.0389, R2 = 0.27)
(Table 1).
We found a significant effect of reproductive mode on rela-
tive testes mass, with oviparous species showing relatively larger
testes than viviparous species (Table 1). Additionally, reproduc-
tive mode affected TSL and MPL in the same manner (Table 1):
oviparous species had longer sperm (Fig. 4A) and longer mid-
pieces (Fig. 4C) than viviparous species. However, no differences
in HL (Table 1; Fig. 4B) or PPL (Table 1; Fig. 4D) were found
between the two groups.
6 EVOLUTION 2009
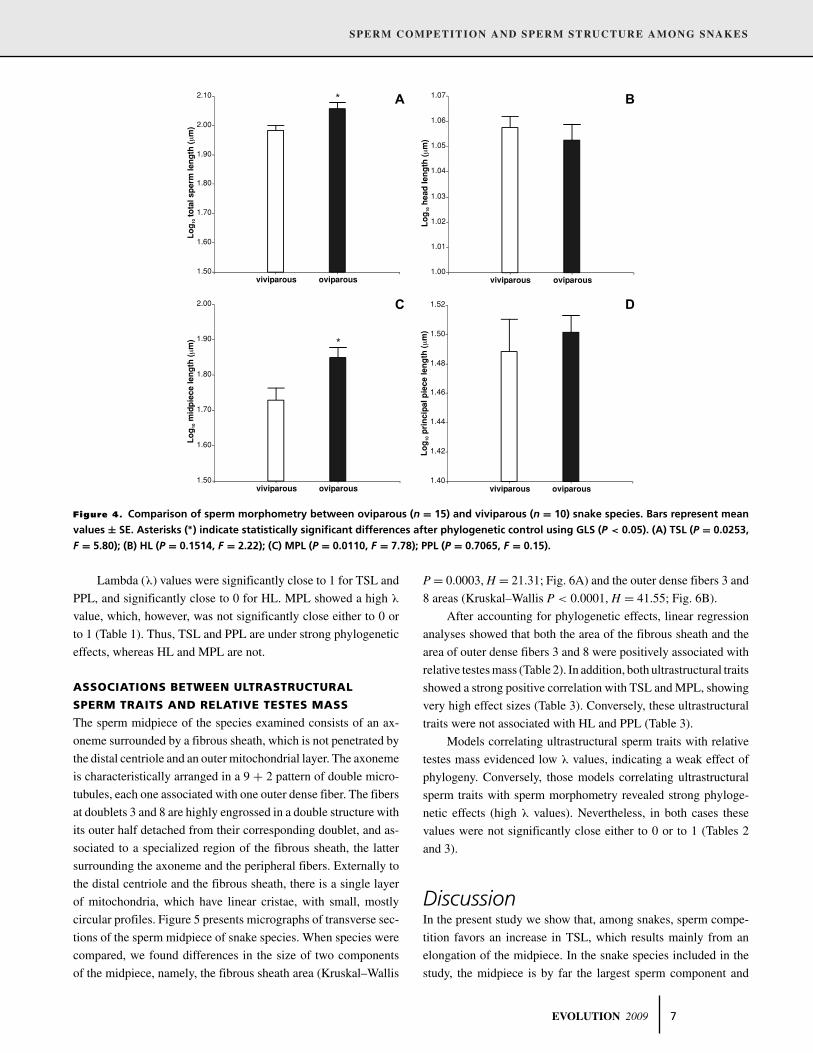
SPERM COMPETITION AND SPERM STRUCTURE AMONG SNAKES
Figure 4. Comparison of sperm morphometry between oviparous (n = 15) and viviparous (n = 10) snake species. Bars represent mean
values ± SE. Asterisks (∗) indicate statistically significant differences after phylogenetic control using GLS (P < 0.05). (A) TSL (P = 0.0253,
F = 5.80); (B) HL (P = 0.1514, F = 2.22); (C) MPL (P = 0.0110, F = 7.78); PPL (P = 0.7065, F = 0.15).
Lambda (λ) values were significantly close to 1 for TSL and
PPL, and significantly close to 0 for HL. MPL showed a high λ
value, which, however, was not significantly close either to 0 or
to 1 (Table 1). Thus, TSL and PPL are under strong phylogenetic
effects, whereas HL and MPL are not.
ASSOCIATIONS BETWEEN ULTRASTRUCTURAL
SPERM TRAITS AND RELATIVE TESTES MASS
The sperm midpiece of the species examined consists of an ax-
oneme surrounded by a fibrous sheath, which is not penetrated by
the distal centriole and an outer mitochondrial layer. The axoneme
is characteristically arranged in a 9 + 2 pattern of double micro-
tubules, each one associated with one outer dense fiber. The fibers
at doublets 3 and 8 are highly engrossed in a double structure with
its outer half detached from their corresponding doublet, and as-
sociated to a specialized region of the fibrous sheath, the latter
surrounding the axoneme and the peripheral fibers. Externally to
the distal centriole and the fibrous sheath, there is a single layer
of mitochondria, which have linear cristae, with small, mostly
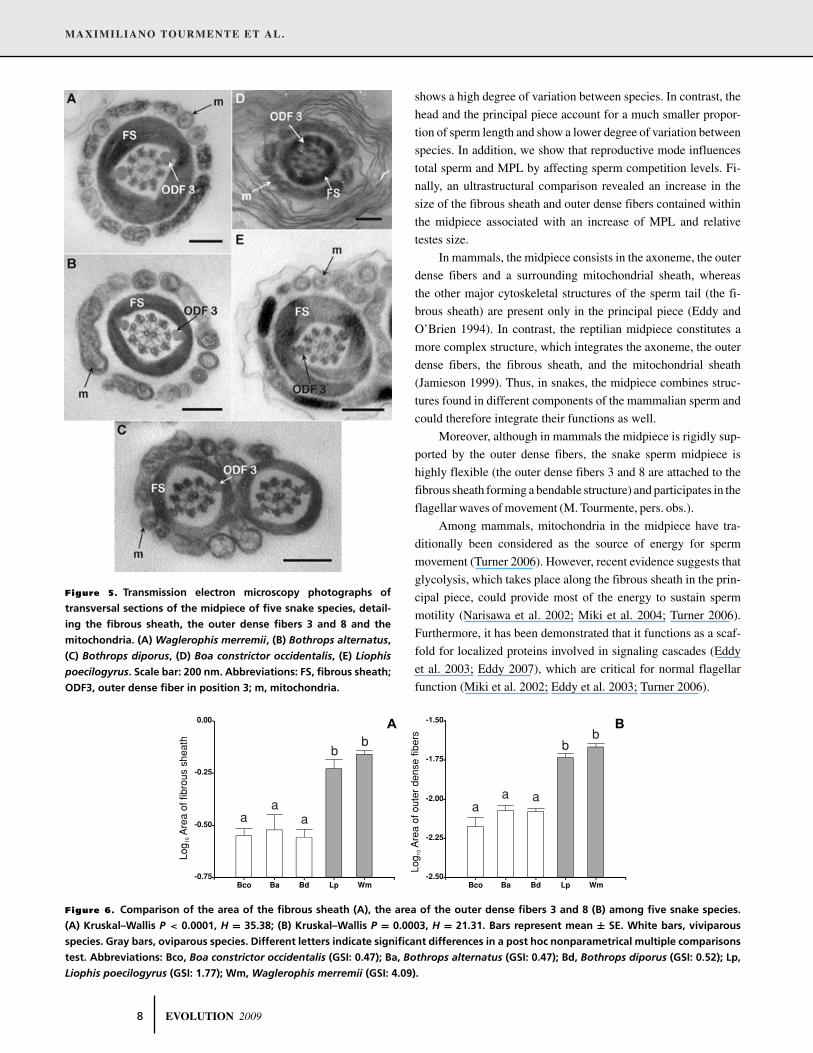
circular profiles. Figure 5 presents micrographs of transverse sec-
tions of the sperm midpiece of snake species. When species were
compared, we found differences in the size of two components
of the midpiece, namely, the fibrous sheath area (Kruskal–Wallis
P = 0.0003, H = 21.31; Fig. 6A) and the outer dense fibers 3 and
8 areas (Kruskal–Wallis P < 0.0001, H = 41.55; Fig. 6B).
After accounting for phylogenetic effects, linear regression
analyses showed that both the area of the fibrous sheath and the
area of outer dense fibers 3 and 8 were positively associated with
relative testes mass (Table 2). In addition, both ultrastructural traits
showed a strong positive correlation with TSL and MPL, showing
very high effect sizes (Table 3). Conversely, these ultrastructural
traits were not associated with HL and PPL (Table 3).
Models correlating ultrastructural sperm traits with relative
testes mass evidenced low λ values, indicating a weak effect of
phylogeny. Conversely, those models correlating ultrastructural
sperm traits with sperm morphometry revealed strong phyloge-
netic effects (high λ values). Nevertheless, in both cases these
values were not significantly close either to 0 or to 1 (Tables 2
and 3).
DiscussionIn the present study we show that, among snakes, sperm compe-
tition favors an increase in TSL, which results mainly from an
elongation of the midpiece. In the snake species included in the
study, the midpiece is by far the largest sperm component and
EVOLUTION 2009 7
MAXIMILIANO TOURMENTE ET AL.
Figure 5. Transmission electron microscopy photographs of
transversal sections of the midpiece of five snake species, detail-
ing the fibrous sheath, the outer dense fibers 3 and 8 and the
mitochondria. (A) Waglerophis merremii, (B) Bothrops alternatus,
(C) Bothrops diporus, (D) Boa constrictor occidentalis, (E) Liophis
poecilogyrus. Scale bar: 200 nm. Abbreviations: FS, fibrous sheath;
ODF3, outer dense fiber in position 3; m, mitochondria.
Figure 6. Comparison of the area of the fibrous sheath (A), the area of the outer dense fibers 3 and 8 (B) among five snake species.
(A) Kruskal–Wallis P < 0.0001, H = 35.38; (B) Kruskal–Wallis P = 0.0003, H = 21.31. Bars represent mean ± SE. White bars, viviparous
species. Gray bars, oviparous species. Different letters indicate significant differences in a post hoc nonparametrical multiple comparisons
test. Abbreviations: Bco, Boa constrictor occidentalis (GSI: 0.47); Ba, Bothrops alternatus (GSI: 0.47); Bd, Bothrops diporus (GSI: 0.52); Lp,
Liophis poecilogyrus (GSI: 1.77); Wm, Waglerophis merremii (GSI: 4.09).
shows a high degree of variation between species. In contrast, the
head and the principal piece account for a much smaller propor-
tion of sperm length and show a lower degree of variation between
species. In addition, we show that reproductive mode influences
total sperm and MPL by affecting sperm competition levels. Fi-
nally, an ultrastructural comparison revealed an increase in the
size of the fibrous sheath and outer dense fibers contained within
the midpiece associated with an increase of MPL and relative
testes size.
In mammals, the midpiece consists in the axoneme, the outer
dense fibers and a surrounding mitochondrial sheath, whereas
the other major cytoskeletal structures of the sperm tail (the fi-
brous sheath) are present only in the principal piece (Eddy and
O’Brien 1994). In contrast, the reptilian midpiece constitutes a
more complex structure, which integrates the axoneme, the outer
dense fibers, the fibrous sheath, and the mitochondrial sheath
(Jamieson 1999). Thus, in snakes, the midpiece combines struc-
tures found in different components of the mammalian sperm and
could therefore integrate their functions as well.
Moreover, although in mammals the midpiece is rigidly sup-
ported by the outer dense fibers, the snake sperm midpiece is
highly flexible (the outer dense fibers 3 and 8 are attached to the
fibrous sheath forming a bendable structure) and participates in the
flagellar waves of movement (M. Tourmente, pers. obs.).
Among mammals, mitochondria in the midpiece have tra-
ditionally been considered as the source of energy for sperm
movement (Turner 2006). However, recent evidence suggests that
glycolysis, which takes place along the fibrous sheath in the prin-
cipal piece, could provide most of the energy to sustain sperm
motility (Narisawa et al. 2002; Miki et al. 2004; Turner 2006).
Furthermore, it has been demonstrated that it functions as a scaf-
fold for localized proteins involved in signaling cascades (Eddy
et al. 2003; Eddy 2007), which are critical for normal flagellar
function (Miki et al. 2002; Eddy et al. 2003; Turner 2006).
8 EVOLUTION 2009
SPERM COMPETITION AND SPERM STRUCTURE AMONG SNAKES
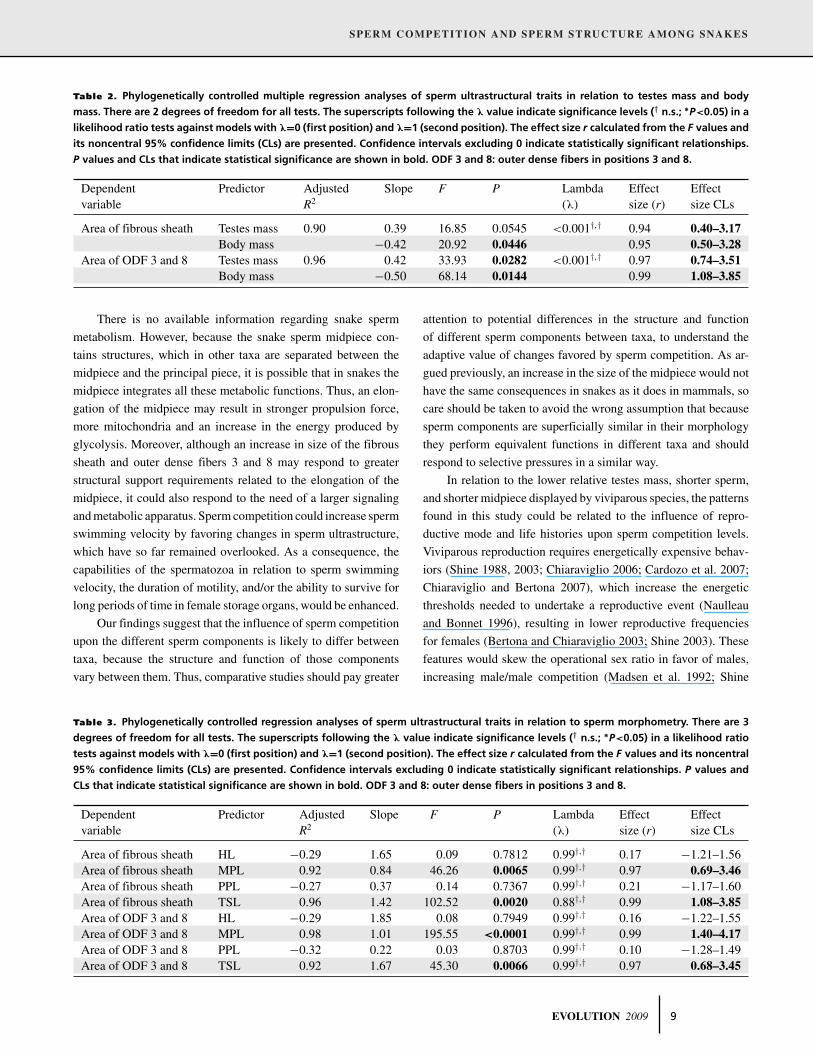
Table 2. Phylogenetically controlled multiple regression analyses of sperm ultrastructural traits in relation to testes mass and body
mass. There are 2 degrees of freedom for all tests. The superscripts following the λ value indicate significance levels († n.s.; ∗P<0.05) in a
likelihood ratio tests against models with λ=0 (first position) and λ=1 (second position). The effect size r calculated from the F values and
its noncentral 95% confidence limits (CLs) are presented. Confidence intervals excluding 0 indicate statistically significant relationships.
P values and CLs that indicate statistical significance are shown in bold. ODF 3 and 8: outer dense fibers in positions 3 and 8.
Dependent Predictor Adjusted Slope F P Lambda Effect Effectvariable R2 (λ) size (r) size CLs
Area of fibrous sheath Testes mass 0.90 0.39 16.85 0.0545 <0.001†,† 0.94 0.40–3.17Body mass −0.42 20.92 0.0446 0.95 0.50–3.28
Area of ODF 3 and 8 Testes mass 0.96 0.42 33.93 0.0282 <0.001†,† 0.97 0.74–3.51Body mass −0.50 68.14 0.0144 0.99 1.08–3.85
There is no available information regarding snake sperm
metabolism. However, because the snake sperm midpiece con-
tains structures, which in other taxa are separated between the
midpiece and the principal piece, it is possible that in snakes the
midpiece integrates all these metabolic functions. Thus, an elon-
gation of the midpiece may result in stronger propulsion force,
more mitochondria and an increase in the energy produced by
glycolysis. Moreover, although an increase in size of the fibrous
sheath and outer dense fibers 3 and 8 may respond to greater
structural support requirements related to the elongation of the
midpiece, it could also respond to the need of a larger signaling
and metabolic apparatus. Sperm competition could increase sperm
swimming velocity by favoring changes in sperm ultrastructure,
which have so far remained overlooked. As a consequence, the
capabilities of the spermatozoa in relation to sperm swimming
velocity, the duration of motility, and/or the ability to survive for
long periods of time in female storage organs, would be enhanced.
Our findings suggest that the influence of sperm competition
upon the different sperm components is likely to differ between
taxa, because the structure and function of those components
vary between them. Thus, comparative studies should pay greater
Table 3. Phylogenetically controlled regression analyses of sperm ultrastructural traits in relation to sperm morphometry. There are 3
degrees of freedom for all tests. The superscripts following the λ value indicate significance levels († n.s.; ∗P<0.05) in a likelihood ratio
tests against models with λ=0 (first position) and λ=1 (second position). The effect size r calculated from the F values and its noncentral
95% confidence limits (CLs) are presented. Confidence intervals excluding 0 indicate statistically significant relationships. P values and
CLs that indicate statistical significance are shown in bold. ODF 3 and 8: outer dense fibers in positions 3 and 8.
Dependent Predictor Adjusted Slope F P Lambda Effect Effectvariable R2 (λ) size (r) size CLs
Area of fibrous sheath HL −0.29 1.65 0.09 0.7812 0.99†,† 0.17 −1.21–1.56Area of fibrous sheath MPL 0.92 0.84 46.26 0.0065 0.99†,† 0.97 0.69–3.46Area of fibrous sheath PPL −0.27 0.37 0.14 0.7367 0.99†,† 0.21 −1.17–1.60Area of fibrous sheath TSL 0.96 1.42 102.52 0.0020 0.88†,† 0.99 1.08–3.85Area of ODF 3 and 8 HL −0.29 1.85 0.08 0.7949 0.99†,† 0.16 −1.22–1.55Area of ODF 3 and 8 MPL 0.98 1.01 195.55 <0.0001 0.99†,† 0.99 1.40–4.17Area of ODF 3 and 8 PPL −0.32 0.22 0.03 0.8703 0.99†,† 0.10 −1.28–1.49Area of ODF 3 and 8 TSL 0.92 1.67 45.30 0.0066 0.99†,† 0.97 0.68–3.45
attention to potential differences in the structure and function
of different sperm components between taxa, to understand the
adaptive value of changes favored by sperm competition. As ar-
gued previously, an increase in the size of the midpiece would not
have the same consequences in snakes as it does in mammals, so
care should be taken to avoid the wrong assumption that because
sperm components are superficially similar in their morphology
they perform equivalent functions in different taxa and should
respond to selective pressures in a similar way.
In relation to the lower relative testes mass, shorter sperm,
and shorter midpiece displayed by viviparous species, the patterns
found in this study could be related to the influence of repro-
ductive mode and life histories upon sperm competition levels.
Viviparous reproduction requires energetically expensive behav-
iors (Shine 1988, 2003; Chiaraviglio 2006; Cardozo et al. 2007;
Chiaraviglio and Bertona 2007), which increase the energetic
thresholds needed to undertake a reproductive event (Naulleau
and Bonnet 1996), resulting in lower reproductive frequencies
for females (Bertona and Chiaraviglio 2003; Shine 2003). These
features would skew the operational sex ratio in favor of males,
increasing male/male competition (Madsen et al. 1992; Shine
EVOLUTION 2009 9

MAXIMILIANO TOURMENTE ET AL.
2003). However, five out of 10 viviparous species analyzed in
the present study (the genera Boa, Bothrops, and Crotalus) are
included in taxa, which are associated with the “prolonged mate
searching polygyny” (the mating system of the other viviparous
species are unknown). This mating pattern selects for males that
have the skill and endurance to transverse long distances (Duvall
et al. 1992; Rivera et al. 2006) searching for widely distributed,
spatially unpredictable, and/or scarce females (Duvall et al. 1992),
while forgoing feeding (Madsen and Shine 2000) and exposing
themselves to high mortality risks (Aldridge and Brown 1995;
Bonnet et al. 1999). As these activities are extremely costly for
males in terms of energy utilization (Olsson et al. 1997; Aldridge
and Duvall 2002) and females have such low spatial predictabil-
ity, the rate of male–female encounter is likely to be relatively
low. Because males are solitary searchers in snake species with
prolonged mate searching polygyny (Aldridge and Duvall 2002;
Shine 2003), this scenario would also decrease the rate of male–
male encounters, thus relaxing sperm competition pressures.
The relationships between sperm competition and sperm
morphology, ultrastructure and physiology appear extremely com-
plex and the adaptive significance of the patterns so far described
remains unclear. Studying a greater diversity of taxa will con-
tribute to provide a clearer picture of how sperm competition
shapes sperm morphology and function, and the adaptive conse-
quences of such changes.
ACKNOWLEDGMENTSWe are grateful to G. Cardozo, V. Di Cola, S. Naretto, D. Britos,and G. Bertone (FCEFyN—UNC), for helping with the capture ofliving specimens and semen extraction; to G. Leynaud (Centro deZoologıa Aplicada—UNC) for the provision of specimens; to M. Cabrera(FCEFyN—UNC), B. Alvarez and C. Calamante (FaCENA—UNNE), J.Faivovich and B. Blotto (MACN, Bernardino Rivadavia), J. Williams andE. Kubisch (Museo de Ciencias Naturales Perito Moreno) for grantingus access to the corresponding collections. We also thank C. Maldonadofor her assistance with electron microscopy and micrographic techniques,and S. Immler for her assistance with phylogenetic control methods. Thisstudy was funded by CONICET and SeCyT—UNC, Argentina, and bythe MEC, Spain. MT is a fellow of CONICET, a student of the Doc-torado en Ciencias Biologicas (FCEFyN—UNC) and the recipient of aBoehringer Ingelheim Travel Grant. ERSR is the recipient of a RoyalSociety Wolfson Research Merit Award.
LITERATURE CITEDAldridge, R. D., and W. S. Brown. 1995. Male reproductive cycle, age at
maturity, and cost of reproduction in the timber rattlesnake (Crotalushorridus). J. Herpetol. 29:399–407.
Aldridge, R. D., and D. Duvall. 2002. Evolution of the mating season in thepitvipers of North America. Herpetological Monographs 16:1–25.
Anderson, M. J., and A. F. Dixson. 2002. Sperm competition: motility and themidpiece in primates. Nature 416:496.
Anderson, M. J., J. Nyholt, and A. F. Dixson. 2005. Sperm competition andthe evolution of sperm midpiece volume in mammals. J. Zool. 267:135–145.
Anderson, M. J., A. S. Dixson, and A. F. Dixson. 2006. Mammalian spermand oviducts are sexually selected: evidence for co-evolution. J. Zool.270:682–686.
Balshine, S., B. J. Leach, F. Neat, N. Y. Werner, and R. Montgomerie. 2001.Sperm size of African cichlids in relation to sperm competition. Behav.Ecol. 12:726–731.
Bertona, M., and M. Chiaraviglio. 2003. Reproductive biology, mating aggre-gations, and sexual dimorphism of the argentine boa constrictor (Boa
constrictor occidentalis). J. Herpetol. 37:510–516.Birkhead, T. R., J. G. Martinez, T. Burke, and D. P. Froman. 1999.
Sperm mobility determines the outcome of sperm competition inthe domestic fowl. Proc. R. Soc. Lond., B, Biol. Sci. 266:1759–1764.
Bonnet, X., G. Naulleau, and R. Shine. 1999. The dangers of leaving home:dispersal and mortality in snakes. Biol. Conserv. 89:39–50.
Briskie, J. V., and R. Montgomerie. 1992. Sperm size and sperm competitionin birds. Proc. R. Soc. Lond., B, Biol. Sci. 247:89–95.
Briskie, J. V., R. Montgomerie, and T. R. Birkhead. 1997. The evolution ofsperm size in birds. Evolution 51:937–945.
Brown, G. P., and R. Shine. 2002. Reproductive ecology of a tropical natricinesnake, Tropidonophis mairii (Colubridae). J. Zool. 258:63–72.
Byrne, P. G., J. D. Roberts, and L. W. Simmons. 2002. Sperm competi-tion selects for increased testes mass in Australian frogs. J. Evol. Biol.15:347–355.
Byrne, P. G., L. W. Simmons, and R. J. Dale. 2003. Sperm competition andthe evolution of gamete morphology in frogs. Proc. R. Soc. Lond., B,Biol. Sci. 270:2079–2086.
Cardozo, G., P. C. Rivera, M. Lanfri, M. Scavuzzo, C. N. Gardenal, and M.Chiaraviglio. 2007. Effects of habitat loss on the genetic structure ofpopulations of the Argentine boa constrictor (Boa constrictor occiden-
talis). Pp. 328–339 in R. W. Henderson and R. Powell, eds. Biologyof the Boas and Pythons. Eagle Mountain Publishing, Eagle Mountain,Utah.
Carpenter, J. W., T. Y. Mashima, and D. J. Rupiper. 2001. Exotic AnimalFormulary. Saunders Co., Philadelphia, PA.
Chiaraviglio, M. 2006. The effects of reproductive condition on thermoreg-ulation in the Argentina boa constrictor (Boa constrictor occidentalis)(Boidae). Herpetological Monographs 20:172–177.
Chiaraviglio, M., and M. Bertona. 2007. Reproduction and thermoregulationare principal factors that influence habitat selection in the Argentine boaconstrictor (Boa constrictor occidentalis). Pp. 76–87 in R. W. Hendersonand R. Powell, eds. Biology of the Boas and Pythons. Eagle MountainPublishing, Eagle Mountain, Utah.
Clark, A. G., D. J. Begun, and T. Prout. 1999. Female x male interactions inDrosophila sperm competition. Science 283:217–220.
Cohen, J. 1988. Statistical power analysis for the behavioral sciences.Erlbaum, Hillsdale, NJ.
Cook, P. A., and M. J. G. Gage. 1995. Effects of risks of sperm competitionon the numbers of eupyrene and apyrene sperm ejaculated by the mothPlodia interpunctella (Lepidoptera: Pyralidae). Behav. Ecol. Sociobiol.36:261–268.
Duvall, D., S. J. Arnold, and G. W. Schuett. 1992. Pitviper mating systems:Ecological potential, sexual selection and microevolution. Pp. 321–336in J. A. Campbell and E. D. Brodie Jr., eds. Biology of the Pitvipers.Selva Press, Tyler, TX.
Eddy, E. M. 2007. The scaffold role of the fibrous sheath. Pp. 45–62 in E.R. S. Roldan and M. Gomendio, eds. Spermatology. Nottingham Univ.Press, Nottingham, U.K.
Eddy, E. M., and D. A. O’Brien 1994. The spermatozoon. Pp. 29–77 in E.Knobil and J. D. Neil, eds. The physiology of reproduction. Raven Press,New York.
1 0 EVOLUTION 2009

SPERM COMPETITION AND SPERM STRUCTURE AMONG SNAKES
Eddy, E. M., K. Toshimori, and D. A. O’Brien. 2003. Fibrous sheath ofmammalian spermatozoa. Microsc. Res. Tech. 61:103–115.
Felsenstein, J. 1985. Phylogenies and the comparative method. Am. Nat.125:1–15.
Fitzpatrick, J. L., R. Montgomerie, J. K. Desjardins, K. A. Stiver, N. Kolm, andS. Balshine. 2009. Female promiscuity promotes the evolution of fastersperm in cichlid fishes. Proc. Natl. Acad. Sci. U.S.A. 106:1128–1132.
Freckleton, R., P. H. Harvey, and M. D. Pagel. 2002. Phylogenetic analysisand comparative data: a test and review of evidence. Am. Nat. 160:712–726.
Froman, D. P., T. Pizzari, A. J. Feltmann, H. Castillo, and T. R. Birkhead. 2002.Sperm mobility: mechanisms of fertilizing efficiency, genetic variationand phenotypic relationship with male status in the domestic fowl, Gallus
gallus domesticus. Proc. R. Soc. Lond., B, Biol. Sci. 269:607–612.Gage, M. J. G. 1994. Associations between body size, mating pattern, testis
size and sperm lengths across butterflies. Proc. R. Soc. Lond., B, Biol.Sci. 258:247–254.
Gage, M. J. G., C. P. Macfarlane, S. Yeates, R. G. Ward, J. B. Searle, and G.A. Parker. 2004. Spermatozoal traits and sperm competition in atlanticsalmon: relative sperm velocity is the primary determinant of fertilizationsuccess. Curr. Biol. 14:44–47.
Gomendio, M., and E. R. S. Roldan. 1991. Sperm competition influencessperm size in mammals. Proc. R. Soc. Lond., B, Biol. Sci. 243:181–185.
———. 1993. Coevolution between male ejaculates and female reproductivebiology in eutherian mammals. Proc. R. Soc. Lond., B, Biol. Sci. 252:7–12.
———. 2008. Implications of diversity in sperm size and function for spermcompetition and fertility. Int. J. Dev. Biol. 52:439–447.
Gomendio, M., A. H. Harcourt, and E. R. S. Roldan. 1998. Sperm competitionin mammals. Pp. 667–756 in T. R. Birkhead and A. P. Moller, eds. Spermcompetition and sexual selection. Academic Press, San Diego, CA.
Gomendio, M., J. Martin-Coello, C. Crespo, C. Magana, and E. R. S. Roldan.2006. Sperm competition enhances functional capacity of mammalianspermatozoa. Proc. Natl. Acad. Sci. U.S.A. 103:15113–15117.
Harvey, P. H., and M. D. Pagel. 1991. The comparative method in evolutionarybiology. Oxford Univ. Press, Oxford, U.K.
Holman, L., R. Freckleton, and R. R. Snook. 2008. What use is an infer-tile sperm? A comparative study of sperm-heteromorphic Drosophila.Evolution 62:374–385.
Hosken, D. J. 1997. Sperm competition in bats. Proc. R. Soc. Lond., B, Biol.Sci. 264:385–392.
Hunter, F. M., and T. R. Birkhead. 2002. Sperm viability and sperm competi-tion in insects. Curr. Biol. 12:121–123.
Immler, S., and T. R. Birkhead. 2007. Sperm competition and sperm midpiecesize: no consistent pattern in passerine birds. Proc. R. Soc. Lond., B,Biol. Sci. 274:561–568.
Immler, S., M. Saint-Jalme, L. Lesobre, G. Sorci, Y. Roman, and T. R.Birkhead. 2007. The evolution of sperm morphometry in pheasants.J. Evol. Biol. 20:1008–1014.
Jamieson, B. G. M. 1999. Spermatozoal phylogeny of the vetebrata. Pp. 303–331 in C. Glagnon, ed. The male gamete: from basic science to clinicalapplications. Cache River Press, Vienna, IL.
Jennions, M. D., and N. I. Passmore. 1993. Sperm competition in frogs:testis size and “sterile male” experiment on Chiromantis xerampelina
(Rhacophoridae). Biol. J. Linn. Soc. Lond. 50:211–220.Johnson, D. P., and J. V. Briskie. 1999. Sperm competition and sperm length
in shorebirds. Condor 101:848–854.LaMunyon, C. W., and S. Ward. 1998. Larger sperm outcompete smaller
sperm in the nematode Caenorhabditis elegans. Proc. R. Soc. Lond., B,Biol. Sci. 265:1997–2002.
———. 1999. Evolution of sperm size in nematodes: sperm competitionfavours larger sperm. Proc. R. Soc. Lond., B, Biol. Sci. 266:263–267.
———. 2002. Evolution of larger sperm in response to experimentally in-creased sperm competition in Caenorhabditis elegans. Proc. R. Soc.Lond., B, Biol. Sci. 269:1125–1128.
Lupold, S., G. M. Linz, J. W. Rivers, D. F. Westneat, and T. R. Birkhead. 2008.Sperm competition selects beyond relative testes size in birds. Evolution63:391–402.
Lupold, S., S. Calhim, S. Immler, and T. R. Birkhead. 2009. Sperm morphol-ogy and sperm velocity in passerine birds. Proc. R. Soc. Lond., B, Biol.Sci. 276:1175–1181.
Mader, D. R. 2005. Reptile Medicine and Surgery. Saunders, Philadelphia,PA.
Madsen, T., and R. Shine. 2000. Energy versus risk: costs of reproduction infree-ranging pythons in tropical Australia. Aust. Ecol. 25:670–675.
Madsen, T., R. Shine, J. Loman, and T. Hakansson. 1992. Why do femaleadders copulate so frequently? Nature 355:440–441.
Malo, A. F., M. Gomendio, J. Garde, B. Lang-Lenton, A. J. Soler, and E.R. S. Roldan. 2006. Sperm design and sperm function. Biol. Lett. 2:246–249.
Martins, M., O. A. V. Marques, and I. Sazima. 2002. Ecological and phy-logenetic correlates of feeding habits in Neotropical Pitvipers of thegenus Bothrops. Pp. 307–328 in G. W. Schuett, M. Hoggren, M. E.Douglas, and H. W. Greene, eds. Biology of the Vipers. Eagle MountainPublishing, Eagle Mountain, Utah.
Mendez, F. R., and M. Villagran. 1998. Reproduccion asincronica de Scelo-
porus palaciosi (Sauria: Phrynosomatidae) en Mexico, con comentariossobre sus ventajas y regulacion. Rev. Biol. Trop. 46:1159–1161.
Miki, K., W. Willis, P. Brown, E. Goulding, K. Fulcher, and E. M. Eddy.2002. Targeted disruption of the AKAP4 gene causes defects in spermflagellum and motility. Dev. Biol. 248:331–342.
Miki, K., W. Qu, E. H. Goulding, W. D. Willis, D. O. Bunch, L. F. Strader,S. D. Perreault, E. M. Eddy, and D. A. O’Brien. 2004. Glyceraldehyde3-phosphate dehydrogenase-S, a sperm-specific glycolytic enzyme, isrequired for sperm motility and male fertility. Proc. Natl. Acad. Sci.U.S.A. 101:16501–16506.
Miller, G. T., and S. Pitnick. 2002. Sperm-Female Coevolution in Drosophila.Science 298:1230–1233.
Minder, A. M., D. J. Hosken, and P. I. Ward. 2005. Co-evolution of male andfemale reproductive characters across the Scatophagidae (Diptera). J.Evol. Biol. 18:60–69.
Morrow, E. H., and M. J. G. Gage. 2000. The evolution of sperm length inmoths. Proc. R. Soc. Lond., B, Biol. Sci. 267:307–313.
Nagakawa, S. 2004. A farewell to Bonferroni: the problems of low statisticalpower and publication bias. Behav. Ecol. 15:1044–1045.
Narisawa, S., N. B. Hecht, E. Goldberg, K. M. Boatright, J. C. Reed, and J. L.Millan. 2002. Testis-specific cytochrome c-null mice produce functionalsperm but undergo early testicular atrophy. Mol. Cell. Biol. 22:5554–5562.
Naulleau, G., and X. Bonnet. 1996. Body condition threshold for breeding ina viviparous snake. Oecologia 107:301–306.
Oliver, S. C., B. G. M. Jamieson, and D. M. Scheltinga. 1996. The ultrastruc-ture of spermatozoa of Squamata. II. Agamidae, Varanidae, Colubridae,Elapidae, and Boidae (Reptilia). Herpetologica 52:216–241.
Olsson, M., and T. Madsen. 1998. Sexual selection and sperm competition inreptiles. Pp. 503–578 in T. R. Birkhead and A. P. Moller, eds. SpermCompetition and Sexual Selection. Academic Press, San Diego, CA.
Olsson, M., T. Madsen, and R. Shine. 1997. Is sperm really so cheap? Costsof reproduction in male adders, Vipera berus. Proc. R. Soc. Lond., B,Biol. Sci. 264:455–459.
EVOLUTION 2009 1 1

MAXIMILIANO TOURMENTE ET AL.
Parker, G. A. 1998. Sperm competition and the evolution of ejaculates: towardsa theory base. Pp. 3–54 in T. R. Birkhead and A. P. Moller, eds. Spermcompetition and sexual selection. Academic Press, San Diego, CA.
Pitnick, S., G. T. Miller, M. Schneider, and T. A. Markow. 2003. Ejaculate-female coevolution in Drosophila mojavensis. Proc. R. Soc. Lond., B,Biol. Sci. 270:1507–1512.
Presgraves, D. C., R. H. Baker, and G. S. Wilkinson. 1999. Coevolution ofsperm and female reproductive tract morphology in stalk-eyed flies.Proc. R. Soc. Lond., B, Biol. Sci. 266:1041–1047.
Radwan, J. 1996. Intraspecific variation in sperm competition success in thebulb mite: a role for sperm size. Proc. R. Soc. Lond., B, Biol. Sci.263:855–859.
Rivera, P. C., C. N. Gardenal, and M. Chiaraviglio. 2006. Sex-biased dispersaland high levels of gene flow among local populations in the argentineboa constrictor, Boa constrictor occidentalis. Aust. Ecol. 31:948–955.
Roldan, E. R. S., M. Gomendio, and A. D. Vitullo. 1992. The evolution ofeuhterian spermatozoa and underlying selective forces: female selectionand sperm competition. Biol. Rev. 67:551–593.
Rosenthal, R. 1991. Meta-analytic procedures for social research. SAGE Pub-lications, Newbury Park, CA.
———. 1994. Parametric measures of effect size. Pp. 231–244 in H. Cooperand L. Hedges, eds. The handbook of research synthesis. SAGE Publi-cations, New York, NY.
Rosnow, R., and R. Rosenthal. 2003. Effect sizes for experimenting psychol-ogists. Can. J. Exp. Psychol. 57:221–237.
Scrocchi, G. J., I. Aguero, V. Arzamendia, P. Cacivio, H. Carcacha, M. Chiar-aviglio, A. Giraudo, S. Kretzschmar, G. Leynaud, M. S. Lopez, et al.2000. Categorizacion de las serpientes de Argentina. Pp. 76–93 in E.O. Lavilla, E. Richard, and G. Scrocchi, eds. Categorizacion de los an-fibios y reptiles de la Republica Argentina. Asociacion HerpetologicaArgentina, San Miguel de Tucuman, Argentina.
Sever, D. M., and W. C. Hamlett. 2002. Female sperm storage in reptiles. J.Exp. Zoolog. 292:187–199.
Shine, R. 1988. The evolution of large body size in females: a critique ofDarwin’s “fecundity advantage” model. Am. Nat. 131:124–131.
———. 2003. Reproductive strategies in snakes. Proc. R. Soc. Lond., B, Biol.Sci. 270:995–1004.
Shine, R., and T. Madsen. 1997. Prey abundance and predator reproduction:rats and pythons on a tropical Australian floodplain. Ecology 78:1078–1086.
Slowinski, J. B., and R. Lawson. 2002. Snake phylogeny: evidence fromnuclear and mitochondrial genes. Mol. Phylogenet. Evol. 24:194–202.
Smithson, M. 2003. Confidence intervals. SAGE Publications, London, U.K.Snook, R. R. 2005. Sperm in competition: not playing by the numbers. Trends
Ecol. Evol. 20:46–53.Stockley, P., M. J. G. Gage, G. A. Parker, and A. P. Møller. 1997. Sperm com-
petition in fishes: the evolution of testis size and ejaculate characteristics.Am. Nat. 149:933–954.
Tomkins, J. L., and L. W. Simmons. 2002. Measuring relatie investment: a casestudy of testes investment in species with alternative male reproductivetactics. Anim. Behav. 63:1009–1016.
Tourmente, M., G. Cardozo, M. Bertona, A. Guidobaldi, L. Giojalas, andM. Chiaraviglio. 2006. The ultrastructure of the spermatozoa of Boa
constrictor occidentalis, with considerations on its mating system andsperm competition theories. Acta Zool. 87:25–32.
Tourmente, M., L. Giojalas, and M. Chiaraviglio. 2008. Sperm ultrastructureof Bothrops alternatus and Bothrops diporus (Viperidae, Serpentes),and its possible relation to the reproductive features of the species.Zoomorphology 127:241–248.
Turner, R. M. 2006. Moving to the beat: a review of mammalian sperm motilityregulation. Reprod. Fertil. Dev. 18:25–38.
Vidal, N., and S. B. Hedges. 2002. Higher-level relationships of snakes inferredfrom four nuclear and mitochondrial genes. C. R. Biol. 325:977–985.
Vidal, N., S. G. Kindl, A. Wong, and S. B. Hedges. 2000. Phylogenetic rela-tionships of xenodontine snakes inferred from 12S and 16S ribosomalRNA sequences. Mol. Phylogenet. Evol. 14:389–402.
Zaher, H. 1999. Hemipenial morphology of the South American xenodon-tine snakes: with a proposal for a monophyletic Xenodontinae and areappraisal of colubroid hemipenes. Bulletin of the AMNH 240.
Zug, G. R., L. J. Vitt, and J. P. Caldwell. 2001. Herpetology: an introductorybiology of amphibians and reptiles. Academic Press, San Diego, CA.
Associate Editor: Prof. R. Snook
Supporting InformationThe following supporting information is available for this article:
Table S1. Mean values of corporal and sperm dimensions for the 25 analyzed snake species.
Supporting Information may be found in the online version of this article.
(This link will take you to the article abstract).
Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting information supplied by the
authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
Additional results and discussion can be found in a document at http://www.repository.naturalis.nl/record/289893.
1 2 EVOLUTION 2009