sperm chromatin released by nucleases
TRANSCRIPT

Archives of Andrology: Journal of Reproductive Systems, 54:1–10, 2008Copyright r Informa Healthcare USA, Inc.ISSN: 0148-5016 print/1521-0375 onlineDOI: 10.1080/19396360701876849
Research Article
5Sperm Chromatin Released by Nucleases
In human spermatozoa, 15–20% of histones are retained in the nucleus to
coexist with protamines. Hypothetically, nucleohistone regions of sperm
chromatin mark DNA sequences for distinctive processing during fertilization
10and early embryogenesis. The structural organization and molecular composi-
tion of nucleohistones in human spermatozoa is poorly studied. Here, we
isolate and characterize fractions of sperm chromatin that are solubilized by
endogenous and micrococcal nucleases. Chromatin isolated by either nuclease
have a nucleosomal organization with the periodicity of B195bp (endogenous
15nuclease digest) and B189bp (micrococcal nuclease digest), which is similar to
that of somatic cells. A distinct feature of sperm nucleohistone is its specific
compact supra-nucleosomal organization that was demonstrated by two-
dimensional electrophoresis and by atomic force microscopy. The latter tech-
nique showed compacted fiber arrays composed of globular particles with the
20prevailing diameter of B16nm. A rough estimation indicates that histones may
cover continuous stretches of 450kbp of sperm DNA. This initial character-
ization of sperm chromatin solubilized by nucleases is important for our
understanding of the bipartite structural organization of the paternal genome.
KEYWORDS chromatin, endogenous nucleases, human sperm25
INTRODUCTION
In the nucleus of mammalian sperm, DNA is tightly packed by prota-
mines, forming a protected and inert conformation of chromatin ready for
30fertilization. Recent studies have demonstrated that an elementary structural
unit of the sperm nucleoprotamine contains B50 kb of DNA compacted by
protamines into a toroid with outer diameter of approximately 900 A, a
thickness of 200 A, and a central hole with a diameter of 150 A [Balhorn et al.
1999]. Human spermatozoa are unique among the mammals in that along
35with protamines, a significant amount (15–20%) of histones are retained in
the nucleus [Puwaravutipanich and Panyim 1975; Gusse et al. 1986;
Gatewood et al. 1990]. Accordingly, a small fraction of chromatin organized
into nucleosomes has been detected in human spermatozoa using digestion
with micrococcal nuclease [Banarjee et al. 1995; Zalenskaya et al. 2000].
40This dual organization of the chromatin in the human sperm has evoked
considerable interest. It has been proposed that the distribution of histones
Igor B. Nazarov
The Jones Institute for
Reproductive Medicine, Eastern
Virginia Medical School, 601
Colley Ave., Norfolk VA, 23507
Luda S. Shlyakhtenko and
Yuri L. Lyubchenko
The UNMC Nanoimaging Core
Facility, Department of
Pharmaceutical Sciences
College of Pharmacy, University
of Nebraska Medical Center,
986025 Nebraska Medical
Center, Omaha, NE 68198-6025
Irina A. Zalenskaya and
Andrei O. Zalensky
The Jones Institute for
Reproductive Medicine, Eastern
Virginia Medical School, 601
Colley Ave.,Q1 Norfolk VA, 23507
Abbreviations: AFM: atomic forcemicroscopy; EDTA: ethylenediaminetetraacetic acid; EN: endogenousnuclease; MN: micrococcal nuclease;PAGE: polyacrylamide gelelectrophoresis; PBS: phosphatebuffered saline; PMSF:phenylmethylsulphonylflouride; SDS:sodium dodecyl sulfate; TRIS:trishydroxymethylaminomethane.
Received 26 July 2007; accepted13 October 2007.
Address correspondence to Andrei O.Zalensky, The Jones Institute forReproductive Medicine, EasternVirginia Medical School, 601 ColleyAve., Norfolk VA, 23507. E-mail:[email protected]
1

and protamines along paternal DNA is not random,
and this may provide a potential mechanism for
differential gene expression during fertilization and
45 early embryogenesis [Gatewood et al. 1987; Wykes
and Krawetz 2003; Miller et al. 2005]. This hypothesis
is corroborated by a detailed study of the histone
distribution among the members of the b-globin
gene family in the sperm chromatin, in which
50 histones were found to be associated only with
the genes that are expressed in the embryo
[Gardiner-Garden et al. 1998].
Digestion of hamster sperm chromatin with
DNAse I [Sotolongo et al. 2003] or digestion of
55 mouse, hamster, and human sperm with endogen-
ous nucleases [Pittoggi et al. 1999; Sotolongo et al.
2003; 2005] revealed long stretches of sheltered DNA
40–50 kb long. It has been suggested that the
nuclease resistant regions of sperm chromatin corres-
60 pond to the microscopically observed nucleo-
protamine toroids, while the residual sperm histones
are associated with nuclease sensitive inter-toroid
linkers [reviewed in Ward and Ward 2004].
Major efforts have been directed to the portrayal
65 of limited DNA digests remaining in the sperm
nuclei pellet and characterization of endogenous
nucleases (EN) involved [Shaman et al. 2006].
Molecular and structural characterization of the
soluble chromatin has not been performed, and this
70 is the subject of the current study.
RESULTS
Soluble Chromatin Released byEndogenous Nucleases
Permeabilized human sperm cells were incubated
75 at 371C in the presence of divalent cations
(Caþþ þMgþþ ) for different periods of time, which
resulted in auto-digestion of the nuclear DNA.
Chromatin release in low ionic strength buffer was
not observed before 30min of incubation. Maximum
80 release (1–3% of total DNA) was at 2 h of incubation.
Electrophoretic separation of the liberated chromatin
(fractions S1 and S2, see scheme of experimental
design Fig. 1) under nondenaturing conditions
revealed a smear of chromatin with a low mobility
85 (Fig. 2A). In these conditions, S2 chromatin demon-
strated noticeably higher mobility than S1 chromatin.
Second dimension electrophoresis of S1/S2 chro-
matin in the presence of SDS exposed a nucleo-
some-like ladder of DNA fragments (thin arrows in
90Fig. 2B) emerging from the leading region of the
chromatin smear. We conclude that the chromatin in
S1 and S2 fractions have been cut into nucleosome-
like particles, which under conditions of native
electrophoresis were kept together in a large
95nucleoprotein complex. Hidden double-stranded
DNA cuts become detectable during electrophoresis
in the second dimension due to disruption of DNA-
protein interactions by SDS. Such behavior of sperm
chromatin is dissimilar to that of a fragmented
100chromatin isolated from somatic cells. High mole-
cular weight complex of sperm chromatin estimated
by electrophoretic mobility, may contain about 50
nucleosome-like particles.
In contrast, the trailing component of the chro-
105matin smear in the 1st dimension does not show
periodical DNA fragmentation in the 2nd dimension,
but produces an extended DNA streak (thick arrows
in Fig. 2B). Thus, EN revealed two types of the
soluble sperm chromatin that differ in their structural
110characteristics.
To estimate the periodicity of DNA fragments in
nucleosomal-type chromatin, DNA isolated from S1
and S2 fractions was separated in 1.5% agarose gel
(Fig. 2C). The values for the nucleosomal DNA
115repeat lengths were obtained from the slope of the
regression line in a plot of fragment size versus band
number. 100 bp DNA ladder was used as the
standard. The average from 10 experiments is
FIGURE 1 Experimental scheme of soluble chromatin frag-
ment isolation from human sperm.
I. B. Nazarov et al. 2

195.3� 3.5SD bp (p<0.0001), a value close to the
120 nucleosomal repeat in human somatic cells.
Chromatin protein composition was analyzed
using SDS electrophoresis. Separation of proteins
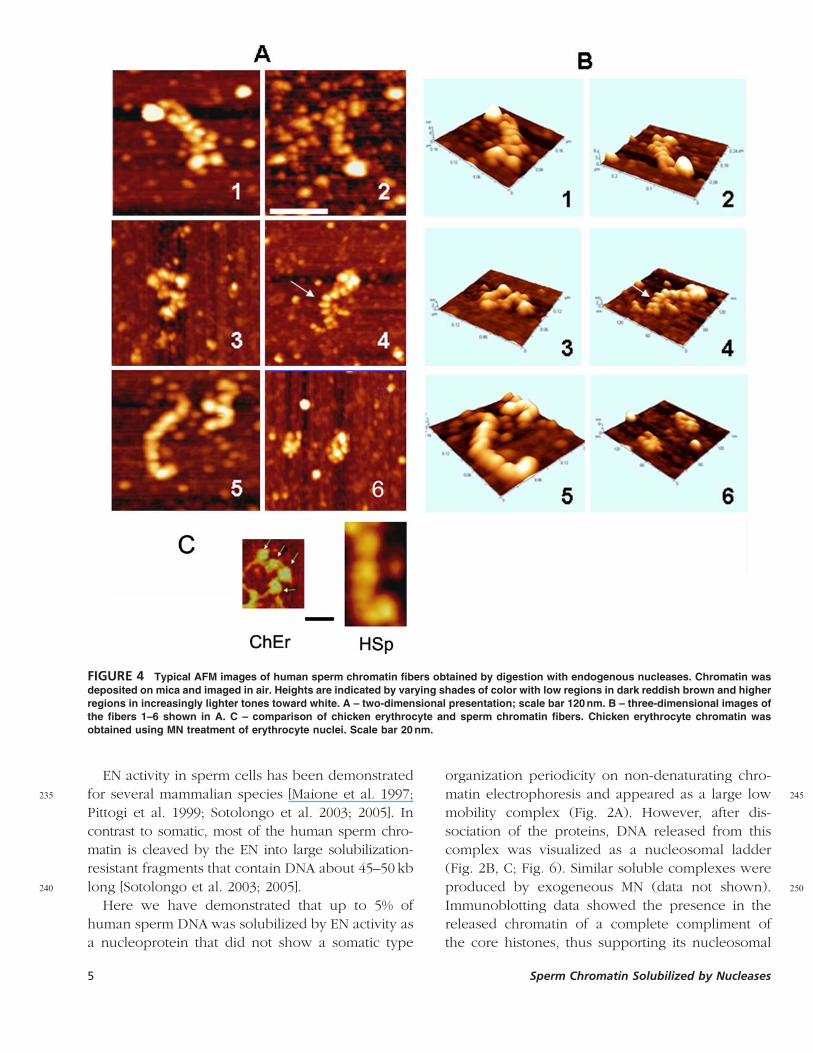
from S1 and S2 chromatin did not reveal noticeable
differences between them, therefore only data for
125 the S2 fraction are provided. Figure 3A shows
proteins of the S2 fraction separated by the 2D
electrophoresis (first dimension: native gel; second:
15% PAGE-SDS gel). This gel demonstrated the
presence of proteins with the range of molecular
130 weights corresponding to histones, shown by a
bracket in Fig. 3A. Histones present in the S2
fraction were confirmed by immunoblotting with
antibodies against core histones H4 and H3 after
one-dimensional SDS-PAGE separation (Fig. 3B).
135 Importantly, chromatin fractions solubilized by EN
were free of protamines, which remained associated
with the nuclear pellet, as was demonstrated by
electrophoresis in acetic acid/urea gel (Fig. 3C).
Imaging of Chromatin Solubilized by140 EN using Atomic Force Microscopy
To acquire more information about structural
organization of the sperm chromatin released by
EN, we performed imaging of soluble sperm
chromatin using atomic force microscopy (AFM).
145Figure 4A demonstrates typical images of the S2
fraction in low ionic strength buffer (10 mM TE, Q21 mM
EDTA), Fig. 4B presents matching three-dimensional
images. AFM revealed compact chromatin fibers of
different length. The fibers are 27� 8 SD nm thick
150(range15–42nm) and composed of single (Fig. 4A,
B-5) or double (Fig. 4A, B-1, 2) rows of spherical
globular particles. A majority of the globes have a
mean diameter of 16� 2nm. The globes are tightly
packed and no linker DNA is visualized in the AFM
155images of sperm chromatin, in contrast to relaxed
reference chromatin of chicken erythrocytes imaged
using the same conditions (Fig. 4C).
Spheres, located at the less condensed areas of the
sperm chromatin (arrows in Fig. 4A, B-4) have dia-
160meters of B8 nm and lower in height. We propose
that these smaller particles are nucleosomes since
they are similar to the B8–10 nm diameter globules
in erythrocyte chromatin (thin arrows in Fig. 4C).
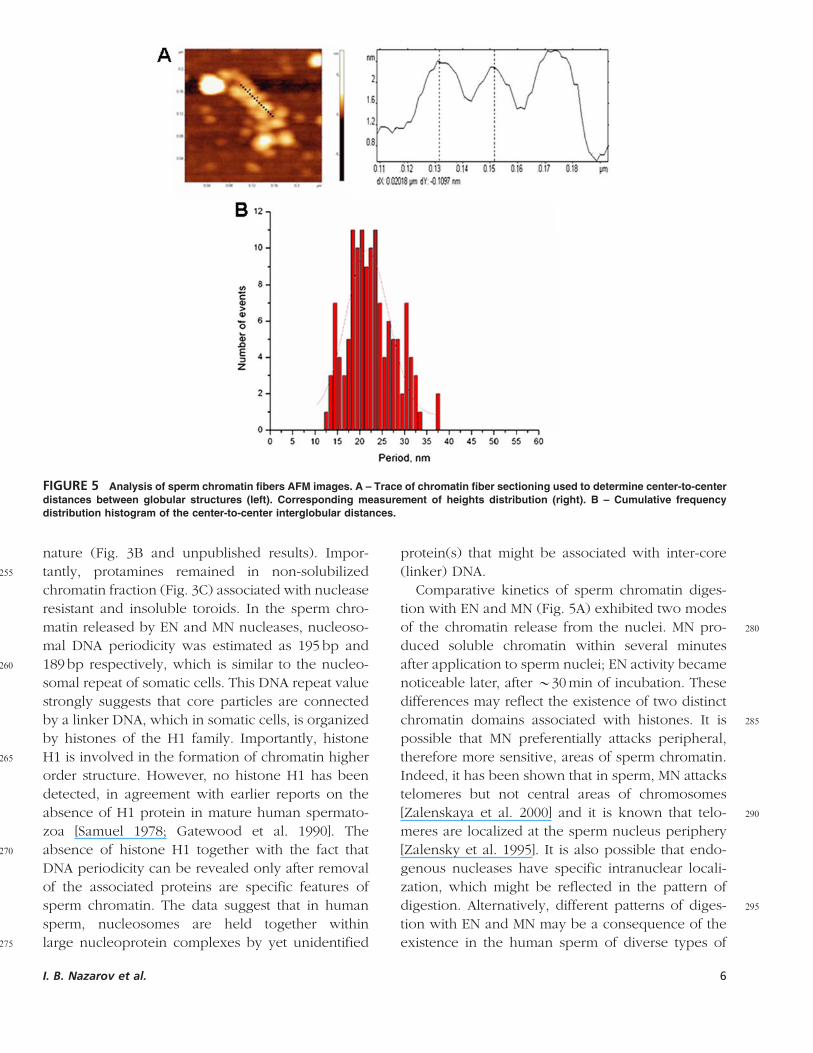
Figure 5A illustrates a cross-section examination
165of the chromatin fragments. Measurements of
center-to-center distance of adjacent particles along
the chromatin fiber path demonstrated a pronounced
periodicity within a fiber. Frequency distribution
histogram of the ‘‘center-to-center’’ distance
170(Fig. 5B) showed a Gaussian distribution with the
FIGURE 2 Characterization of chromatin solubilized by endogenous nucleases. Sperm nuclei were incubated in the presence of 2 mM
CaCl2, 2 mM MgCl2 during 2 h at 371C. A – Electrophoresis of chromatin in native conditions: 0.6% agarose gel on 0.5�TBE buffer;
Ethidium Bromide staining. Molecular weight markers (two left lanes): 100 bp Ladder (Bioline) and l/Hind DNA digest. B – Two-
dimensional electrophoresis. First dimension (top), native gel as in A; second dimension, 1.5% agarose gel. Ethidium Bromide staining.
Thin arrows show fragments of nucleosomal DNA, thick arrows – DNA lacking nucleosomal periodicity. C – Typical electrophoretic
patterns of DNA isolated from S1 and S2 fractions, which were used to determine nucleosomal repeat lengths: 1.5% agarose gel; DNA
marker is 100 bp Ladder (Bioline).
3 Sperm Chromatin Solubilized by Nucleases

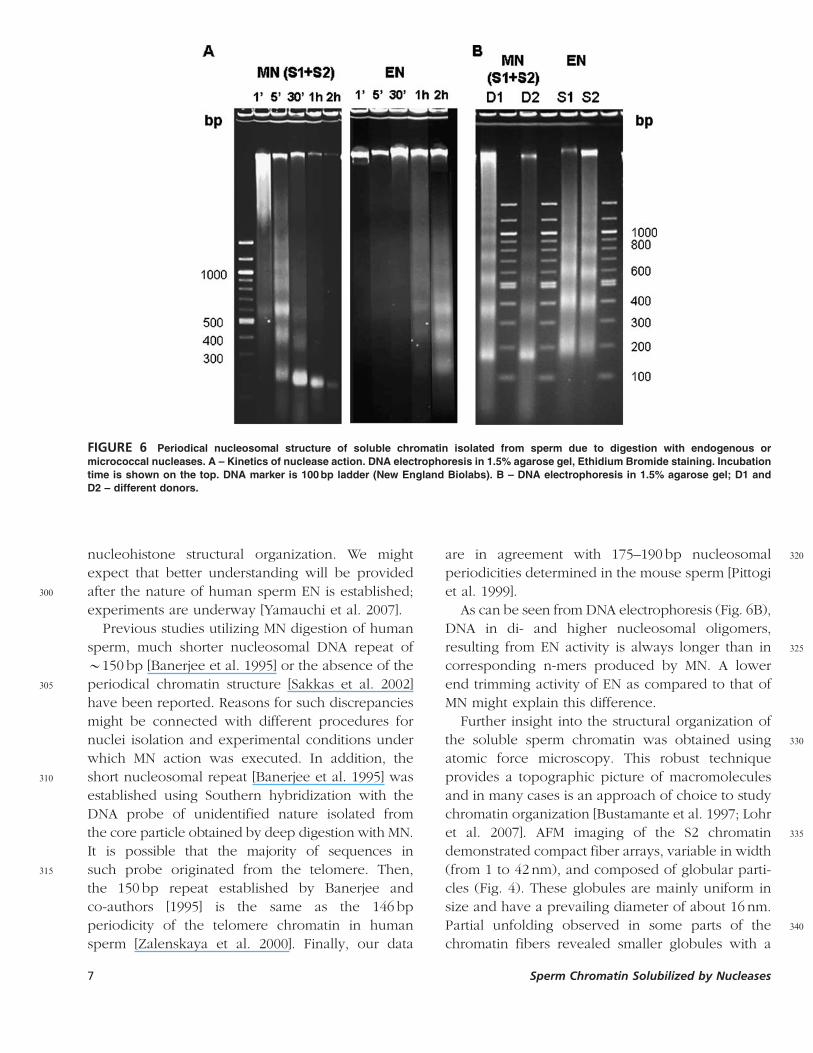
major peak corresponding to 21.7� 0.5 nm and
smaller satellite peaks at 14nm and 31nm.
Sperm Chromatin Accessible toMicrococcal Nuclease
175 We have demonstrated that in human sperm, EN
action became noticeable after an extended lag
period, about 30 min–1 h in our experimental con-
ditions. This is in agreement with the timing of the
nuclease resistant DNA accumulation in sperm
180 reported earlier [Sotolongo et al. 2005]. At the next
step, we were interested in comparing the EN
pattern of digestion with that of micrococcal
nuclease (MN), traditionally used for analysis of
chromatin structure in somatic nuclei. In contrast to
185 EN, MN activity manifested almost immediately.
After the first minute of sperm nuclei incubation
with nuclease, a smear of high molecular weight
DNA was registered, which was quickly processed
into a nucleosomal-like DNA ladder after five
190 minutes (Fig. 6A, left panel). DNA of only mono-
nucleosomal size was present (Fig. 6A) after more
prolonged treatment with MN (30 min–1 h). The
amount of DNA in the soluble chromatin after
treatment with MN was 1–4% of total nuclear DNA
195 and varied between donors (lanes D1 and D2 in Fig.
6B). In a parallel experiment using the same sample
of sperm nuclei, a nucleosome-like ladder generated
by EN appeared much later, after 1 h of incubation
(Fig. 6A, right panel).
200 Nucleosomal DNA repeat lengths calculated from
several independent experiments were 189� 5 bp
and 195� 5 bp for chromatin solubilized by MN and
EN, respectively. While these values of nucleosomal
repeat are very close, it is quite clear from the
205electrophoretic analysis (Fig. 6B) that the mobility of
the equivalent particles (e.g. di- tri- nucleosomes,
etc.) in MN and EN digested chromatin are different.
DNA of the corresponding chromatin fragments
released by digestion with EN was consistently
210longer, which may indicate that EN trims the ends of
nucleosomal fiber to a less extent.
DISCUSSION
In the early 1970s, it was found that in various
types of somatic cells Caþþ/Mgþþ dependent
215endogenous nucleases cleave chromosomal DNA
in a periodical manner: the cleavage sites being
separated by B200 bp [Hewish and Burgoyne 1973].
The same results were obtained using micrococcal
nuclease [reviewed in Felsenfeld 1978]. These
220periodical particles, or nucleosomes, appeared
under electron microscopy as beads on a string.
The nucleosome contains core and linker domains
that are structurally distinct. In the core, DNA of
invariable 146 bp lengths is wrapped around histone
225octamer. Linker DNA is associated with histones
belonging to the H1 family and its length varies
depending on participating histone H1 variant
[reviewed in Woodcock et al. 2006]. Chromatin
fragmented by EN or MN, as well as DNA isolated
230from the fragments, can be resolved by electrophor-
esis and visualized as nucleosomal particles or a
DNA nucleosomal ladder [Albright et al. 1980;
Pennings 1997].
FIGURE 3 Proteins in chromatin solubilized by endogenous nucleases. A – Chromatin proteins separated using two-dimensional
electrophoresis. First dimension – native agarose gel; second dimension – 15% SDS PAGE. Silver staining. B – Immunoblotting of
chromatin proteins separated in 15% SDS PAGE using antibodies against histones H3 and H4. C – Separation of chromatin proteins in
acetic acid/urea PAGE. Coomassie staining.
I. B. Nazarov et al. 4
EN activity in sperm cells has been demonstrated
235 for several mammalian species [Maione et al. 1997;
Pittogi et al. 1999; Sotolongo et al. 2003; 2005]. In
contrast to somatic, most of the human sperm chro-
matin is cleaved by the EN into large solubilization-
resistant fragments that contain DNA about 45–50 kb
240 long [Sotolongo et al. 2003; 2005].
Here we have demonstrated that up to 5% of
human sperm DNA was solubilized by EN activity as
a nucleoprotein that did not show a somatic type
organization periodicity on non-denaturating chro-
245matin electrophoresis and appeared as a large low
mobility complex (Fig. 2A). However, after dis-
sociation of the proteins, DNA released from this
complex was visualized as a nucleosomal ladder
(Fig. 2B, C; Fig. 6). Similar soluble complexes were
250produced by exogeneous MN (data not shown).
Immunoblotting data showed the presence in the
released chromatin of a complete compliment of
the core histones, thus supporting its nucleosomal
FIGURE 4 Typical AFM images of human sperm chromatin fibers obtained by digestion with endogenous nucleases. Chromatin was
deposited on mica and imaged in air. Heights are indicated by varying shades of color with low regions in dark reddish brown and higher
regions in increasingly lighter tones toward white. A – two-dimensional presentation; scale bar 120 nm. B – three-dimensional images of
the fibers 1–6 shown in A. C – comparison of chicken erythrocyte and sperm chromatin fibers. Chicken erythrocyte chromatin was
obtained using MN treatment of erythrocyte nuclei. Scale bar 20 nm.
5 Sperm Chromatin Solubilized by Nucleases
nature (Fig. 3B and unpublished results). Impor-
255 tantly, protamines remained in non-solubilized
chromatin fraction (Fig. 3C) associated with nuclease
resistant and insoluble toroids. In the sperm chro-
matin released by EN and MN nucleases, nucleoso-
mal DNA periodicity was estimated as 195 bp and
260 189 bp respectively, which is similar to the nucleo-
somal repeat of somatic cells. This DNA repeat value
strongly suggests that core particles are connected
by a linker DNA, which in somatic cells, is organized
by histones of the H1 family. Importantly, histone
265 H1 is involved in the formation of chromatin higher
order structure. However, no histone H1 has been
detected, in agreement with earlier reports on the
absence of H1 protein in mature human spermato-
zoa [Samuel 1978; Gatewood et al. 1990]. The
270 absence of histone H1 together with the fact that
DNA periodicity can be revealed only after removal
of the associated proteins are specific features of
sperm chromatin. The data suggest that in human
sperm, nucleosomes are held together within
275 large nucleoprotein complexes by yet unidentified
protein(s) that might be associated with inter-core
(linker) DNA.
Comparative kinetics of sperm chromatin diges-
tion with EN and MN (Fig. 5A) exhibited two modes
280of the chromatin release from the nuclei. MN pro-
duced soluble chromatin within several minutes
after application to sperm nuclei; EN activity became
noticeable later, after B30 min of incubation. These
differences may reflect the existence of two distinct
285chromatin domains associated with histones. It is
possible that MN preferentially attacks peripheral,
therefore more sensitive, areas of sperm chromatin.
Indeed, it has been shown that in sperm, MN attacks
telomeres but not central areas of chromosomes
290[Zalenskaya et al. 2000] and it is known that telo-
meres are localized at the sperm nucleus periphery
[Zalensky et al. 1995]. It is also possible that endo-
genous nucleases have specific intranuclear locali-
zation, which might be reflected in the pattern of
295digestion. Alternatively, different patterns of diges-
tion with EN and MN may be a consequence of the
existence in the human sperm of diverse types of
FIGURE 5 Analysis of sperm chromatin fibers AFM images. A – Trace of chromatin fiber sectioning used to determine center-to-center
distances between globular structures (left). Corresponding measurement of heights distribution (right). B – Cumulative frequency
distribution histogram of the center-to-center interglobular distances.
I. B. Nazarov et al. 6
nucleohistone structural organization. We might
expect that better understanding will be provided
300 after the nature of human sperm EN is established;
experiments are underway [Yamauchi et al. 2007].
Previous studies utilizing MN digestion of human
sperm, much shorter nucleosomal DNA repeat of
B150 bp [Banerjee et al. 1995] or the absence of the
305 periodical chromatin structure [Sakkas et al. 2002]
have been reported. Reasons for such discrepancies
might be connected with different procedures for
nuclei isolation and experimental conditions under
which MN action was executed. In addition, the
310 short nucleosomal repeat [Banerjee et al. 1995] was
established using Southern hybridization with the
DNA probe of unidentified nature isolated from
the core particle obtained by deep digestion with MN.
It is possible that the majority of sequences in
315 such probe originated from the telomere. Then,
the 150 bp repeat established by Banerjee and
co-authors [1995] is the same as the 146 bp
periodicity of the telomere chromatin in human
sperm [Zalenskaya et al. 2000]. Finally, our data
320are in agreement with 175–190 bp nucleosomal
periodicities determined in the mouse sperm [Pittogi
et al. 1999].
As can be seen from DNA electrophoresis (Fig. 6B),
DNA in di- and higher nucleosomal oligomers,
325resulting from EN activity is always longer than in
corresponding n-mers produced by MN. A lower
end trimming activity of EN as compared to that of
MN might explain this difference.
Further insight into the structural organization of
330the soluble sperm chromatin was obtained using
atomic force microscopy. This robust technique
provides a topographic picture of macromolecules
and in many cases is an approach of choice to study
chromatin organization [Bustamante et al. 1997; Lohr
335et al. 2007]. AFM imaging of the S2 chromatin
demonstrated compact fiber arrays, variable in width
(from 1 to 42 nm), and composed of globular parti-
cles (Fig. 4). These globules are mainly uniform in
size and have a prevailing diameter of about 16 nm.
340Partial unfolding observed in some parts of the
chromatin fibers revealed smaller globules with a
FIGURE 6 Periodical nucleosomal structure of soluble chromatin isolated from sperm due to digestion with endogenous or
micrococcal nucleases. A – Kinetics of nuclease action. DNA electrophoresis in 1.5% agarose gel, Ethidium Bromide staining. Incubation
time is shown on the top. DNA marker is 100 bp ladder (New England Biolabs). B – DNA electrophoresis in 1.5% agarose gel; D1 and
D2 – different donors.
7 Sperm Chromatin Solubilized by Nucleases

diameter B8 nm. These globules most likely corre-
spond to the core nucleosome particles. We propose
that large globes, such as presented in Fig. 4A, B-5,
345 are composed of 4–6 nucleosomes as visualized in
the areas where chromatin is unfolded (Fig. 4A, B-4).
In the compact fibrils of sperm chromatin, a
center-to-center distance of 21.7� 0.5 nm was
observed for adjacent globules (Fig. 5). In the fibers
350 of chicken erythrocyte chromatin, particle center-to-
center distance is 31.4 nm, of which B22 nm
belongs to internucleosomal linker DNA and
B10 nm is the diameter of the nucleosome core
particle [D’Erme et al. 2001]. Thus, histone contain-
355 ing chromatin of human sperm nuclei is organized
into much more compact structures as compared to
chromatin of somatic cells.
Chromatin fragments observed in AFM experi-
ments are heterogeneous in size and estimates sug-
360 gest that more than 50 nucleosomes may be
involved. This covers over 12 kbp of DNA, which
correlates well with the estimation of the size of the
EN-released complexes by electrophoretic mobility
on DNP electrophoresis (Fig. 2).
365 It is not known if the endogenous and exoge-
neous nucleases target the same areas of chromatin,
and the important question to be answered is
how the nuclease sensitive (nucleohistone) areas
are distributed along the human genome in
370 spermatozoa. Research using microarray profiling of
histone/protamine distribution in spermatozoa is in
progress (D. Miller, University of Leeds, personal
communication) and experiments using FISH
localization of the histone-associated sequences
375 are underway in our laboratory. It is anticipated that
the combination of these approaches will help to
define more accurately molecular organization of
DNA in spermatozoa and shed some light on its
significance for fertilization and early embryonic
380 development.
MATERIALS AND METHODS
Biological Material
Semen samples were collected from healthy
donors after at least 2 days of abstinence. Written
385 informed consent was obtained from all participants.
The samples were collected by masturbation and
allowed to liquefy at room temperature. The
samples were kept frozen (at �801C) until processed
for analyses.
390Isolation of Sperm Cells
All procedures were performed at 41C. Frozen
semen samples were thawed, diluted with ice cold
phosphate buffered saline (PBS) containing 0.5 mM
phenylmethylsulphonylflouride (PMSF), vortexed,
395filtered through 2 layers of Miracloth, and pelleted
by centrifugation (850� g for 10 min). The pellet
was washed with PBS two more times. Presence of
contaminating cells was evaluated by microscopic
analysis. Samples, which had more than 1% of
400somatic or immature germ cells, were excluded from
experiments.
Isolation of Chromatin Solubilizedby Nucleases
The general experimental scheme is outlined in
405Fig. 1. Washed sperm cells were resuspended in
PBS/PMSF containing 0.5% Triton X-100, incubated
on ice for 10 min, and washed twice with PBS/PMSF.
Partially demembranized cells were resuspended in
the reaction buffer comprised of 10 mM Tris-HCl, pH
4108.0, 2 mM CaCl2, 2 mM MgCl2, 10 mM dithiothreitol,
protease inhibitor cocktail (Roche) at a DNA
concentration of 3–5 mg/ml. Cells were incubated
at 371C for the times indicated, aliquots were
immediately cooled on ice, then centrifuged for
4155 min at 500 g, 401C. Supernatant (S1) was collected
and EDTA was added to 5 mM. The pellet was
resuspended in 2 volumes of TED buffer (10 mM
Tris-HCl, Q3pH 8.0, 5 mM EDTA, 10 mM dithiothreitol)
and extracted on ice for 0.5 h. The sample was
420centrifuged for 5 min at 500 g at 401C and super-
natant (S2) was collected.
In the micrococcal nuclease (MN) experiments,
20 U of the enzyme was added per mg of DNA,
incubation was at 401C. The reaction was stopped by
425the addition of EDTA, digested nuclei were cen-
trifuged, and supernatant (S1þ S2) was collected.
DNA concentrations in cell suspensions and S1/S2
extracts were determined by spectrometry measur-
ing OD at 260 nm of the alkali-denatured samples
430and by fluorimetry using emission at 458 nm in the
presence of Hoechst 33258.
I. B. Nazarov et al. 8

One-dimensional DNAElectrophoresis and Determination
of Nucleosomal DNA Repeat435 Length
DNA was isolated by a standard phenol-
chloroform extraction. In brief, samples were treated
at 371C with 100mg/ml proteinase K in the presence of
0.5% SDS overnight, followed by phenol/chloroform
440 extraction and ethanol precipitation. DNA was
dissolved in 10 mM Tris-HCl, pH 7.5, 1 mM EDTA
(TE buffer) and subjected to electrophoreses in 1.5%
agarose gel in 1�TAE buffer (40 mM Tris-HClQ4 , pH
8.0, 20 mM acetic acid, 1 mM EDTA). DNA was
445 stained with ethidium bromide; DNA fluorescence
was recorded using Kodak Image station 440. Sizes
of DNA fragments were determined from calibration
curve generated using 100 bp DNA ladder. Calcula-
tion of nucleosomal periodicity was described in
450 Results.
Protein Electrophoresis andWestern-blotting
Proteins were analyzed using two electrophoretic
systems: 15% SDS-PAGE according to Laemmli [1970]
455 or 15% acetic acid/urea PAGE according to a
modified method of Panyim and Chalkley [Hurley
1977]. The latter allows visualizing protamines that
precipitate in the presence of SDS and therefore
cannot be resolved in SDS-PAGE. Proteins were
460 stained with Coomassie R-250. For identification of
histones, the proteins separated in 15% SDS-PAGE
were transferred to a PVDF membrane. Immunode-
tection of histone fractions was performed using
polyclonal monospecific antibodies as described
465 [Zalensky et al. 2002].
Native Electrophoresis of SolubleChromatin
Native electrophoresis of chromatin was per-
formed using 0.6% agarose gel on 0.5�TBE bufferQ5 .
470 Samples were prepared by adding 6�DNA loading
buffer to fractions of soluble chromatin. The gel was
run at 6 V per cm of length for 3 h. DNA was stained
with ethidium bromide and image acquisition was
performed as described above.
475Two-dimensional Electrophoresis
After electrophoresis, agarose strips were cut out,
incubated in 1X TAE containing 1% SDS during
15 min at room temperature, and were placed on the
start of the 1.5% agarose/1X TAE gel. Electrophoresis
480was performed at 8 V/cm during 1–2 h, and DNA
was stained with ethidium bromide.
Protein analysis used 15% SDS PAGE for separa-
tion in the second dimension. Proteins were silver
stained using SilverQuest staining kit (Invitrogen
485Inc). Images of DNA or proteins were acquired using
Kodak Image Station 440.
Atomic Force Microscopy
Samples were prepared using mica surface (APS
mica), modified with 1-(3-aminopropyl) silatrane as
490described earlier [Shlyakhtenko et al. 2003]. The
chromatin samples were diluted 10 to 20 fold in TE
and 5 mL of the sample was deposited on APS mica.
After 2 min, the sample was rinsed with deionized
water and dried with argon. Images were acquired
495in air using a MultiMode SPM NanoScope IV system
(Veeco/Digital Instruments, Santa Barbara, CA)
operating in tapping mode. Tapping Mode Etched
Silicon Probes (TESP; Veeco/Digital Instruments,
Inc.) with a spring constant of about 42 N/m and a
500resonant frequency between 270–320 kHz were
used. Image processing and cross-section measure-
ments were performed using Femtoscan (Advanced
Technologies Center, Moscow, Russia).
ACKNOWLEDGEMENTS
505This work has been supported by an EVMS
Institutional Grant and in part by National Institutes
of Health Grant HD-042748 to A. O. Z. We thank
Dr. S. Gitlin for proofreading the manuscript.
REFERENCES510Albright, S. C., Wiseman, J. M., Lange, R. A. and Garrard, W. T. (1980)
Subunit structures of different electrophoretic forms of nucleosomes.J Biol Chem 255:3673–3684.
Balhorn, R., Cosman, M., Thornton, K., Krishnan, V. V., Corzett, M.,Bench, G., Kramer, C., Lee, J., Hud, N. V., Allen, M., et al. (1999) In:
515The Q6Male Gamete: From Basic Knowledge to Clinical Applications,Gagnon, C., (Ed.), St. Louis: Cache River Press; pp 55–70.
Banerjee, S., Smallwood, A. and Hulten, M. (1995) ATP-dependentreorganization of human sperm nuclear chromatin. J Cell Sci108:755–765.
9 Sperm Chromatin Solubilized by Nucleases

520 Bustamante, C., Zuccheri, G., Leuba, S. H., Yang, G. and Samori, B.(1997) Visualization and analysis of chromatin by scanning forcemicroscopy. Methods 12:73–83.
D’Erme, M., Yang, G., Sheagly, E., Palitti, F. and Bustamante, C. (2001)Effect of poly(ADP-ribosyl)ation and Mg2þ ions on chromatin
525 structure revealed by scanning force microscopy. Biochemistry40:10947–10955.
Felsenfeld, G. (1978) Chromatin. Nature 271:115–122.Gardiner-Garden, M., Ballesteros, M., Gordon, M. and Tam, P. P. (1998)
Histone- and protamine-DNA association: conservation of different530 patterns within the beta-globin domain in human sperm. Mol Cell
Biol 18:3350–3356.Gatewood, J. M., Cook, G. R., Balhorn, R., Bradbury, E. M. and Schmid,
C. W. (1987) Sequence-specific packaging of DNA in human spermchromatin. Science 236:962–964.
535 Gatewood, J. M., Cook, G. R., Balhorn, R., Schmid, C. W. and Bradbury,E. M. (1990) Isolation of four core histones from human spermchromatin representing a minor subset of somatic histones. J BiolChem 265:20662–20666.
Gusse, M., Sautiere, P., Belaiche, D., Martinage, A., Roux, C., Dadoune, J. P.540 and Chevaillier, P. (1986) Purification and characterization of
nuclear basic proteins of human sperm. Biochim Biophys Acta884:124–134.
Hewish, D. R. and Burgoyne, L. A. (1973) Chromatin sub-structure. Thedigestion of chromatin DNA at regularly spaced sites by a nuclear
545 deoxyribonuclease. Biochem Biophys Res Commun 52:504–510.Hurley, C. K. (1977) Electrophoresis of histones: a modified Panyim and
Chalkley system for slab gels. Anal Biochem 80:624–626.Laemmli, U. K. (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:680–685.550 Lohr, D., Bash, R., Wang, H., Yodh, J. and Lindsay, S. (2007) Using atomic
force microscopy to study chromatin structure and nucleosomeremodeling. Methods 41:333–341.
Maione, B., Pittoggi, C., Achene, L., Lorenzini, R. and Spadafora, C.(1997) Activation of endogenous nucleases in mature sperm cells
555 upon interaction with exogenous DNA. DNA Cell Biol 16:1087–1097.Miller, D., Ostermeier, G. C. and Krawetz, S. A. (2005) The controversy,
potential and roles of spermatozoal RNA. Trends Mol Med 11:156–163Pennings, S. (1997) Nucleoprotein gel electrophoresis for the analysis of
nucleosomes and their positioning and mobility on DNA. Methods560 12:20–27.
Pittoggi, C., Renzi, L., Zaccagnini, G., Cimini, D., Degrassi, F., Giordano, R.,Magnano, A. R., Lorenzini, R., Lavia, P. and Spadafora, C. (1999) Afraction of mouse sperm chromatin is organized in nucleosomalhypersensitive domains enriched in retroposon DNA. J Cell Sci
565 112:3537–3548.
Puwaravutipanich, T. and Panyim, S. (1975) The nuclear basic proteins ofhuman testes and ejaculated spermatozoa. Exp Cell Res 90:153–158.
Richmond, T. J., Finch, J. T., Rushton, B., Rhodes, D. and Klug, A. (1984)Structure of the nucleosome core particle at 7 A resolution Q7. Nature
570311:532–537.Sakkas, D., Moffatt, O., Manicardi, G. C., Mariethoz, E., Tarozzi, N. and
Bizzaro, D. (2002) Nature of DNA damage in ejaculated humanspermatozoa and the possible involvement of apoptosis. Biol Reprod66:1061–1067.
575Samuel, T. (1978) Differentiation between antibodies to protamines andsomatic nuclear antigens by means of a comparative fluorescencestudy on swollen nuclei of spermatozoa and somatic cells. Clin ExpImmunol 32:290–298.
Shaman, J. A., Prisztoka, R. and Ward, W. S. (2006) Topoisomerase IIB580and an extracellular nuclease interact to digest sperm DNA in an
apoptotic-like manner. Biol Reprod 75:741–748.Shlyakhtenko, L. S., Gall, A. A., Filonov, A., Cerovac, Z., Lushnikov, A.
and Lyubchenko, Y. L. (2003) Silatrane-based surface chemistry forimmobilization of DNA, protein-DNA complexes and other biological
585materials. Ultramicroscopy 97:279–287.Sotolongo, B., Huang, T. T., Isenberger, E. and Ward, W. S. (2005) An
endogenous nuclease in hamster, mouse, and human spermatozoacleaves DNA into loop-sized fragments. J Androl 26:272–280.
Sotolongo, B., Lino, E. and Ward, W. S. (2003) Ability of590hamster spermatozoa to digest their own DNA. Biol Reprod
69:2029–2035.Ward, M. A. and Ward, W. S. (2004) A model for the function of sperm
DNA degradation. Reprod Fertil Dev 16:547–554.Woodcock, C. L., Skoultchi, A. I. and Fan, Y. (2006) Role of linker histone
595in chromatin structure and function: H1 stoichiometry and nucleo-some repeat length. Chromosome Res 14:17–25.
Wykes, S. M. and Krawetz, S. A. (2003) The structural organization ofsperm chromatin. J Biol Chem 278:29471–29477.
Yamauchi, Y., Shaman, J. A. and Ward, W. S. (2007) Topoisomerase600II-mediated breaks in spermatozoa cause the specific degradation of
paternal DNA in fertilized oocytes. Biol Reprod 76:666–672.Zalenskaya, I. A., Bradbury, E. M. and Zalensky, A. O. (2000) Chromatin
structure of telomere domain in human sperm. Biochem Biophys ResCommun 279:213–218.
605Zalensky, A. O., Allen, M. J., Kobayashi, A., Zalenskaya, I. A., Balhorn, R.and Bradbury, E. M. (1995) Well-defined genome architecture in thehuman sperm nucleus. Chromosoma 103:577–590.
Zalensky, A. O., Siino, J., Gineitis, A., Tomilin, N. V., Zalenskaya, I. A.,Yau, P. and Bradbury, E. M. (2002) Human testis/sperm specific
610histone H2B (hTSH2B). Molecular cloning and characterization. J BiolChem 8:43474–43480.
I. B. Nazarov et al. 10

QUERY FORM
UAAN
Manuscript ID [Art. Id: 287853]
Author
Editor
Publisher
Journal: UAAN
Disk usage:-
Disk not provided
Disk used
Disk not used
Disk is corrupt
Unknown file format
Virus found
Additional work done:
AUTHOR: The following queries have arisen during the editing of your manuscript. Please answer queries by making
the requisite corrections at the appropriate positions in the text.
Query No. Nature of Query Author’s Response
Q1 please indicate which author(s) are affiliated with the following affiliation.
Q2 please provide pH.
Q3 confirm the following pH buffer.
Q4 confirm pH.
Q5 define and include pH.
Q6 chapter/paper title?
Q7 plese confirm is referenced in body.