bulky dna adducts in human sperm: relationship with fertility, semen quality, smoking, and...
TRANSCRIPT

Mutation Research 537 (2003) 53–65
Bulky DNA adducts in human sperm: relationship with fertility,semen quality, smoking, and environmental factors
Stanislaw Horaka, Joanna Polanskab, Piotr Widlakc,∗a First Chair and Clinical Department of Obstetrics and Gynecology, Silesian Medical Academy, Bytom, Poland
b Department of Automatic Control, Institute of Automation, Silesian Technical University, Gliwice, Polandc Department of Experimental and Clinical Radiobiology, Center of Oncology, Gliwice 44 100, Poland
Received 30 July 2002; received in revised form 20 February 2003; accepted 21 February 2003
Abstract
The integrity of DNA of spermatogenic cells can be affected by endogenous and exogenous genotoxic factors. ResultingDNA damage in spermatozoa may significantly contribute to impaired fertility. Here, the32P-postlabeling method was usedto analyze the levels of bulky DNA adducts in sperm cells in a group of 179 males, either healthy donors or patients withan impaired fertility. When all donors were analyzed, the levels of bulky DNA adducts was 1.2-fold higher in smokersthan in non-smokers, but the difference was not statistically significant (P = 0.054). However, a statistically significantdifference existed between current smokers and never smokers among the healthy individuals (1.7-fold increase,P = 0.008).No correlation between alcohol or coffee consumption and sperm DNA adducts was found. The levels of DNA adducts insperm seemed to be unaffected by environmental and occupational factors. On the other hand, groups of healthy personsand patients with male-factor infertility differed significantly with respect to the level of bulky DNA adducts (P = 0.012).A significant negative correlation between DNA adducts and sperm concentration or sperm motility existed among patientswith an impaired fertility (n = 93;P < 0.029,rS = −0.225). These results suggest that DNA adducts in sperm cells can beapplied as potential biomarkers in studies of human infertility.© 2003 Elsevier Science B.V. All rights reserved.
Keywords: Adducts; DNA damage; Humans;32P-postlabeling; Sperm
1. Introduction
According to estimations of the World Health Orga-nization, about 8% of couples worldwide (10–15% inindustrialized nations) experience some form of infer-tility. The incidence of infertility is equal among malesand females[1]. Infertility can be caused by manydifferent factors (e.g. abnormal oocyte and/or spermproduction, tubal defects, or endometriosis); however,
∗ Corresponding author. Tel.:+48-32-278-9672;fax: +48-32-231-3512.E-mail address: [email protected] (P. Widlak).
the etiology of a large proportion of cases remains ob-scure. Although controversies exist in the field, severalstudies have reported a progressive decline in spermquality and fertility over the last few decades, and ithas been speculated that this was a combined effect ofincreased stress and environmental pollution[2]. Ex-posure to several environmental, occupational, and be-havioral (e.g. smoking) factors, of either a chemical orphysical nature, has been reported as a risk factor formale infertility. These factors include heat, ionizingand non-ionizing electromagnetic waves, heavy met-als, pesticides, organic solvents, and aromatic com-pounds. They can affect the male reproductive system
1383-5718/03/$ – see front matter © 2003 Elsevier Science B.V. All rights reserved.doi:10.1016/S1383-5718(03)00051-2

54 S. Horak et al. / Mutation Research 537 (2003) 53–65
either directly, causing decreased or altered sperm pro-duction in the testis, and/or indirectly through the en-docrine system causing hormonal imbalance. Severalinfertility factors are known or putative mutagens andcarcinogens, and induction of mutagenic events canbe involved in the etiology of some infertility cases[3–5].
Human semen parameters, including volume, spermconcentration, motility and morphology, are valuableand generally accepted indicators of exposure to oc-cupational and environmental factors affecting malefertility. Cigarette smoking is among the best studiedfactors affecting reproduction in humans. The major-ity of the data show that smoking is associated with thereduction in sperm concentration and motility, and thisassociation is stronger in healthy men than in patientssuffering from infertility. Despite a modest reductionin sperm quality, studies have not shown a reductionin fertility associated with paternal smoking (reviewedin ref. [6]). It is well recognized that cigarette smokecontains numerous known or putative mutagens andcarcinogens (e.g. nitrosoamines or polycyclic aro-matic hydrocarbons—PAHs) that can induce DNAdamage[7]. Some evidence suggests that paternalsmoking can be associated with congenital anoma-lies and childhood cancers in offspring as well asalterations in male hormone levels. It has been postu-lated that genotoxic agents might induce mutations insperm cells, which are then passed on to the offspringaffecting their development and the occurrence of can-cer [6]. It has been shown that active DNA-adductingmetabolites of chemical carcinogens could be trans-ported in the serum to different organs and tissues[8].Because of the existence of a blood–testis barrier, themajority of developing germ cells are not in directcontact with the circulation. However, detection ofnicotine and its metabolites in the semen of smokers[9] suggests that DNA-adducting metabolites mightpass through this barrier and react with germ cells aswell. It has been shown that DNA repair induced byseveral chemicals and radiation occurs early duringspermatogenesis (spermatocytes and early spermatids)but not in mature spermatids and spermatozoa[10],raising the possibility of accumulation of non-repairedDNA damage during spermiogenesis.
Covalent modifications of nucleotides, termed DNAadducts, are widely accepted as biomarkers of ex-posure to chemical mutagens and carcinogens in
molecular epidemiology studies[11]. 32P-postlabelingis a method of choice for analyzing DNA adductsin humans exposed to complex mixtures like to-bacco smoke, air pollutants, engine exhausts, or cokeoven emissions. Numerous studies have shown ele-vated levels of DNA adducts in individuals exposedto behavior-, occupation- and environment-relatedgenotoxic agents, in either surrogate cells (e.g. leuko-cytes, the most widely used) or target tissues[12,13].However, only limited data concerning DNA adductsin human spermatogenic cells and spermatozoa areavailable at the moment. The level of 8-hydroxydeo-xyguanosine, a major product of oxidative DNAdamage, was 1.6-fold higher in sperm of smokerscompared with non-smokers, when determined in agroup of 60 healthy volunteers[14]. The immunocy-tochemical analysis of benzo(a)pyrene diol epoxide(BPDE)–DNA adducts in sperm of 23 patients of aninfertility clinic revealed a higher level of DNA dam-age (1.9-fold) in smokers versus non-smokers[9]. Onthe other hand, no significant difference in the level ofDNA breaks in spermatozoa was found between smok-ers and non-smokers in a group of 97 healthy individ-uals [15]. The only analysis of bulky DNA adductsin human sperm determined by the32P-postlabelingtechnique, in 36 healthy volunteers, revealed no dif-ferences between smokers and non-smokers. How-ever, the adduct levels detected in this study were atthe detection limit of the method used[16].
The aim of the present study was to reevaluate thefeasibility of using the32P-postlabeling method to an-alyze bulky DNA adducts in human sperm cells as ameasure of DNA lesions, and to relate these findingsto possible defects of human fertility. Analysis of a rel-atively large group of individuals with different expo-sures to potent genotoxic factors has the possibility ofestablishing a correlation between DNA adducts andquality of sperm, should one exist. We demonstratehere that this appears to be indeed the case.
2. Materials and methods
2.1. Study participants
Semen samples were collected from 179 volun-teers aged between 22 and 47 years (median= 35,mean= 35.15). Each individual signed an informed

S. Horak et al. / Mutation Research 537 (2003) 53–65 55
consent form approved by the Ethics Committeeof the Silesian Medical Academy. Twenty subjectswere healthy sperm donors with proven fertility and159 subjects were patients of our infertility clinic.Male-factor infertility was suspected in 94 of them,based on case records and semen analyses. Sixty-fivepatients, who were not suspected of male-factor in-fertility were joined to sperm donors as the healthygroup. These patients do not differ significantly fromsperm donors with respect to parameters of spermquality. Information on smoking and drinking habits,occupational, and demographic data were obtainedthrough a questionnaire. The questionnaire and mor-phological data were not complete in a few cases and,therefore, the number of subjects was less than 179in several statistical analyses.
2.2. Semen collection and examination
Semen samples were obtained by masturbation af-ter 2–7 days of abstinence. Semen analysis (i.e. vol-ume, sperm concentration, motile sperm percentage,amount of leukocytes in semen) was performed ac-cording to standard methods[17]. Samples were di-vided in aliquots and stored in liquid nitrogen. Inthe majority of semen samples, some white bloodcells could be detected, whose relative proportion tospermatozoa ranged from 0 to 250% (median 2.3%,mean 8.91%). The amounts of leukocytes in semenwere significantly higher (P < 0.00001) in individu-als with predicted male-factor infertility (median 5%,mean 14.66%) than in semen of healthy subjects (me-dian 1.1%, mean 2.41%). There was also a strongnegative correlation between the amounts of leuko-cytes in semen and the concentration of spermatozoa(rS = −0.6377,P < 0.0001) or their motility (rS =−0.516,P < 0.0001). Granulocytes were identified asperoxidase-positive cells[17] and were the major com-ponent of leukocytes detected in semen (about 90%,on the average).
2.3. DNA isolation
Sperm cells washed in PBS were incubated withproteinase K (Sigma; final concentration 0.2 mg/ml)in the presence of 0.5% SDS and 10 mM EDTA for12–16 h at 37◦C (all reagents from Sigma). DNAwas then purified by 2–3 repeated extractions with
phenol/chloroform and recovered by ethanol precipi-tation. Precipitated DNA was dissolved in TE buffer(10 mM Tris, 1 mM EDTA; pH 8.0), incubated withRNaseA to remove traces of RNA, then re-purifiedby phenol/chloroform extraction and ethanol precipi-tation. DNA concentration was measured spectropho-tometrically and its integrity examined by agarose gelelectrophoresis. In all samples analyzed for adducts,the majority of DNA (>95%) was of high molecularweight (>25 kb).
2.4. Adduct analysis
Bulky DNA adducts were assayed according to the32P-postlabeling method[18] with some modifica-tions as described in detail elsewhere[19]. Briefly,10�g of DNA was digested to mononucleotides withmicrococcal nuclease (for 3 h at 37◦C) followed bydigestion with spleen phosphodiesterase (for an ad-ditional 3 h) and then non-modified mononucleotideswere removed by digestion with nuclease P1 (foran additional 1 h; all enzymes from Sigma). Mod-ified nucleotides were radiolabeled for 30 min at37◦C in a 3-�l reaction mixture containing 3 pmol32P-�-ATP (ICN; 3000 Ci/mmol) and 3 units of T4polynucleotide kinase (USB), and then excess ATPwas removed with apyrase. The labeled nucleotideswere applied to pre-washed PEI-cellulose TLC plates(Macherey-Nagel). The chromatography was carriedout in 1 M sodium phosphate, pH 6.0, for 6 h onto apaper wick (D1). Modified nucleotides were separatedby chromatography in 8.5 M urea, 3.5 M lithium for-mate, pH 3.5 (D2, direction opposite to D1) and then8.5 M urea, 0.8 M LiCl2, 0.5 M Tris, pH 8.0 (D3, di-rection turned 90◦ to D2). D2 and D3 were developedonto paper wicks, and the time of chromatographyrequired for optimal and reproducible resolution ofadducts was determined experimentally for each batchof TCL plates. The adduct spots were visualized byautoradiography at−70◦C for 2–4 days. Radioactiv-ity of excised areas of TLC plates (adduct spots andplate background) was counted by Cerenkov radiationto calculate the level of DNA adducts. Calf thymusDNA modified by a microsomal-activated mixtureof 10 PAHs (PAH–DNA;[19]) and deoxyguanosinemodified by BPDE (dG–BPDE) were used as stan-dards with known levels of adducts. The levels ofadducts were standardized according to PAH–DNA,

56 S. Horak et al. / Mutation Research 537 (2003) 53–65
which was assayed in parallel to each set (10–12)of analyzed samples. Initially, both nuclease P1 di-gestion and butanol extraction were used to analyzeseveral samples of DNA from sperm cells, and therelative adduct levels (RAL) obtained with either ofthese procedures were compared. Since adduct levelswere two- to five-fold higher in samples treated withnuclease P1, this procedure was chosen for furtheranalysis of all sperm samples.
2.5. Statistical analyses
S-PLUS 4.5 software of MathSoft was used for dataanalysis. Since most of the data were not distributednormally, a non-parametric approach was preferred.The Box–Cox algorithm was applied when the datatransformation into normal distribution was necessary(data cleaning procedure, linear regression). To deter-mine differences between subject groups with respectto adduct levels and sperm parameters, the ANOVAKruskal–Wallis, Mann–WhitneyU or G test was used.The correlations between adduct levels and sperm pa-rameters or measures of smoking, alcohol, and coffeehabits were estimated by the Spearman rank correla-tion coefficient. For all analyses,P < 0.05 was con-sidered statistically significant.
3. Results
3.1. Characterization of DNA adducts in sperm
To characterize bulky DNA adducts in human spermcells, samples were analyzed by the32P-postlabelingmethod. DNA from total blood and DNA damagedby a mixture of PAHs were also analyzed in parallel(Fig. 1). RAL determined in sperm DNA by nucleaseP1 treatment were at least five-fold lower as comparedto blood DNA from the same donor (approximately1 and 5 adducts per 108 nucleotides or 0.03 and0.15 fmol adducts/�g DNA, respectively). Althoughdetermined for only three donors, these data are inagreement with our previous results showing similaradduct levels in lymphocytes or granulocytes of malesfrom the Silesian population[19]. When DNA sam-ples from either sperm or blood were extracted withbutanol to enrich non-polar nucleotides, the patterns of
DNA adducts were similar to that of the PAH-treatedDNA standard. However, after treatment with nucle-ase P1, the patterns of DNA adducts in sperm andblood were different from that of PAH–DNA. Further-more, these more polar DNA adducts were differentin sperm and blood cells (Fig. 1). We have not de-tected any adduct pattern that would be common toall sperm samples. However, several more “typical”patterns were detected, and some of them are pre-sented inFig. 2B. One particular adduct spot (spot“S” in Fig. 1; arrowheads inFig. 2B) was detectedin the majority of the sperm samples (140 out of179) treated with nuclease P1. This particular adductspot contributed 10–90% of the total adducts in dif-ferent samples. The relative contribution of adductsin spot S (percentage of total adducts) positivelycorrelated to total amount of adducts (rS = 0.2674,P = 0.0003).
Adduct levels in each of 179 individuals are de-picted in Fig. 2A. These ranged from 0.34 × 10−2
to 18.5 × 10−2 fmol/�g DNA, with a mean of3.2 × 10−2 ± 2.7 × 10−2 (standard deviation). Thevalues did not have a normal distribution (P < 0.001,Shapiro–Wilk test). Adduct levels were comparedbetween groups of individuals differently exposedto environmental and occupational factors. To deter-mine seasonal variations in adduct levels, sampleswere divided into “summer” (collected from Mayto October) and “winter” (collected from Novem-ber to April). No significant difference was foundbetween these groups (P = 0.626; Fig. 3A). Inhabi-tants of Silesia or large cities (with populations largerthan 100,000) were classified as an “environmentallyexposed” group. No difference (P = 0.541) wasfound between this group and a group of inhabitantsof “unpolluted” areas (Fig. 3B). Individuals workingsuch as drivers, miners, farmers, or others exposedto chemicals or sources of electromagnetic fields inthe workplace were classified as an “occupationallyexposed” group. No difference (P = 0.496) wasfound between this group and a group of “unexposed”persons (Fig. 3C). In addition, no significant dif-ferences were found between these same groups ofsubjects when sperm parameters (sperm concentra-tion and motility) were analyzed (data not shown),except an “occupationally exposed” group thatshows lower sperm mobility compared to unexposedpersons (P = 0.038).

S. Horak et al. / Mutation Research 537 (2003) 53–65 57
Fig. 1. Thin-layer chromatograms of DNA samples analyzed by the32P-postlabeling method. DNA was either treated in vitro with amixture of PAHs (PAH–DNA), or purified from total human blood (blood DNA) or human semen (sperm DNA). Modified nucleotideswere enriched by either nuclease P1 treatment or butanol extraction. Shown is an enzyme-only control, a chromatogram of dG–BPDEadduct, and the directions of chromatography. A well-defined spot typical for sperm DNA is marked “S”.
3.2. Correlation between DNA adducts, fertility,and semen parameters
When adduct levels were compared between agroup of patients with male-factor infertility andhealthy individuals, higher amounts of adductswere found in the “infertile” group (P = 0.012;
Fig. 4A). Similar to the total amounts of adducts,the amounts of adducts in the spot S were signif-icantly higher in the infertile group (P = 0.0014).Because the “infertile” and “healthy” groups of sub-jects differed significantly (P < 0.0001) in respectof sperm concentration and motility, the correla-tion between these sperm parameters and adduct

58 S. Horak et al. / Mutation Research 537 (2003) 53–65
Fig. 2. Bulky adducts in DNA from human sperm cells. (A) Levels of DNA adducts in each of 179 analyzed samples. The median isrepresented by the heavy horizontal line. Healthy smokers are marked by filled circles, healthy never-smokers are marked by empty circles.(B) Patterns of adduct spots representative for DNA from human sperm. Spot “S” has been marked with arrowheads.
levels were determined in further more detailedanalyses.
Significantly higher levels of DNA adducts (P =0.001) were determined in a group of subjectswith oligozoospermia (sperm concentration below
20 × 106 ml−1) than in individuals with a normalsperm concentration (Fig. 4B). The relationshipbetween the levels of DNA adducts and parame-ters of semen quality was analyzed separately forhealthy persons (n = 83) and patients with suspected

S. Horak et al. / Mutation Research 537 (2003) 53–65 59
Fig. 3. Differences in levels of sperm DNA adduct between differently exposed individuals. (A) The levels of DNA adducts in spermcollected during summer and winter. (B) The levels of DNA adducts in sperm of inhabitants of large cities or Silesia (exposed) andinhabitants of unpolluted rural areas (non-exposed). (C) The levels of DNA adducts in sperm of individuals exposed and non-exposed topotential genotoxic factors in the work place. Significance levels (P) of the difference were determined by Mann–WhitneyU test. DNAadduct levels are depicted as femtomoles per microgram of DNA× 102. Shown are the median, the maximum and minimum values, andupper and lower quartiles.
male-factor infertility (n = 93). A significant neg-ative correlation between sperm concentration andthe levels of DNA adducts was found for the infer-tile group (rS = −0.228, P = 0.027; Fig. 5A). Asignificant negative correlation between the levels ofDNA adducts and sperm motility (percentage of sper-matozoa with rapid forward progression) was alsodetermined for this group (rS = −0.225,P = 0.029;Fig. 5B). The quality of sperm can be also expressedas the concentration of motile spermatozoa (con-centration of spermatozoa× percentage of rapidlymoving spermatozoa). A highly significant negativecorrelation between this parameter and the levels ofDNA adducts was observed in the infertile group
(rS = −0.285, P = 0.005; Fig. 5C). In contrast, amuch weaker correlation between the levels of DNAadducts and parameters of semen quality was foundwhen the group of healthy persons was analyzed. Thefollowing correlation was established:rS = −0.173and P = 0.118; rS = −0.066 andP = 0.555;rS = −0.166 andP = 0.132, respectively for spermconcentration, motility, and concentration of motilespermatozoa. Semen volume of the vast majority ofsamples (98%) was at its normal value (2.0 ml ormore, according to WHO standards[17]) and thisparameter was not studied in further analyses.
Correlation between the amount of leukocytes de-tected in semen and DNA adduct levels in sperm was

60 S. Horak et al. / Mutation Research 537 (2003) 53–65
Fig. 4. The levels of sperm DNA adduct in relation to fertility and semen parameters. (A) The levels of DNA adducts in sperm ofhealthy individuals and patients with suspected male-factor infertility. (B) The levels of DNA adducts in sperm of individuals sufferingfrom oligozoospermia and individuals with normal concentrations of spermatozoa (control). (C) The levels of DNA adducts in spermof individuals with low and high concentrations of leukocytes in semen. Significance levels (P) of the difference were determined byMann–WhitneyU test. DNA adducts levels are depicted as femtomoles per microgram of DNA×102. Shown are the median, the maximumand minimum values, and upper and lower quartiles.
also determined. A significant positive correlation(rS = 0.209, P = 0.006) between the concentra-tion of leukocytes and the adduct level in spermwas found. Two groups of samples were analyzedin parallel: samples of semen where the leukocyteconcentrations were below the median (2.3% of sper-matozoa) and samples where the leukocyte concen-trations were equal or above the median (referred aslow- and high-leukocyte groups, respectively). The“infertile” group and “high-leukocyte” group par-tially overlapped (70% of infertile patients belong tothe high-leukocyte group). The “low-leukocyte” and“high-leukocyte” groups differed in respect to thelevels of DNA adducts (P = 0.044;Fig. 4C). Similarcorrelations between the levels of DNA adducts and
the concentration of motile spermatozoa were detectedwhen both groups were analyzed (rS = −0.253,P =0.019 andrS = −0.252,P = 0.016, respectively, forlow- and high-leukocyte groups). Re-evaluation ofdifferences in adduct levels between different occu-pationally and environmentally exposed subjects hasnot revealed a statistical significance when the low-and high-leukocyte group were analyzed.
3.3. Effects of cigarette smoking and coffee oralcohol consumption
To determine possible effects of behavior-relatedexposure to genotoxic factors upon formation ofDNA adducts in sperm, adduct levels were compared

S. Horak et al. / Mutation Research 537 (2003) 53–65 61
Fig. 5. Correlation between the levels of sperm DNA adducts and measures of sperm quality in the group of infertile patients. (A)Correlation between levels of DNA adducts in sperm cells and sperm concentration (amount of spermatozoa× 106 ml−1). (B) Correlationbetween levels of DNA adducts and sperm motility (percentage of rapidly moving spermatozoa). (C) Correlation between levels of DNAadducts in sperm cells and the concentration of motile spermatozoa (amount of rapidly moving spermatozoa× 106 ml−1). DNA adductslevels are shown as femtomoles per microgram of DNA× 102. Presented are the values of SpearmanR correlation coefficient (rS) and itssignificance level (P); n = 93.
among groups of subjects that differed with respectto cigarette smoking and alcohol or coffee drinking.The non-smokers group included persons who eitherhad never smoked or had stopped smoking at least 2years before enrolment in this study. All smokers hadsmoked cigarettes for at least 2 years before the study.Only modest differences (P = 0.054) in the levelsof adducts was detected between smokers (n = 52)and non-smokers (n = 90), when the total group wasanalyzed. When the relationship between the levelsof DNA adducts and either the number of cigarettesconsumed daily, the amount of nicotine and tar sub-stances in consumed cigarettes, or years of smokingwas analyzed, no statistical significant correlation was
found (P > 0.25 in all cases, data not shown). Thedifference in levels of DNA adducts between smok-ers and non-smokers was also determined separatelyin a group of subjects with suspected male-factorinfertility and in a group of presumably healthy indi-viduals. No significant differences between smokersand non-smokers with suspected male-factor infertil-ity was observed (P = 0.646). However, in the groupof healthy individuals, the level of DNA adducts inthe smokers subgroup (n = 22) was significantlyhigher than in the non-smokers subgroup (n = 48)(P = 0.026); both the mean value and the medianwere about 1.5-fold higher in the group of smokers.When 15 healthy ex-smokers were excluded from the
62 S. Horak et al. / Mutation Research 537 (2003) 53–65
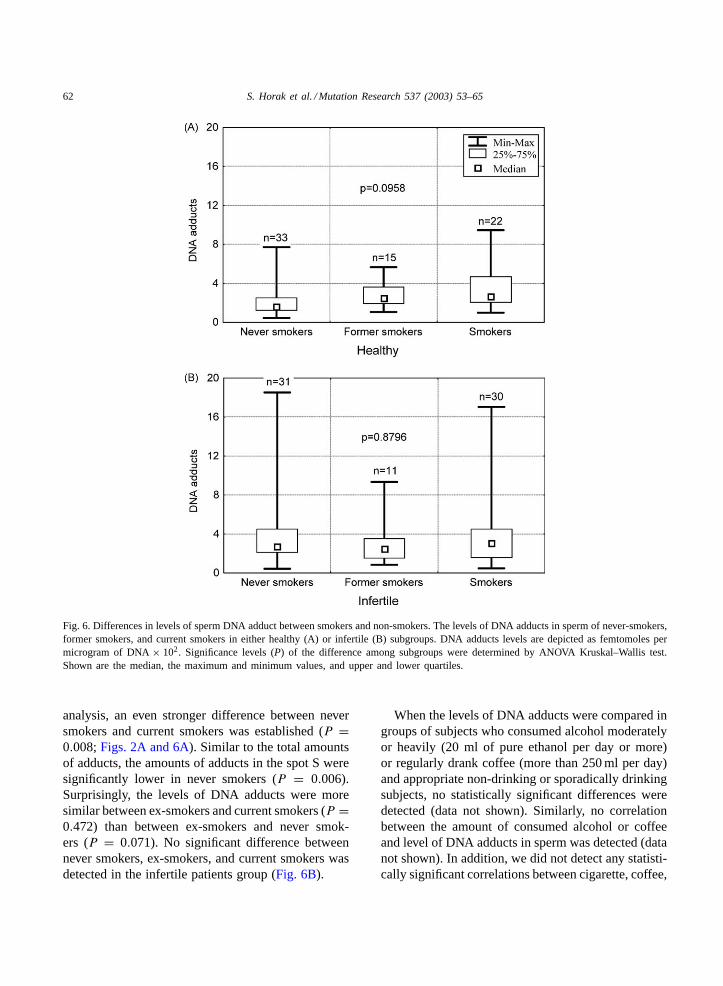
Fig. 6. Differences in levels of sperm DNA adduct between smokers and non-smokers. The levels of DNA adducts in sperm of never-smokers,former smokers, and current smokers in either healthy (A) or infertile (B) subgroups. DNA adducts levels are depicted as femtomoles permicrogram of DNA× 102. Significance levels (P) of the difference among subgroups were determined by ANOVA Kruskal–Wallis test.Shown are the median, the maximum and minimum values, and upper and lower quartiles.
analysis, an even stronger difference between neversmokers and current smokers was established (P =0.008;Figs. 2A and 6A). Similar to the total amountsof adducts, the amounts of adducts in the spot S weresignificantly lower in never smokers (P = 0.006).Surprisingly, the levels of DNA adducts were moresimilar between ex-smokers and current smokers (P =0.472) than between ex-smokers and never smok-ers (P = 0.071). No significant difference betweennever smokers, ex-smokers, and current smokers wasdetected in the infertile patients group (Fig. 6B).
When the levels of DNA adducts were compared ingroups of subjects who consumed alcohol moderatelyor heavily (20 ml of pure ethanol per day or more)or regularly drank coffee (more than 250 ml per day)and appropriate non-drinking or sporadically drinkingsubjects, no statistically significant differences weredetected (data not shown). Similarly, no correlationbetween the amount of consumed alcohol or coffeeand level of DNA adducts in sperm was detected (datanot shown). In addition, we did not detect any statisti-cally significant correlations between cigarette, coffee,

S. Horak et al. / Mutation Research 537 (2003) 53–65 63
or alcohol consumption and measures of sperm qual-ity (data not shown). The concentration of sperm wasslightly lower in smokers compared to non-smokers,but the difference was not statistically significant (P =0.075).
4. Discussion
The majority of individuals in an environmentallyexposed group studied in this project were inhabi-tants of Silesia, a heavily industrialized and pollutedarea in the south of Poland. It has been reportedthat the Silesian population showed elevated levelsof bulky DNA adducts in white blood cells[20].That finding was followed by reports of large sea-sonal variations in adduct levels that matched theseasonal variations in the air concentrations of PAHs.The effect was seen mostly in lymphocytes, wherelevels of adduct in “winter” samples were two- tothree-fold higher than in “summer” samples from thesame individuals[21,22]. When levels of bulky DNAadducts in sperm were analyzed in the present work,neither differences between inhabitants of pollutedand unpolluted areas nor seasonal variations werefound, which is in contrast to the lymphocyte-basedstudies.
The levels of bulky DNA adducts are known to behigher in blood cells of smokers than of non-smokers[13]. Although actual values given in different stud-ies have varied, about a 2.5-fold difference had beenreported when purified lymphocytes from smokersand non-smokers were analyzed[23]. In this work,we observed only marginally (∼1.2-fold) higher lev-els of bulky DNA adducts in sperm of smokers thannon-smokers when all donors were tested. A higherdifference (∼1.7-fold increase) was found betweensmokers and never smokers in a group of healthydonors only. Interestingly, published work shows thatsmoking is more likely to be associated with de-creased sperm quality when healthy volunteers but notinfertility clinic patients are tested (reviewed in ref.[6]). An earlier study of Gallagher et al.[16], whichanalyzed bulky DNA adducts in sperm of a smallgroup of healthy donors, revealed no difference be-tween smokers and non-smokers. On the other hand,Zenzes et al.[9] reported about two-fold higher levelsof PAH–DNA adducts in the sperm of smokers com-
pared to that of non-smokers. The immunocytochem-ical method used in that study allowed specific detec-tion of DNA adducts induced by BPDE and relatedcompounds. However, data presented in this papershow that only a small fraction of the DNA adductsdetected in sperm are of the non-polar class, which arepotentially recognized by anti-BPDE–DNA antibody[19]. This suggests that PAH-induced DNA damagedoes not contribute significantly to the total level ofbulky DNA adducts detected by32P-postlabeling insperm. It clearly appears that results of studies onsmoking and DNA damage in sperm may be greatlyaffected by the choice of the analytical method andthe selection of subjects.
Our data demonstrate a lack of correlation be-tween environmentally related exposure to genotoxiccompounds and the levels of bulky DNA adductsin sperm, and relatively weak effects of compoundspresent in tobacco smoke on the levels of suchadducts. This finding is in clear contrast with datashowing an influence of environmental- and cigarettesmoke-related factors upon DNA damage in whiteblood cells. Furthermore, the levels of bulky DNAadducts in sperm are markedly lower than those inwhite blood cells, suggesting that germ cells are pro-tected against environmental factors. Several mecha-nisms may contribute to resistance of sperm cells toexogenous genotoxic factors: effective restriction ofDNA-reacting compounds by the blood–testis barrier,effective repair of DNA damage during early stages ofspermatogenesis and/or ineffective formation of DNAdamage due to the compact nature of sperm chro-matin. The relative resistance of sperm DNA againstexogenous genotoxic factors, as well as the differentpatterns of adduct spots detected here in blood andsperm cells, suggests that endogenous tissue-specificfactors may play an important role in induction ofDNA adducts detected in spermatozoa. Several bulkyDNA modifications termed I-compounds (or I-spots,from “Indigenous”) have been detected in rodents notexposed to known genotoxic chemicals and their for-mation is strain-, age-, gender-, and tissue-dependent[24]. Steroid estrogens and their metabolites areamong the endogenous DNA-reactive molecules thathave been identified. Estrogen quinone metabolitesbind covalently to DNA and chronic exposure ofrodents to either synthetic or natural (e.g. estradiol)estrogens induces formation of bulky DNA adducts

64 S. Horak et al. / Mutation Research 537 (2003) 53–65
in hormone-responsive tissues that can be detectedby 32P-postlabeling[25]. The identities of the DNAadducts detected in human spermatozoa remain tobe established. However, similar levels (about 1 per108 nucleotides) and structural features (resistance tonuclease P1) between I-spots and adducts detectedin human sperm should be noted, and one may alsosuggest endogenous compounds related to hormonemetabolites as potential inducers of sperm adducts.
Epidemiological and experimental studies indicatethat DNA lesions reflect exposure to carcinogenicfactors and there is no doubt that carcinogen-inducedDNA adducts are involved in tumor initiation. How-ever, much less is known about the role of bulky DNAmodifications induced by endogenous sources. Herewe show that the levels of covalent modifications inhuman sperm DNA are negatively correlated withspermatozoa concentration and motility, measures ofsperm quality. Similarly, the levels of bulky DNAadducts were higher in sperm of patients with sus-pected male-factor infertility than in sperm of healthyindividuals, and several mechanisms may contributeto this correlation. One could suggest an indirect re-lationship between elevated levels of DNA adductsand decreased sperm quality. For example, the de-tected bulky DNA modifications may reflect the in-fluence of endo- or exogenous factors, whose effectson germ-line cells involve other mechanisms (e.g.general cytotoxicity, hormonal imbalance). A directrelationship between both factors (e.g. DNA adductsbeing a causative factor of low sperm quality) seemsmuch less probable. Since a high level of leuko-cytes (where adducts were presumably more frequentthan in spermatozoa) was often observed in semenof patients with low concentrations and/or motil-ity of spermatozoa, these might partially contributeto elevated levels of DNA adducts in “low-quality”sperm cells. However, a significant negative correla-tion between level of DNA adducts and measures ofsperm quality was observed also in samples with lowleukocyte content, showing that an elevated level ofDNA adducts in “low-quality” sperm is an intrinsicfeature of spermatozoa. Although a functional cor-relation between bulky modifications of sperm DNAand quality of spermatozoa is not clear at the mo-ment, our results imply that DNA adducts can beapplied as a potential biomarkers in studies of humaninfertility.
Acknowledgements
We thank Ms. Lucyna Ponge for excellent technicalassistance. This work was supported in part by thePolish State Committee for Scientific Research KBN,Grant 8T11E01319 (to J.P. and P.W.).
References
[1] World Health Organization, Infections, pregnancies andinfertility: perspectives on prevention, Fertil. Steril. 47 (1987)964–968.
[2] A. Giwercman, J.P. Bonde, Declining male fertility andenvironmental factors, Endocrinol. Metab. Clin. North Am.27 (1998) 807–830.
[3] J. Lahdetie, Occupation- and exposure-related studies onhuman sperm, J. Occup. Environ. Med. 37 (1995) 922–930.
[4] S. Tas, R. Lauwerys, D. Lison, Occupational hazards for themale reproductive system, Crit. Rev. Toxicol. 26 (1996) 261–307.
[5] K.S. Hruska, P.A. Furth, D.B. Seifer, F.I. Sharara, J.A. Flaws,Environmental factors in infertility, Clin. Obstet. Gynecol. 43(2000) 821–829.
[6] M.F. Vine, Smoking and male reproduction: a review, Int. J.Androl. 19 (1996) 323–337.
[7] International Agency for Research on Cancer, TobaccoSmoking. IARC Monographs on the Evaluation of theCarcinogenic Risks of Chemicals to Humans, vol. 38, IARC,Lyon, France, 1986.
[8] G.L. Ginsberg, B. Atherholt, Transport of DNA-adductingmetabolites in mouse serum following benzo(a)pyreneadministration, Carcinogenesis 10 (1989) 673–679.
[9] M.T. Zenzes, R. Bielecki, T.E. Reed, Detection ofbenzo(a)pyrene diol epoxide–DNA adducts in sperm of menexposed to cigarette smoke, Fertil. Steril. 72 (1999) 330–335.
[10] R.E. Sotomayor, G.A. Sega, Unscheduled DNA synthesisassay in mammalian spermatogenic cells: an update, Environ.Mol. Mutagen. 36 (2000) 255–265.
[11] C.C. Harris, Future directions in the use of DNA adductsas internal dosimeters for monitoring human exposure toenvironmental mutagens and carcinogens, Environ. HealthPerspect. 62 (1986) 185–191.
[12] A.C. Beach, R.C. Gupta, Human biomonitoring and the32P-postlabeling assay, Carcinogenesis 13 (1992) 1053–1074.
[13] J. Lewtas, J. Mumford, R.B. Everson, B. Hulka, T. Wilcosky,W. Kozumbo, C. Thompson, M. George, L. Dobias, R. Sram,X. Li, J.E. Gallagher, Comparison of DNA adducts fromexposure to complex mixtures in various human tissues andexperimental systems, Environ. Health Perspect. 99 (1993)89–97.
[14] H.M. Shen, S.E. Chia, Z.Y. Ni, A.L. New, B.L. Lee, C.N.Ong, Detection of oxidative DNA damage in human spermand the association with cigarette smoking, Reprod. Toxicol.11 (1997) 675–680.

S. Horak et al. / Mutation Research 537 (2003) 53–65 65
[15] M. Sergerie, S. Ouhilal, F. Bissonnette, J. Brodeur, G. Bleau,Lack of association between smoking and DNA fragmentationin the spermatozoa of normal men, Hum. Reprod. 15 (2000)1314–1321.
[16] J.E. Gallagher, M.F. Vine, M.M. Schramm, J. Lewtas, M.H.George, B. Hulka, R.B. Everson,32P-postlabeling analysisof DNA adducts in human sperm cells from smokers andnonsmokers, Cancer Epidemiol. Biomarkers Prev. 2 (1993)581–585.
[17] World Health Organization, WHO Laboratory Manual for theExamination of Human Semen and Sperm-Cervical MucusInteractions, 4th ed., Cambridge University Press, Cambridge,1999.
[18] R.C. Gupta, M.V. Reddy, K. Randerath,32P-postlabelinganalysis of non-radioactive aromatic carcinogen–DNAadducts, Carcinogenesis 3 (1982) 1081–1092.
[19] P. Widlak, E. Grzybowska, K. Hemminki, R. Santella, M.Chorazy, 32P-postlabelling of bulky human DNA adductsenriched by diffrent methods including immunoaffinitychromatography, Chem. Biol. Interact. 99 (1996) 99–107.
[20] F.P. Perera, K. Hemminki, E. Grzybowska, G. Motykiewicz,J. Michalska, R.M. Santella, T.L. Young, C. Dickey, P.
Brandt-Rauf, I. De Vivo, W. Blaner, W.Y. Tsai, M. Chorazy,Molecular and genetic damage in humans from environmentalpollution in Poland, Nature 360 (1992) 256–258.
[21] E. Grzybowska, K. Hemminki, J. Szeliga, M. Chorazy,Seasonal variation of aromatic DNA adducts in humanlymphocytes and granulocytes, Carcinogenesis 14 (1993)2523–2526.
[22] K. Hemminki, E. Grzybowska, P. Widlak, M. Chorazy, DNAadducts in environmental, occupational and life-style studiesin human biomonitoring, Acta Biochim. Pol. 43 (1996) 305–312.
[23] K. Savela, K. Hemminki, DNA adducts in lymphocytesand granulocytes of smokers and nonsmokers detected bythe 32P-postlabeling assay, Carcinogenesis 12 (1991) 503–508.
[24] K. Randerath, E. Randerath, G.D. Zhou, D. Li, Bulkyendogenous DNA modifications (I-compounds)—possiblestructural origins and functional implications, Mutat. Res. 424(1999) 183–194.
[25] D. Roy, J.G. Liehr, Estrogen, DNA damage and mutations,Mutat. Res. 424 (1999) 107–115.