cloned embryos from semen. part 2: intergeneric nuclear transfer of semen-derived eland (...
TRANSCRIPT

CLONING AND STEM CELLSVolume 10, Number 1, 2008© Mary Ann Liebert, Inc.DOI: 10.1089/clo.2007.0069
Cloned Embryos from Semen. Part 2: IntergenericNuclear Transfer of Semen-Derived Eland
(Taurotragus oryx) Epithelial Cells into Bovine Oocytes
LIESL NEL-THEMAAT,1,2 MARTHA C. GÓMEZ,2 C. EARLE POPE,2 MONICA LOPEZ,2,3
GEMECHU WIRTU,2,4 JILL A. JENKINS,5 ALEX COLE,2 BETSY L. DRESSER,2,3
KENNETH R. BONDIOLI,1 and ROBERT A. GODKE1
ABSTRACT
The production of cloned offspring by nuclear transfer (NT) of semen-derived somatic cellsholds considerable potential for the incorporation of novel genes into endangered speciespopulations. Because oocytes from endangered species are scarce, domestic species oocytesare often used as cytoplasts for interspecies NT. In the present study, epithelial cells isolatedfrom eland semen were used for intergeneric transfer (IgNT) into enucleated bovine oocytesand compared with bovine NT embryos. Cleavage rates of bovine NT and eland IgNT em-bryos were similar (80 vs. 83%, respectively; p � 0.05); however, development to the morulaand blastocyst stage was higher for bovine NT embryos (38 and 21%, respectively; p � 0.0001),than for eland IgNT embryos (0.5 and 0%, respectively). DNA synthesis was not observed ineither bovine NT or eland IgNT cybrids before activation, but in 75 and 70% of bovine NTand eland igNT embryos, respectively, cell-cycle resumption was observed at 16 h postacti-vation (hpa). For eland IgNT embryos, 13% had �8 cells at 84 hpa, while 32% of the bovineNT embryos had �8 cells at the same interval. However, 100 and 66% of bovine NT and elandIgNT embryos, respectively, that had �8 cells synthesized DNA. From these results we con-cluded that (1) semen-derived epithelial cell nuclei can interact and be transcriptionally con-trolled by bovine cytoplast, (2) the first cell-cycle occurred in IgNT embryos, (3) a high fre-quency of developmental arrest occurs before the eight-cell stage in IgNT embryos, and (4)IgNT embryos that progress through the early cleavage stage arrest can (a) synthesize DNA,(b) progress through subsequent cell cycles, and (c) may have the potential to develop fur-ther.
161
INTRODUCTION
SINCE THE BIRTH OF THE FIRST LAMB cloned froman adult somatic cell (Wilmut et al., 1997),
cloning technology has been used not only in ba-
sic research, but also for the multiplication of elitelivestock, production of transgenic animals, andconservation of endangered species (Campbell etal., 2005). Nuclear transfer (NT) could also bevaluable in conservation of species that are en-
1Department of Animal Sciences, Louisiana State University Agricultural Center, Baton Rouge, Louisana.2Audubon Center for Research of Endangered Species, New Orleans, Louisiana.3Department of Biological Sciences, University of New Orleans, New Orleans, Louisana.4National Wetlands Research Center, US Geological Survey, Lafayette, Louisana.

dangered due to inbreeding. The introduction ofnovel genes would dilute the effects of deleteri-ous recessive genes (Ryder and Benirschke, 1997).
Somatic cells derived from a variety of donortissues have been used to create cloned animals(Brem and Kuhholzer, 2002; Galli et al., 1999; Katoet al., 1998; Wayakama et al., 1998; Wilmut et al.,1997). It is a common practice to reconstructcloned embryos with either fresh donor cells dis-aggregated from a monolayer, or with frozendonor cells thawed just prior NT. If cell lines werenot derived and cryopreserved from either thelive or recently deceased animal, no nucleardonor material would be available. Therefore, thepossibility of using somatic cells isolated fromcryopreserved semen for NT would enable thereintroduction of previously inaccessible genesfrom earlier generations. Therefore, banks of cry-opreserved semen are a potential source of novelgenes that could be reincorporated into an ani-mal population with a limited gene pool. Re-cently, we demonstrated that epithelial cells canbe successfully isolated from ram (Ovis aries) andCommon eland antelope (Taurotragus oryx) se-men and used to establish cell lines in vitro (Nel-Themaat et al., 2007).
Usually, cloned embryos are derived by thetransfer of an adult or fetal somatic cell into a con-specific oocyte (intraspecies) (Baguisi et al., 1999;Galli et al., 2003; Kato et al., 1998; Polejaeva et al.,2000; Wayakama et al., 1998; Wilmut et al., 1997).However, because oocytes from endangeredspecies are rarely available, oocytes of closely re-lated domestic or laboratory animals are used asrecipient cytoplasts. Pregnancies and live birthsof cloned endangered mammals such as gaur(Lanza et al., 2000), mouflon (Loi et al., 2001), ban-teng (Janssen et al., 2004; Sansinena et al., 2005),and African wildcats (Gómez et al., 2004) havebeen produced by interspecies NT. In contrast, in-tergeneric nuclear transfer (IgNT) has been lesssuccessful, and viable mammalian progeny havenot been produced. Even though no live mam-mals have been produced by IgNT, early embryodevelopment has been achieved after IgNT of so-matic cells of endangered mammals into bovineoocytes with cleavage rates similar to that ofbovine NT embryos (Bui et al., 2002; Lee et al.,2003; Lu et al., 2005; Murakami et al., 2005; Ty etal., 2003). Development was arrested in a high fre-quency of IgNT cloned embryos, and was coinci-dent with embryonic genome activation (Lu et al.,2005; Matshikiza et al., 2004; Murakami et al.,
2005; Ty et al., 2003). Thus, few IgNT embryos de-veloped to the blastocyst stage (Bui et al., 2002;Lee et al., 2003; Lu et al., 2005; Matshikiza et al.,2004; Murakami et al., 2005; Ty et al., 2003), likelydue to multiple factors, but arrest at the time ofembryonic genome activation possibly is associ-ated with improper early reprogramming due tothe genetic distance between donor nucleus andrecipient oocyte, as well as abnormal nuclear re-modeling (Hamilton et al., 2004).
The oocyte cytoplasm controls nuclear eventsand DNA replication of the transplanted nucleus(Campbell et al., 1993); therefore, cytoplasmic in-compatibilities between the nucleus and somaticcell may contribute to the abnormal nuclear re-modeling.
In the present study, we evaluated the abilityof eland epithelial cells isolated from semen tosupport early in vitro embryonic developmentwhen transferred to bovine cytoplasts. In addi-tion, we compared the ability of eland semen-de-rived and bovine fibroblast cells to initiate DNAreplication after nuclear transfer.
MATERIALS AND METHODS
Chemical reagents
All chemicals were obtained from SigmaChemical Co. (St Louis, MO), unless otherwisestated.
Animals
A 7-year-old common eland bull was main-tained on a diet consisting of 8 kg of herbivorefeed (Purina ADF-16) at the Freeport-McMoRanAudubon Species Survival Center (FMA-SSC)near New Orleans, Louisiana. The InstitutionalAnimal Care and Use Committee of AudubonNature Institute approved all eland procedures.
Establishment of donor cell line
An eland epithelial cell line was established asdescribed previously (Nel-Themaat et al., 2005).Briefly, eland semen was collected by a combi-nation of rectal massage and electroejaculation(Wirtu et al., 2002, 2007) diluted in 10 mL of Ca2�
and Mg2�-free Dulbecco’s phosphate-bufferedsaline (DPBS, Gibco, Grand Island, NY), and cen-trifuged at 500 � g for 10 min. Samples werewashed two more times and the final pellet was
NEL-THEMAAT ET AL.162

resuspended in 2.5 mL of culture medium (CM)consisting of Minimum Essential Medium alphamedium (�-MEM; Gibco) supplemented with15% newborn calf serum (NCS), 1% MEMnonessential amino acids (NEAA), 500 IU/mL ofpenicillin, and 0.5 mg/mL of streptomycin (5%Pen/Strep; Cellgro, Herndon, VA), and 250�g/mL of gentamicin (Gibco). The resuspendedpellet was plated on collagen-coated (Type 1 fromcalf skin) 35-mm culture plates and cultured for24 h before rinsing with CM containing 1%Pen/Strep and no gentamicin. In vitro culture wascontinued until cell colonies covered �0.5 cm2 ofthe culture surface. Then, cell colonies were dis-sociated for subpassages using 0.125% trypsin inDPBS and cultured until passage 3 to 5 before re-suspension in CM supplemented with 10% (v/v)dimethyl sulfoxide (DMSO), and cooled at1.0°C/min to �80°C (Mr. Frosty; Nalgene, Roch-ester, NY) before storage in liquid nitrogen (LN2).
The bovine fibroblast cell line was obtainedfrom an ear biopsy of a domestic cow. Tissue wasminced and cultured in a 35-mm tissue cultureplate in CM until fibroblasts reached �100% con-fluency. Fibroblasts cells were subsequently dis-sociated with 0.25% Trypsin in DPBS and cul-tured in 25-cm2 flasks. At passage 3 to 5, confluentcells were dissociated and cryopreserved in CMplus 10% DMSO in aliquots for NT.
In the evening before eland or bovine NT ses-sions, one vial of frozen cells was thawed andplated on a 35 mm collagen-coated plate (elandepithelial cells) or an uncoated plate (bovine fi-broblasts) containing 3 mL of CM. Immediatelybefore NT, cells were dissociated with 0.125%(eland epithelial cells) or 0.25% (bovine fibro-blasts) trypsin and resuspended in CM.
Analysis of cell-cycle phase by flow cytometry
Eland epithelial cells (passage 3 to 5, 80–100%confluence) were dissociated from the cultureflasks, centrifuged at 300 � g for 5 min and di-luted to 1 � 106 cells/mL. Duplicate sampleswere stained with 50 �g/mL of propidium iodide(PI), 650,000 units/mL of RNAse, and 0.1% (v/v)Triton X-100 (Crissman and Steinkamp, 1973) for20 min at 24°C.
Data from �10,000 nuclei per sample were ac-quired in doublet discrimination mode using aFACScan [Becton Dickinson (BD) Immunocy-tometry Systems, San Jose, CA] flow cytometerequipped with an argon laser. Propidium iodide
is excited at 488 nm, and after binding with DNA,emits red fluorescence at 617 nm and is measuredin FL2. Data were collected using CELLQuestsoftware (BD) with FL2-area (FL-2A) and a linearscale of fluorescence. One parameter (FL2-A)1024-channel histograms and two parameter (FL-2 A and FL-2 width) 1024-channel dot plotswere generated and analyzed with Modfit LT cellcycle analysis software (Verity Software House,Inc., Topsham, ME) using F_DIP_T1.mod.
Oocyte maturation
Bovine ovaries were obtained from a localslaughterhouse and transported to the laboratoryat 30°C in saline solution (0.9%) containing 1%Pen/Strep. Cumulus–oocyte complexes (COCs)were aspirated from follicles (3–10 mm in diam-eter) and cultured in modified TCM-199 contain-ing 1 IU/mL human Chorionic Gonadotropin(hCG), 0.5 IU/mL equine Chorionic Gonadotro-pin (eCG), 10 �g/mL Epidermal Growth Factor(EGF), and 3 mg/mL fatty acid free bovine serumalbumin (BSA, Fraction V; Serological Proteins,Kankakee, IL) (Gómez et al., 2003) for 19 h in 5%CO2 in air at 38.5°C.
Cumulus cells of in vitro-matured oocytes were removed by vortexing in 1 mg/mL ofhyaluronidase for 3 min. Denuded oocytes wereplaced in CR1aa medium supplemented with 5%NCS, 1% NEAA, 2% essential amino acids (EAA),1 mM glutamine, and 50 �g/mL of gentamicin at38.5°C in 5% CO2 in air until further use.
Nuclear transfer
To eliminate the risk of crosscontamination be-tween eland and bovine cells and to reduce vari-ability in the time after maturation at which nu-clear transfer is performed, NT for each specieswas done on alternating days using oocytes fromthe same source. For the bovine intraspecies NTand parthenogenetic activation, a total of threereplicates of each were performed, whereas fourreplicates of eland igNT were performed.
Before enucleation (�20 h postmaturation), de-nuded metaphase II (M-II) oocytes were incu-bated for 10 to 30 min in CR1aa medium con-taining 10 �g/mL of Hoechst 33342 and 7.5�g/mL of cytochalasin B (CCB). After incubation,oocytes were enucleated in TCM-199 supple-mented with 15 mM HEPES, 15 mM NaHCO3,0.36 mM pyruvate, 1 mM glutamine, 2.2 mM cal-cium lactate, 50 �g/mL of gentamicin, 4 mg/mL
NUCLEAR TRANSFER OF SEMEN EPITHELIAL CELLS 163

BSA, and 10 �g/mL of CCB. The metaphase-IIplate was aspirated into an enucleation pipette(20–25 �m outer diameter), and where possible,the first polar body was removed. Removal of themetaphase spindle was confirmed by brief expo-sure (1 sec) to ultraviolet (UV) light using epiflu-orescence microscopy.
A single eland epithelial or bovine fibroblastcell was introduced into the perivitelline space ofeach enucleated oocyte. For fusion, each coupletwas placed in a solution of 0.3 M mannitol and0.1 mM Mg2� between two stainless steel elec-trodes spaced 120 �m apart (LF-101; Nepa Gene,Tokyo, Japan). Membrane fusion was induced byapplying a 0.2-sec AC prepulse of 17 V, 1 MHz;followed by two 35-�sec DC pulses of 14 V at in-tervals of 1 sec. Following the fusion pulses, cou-plets were cultured in CR1aa, and after 20 to 30min, fusion was assessed visually by confirmingthe presence or absence of the donor cell in theperivitelline space. Fused couplets were culturedfor 2 to 3 h in CR1aa medium supplemented with7.8 mM calcium lactate and 2.5 �g/mL of CCB.Activation of fused couplets was performed28–30 h post maturation by incubation in 10 �Mcalcium ionophore for 5 min and subsequent cul-ture for 4 h in 2 mM dimethylaminopurine at38°C in 5% CO2 in air under mineral oil. As a con-trol for the activation protocol, denuded M-IIoocytes were parthenogenetically activated bythe same procedure as described for fused NTcouplets.
Following activation, NT couplets and partheno-genetically activated oocytes (PAOs) were cul-tured in CR1aa medium under mineral oil in 5%CO2, 5% O2 and 90% N2 at 38.5°C for 9 days.
Embryo evaluation
Fused couplets and PAOs were evaluated forfrequency of cleavage on day 3 of in vitro cultureand development to the blastocyst stage on days7, 8, and 9. Evaluations were made using bright-field and fluorencence microscopy. Blastocyst cell numbers, both total and inner cell mass/trophodectoderm cells (ICM/TE), was deter-mined by differential staining of cells with HXTand PI, respectively and fluorescent microscopy.
Couplets that did not cleave by day 3 werefixed at 4°C for 24 to 48 h in ethanol and aceticacid (3:1) and stained with aceto-orcein. The nu-clear status were examined using phase-contrastmicroscopy (400�).
Cleaved NT embryos that had less than fourcells on day 3 and parthenogenetically activatedand NT embryos that did not develop to the blas-tocyst stage by day 9 were incubated for 30 to 60min at 38°C in 20 �g/mL of Hoechst 33342.Stained embryos were placed in a drop of an-tifade reagent (FluoroGuard, Bio-Rad Laborato-ries; Hercules, CA) on a clean glass microslideand cell numbers were counted by epifluores-cence microscopy.
NT and parthenogenetic blastocysts on days 7,8, and 9 of culture were incubated for 1 h inHoechst 33342 as previously described. After in-cubation, blastocysts were exposed to 0.04% Tri-ton X-100 in PBS for 30 sec and then cultured for15 min at 38°C in 25 �g/mL of PI. Stained blas-tocysts were then mounted on microslides as de-scribed above and the ratio of ICM cells to TPDcells per embryo was determined by counting thenumber of blue (ICM) and red (TE) cells stainedby Hoechst and PI, respectively.
Determination of DNA synthesis
Replication potential of eland epithelial orbovine fibroblast cells transferred into in vitro ma-tured bovine oocytes was evaluated by immuno-cytochemical detection of 5-Bromodeoxyuridine(BrdU; Roche Diagnostics Corporation, Indi-anapolis, IN) incorporated into newly synthe-sized DNA strands.
To detect DNA replication, eland and bovineNT cybrids (fused couplets that contain cyto-plasm from the oocyte and the somatic cell) orcleaved embryos were incubated for 12 h in BrdUat different times after NT: (1) nonactivated cy-brids (2 to 3 h after fusion), (2) activated cybrids(4 h postactivation (hpa) fixed at 16 hpa), (3)cleaved NT embryos (72 hpa, fixed at 84 hpa), and(4) bovine PAOs (4 or 72 hpa, fixed at 16 and 84hpa, respectively).
Oocytes, cybrids, and embryos (OCEs) in eachtreatment were incubated in 20 �M BrdU at 38°Cfor 2 h (treatment 1) or 12 h (treatments 2, 3, and4), then washed three times in PBS plus 5% NCS(PBS-S). After labeling, the zona pellucida was re-moved by incubation in 0.3% pronase for 3 to 5min and rinsed in PBS-S. Zona-free OCEs wereimmediately fixed in a solution of 2.5% parafor-maldehyde for 24 h at 4°C. Fixed OCEs wererinsed in PBS-S and membranes permeabilized inPBS supplemented with 0.1% of Triton X-100 for10 min. After permeabilization, the OCEs were
NEL-THEMAAT ET AL.164
rinsed in PBS supplemented with 0.1% Tween 20(PBS-T20) and finally cultured in 7.5 mg/mL ofglycine in PBS-T20 for 10 min to prevent non-specific binding. Then, OCEs were rinsed in PBS-T20 and DNA denaturation was performed by 30min incubation in 2N HCl.
After denaturation OCEs were incubated inPBS-S for 5 min followed by incubation in bufferconsisting of PBS-T20 plus 0.5% BSA for 10 minbefore exposure to 50 �L of monoclonal anti-BrdU antibody conjugated with FITC for 45 minat 38°C. Finally, chromatin was stained with 25�g/mL PI in PBS-S for 5 min and OCEs wererinsed twice for 3 and 15 min before beingmounted on clean glass microslides and coveredwith mounting medium before viewing with flu-orescence microscopy.
When DNA synthesis was evaluated by epi-fluoresence microscopy, nuclei that incorporatedBrdU (S-phase) fluoresced green while all nucleifluoresced red. Nuclear stages of embryos werealso assessed and total cell numbers werecounted.
Statistical analysis
Results were analyzed using Graphpad Instatsoftware (Graphpad Software, Inc., San Diego,
CA) and, for all experiments, a p-value of lessthan 0.05 was considered to be significant. De-velopmental comparisons between bovine NTand eland IgNT embryos were performed usingthe chi-square test, while percentage of blas-tomeres that contributed to the ICM and tro-phoblast were compared by unpaired t-tests withWelch correction. Cell numbers that incorporatedBrdU between cell number groups were com-pared using Fisher’s exact test.
RESULTS
Analysis of cell cycle by flow cytometry
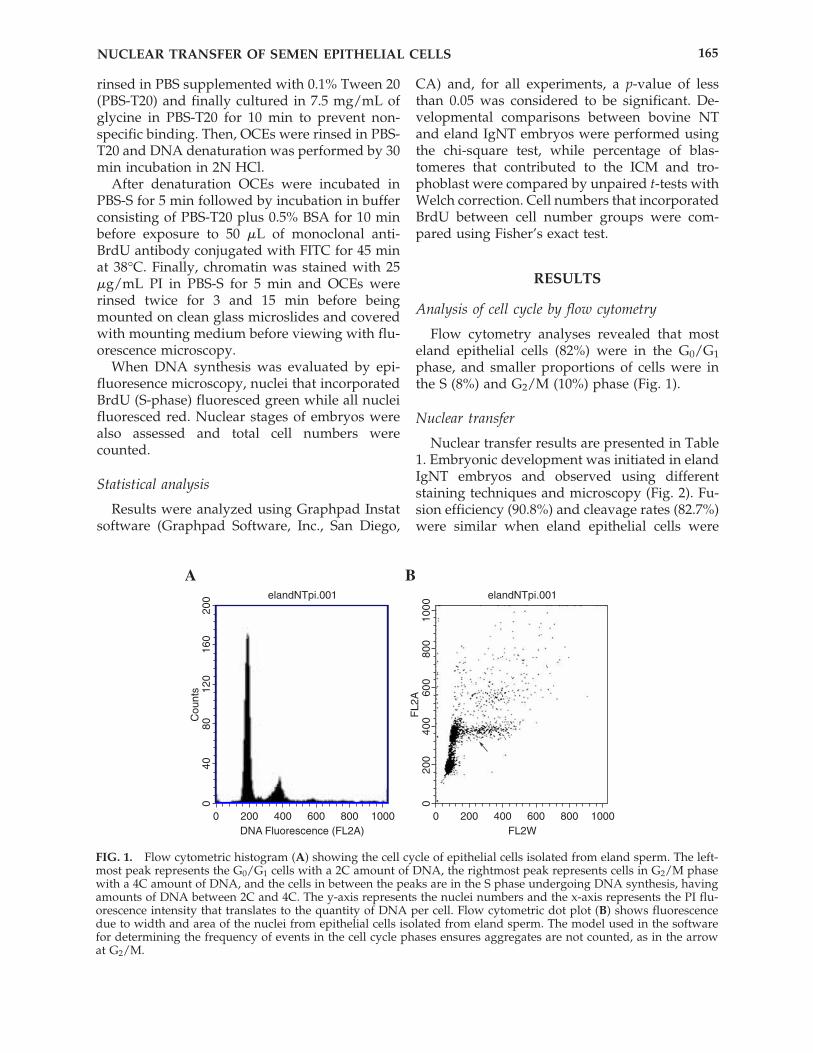
Flow cytometry analyses revealed that mosteland epithelial cells (82%) were in the G0/G1phase, and smaller proportions of cells were inthe S (8%) and G2/M (10%) phase (Fig. 1).
Nuclear transfer
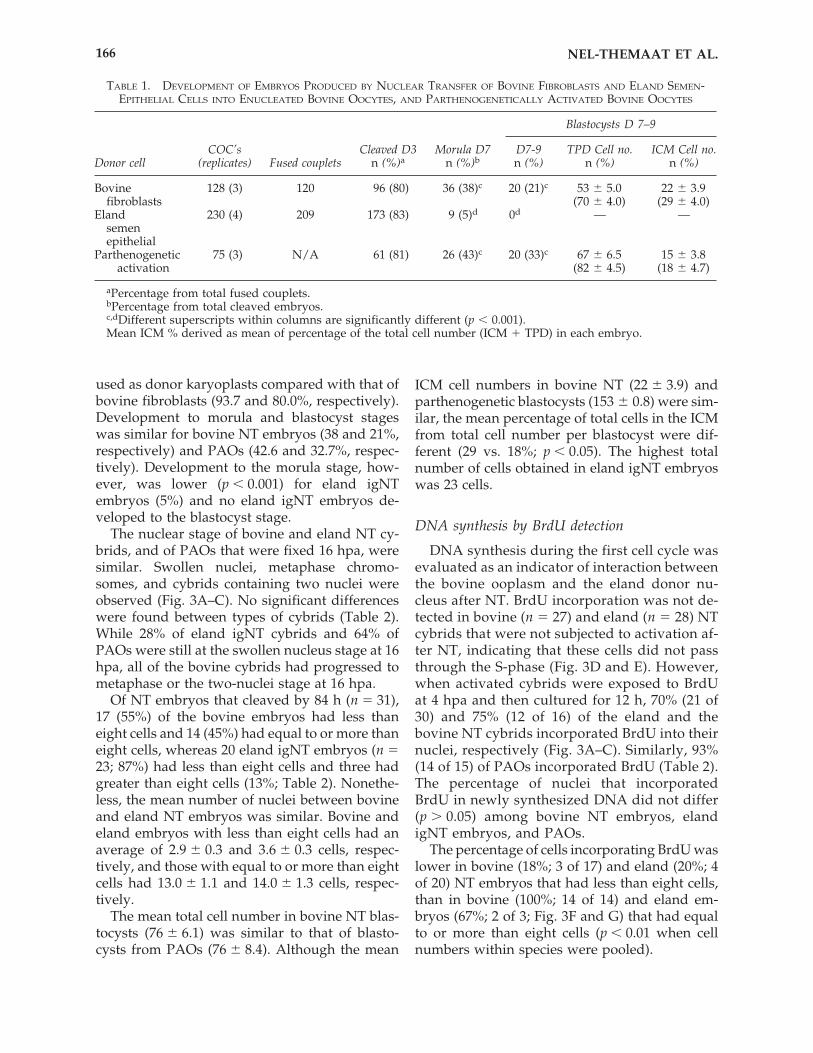
Nuclear transfer results are presented in Table1. Embryonic development was initiated in elandIgNT embryos and observed using differentstaining techniques and microscopy (Fig. 2). Fu-sion efficiency (90.8%) and cleavage rates (82.7%)were similar when eland epithelial cells were
NUCLEAR TRANSFER OF SEMEN EPITHELIAL CELLS 165
A
0
0
DNA Fluorescence (FL2A)
elandNTpi.001
Cou
nts
200 400 600 800 1000
4080
120
160
200
B
0
0
FL2W
elandNTpi.001
FL2
A
200 400 600 800 1000
200
400
600
800
1000
FIG. 1. Flow cytometric histogram (A) showing the cell cycle of epithelial cells isolated from eland sperm. The left-most peak represents the G0/G1 cells with a 2C amount of DNA, the rightmost peak represents cells in G2/M phasewith a 4C amount of DNA, and the cells in between the peaks are in the S phase undergoing DNA synthesis, havingamounts of DNA between 2C and 4C. The y-axis represents the nuclei numbers and the x-axis represents the PI flu-orescence intensity that translates to the quantity of DNA per cell. Flow cytometric dot plot (B) shows fluorescencedue to width and area of the nuclei from epithelial cells isolated from eland sperm. The model used in the softwarefor determining the frequency of events in the cell cycle phases ensures aggregates are not counted, as in the arrowat G2/M.
used as donor karyoplasts compared with that ofbovine fibroblasts (93.7 and 80.0%, respectively).Development to morula and blastocyst stageswas similar for bovine NT embryos (38 and 21%,respectively) and PAOs (42.6 and 32.7%, respec-tively). Development to the morula stage, how-ever, was lower (p � 0.001) for eland igNT embryos (5%) and no eland igNT embryos de-veloped to the blastocyst stage.
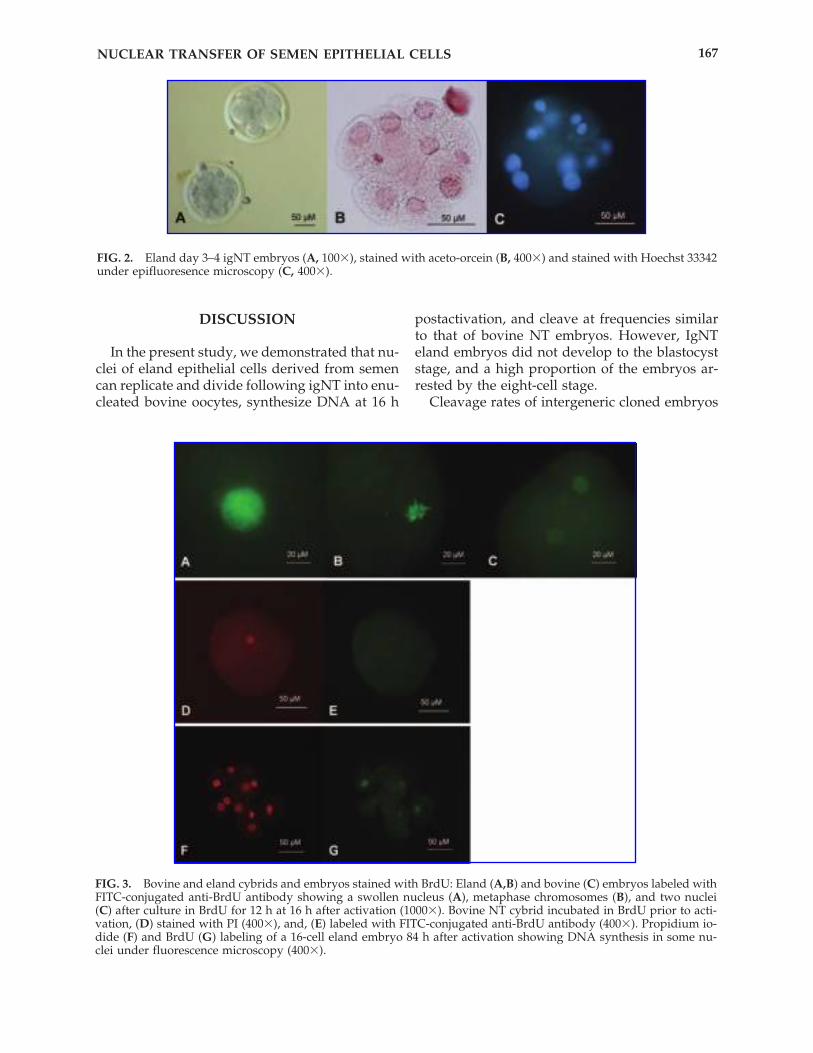
The nuclear stage of bovine and eland NT cy-brids, and of PAOs that were fixed 16 hpa, weresimilar. Swollen nuclei, metaphase chromo-somes, and cybrids containing two nuclei wereobserved (Fig. 3A–C). No significant differenceswere found between types of cybrids (Table 2).While 28% of eland igNT cybrids and 64% ofPAOs were still at the swollen nucleus stage at 16hpa, all of the bovine cybrids had progressed tometaphase or the two-nuclei stage at 16 hpa.
Of NT embryos that cleaved by 84 h (n � 31),17 (55%) of the bovine embryos had less thaneight cells and 14 (45%) had equal to or more thaneight cells, whereas 20 eland igNT embryos (n �23; 87%) had less than eight cells and three hadgreater than eight cells (13%; Table 2). Nonethe-less, the mean number of nuclei between bovineand eland NT embryos was similar. Bovine andeland embryos with less than eight cells had anaverage of 2.9 � 0.3 and 3.6 � 0.3 cells, respec-tively, and those with equal to or more than eightcells had 13.0 � 1.1 and 14.0 � 1.3 cells, respec-tively.
The mean total cell number in bovine NT blas-tocysts (76 � 6.1) was similar to that of blasto-cysts from PAOs (76 � 8.4). Although the mean
ICM cell numbers in bovine NT (22 � 3.9) andparthenogenetic blastocysts (153 � 0.8) were sim-ilar, the mean percentage of total cells in the ICMfrom total cell number per blastocyst were dif-ferent (29 vs. 18%; p � 0.05). The highest totalnumber of cells obtained in eland igNT embryoswas 23 cells.
DNA synthesis by BrdU detection
DNA synthesis during the first cell cycle wasevaluated as an indicator of interaction betweenthe bovine ooplasm and the eland donor nu-cleus after NT. BrdU incorporation was not de-tected in bovine (n � 27) and eland (n � 28) NTcybrids that were not subjected to activation af-ter NT, indicating that these cells did not passthrough the S-phase (Fig. 3D and E). However,when activated cybrids were exposed to BrdUat 4 hpa and then cultured for 12 h, 70% (21 of30) and 75% (12 of 16) of the eland and thebovine NT cybrids incorporated BrdU into theirnuclei, respectively (Fig. 3A–C). Similarly, 93%(14 of 15) of PAOs incorporated BrdU (Table 2).The percentage of nuclei that incorporatedBrdU in newly synthesized DNA did not differ(p � 0.05) among bovine NT embryos, elandigNT embryos, and PAOs.
The percentage of cells incorporating BrdU waslower in bovine (18%; 3 of 17) and eland (20%; 4of 20) NT embryos that had less than eight cells,than in bovine (100%; 14 of 14) and eland em-bryos (67%; 2 of 3; Fig. 3F and G) that had equalto or more than eight cells (p � 0.01 when cellnumbers within species were pooled).
NEL-THEMAAT ET AL.166
TABLE 1. DEVELOPMENT OF EMBRYOS PRODUCED BY NUCLEAR TRANSFER OF BOVINE FIBROBLASTS AND ELAND SEMEN-EPITHELIAL CELLS INTO ENUCLEATED BOVINE OOCYTES, AND PARTHENOGENETICALLY ACTIVATED BOVINE OOCYTES
Blastocysts D 7–9
COC’s Cleaved D3 Morula D7 D7-9 TPD Cell no. ICM Cell no.Donor cell (replicates) Fused couplets n (%)a n (%)b n (%) n (%) n (%)
Bovine 128 (3) 120 96 (80) 36 (38)c 20 (21)c 53 � 5.0 22 � 3.9fibroblasts (70 � 4.0) (29 � 4.0)
Eland 230 (4) 209 173 (83) 9 (5)d 0d — —semenepithelial
Parthenogenetic 75 (3) N/A 61 (81) 26 (43)c 20 (33)c 67 � 6.5 15 � 3.8activation (82 � 4.5) (18 � 4.7)
aPercentage from total fused couplets.bPercentage from total cleaved embryos.c,dDifferent superscripts within columns are significantly different (p � 0.001).Mean ICM % derived as mean of percentage of the total cell number (ICM � TPD) in each embryo.
DISCUSSION
In the present study, we demonstrated that nu-clei of eland epithelial cells derived from semencan replicate and divide following igNT into enu-cleated bovine oocytes, synthesize DNA at 16 h
postactivation, and cleave at frequencies similarto that of bovine NT embryos. However, IgNTeland embryos did not develop to the blastocyststage, and a high proportion of the embryos ar-rested by the eight-cell stage.
Cleavage rates of intergeneric cloned embryos
NUCLEAR TRANSFER OF SEMEN EPITHELIAL CELLS 167
FIG. 2. Eland day 3–4 igNT embryos (A, 100�), stained with aceto-orcein (B, 400�) and stained with Hoechst 33342under epifluoresence microscopy (C, 400�).
FIG. 3. Bovine and eland cybrids and embryos stained with BrdU: Eland (A,B) and bovine (C) embryos labeled withFITC-conjugated anti-BrdU antibody showing a swollen nucleus (A), metaphase chromosomes (B), and two nuclei(C) after culture in BrdU for 12 h at 16 h after activation (1000�). Bovine NT cybrid incubated in BrdU prior to acti-vation, (D) stained with PI (400�), and, (E) labeled with FITC-conjugated anti-BrdU antibody (400�). Propidium io-dide (F) and BrdU (G) labeling of a 16-cell eland embryo 84 h after activation showing DNA synthesis in some nu-clei under fluorescence microscopy (400�).
created by using bovine oocytes as recipient cy-toplasts were similar to the cleavage rates ob-served by intraspecies bovine NT (Dominko et al.,1999; Lu et al., 2005; Murakami et al., 2005). Inaddition, higher cleavage rates have been re-ported after transfer of sheep (66%), monkey(85.7%), and rat (90.2%) somatic cells into bovinecytoplasts when compared with cleavage rates re-sulting from the transfer of bovine cells (55.8%)(Dominko et al., 1999). Even though bovineoocytes can induce early nuclear remodeling,blastocyst development of IgNT embryos is usu-ally reduced when compared with that reportedafter intraspecies NT (Matshikiza et al., 2004).
In the present study, we observed that elandIgNT and intraspecies bovine NT embryoscleaved at similar frequencies (83 and 80%, re-spectively), further confirmation that bovineoocytes can interact with and induce some levelof nuclear reorganization of somatic cells from adifferent genus. However, eland IgNT embryosdid not develop to the blastocyst stage. Develop-ment to the blastocyst stage of cloned embryosreconstructed with somatic cells of other an-telopes, such as the bongo (21%; Tragelaphus eu-
ryceros) and the Giant eland (27%; Taurotragus der-bianus), with bovine cytoplasts (Damiani et al.,2003; Lee et al., 2003) have been reported. The fre-quency of blastocyst development (28%) ofcloned embryos reconstructed with saola (Pseudo-ryx nghetinhensis) donor cells was similar to thatof bovine NT (23%) (Bui et al., 2002). Althoughsaola is classified under the subfamily Bovinae,little is known about the phylogeny of thisspecies, and their relationship to domestic cattleis unclear. The bongo and Giant eland are mem-bers of the Bovinae family, and are relativelyclosely related to Common eland antelope (Tau-rotragus oryx). Interestingly, 2% (one embryo) of the Common eland IgNT produced embryosreached the blastocyst stage following transfer ofCommon eland fibroblasts into bovine cytoplasts(Matshikiza et al., 2004). Although comparisonsbetween experiments are confounded by varia-tions in the type of cells, embryo culture systemsand criteria used to determine development, apossible explanation for the contrasting results inblastocyst development of antelope NT embryoslikely are associated with species-specific differ-ences.
NEL-THEMAAT ET AL.168
TABLE 2. BRDU INCORPORATION OF BOVINE AND ELAND NONACTIVATED ACTIVATED AND CLEAVED NT CYBRIDS
I: Nonactivated NT cybrids
Bovine Eland
NT cybrids, n 27 28BrdU Incorporation 0 0
II: Activated NT cybrids (16 hpa)
Bovine Eland Parthenogenetic
NT cybrids, n 16 30 15BrdU incorporation 12/16 (75%) 21/30 (70%) 14/15 (93%)Swollen nucleus 0 6/21 (28%) 9/14 (64%)Metaphase 8/12 (66%) 7/21 (33%) 2/14 (14%)2-Cell 4/12 (33%) 8/21 (38%) 3/14 (21%)
III: Cleaved NT embryos (84 hpa)
Two to seven cells �8 cells
PI BrdU incorporation PI BrdU incorporation
Total Mean no. Mean no. Mean no. Mean no.embryosa Embryos nuclei Embryos nuclei Embryos nuclei Embryos nuclei
Species n n (%) � SEM n (%) � SEM n (%) � SEM n (%) � SEM
Bovine 31 17/31 (55) 2.9 � 0.3 3/30 (10) 3 � 0.6 14 (45) 13 � 1.1 14 (100) 12 � 1.0Eland 23 20/23 (87) 3.5 � 0.3 4/20 (20) 3 � 0.8 3 (13) 14 � 1.3 2 (67)0 14 � 2.0
aOnly embryos with at least two nuclei as indicated by PI staining.

Although live cloned calves have been pro-duced after transfer of embryos derived from ep-ithelial cells (Gong et al., 2004; Kishi et al., 2000;Miyashita et al., 2002), there are differences be-tween cell lines and cell types in developmentalpotential of the resulting cloned embryos(Batchelder et al., 2005; Kato et al., 2000; Servelyet al., 2003). For example, live offspring have beenproduced after the transfer of cloned NT embryosderived from mammary epithelial cells (http://www.d-b.net/dti/news/990427-calves-cloned-from-milk-cells.html), but embryos derived fromactively proliferating immortalized mammaryepithelial cells failed to develop to the blastocyststage (Zakhartchenko et al., 1999). The authorssuggested that chromatin modifications in somecell types may prevent nuclear reprogramming,and disruption of gene regulation caused by invitro culture also may be involved in failure ofembryo development. Gene expression patternsof cells change over time during in vitro culture,as exemplified by the report that cytokeratin andvimentin expression in epithelial and fibroblastcells was different in early compared with laterpassages (Zakhartchenko et al., 1999).
In the present study, we used epithelial cellsthat were isolated from eland semen and culturedin vitro for a prolonged period (three to five pas-sages). Although we do not know how in vitroculture conditions affected the donor nucleus,epigenetic modifications in the donor cells possi-bly influenced nuclear remodeling and repro-gramming of derived cloned embryos (Santos etal., 2003). Notably, blastocysts (8.3%) have beenobtained after transfer of the same line of elandepithelial cells into eland oocytes (Wirtu, 2004).For these reasons, we surmize that both epige-netic modifications of the somatic cell and speciesincompatibilities between the eland donor nu-cleus and bovine cytoplasm probably contributedto the development failure of the IgNT eland em-bryos.
In vitro-derived (IVF, ICSI) eland embryos ar-rested at the eight-cell stage regardless of the cul-ture medium used, and only one and three clonedblastocysts were obtained after culturing em-bryos in CR1aa medium supplemented with 5%eland serum or alpha-MEM supplemented with5% bovine calf serum, respectively (Wirtu, 2004).Therefore, it is possible that the medium used inthis study for culturing IgNT embryos affectedembryo development. In fact, culture medium isknown to affect in vitro development of cloned
embryos (Choi et al., 2002; Heindryckx et al.,2001). Bongo IgNT embryos developed to theblastocyst stage at higher rates when they werecultured for the first 3 days in hamster embryoculture medium (HECM-6) and the last 5 days inTCM-199 � 10% FBS, than when they were cul-tured in modified synthetic oviduct medium (m-SOF) (Lee et al., 2003). Therefore it is likely thatimprovements in the culture medium would af-fect development of IgNT eland embryos.
The coordination between the cell-cycle stageof the donor nucleus and the recipient oocyte isessential for successful development of recon-structed embryos (Campbell et al., 1993). Analy-sis of the cell-cycle distribution demonstrated thatthe majority (�82%) of eland epithelial cells werein G0/G1 phase, and thus, were compatible withnuclear transfer.
Mature oocytes contain high levels of matura-tion promoter factor (MPF), which is responsiblefor maintaining arrest of oocytes at metaphase-II.Proper activation of fused couplets is required toinduce the events that will result in initiation ofthe first cell-cycle. Results from the BrdU label-ing of cybrids demonstrated that DNA synthesisdid not take place before activation. Because mostof the transferred nuclei were in the G0/G1 phaseof the cycle, we can assume that the cell cycle ofthe transferred bovine or eland nucleus does notresume without oocyte activation. Accordingly,when IgNT eland and bovine NT cybrids wereexposed to BrdU after 4 h of activation, 70 and75%, respectively, of the cybrids synthesizedDNA, indicating that MPF activity in the oocytecytoplasm declined and that DNA synthesis dur-ing the first cell cycle was similar in both typesof embryos.
Alberio et al. (2001) reported that 80% of clonedbovine embryos synthesized DNA at 2 h after re-moval (5–7 hpa) of bohemine. Similarly, in sheepNT embryos, DNA synthesis was initiated �5hpa, peaked at �10 hpa, and completed at 18 hpa(Liu et al., 1997). Although, we did not evaluatethe exact time at which cloned embryos startedDNA synthesis, our results indicated that mostbovine NT and eland IgNT embryos synthesizedDNA by 16 hpa.
Evaluation of nuclear status (swollen nucleus,condensed chromatin or two nuclei) in clonedembryos that stained positive for BrdU revealedthat 28% of the eland IgNT embryos had aswollen nucleus, whereas all of the bovine NTembryos had progressed to at least the condensed
NUCLEAR TRANSFER OF SEMEN EPITHELIAL CELLS 169

chromatin stage. The number of embryos withtwo nuclei was similar in both eland (38%) andbovine (33%) NT embryos, suggesting that someeland IgNT embryos were delayed in develop-ment. However, these “delayed” embryos didsynthesize DNA, and the developmental dis-crepancy may have been a function of the differ-ence in cell types between the two species. It isunclear why the bovine PAOs, despite progress-ing through the S-phase within 16 h, progressedmore slowly through the different nuclear stagesthan did the nuclear transfer groups.
Most eland IgNT embryos arrested at thetwo- to seven-cell stages (87%) and only 20% ofthem entered DNA synthesis during the 12 hBrdU incubation. In contrast, 67% (2 of 3) of theeland IgNT embryos that progressed to theeight-cell stage by 84 hpa synthesized DNA inblastomeres. Also, 100% of the cells from equalto or more than eight-cell embryos in the bovineNT group stained positive for BrdU. Thus, al-though most eland IgNT embryos arrested ear-lier than bovine NT embryos, a few IgNT em-bryos continued dividing up to 84 hpa.Improvements of the in vitro culture system mayimprove the potential for blastocyst develop-ment of these embryos.
Potentially, nuclear transfer technology canhave a positive impact on endangered speciesconservation and the production of elite ani-mals. Semen-derived somatic cells could be par-ticularly valuable when semen from deceasedanimals is available but of limited supply. Byproducing cloned offspring, the genome of theanimal of interest could be introduced into thegene pool by natural breeding and, therefore,aid in reducing the negative consequences of in-breeding, a concern in zoo populations and in-tensively managed livestock breeds. Cloning ef-forts in endangered species often rely oninterspecies nuclear transfer because oocytesfrom the endangered animal are seldom avail-able. Thus, it is important to further examine themechanisms involved during nuclear remodel-ing and reprogramming after igNT to aid in un-derstanding factors that affect these processes.Although the early development of IgNT em-bryos appears to be similar initially to that of NTembryos, subsequent embryonic development isseverely impaired, especially after genomic ac-tivation. Nonetheless, with further research, theuse of semen-derived somatic cells for nucleartransfer holds promise as a method for aiding in
the conservation of endangered species by in-corporating otherwise inaccessible genetic ma-terial from previous generations.
ACKNOWLEDGMENTS
The authors would like to thank the staff ofFMA-SSC and Audubon Center for Research ofEndangered Species veterinary department forassisting in semen collections, as well as HydeMeat packers for supplying the ovaries. We alsothank Dr. Stanley Leibo for intellectual input.This article was approved for publication by theDirector of the Louisiana Agricultural Experi-mental Station as manuscript No. 07-18-0347. Thisresearch was supported, in part, by funds fromthe Audubon Center for Research of EndangeredSpecies, the Louisiana Agricultural ExperimentStation, and the Multistate Federal Regional Proj-ect No. 1171.
REFERENCES
Alberio, R., Brero, A., Motlik, J., et al. (2001). Remodelingof donor nuclei, DNA-synthesis, and ploidy of bovinecumulus cell nuclear transfer embryos: effect of activa-tion protocol. Mol. Reprod. Dev. 59, 371–379.
Baguisi, A., Behboodi, E., Melican, D.T., et al. (1999). Pro-duction of goats by somatic cell nuclear transfer. Nat.Biotechnol. 17, 456–461.
Batchelder, C.A., Hoffert, K.A., Bertolini, M., et al. (2005).Effect of the nuclear-donor cell lineage, type, and celldonor on development of somatic cell nuclear transferembryos in cattle. Cloning Stem Cells 7, 238–254.
Brem, G., and Kuhholzer, B. (2002). The recent history ofsomatic cloning in mammals. Cloning Stem Cells 4,57–63.
Bui, L.C., Vignon, X., Campion, E., et al. (2002). Use of in-terspecies nuclear transfer to study the early embryonicdevelopment and nuclear activities of the endangeredspecies Pseudoryx nghetinhensis (saola). Theriogenology57, 427.
Campbell, K.H., Ritchie, W.A., and Wilmut, I. (1993). Nu-clear–cytoplasmic interactions during the first cell cy-cle of nuclear transfer reconstructed bovine embryos:implications for deoxyribonucleic acid replication anddevelopment. Biol. Reprod. 49, 933–942.
Campbell, K.H.S., Alberio, R., Choi, I., et al. (2005).Cloning: eight years after Dolly. Reprod. Domest.Anim. 40, 256–268.
Choi, Y.H., Lee, B.C., Lim, J.M., et al. (2002). Optimiza-tion of culture medium for cloned bovine embryos andits influence on pregnancy and delivery outcome. The-riogenology 58, 1187–1197.
NEL-THEMAAT ET AL.170

Crissman, H.A., and Steinkamp, J.A. (1973). Rapid, si-multaneous measurement of DNA, protein, and cellvolume in single cells from large mammalian cell pop-ulations. J. Cell Biol. 59, 766–771.
Damiani, P., Wirtu, G., Miller, F., et al. (2003). Develop-ment of giant eland (Taurotragus oryx) and bovine (Bos taurus) oocytes. Theriogenology 58, 390.
Dominko, T., Mitalipova, M., Haley, B., et al. (1999).Bovine oocyte cytoplasm supports development of em-bryos produced by nuclear transfer of somatic cell nu-clei from various mammalian species. Biol. Reprod. 60,1496–1502.
Galli, C., Duchi, R., Moor, R.M., et al. (1999). Mammalianleukocytes contain all the genetic information necessaryfor the development of a new individual. Cloning 1,161–170.
Galli, C., Lagutina, I., Crotti, G., et al. (2003). Pregnancy:a cloned horse born to its dam twin. Nature 424, 635.
Gómez, M.C., Pope, E., Harris, R., et al. (2003). Develop-ment of in vitro matured, in vitro fertilized domesticcat embryos following cryopreservation, culture andtransfer. Theriogenology 60, 239–251.
Gómez, M.C., Pope, C.E., Giraldo, A., et al. (2004). Birthof African wildcat cloned kittens born from domesticcats. Cloning Stem Cells 6, 247–258.
Gong, G., Dai, Y., Zhu, H., et al. (2004). Generation ofcloned calves from different types of somatic cells. Sci.China C Life Sci. 47, 470–476.
Hamilton, H.M., Peura, T.T., Laurincik, J., et al. (2004).Ovine ooplasm directs initial nucleolar assembly in em-bryos cloned from ovine, bovine, and porcine cells. Mol.Reprod. Dev. 69, 117–125.
Heindryckx, B., Rybouchkin, A., Van der, E.J., et al. (2001).Effect of culture media on in vitro development ofcloned mouse embryos. Cloning 3, 41–50.
Janssen, D.L., Edwards, M.L., Koster, J.A., et al. (2004).Postnatal management of chryptorchid banteng calvescloned by nuclear transfer utilizing frozen fibroblastcultures and enucleated cow ova. Reprod. Fertil. Dev.16, 224.
Kato, Y., Tani, T., Sotomaru, Y., et al. (1998). Eight calvescloned from somatic cells of a single adult. Science 282,2095–2098.
Kato, Y., Tani, T., and Tsunoda, Y. (2000). Cloning ofcalves from various somatic cell types of male and fe-male adult, newborn and fetal cows. J. Reprod. Fertil.120, 231–237.
Kishi, M., Itagaki, Y., Takakura, R., et al. (2000). Nucleartransfer in cattle using colostrum-derived mammarygland epithelial cells and ear-derived fibroblast cells.Theriogenology 54, 675–684.
Lanza, R.P., Cibelli, J.B., Diaz, F., et al. (2000). Cloning ofan endangered species (Bos gaurus) using interspeciesnuclear transfer. Cloning 2, 79–90.
Lee, B., Wirtu, G.G., Damiani, P., et al. (2003). Blastocystdevelopment after intergeneric nuclear transfer ofmountain bongo antelope somatic cells into bovineoocytes. Cloning Stem Cells 5, 25–33.
Liu, L., Dai, Y., and Moor, R.M. (1997). Nuclear transferin sheep embryos: the effect of cell-cycle coordination
between nucleus and cytoplasm and the use of in vitromatured oocytes. Mol. Reprod. Dev. 47, 255–264.
Loi, P., Ptak, G., Barboni, B., et al. (2001). Genetic rescueof an endangered mammal by cross-species nucleartransfer using post-mortem somatic cells. Nat. Biotech-nol. 19, 962–964.
Lu, F., Shi, D., Wei, J., et al. (2005). Development of em-bryos reconstructed by interspecies nuclear transfer ofadult fibroblasts between buffalo (Bubalus bubalis) andcattle (Bos indicus). Theriogenology 64, 1309–1319.
Matshikiza, M., Bartels, P., Vajta, G., et al. (2004). Embryodevelopment following interspecies nuclear transfer ofAfrican buffalo ( Syncerus caffer ), bontebok ( Damalis-cus dorcus dorcus ) and eland ( Taurotragus oryx ) somaticcells into bovine cytoplasts. Reprod. Fertil. Dev. 16,150–151.
Miyashita, N., Shiga, K., Yonai, M., et al. (2002). Remark-able differences in telomere lengths among cloned cat-tle derived from different cell types. Biol. Reprod. 66,1649–1655.
Murakami, M., Otoi, T., Wongsrikeao, P., et al. (2005). De-velopment of interspecies cloned embryos in yak anddog. Cloning Stem Cells 7, 77–81.
Nel-Themaat, L., Gómez, M.C., Wirtu, G., et al. (2005). So-matic cell isolation from semen by percoll gradients.Reprod. Fert. Dev. 17, 314–315.
Nel-Themaat, L., Gómez, M.C., Damiani, P., et al. (2007).Isolation, culture and characterization of somatic cellsderived from semen and milk of endangered sheep andeland antelope. Reprod. Fertil. Dev. 19, 576–584.
Polejaeva, I.A., Chen, S.H., Vaught, T.D., et al. (2000).Cloned pigs produced by nuclear transfer from adultsomatic cells. Nature 407, 86–90.
Ryder, O.A., and Benirschke, K. (1997). The potential useof “cloning” in the conservation effort. Zoo Biol. 16,295–300.
Sansinena, M.J., Hylan, D., Hebert, K., et al. (2005). Ban-teng (Bos javanicus) embryos and pregnancies producedby interspecies nuclear transfer. Theriogenology 63,1081–1091.
Santos, F., Zakhartchenko, V., Stojkovic, M., et al. (2003).Epigenetic marking correlates with developmental po-tential in cloned bovine preimplantation embryos.Curr. Biol. 13, 1116–1121.
Servely, J.L., Le Bourhis, D., Freret, S., et al. (2003). Com-parison of developmental potential of keratinocytesand fibroblasts after nuclear transfer in bovine. Therio-genology 59, 288.
Ty, L.V., Hahn, N.V., Uoc, N.T., et al. (2003). Preliminaryresults of cell cryobanking and embryo production ofblack bear (Ursus thibetanus) by interspecies somatic cellnuclear transfer. Theriogenology 59, 290.
Wayakama, T., Perry, A.C., Zuccotti, M., et al. (1998). Full-term development of mice from enucleated oocytes in-jected with cumulus cell nuclei. Nature 394, 369–374.
Wilmut, I., Schnieke, A.E., McWhir, J., et al. (1997). Viableoffspring derived from fetal and adult mammaliancells. Nature 385, 810–813.
Wirtu, G. (2004). Developing embryo technologies for theeland antelope (Taurotragus oryx) Louisiana State
NUCLEAR TRANSFER OF SEMEN EPITHELIAL CELLS 171

University and Agricultural and Mechanical College.Thesis.
Wirtu, G., Pope, C.E., Lee, B.C., et al. (2002). Collection ofsemen by rectal massage from an eland (Taurotragusoryx) bull. Proceedings of the 2nd International Symposiumon Assisted Reproductive Technologies for the Conservationand Genetic Management of Wildlife, September 28–29,2002, pp. 318–320.
Wirtu, G., Pope, C.E., Maclean, R.A., et al. (2008). Rever-sal of motility loss in bongo antelope (Tragelaphus eu-rycerus isaaci) spermatozoa contaminated with hypos-motic urine during electroejaculation. Anim. Reprod.Sci. 103, 392–397.
Zakhartchenko, V., Alberio, R., Stojkovic, M., et al. (1999).Adult cloning in cattle: potential of nuclei from a per-manent cell line and from primary cultures. Mol. Re-prod. Dev. 54, 264–272.
Address reprint requests to:Dr. Kenneth Bondioli
School of Animal SciencesLSU Agricultural CenterBaton Rouge, LA 70803
E-mail: [email protected]
NEL-THEMAAT ET AL.172