staphylococcal colonization of e-beam patterned surfaces
TRANSCRIPT

lable at ScienceDirect
Biomaterials xxx (2014) 1e7
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
Conditions of lateral surface confinement that promote tissue-cellintegration and inhibit biofilm growth
Yi Wang a,b,1, Joana F. da Silva Domingues b,1, Guruprakash Subbiahdoss b,Henny C. van der Mei b, Henk J. Busscher b, Matthew Libera a,*
aDepartment of Chemical Engineering and Materials Science, Stevens Institute of Technology, Hoboken, NJ, USAbUniversity of Groningen, University Medical Center Groningen, Department of Biomedical Engineering, Antonius Deusinglaan 1, 9713 AV Groningen,The Netherlands
a r t i c l e i n f o
Article history:Received 29 January 2014Accepted 21 March 2014Available online xxx
Keywords:BiofilmCell adhesionInfectionMicropatterningOsteoblastPolyethylene oxide
* Corresponding author. Tel.: þ1 201 216 5259.E-mail address: [email protected] (M. Libera).
1 These authors contributed equally to this work.
http://dx.doi.org/10.1016/j.biomaterials.2014.03.0570142-9612/� 2014 Elsevier Ltd. All rights reserved.
Please cite this article in press as: Wang Y,biofilm growth, Biomaterials (2014), http://d
a b s t r a c t
Surfaces with cell adhesiveness modulated at micro length scales can exploit differences between tissue/bacterial cell size, membrane/wall plasticity, and adhesion mechanisms to differentially control tissue-cell/material and bacteria/material interactions. This study explores the short-term interactions ofStaphylococcus aureus and osteoblast-like cells with surfaces consisting of cell-adhesive circular patches(1e5 mm diameter) separated by non-adhesive electron-beam patterned poly(ethylene glycol) hydrogelthin films at inter-patch distances of 0.5e10 mm. Osteoblast-like U2OS cells both bind to and spread onthe modulated surfaces, in some cases when the cell-adhesive area comprises only 9% of the total surfaceand in several cases at least as well as on the continuously adhesive control surfaces. In contrast, S. aureusadhesion rates are 7e20 times less on the modulated surfaces than on the control surfaces. Furthermore,the proliferation of those bacteria that do adhere is inhibited by the lateral confinement imposed by thenon-adhesive boundaries surrounding each patch. These findings suggest a new approach to createbiomaterial surfaces that may promote healing while simultaneously reducing the probability ofinfection.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
The underlying objective of any tissue-contacting biomedicaldevice is to promote healing after injury or disease. Such devicesare designed to interact with tissue cells in a variety of ways,including supporting tissue-cell adhesion and tissue integration.The scientific and clinical communities have, however, longrecognized that implantable devices provide foreign surfaces towhich bacteria can adhere and colonize. Hence, in addition to theirprimary healing function, tissue-contacting biomedical devices alsobring a heightened probability of introducing a chronic infectionwith potentially severe consequences to the patient. An importantchallenge facing next-generation biomaterials is thus to preserve orenhance the ability of a device to facilitate desirable tissue in-teractions while simultaneously inhibiting bacterial colonization[1,2].
et al., Conditions of lateral sx.doi.org/10.1016/j.biomater
Many approaches have been explored to inhibit the bacterialcolonization of synthetic implant surfaces. Foremost among theseare antifouling surface coatings that resist protein and cell adhe-sion, such as poly(ethylene glycol) [PEG]. PEGylated surfaces, forexample, have been extensively studied [3e5], and a number ofanalogous gel-like coatings are increasingly being explored. Sur-faces with micro/nano-patterned topography have also beenshown to hinder bacterial adhesion and alter biofilm development[6e9], and topographic modulations at even smaller length scalescan force individual bacteria to adopt positions and orientationsdefined by the surface patterning [10]. While successful at reducingbacterial colonization, tissue-cell interactions with these samesurfaces are either simultaneously inhibited or have not beenstudied.
Surfaces with adhesiveness laterally modulated over micro-scopic length scales present an alternate between the extremes offully adhesive or fully non-adhesive surfaces with which to differ-entially control cell/material and bacteria/material interactions.There is good evidence that tissue cells are able to adhere to andspread on such modulated surfaces. Chen et al. [11], for example,showed that individual endothelial cells are able to bind to a
urface confinement that promote tissue-cell integration and inhibitials.2014.03.057

Y. Wang et al. / Biomaterials xxx (2014) 1e72
surface modulated at sub-cellular length scales with adhesivepatches patterned on otherwise non-adhesive surface. The cellsstraddle the non-adhesive surface areas and bind to one or moreadhesive patches depending on the patch size and spacing. Spatzet al. [12e14] have done similar experiments that explore fibroblastadhesion and spreading on surfaces with square adhesive patcheshaving edge sizes of 0.1e3 mmand patch spacings equal to the patchsize. Malmstrom et al. [15] used colloidal lithography to patternsurfaces with randomly distributed adhesive patches ranging indiameter from 0.1 to 1.0 mm and showed that mammalian breastcancer cells can adhere to them all by bridging across the non-adhesive surface. The adhesion is less effective as the patch diam-eter decreases, an effect they attribute to a disruption of focaladhesion formation.
Less is known about how bacteria respond to surfaces withlaterally modulated adhesiveness. Wang et al. [16,17] have recentlystudied surfaces whose adhesiveness is modulated by submicronnon-adhesive microgels and showed that bacterial adhesion isreduced as the surface concentration of microgels increases. At theopposite extreme, cell-adhesive patches patterned on an otherwisenon-adhesive surface introduce a form of spatial confinement thatcan both localize where bacteria adhere as well as limit the physicalsize of growing colonies [18]. Furthermore, the fact that adjacentbut separately growing bacterial colonies are able to communicatevia the diffusion of quorum-sensing agents [19,20] indicates thatthe proximity of growing colonies on a surface may influence boththeir individual and collective development into biofilms. The ideathat spatial confinement influences biofilm development has beenfurther reinforced by recent experiments, which demonstratequorum-sensing behavior in individual bacteria when they areconfined within small droplets of growth medium on a surface[21e23].
The question of how micro-patterned cell adhesiveness affectsthe surface interactions both with tissue cells and with bacteriaraises the hypothesis that there is a window of patch sizes andpatch spacings that promotes tissue-cell interactions and also in-hibits bacterial colonization. This study thus reports on experi-ments that test how tissue cells and staphylococcal bacteria interactwith surfaces that have circular adhesive patches with diameters of1, 2, 3, or 5 mm patterned within otherwise non-adhesive surface.
2. Materials and methods
2.1. Surface patterning
Microgel patterned glass slides were prepared using established procedures[24,25]. Glass microscope slides were sonicated in ethanol (96%), cleaned in aPiranha solution (3:1 98% sulfuric acid and 30% H2O2) for 30 min, rinsed withdeionized water, dried, and exposed to low-pressure O2 plasma (w300 mTorr,1.75 W) for 10 min. The slides were then silanized with 2% [v/v] vinyl-methoxysiloxane homopolymer (Gelest Inc., Morrisville, PA) in ethanol for 10 min, rinsed,dried, and baked at 110 �C for 2 h. After cooling, thin films of PEG (w100 nm thick)were spin cast onto these substrata using a solution of 2 wt% PEG (6 kDa; Fluka) intetrahydrofuran. Samples were stored under vacuum (w10�3 Torr) until they wereused for patterning. E-beam patterning used a Zeiss Auriga Scanning Electron Mi-croscope (SEM) with a Schottky field-emission electron source with a point dose of10 fC and an incident electron energy of 2 keV. Such irradiation conditions convertthe PEG homopolymer precursor into microgels grafted to the underlying substrate[25]. The e-beam position and dwell timewere controlled using a Nabity NanometerPattern Generation System. Individual PEG microgels with diameters of about400 nm were created by single point irradiation, and the spacing between adjacentirradiation points was set to 50 nm so individual microgels would overlap to formcontinuous thin-film gels. After patterning, the slides were rinsed in deionizedwaterfor 30 min to remove unirradiated PEG and expose the silanized glass in the unir-radiated areas.
Arrays of adhesive patches were patterned on an otherwise non-adhesive sur-face by creating areas of PEG microgel films containing circular patches of exposedsilanized glass. Patch diameters, a, of 1, 2, 3 or 5 mmwith inter-patch (patch-edge topatch-edge) distances, b, of a/2, a and 2a were studied. These two parameters werecombined to create surfaces with adhesive area fractions, c, of 0.09, 0.20, and 0.35where the adhesive area fraction is defined as:
Please cite this article in press as: Wang Y, et al., Conditions of lateral sbiofilm growth, Biomaterials (2014), http://dx.doi.org/10.1016/j.biomater
c ¼ p�a2
�2
ðaþ bÞ2(1)
Twelve 200 mm � 200 mm patch arrays were patterned on a single glass slide,with the arrays separated from each other by 100 mm-wide strips of silanized glass.After patterning, substrata were stored under vacuum (w10�3 Torr). Light micro-scopic images of the arrays were taken using a Nikon Eclipse E1000 upright opticalmicroscope. The final cross-linked PEG film thickness was determined by atomicforce microscopy (AFM; Bruker Bioscope Catalyst).
Fibronectin (Fn) was adsorbed onto exposed silanized glass surfaces byimmersing the patterned slides in a 25 mg/mL solution of human Fn (SigmaeAldrichBV, Zwijndrecht, The Netherlands) in phosphate buffered saline (PBS, 10 mM po-tassium phosphate, 0.15 M NaCl, pH 7.0) for 30 min at room temperature and thenwashed three times with PBS. To confirm the preferential Fn adsorption on theexposed silanized glass, a slide was immersed in PBS containing 1% BSA for 1 min toblock non-specific protein adsorption, rinsed three times in PBS, exposed to a pri-mary antibody (rabbit-anti-human fibronectin Ab, polyclonal, dilution 1:400 in PBS)for 30 min, rinsed, and then exposed to a fluorescent secondary antibody (FITC-conjugated donkey-anti-rabbit IgG, dilution 1:100 in PBS). After a final rinse, thepatterned glass slide was placed in a petri dish filled with PBS and examined, whilefully hydrated, by confocal laser scanning microscopy (CLSM, Leica DMRXE withconfocal TCS SP2 unit). Fn-treated silanized glass is henceforth referred to heresimply as glass unless otherwise indicated.
2.2. Tissue-cell culture
U2OS osteosarcoma cells were cultured in Dulbecco’s Modified Eagle’s Medium(DMEM)-low glucose supplemented with 10% fetal bovine serum (FBS) and 0.2 mM
ascorbic acid-2-phosphate (AA2P), denoted as DMEM þ FBS. U2OS cells weremaintained in tissue culture polystyrene flasks (TCPS, Greiner Bio-One) at 37 �C inhumidified air with 5% CO2 and harvested at 90% confluence using trypsine/ethyl-enediamineetetra-acetic acid. The harvested cells were diluted to 6 � 105 cells/mLin DMEM þ FBS. U2OS is an immortalized human cell line derived from osteosar-coma cells and was chosen from a broad selection of human osteoblastic cell linesused previously [26]. It has been demonstrated that osteosarcoma cell lines exhibitmeaningful osteoblastic phenotypes [27].
U2OS cell adhesion and spreading on patterned surfaces were studied by in situdigital phase-contrast microscopy (Olympus BH-2; 10� objective) in a parallel-plateflow chamber (175� 17 � 0.75 mm3) and by ex situ immunofluorescence imaging.The flow chamber was maintained at 37 �C throughout the experiments. Once fullyfilled and free of air-bubbles, a U2OS cell suspensionwas introduced in the chamber.Flow was stopped for 1.5 h to allow the cells to settle and adhere. Phase-contrastimages were taken at this time point from each patterned array and also from theunpatterned glass to determine the initial surface coverage of adhered cells.DMEM þ FBS supplemented with 2% HEPES buffer was then perfused through thechamber at 0.14 s�1 shear rate. After 48 h of additional culture beyond the 1.5 hsettling period the substrata were removed and fixed in 3.7% formaldehyde incytoskeleton stabilization buffer (CS; 0.1 M Pipes, 1 mM EGTA, 4% (w/v) PEG 8000, pH6.9). After fixation, samples were incubated in 0.5% Triton X-100 for 3 min, rinsedwith PBS, and stained for 30 min with 4 mg/mL of DAPI and 2 mg/mL of TRITCePhalloidin. Slides were examined by fluorescence microscopy (Leica DM4000B).Each of these experiments was performed in triplicate. The total surface coverage ofadhering cells on the patterned surfaces was determined by Scion image software.The surface coverage during the spreading period was defined as:
F ¼ f48 � f1:5f1:5
(2)
where fi is the area fraction of surface covered by U2OS cells after 1.5 or 48 h. Apositive value ofF indicates that the cells spread. A negative value ofF indicates thatthe cells either contracted or, more likely, migrated off the patterned surface.
2.3. Bacterial culture
Staphylococcus aureus NCTC 8325-4 were provided by T.J. Foster (Moyne Insti-tute of Preventive Medicine, Dublin, Ireland). This strain possesses fibronectin-binding proteins (FnBPs). Bacteria were inoculated on trypsine soy broth (TSB,Oxoid) agar plates and incubated overnight at 37 �C. One colony was grown in 10 mLTSB overnight with constant rotation (120 rpm) and subsequently used to inoculate190 mL TSB. After 2 h of incubation, when S. aureusNCTC 8325-4 was expected to bein the exponential phase of growth with the peak of FnBPs expression, bacteriawereharvested by centrifugation (6500 g, 5 min, 10 �C) and washed twice in sterile PBS.Bacterial aggregates were broken by mild, intermittent sonication on ice (3 times,10 s, 30 W, Wibra Cell model 375, Sonics and Materials Inc., Danbury, CT, USA) andre-suspended to a concentration of 3 � 108 bacteria/mL in PBS. Bacterial adhesion topatterned glass slides was studied in a parallel-plate flow chamber andmonitored insitu by digital phase-contrast microscopy. After removing air bubbles in the tubingby flowing PBS, the S. aureus suspension was perfused through the chamber (shearrate of 11 s�1) for 30 min at room temperature. After bacterial deposition, sterile PBSwas flowed through the system (shear rate of 11 s�1) for 30 min to remove non-
urface confinement that promote tissue-cell integration and inhibitials.2014.03.057
= 1 µm = 1 µm
= 5 µm= 5 µm
β
β
α
00 5 10 15 20 25
50100
Distance (µm)Hei
ght (
nm)
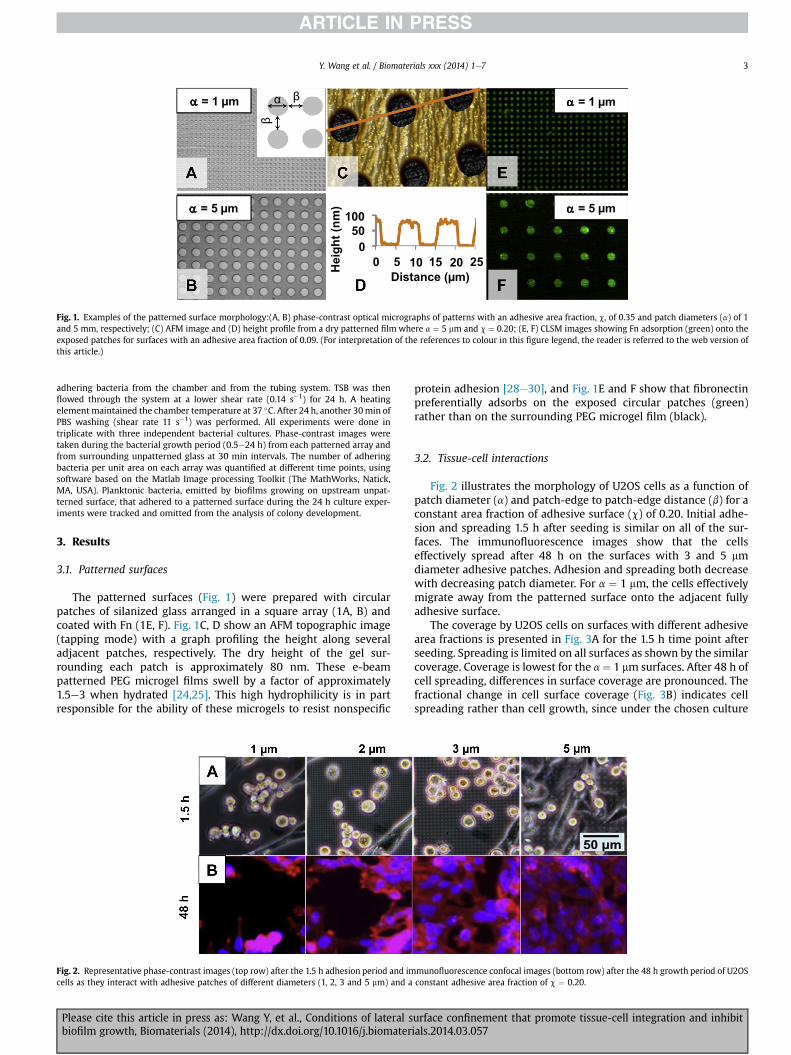
Fig. 1. Examples of the patterned surface morphology:(A, B) phase-contrast optical micrographs of patterns with an adhesive area fraction, c, of 0.35 and patch diameters (a) of 1and 5 mm, respectively; (C) AFM image and (D) height profile from a dry patterned film where a ¼ 5 mm and c ¼ 0.20; (E, F) CLSM images showing Fn adsorption (green) onto theexposed patches for surfaces with an adhesive area fraction of 0.09. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version ofthis article.)
Y. Wang et al. / Biomaterials xxx (2014) 1e7 3
adhering bacteria from the chamber and from the tubing system. TSB was thenflowed through the system at a lower shear rate (0.14 s�1) for 24 h. A heatingelementmaintained the chamber temperature at 37 �C. After 24 h, another 30min ofPBS washing (shear rate 11 s�1) was performed. All experiments were done intriplicate with three independent bacterial cultures. Phase-contrast images weretaken during the bacterial growth period (0.5e24 h) from each patterned array andfrom surrounding unpatterned glass at 30 min intervals. The number of adheringbacteria per unit area on each array was quantified at different time points, usingsoftware based on the Matlab Image processing Toolkit (The MathWorks, Natick,MA, USA). Planktonic bacteria, emitted by biofilms growing on upstream unpat-terned surface, that adhered to a patterned surface during the 24 h culture exper-iments were tracked and omitted from the analysis of colony development.
3. Results
3.1. Patterned surfaces
The patterned surfaces (Fig. 1) were prepared with circularpatches of silanized glass arranged in a square array (1A, B) andcoated with Fn (1E, F). Fig. 1C, D show an AFM topographic image(tapping mode) with a graph profiling the height along severaladjacent patches, respectively. The dry height of the gel sur-rounding each patch is approximately 80 nm. These e-beampatterned PEG microgel films swell by a factor of approximately1.5e3 when hydrated [24,25]. This high hydrophilicity is in partresponsible for the ability of these microgels to resist nonspecific
Fig. 2. Representative phase-contrast images (top row) after the 1.5 h adhesion period and imcells as they interact with adhesive patches of different diameters (1, 2, 3 and 5 mm) and a
Please cite this article in press as: Wang Y, et al., Conditions of lateral sbiofilm growth, Biomaterials (2014), http://dx.doi.org/10.1016/j.biomater
protein adhesion [28e30], and Fig. 1E and F show that fibronectinpreferentially adsorbs on the exposed circular patches (green)rather than on the surrounding PEG microgel film (black).
3.2. Tissue-cell interactions
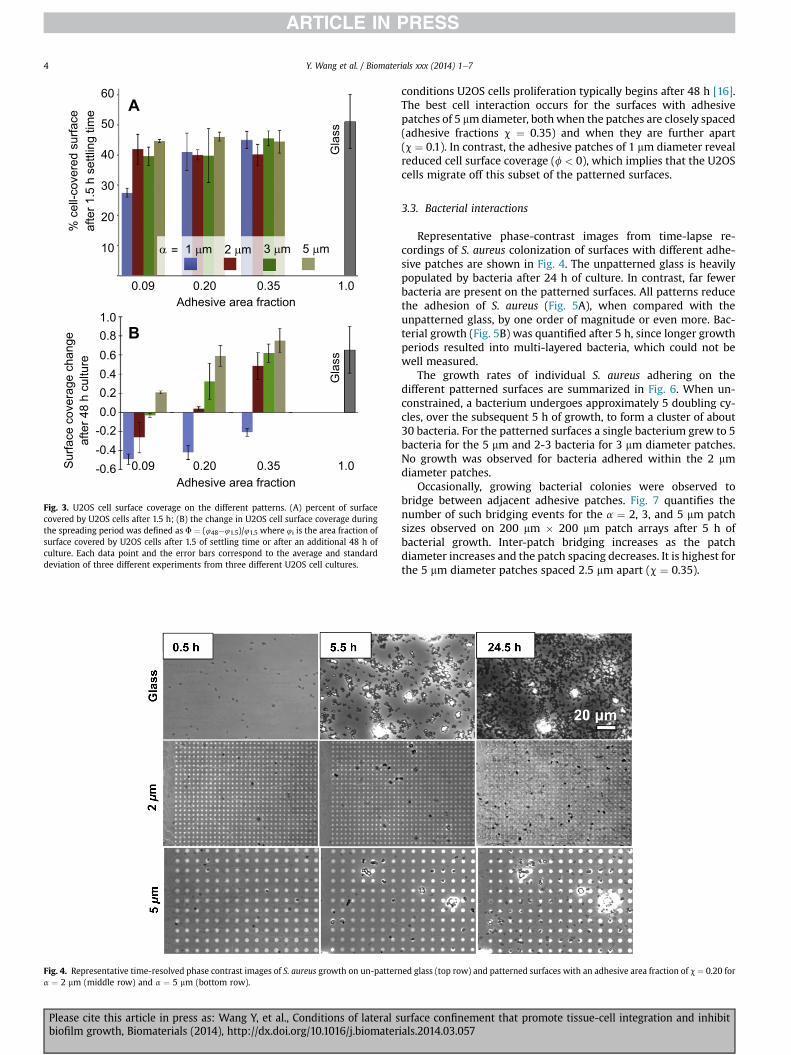
Fig. 2 illustrates the morphology of U2OS cells as a function ofpatch diameter (a) and patch-edge to patch-edge distance (b) for aconstant area fraction of adhesive surface (c) of 0.20. Initial adhe-sion and spreading 1.5 h after seeding is similar on all of the sur-faces. The immunofluorescence images show that the cellseffectively spread after 48 h on the surfaces with 3 and 5 mmdiameter adhesive patches. Adhesion and spreading both decreasewith decreasing patch diameter. For a ¼ 1 mm, the cells effectivelymigrate away from the patterned surface onto the adjacent fullyadhesive surface.
The coverage by U2OS cells on surfaces with different adhesivearea fractions is presented in Fig. 3A for the 1.5 h time point afterseeding. Spreading is limited on all surfaces as shown by the similarcoverage. Coverage is lowest for the a¼ 1 mm surfaces. After 48 h ofcell spreading, differences in surface coverage are pronounced. Thefractional change in cell surface coverage (Fig. 3B) indicates cellspreading rather than cell growth, since under the chosen culture
munofluorescence confocal images (bottom row) after the 48 h growth period of U2OSconstant adhesive area fraction of c ¼ 0.20.
urface confinement that promote tissue-cell integration and inhibitials.2014.03.057
1.00.80.60.40.20.0
-0.2-0.4-0.6S
urfa
ce c
over
age
chan
geaf
ter 4
8 h
cultu
re
0.09 0.20 0.35 1.0
0.09 0.20 0.35 1.0
Adhesive area fraction
Adhesive area fraction
Gla
ssG
lass
10
20
30
40
50
60
% c
ell-c
over
ed s
urfa
ceaf
ter 1
.5 h
set
tling
tim
e
α = 1 μm 2 μm 3 μm 5 μm
A
B
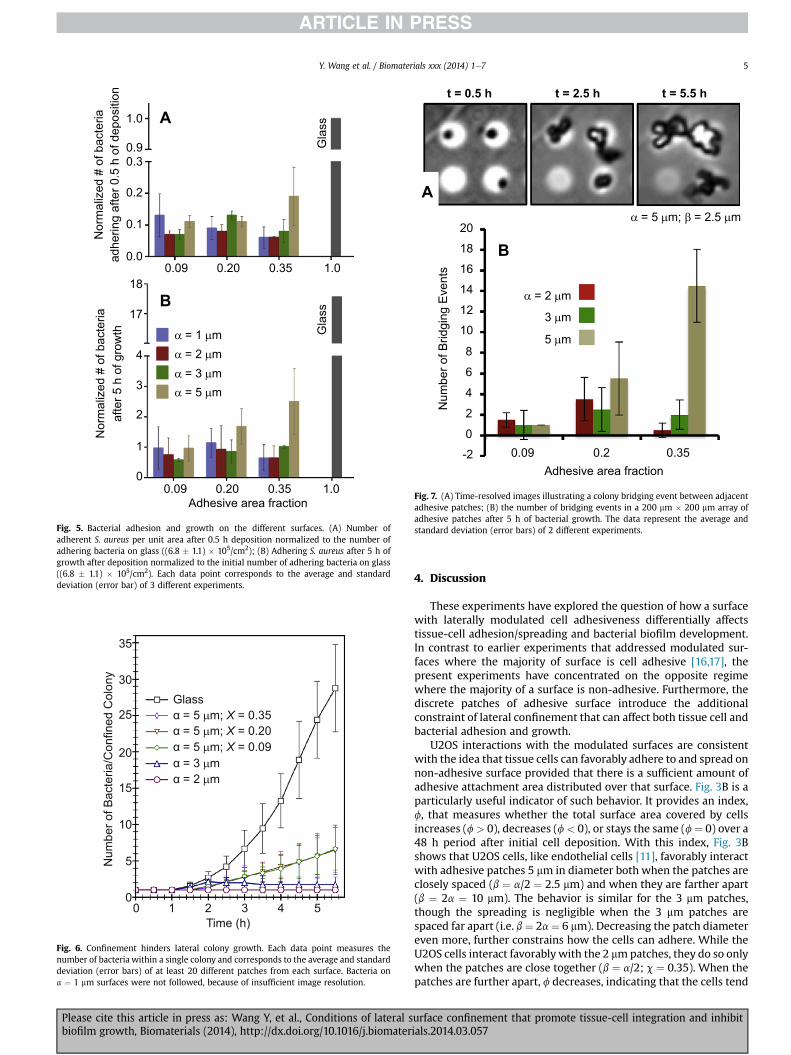
Fig. 3. U2OS cell surface coverage on the different patterns. (A) percent of surfacecovered by U2OS cells after 1.5 h; (B) the change in U2OS cell surface coverage duringthe spreading period was defined as F ¼ (448�41.5)/41.5 where 4i is the area fraction ofsurface covered by U2OS cells after 1.5 of settling time or after an additional 48 h ofculture. Each data point and the error bars correspond to the average and standarddeviation of three different experiments from three different U2OS cell cultures.
Fig. 4. Representative time-resolved phase contrast images of S. aureus growth on un-patterna ¼ 2 mm (middle row) and a ¼ 5 mm (bottom row).
Y. Wang et al. / Biomaterials xxx (2014) 1e74
Please cite this article in press as: Wang Y, et al., Conditions of lateral sbiofilm growth, Biomaterials (2014), http://dx.doi.org/10.1016/j.biomater
conditions U2OS cells proliferation typically begins after 48 h [16].The best cell interaction occurs for the surfaces with adhesivepatches of 5 mmdiameter, both when the patches are closely spaced(adhesive fractions c ¼ 0.35) and when they are further apart(c ¼ 0.1). In contrast, the adhesive patches of 1 mm diameter revealreduced cell surface coverage (f < 0), which implies that the U2OScells migrate off this subset of the patterned surfaces.
3.3. Bacterial interactions
Representative phase-contrast images from time-lapse re-cordings of S. aureus colonization of surfaces with different adhe-sive patches are shown in Fig. 4. The unpatterned glass is heavilypopulated by bacteria after 24 h of culture. In contrast, far fewerbacteria are present on the patterned surfaces. All patterns reducethe adhesion of S. aureus (Fig. 5A), when compared with theunpatterned glass, by one order of magnitude or even more. Bac-terial growth (Fig. 5B) was quantified after 5 h, since longer growthperiods resulted into multi-layered bacteria, which could not bewell measured.
The growth rates of individual S. aureus adhering on thedifferent patterned surfaces are summarized in Fig. 6. When un-constrained, a bacterium undergoes approximately 5 doubling cy-cles, over the subsequent 5 h of growth, to form a cluster of about30 bacteria. For the patterned surfaces a single bacterium grew to 5bacteria for the 5 mm and 2-3 bacteria for 3 mm diameter patches.No growth was observed for bacteria adhered within the 2 mmdiameter patches.
Occasionally, growing bacterial colonies were observed tobridge between adjacent adhesive patches. Fig. 7 quantifies thenumber of such bridging events for the a ¼ 2, 3, and 5 mm patchsizes observed on 200 mm � 200 mm patch arrays after 5 h ofbacterial growth. Inter-patch bridging increases as the patchdiameter increases and the patch spacing decreases. It is highest forthe 5 mm diameter patches spaced 2.5 mm apart (c ¼ 0.35).
ed glass (top row) and patterned surfaces with an adhesive area fraction of c ¼ 0.20 for
urface confinement that promote tissue-cell integration and inhibitials.2014.03.057
0
1
2
3
4
17
18
0.0
0.09 0.20 0.35 1.0
0.09 0.20 0.35 1.0
0.1
0.2
0.30.9
1.0
Gla
ssG
lass
Adhesive area fraction
Nor
mal
ized
# o
f bac
teria
Nor
mal
ized
# o
f bac
teria
afte
r 5 h
of g
row
thad
herin
g af
ter 0
.5 h
of d
epos
ition
α = 1 μmα = 2 μmα = 3 μmα = 5 μm
B
A
Fig. 5. Bacterial adhesion and growth on the different surfaces. (A) Number ofadherent S. aureus per unit area after 0.5 h deposition normalized to the number ofadhering bacteria on glass ((6.8 � 1.1) � 105/cm2); (B) Adhering S. aureus after 5 h ofgrowth after deposition normalized to the initial number of adhering bacteria on glass((6.8 � 1.1) � 105/cm2). Each data point corresponds to the average and standarddeviation (error bar) of 3 different experiments.
0 1 2 3 4 50
5
10
15
20
25
30
35
Time (h)
Num
ber o
f Bac
teria
/Con
fined
Col
ony
Glassα = 5 μm; X = 0.35α = 5 μm; X = 0.20α = 5 μm; X = 0.09α = 3 μmα = 2 μm
Fig. 6. Confinement hinders lateral colony growth. Each data point measures thenumber of bacteria within a single colony and corresponds to the average and standarddeviation (error bars) of at least 20 different patches from each surface. Bacteria ona ¼ 1 mm surfaces were not followed, because of insufficient image resolution.
-2
0
2
4
6
8
10
12
14
16
18
20
0.09 0.350.2Adhesive area fraction
Num
ber o
f Brid
ging
Eve
nts
α = 2 μm
α = 5 μm; β = 2.5 μm
3 μm
5 μm
B
A
t = 0.5 h t = 2.5 h t = 5.5 h
Fig. 7. (A) Time-resolved images illustrating a colony bridging event between adjacentadhesive patches; (B) the number of bridging events in a 200 mm � 200 mm array ofadhesive patches after 5 h of bacterial growth. The data represent the average andstandard deviation (error bars) of 2 different experiments.
Y. Wang et al. / Biomaterials xxx (2014) 1e7 5
Please cite this article in press as: Wang Y, et al., Conditions of lateral sbiofilm growth, Biomaterials (2014), http://dx.doi.org/10.1016/j.biomater
4. Discussion
These experiments have explored the question of how a surfacewith laterally modulated cell adhesiveness differentially affectstissue-cell adhesion/spreading and bacterial biofilm development.In contrast to earlier experiments that addressed modulated sur-faces where the majority of surface is cell adhesive [16,17], thepresent experiments have concentrated on the opposite regimewhere the majority of a surface is non-adhesive. Furthermore, thediscrete patches of adhesive surface introduce the additionalconstraint of lateral confinement that can affect both tissue cell andbacterial adhesion and growth.
U2OS interactions with the modulated surfaces are consistentwith the idea that tissue cells can favorably adhere to and spread onnon-adhesive surface provided that there is a sufficient amount ofadhesive attachment area distributed over that surface. Fig. 3B is aparticularly useful indicator of such behavior. It provides an index,f, that measures whether the total surface area covered by cellsincreases (f> 0), decreases (f< 0), or stays the same (f¼ 0) over a48 h period after initial cell deposition. With this index, Fig. 3Bshows that U2OS cells, like endothelial cells [11], favorably interactwith adhesive patches 5 mm in diameter both when the patches areclosely spaced (b ¼ a/2 ¼ 2.5 mm) and when they are farther apart(b ¼ 2a ¼ 10 mm). The behavior is similar for the 3 mm patches,though the spreading is negligible when the 3 mm patches arespaced far apart (i.e. b¼ 2a¼ 6 mm). Decreasing the patch diametereven more, further constrains how the cells can adhere. While theU2OS cells interact favorably with the 2 mmpatches, they do so onlywhen the patches are close together (b ¼ a/2; c ¼ 0.35). When thepatches are further apart, f decreases, indicating that the cells tend
urface confinement that promote tissue-cell integration and inhibitials.2014.03.057

Y. Wang et al. / Biomaterials xxx (2014) 1e76
to migrate away from the patterned surface and onto the adjacentunpatterned glass. Such migration is observed for all three caseswhere a ¼ 1 mm, indicating that these surfaces are unable to sup-port U2OS adhesion and spreading.
The findings involving smaller patch diameters are less consis-tent with published reports on cell adhesion to nanopatternedsurfaces. Spatz et al. [31], for example, have shown that fibroblastscan adhere to adhesive patches as small as 100 nm in size whenthey are closely spaced. In the present experiments, however, thetopography introduced by the PEG microgel thin films may hindercell adhesionwhen the adhesive patch size is small and the patchesare close together. Since the hydrated PEG microgel thin films arew120 nm thick, changes in membrane curvature, including in-flection points, would be required at the edge of each patch in orderfor a cell to adhere to one adhesive patch and bridge across the PEGto an adjacent adhesive patch. The total amount of such curvedmembrane would increase with decreasing patch size. When thepatch size falls below some critical dimension (e.g. 1e2 mm), theadded membrane curvature may create a morphology that isenergetically unfavorable for adhesion. This observation furthersuggests that adhesive patches which slightly protrude above sur-rounding non-adhesive area may be more effective at promotingcell adhesion than ones that lie slightly below the surrounding non-adhesive area.
Because of their size and their non-conformable cell walls,staphylococci should interact with patterned surfaces differentlythan tissue cells. Indeed, all of the patterns studied here reduce theadhesion of S. aureus relative to unpatterned glass by at least afactor of seven or more (Fig. 5A). Importantly, the bacterial adhe-sion rates do not scale with the patch area. This finding indicatesthat the bacteria do not adhere immediately upon contact to anadhesive patch. Because of the confinement imposed by the size ofthe adhesive patch, bacteria that are close to the adhesive surfacemust adhere within a finite time window determined by theculture-medium flow rate and by the patch size. In the event thatstable binding has not occurred by the time the flowing bacteriumencounters the far end of an adhesive patch, the bacterium will beforced to move away from the surface. In contrast, on an unpat-terned surface, the bacteriumwill have an almost unlimited time tocreate a binding event. Relatively large adhesive patches that areclosely spaced together would be more susceptible to bacterialadhesion, and the fact that the a ¼ 5 mm, b ¼ a/2 (c ¼ 0.35) surfacehas a slightly higher adhesion rate than the other surfaces (Fig. 5A)supports this hypothesis.
Those bacteria that stably adhere to an adhesive patch aresubjected to additional confinement effects that hinder their sub-sequent growth. The growth rates for individual colonies (Fig. 6)confined on 2, 3, and 5 mm diameter patches are all substantiallylower than unconstrained colonies. In fact, 2 and 3 mm diameterpatches stop the lateral colony growth altogether. This, too, is sig-nificant from the point of view of inhibiting infection, because itsuggests that the development of a growing bacterial colony into amature biofilm can be inhibited by spatial confinement. Arrestingbiofilm formation may then prevent the development of hierar-chical structure and varying phenotypes, including persisters,which are involved in the well-known resistance of biofilms toantimicrobials [32,33].
The lateral confinement effects on bacterial growth can beovercome if a growing colony is able to bridge from one adhesivepatch to another. This behavior is illustrated by Fig. 7A and can alsobe seen in some of the colonies on the 5 mm patches in Fig. 4. Thebridging statistics (Fig. 7B) indicate that colonies growing onclosely spaced, larger patches bridge more frequently than those onsmaller patches or those on widely separated patches. Sometimesthe bridging occurs between patches that are both occupied with
Please cite this article in press as: Wang Y, et al., Conditions of lateral sbiofilm growth, Biomaterials (2014), http://dx.doi.org/10.1016/j.biomater
growing colonies (e.g. Fig. 7A), suggesting that there may be somesort of communication between these colonies such as the diffusionof quorum-sensing agents. However, bridging can also occur froman isolated colony to an adjacent unoccupied patch. While bridgingoccurs in a relatively small fraction of patches (e.g. w10% and w1%for the 5 mm patches on the c ¼ 0.35 and c ¼ 0.20 surfaces,respectively, and much less for the other surfaces), it is a mecha-nism by which a colony can grow despite the underlying non-adhesive PEG surface.
From a practical point of view, the results of these experimentsindicate that laterally modulated surfaces can be structured in or-der to preserve, or possibly even enhance, tissue-cell interactionswhile reducing bacterial colonization. Both the bacterial adhesionrate as well as the bacterial lateral growth rate are substantiallylower on micro-patterned surfaces than on unpatterned controlsurfaces. Nevertheless, on many of these same surfaces, tissue cellsare able to adhere and spread in a manner comparable to that on anunpatterned surface. These results thus suggest that patterning thesurface of a tissue-contacting biomedical device may be able to notonly promote healing but may also help reduce the probability ofthat device developing a biomaterials-associated infection. In thespecific case of an orthopedic application, these results suggest thatthere is a window of surfaces with adhesive patches on the order of2e5 mm separated by distances greater than the patch diameterthat can promote osteoblast interactions and simultaneouslyreduce staphylococcal colonization.
5. Conclusion
The relative effects of lateral confinement on bacterial coloni-zation and tissue-cell integration were studied by modulating thediameter and spacing between micro-scale circular cell-adhesivepatches (1, 2, 3, and 5 mm) separated by continuous PEG thinfilms such that the fraction of adhesive surface area was 0.09, 0.20,or 0.35. Osteoblast-like U2OS cells can adhere to and spread onmany of the patterned surfaces despite the fact that the majority ofthe surface is non-adhesive. In contrast, under the bacterial depo-sition conditions used here, the rate of S. aureus adhesion onto anyof these patterned surfaces is about an order of magnitude less onthe patterned surfaces than that on fully adhesive surfaces. There isalso little or no dependence of bacterial adhesion on patch size,suggesting that an adhesive patch of 5 mmdiameter or less providesinsufficient interaction between a flowing bacterium and the sur-face before that bacterium encounters non-adhesive surface at theedge of the patch and is repelled from the surface. Those bacteriathat adhere within an adhesive patch develop over the 5 h cultureperiod studied within the present experiments into multicellularcolonies at a rate much less than those growing in an unconfinedway on a fully adhesive surface, and the diameter of a confinedcolony grows to about the diameter of the underlying adhesivepatch. The lateral confinement of a bacterial colony can be over-come if the spacing between adjacent adhesive patches is smallenough to enable bridging across the non-adhesive surface be-tween the patches. When surface interactions with both tissue cellsand with bacteria are considered, the results of these experimentsindicate that there is a window of adhesive patch diameters (w2 to5 mm) and patch spacings (1e2� the patch diameter) that canpromote tissue-cell interactions while simultaneously inhibitingbacterial adhesion and biofilm development.
Acknowledgments
This project was partially supported by the U.S. Army ResearchOffice via grant #W911NF-12-1-0331. H.J. Busscher is also directorof a consulting company, SASA BV (GN Schutterlaan 4, 9797 PC
urface confinement that promote tissue-cell integration and inhibitials.2014.03.057

Y. Wang et al. / Biomaterials xxx (2014) 1e7 7
Thesinge, The Netherlands). The authors declare no potential con-flicts of interest with respect to authorship and/or publication ofthis article. Opinions and assertions contained herein are those ofthe authors and are not construed as necessarily representingviews of the funding organization or their respective employers.
References
[1] Busscher HJ, Van der Mei HC, Subbiahdoss G, Jutte PC, Van Den Dungen JJAM,Zaat SAJ, et al. Biomaterial-associated infection: locating the finish line in therace for the surface. Sci Transl Med 2012;4:153RV10.
[2] Gristina AG. Biomaterial-centered infection: microbial adhesion versus tissueintegration. Science 1987;237:1588e95.
[3] Nejadnik MR, Van der Mei HC, Norde W, Busscher HJ. Bacterial adhesion andgrowth on a polymer brush-coating. Biomaterials 2008;29:4117e21.
[4] Park KD, Kim YS, Han DK, Kim YH, Lee EH, Suh H, et al. Bacterial adhesion onPEG modified polyurethane surfaces. Biomaterials 1998;19:851e9.
[5] Kingshott P, Wei J, Bagge-Ravn D, Gadegaard N, Gram L. Covalent attachmentof poly(ethylene glycol) to surfaces, critical for reducing bacterial adhesion.Langmuir 2003;19:6912e21.
[6] Ivanova EP, Hasan J, Webb HK, Gervinskas G, Juodkazis S, Truong VK, et al.Bactericidal activity of black silicon. Nat Commun 2013/11/26;4. online.
[7] Kappell GM, Grover JP, Chrzanowski TH. Micro-scale surface-patterning in-fluences biofilm formation. Electron J Biotechnol 2009;12:1215.
[8] Magin CM, Cooper SP, Brennan AB. Non-toxic antifouling strategies. MaterToday 2010;13:36e44.
[9] Schumacher JF, Carman ML, Estes TG, Feinberg AW, Wilson LH, Callow ME,et al. Engineered antifouling microtopographies e effect of feature size, ge-ometry, and roughness on settlement of zoospores of the green alga Ulva.Biofouling 2007;23:55e62.
[10] Hochbaum AI, Aizenberg J. Bacteria pattern spontaneously on periodicnanostructure arrays. Nano Lett 2010;10:3717e21.
[11] Chen CS, Mrksich M, Huang S, Whitesides GM, Ingber DE. Geometric control ofcell life and death. Science 1997;276:1425e8.
[12] Arnold M, Schwieder M, Blummel J, Cavalcanti-Adam EA, Lopez-Garcia M,Kessler H, et al. Cell interactions with hierarchically structured nano-patterned adhesive surfaces. Soft Matter 2009;5:72e7.
[13] Cavalcanti-Adam EA, Volberg T, Micoulet A, Kessler H, Geiger B, Spatz JP. Cellspreading and focal adhesion dynamics are regulated by spacing of integrinligands. Biophys J 2007;92:2964e74.
[14] Glass R, Arnold M, Cavalcanti-Adam EA, Blummel J, Haferkemper C, Dodd C,et al. Block copolymer micelle nanolithography on non-conductive substrates.New J Phys 2004;6:1e17.
[15] Malmstrom J, Christensen B, Jakobsen HP, Lovmand J, Foldbjerg R,Sorensen ES, et al. Large area protein patterning reveals nanoscale control offocal adhesion development. Nano Lett 2010;10:686e94.
Please cite this article in press as: Wang Y, et al., Conditions of lateral sbiofilm growth, Biomaterials (2014), http://dx.doi.org/10.1016/j.biomater
[16] Wang Y, Subbiahdoss G, Swartjes J, Van der Mei HC, Busscher HJ, Libera M.Length-scale mediated differential adhesion of mammalian cells and mi-crobes. Adv Funct Mater 2011;21:3916e23.
[17] Wang Q, Uzunoglu E, Wu Y, Libera M. Self-assembled poly(ethylene glycol)-co- acrylic acid microgels to inhibit bacterial colonization of synthetic sur-faces. ACS Appl Mater Interfaces 2012;4:2498e506.
[18] Weibel DB, DiLuzio WR, Whitesides GM. Microfabrication meets microbi-ology. Nat Rev Microbiol 2007;5:209e18.
[19] Hyun JK, Boedicker JQ, Jang WC, Ismagilov RF. Defined spatial structure sta-bilizes a synthetic multispecies bacterial community. Proc Natl Acad Sci U S A2008;105:18188e93.
[20] Flickinger ST, Copeland MF, Downes EM, Braasch AT, Tuson HH, Eun YJ, et al.Quorum sensing between Pseudomonas aeruginosa biofilms accelerates cellgrowth. J Am Chem Soc 2011;133:5966e75.
[21] Carnes EC, Lopez DM, Donegan NP, Cheung A, Gresham H, Timmins GS, et al.Confinement-induced quorum sensing of individual Staphylococcus aureusbacteria. Nat Chem Biol 2010;6:41e5.
[22] Boedicker JQ, Vincent ME, Ismagilov RF. Microfluidic confinement of singlecells of bacteria in small volumes initiates high-density behavior of quorumsensing and growth and reveals its variability. Angew Chem Int Ed 2009;48:5908e11.
[23] Vincent ME, Liu W, Haney EB, Ismagilov RF. Microfluidic stochastic confine-ment enhances analysis of rare cells by isolating cells and creating highdensity environments for control of diffusible signals. Chem Soc Rev 2010;39:974e84.
[24] Krsko P, Mansfield M, Sukhishvili S, Clancy R, Libera M. Electron-beampatterned poly(ethylene glycol) microhydrogels. Langmuir 2003;19:5618e25.
[25] Wang Y, Firlar E, Dai X, Libera M. Poly(ethylene glycol) as a biointeractiveelectron-beam resist. J Polym Sci Part B: Polym Phys 2013;51:1543e54.
[26] De Ruijter JE, Ter Brugge PJ, Dieudonne SC, Van Vliet SJ, Torensma R, Jansen JA.Analysis of integrin expression in U2OS cells cultured on various calciumphosphate ceramic substrates. Tissue Eng 2001;7:279e89.
[27] Clover J, Gowen M. Are MG-63 and HOS TE85 human osteosarcoma cell linesrepresentative models of the osteoblastic phenotype? Bone 1994;15:585e91.
[28] Cole MA, Voelcker NH, Thissen H, Griesser HJ. Stimuli-responsive interfacesand systems for the control of protein-surface and cell-surface interactions.Biomaterials 2009;30:1827e50.
[29] Rabe M, Verdes D, Seeger S. Understanding protein adsorption phenomena atsolid surfaces. Adv Colloid Interface Sci 2011;162:87e106.
[30] Zhang M, Desai T, Ferrari M. Proteins and cells on PEG immobilized siliconsurfaces. Biomaterials 1998;19:953e60.
[31] Cavalcanti-Adam EA, Aydin D, Hirschfeld-Warneken VC, Spatz JP. Cell adhe-sion and response to synthetic nanopatterned environments by steering re-ceptor clustering and spatial location. HFSP J 2008;2:276e85.
[32] Keren I, Kaldalu N, Spoering A, Wang Y, Lewis K. Persister cells and toleranceto antimicrobials. FEMS Microbiol Lett 2004;230:13e8.
[33] Lewis K. Persister cells: molecular mechanisms related to antibiotic tolerance.Handb Exp Pharmacol; 2012:121e33.
urface confinement that promote tissue-cell integration and inhibitials.2014.03.057