staphylococcal toxins and sudden infant death syndrome
TRANSCRIPT

76 Clin Pathol 1992;45:716-721
Staphylococcal toxins and sudden infant deathsyndrome
J E Malam, G F Carrick, D R Telford, J A Morris
AbstractAims: To investigate the hypothesis thatcommonly occurring bacterial toxinscause sudden infant death syndrome(SIDS) by (1), determining in which tis-sues bacterial toxins are concentratedafter intravenous injection in rats; and (2),seeing if the same tissues contain detect-able toxins in cases of SIDS.Methods: The tissue distribution of intra-venously injected staphylococcal entero-toxin A (SEA), enterotoxin B (SEB),enterotoxin C (SEC), enterotoxin D(SED), toxic shock syndrome toxin(TSST-1), and a-haemolysin was studiedin rats using immunohistology and poly-acrylamide gel electrophoresis withimmunoblotting. Immunostaining wasalso carried out on formalin fixed kidneysfrom cases of SIDS and a comparisonseries of necropsy cases using anti-SEA,anti-SEB, anti-SEC2 and anti-SED.Results: Immunohistology showed thatSEB, SEC, SED and TSST-1 were allconcentrated in the proximal convolutedtubular cells ofthe kidney. The presence ofthese toxins was confirmed in kidneyhomogenates using electrophoresis andimmunoblotting. There was positive gran-ular staining in the proximal convolutedtubular cells of the kidney in 36% of SIDScases and 12% of the comparison serieswith anti SEC2 (X2 = 6; p < 0.025).Conclusion: SEC, or a bacterial toxin withepitopes in common, could have a patho-genic role in SIDS.
Department ofPathology, LancasterMoor Hospital,Lancaster LAI 3JRJ E MalamG F CarrickD RTelfordJ A MorrisCorrespondence to:Dr J A Morris
Accepted for publication5 February 1992
The idea that commonly occurring bacterialtoxins have a pathogenic role in some cases ofsudden infant death syndrome (SIDS) is gain-ing support. It is consistent with the epidemio-logical features of SIDS, including the winterexcess of cases, the characteristic age distribu-tion, and the association with a prone sleepingposition.' - There is also experimental supportfrom a number of studies of the naso-pharyngeal4 and gut flora,5'- and from animalmodels of bacterially induced suddendeath.80
In a recent immunohistological study weshowed that staphylococcal toxic shock syn-drome toxin (TSST-1) is concentrated in theproximal convoluted tubular cells of the kidneyin rats following intravenous injection." Thesame antiserum gave positive staining in theproximal convoluted tubular cells of humankidney in 18% of SIDS cases. We now describe
the results of work designed to extend theseobservations.
MethodsTOXIN PREPARATIONS
Samples of Staphylococcus aureus from thenasopharynges of babies attending a well babyclinic were phage typed and tested for toxinproduction at the Food Hygiene Laboratory,Central Public Health Laboratory, Colindale.The methods of obtaining the samples and theresults obtained have been published else-where.4 Bacteria which produced one or moreof the enterotoxins A, B, C, D and TSST-1were used for subsequent study. Alpha-haemo-lysin was obtained from Wood 46 strain pur-chased from the Central Public HealthLaboratory.Bead cultures stored at - 70'C were grown
on blood agar overnight at 37'C, then trans-ferred to bijou bottles containing 5 ml brain-heart infusion (BHI) medium, and again leftovernight at 37'C. Two drops of the culturewere spread on dialysis membrane overlyingBHI-1 5% agar plates and incubated for threeto four days at 37'C in 5% CO2. Phosphatebuffered saline (PBS) (2 ml) was used to washthe growth from the membrane and the sus-pension was centrifuged at 4000 x g for 15minutes. The supernatant was sterilised bypassing through a 0 2 ,um Minisart disposablefilter and stored at - 20°C.
ELECTROPHORESIS AND IMMUNOBLOTIINGToxin preparations were examined by sodiumdodecyl sulphate polyacrylamide gel electro-phoresis (SDS-PAGE), using a 1-5 mm slabgel containing 12% (WY) acrylamide for theseparating gel and a 4% acrylamide for thestacking gel. The buffer system was a slightlymodified version of Laemmli. 12 The samplebuffer contained 2% SDS, 10% sucrose,002 M TRIS-HCI (pH 8 0), 0-002 M EDTAand 2 mg Pyronin Y as marker dye. (Mercap-to-ethanol was added to the sample buffer formost toxins but was found to break down SEBinto small fractions.) Equal quantities of sam-ple and sample buffer were used at25 ,um/well.Electrophoresis was carried out in"The Sturdier" vertical slab unit, SE 400(Hoefer Scientific Instruments).Immediately after electrophoresis the sepa-
rating gels were soaked in transfer buffer(0-25 MTRIS, 0-192 M glycine in 20% meth-anol) for 15 minutes before blotting. Theseparated proteins were transferred to nitro-cellulose membrane for one hour at 100 mA,
716
group.bmj.com on July 11, 2011 - Published by jcp.bmj.comDownloaded from
Staphylococcal toxins and SIDS
using the TE 70 Semiphor semi-dry transferunit (Hoefer Scientific Instruments). Aftertransfer the nitrocellulose membrane wasimmersed in PBS-5% normal swine serum(NSS) overnight at 4°C. The following day themembrane was incubated in turn with therelevant polyclonal rabbit antitoxin at 1 in1000 in PBS-5% NSS for one hour at 37°C,swine anti-rabbit antibody at 1 in 500 PBS-5%NSS for one hour at 37°C, and horseradishperoxidase complexed with rabbit anti-perox-idase at 1 in 1000 in PBS-5% NSS for onehour at 37°C. Between each incubation themembrane was washed three times for 10minutes in PBS. The final washes were 10minutes each in PBS, distilled water, and0-05 M acetate buffer (pH 5 0), followed byvisualisation of the antigen and complexedantibody with 0-06 g 3-amino9-ethyl carbazole(AEC) in 15 ml dimethyl formamide diluted to300 ml with acetate buffer plus 5 drops of30%hydrogen peroxide. The primary antisera usedwere directed against staphylococcal entero-toxins A, B., CI, C2, C3, D a-haemolysin andTSST-1 (Toxin Technology, Florida, USA).Comparison gels were stained with Coomassieblue.
ANIMAL TISSUE PREPARATION
Female Wistar rats weighing 240-330 g wereanaesthetised by intraperitoneal injection ofNembutal (0-75 ml/100 g body weight of asolution of 10% Nembutal in 10% ethylalcohol). The toxin preparation from a singleorganism, which varied in volume from1 0-1 5 ml was given by slow intravenousinjection into the tail vein of one test animal.An equal volume of sterile saline was injectedinto control animals. A total of 42 toxinpreparations were tested with at least five
repetitions of each toxin. (Some preparationscontained more than one toxin). The rats wereobserved either until death or until the begin-ning of recovery from the anaesthetic. The ratswere then sacrificed by cervical dislocation.Urine samples were collected if possible. Dis-section was carried out immediately, andsamples of kidney, heart, lung, liver and spleenwere obtained. From some animals, a portionof gut, skin and thymus was also removed. Aportion of each sample was stored frozen at- 20°C and the rest fixed in 10% formalin.
IMMUNOSTAINING OF TISSUESImmunostaining was performed on formalinfixed, paraffin wax embedded blocks. A stan-dard avidin-biotin peroxidase technique wasused. The appropriate polyclonal rabbit anti-sera at a dilution of 1 in 300 for anti-B andanti-C2, 1 in 150 for anti-A and anti-D and 1in 240 for a-haemolysin was applied for threehours, followed by biotinylated swine anti-rabbit antibody at a dilution of 1 in 400 for 30minutes, then avidin-biotin complex for 30minutes. The antigen and complexed antibodywere visualised with AEC, as previously descri-bed. For each test animal and its paired controlthe primary antisera were appropriate to thetoxin given. In addition, a selection of posi-tively stained kidneys were tested with allantisera to check for cross-reaction. In all casesthe sections from both test and control animalswere stained in duplicate with one acting as anegative control in which specific antiserumwas replaced by normal rabbit serum (immu-noglobulin fraction) at a similar protein dilu-tion.
Selected tissues from the test and controlrats were examined by electrophoresis andimmunoblotting as described above. Approxi-
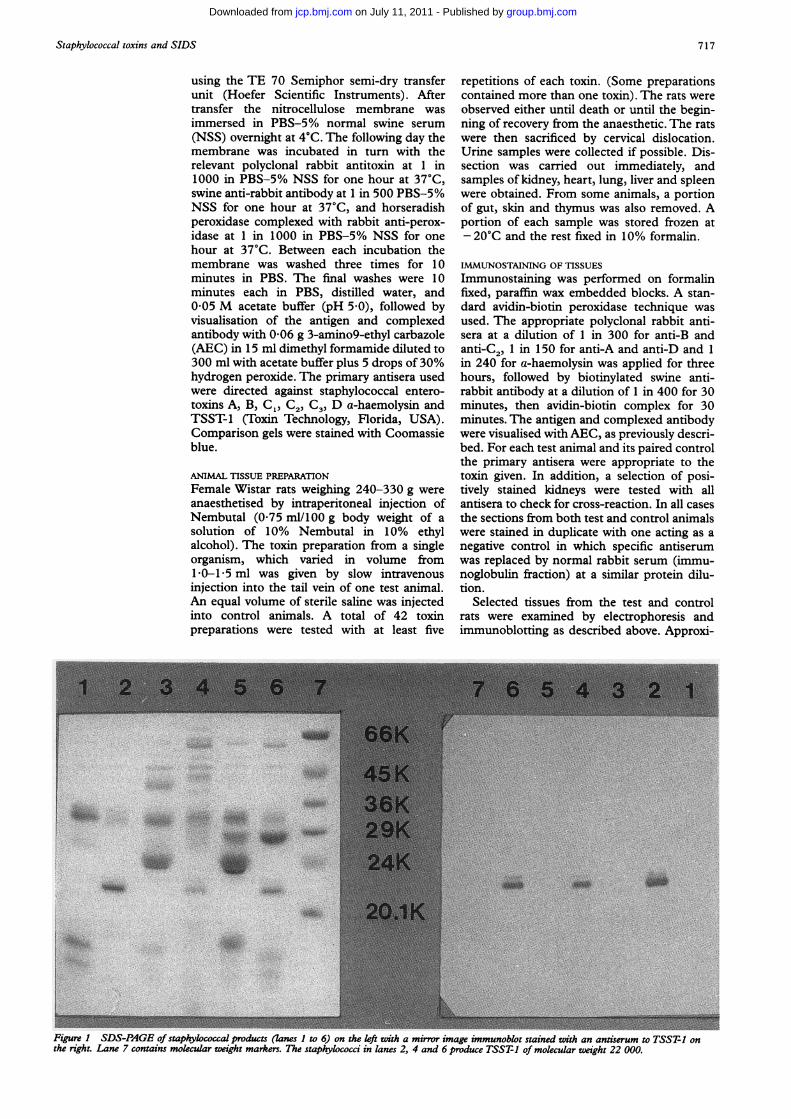
Figure I SDS-PAGE of staphylococcal products (lanes 1 to 6) on the left with a mirror image immunoblot stained with an antiserum to TSST-1 onthe right. Lane 7 contains molecular weight markers. The staphylococci in lanes 2, 4 and 6 produce TSST-1 of molecular weight 22 000.
717
.... .7
group.bmj.com on July 11, 2011 - Published by jcp.bmj.comDownloaded from
Malam, Carrick, Telford, Morris
mately 1 g of tissue was frozen, slowly thawedand ground. PBS (2 ml) were added and thefreezing, thawing, and grinding repeated up tothree times. The resulting suspension wascentrifuged at 4000 x g for 30 minutes andthe supernatant filtered through a 0-2 ,um filterand stored at - 20°C.
ResultsELECTROPHORESIS OF BACTERIAL PRODUCTS
The mixtures of bacterial toxins produced bygrowth of staphylococci on cellophane overagar were examined by SDS-PAGE followedby immunoblotting (fig 1). There was clearseparation of the polypeptides secreted by thestaphylococci. The TSST-1 band could easilybe identified by its position which correspondsto a molecular weight of 22 000 daltons. Thetoxins, SEA, SEB, SEC, and SED have molec-ular weights in the range 26 000-30 000, as domany other products of the bacteria, thereforethey could not be distinguished by theirposition alone. During the course of the work itwas found that incubating the bacteria in acO2 enriched atmosphere increased toxin pro-duction, as assessed by staining intensity of thebands on electrophoresis.
Polypeptide bands on electrophoresis
Antisera 35000 SEA SEB SEC SED 24000 TSST-I
anti-SEA - + ± ± ± ianti-SEB - +++ ++ ++anti SEC, + + + +anti-SEC2 - - ++ + +anti-SEC, - - + + + ++anti-SED - + i + + + +anti-TSST-1 - + + +anti-a-haemolysin - + ++
Reaction between antisera and polypeptide bands as assessed by immunoblotting followingpolyacrylamide gel electrophoresis+ + + is a strong reaction, - is a negative reaction.
..A*@ ...
I ll -5...
Xe
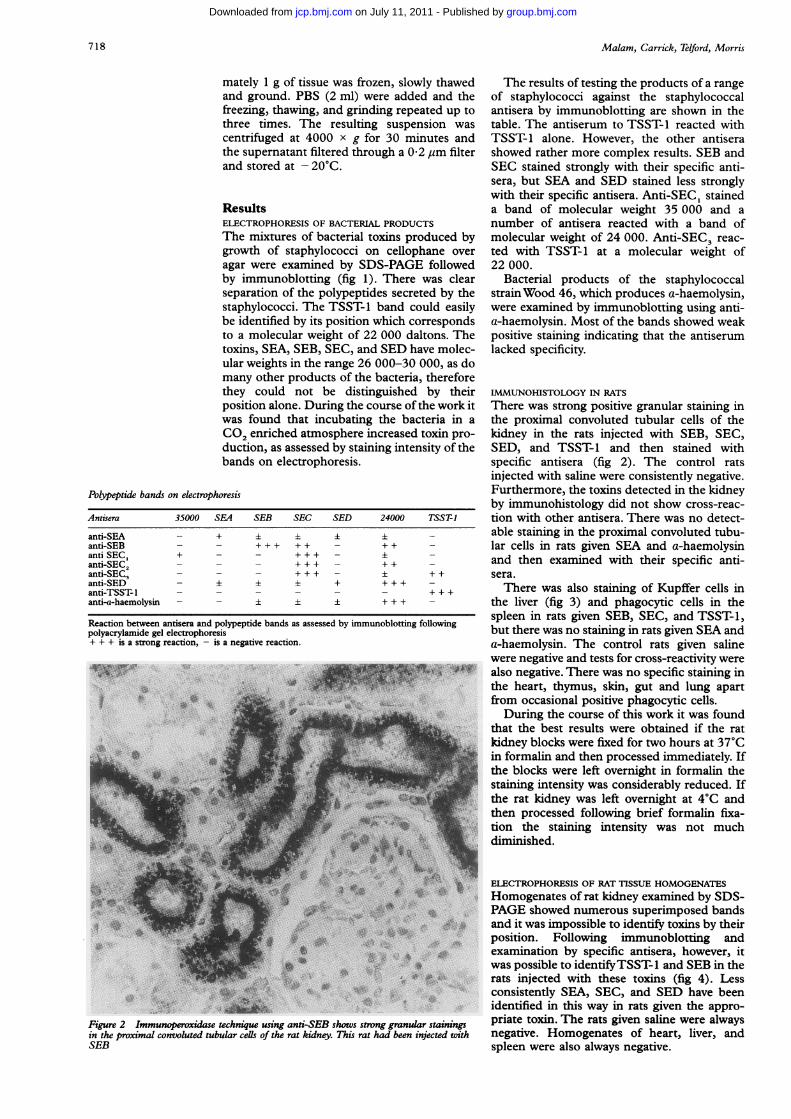
Figure 2 Immunoperoxidase technique using anti-SEB shows strong granular stainingsin the proximal convoluted tubular cells of the rat kidney. This rat had been injected withSEB
The results of testing the products of a rangeof staphylococci against the staphylococcalantisera by immunoblotting are shown in thetable. The antiserum to TSST-1 reacted withTSST- 1 alone. However, the other antiserashowed rather more complex results. SEB andSEC stained strongly with their specific anti-sera, but SEA and SED stained less stronglywith their specific antisera. Anti-SEC, staineda band of molecular weight 35 000 and anumber of antisera reacted with a band ofmolecular weight of 24 000. Anti-SEC3 reac-ted with TSST-1 at a molecular weight of22 000.
Bacterial products of the staphylococcalstrainWood 46, which produces a-haemolysin,were examined by immunoblotting using anti-a-haemolysin. Most of the bands showed weakpositive staining indicating that the antiserumlacked specificity.
IMMUNOHISTOLOGY IN RATS
There was strong positive granular staining inthe proximal convoluted tubular cells of thekidney in the rats injected with SEB, SEC,SED, and TSST-1 and then stained withspecific antisera (fig 2). The control ratsinjected with saline were consistently negative.Furthermore, the toxins detected in the kidneyby immunohistology did not show cross-reac-tion with other antisera. There was no detect-able staining in the proximal convoluted tubu-lar cells in rats given SEA and a-haemolysinand then examined with their specific anti-sera.There was also staining of Kupffer cells in
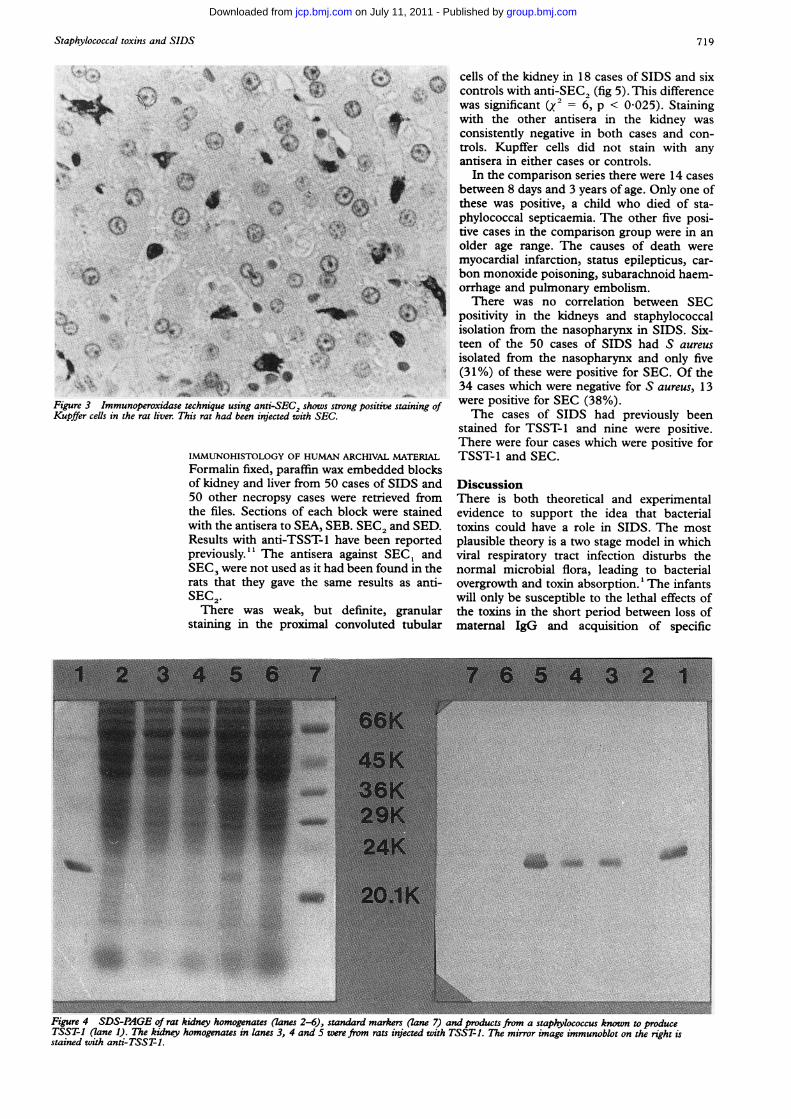
the liver (fig 3) and phagocytic cells in thespleen in rats given SEB, SEC, and TSST-1,but there was no staining in rats given SEA anda-haemolysin. The control rats given salinewere negative and tests for cross-reactivity werealso negative. There was no specific staining inthe heart, thymus, skin, gut and lung apartfrom occasional positive phagocytic cells.During the course of this work it was found
that the best results were obtained if the ratkidney blocks were fixed for two hours at 37°Cin formalin and then processed immediately. Ifthe blocks were left overnight in formalin thestaining intensity was considerably reduced. Ifthe rat kidney was left overnight at 4°C andthen processed following brief formalin fixa-tion the staining intensity was not muchdiminished.
ELECTROPHORESIS OF RAT TISSUE HOMOGENATES
Homogenates of rat kidney examined by SDS-PAGE showed numerous superimposed bandsand it was impossible to identify toxins by theirposition. Following immunoblotting andexamination by specific antisera, however, itwas possible to identifyTSST-l and SEB in therats injected with these toxins (fig 4). Lessconsistently SEA, SEC, and SED have beenidentified in this way in rats given the appro-priate toxin. The rats given saline were alwaysnegative. Homogenates of heart, liver, andspleen were also always negative.
718
group.bmj.com on July 11, 2011 - Published by jcp.bmj.comDownloaded from
Staphylococcal toxins and SIDS
..
.: ..
._ . I._~
_. t
C
*Yb o.
4 .._lo"*r
Figure 3 Immunoperoxidase technique using anti-SEKupffer ceUls in the rat liver. This rat had been injectec
IMMUNOHISTOLOGY
0X; = cells of the kidney in 18 cases of SIDS and sixv controls with anti-SEC, (fig 5). This difference
was significant (X~= 6, p < 0-025). Staining> jI~* with the other antisera in the kidney was< 9>s ~ 7 consistently negative in both cases and con-I t., ,X f3etrols. Kupffer cells did not stain with any
hn iantisera in either cases or controls.In the comparison series there were 14 cases
< > f i Oo between 8 days and 3 years of age. Only one ofthese was positive, a child who died of sta-phylococcal septicaemia. The other five posi-
'nv- tive cases in the comparison group were in anolder age range. The causes of death were
V+-- myocardial infarction, status epilepticus, car-M 0;^ bon monoxide poisoning, subarachnoid haem-
orrhage and pulmonary embolism.There was no correlation between SEC
positivity in the kidneys and staphylococcalisolation from the nasopharynx in SIDS. Six-
@*$£03W$'$4w teen of the 50 cases of SIDS had S aureusisolated from the nasopharynx and only five
es*...eI}e;*' (31%) of these were positive for SEC. Of the34 cases which were negative for S aureus, 13
^=.show. were positive for SEC (38%).C2, shows strong positiVe staining ofi with SEC. The cases of SIDS had previously been
stained for TSST-1 and nine were positive.There were four cases which were positive for
OF HUMAN ARCHIVAL MATERIAL TSST-1 and SEC.Formalin fixed, paraffin wax embedded blocksof kidney and liver from 50 cases of SIDS and50 other necropsy cases were retrieved fromthe files. Sections of each block were stainedwith the antisera to SEA, SEB. SEC2 and SED.Results with anti-TSST-1 have been reportedpreviously." The antisera against SEC, andSEC3 were not used as it had been found in therats that they gave the same results as anti-SEC2.There was weak, but definite, granular
staining in the proximal convoluted tubular
DiscussionThere is both theoretical and experimentalevidence to support the idea that bacterialtoxins could have a role in SIDS. The mostplausible theory is a two stage model in whichviral respiratory tract infection disturbs thenormal microbial flora, leading to bacterialovergrowth and toxin absorption.' The infantswill only be susceptible to the lethal effects ofthe toxins in the short period between loss ofmaternal IgG and acquisition of specific
Figure 4 SDS-PAGE of rat kidney homogenates (lanes 2-6), standard markers (lane 7) and products from a staphylococcus known to produceTSST-1 (lane 1). The kidney homogenates in lanes 3, 4 and 5 were from rats injected with TSST-1. The mirror image immunoblot on the right isstained with anti-TSST-1.
4I.+
719
0.""t., .-
.11Wf.,odlli. &
group.bmj.com on July 11, 2011 - Published by jcp.bmj.comDownloaded from
Malam, Carrick, Telford, Morris
'J~~~~~~~~~~~~~~~~~~~~~~~j
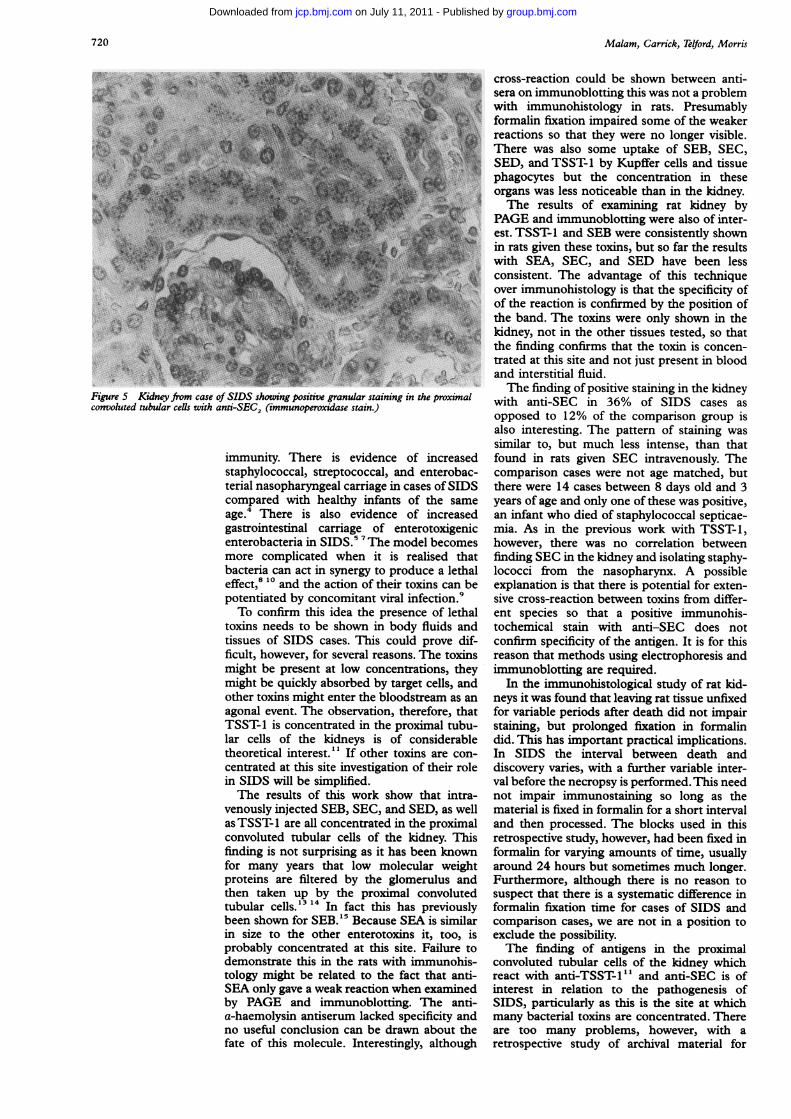
Figure 5 Kidney from case of SIDS showing positive granular staining in the proximalconvoluted tubular cells with anti-SEC2 (immunoperoxidase stain.)
immunity. There is evidence of increasedstaphylococcal, streptococcal, and enterobac-terial nasopharyngeal carriage in cases of SIDScompared with healthy infants of the sameage.4 There is also evidence of increasedgastrointestinal carriage of enterotoxigenicenterobacteria in SIDS.5"Me model becomesmore complicated when it is realised thatbacteria can act in synergy to produce a lethaleffect,8 10 and the action of their toxins can bepotentiated by concomitant viral infection.9To confirm this idea the presence of lethal
toxins needs to be shown in body fluids andtissues of SIDS cases. This could prove dif-ficult, however, for several reasons. The toxinsmight be present at low concentrations, theymight be quickly absorbed by target cells, andother toxins might enter the bloodstream as anagonal event. The observation, therefore, thatTSST-1 is concentrated in the proximal tubu-lar cells of the kidneys is of considerabletheoretical interest." If other toxins are con-centrated at this site investigation of their rolein SIDS will be simplified.The results of this work show that intra-
venously injected SEB, SEC, and SED, as wellas TSST-1 are all concentrated in the proximalconvoluted tubular cells of the kidney. Thisfinding is not surprising as it has been knownfor many years that low molecular weightproteins are filtered by the glomerulus andthen taken up by the proximal convolutedtubular cells.'3 "1 In fact this has previouslybeen shown for SEB.'5 Because SEA is similarin size to the other enterotoxins it, too, isprobably concentrated at this site. Failure todemonstrate this in the rats with immunohis-tology might be related to the fact that anti-SEA only gave a weak reaction when examinedby PAGE and immunoblotting. The anti-a-haemolysin antiserum lacked specificity andno useful conclusion can be drawn about thefate of this molecule. Interestingly, although
cross-reaction could be shown between anti-sera on immunoblotting this was not a problemwith immunohistology in rats. Presumablyformalin fixation impaired some of the weakerreactions so that they were no longer visible.There was also some uptake of SEB, SEC,SED, and TSST-1 by Kupffer cells and tissuephagocytes but the concentration in theseorgans was less noticeable than in the kidney.The results of examining rat kidney by
PAGE and immunoblotting were also of inter-est. TSST-1 and SEB were consistently shownin rats given these toxins, but so far the resultswith SEA, SEC, and SED have been lessconsistent. The advantage of this techniqueover immunohistology is that the specificity ofof the reaction is confirmed by the position ofthe band. The toxins were only shown in thekidney, not in the other tissues tested, so thatthe finding confirms that the toxin is concen-trated at this site and not just present in bloodand interstitial fluid.The finding of positive staining in the kidney
with anti-SEC in 36% of SIDS cases asopposed to 12% of the comparison group isalso interesting. The pattern of staining wassimilar to, but much less intense, than thatfound in rats given SEC intravenously. Thecomparison cases were not age matched, butthere were 14 cases between 8 days old and 3years of age and only one of these was positive,an infant who died of staphylococcal septicae-mia. As in the previous work with TSST-1,however, there was no correlation betweenfinding SEC in the kidney and isolating staphy-lococci from the nasopharynx. A possibleexplanation is that there is potential for exten-sive cross-reaction between toxins from differ-ent species so that a positive immunohis-tochemical stain with anti-SEC does notconfirm specificity of the antigen. It is for thisreason that methods using electrophoresis andimmunoblotting are required.
In the immunohistological study of rat kid-neys it was found that leaving rat tissue unfixedfor variable periods after death did not impairstaining, but prolonged fixation in formalindid. This has important practical implications.In SIDS the interval between death anddiscovery varies, with a further variable inter-val before the necropsy is performed. This neednot impair immunostaining so long as thematerial is fixed in formalin for a short intervaland then processed. The blocks used in thisretrospective study, however, had been fixed informalin for varying amounts of time, usuallyaround 24 hours but sometimes much longer.Furthermore, although there is no reason tosuspect that there is a systematic difference informalin fixation time for cases of SIDS andcomparison cases, we are not in a position toexclude the possibility.The finding of antigens in the proximal
convoluted tubular cells of the kidney whichreact with anti-TSST-1" and anti-SEC is ofinterest in relation to the pathogenesis ofSIDS, particularly as this is the site at whichmany bacterial toxins are concentrated. Thereare too many problems, however, with aretrospective study of archival material for
720
group.bmj.com on July 11, 2011 - Published by jcp.bmj.comDownloaded from

Staphylococcal toxins and SIDS
definite conclusions, but there is sufficientcircumstantial evidence to warrant an intensivestudy of the kidney in SIDS. This should bedone prospectively, with optimal fixation, andusing a wide range of antisera to bacterialtoxins. In parallel, kidney homogenates shouldbe examined by electrophoresis and immuno-blotting to confirm specificity of staining.Furthermore, because antisera to all poten-tially important bacterial toxins are not avail-able, it will be important to develop bioassaysystems for lethal toxins using kidney homo-genates.
This work was funded by a research grant from North WesternRegional Research Committee.
1 Morris JA, Haran D, Smith A. Hypothesis: Commonbacterial toxins are a possible cause of the sudden infantdeath syndrome. Medical Hypotheses 1987;22:211-22.
2 Morris JA. Increased risk of sudden infant death syndromein older infants at weekends. Br Med 1986;293:566.
3 Morris JA. Sudden infant death syndrome. Br Med1989;298:958.
4 Telford DR, Morris JA, Hughes P, et al. The nasopharyngealbacterial flora in sudden infant death syndrome. Infect1989;18: 125-30.
5 Bettelheim KA, Goldwater PN, Dwyer BW, Bourne AJ,Smith DL. Toxigenic Escherichia coli associated withsudden infant death syndrome. Scand Infect Dis1990;22:467-76.
6 Arnon SS, Midura TF, Damus K, Wood RM, Chin J.Intestinal infection and toxin production by Clostridiumbotulinum as one cause ofsudden infant death syndrome.Lancet 1978;i: 1273-7.
7 Sonnabend OAR, Sonnabend WFF, Krech U, Molz G,Sigrist T. Continuous microbiological and pathologicalstudy of 70 sudden and unexpected infant deaths:Toxigenic intestinal Clostridium botulinum infection in 9cases of sudden infant death syndrome. Lancet1985;i:237-41.
8 Lee S, Barson AJ, Drucker DB, Morris JA, Telford DR.Lethal challenge of gnotobiotic weanling rats with bacte-rial isolates from cases of sudden infant death syndrome.J Clin Pathol 1987;40:1393-6.
9 Jakeman KJ, Rushton DI, Smith H, Sweet C. Exacerbationof bacterial toxicity to infant ferrets by influenza virus:Possible role in sudden infant death syndrome. J Infect Dis1991;163:35-40.
10 De Azavedo JCS, Lucken RN, Arbuthnott JP. Effect of toxicshock syndrome Toxin- I on chick embryos. Infect Immun1985;47:710-12.
11 Newbould MJ, Malam J, Mclllmurray JM, Morris JA,Telford DR, Barson AJ. Immunohistological localisationof staphylococcal toxic shock syndrome toxin (TSST-1)antigen in sudden infant death syndrome. J Clin Pathol1989;42:935-9.
12 Laemmli UK. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature1970;227:680-5.
13 Straus W. Cytochemical observations on the relationshipbetween lysosomes and phagosomes in kidney and liverby combined staining for acid phosphatase and intra-venously injected horseradish peroxidase. J Cell Biol1964;20:497-507.
14 Norman SJ. Renal fate of the staphylococcal enterotoxin B.Lab Invest 1971;25:126-32.
15 Norman SJ, Jaeger RF, Johnsey RT. Pathology of experi-mental enterotoxaemia: the in vivo localisation of staphy-lococcal enterotoxin B. Lab Invest 1969;20: 17-25.
721
group.bmj.com on July 11, 2011 - Published by jcp.bmj.comDownloaded from

doi: 10.1136/jcp.45.8.716 1992 45: 716-721J Clin Pathol
J E Malam, G F Carrick, D R Telford, et al. death syndrome.Staphylococcal toxins and sudden infant
http://jcp.bmj.com/content/45/8/716Updated information and services can be found at:
These include:
References http://jcp.bmj.com/content/45/8/716#related-urls
Article cited in:
serviceEmail alerting
the box at the top right corner of the online article.Receive free email alerts when new articles cite this article. Sign up in
Notes
http://group.bmj.com/group/rights-licensing/permissionsTo request permissions go to:
http://journals.bmj.com/cgi/reprintformTo order reprints go to:
http://group.bmj.com/subscribe/To subscribe to BMJ go to:
group.bmj.com on July 11, 2011 - Published by jcp.bmj.comDownloaded from