calcium-mediated modulation of staphylococcal bacterial biofilms
TRANSCRIPT

Indian Journal of Geo-Marine Sciences Vol. 43(11), November 2014, pp. 2107-2116
Calcium-Mediated Modulation of Staphylococcal Bacterial Biofilms
Sudhir K. Shukla1,2, T. Subba Rao1,2,*
1Biofouling & Biofilm Processes Section,Water and Steam Chemistry Division, Bhabha Atomic Research Centre, Kalpakkam – 603102, Tamil nadu, India
2Homi Bhabha National Institute, Mumbai 400094, India [E-mail: [email protected] ]
Received 20 June 2014; revised 16 September 2014
Studies on the effect of calcium on Bap-mediated inhibition and modulation of Staphylococcus aureus biofilm was the motivation behind this study. In this study, the effect of calcium on modulation of Staphylococcus aureus biofilm has been studied. Four S. aureus bovine mastitis isolates viz. SA7, SA10, SA33 and bap-positive S. aureus V329 (positive control) along with a bap-isogenic mutant M556 (negative control) were used in this study. PCR was used for detection of bap, biofilm assays were done in microtitre plates and confocal laser scanning microscopy (CLSM) was used to study the biofilm architecture. PCR mediated detection identified SA7, SA10 and SA33 as bap-negative strains. Biofilm assay showed that Ca2+ inhibited the biofilm growth of SA10 and SAV329 in a dose dependent manner whereas SA7 and SA33 did not show any inhibition. Strain M556 was found to be a weak biofilm former and showed no significant change in the presence of Ca2+. The planktonic growth study results showed that Ca2+ did not influence planktonic growth in any of the S. aureus strains except SA10 where ≥ 5 mM Ca2+ had an inhibitory effect. Interaction of Bap and Ca2+ was found to be specific, since studies with Mg2+ did not show any inhibitory effect on V329 biofilm formation. CLSM images of V329, SA7 and SA10 biofilms showed reduction in biofilm thickness as well as altered biofilm topography. The inhibition effect of Ca2+ on V329 and SA10 biofilms disappeared in the presence of chelating agent EDTA at a sub-minimum inhibitory concentration (0.15 mM). The paper elaborates the role of Ca2+ in the biofilm architecture of S. aureus.
[Keywords: Staphylococcus aureus; calcium; biofilm]
Introduction
Staphylococcus aureus is predominant among the various infections causing bacteria1. Persistent infection by S. aureus is due to its pronounced ability to develop biofilms that confers incredible tolerance to antimicrobial agents1-3. Various methods have been tried to prevent and control the S. aureus infection but had limited success due to ill-discerned fundamentals of primary attachment and biofilm development. S. aureus uses multiple mechanisms involved in primary adhesion and biofilm development. In S. aureus, interactions with abiotic hydrophilic surfaces are mediated by polysaccharide intracellular adhesion (PIA), which is encoded by the ica operon (icaABCD)4,5. It was shown that ica- dependent biofilm could be easily dispersed by DispersinB6,7 whereas ica-independent biofilms (absence of PIA) were easily disrupted by proteinase K8,9. This indicated that apart from PIA, cell surface proteins also play a major role in cell adhesion and biofilm development. Developments of these two types of S. aureus biofilms are currently being actively studied as their regulation and respective roles during infection are fully not understood. Thus, it is
necessary to develop better understanding of fundamentals of S. aureus biofilm development especially; factors that are involved in primary attachment to devise better strategies for the treatment of S. aureus infections.
Many surface proteins such as Bap (biofilm-
associated protein), Eap (extracellular adherence protein) and SasG (S. aureus surface protein G) are known to promote S. aureus adherence to host cells and surfaces as well as cell aggregation10-13. Among which Bap, a large cell surface protein, was isolated from bovine mastitis S. aureus V329. It was found to be involved in initial adhesion and biofilm formation14. Bap protein contains four potential calcium binding EF domains15. In general, extracellular Ca2+plays an important role in maintaining the integrity of the cell wall16and strengthening the biofilm matrix by cross-linking the extracellular polymeric substances and sometimes enhances the EPS production17. However, some recent reports on effect of Ca2+ on Bap-mediated S. aureus biofilm had opposite effect. It has been demonstrated that > 5 mM Ca2+ concentration had inhibitory as well as modulatory effect on biofilm architecture of

INDIAN J. MAR. SCI., VOL 43, NO.11 NOVEMBER 2014 2108
bap-positive S. aureus15,18. Our earlier report showed that Ca2+had dose dependent effect on S. aureus biofilm parameters such as biofilm thickness, biomass, roughness coefficient and surface to bio-volume ratio18.
Since milk contains high concentration of
Ca2+ and bap-positive S. aureus isolates are mostly found at mastitis infection sites19, it was hypothesized that Ca2+ present in milk may play a critical role in bap-positive S. aureus biofilm development or inhibition. In the present study, we investigated the influence of Ca2+on biofilm development of four bovine mastitis isolates of S. aureus, viz., one bap-positive S. aureus V329, three clinical mastitis isolates and one laboratory constructed bap-isogenic mutant of S. aureus M556.The objective of the study was to detect the presence of bap gene in three mastitis S. aureus isolates and to investigate whether calcium had any positive/negative effect on their biofilm forming capacity. Materials and Methods Microorganisms and culture conditions
Five S. aureus strains were used in the study. Details of the strains are given as below (Table 1). Before each experiment the inocula from the stock cultures were added to the freshly prepared media and allowed to grow overnight in an incubator at 37oC. Two ml of overnight grown S. aureus cultures were inoculated in 50 ml of tryptic soy broth (TSB) supplemented with 0.25% glucose (w/v) in 250 ml flasks and were incubated at 37oC in an incubator at 150 rpm. Mid-log phase cells were harvested, washed two times with phosphate buffer saline (PBS) and used for the biofilm studies after adjusting the absorbance (O.D.) to 0.5 at 600nm. TRIS-G minimal media supplemented with 1% glucose (w/v) was used in some of the experiments. Constituents of the one litre TRIS medium were as follows; Tris (hydroxymethyl) aminomethan 6.057 g, Tris HCl 7.88 g, NaCl (4.675 g), KCl (1.4912 g), NH4Cl (1.0698 g), Na2SO4 (4.261 g), MgCl2.6H2O (2.033 g), CaCl2.2H2O (0.294 g), NaH2PO4..2H2O (40.mg), Ferrous ammonium-citrate (4.8 mg/l) and 100 µl of trace elements stock solution [5 mM Zinc sulphate heptahydrate, 5 mM manganese chloride-Tetrahydrate, 10 mM Boric acid, 1 mM Nickel chloride hexahydrate, 1,5 mM sodium molybdate- dehydrate]. Filter sterile CaCl2 and
EDTA were added to culture media as per the experimental plans.
Table 1: Details of S. aureus strains used in this study Strain Details Source S. aureus V329
V329, bap-positive mastitis isolates
Prof. I. Lasa, Spain
S. aureus M556
M556, bap-isogenic mutant of V329
Prof. J.R. Penades, Spain
S. aureus 7 SA7, mastitis isolate Department of Microbiology, Karnataka veterinary college, India
S. aureus 10
SA10, mastitis isolate
-Do-
S. aureus 33
SA33, mastitis isolate
-Do-
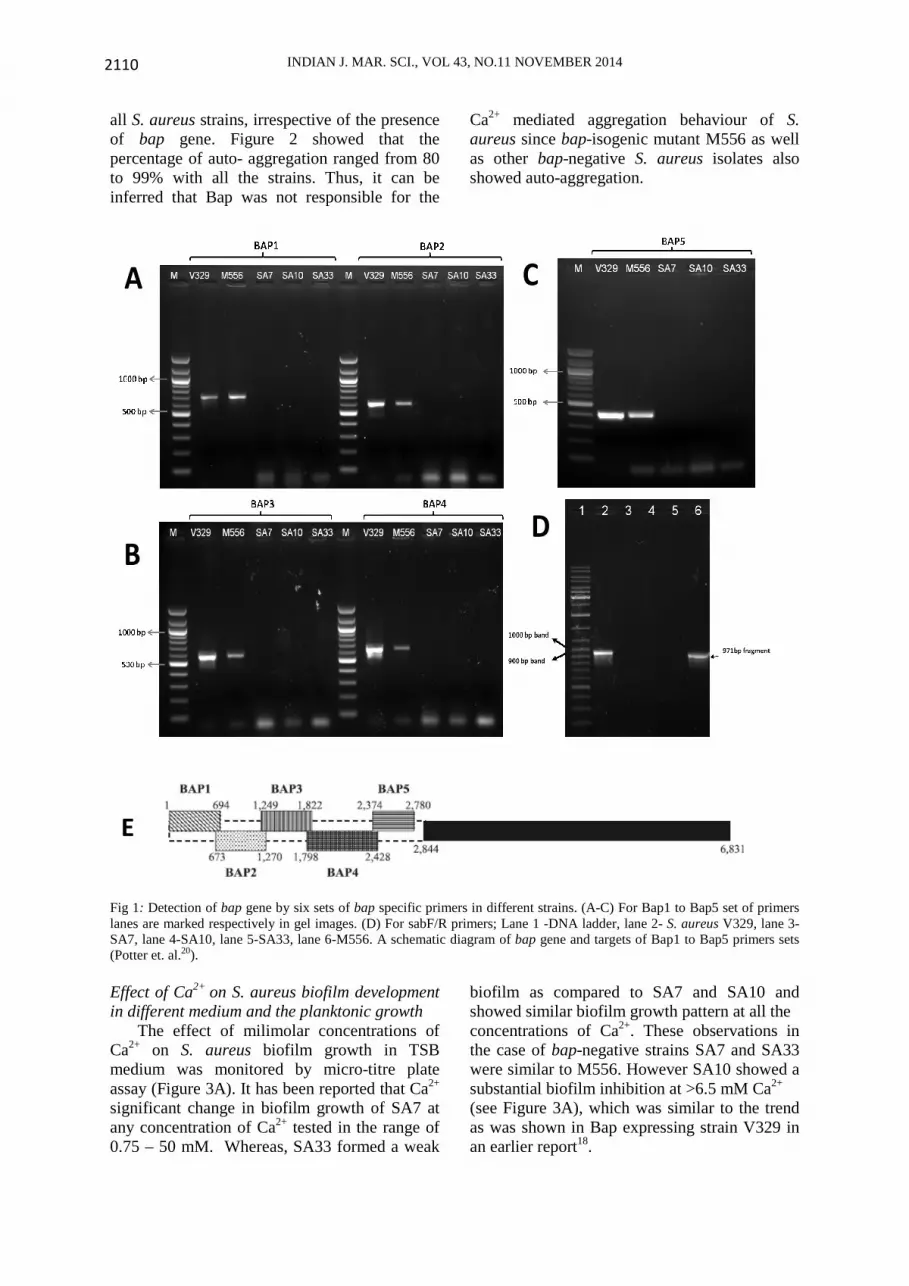
PCR for bap gene detection
Bacterial cultures were grown overnight and genomic DNA was isolated from 3 ml mid log phase culture using a Qiagen DNA isolation kit according to manufacturer’s protocol except that the bacterial cells were lysed by 5 µl of a 10 mg ml-1 solution of lysostaphin (Sigma) and incubated at 37°C for 2 h. For the detection of the bap gene, a set of previously published primers were selected (Table 2)20,21. This ensures specificity and minimum bias in PCR amplification. Each primer set was targeted to different region of bap gene. PCR program was set up as follows; an initial denaturation at 94oC for 5 min was followed by 40 cycles of 94oC for 1 min, Tm
oC for 30 sec, and 72oC for 1 min with a final step at 72oC for 5 min. The size of the PCR products was analysed by electrophoresis on 1% (w/v) agarose gel. Auto-aggregation assay
This assay was performed as per the standard procedure detailed by Beloin et. al.,22. The S. aureus stock cultures were sub cultured; the overnight grown broth (2 ml) was inoculated in 50 ml of TSB medium. Mid log phase cells were harvested and washed three times in PBS. Final OD was set to 1.0 in the PBS at 600 nm. The cell suspension was incubated at 37oC for 4 h. Filter sterile Calcium chloride was added to achieve different concentration of Ca2+. After 4 h of incubation absorbance was measured at 600 nm and the percentage of auto-aggregation was calculated by using the following formula:
%Autoaggregation =��� − ���
���X100
Where, OD0 is OD at zero hour and ODt is OD at t hours (i.e., given time)

SUDHIR et al.: CALCIUM-MEDIATED MODULATION OF STAPHYLOCOCCAL BACTERIAL BIOFILMS
2109
Microtitre plate assay for biofilm formation SA7, SA10 andSA33 biofilms were formed
on pre-sterilized 96 well flat bottom polystyrene microtitre plates in triplicates. Cell suspension (10 µl) having 0.5 OD was inoculated in 190 µl TSB medium in each well and incubated for 16 h at 37oC.The biofilm formation was quantified by classical crystal violet assay23. For dissolution of bound crystal violet 33% acetic acid was used18. Biofilm growth was monitored in terms of absorbance at 570nm using micro plate reader (Multiskan, Thermo Labsystems). Planktonic growth studies
The experiments were carried out with overnight grown bacterial cultures by harvesting the cells and rinsing them twice with PBS and OD of each culture was set to 0.5 at 600 nm wherein the cell concentration was 1x108cfu ml-1. 100 µl of each S. aureus culture was inoculated in 1900 µl of TSB medium supplemented with 0.25% glucose. Cultures were incubated at 37oC and 150 rpm. Absorbance of each culture was recorded at different time intervals after vortexing for 5 sec, to re-suspend the settled cells. Confocal laser scanning microscopy (CLSM) studies and image analysis
Biofilms grown on pre-sterilized microscopic glass slides were studied using CSLM18. Briefly, 2 ml of 0.5 OD cultures of S. aureus was inoculated in sterile petri plates containing 20 ml of TSB medium supplemented with 0.25% glucose. Sterile glass slides were immersed into the medium as substratum for biofilm growth. The petriplates were incubated at 80 rpm for 15 min on shaker for proper mixing of the CaCl2 and EDTA and incubated for 24 h at 37 ◦C. The glass slides were gently washed with PBS to remove loosely attached cells and stained with 0.2% acridine orange for 5 min, thereafter the slide is thoroughly washed with PBS. A thin cover slide was mounted over the stained biofilm and observed by keeping the slide upside down on objective lens of the confocal laser scanning microscope (TCS SP2 AOBS) equipped with DM IRE 2-inverted microscope (Leica Microsystems, Hessen Wetzlar, Germany). Image stacks were collected from 20 random points of the biofilms in order to get an accurate mean value of the biofilm parameters.
Statistical Analysis All data are expressed as mean standard deviation (SD) of the triplicate experimental data. A two-tailed Student’s t-test was used to determine the differences in biofilm formation between the control and each group. The P value < 0.05 was taken as significant. Results Detection of bap gene by PCR in S. aureus isolates All S. aureus strains were screened for the presence of bap gene, S. aureus V329 a bap-positive strain was used as positive control. PCR mediated search for bap gene with the help of six sets bap-specific primers (Table 2) targeting different regions of bap was performed. Table 2: Primers used to detect bap gene in this study. Primers
Sequence (5’-3’)
Ampli-fied Product (bp)
Refe-rence
Bap1 F Bap1 R
ATGGGAAATAAACAAGGTTTTTTACC CTTCTTGTTGTTTATCTGGCTC
694 Potter et. al.20
Bap2 F Bap2 R
GAGCCAGATAAACAACAAGAAG CATGCTCAGCAATAATTGGATC
598 -do-
Bap3 F Bap3 R
GATCCAATTATTGCTGAGCATG CACCTTCGATATATGGTAGTAAGTC
574 -do-
Bap4 F Bap4 R
GACTTACTACCATATATCGAAGGTG CCTCTGCATTAATTACTTTAGC
631 -do-
Bap5 F Bap5 R
GTTCCTCTTAAAGAAGGTGCAG CTAGCTGTTGAAGTTAATACTG
407 -do-
sab F sab R
CCCTATATCGAAGGTGTAGAATTGCAC GCTGTTGAAGTTAATACTGTACCTGC
971 Vautor et.al.21
Sizes of the anticipated amplicons were
present in V329 and M556 (as shown in Table 2). M556 strain also showed the presence of bap gene, since it was constructed by insertion of transposon in to downstream of bap gene, resulting in the production of truncated protein lacking cell wall anchoring region. Effect of Ca2+ on auto-aggregation in S. aureus
To study the effect of Ca2+on auto-aggregation milimolar (mM) concentrations of Ca2+ were added and tested with S. aureus cultures. Ca2+enhanced the auto-aggregation in
INDIAN J. MAR. SCI., VOL 43, NO.11 NOVEMBER 2014 2110
all S. aureus strains, irrespective of the presence of bap gene. Figure 2 showed that the percentage of auto- aggregation ranged from 80 to 99% with all the strains. Thus, it can be inferred that Bap was not responsible for the
Ca2+ mediated aggregation behaviour of S. aureus since bap-isogenic mutant M556 as well as other bap-negative S. aureus isolates also showed auto-aggregation.
Fig 1: Detection of bap gene by six sets of bap specific primers in different strains. (A-C) For Bap1 to Bap5 set of primers lanes are marked respectively in gel images. (D) For sabF/R primers; Lane 1 -DNA ladder, lane 2- S. aureus V329, lane 3- SA7, lane 4-SA10, lane 5-SA33, lane 6-M556. A schematic diagram of bap gene and targets of Bap1 to Bap5 primers sets (Potter et. al.20). Effect of Ca2+ on S. aureus biofilm development in different medium and the planktonic growth The effect of milimolar concentrations of Ca2+ on S. aureus biofilm growth in TSB medium was monitored by micro-titre plate assay (Figure 3A). It has been reported that Ca2+ significant change in biofilm growth of SA7 at any concentration of Ca2+ tested in the range of 0.75 – 50 mM. Whereas, SA33 formed a weak
biofilm as compared to SA7 and SA10 and showed similar biofilm growth pattern at all the concentrations of Ca2+. These observations in the case of bap-negative strains SA7 and SA33 were similar to M556. However SA10 showed a substantial biofilm inhibition at >6.5 mM Ca2+ (see Figure 3A), which was similar to the trend as was shown in Bap expressing strain V329 in an earlier report18.
A
B
C
D
E

SUDHIR et al.: CALCIUM-MEDIATED MODULATION OF STAPHYLOCOCCAL BACTERIAL BIOFILMS
2111
0 1 2.5 5 10 25 500
20
40
60
80
100
120
% A
uto-
aggr
egat
ion
Calcium Conc. (mM)
V329 SA7 SA10 SA33 M556
Fig. 2: Effect of different concentration of Ca2+ on auto-aggregation behaviour of different S. aureus strains. Error bars at some points are not visible due to very small variation.
0 0.75 1.56 3.12 6.25 12.5 25 500.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Bio
film
gro
wth
(O
D 5
70nm
)
Calcium Concentartion (mM)
SA7 SA10 SA33 V329 M556
(A) TSB medium
0 0.75 1.56 3.12 6.25 12.5 25 500.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Bio
film
Gro
wth
(O
D 5
70nm
)
Calcium Concentration (mM)
SA7 SA10 SA33 V329 M556
(B) TRIS-G medium
Fig. 3: Comparative study of biofilm forming capacity among bap-positive and bap-negative strains of S. aureus in the presence of different concentration of Ca2+. Biofilm was evaluated after 16 h of growth. Error bars at some points are not visible due to very small variation.
0 1 2 3 4 5 6 7 8 9 100.0
0.2
0.4
0.6
0.8
1.0
1.2
Pla
nkto
nic
grow
th (
OD
600)
Time (h)
Control 1 mM Ca2+
2 mM Ca2+
5 mM Ca2+
10 mM Ca2+
SA7
0 1 2 3 4 5 6 7 8 9 100.0
0.2
0.4
0.6
0.8
1.0
1.2
Time (h)
Control 1 mM Ca2+
2 mM Ca2+
5 mM Ca2+
10 mM Ca2+
SA10
0 1 2 3 4 5 6 7 8 9 100.0
0.2
0.4
0.6
0.8
1.0
1.2
Time (h)
Control 1 mM Ca2+
2 mM Ca2+
5 mM Ca2+
10 mM Ca2+
SA33
Fig. 4: Effect of different concentration of Ca2+ on the planktonic growth of S. aureus strains (SA7, SA10, SA33).

INDIAN J. MAR. SCI., VOL 43, NO.11 NOVEMBER 2014 2112
Biofilm assay using TRIS-G (Figure 3b) showed that biofilm formation capability of SA10 was significantly reduced in the minimal medium as compared to nutrient rich and complex TSB medium and apparently had no effect of increasing concentrations of Ca2+. Whereas SA7 and SA33 strains did not show any response in the presence of Ca2+and exhibited similar behaviour as shown in complex medium (TSB).
Planktonic growth studies were carried out for SA7, SA10 and SA33 to evaluate whether inhibition of SA10 biofilm in the presence of Ca2+ was due to the hampered planktonic growth. Planktonic growth studies (Figure 4) showed that Ca2+ did not decrease/inhibit the planktonic growth of the cells of SA7 and SA33 but in the case of SA10, planktonic growth was
hampered at ≥ 5 mM Ca2+, which resulted in inhibition of biofilm formation. This observation also suggested that calcium-mediated biofilm inhibition in SA10 strain was essentially Bap independent. Effect of Mg2+ on V329 biofilm formation
In this experiment, we studied whether the biofilm inhibition in V329 was Ca2+ specific or any other divalent ion can manifest similar effect. To test this assumption, similar concentrations of Mg2+ ions i.e. 0.78 mM to 50 mM were used in biofilm assay. Figure 5 describes that Mg2+ did not show any significant change in biofilm forming capacity of V329 even up to 50 mM concentration, while Ca2+showed a dose dependent inhibition.
Fig. 5: Effect of different concentrations of divalent ions (Mg2+ and Ca2+) on biofilm forming capacity of bap –positive S.
aureus V329.
Effect of Ca2+ on S. aureus biofilms; A Confocal Laser Scanning Microscopy study
CSLM studies were also performed to investigate the effect of Ca2+ on S. aureus biofilms. S. aureus biofilms (SA7, SA10 and SA V329) were grown in the presence of critical inhibitory concentrations of Ca2+ (5 mM) in one set and in second set, 5 mM Ca2+ along with sub-inhibitory concentration of EDTA (0.15 mM), which chelates the Ca2+ ions. SA33 and M556 were excluded from this study as they were found to be a very weak biofilm producer on glass slides. Table 3 shows maximum biofilm thickness and average biofilm thickness observed in each case. CLSM images of control V329 biofilms and in the presence of Ca2+
(Figure 5 and Figure 6) show that there was a significant decrease in biofilm thickness in the presence of 5 mM Ca2+. As a control set V329 biofilm formed in the presence of Ca2+ showed a significant difference in biofilm thickness from the control biofilm (P <0.001) and when Ca2+ was chelated by EDTA, biofilm forming capacity of V329 strain was restored (P = 0.3264). Identical treatment of Ca2+ was given to S. aureus SA7 and SA10, where no substantial differences were observed in the case of SA7. Whereas similar to V329 biofilm, a decrease in the biofilm thickness was also observed in SA10 biofilm in the presence of Ca2+(10.33 ± 2.35 µm) as compared to control biofilm (19.91 ± 3.21 µm; P< 0.001).
0 0.78 1.56 3.12 6.25 12.5 25 50
0.0
0.5
1.0
1.5
2.0
Bio
film
gro
wth
O
D a
t 570
nm
Conc. of Calcium (mM)
Calcium Magnesium

SUDHIR et al.: CALCIUM-MEDIATED MODULATION OF STAPHYLOCOCCAL BACTERIAL BIOFILMS
2113
Table 3. A comparative CLSM study of 24 h old biofilms of bap-positive V329 and bap-negative SA7
and SA10 strains in the presence of Ca2+ in one set and EDTA + Ca2+ in another set.
Strains Control CaCl2
(5 mM) CaCl2 (5 mM) + EDTA (0.15 mM)
Mea
n
thic
kne
ss
(µm
±S
D)
SA V329 23.43 ± 4.82 12.95 ± 2.58 25.28 ± 6.71 SA7 17.12 ± 2.07 15.35 ± 2.41 10.66 ± 1.60 SA10 19.91 ± 3.21 10.33 ± 3.35 23.98 ± 4.59
Max
imum
T
hick
nes
s(µ
m) SA V329 31.89 21.60 32.31
SA7 22.76 22.57 11.97 SA10 24.94 18.92 28.89
Control Ca2+ (5 mM)
Fig. 5: CSLM images of 24 old control biofilms (V329 SA7 and SA10) and biofilms grown in the presence of 5 mM Ca2+.
V329
SA7
SA10

INDIAN J. MAR. SCI., VOL 43, NO.11 NOVEMBER 2014 2114
Figure 6 and Table 3 indicate that addition of EDTA had reversed the Ca2+ inhibition effect in both V329 as well as SA10 biofilms. SA10 biofilm grown in Ca2+ along with EDTA did not show significant difference (P > 0.05) as compared to control biofilm. Discussion
Biofilm development is widely believed to be initiated by bacteria sensing the surface that trigger the transition from a planktonic state to a biofilm mode of growth24. Adhesion is a threshold event in formation of biofilms, studies on molecular and biochemical aspects have highlighted that the protein matrix of the biofilm structure is of interest in developing strategies to combat microbial fouling /chronic biofilm infections14. Microbiota acquires diverse mechanisms to adhere and initiate the biofilm development in various
environments25. In the pioneering study by Arrizubieta et al.18 on the effect of Ca2+ on S. aureus biofilms they that
Control Ca2+ EDTA + Ca2+0
5
10
15
20
25
30
35
40
Avg
. bio
film
thic
knes
s (µ
m)
V329 SA7 SA10
Fig. 6: Comparison of biofilm thickness of 24 old biofilmsof bap-positive, bap-negative strains of S. aureus in the presence of Ca2+ in one set and EDTA + Ca2+ in another set. Error bars are shown as ±1SD.
Ca2+affected the multi-cellular behaviour of Bap expressing S. aureus cells in cell suspension, where all experiments were carried out in TSB-glucose. However, we observed that addition of Ca2+ to TSB medium increased the turbidity and it was difficult to interpret the aggregation behaviour in turbid conditions. Therefore, PBS was used to observe auto-aggregation of S. aureus cells in the presence of Ca2+. The results showed that auto aggregative behaviour was independent of the Bap presence. This observation was found to be contradictory to the previous findings where Bap was claimed to be an essential factor for aggregative behaviour of S. aureus V329 strain15.
Biofilm assay for SA7, SA10, SA33 along with positive control (V329) and negative control (M556) showed that there was no effect of increasing concentration of Ca2+ on SA7 and SA33 but SA10 biofilm growth was inhibited. Effect of Ca2+ on bap-negative SA10 biofilm was found to be very similar to bap- +ve V329. The biofilm assay results strengthen the hypothesis that putative presence of EF hand motifs for Ca2+ in Bap might influence the biofilm development in bap-positive S. aureus. This inhibitory effect in V329 was due to the presence of Bap as bap-isogenic mutant M556 did not manifest similar response. In our previous study, it was found that the inherent concentration of Ca2+
in TSB medium (Difco make) was 7.36 ± 0.85 mM18. It was also observed that addition of Ca2+into TSB enhanced the turbidity of the medium. This observation led us to contemplate that some essential factor/s, which might be required for SA10 biofilm formation, could have been complexed with Ca2+ and became unavailable to the bacterium. To rule out this possibility, we used TRIS minimal medium supplemented with 1% glucose (Tris-G) and monitored the effect of Ca2+on biofilm formation of bap-negative S. aureus strains. Results showed that SA7 and SA33 strains did not show any biofilm inhibition in the presence of Ca2+, however SA10 showed reduced biofilm formation in TRIS-G as compared to TSB-G, with no Ca2+inhibition effect (Figure 3B). Biofilm assay using minimal medium suggested two points; (1) biofilm inhibition by Ca2+ in V329 was not due to any complexion of Ca2+with medium component. (2)S. aureus strain SA10 needs an unknown factor present in complex media for growth which also competes with Ca2+. Higher concentrations of Ca2+ out compete (or block the uptake of) the required unknown factor hence reduced the growth of SA10 biofilm. In minimal medium the unknown factor was absent all together hence biofilm formation was significantly decreased and we did not see any competing effect at any concentrations of Ca2+. At this stage, it is suggested that the significant variation in SA10 biofilm growth in

SUDHIR et al.: CALCIUM-MEDIATED MODULATION OF STAPHYLOCOCCAL BACTERIAL BIOFILMS
2115
the two different media needs further investigation to prove above speculations.
In order to evaluate, whether inhibition of
biofilm was due to the hampered growth or due to the life style switch over of the cells to planktonic form, growth studies were carried out. The growth study results indicated that Ca2+did not inhibit the cell division of the SA7 and SA33 bacterial cells but had inhibitory effect on SA10. Thus, it was concluded that decrease in biofilm forming capacity of SA10 was due to the inhibition of cell division, unlike bap-positive V329 where calcium inhibition of biofilm was Bap-mediated15,18.
According to Michiels et. al.16, EF hand motifs are known to have very specific interaction with Ca2+. Biofilm assay in presence of Mg2+ showed that Bap-dependent Ca2+ inhibition of V329biofilm development is highly specific. If other surface proteins or Bap had non-specific or electrostatic interactions with divalent ions then high concentrations of Mg2+ had resulted in the inhibition of biofilm forming capacity. Biofilm assay in the presence of high salt concentration showed that Bap-dependent Ca2+ inhibition of biofilm development in V329 is highly specific. Therefore, the probability of inhibition of biofilm formation because of nonspecific electrostatic interactions between Ca2+ and Bap as well as other surface proteins was ruled out.
CSLM analysis of S. aureus biofilms development at critical concentrations of Ca2+showed a decrease in biofilm formation and the presence of Ca2+ substantially affected the architecture of V329 biofilm (positive control) and SA10. CSLM analysis showed that there was a gradual decrease in average thickness of SA10 and V329 biofilms, which reconfirmed that Ca2+ inhibited the SA10 biofilm development. Reduction of pillar size, flattening of biofilm and reduction of the biomass was observed when SA10 biofilm was grown with 5 mM Ca2+. The effect of Ca2+on S. aureus biofilm inhibition was further investigated by adding EDTA to the medium. Addition of divalent chelator EDTA confirmed that observed architectural change was due to the presence of Ca2+only as addition of EDTA restored the thickness of both the S. aureus biofilms i.e. V329 and SA10 biofilms.
The present study supports earlier proposed hypothesis that multiple mechanisms are adopted by different staphylococci for biofilm formation26-28. It was postulated that adherence of S. aureus involves multiple mechanism; some of them involve surface proteins that bind Ca2+ and others involve surface proteins that are independent of Ca2+. This study highlights that Ca2+ ion can affect S. aureus biofilm formation by affecting its planktonic growth. However, at this stage with limited knowledge on adherence mechanism of bacteria, we cannot predict that what are the other factors involved in Ca2+ mediated modulation of bap-negative staphylococcal biofilms.
In brief, this study establishes the following salient points ;( 1) PCR mediated detection approach identifies SA7, SA10 and SA33 as bap negative S. aureus strains. (2) auto-aggregation could also be due to nonspecific bridging of acidic functional groups of cell surface polymers with Ca2+and was found to be independent of Bap presence. (3) Ca2+ inhibited the biofilm formation in bap-negative SA10 and bap- positive V329, but did not affect biofilm formation in bap-negative SA7 and SA33 even up to 50 mM. (4) In V329 Ca2+ inhibition was found to be Bap mediated whereas in SA10 it was found to be due to the inhibition of planktonic growth. (5) CLSM study showed that Ca2+ influenced the biofilm development of the SA10, which was reversed by chelation of EDTA. Although, our studies were carried out using a clinical isolate of S. aureus as model organism it establishes the fact that cell surface proteins play important role in biofilm establishment and presence of Ca2+might affect its growth. A better understanding of molecular aspects of adhesion and biofilm phenomenon will certainly help in developing control strategies for clinical manifestations as well as aid in designing better industrial cooling water treatment. Acknowledgements Authors are grateful to Prof. Inigo Lasa, Laboratory of Microbial Biofilms, Institute of Agro-biotechnology, Spain, for providing bap-positive S. aureus strain, and José R Penadés, Instituto Valenciano de Investigaciones Agrarias (IVIA)-Universidad Cardenal Herrera-CEU, Moncada, Spain, for providing

INDIAN J. MAR. SCI., VOL 43, NO.11 NOVEMBER 2014 2116
bap-isogenic mutant of S. aureus. Authors also acknowledge Prof. S. K. Isloor, Department of Microbiology, KVC, Bangalore, India, for providing other mastitis isolates of S. aureus. References 1. Donlan, R.M. Biofilms and device-associated
infections. Emerg. Infect. Dis.7 (2001) 277-281. 2. Sutra, L. & Poutrel, B. Detection of capsular
polysaccharide in milk of cows with natural intramammary infection caused by Staphylococcus aureus. Am. J. Vet. Res.51 (1990) 1857-1859.
3. Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med.339 (1998) 520-532.
4. Gotz, F. (2002) Staphylococcus and biofilms. Mol.Microbiol.43, 1367-1378.
5. Cramton, S. E., Gerke, C., Schnell, N. F., Nichols, W. W., & Götz, F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect. Immun. 67(1999) 5427-5433.
6. Kaplan, J.B., Ragunath, C., Velliyagounder, K., Fine, D.H. & Ramasubbu, N. Enzymatic detachment of Staphylococcus epidermidis biofilms. Antimicrob agents chemotherapy48 (2004) 2633-2636.
7. Kogan, G., Sadovskaya, I., Chaignon, P., Chokr, A. & Jabbouri, S. Biofilms of clinical strains of Staphylococcus that do not contain polysaccharide intercellular adhesin. FEMS Microbiol. Lett.255 (2006) 11-16.
8. Chaignon, P., Sadovskaya, I., Ragunah, C., Ramasubbu, N., Kaplan, J.B. & Jabbouri, S. Susceptibility of staphylococcal biofilms to enzymatic treatments depends on their chemical composition. Appl. Microbiol. Biotechnol.75 (2007) 125-132.
9. Shukla, S.K. & Rao, T.S. Dispersal of Bap-mediated Staphylococcus aureus biofilm by proteinase K. J. Antibiot. 66 (2013) 55-60.
10. Cucarella, C., Tormo, M.A., Ubeda, C., Trotonda, M.P., Monzon, M., Peris, C., Amorena, B., Lasa, I. & Penades, J.R. Role of biofilm-associated protein bap in the pathogenesis of bovine Staphylococcus aureus. Infect. Immun.72 (2004) 2177-2185.
11. Hussain, M., A. Haggar, C. Heilmann, G. Peters, J. I. Flock, & M. Herrmann. Insertional inactivation of eap in Staphylococcus aureus strain Newman confers reduced staphylococcal binding to fibroblasts. Infect. Immun. 70 (2002) 2933-2940.
12. Stoodley, P., Sauer, K., Davies, D.G. &Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol.56 (2002) 187-209.
13. Hussain, M., Haggar, A., Peters, G., Chhatwal, G.S., Herrmann, M., Flock, J.-I. & Sinha, B. More than one tandem repeat domain of the extracellular adherence protein of Staphylococcus aureus is required for aggregation, adherence, and host cell invasion but not for leukocyte activation. Infect. Immun. 76 (2008) 5615-5623.
14. Corrigan, R. M., Rigby, D., Handley, P., & Foster, T. J. The role of Staphylococcus aureus surface protein SasG in adherence and biofilm formation. Microbiology 153 (2007) 2435-2446.
15. Cucarella, C., Solano, C., Valle, J., Amorena, B., Lasa, I. & Penades, J.R. Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J. Bacteriol. 183 (2001) 2888-2896.
16. Arrizubieta, M.J., Toledo-Arana, A., Amorena, B., Penades, J.R. & Lasa, I. Calcium inhibits bap-dependent multicellular behavior in Staphylococcus aureus. J. Bacteriol.186 (2004) 7490-7498.
17. Michiels, J., Xi, C., Verhaert, J. & Vanderleyden, J. The functions of Ca (2+) in bacteria: a role for EF-hand proteins? Trends Microbiol.10 (2002) 87-93.
18. Mangwani, N., Kumari, S., Shukla, S.K., Rao, T. & Das, S. Phenotypic Switching in Biofilm-Forming Marine Bacterium Paenibacillus lautus NE3B01. Curr. Microbiol.68 (2014), 648-656.
19. Shukla, S.K. & Rao, T.S. Effect of calcium on Staphylococcus aureus biofilm architecture: A confocal laser scanning microscopic study. Colloids Surf B: Biointerfaces103 (2013), 448-454.
20. Tormo, M.A., Knecht, E., Gotz, F., Lasa, I. & Penades, J.R. Bap-dependent biofilm formation by pathogenic species of Staphylococcus: evidence of horizontal gene transfer? Microbiology151 (2005) 2465-2475.
21. Potter, A., Ceotto, H., Giambiagi-Demarval, M., dos Santos, K.R., Nes, I.F. & BastosMdo, C. The gene bap, involved in biofilm production, is present in Staphylococcus spp. strains from nosocomial infections. J. Microbiol.47 (2009) 319-326.
22. Vautor, E., Abadie, G., Pont, A. & Thiery, R. Evaluation of the presence of the bap gene in Staphylococcus aureus isolates recovered from human and animals species. Vet. Microbiol.127 (2008) 407-411.
23. Beloin, C., Michaelis, K., Lindner, K., Landini, P., Hacker, J., Ghigo, J.M. & Dobrindt, U. The transcriptional antiterminatorRfaH represses biofilm formation in Escherichia coli. J. Bacteriol.188 (2006) 1316-1331.
24. O'Toole, G.A. & Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: a genetic analysis. Mol. Microbiol.28 (1998) 449-461.
25. Johnson, M., Cockayne, A., Williams, P.H. & Morrissey, J.A. Iron-responsive regulation of biofilm formation in Staphylococcus aureus involves fur-dependent and fur-independent mechanisms. J. Bacteriol.187 (2005) 8211-8215.
26. Thomas, V.L., Sanford, B.A. & Ramsay, M.A. Calcium- and mucin-binding proteins of staphylococci. J. Gen. Microbiol.139 (1993) 623-629.
27. Davey, M.E. & O'Toole G, A. Microbial biofilms: from ecology to molecular genetics. Microbiol. Mol. Biol. Rev.64 (2000) 847-867.
28. O'Toole, G., Kaplan, H.B. & Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol.54 (2000) 49-79.