injectable plga microspheres encapsulating wkymvm peptide for neovascularization
TRANSCRIPT

Acta Biomaterialia xxx (2015) xxx–xxx
Contents lists available at ScienceDirect
Acta Biomaterialia
journal homepage: www.elsevier .com/locate /actabiomat
Injectable PLGA microspheres encapsulating WKYMVm peptidefor neovascularization
http://dx.doi.org/10.1016/j.actbio.2015.07.0331742-7061/� 2015 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved.
⇑ Corresponding authors at: School of Chemical and Biological Engineering, SeoulNational University, 1 Gwanak-ro, Gwanak-gu, Seoul 151-742, Republic of Korea(N.S. Hwang).
E-mail addresses: [email protected] (J.H. Kim), [email protected] (N.S. Hwang).1 These authors contributed equally.
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microspheres encapsulating WKYMVm peptide for neovascularization, Acta Bio(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
Young Hwan Choi a,1, Soon Chul Heo b,1, Yang Woo Kwon b, Hwan D. Kim a, Seung Hyun L. Kim a, Il Ho Jang b,Jae Ho Kim b,⇑, Nathaniel S. Hwang a,c,⇑a School of Chemical and Biological Engineering, Institute of Chemical Processes, Seoul National University, Seoul 152-742, Republic of Koreab Department of Physiology, School of Medicine, Pusan National University, Yangsan 626-870, Gyeongsangnam-do, Republic of Koreac N-Bio Institute, Seoul National University, Seoul 152-742, Republic of Korea
a r t i c l e i n f o
Article history:Received 5 March 2015Received in revised form 24 June 2015Accepted 23 July 2015Available online xxxx
Keywords:WKYMVm peptidePLGA microsphereDrug deliveryOutgrowth endothelial cellsNeovascularization
a b s t r a c t
Formyl peptide receptor-2 (FPR-2) is expressed in various cell types, such as phagocytes, fibroblasts, andendothelial cells. FPR-2 has been reported to play a significant role in inflammation and angiogenicresponse, and synthetic WKYMVm peptide has been identified as a novel peptide agonist for the FPR-2.In this study, we demonstrate that WKYMVm peptides stimulate the angiogenic potential of outgrowthendothelial cells (OECs). Upon WKYMVm peptide exposure, migration and proliferation of OECs werestimulated. WKYMVm effectively stimulated angiogenesis in tube formation assay and aortic ring assay.Furthermore, we fabricated injectable poly (lactide-co-glycolide) (PLGA) microspheres encapsulatingWKYMVm peptides, which showed sustained release of cargo molecule. When WKYMVm peptide encap-sulated microspheres were injected into the hind limb ischemia model, a single injection of microsphereswas as effective as multiple injections of WKYMVm peptide in restoring blood flow from ischemic injuryand promoting capillary growth. These results demonstrate that sustained release of WKYMVm peptidefrom microspheres in the application to ischemic hind limb extended angiogenic stimulation.
Statement of Significance
Formyl peptide receptor (FPR) has been reported to play an important role in inflammation and angio-genic response. A synthetic WKYMVm peptide has been identified as a novel peptide activating theFPR-2 that is expressed in a various cell types, such as phagocytes, fibroblasts, and endothelial cells. Inthis manuscript we explored a unique property of high-affinity ligand for formyl peptide receptors-2(FPR-2) (i.e., WKYMVm). WKYMVm-induced activation of FPR2 has been reported to be crucial in hostdefense and inflammation by activation of phagocytes, monocytes, and lymphocytes. In this study, high-light the efficacy of WKYMVm peptide’s role in inducing neovascularization in vivo hind limb ischemiamodel when the peptide was released from injected PLGA microspheres in sustained manner. Our resultsdemonstrate that sustained release of WKYMVm peptide from microspheres have extended angiogenicstimulation capacity.
� 2015 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Angiogenesis, defined as functional collateral vessel formationout of preexisting vessels, contributes to enhanced tissue perfu-sion. Factors such as smoking, hypertension, hypercholesterolemia,
atherosclerosis, and diabetes can cause insufficient blood flow totissue and abnormal collateral vessel formation in ischemic tissue[1–3]. Among the disease caused by risk factors, peripheral arterydiseases (PAD) is a narrowing of the arteries that supply the heart,brain or legs. PAD has been commonly known to affect the legs. Inperipheral vascular defects, limbs that do not receive enough bloodflow results in limb pain accompanied by necrosis and ulceration.Finally, amputation of the ischemic limb may be required [4]. Toinduce neovascularization in ischemic tissues, previous studieshave utilized therapeutic angiogenic growth factors. To date,
mater.

2 Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx
vascular endothelial growth factor (VEGF), hepatocyte growth fac-tor (HGF), and basic fibroblast growth factor (bFGF) have been uti-lized to induce blood vessel formation [5,6]. However, there stillexists the safety and cost issue associated with expression andpurification of growth factor proteins.
Formyl peptide receptors-2 (FPR-2) has been reported to becrucial in host defense and inflammation by activation ofphagocytes, monocytes, and lymphocytes [7–9]. WKYMVm peptideworks as a high-affinity ligand for FPR-2. Recently, severalstudies identified that the synthetic peptide WKYMVm(Trp-Lys-Tyr-Met-Val-D-Met-NH2) can induce angiogenesis ofendothelial cells via FPR-2 dependent stimulation of proliferation,migration, tube formation, and sprouting activity [10,11]. In addi-tion, a recent study reported that the WKYMVm-induced activationof FPR-2 plays a role in ischemic tissue regeneration and neovascu-larization by homing and angiogenic activation of outgrowthendothelial cells (OECs) [12]. However, therapeutic activities ofinjected peptides may be diminished by several factors, includingthat of rapid degradation and secretion by the circulation system.To provide a large pharmacokinetic window for delivered peptidesin ischemic tissues, a controlled delivery of peptides in therapeuticsconcentration to the ischemic region would be desirable. Previously,biodegradable polymer-based delivery systems have been devel-oped for sustained peptide deliveries [13]. Many injectable drugdelivery systems utilizing biocompatible and biodegradable micro-spheres have been fabricated for peptide deliveries [14–16]. Polylactic-co-glycolic acid (PLGA) is a biocompatible member of the ali-phatic polyester family of biodegradable polymers. This biomateri-als exhibits a broad range of erosion times and has tunablemechanical properties [17]. This FDA approved polymer had beenextensively studied for peptide, protein, and drug carriers [18,19].PLGA degrades slowly via hydrolysis in an aqueous environment.In this study, we describe the formation and characterization ofPLGA microspheres form by double-emersion. Particle size and mor-phology are determined via scanning electron microscopy (SEM).Furthermore, release kinetics and therapeutic efficacy ofWKYMVm peptides from PLGA microspheres were investigatedboth in vitro and in vivo.
2. Materials and methods
2.1. Cell culture
Human outgrowth endothelial cells (OECs) were isolated fromhuman umbilical cord blood as previously described [12]. Cellswere seeded on to the gelatin coated tissue culture plate(Millipore, Embryomax�, Ultrapure water with 0.1% gelatin) andmaintained with endothelial cell basal medium-2 (EBM-2; Lonza,Walkersville, MD) supplemented with EGM™-2 singleQuots�
except hydrocortisone. EBM-2 medium was exchanged 24 h afterthe initial plating to remove non-adherent cells. EBM-2 mediumwas changed every day, and OECs with passage numbers between5 and 10 were used for all experiments.
2.2. Synthesis of WKYMVm peptides
WKYMVm (Trp-Lys-Tyr-Met-Val-D-Met) was synthesized bythe solid phase peptide synthesis (SPPS) according to a classicalFmoc/tBu method. WKYMVm peptide purity was checked by ana-lytical HPLC with a single peak. The mass was confirmed by massspectroscopy with the peptide mass of 856.50 (Molecular weightof WKYMVm peptide, Supplementary Fig. 1).
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
2.3. PLGA microsphere fabrication and WKYMVm peptideencapsulation
180 mg of PLGA (Sigma–Aldrich, Mw: 50,000–75,000, lac-tide:glycolide (85:15)) was dissolved in 2 ml methylene chlorideby using vortex for 15 min. After the polymer was completely dis-solved, 180 ll of 1.2 mM WKYMVm peptide in water (W1) wasadded to polymer solution, and the mixture was sonicated for a1 min (Branson Sonifier 250, Output control; 2. Duty cycle; 30%).Then the W1 in oil (W1/O) phase solution was added to 50 ml of1% w/w Poly (vinyl alcohol) (Sigma–Aldrich, Mw: 89,000–98,000)in distilled water, and emulsified for 1 min by homogenizer(VWR PowerMax AHS 250, 10,000 rpm), and the homogenizedmixture was set under a fume hood and stirred by magnetic stirrerfor 3 h to evaporate off the methylene chloride solution and hardenthe particles. W1/O/W2 double emulsion PLGA microsphere wasfinally fabricated. PLGA microspheres encapsulating WKYMVmpeptide were diluted to adjust the same concentration of 10 lMWKYMVm peptide in vitro experiments and 120 lM WKYMVmpeptide in vivo experiments.
2.4. Micro particles characterization
The PLGA microspheres were lyophilized for 48 h before beingexamined. Then, the particles were mounted on a SEM sampleholder with carbon tape and were sputter-coated with platinum.All images were captured using an acceleration voltage of 10 kV.To examine the degradation of microspheres, the particles weresuspended in a 10 ml phosphorus buffer solution (PBS) in 50 mlconical tube (Falcon™) for 50 days. Coulter Multisizer (BECKMANCOULTER. Multisizer™ 3 COULTER COUNTER�) was used to deter-mine the mean particle diameter and distribution.
2.5. Cell proliferation assay and migration analysis
OECs were incubated in EBM-2 with 10% FBS with WKYMVm(10�6 mol/L or 10�5 mol/L) or with VEGF (10 ng/ml) (rhVEGF,R&D system, Cat No. 290-VS/CF) for 48 h. Then, EdU (Click-iT�
EdU Flow Cytometry Assay Kits, Invitrogen, Cat No. C10425) wasadded to the well (10 lM). After 3 h of incubation, cells were thenfixed, permeabilized, and stained with Alexa Fluor� 488 azide (CatNo. C10425) for 30 min and stained with 40,6-diamidino-2-phenylindole (DAPI; Sigma–Aldrich) for 10 min. S phase cells’ ratio wasanalyzed by quantifying the percentage of EdU+ cells per totalDAPI+ cells. 1/10th volume of AlamarBlue (AlamarBlue� cell viabil-ity reagent, Cat No. DAL1025) reagent was added to cells in culturemedium, and Cells were incubated for 2 h at 37 �C in a cell cultureincubator, protected from direct light. Then the chemical reductionof alamar blue was measured using fluorescence spectroscopy(kex = 550 nm, kem = 600 nm) using Infinite� 200 PRO TECAN Ltd.Cellular migration was performed via wound scratch assay, as pre-viously described [20,21].
2.6. In vitro controlled release analysis
The release profile of fluorescein isothiocyanate (FITC, Sigma–Aldrich) from the PLGA microspheres was measured using fluores-cence spectroscopy (kex = 495 nm, kem = 520 nm) using Infinite�
200 PRO TECAN Ltd. The microsphere (1.2 mM FITC) samples wereincubated in PBS at 37 �C for up to 600 h. Supernatants at differenttime points (24, 48, 120, 168, 360, 480, 600 h) was collected andanalyzed. Relative fluorescence intensity values were convertedinto the actual weight of FITC released by correlation with a stan-dard curve.
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.

Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx 3
2.7. Tube formation assay and aorta ring assay
300 ll of chilled BD Matrigel™ matrix (BD Biosciences, BD CatNo. 354234) were added to each well of 24-well culture plates,and plates were incubated at 37 �C for 30 min. Membrane labeledOECs (PKH26 red fluorescent cell linker kits for general cell mem-brane labeling, Sigma–Aldrich, Cat No. MINI26 and PKH26GL) wereseeded onto a layer of Matrigel within 24-well plates at a density of10 � 104 cells per well. Then they were incubated with EBM-2 con-taining 5% FBS or supplemented with WKYMVm (10�6 mol/L) orWKYMVm (10�5 mol/L) or VEGF (10 ng/ml). After 12 h of incuba-tion, tube formation was analyzed by quantifying the area of tubenetworks in three random microscope fields (Image J, MATLAB).Aortic rings were isolated from BALB/c mouse (8 weeks old), andcut into 1–1.5 mm segments, and placed on 24-well plates coatedwith 100 ll of Matrigel, and add another 100 ll of Matrigel tocover each ring. Then they were incubated with EBM-2 containing5% FBS or supplemented with WKYMVm (10�6 mol/L) orWKYMVm (10�5 mol/L) or VEGF (10 ng/ml). Mouse aorta ringassay with PLGA microsphere encapsulating WKYMVm peptides,WKYMVm (10�5 mol/L) or PLGA microspheres or PLGA micro-spheres encapsulating WKYMVm peptides (10�5 mol/L ofWKYMVm peptide) were incubated mouse aorta. After 3 days ofincubation, neovascularization was analyzed by quantifying thenumber of sprouts per aorta segment.
2.8. Real-time PCR analysis
Cells from each group were collected to extract RNAs withTrizol. cDNA reverse transcription was performed using theSuperScript� First-Strand Synthesis System for RT-PCR(Invitrogen™). Real Time-PCR was performed using the SYBRGreen PCR Master mix and the ABI StepOnePlus™ Real-Time PCRSystem (Applied Biosystems). The synthesized cDNA samples wereanalyzed for the genes of interest which are Angiogenin,Angiopoietin-1, and VCAM-1 with b-Actin as a reference gene.Angiogenin (Forward: 50-CATCATGAGGAGACGGGG-30, Reverse: 50
-TCCAAGTGGACAGGTAAGCC-30), Angiopoietin-1 (Forward: 50-AGCTGTGATCTTGTCTTGGC-30, Reverse: 50-GTTCAAGTCTCGTGGTCTGA-30), VCAM-1 (Forward: 50-TAAAATGCCTGGGAAGATGG-30,Reverse: 50-GGTGCTGCAAGTCAATGAGA-30).
2.9. Hind limb ischemia model and in vivo PLGA microsphere injection
Animal experiments were performed using protocols approvedby the Pusan National University Institutional Animal Use and CareCommittee. BALB/C mice (male, age 8–10 weeks, weighing 22–24 g) were anesthetized with an intraperitoneal injection of400 mg/kg 2,2,2,-tribromoethanol (Avertin; Sigma–Aldrich, St.Louis, MO), and the skin was incised. The femoral artery wasexcised from the external iliac artery to its branches. After excisionof arteries, hind limb ischemia induced mice was treated withHBSS (Hank’s Balanced Salt Solution) or WKYMVm (Wm,10�5 mol/L) 3 times per week. PLGA microspheres (Control bead)or PLGA microspheres containing WKYMVm (Wm bead,12 � 10�5 mol/L) was injected into ischemic hind limb with a sin-gle injection on day 0. PLGA microspheres encapsulatingWKYMVm peptides were diluted to adjust the same concentrationof Wm group (injection of 10�5 mol/L of WKYMVm peptides3 times per week for 4 weeks). All groups were injected into threesites (20 ll/each location) of the gracilis muscle in the medialthigh. The extent of necrosis in the ischemic hind limb wasrecorded on day 28 after surgery. Scores for necrosis were assessed(0: limb salvage, 1: toes amputation, 2: foot amputation, 3: limbamputation). Blood perfusion of the hind limb was evaluated usinga laser Doppler perfusion imaging (LDPI) analyzer (Moor
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
instruments, Devon, UK) on days 0 and every week for 4 weeksafter induction of hind limb ischemia. Blood perfusion is expressedas the LDPI index representing the ratio of ischemic versusnon-ischemic limb blood flow. A ratio of 1 before surgery indicatesequal blood perfusion of both legs.
2.10. Histological analysis and immunostaining
For histological and immunostaining of the tissue specimens,hind limb muscles were removed, formalin-fixed, andparaffin-embedded. Three sections measuring 6 lm in thicknesswere taken from the paraffin-embedded specimens at 150 lmintervals. Endothelial cells and smooth muscle cells wereimmunostained with biotinylated isolectin B4 (ILB4; VectorLaboratories, Burlingame, CA) and rabbit anti-a-SMA (Abcam,Cambridge, U.K.) antibodies. The specimens were incubated withstreptavidin conjugated Alexa Fluor 488 or Alexa Fluor 568 goatanti-rabbit secondary antibodies (Life Technologies, Carlsbad,CA), followed by washing and mounting in Vectashield medium(Vector Laboratories) with 40,6-diamidino-2-phenylindole (DAPI)for visualization of nuclei. The stained sections were visualizedusing laser scanning confocal microscopy (Olympus FluoViewFV1000). Capillary density and the number of arterioles/arterieswere assessed by counting the number of ILB4-positive anda-SMA-positive features per high power field (�800). Five ran-domly selected microscopic fields from five serial sections in eachtissue block were examined for the numbers of capillary densityand a-SMA-positive arteries for each mouse.
2.11. Statistical analysis
All data are represented as mean ± standard deviation (SD).Statistical significance was determined by an analysis of variance(ANOVA single factor) with *P < 0.05, **P < 0.01.
3. Results
3.1. WKYMVm peptides stimulate migration, proliferation of OECs
Human blood outgrowth endothelial cells (OECs) were isolatedfrom human umbilical cord blood and maintained with EBM con-taining 5% FBS. OECs were maintained endothelial cell morpholo-gies. In addition, surfaces analysis showed that OECs retainedendothelial cell surface makers (Supplementary Fig. 2). Most ofOECs were positive ECFC markers such as CD31 and CD144. Inaddition, over 50% of cells were positive for Flk1 and 22.5% of cellswere positive for CXCR4. Fraction of the cells displayed CD133(1.2%), c-kit (10.4%), and CD34 (17.3%). Isolated OECs were negativefor hematopoietic lineage markers. Immunostaining confirmedthat these cells take up acetylated low-density lipoprotein(Dil-acLDL), and express multiple endothelial cell markers suchas CD31, Flk1, phospo-eNOS, and vWF (Supplementary Fig. 3).
In order to explore the functional effects of the WKYMVm pep-tides on OECs migration, we treated the cells with EBM containing5% FBS and WKYMVm (10�6 mol/L) or WKYMVm (10�5 mol/L) andcompared to that with EBM containing VEGF (10 ng/ml). OECs cul-tured with EBM containing 5% FBS was utilized for negative con-trol. Cells were plated at 2.5 � 105 cells/cm2 gelatin coated tissueculture plates, and cells were grown to confluence. A thin ‘wound’was introduced by scratching with a 1 ml micropipette tip.Washing with PBS twice removed detached cells from scratching.Then, they were incubated in EBM-2 with 5% FBS, withWKYMVm (10�6 mol/L), with WKYMVm (10�5 mol/L) or withVEGF (10 ng/ml). After 10 h of incubation, cell migration was ana-lyzed by measuring the relative area covered by cells to compare
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.

Fig. 1. Effects of WKYMVm peptide on migration and proliferation of OECs in vitro. WKYMVm peptide resulted in enhanced migration and proliferation in concentrationdependent manner. (A) Representative images of scratch wounding migration. (Scale bar = 500 lm). (B) Representative images of OECs in S phase (EdU+, as indicated ingreen), Nucleus (DAPI+, as indicated in blue). (Scale bar = 100 lm). (C) Migration of OECs was measured in response to WKYMVm (10�6 mol/L), WKYMVm (10�5 mol/L) orVEGF (10 ng/ml). Scratch wounding migration was quantified by measuring the area beyond the reference line. (D–E) To evaluate the proliferative effect of WKYMVm peptideon OECs, immunocytochemistry and cell viability assay were performed. (D) Expression levels of EdU were quantified by counting of EdU positive cells among the totalAppositive cells and presented as a percentage of EdU positive cells. (E) Innate metabolic activity was represented by the chemical reduction of alamarblue reagent. Thechemical reduction of alamarblue was measured using fluorescence spectroscopy. (For interpretation of the references to color in this figure legend, the reader is referred tothe web version of this article.)
4 Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx
with the initial gap. Quantification was performed by image J. Asshown in Fig. 1(A and C), the control group (EBM-2) andWKYMVm (10�6 mol/L) group had no significant impact on migra-tion of OECs. On the other hand, not only VEGF, but also WKYMVm(10�5 mol/L) stimulated migration of OECs. However WKYMVm(10�5 mol/L) group had less effect than VEGF (10 ng/ml) group.
To assess whether WKYMVm peptides can improve the prolifer-ation of OECs, cells in S phase were counted after treatment of cellswith WKYMVm peptides for 48 h. WKYMVm peptides resulted inan increase of EdU positive cells in a dose-dependent manner. Inparticular, the proliferative effect of WKYMVm peptides at10�5 mol/L concentration was as potent as that of VEGF(Fig. 1B and D). Additionally, we incubated OECs with EBM-2 con-taining 10% FBS, WKYMVm (10�6 mol/L), WKYMVm (10�5 mol/L)or VEGF (10 ng/ml). We treated alamar blue with culture mediumfor 2 h each day in a cell culture incubator. The innate metabolicactivity resulted in a chemical reduction of alamar blue.WKYMVm-induced proliferation of OECs was increased in timedependent manner. As shown in Fig. 1E, WKYMVm peptides at
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
10�6 mol/L and 10�5 mol/L concentration stimulated proliferationof OECs better than the control group at day 3 and day 4.
3.2. WKYMVm peptides stimulate angiogenesis of aorta ring, and tubeformation, gene expression of OECs
To evaluate whether WKYMVm peptides can inducemicro-vessel formations, we incubated aorta ring in EBM-2 with5% FBS, with WKYMVm (10�6 mol/L), with WKYMVm(10�5 mol/L), or with VEGF (10 ng/ml). The angiogenic effect ofWKYMVm peptides was assessed by the endothelial cell sproutingfrom mouse aortic rings. As shown in Fig. 2(A and C), the sproutingof endothelial cells was increased as the result of WKYMVm pep-tides treatment, in a dose-dependent manner, whereas it was rarelyobserved in the absence of WKYMVm peptides. The largest numberof micro-vessel endothelial networks and branches sprouting fromthe aortic ring was observed in the group treated with theWKYMVm (10�5 mol/L). We further evaluated whether WKYMVmpeptides can stimulate tube formation of OECs. OEC cells were
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.

Fig. 2. Effects of WKYMVm peptide on angiogenesis of aorta ring and tube formation and angiogenic gene expression level of OECs in vitro. WKYMVm peptide stimulatedangiogenic sprouting from aortic ring, and improved tube formation and angiogenic gene expression level of OECs. (A) Vessel images depicting sprouting from aorta ring.(Scale bar = 500 lm). (B) Images depicting tube formation of OECs. (Scale bar = 500 lm). (C) Aortic ring, which were extracted from BALB/c mouse, were cultured in thematrigel with EBM, WKYMVm (10�6 mol/L), WKYMVm (10�5 mol/L) or VEGF (10 ng/ml). This treatment lasted for 3 days and then photographs were taken of representativeaortic rings. Mouse aorta ring in response to WKYMVm (10�5 mol/L) showed a higher number of angiogenic sprouts. (D and E) WKYMVm-induced formation of tubulestructure of OECs. Tube formation was quantified by measuring the area of tubule structure in three random fields from each well by Image J and MATLAB, and normalizingthe values relative to those of the corresponding control. The number of tube loops was quantified by counting the rings of tubule structure. (F–H) To determine theangiogenic effects of WKYMVm peptides, angiogenic specific gene expressions (Angiogenin (ANG), Angiopoietin-1 (ANGPT-1), Vascular cell adhesion molecule 1(VCAM-1))were investigated by real-time PCR. OECs were cultured for 7 days with EBM, WKYMVm (10�6 mol/L), WKYMVm (10�5 mol/L) or VEGF (10 ng/ml).
Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx 5
seeded onto Matrigel-coated 24 well plate at 5 � 105 cells/well.Treatment with WKYMVm peptides resulted in dose-dependentstimulation of tube formation (Fig. 2B, D, and E). WKYMVm(10�5 mol/L) group formed the largest area of tubule structure(Fig. 2D) and the highest number of tube loops (Fig. 2E).WKYMVm (10�5 mol/L) group facilitated the formation of tubule
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
structures about twice the number compared to the control group.The quantifications of area and number of tube structures were ana-lyzed using Image J and MATLAB. To confirm the pro-angiogeniceffects of WKYMVm peptides, blood vessel specific gene expres-sions such as angiogenin (ANG), angiopoietin-1 (ANGPT-1), and vascu-lar cell adhesion molecule 1 (VCAM-1) were analyzed by real-time
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.

Fig. 3. Fabrication of PLGA microspheres containing WKYMVm peptides in a double emulsion process. Microspheres encapsulating WKYMVm peptides were fabricated viadouble emulsion methods, and size distribution. (A) The scheme of PLGA microsphere fabrication and WKYMVm peptides encapsulation. (B) Microscopic image and scanningelectron microscope (SEM) Image of PLGA microspheres after the fabrication. The PLGA microspheres were lyophilized before being imaged by SEM. (scale bar = 200 lm). (C)The size distribution of PLGA microspheres. The size of PLGA microsphere was measured by Coulter Multisizer.
6 Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx
PCR in each group at day 7. All gene expressions were internally nor-malized to housekeeping b-Actin gene. As shown in Fig. 2(F–H), thegene expressions of ANGPT-1, and VCAM-1 were up-regulated in allthree groups compared to the control group. On the contrary, thegene expression level of ANG was similar in all four groups. Theresults showed that the gene expressions of ANGPT-1 and VCAM-1were up-regulated in the groups treated with the WKYMVm pep-tides compared to the control group. The groups treated with the10 lM WKYMVm peptides showed approximately the 5 times inthe expression of ANGPT-1 and the 10 times in the expression ofVCAM-1 than the control groups.
3.3. PLGA microspheres encapsulating WKYMVm peptides
For sustained release application, WKYMVm peptide loadedmicrospheres were successfully prepared by the W1/O/W2 doubleemulsion solvent evaporation method (Fig. 3A). The sizes and mor-phologies of the PLGA microspheres were examined with a
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
scanning electron microscope (SEM). As shown in Fig. 3B, micro-scopic image and SEM image showed that the microspheres aresmooth and spherical shape. Through size distribution of micro-spheres, the mean size of the microspheres was confirmed to be2.784 lm (total number: 10138) (Fig. 3C). The sustained peptidesrelease from the PLGA microspheres depended on the degradationof PLGA microspheres (Fig. 4A). Microspheres that were placedunder the hydrolytic degradation condition in PBS for 50 daysshowed significant deformation with large holes were remnantthroughout (Fig. 4B). In order to investigate the release rate fromPLGA microspheres, we encapsulated FITC via double emersionmethods and monitored the release via fluorescence measurement.Light microscopy conformed the size and the morphology of theFITC encapsulated PLGA microspheres. Furthermore, fluorescencemicroscopy confirmed the presence of FITC molecules within themicrospheres (Fig. 4C). FITC encapsulated PLGA microspheres wereincubated under the hydrolytically degrading condition up to25 days, and released FITC molecules were measured and
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.

Fig. 4. Release of WKYMVm peptides from PLGA microspheres with time dependent degradation of PLGA and effects on angiogenesis of aorta ring. WKYMVm peptides, whichreleased from microspheres facilitated sprouting of micro vessel from aorta ring as much as WKYMVm peptides which just treated with media did. (A) Scheme of thedegradation of PLGA microspheres and sustained release of WKYMVm peptides from PLGA microspheres. (B) To identify the degradation of PLGA microspheres, microsphereswere suspended in PBS for 50 days, and then microspheres were lyophilized. Degradation images of PLGA microspheres were taken by using Scanning electron microscope(SEM). (C) The fluorescence images of PLGA microspheres encapsulating fluorescein isothiocyanate (FITC). (Scale bar = 50 lm). (D) The cumulative release profile of FITC frommicrospheres. The amount of FITC was measured using fluorescence spectroscopy for 600 h. (E and F) Aortic ring, were cultured in the matrigel with EBM, WKYMVm(10�5 mol/L), PLGA microspheres, PLGA microspheres containing WKYMVm (10�5 mol/L). This treatment lasted for 3 days and then photographs were taken of representativeaortic rings. Mouse aorta ring in response to PLGA microspheres containing WKYMVm showed a higher number of angiogenic sprouts. (Scale bar = 500 lm).
Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx 7
accumulated FITC molecules were quantified by a microplatereader. We observed sustained FITC release after an initial burstrelease at the beginning. (Fig. 4C). FITC molecules on the surfaceof PLGA microspheres which were not completely encapsulatedwithin the microspheres may have contributed to the initial burstrelease.
3.4. Mouse aorta ring assay with PLGA microspheres encapsulatingWKYMVm peptides
In order to evaluate whether PLGA microspheres encapsulatingWKYMVm peptides can induce angiogenic sprout formations frommouse aorta, we incubated aorta in EBM-2 with 5% FBS (Control),10�5 mol/L WKYMVm (Wm), empty PLGA microsphere (Controlbead) or PLGA microspheres encapsulating 10�5 mol/L WKYMVmpeptides (Wm bead) for 3 days. The angiogenic effect of each groupwas assessed by counting the number of sprouts from mouse aorticrings. As shown in Fig. 4E and F, the control bead group had no sig-nificant impact on the sprouting when compared to the control. Onthe other hand, both WKYMVm (10�5 mol/L) and PLGA micro-spheres encapsulating WKYMVm peptides group (10�5 mol/L)stimulated angiogenic sprouts from aorta ring. The group treated
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
with the 10�5 mol/L of WKYMVm had the largest number of microvessel networks and branches sprouting from the aortic ring.
3.5. PLGA microspheres encapsulating WKYMVm peptides improvesblood perfusion and ischemic limb salvage in a murine ischemic hindlimb animal model
To assess whether the WKYMVm peptides and the PLGA micro-spheres containing WKYMVm peptides can stimulate neovascular-ization in vivo, hind limb ischemia was induced in BALB/c mice.Femoral artery was ligated surgically, and blood perfusion wasmeasured for 4 weeks with intramuscular injections of HBSS,WKYMVm peptide, control PLGA microspheres, or PLGA micro-spheres encapsulating WKYMVm peptides into the ischemic hindlimb. Intramuscular injection of WKYMVm (Wm, 10�5 mol/L,3 times per week) or PLGA microspheres containing WKYMVm(Wm bead, 12 � 10�5 mol/L) significantly increased blood flow inthe ischemic hind limb compared with the HBSS-injected controlgroup or control PLGA microsphere-injected group(Fig. 5A and B). Injection of WKYMVm (Wm, 10�5 mol/L, 3 timesper week) or PLGA microspheres containing WKYMVm (Wm bead,12 � 10�5 mol/L) significantly reduced tissue necrosis and limb
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.
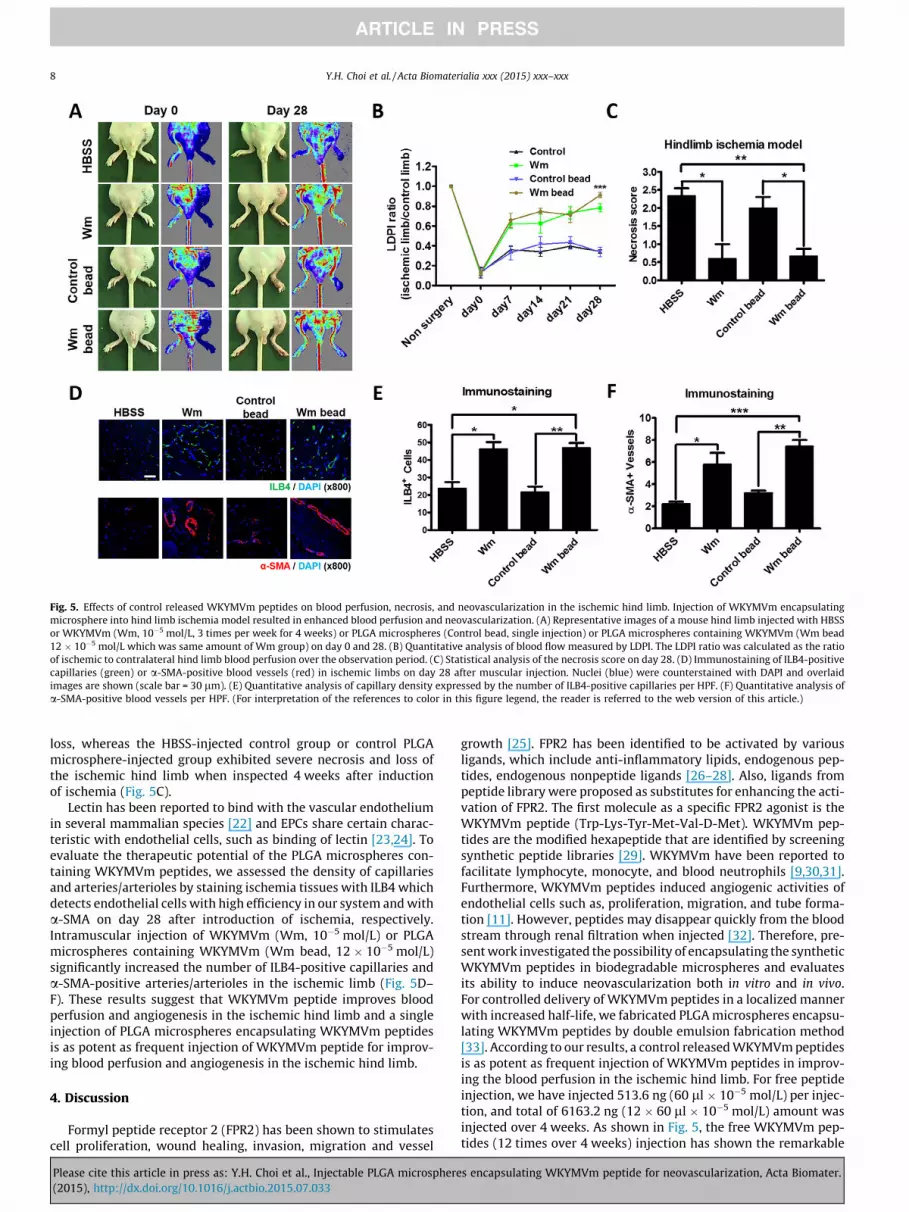
Fig. 5. Effects of control released WKYMVm peptides on blood perfusion, necrosis, and neovascularization in the ischemic hind limb. Injection of WKYMVm encapsulatingmicrosphere into hind limb ischemia model resulted in enhanced blood perfusion and neovascularization. (A) Representative images of a mouse hind limb injected with HBSSor WKYMVm (Wm, 10�5 mol/L, 3 times per week for 4 weeks) or PLGA microspheres (Control bead, single injection) or PLGA microspheres containing WKYMVm (Wm bead12 � 10�5 mol/L which was same amount of Wm group) on day 0 and 28. (B) Quantitative analysis of blood flow measured by LDPI. The LDPI ratio was calculated as the ratioof ischemic to contralateral hind limb blood perfusion over the observation period. (C) Statistical analysis of the necrosis score on day 28. (D) Immunostaining of ILB4-positivecapillaries (green) or a-SMA-positive blood vessels (red) in ischemic limbs on day 28 after muscular injection. Nuclei (blue) were counterstained with DAPI and overlaidimages are shown (scale bar = 30 lm). (E) Quantitative analysis of capillary density expressed by the number of ILB4-positive capillaries per HPF. (F) Quantitative analysis ofa-SMA-positive blood vessels per HPF. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
8 Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx
loss, whereas the HBSS-injected control group or control PLGAmicrosphere-injected group exhibited severe necrosis and loss ofthe ischemic hind limb when inspected 4 weeks after inductionof ischemia (Fig. 5C).
Lectin has been reported to bind with the vascular endotheliumin several mammalian species [22] and EPCs share certain charac-teristic with endothelial cells, such as binding of lectin [23,24]. Toevaluate the therapeutic potential of the PLGA microspheres con-taining WKYMVm peptides, we assessed the density of capillariesand arteries/arterioles by staining ischemia tissues with ILB4 whichdetects endothelial cells with high efficiency in our system and witha-SMA on day 28 after introduction of ischemia, respectively.Intramuscular injection of WKYMVm (Wm, 10�5 mol/L) or PLGAmicrospheres containing WKYMVm (Wm bead, 12 � 10�5 mol/L)significantly increased the number of ILB4-positive capillaries anda-SMA-positive arteries/arterioles in the ischemic limb (Fig. 5D–F). These results suggest that WKYMVm peptide improves bloodperfusion and angiogenesis in the ischemic hind limb and a singleinjection of PLGA microspheres encapsulating WKYMVm peptidesis as potent as frequent injection of WKYMVm peptide for improv-ing blood perfusion and angiogenesis in the ischemic hind limb.
4. Discussion
Formyl peptide receptor 2 (FPR2) has been shown to stimulatescell proliferation, wound healing, invasion, migration and vessel
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
growth [25]. FPR2 has been identified to be activated by variousligands, which include anti-inflammatory lipids, endogenous pep-tides, endogenous nonpeptide ligands [26–28]. Also, ligands frompeptide library were proposed as substitutes for enhancing the acti-vation of FPR2. The first molecule as a specific FPR2 agonist is theWKYMVm peptide (Trp-Lys-Tyr-Met-Val-D-Met). WKYMVm pep-tides are the modified hexapeptide that are identified by screeningsynthetic peptide libraries [29]. WKYMVm have been reported tofacilitate lymphocyte, monocyte, and blood neutrophils [9,30,31].Furthermore, WKYMVm peptides induced angiogenic activities ofendothelial cells such as, proliferation, migration, and tube forma-tion [11]. However, peptides may disappear quickly from the bloodstream through renal filtration when injected [32]. Therefore, pre-sent work investigated the possibility of encapsulating the syntheticWKYMVm peptides in biodegradable microspheres and evaluatesits ability to induce neovascularization both in vitro and in vivo.For controlled delivery of WKYMVm peptides in a localized mannerwith increased half-life, we fabricated PLGA microspheres encapsu-lating WKYMVm peptides by double emulsion fabrication method[33]. According to our results, a control released WKYMVm peptidesis as potent as frequent injection of WKYMVm peptides in improv-ing the blood perfusion in the ischemic hind limb. For free peptideinjection, we have injected 513.6 ng (60 ll � 10�5 mol/L) per injec-tion, and total of 6163.2 ng (12 � 60 ll � 10�5 mol/L) amount wasinjected over 4 weeks. As shown in Fig. 5, the free WKYMVm pep-tides (12 times over 4 weeks) injection has shown the remarkable
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.

Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx 9
efficacy in promoting the recovery of blood flow and the reductionof tissue necrosis and limb loss.
Injected free peptides may experience low therapeutic efficacydue to limited half-life. This may be the sole reason why repeatedinjection of free peptides was needed for therapeutic efficacy.Similarly, Kim et al. have demonstrated large amount of peptides(4 mg/kg/twice per day for 2 days) was required to elicit therapeuticefficacy against poly-microbial sepsis due to its short half-life in vivo[34]. In retrospect, Park et al. demonstrate that WKYMVm peptidemodification by conjugation to cotinine hapten and binding toanti-cotinine antibody significantly enhanced the in vivo stabilityand biological activity [32]. Several other strategies to prolong thepeptides’ half-life have been developed. These technologies includechemical modification, liposomal encapsulation, and polymericencapsulation [35–37]. In this study, we incorporated microspheresencapsulation for WKYMVm peptides for controlled release in local-ized manner. For in vivo application, we have shown that ‘singleinjection’ of PLGA microspheres encapsulating WKYMVm peptides(6163.2 ng, 60 ll � 12 � 10�5 mol/L) demonstrated similar or bet-ter efficacy in comparison to free peptide injected group.
Since most peptides have a short half-life in vivo, direct injec-tion of the drugs at the target site is not appropriate. PLGA is abiodegradable, biocompatible, and FDA approved therapeuticmaterial [17]. For peptide stability, peptides were solubilized inenzyme free distilled water (W1 phase), and this W1 phase wasencapsulated by PLGA polymer (O phase) to inhibit bulk peptidedispersion toward outside. And then, this O phase was stabilizedand hardened by a stabilizing agent PVA (W2 phase) (Fig. 3A).Average diameter of PLGA microspheres ranged from ones to tensof micrometer. This allowed generation of injectable sizes, stabili-ties and large surface areas for drug release. (Fig. 3B and C). In thepresent study, PLGA microspheres demonstrated sustained releaseof the encapsulated FITC. These results demonstrate that peptidesencapsulated within PLGA microspheres could induce enhancedeffects due to the long-lasting, continuous release for a long time.In addition, control released peptides from PLGA microsphereseffectively facilitated angiogenic sprouting of mouse aorta ringin vitro (Fig. 4E and F). Additional studies employing PLGA with dif-ferent lactide–glycolide monomers proportions, variation in micro-spheres particle size and the possible association of differentformulation would optimize a controlled release system for syn-thetic peptides.
Outgrowth endothelial cells (OECs) are kinds of endothelial pro-genitor cells (EPCs) that human circulating cells, and they are alsoreferred to as late-outgrowth EPCs (L-EPCs) [38–40]. OECs aremobilized into the peripheral blood in response to ischemia, andthey are recruited to the ischemic tissue to promote angiogenesisand neovascularization [41]. OECs have a high proliferative capac-ity. Furthermore, OECs facilitate neovascularization of ischemic tis-sue by secreting angiogenic factors and by directly incorporatinginto sites of the newly forming vasculatures [42,43]. In the presentstudy, OECs were characterized by taking up acetylatedlow-density lipoprotein (LDL) and expressing marker proteins ofthe EC lineage such as CD31, CD144 (vascular endothelialVE-cadherin), von Willebrand factor (vWF), endothelial cell nitricoxide synthase (eNOS) (Supplementary Fig. 3). Although manyexperimental studies have shown that the OEC transplantationtherapy was a promising strategy that promoted angiogenesis inanimal models of ischemia, poor engraftment and difficulty ofacquiring the required number of cells limits its use. Rather thantransplanting OECs, recent studies have attempted to recruitendothelial progenitor cell toward ischemic sites for neovascular-ization. A rich variety of proangiogenic molecules have alreadybeen discovered such as vascular endothelial growth factor
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
(VEGF) [44], angiopoietin [45], and erythropoietin [46]. VEGF is apotent inducer of angiogenesis. Signal transduction of VEGFinvolves binding of tyrosine kinase receptors and results inendothelial cell proliferation, migration, and new vessel formation.Previous study utilizing WKYMVm on HUVEC showed that 10 nMconcentration was sufficient to induce cell migration and angio-genic response [11]. However, in the present study, 10 lMWKYMVm peptides was needed to stimulate proliferation, woundmigration, and tube formation of OECs as effective as VEGF(10 ng/ml). Discrepancies in the response may due to the differentserum and medium conditions between two studies.
In pathological conditions, including ischemia, body’s vascularsystem responds mainly two different processes to preserve organfunction and tissue survival, angiogenesis and arteriogenesis [47].Angiogenesis describes the growth of new capillaries by sproutingfrom the pre-existing vasculature [48]. However, angiogenesisalone has a limited capacity to blood perfusion, maintain or restoreoxygen and nutrition supply of hypoxic tissue. In contrast, arterio-genesis is defined by the growth of functional collateral arteriesfrom pre-existing arerio–arteriolar anastomoses [49], and is trig-gered by physical forces such as altered shear forces induced bystenosis or occlusion of a major artery [50]. A number of studiesdemonstrate that tissue resident endothelial cells in the vessel walland circulating progenitor cells in the blood contribute to compen-satory vessel growth [51–53]. In the present study, WKYMVmtreatment enhanced the formation of ILB4-positve capillaries anda-SMA-positive arteries within the ischemic limb, indicating thatWKYMVm contributes not only to the growth of capillaries but alsoto the growth of collateral arteries. Therefore, both tissue residentendothelial cells and circulating progenitor cells could haveresponded to WKYMVm treatment for increased capillary forma-tion and arteriogenesis.
5. Conclusion
In this study, we have shown that WKYMVm peptide effectivelyfacilitated proliferation and angiogenic activities of OECs in vitro.Furthermore, we have engineered biodegradable microsphereencapsulating WKYMVm for controlled release in vivo. Our animalexperiments demonstrated that multiple WKYMVm peptide injec-tion into hind limb ischemia model resulted in a pro-angiogenicresponse. We further showed that WKYMVm peptide loadedmicrosphere led to controlled in vivo release and resulted inincreased therapeutic efficacy of single injection. The resultsobtained in the present study shows the angiogenic responseinduced by the WKYMVm peptides in PLGA microspheres, suggest-ing the possibility of simplifying the peptide-based therapeuticstrategies.
Disclosure
The authors declare no conflict of interest.
Acknowledgements
OECs were kindly provided by Dr. Sang-Mo Kwon’s laboratory(Pusan National University, South Korea). This work was supportedby Brain Fusion Grant of SNU. This research was supported byIndo-Korea SAVI Program through the National ResearchFoundation of Korea (NRF-2013K2A1B9065267). This researchwas also supported by NRF funded by the Ministry of Education,Science and Technology – Republic of Korea (Grant NumbersNRF-2012M3A9C6050102, 2012M3A9C7050184).
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.

10 Y.H. Choi et al. / Acta Biomaterialia xxx (2015) xxx–xxx
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.actbio.2015.07.033.
References
[1] J. Waltenberger, Impaired collateral vessel development in diabetes: potentialcellular mechanisms and therapeutic implications, Cardiovasc. Res. 49 (2001)554–560.
[2] E. Van Belle, A. Rivard, D. Chen, M. Silver, S. Bunting, N. Ferrara, et al.,Hypercholesterolemia attenuates angiogenesis but does not precludeaugmentation by angiogenic cytokines, Circulation 96 (1997) 2667–2674.
[3] H.A. Struijker Boudier, Arteriolar and capillary remodelling in hypertension,Drugs (1999). 59 Spec No: 37–40.
[4] K. Ouriel, Peripheral arterial disease, Lancet 358 (2001) 1257–1264.[5] A. Grochot-Przeczek, J. Dulak, A. Jozkowicz, Therapeutic angiogenesis for
revascularization in peripheral artery disease, Gene 525 (2013) 220–228.[6] T.P. Richardson, M.C. Peters, A.B. Ennett, D.J. Mooney, Polymeric system for
dual growth factor delivery, Nat. Biotechnol. 19 (2001) 1029–1034.[7] Y. Le, W. Gong, B. Li, N.M. Dunlop, W. Shen, S.B. Su, et al., Utilization of two
seven-transmembrane, G protein-coupled receptors, formyl peptide receptor-like 1 and formyl peptide receptor, by the synthetic hexapeptide WKYMVm forhuman phagocyte activation, J. Immunol. 163 (1999) 6777–6784.
[8] Y. Le, P.M. Murphy, J.M. Wang, Formyl-peptide receptors revisited, TrendsImmunol. 23 (2002) 541–548.
[9] S.H. Baek, J.K. Seo, C.B. Chae, P.G. Suh, S.H. Ryu, Identification of the peptidesthat stimulate the phosphoinositide hydrolysis in lymphocyte cell lines frompeptide libraries, J. Biol. Chem. 271 (1996) 8170–8175.
[10] R. Koczulla, G. von Degenfeld, C. Kupatt, F. Krotz, S. Zahler, T. Gloe, et al., Anangiogenic role for the human peptide antibiotic LL-37/hCAP-18, J. Clin.Investig. 111 (2003) 1665–1672.
[11] M.S. Lee, S.A. Yoo, C.S. Cho, P.G. Suh, W.U. Kim, S.H. Ryu, Serum amyloid Abinding to formyl peptide receptor-like 1 induces synovial hyperplasia andangiogenesis, J. Immunol. 177 (2006) 5585–5594.
[12] S.C. Heo, Y.W. Kwon, I.H. Jang, G.O. Jeong, J.W. Yoon, C.D. Kim, et al.,WKYMVm-induced activation of formyl peptide receptor 2 stimulatesischemic neovasculogenesis by promoting homing of endothelial colony-forming cells, Stem Cells 32 (2014) 779–790.
[13] W.R. Gombotz, D.K. Pettit, Biodegradable polymers for protein and peptidedrug delivery, Bioconjug. Chem. 6 (1995) 332–351.
[14] M.F. Goosen, G.M. O’Shea, H.M. Gharapetian, S. Chou, A.M. Sun, Optimizationof microencapsulation parameters: semipermeable microcapsules as abioartificial pancreas, Biotechnol. Bioeng. 27 (1985) 146–150.
[15] L. Degling, P. Stjarnkvist, I. Sjoholm, Interferon-gamma in starchmicroparticles: nitric oxide-generating activity in vitro and antileishmanialeffect in mice, Pharm. Res. 10 (1993) 783–790.
[16] B.J. Ficek, N.A. Peppas, Novel preparation of poly(vinyl alcohol) microparticleswithout cross-linking agent for controlled drug-delivery of proteins, J. Control.Release 27 (1993) 259–264.
[17] H.K. Makadia, S.J. Siegel, Poly lactic-co-glycolic acid (PLGA) as biodegradablecontrolled drug delivery carrier, Polymers 3 (2011) 1377–1397.
[18] R.A. Jain, The manufacturing techniques of various drug loaded biodegradablepoly(lactide-co-glycolide) (PLGA) devices, Biomaterials 21 (2000) 2475–2490.
[19] F. Danhier, E. Ansorena, J.M. Silva, R. Coco, A. Le Breton, V. Preat, PLGA-basednanoparticles: an overview of biomedical applications, J. Control. Release 161(2012) 505–522.
[20] L.G. Rodriguez, X. Wu, J.L. Guan, Wound-healing assay, Methods Mol. Biol. 294(2005) 23–29.
[21] C.C. Liang, A.Y. Park, J.L. Guan, In vitro scratch assay: a convenient andinexpensive method for analysis of cell migration in vitro, Nat. Protoc. 2 (2007)329–333.
[22] J. Alroy, V. Goyal, E. Skutelsky, Lectin histochemistry of mammalianendothelium, Histochemistry 86 (1987) 603–607.
[23] C. Kalka, H. Masuda, T. Takahashi, W.M. Kalka-Moll, M. Silver, M.Kearney, et al., Transplantation of ex vivo expanded endothelialprogenitor cells for therapeutic neovascularization, Proc. Natl. Acad. Sci.U.S.A. 97 (2000) 3422–3427.
[24] S. Rafii, Circulating endothelial precursors: mystery, reality, and promise, J.Clin. Investig. 105 (2000) 17–19.
[25] F. Cattaneo, M. Parisi, R. Ammendola, Distinct signaling cascades elicited bydifferent formyl peptide receptor 2 (FPR2) agonists, Int. J. Mol. Sci. 14 (2013)7193–7230.
[26] Z. Dong, F. An, T. Wu, C. Zhang, M. Zhang, Y. Zhang, et al., PTX3, a keycomponent of innate immunity, is induced by SAA via FPRL1-mediatedsignaling in HAECs, J. Cell. Biochem. 112 (2011) 2097–2105.
Please cite this article in press as: Y.H. Choi et al., Injectable PLGA microsphere(2015), http://dx.doi.org/10.1016/j.actbio.2015.07.033
[27] T. Kucharzik, A.T. Gewirtz, D. Merlin, J.L. Madara, I.R. Williams, Lateralmembrane LXA4 receptors mediate LXA4’s anti-inflammatory actions onintestinal epithelium, Am. J. Physiol. Cell Physiol. 284 (2003) C888–C896.
[28] Q. Chen, D. Wade, K. Kurosaka, Z.Y. Wang, J.J. Oppenheim, D. Yang, Temporin Aand related frog antimicrobial peptides use formyl peptide receptor-like 1 as areceptor to chemoattract phagocytes, J. Immunol. 173 (2004) 2652–2659.
[29] Y.S. Bae, H.Y. Lee, E.J. Jo, J.I. Kim, H.K. Kang, R.D. Ye, et al., Identification ofpeptides that antagonize formyl peptide receptor-like 1-mediated signaling, J.Immunol. 173 (2004) 607–614.
[30] J.K. Seo, S.Y. Choi, Y. Kim, S.H. Baek, K.T. Kim, C.B. Chae, et al., A peptide withunique receptor specificity: stimulation of phosphoinositide hydrolysis andinduction of superoxide generation in human neutrophils, J. Immunol. 158(1997) 1895–1901.
[31] T. Christophe, A. Karlsson, C. Dugave, M.J. Rabiet, F. Boulay, C. Dahlgren, Thesynthetic peptide Trp-Lys-Tyr-Met-Val-Met-NH2 specifically activatesneutrophils through FPRL1/lipoxin A4 receptors and is an agonist for theorphan monocyte-expressed chemoattractant receptor FPRL2, J. Biol. Chem.276 (2001) 21585–21593.
[32] S. Park, S.D. Kim, H.Y. Lee, D. Hwang, J.S. Park, Y.S. Bae, et al., A novel deliveryplatform for therapeutic peptides, Biochem. Biophys. Res. Commun. 450(2014) 13–18.
[33] J.W. McGinity, P.B. O’Donnell, Preparation of microspheres by the solventevaporation technique, Adv. Drug Deliv. Rev. 28 (1997) 25–42.
[34] S.D. Kim, Y.K. Kim, H.Y. Lee, Y.S. Kim, S.G. Jeon, S.H. Baek, et al., The agonists offormyl peptide receptors prevent development of severe sepsis after microbialinfection, J. Immunol. 185 (2010) 4302–4310.
[35] F.M. Veronese, G. Pasut, PEGylation, successful approach to drug delivery, DrugDiscov. Today 10 (2005) 1451–1458.
[36] Y. Malam, M. Loizidou, A.M. Seifalian, Liposomes and nanoparticles: nanosizedvehicles for drug delivery in cancer, Trends Pharmacol. Sci. 30 (2009) 592–599.
[37] C.B. Packhaeuser, J. Schnieders, C.G. Oster, T. Kissel, In situ forming parenteraldrug delivery systems: an overview, Eur. J. Pharm. Biopharm. 58 (2004) 445–455.
[38] J. Hur, C.H. Yoon, H.S. Kim, J.H. Choi, H.J. Kang, K.K. Hwang, et al.,Characterization of two types of endothelial progenitor cells and theirdifferent contributions to neovasculogenesis, Arterioscler. Thromb. Vasc.Biol. 24 (2004) 288–293.
[39] D.A. Ingram, L.E. Mead, H. Tanaka, V. Meade, A. Fenoglio, K. Mortell, et al.,Identification of a novel hierarchy of endothelial progenitor cells using humanperipheral and umbilical cord blood, Blood 104 (2004) 2752–2760.
[40] D.A. Ingram, N.M. Caplice, M.C. Yoder, Unresolved questions, changingdefinitions, and novel paradigms for defining endothelial progenitor cells,Blood 106 (2005) 1525–1531.
[41] G. Krenning, M.J. van Luyn, M.C. Harmsen, Endothelial progenitor cell-basedneovascularization: implications for therapy, Trends Mol. Med. 15 (2009) 180–189.
[42] A. Zampetaki, J.P. Kirton, Q. Xu, Vascular repair by endothelial progenitor cells,Cardiovasc. Res. 78 (2008) 413–421.
[43] S. Xu, J. Zhu, L. Yu, G. Fu, Endothelial progenitor cells: current development oftheir paracrine factors in cardiovascular therapy, J. Cardiovasc. Pharmacol. 59(2012) 387–396.
[44] T. Asahara, T. Takahashi, H. Masuda, C. Kalka, D. Chen, H. Iwaguro, et al., VEGFcontributes to postnatal neovascularization by mobilizing bone marrow-derived endothelial progenitor cells, EMBO J. 18 (1999) 3964–3972.
[45] K. Hattori, S. Dias, B. Heissig, N.R. Hackett, D. Lyden, M. Tateno, et al., Vascularendothelial growth factor and angiopoietin-1 stimulate postnatalhematopoiesis by recruitment of vasculogenic and hematopoietic stem cells,J. Exp. Med. 193 (2001) 1005–1014.
[46] C. Heeschen, A. Aicher, R. Lehmann, S. Fichtlscherer, M. Vasa, C. Urbich, et al.,Erythropoietin is a potent physiologic stimulus for endothelial progenitor cellmobilization, Blood 102 (2003) 1340–1346.
[47] P. Carmeliet, Angiogenesis in health and disease, Nat. Med. 9 (2003) 653–660.[48] P. Carmeliet, Mechanisms of angiogenesis and arteriogenesis, Nat. Med. 6
(2000) 389–395.[49] M. Heil, W. Schaper, Influence of mechanical, cellular, and molecular factors on
collateral artery growth (arteriogenesis), Circ. Res. 95 (2004) 449–458.[50] F. Pipp, S. Boehm, W.J. Cai, F. Adili, B. Ziegler, G. Karanovic, et al., Elevated fluid
shear stress enhances postocclusive collateral artery growth and geneexpression in the pig hind limb, Arterioscler. Thromb. Vasc. Biol. 24 (2004)1664–1668.
[51] T. Asahara, T. Murohara, A. Sullivan, M. Silver, R. van der Zee, T. Li, et al.,Isolation of putative progenitor endothelial cells for angiogenesis, Science 275(1997) 964–967.
[52] T. Asahara, H. Masuda, T. Takahashi, C. Kalka, C. Pastore, M. Silver, et al., Bonemarrow origin of endothelial progenitor cells responsible for postnatalvasculogenesis in physiological and pathological neovascularization, Circ.Res. 85 (1999) 221–228.
[53] N.M. Caplice, B. Doyle, Vascular progenitor cells: origin and mechanisms ofmobilization, differentiation, integration, and vasculogenesis, Stem Cells Dev.14 (2005) 122–139.
s encapsulating WKYMVm peptide for neovascularization, Acta Biomater.