functional modeling of astrocytes in epilepsy: a feedback system perspective
TRANSCRIPT

ISNN 2010
Functional modeling of astrocytes in epilepsy:a feedback system perspective
Mahmood Amiri • Fariba Bahrami •
Mahyar Janahmadi
Received: 3 March 2010 / Accepted: 19 October 2010 / Published online: 9 November 2010
� Springer-Verlag London Limited 2010
Abstract Astrocytes, a subtype of glial cells, in the brain
provide structural and metabolic supports to the nervous
system. They are also active partners in synaptic trans-
mission and neuronal activities. In the present study, a
biologically plausible thalamocortical neural population
model (TCM) originally proposed by Suffczynski et al.
(Neuroscience 126(2):467–484, 2004) is extended by
integrating the functional role of astrocytes in the regula-
tion of synaptic transmission. Therefore, the original TCM
is modified to consider neuron-astrocyte interactions.
Using the modified model, it is demonstrated that the
healthy astrocytes are capable to compensate the variation
of cortical excitatory input by increasing their firing fre-
quency. In this way, they can preserve the attractor corre-
sponding to the normal activity. Furthermore, the
performance of the pathological astrocytes is also investi-
gated. It is hypothesized that one of the plausible causes of
seizures is the malfunction of astrocytes in the regulatory
feedback loop. That is, pathologic astrocytes are not any
more able to regulate and/or compensate the excessive
increase of the cortical input. Therefore, pathologic astro-
cytes lead to the emergence of paroxysmal attractor.
Results demonstrate that disruption of the homeostatic or
signaling function of astrocytes can initiate the synchro-
nous firing of neurons, suggesting that astrocytes might be
one of the potential targets for the treatment of epilepsy.
Keywords Astrocyte � Epilepsy �Thalamocortical model � Functional modeling
1 Introduction
Epilepsy is one of the most prevalent serious neurological
disorders, which affects more than 50 million people
worldwide. It is characterized by a sudden and intermittent
occurrence of synchronous activity which is termed seizure
or ictal event that impairs the normal function of the brain
[1]. These seizures are transient signs of hypersynchronous
neuronal activity in the brain. Several experimental and
clinical findings conducted over the past decade have
revealed several underlying mechanisms involved for epi-
lepsy [1, 2]. However, many issues are still under investi-
gation. There are now increasing evidences that an
improved understanding of the epileptic process can be
achieved through analysis of bidirectional interactions
between astrocytes and neuronal cells [3, 4]. Astrocytes are
the most abundant type and the best studied glial cells
which are connected together by gap junctions forming a
large functional syncytium. In different sites of brain, each
astrocyte can contact 100,000 synapses. Theses synapses
belong to, on average, six neurons and form ‘‘synaptic
islands’’ [3, 5]. Astrocytes regulate neurotransmitter
release, modulate synaptic transmission and control extra-
cellular fluid content and ion homeostasis [6]. Although
astrocytes cannot generate action potentials, they respond
to neuronal activities with an elevation of their intracellular
M. Amiri (&)
School of Electrical and Computer Engineering,
College of Engineering, University of Tehran, Tehran, Iran
e-mail: [email protected]
F. Bahrami
CIPCE, School of Electrical and Computer Engineering,
College of Engineering, University of Tehran, Tehran, Iran
e-mail: [email protected]
M. Janahmadi
Neuroscience Research Center and Department of Physiology,
Medical School, Shahid Beheshti Medical Sciences University,
Tehran, Iran
123
Neural Comput & Applic (2011) 20:1131–1139
DOI 10.1007/s00521-010-0479-0

calcium levels. In this way, not only astrocytes can sense
neuronal transmission but also their calcium elevation
leads to release of gliotransmitters including glutamate or
ATP (adenosine triphosphate), thereby can control synaptic
strength of neighboring neurons [7, 8]. This fact led to the
concept of the ‘‘tripartite synapse’’ [9–11] in which the
astrocyte, a third active element of the synapse, ‘‘listens
and responds’’ to the synapse [12, 13].
Computational models with biological plausibility are
interesting due to their capability to describe neurological
phenomena at various levels of complexity from cellular to
neural population level [14]. They can be utilized to
understand the basic mechanisms of neuron-astrocyte
interactions and for analyzing the effects of varying neu-
rological parameters on the dynamical characteristics of
model. All this will facilitate to get more thorough under-
standing of the behavior of the original system. In addition,
computational models are helpful to design experiments to
validate the resulted novel insights [15]. An increasing
number of models have been proposed to describe the loop
of information exchange between astrocytes and neurons.
Nadkarni and Jung proposed ‘‘dressed neuron’’ model and
provided a mathematical framework for the synaptic inter-
actions between neurons and astrocytes in the tripartite
synapse [16–18]. A general and nondimensional model for
the tripartite synapse is proposed by Postnov and colleagues
[19]. Recently this model was modified in order to apply it
to the spatially extended neuron-astrocyte network [20].
Garbo [21] proposed a minimal model consisting of a
pyramidal neuron, an interneuron and an astrocyte and
studied how the presence of ATP and interneuron affect the
overall neural activities. Ullah et al. considered the gap
junction mediated calcium waves and investigated coordi-
nation of cytosolic calcium oscillations in two coupled
astrocytes [22]. Finally, some of the other researchers
studied a single glial cell dynamics and characterized its
different calcium-signaling responses [23, 24].
Recent physiological findings show how astrocytes
regulate the synaptic strength and synaptic transmission via
uptake of neurotransmitters or release of gliotransmitters
[6–13]. In the light of these findings, in the present study, a
thalamocortical neural population model (TCM) while was
proposed originally by Suffczynski et al. [25] is modified
by integrating the role of astrocytes in the model under
normal and pathological conditions. The modified TCM
(MTCM) is then used to understand one of the basic
functional mechanisms that cause epileptic seizure. The
TCM describes the neuro-electrophysiology of cortical and
thalamic neural populations and was constructed at an
intermediate level. In this way, instead of simulating the
explicit behavior of individual neurons, a network of
interacting populations was considered. This approach
allows investigating the dynamical properties of system
and the role played by different mechanisms in normal and
pathological conditions more easily. The feedback mech-
anism organized by astrocytes in the structure of the
MTCM, ensures normal asynchronous behavior in spite of
being subjected to those abnormal perturbations inducing
synchronous activities. However, disturbing the mecha-
nism of synaptic transmission can lead to epilepsy. Con-
sequently, dysfunction in the feedback actions generated by
astrocytes can also produce abnormal hypersynchronized
oscillations and seizure-like activities.
The rest of the paper is organized as follows: in Sect. 2,
the mathematical description of the astrocyte model is
explained. The original TCM and its modified version i.e.
MTCM which integrates the role of astrocytes in regulation
of synaptic strengths are presented in Sect. 3. The results of
some simulations are discussed in Sect. 4. Finally, Sect. 5
concludes the paper.
2 Astrocyte model
Astrocytes are vital for normal neuronal functioning and
survival. They have a large number of receptors that are
used to get information about synaptic activity. Although
astrocytes do not have adequate voltage-gated sodium
channels to generate action potential, they are excitable
with respect to intracellular calcium [3, 8].
At the cellular level, the main mechanisms underlying
the tripartite synapse are as follows: neurotransmitters
(such as glutamate) released from the presynaptic neuron
are partially bound to the metabotrobic glutamate receptors
(mGluR) of the synaptic astrocytes. This leads to produc-
tion of inositol (1,4,5)-trisphosphate (IP3) which, in turn,
lead to the release of Ca2? into the astrocytic cytoplasm
from endoplasmic reticulum (ER). These calcium eleva-
tions propagate into nearby astrocytes as intercellular cal-
cium waves with the passage of second messengers through
gap junctions [6, 26]. As a consequence of the increased
intracellular Ca2? concentration, astrocyte releases glio-
transmitters including glutamate and ATP into the extra-
cellular space. The released gliotransmitters feedback onto
the pre and postsynaptic terminals, which implies that
astrocytes regulate synaptic information transfer [10–12].
To model the dynamics of the intracellular Ca2? waves
produced by astrocytes, a recently introduced dynamic
model of the astrocyte [19, 20] is used. This is a general
and simplified mathematical model for a small neuron-
astrocyte ensemble and considers the main pathways of
neuron-astrocyte interactions. The model will be useful to
study the main types of astrocyte responses and the
resulting dynamical patterns. This will allow us to predict
the changes in dynamical patterns with varying control
parameters, which will be introduced later in this section.
1132 Neural Comput & Applic (2011) 20:1131–1139
123

This model is explained with the following set of equations
[19, 20]:
scdc
dt¼ �c� c4f ðc; ceÞ þ ðr þ bSmÞ ð1Þ
ec scdce
dt¼ f ðc; ceÞ ð2Þ
f ðc; ceÞ ¼ c1
c2
1þ c2� c2
e
1þ c2e
� �c4
c42 þ c4
� �� c3ce ð3Þ
sSm
dSm
dt¼ 1þ tan h sSmðz� hSmÞ½ �ð Þ � 1� Smð Þ � Sm
dSm
ð4Þ
sGm
dGm
dt¼ 1þ tan h sGmðc� hGmÞ½ �ð Þ � 1� Gmð Þ � Gm
dGm
ð5Þ
where c is the calcium concentration in the astrocytic cyto-
plasm. ce denotes the calcium concentration within the
endoplasmic reticulum. The parameters ec and sc together
define the characteristic time for calcium oscillations. The
calcium influx from the extracellular space is sensitive to the
production of secondary messenger Sm (IP3), which is con-
trolled by the factor b. The initial state of the calcium oscil-
lation is controlled by the parameter r. The calcium exchange
between the cytoplasm and the endoplasmic reticulum is
defined by the nonlinear function f (c, ce). We set the control
parameters sSm; sGm; sSm; sGm; hSm; hGm; dSm; dGm to the
values used by [19]. An increase in the cytoplasmic calcium
concentration causes release of Gm, an astrocytic mediator.
In terms of the thalamocortical network model, Gm is the
output of the astrocyte. The interaction between astrocyte
and subpopulations is denoted with the parameter z that
shows the synaptic activity of the subpopulations of PY
(pyramidal) and IN (interneuron).
3 Modified thalamocortical model
In the present paper, we consider a lumped neural popu-
lation model based on the thalamocortical model (TCM)
proposed by Suffczynski et al. [25]. The TCM considers
the basic components involved in absence seizures and
therefore consists of two main modules: the thalamic
module and the cortical. Each individual module includes
two mutually interconnected neuronal subpopulations. This
model helps to explore the role of astrocytes in epilepsy
from a macroscopic point of view.
The thalamic module consists of RE (reticular thalamic)
and TC (thalamocortical) subpopulations and the cortical
module consists of PY (pyramidal cell) and IN (inter-
neuron) subpopulations. The interaction between these
subpopulations is facilitated via AMPA (a-amino-3-
hydroxy-5-methyl-4isoxazolepropionic acid) mediated the
fast excitatory synapses and GABAA (c-aminobutric acid)
and GABAB mediated both the fast and slow inhibitory
synapses. The schematic diagram of the TCM is shown in
Fig. 1a. Each subpopulation in the model denotes a lump of
neurons that share the same membrane voltage in the cor-
responding area. It is described by two variables: membrane
potential and firing rate. A sigmoid transfer function is used
to convert the mean membrane potential of the subpopu-
lation to its firing rate to make the output of that population.
In the thalamic subpopulation, for transformation between
mean membrane potential and firing density, the low-
threshold calcium current, IT, that underlies burst firing in
the thalamic cells is also considered. This current plays an
important role in the pathophysiology of epileptic seizures
in animals and humans [27, 28]. The intra-TCM inter-
actions, i.e., the interactions between the PY and the IN
subpopulations, are modeled by constant coupling gains.
The network has three external inputs. The pyramidal
population receives excitatory (glutamergic) input from
other cortical pyramidal neurons not included in the popu-
lation. The TC subpopulation receives excitatory input that
represents sensory inputs from the ascending afferents. RE
cells receives inhibitory inputs from the neighboring RE
cells because they are interconnected by mutual inhibitory
synapses. Cortical input PcxðtÞ is modeled by non-zero
mean white Gaussian noise which was mentioned before is
the output of other cortical pyramidal cells not incorporated
in the model. The output of the TCM is the mean membrane
potential of the pyramidal cells which simulates experi-
mental recordings of the local field potentials. Selecting
suitable mean value of the input signal PcxðtÞh ið Þ, the TCM
demonstrates normal activity. For further details, readers
are referred to [25, 27].
As previously was discussed, astrocytes monitor activ-
ity-dependent changes in the chemical environment of the
extracellular space shared with neurons and play a
dynamic role in regulation of synaptic transmission. It is
well established that transmitters released from neurons
can stimulate astrocyte, leading to release of glutamate,
ATP and other neuroactive substances from the astrocyte.
These gliotransmitters can affect neuronal excitability by
facilitating or by suppressing synaptic transmissions
between neurons [6, 7, 12]. To show the astrocyte-
dependent regulation of neural activity, the TCM is
extended and a new block that represents astrocytes is
added to the original TCM. This block indicates a popu-
lation of astrocytes that regulate internal synapses between
pyramidal and interneuron subpopulations. The modified
TCM (called MTCM) is shown in Fig. 1b. It should be
mentioned that all model parameters such as synaptic
kinetics, sigmoid parameters are similar to the original
TCM [25].
Neural Comput & Applic (2011) 20:1131–1139 1133
123

To monitor the synaptic activity, firing rate of the mean
pyramidal neuronal subpopulation FPYðtÞh i is estimated
from the actual firing rate FPYðtÞ as follows [14, 29]:
d FpyðtÞh idt
¼ a FPYðtÞ � FpyðtÞh i� �
ð6Þ
Based on the results of [14, 29], a time constant of 2 s
(a = 0.5) is used. To remove the DC level of the input
signal of the astrocytes, the estimated mean firing rate
FPYðtÞh ið Þ is subtracted from the actual firing rate FPYðtÞð Þ.The obtained signal is multiplied by the gain Ks and
constitutes the input signal of the astrocytes. This, in fact,
is equivalent to monitor the synaptic transmission by the
astrocytes in the nervous system. The astrocyte outputs
should control and regulate the synaptic activities of the
pyramidal and interneuron cells [11, 12]. This is modeled
by adjusting the strength of interneuron to pyramidal
neuron subpopulations inhibitory gain ðCIN!PYÞ and the
strength of pyramidal neuron to interneuron subpopulations
excitatory gain ðCPY!INÞ as follows:
CIN!PYðtÞ ¼ C�IN!PY þ KiðAs OutðtÞÞ ð7Þ
CPY!INðtÞ ¼ C�PY!IN � KeðAs OutðtÞÞ ð8Þ
where As OutðtÞis the output of the astrocytes and C�IN!PY
and C�PY!IN have the same nominal values as in the original
TCM. In other words, in the original TCM, the model
parameters including CIN!PYðtÞ and CPY!INðtÞ are constant
(they are set in nominal values) and the pyramidal neuron
subpopulation output changes with PCxh i; thereby the ori-
ginal TCM is called static: whereas, in the MTCM these
synaptic strengths are dynamically regulated by the astro-
cytes. In the feedback structure of the MTCM, the astro-
cytes control the balance between the excitation and
inhibition levels so that the pyramidal cell firing rate is
maintained in a normal range and accordingly compensates
the increase or decrease of the excitation and inhibition
levels.
4 Simulation results and discussions
In this section the result of the simulations are presented to
explore the role of normal and pathological astrocytes. It
should be pointed out that all the simulations were imple-
mented in Simulink on a personal computer with Intel(R)
Core(TM) 2 Duo CPU 2.66 GHz.
The TCM is a computational model of absence seizures.
Such seizures are paroxysmal losses of consciousness that
start and end suddenly and are accompanied by bilaterally
synchronous rhythmic spike and wave (SW) discharges
that can be recorded on the EEG. Based on the model
simulations and system analysis in [25], it was verified that
the TCM exhibits bistable dynamics, i.e., the normal and
paroxysmal state coexist for the same set of parameters.
Noise present in the network may switch the system
between these two types of behavior. Paroxysmal dis-
charges characterized by large amplitude oscillations take
place and finish spontaneously due to changes in the
intrinsic system parameters of the population [27].
Results from re-simulations of the static TCM
(CIN!PYðtÞ ¼ C�IN!PY and CPY!INðtÞ ¼ C�PY!IN) are shown
in Fig. 2a. In the static TCM, the firing-rate of pyramidal
neuron subpopulation increases and/or decreases with
cortical input PCxh i. Figure 2a clearly shows that changing
PCxh i alters the model output significantly since it has no
alternative to model the neural homeostasis which is
essential for the normal operation of the brain. However,
synaptic transmission and intrinsic properties must be
tuned through release or/and uptake of neurotransmitters
and ions in order to maintain the stable ongoing activity in
Fig. 1 a The original TCM
proposed by Suffczynski et al.
[25]. b The modified TCM
(MTCM) in which astrocytes
dynamically regulate the
synaptic strengths between
pyramidal and interneuron
subpopulations
1134 Neural Comput & Applic (2011) 20:1131–1139
123

neural circuits. Recent studies on communications between
astrocytes and neurons revealed that glutamate release
from single astrocyte may control simultaneously the
excitability of several neighboring pyramidal cells [30].
Figure 2b illustrates the output of MTCM with dynamic
gains CIN!PYðtÞ and CPY!INðtÞ for the changing input
PCxh i. It is observed that the astrocytes modify the synaptic
strengths appropriately and maintain the balance of exci-
tation and inhibition levels in order to track the changes of
PCxh i. This will prevent from the occurrence of spontane-
ous paroxysmal episodes. For the simulations shown in
Fig. 2b the value of the parameters are as follows:
Ks ¼ 3:5, Ki ¼ 3 and Ke ¼ 3:1. For this case, the output of
the astrocytes and the concentration of the intracellular
calcium are shown in Fig. 2c. This figure demonstrates that
by changing the value of PCxh i, the astrocytes regulate and
control the synaptic strengths properly by changing their
firing frequency. Increasing/decreasing PCxh i leads to the
increase/decrease of the firing rate of the astrocyte output.
These simulation data are compatible with experimental
findings showing that astrocytes modulate and regulate the
synaptic activities [6, 11, 12].
In addition to the physiological role of astrocytes in the
control of neuronal excitability and synaptic transmission,
they can contribute to the disorders of nervous system
[4, 30]. Experimental findings demonstrated that patho-
logical astrocytes undergo structural alterations which are
accompanied by variation in their functions [31]. These
findings imply that alterations in astrocyte properties,
specifically glutamate, K? and Ca2? homeostasis, could be
part of the pathophysiology of neuronal hyperexcitability,
which is a characteristic of epilepsy [4, 32]. Hence, here a
pathological condition is considered in which astrocytes
cannot carry out their responsibilities properly. One way to
simulate pathological astrocytes is simply reducing the
capability of astrocytes in monitoring the synaptic activi-
ties. This is physiologically corresponding to deficit in the
structure of astrocyte receptors or scarcity in bindings of
neurotransmitters with astrocyte receptors. This is simu-
lated by decreasing the value of Ks from 3.5 to 1.5 in the
MTCM with the same changing input PCxh i as in Fig. 2b.
The simulation for pathological astrocytes is shown in
Fig. 2d. It is hypothesized that one of the plausible cause of
seizures is pathology in the feedback structure generated by
the astrocytes. In this case, astrocytes cannot regulate and/
or compensate excessive increase of the cortical input
through release of gliotransmitters and/or uptake of neu-
rotransmitters and ions. This is in accordance with the
physiological findings showing that paroxysmal depolar-
ization can be induced by pathological astrocytes in neu-
rons through calcium signaling pathway [3, 30, 33]. Indeed,
before the development of seizures and in the absence of
other pathological changes, hypertrophy of astrocytes
during the process of epileptogenesis has been reported
[34].
To obtain more insights into the role of astrocytes in the
active control of neuronal activity and synaptic neuro-
transmission, the excitation level is plotted versus inhibi-
tion level of pyramidal cells for (a) the MTCM with
healthy astrocytes and (b) the MTCM with pathological
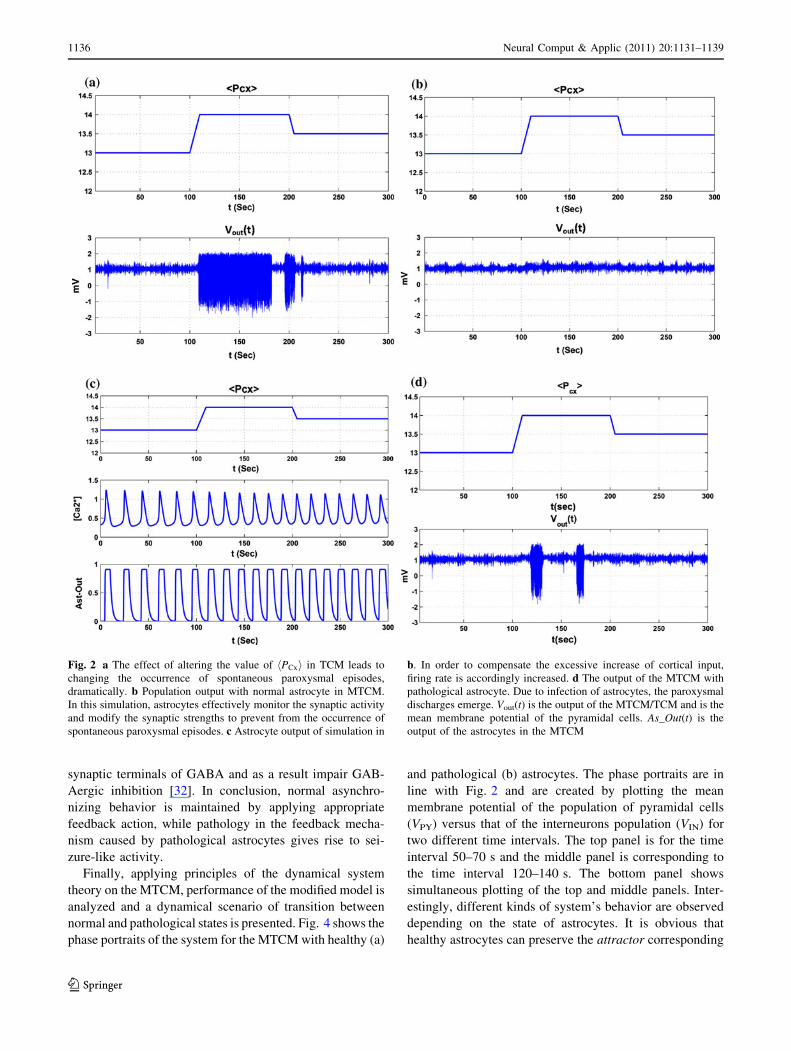
astrocytes in Fig. 3. The top (bottom) panels are related to
the time interval 50–60 s (120–130 s) of the corresponding
model in Fig. 2. In the context of epilepsy, there is a par-
ticular interest to the balance between excitation and
inhibition levels. In fact, numerous researches have shown
that ictal spike discharges initiate in an area of the cortex
that is excessively excitable. This increased excitability is
due to increased excitation, reduced inhibition or both [35,
36]. As a consequence, increasing the ratio of excitation to
inhibition influences the dynamics of the modeled popu-
lation of neurons which can produce sustained discharges
of spikes. In the MTCM, the excitation and inhibition
levels for the cortical module are described by AMPA
mediated excitatory synapses and GABAA and GABAB-
mediated inhibitory synapses. It should be mentioned that
in Fig. 3, for simplicity, only the AMPA excitatory syn-
apses are plotted against the GABAA inhibitory synapses.
As can be seen in Fig. 3a, in spite of abnormal increase
in cortical excitatory input, healthy astrocytes maintain
the balance between excitation and inhibition through
regulation of excitatory and inhibitory neurotransmitter
concentrations in the synaptic cleft. This is achieved via
mechanisms that are responsible for release of gliotrans-
mitters and uptake of neurotransmitters. However, Fig. 3b
shows considerable changes in the level of excitation and
inhibition for the case of pathological astrocytes. Signifi-
cant decrease in the inhibition and to some extent increase
in the excitation levels are the result of the pathological
condition in astrocytes. This is consistent with experi-
mental findings where increased extracellular levels of
glutamate have been found in epileptogenic foci [37].
Hence, it is logical that several drugs impacting
GABAergic synaptic transmission are usually used in the
therapy of epilepsy disorders. In this issue, during gluta-
matergic neurotransmission, glutamate is released by neu-
rons into the synaptic cleft. This glutamate is mainly
exported and taken up into astrocytes by the glutamate
transporters where it is converted into glutamine. Gluta-
mine is then released from astrocytes and in turn is shuttled
back into neurons, where it is converted back to glutamate
[4, 32, 38]. Therefore, due to pathology in astrocytes,
glutamate to glutamine cycling is impaired and this could
contribute to seizures in the epileptic brain. Furthermore,
vesicular GABA levels partially dependent on glutamine
transport into inhibitory neurons. Down-regulation of
glutamine synthetase could partially reduce inhibitory
Neural Comput & Applic (2011) 20:1131–1139 1135
123
synaptic terminals of GABA and as a result impair GAB-
Aergic inhibition [32]. In conclusion, normal asynchro-
nizing behavior is maintained by applying appropriate
feedback action, while pathology in the feedback mecha-
nism caused by pathological astrocytes gives rise to sei-
zure-like activity.
Finally, applying principles of the dynamical system
theory on the MTCM, performance of the modified model is
analyzed and a dynamical scenario of transition between
normal and pathological states is presented. Fig. 4 shows the
phase portraits of the system for the MTCM with healthy (a)
and pathological (b) astrocytes. The phase portraits are in
line with Fig. 2 and are created by plotting the mean
membrane potential of the population of pyramidal cells
(VPY) versus that of the interneurons population (VIN) for
two different time intervals. The top panel is for the time
interval 50–70 s and the middle panel is corresponding to
the time interval 120–140 s. The bottom panel shows
simultaneous plotting of the top and middle panels. Inter-
estingly, different kinds of system’s behavior are observed
depending on the state of astrocytes. It is obvious that
healthy astrocytes can preserve the attractor corresponding
Fig. 2 a The effect of altering the value of PCxh i in TCM leads to
changing the occurrence of spontaneous paroxysmal episodes,
dramatically. b Population output with normal astrocyte in MTCM.
In this simulation, astrocytes effectively monitor the synaptic activity
and modify the synaptic strengths to prevent from the occurrence of
spontaneous paroxysmal episodes. c Astrocyte output of simulation in
b. In order to compensate the excessive increase of cortical input,
firing rate is accordingly increased. d The output of the MTCM with
pathological astrocyte. Due to infection of astrocytes, the paroxysmal
discharges emerge. Vout(t) is the output of the MTCM/TCM and is the
mean membrane potential of the pyramidal cells. As_Out(t) is the
output of the astrocytes in the MTCM
1136 Neural Comput & Applic (2011) 20:1131–1139
123
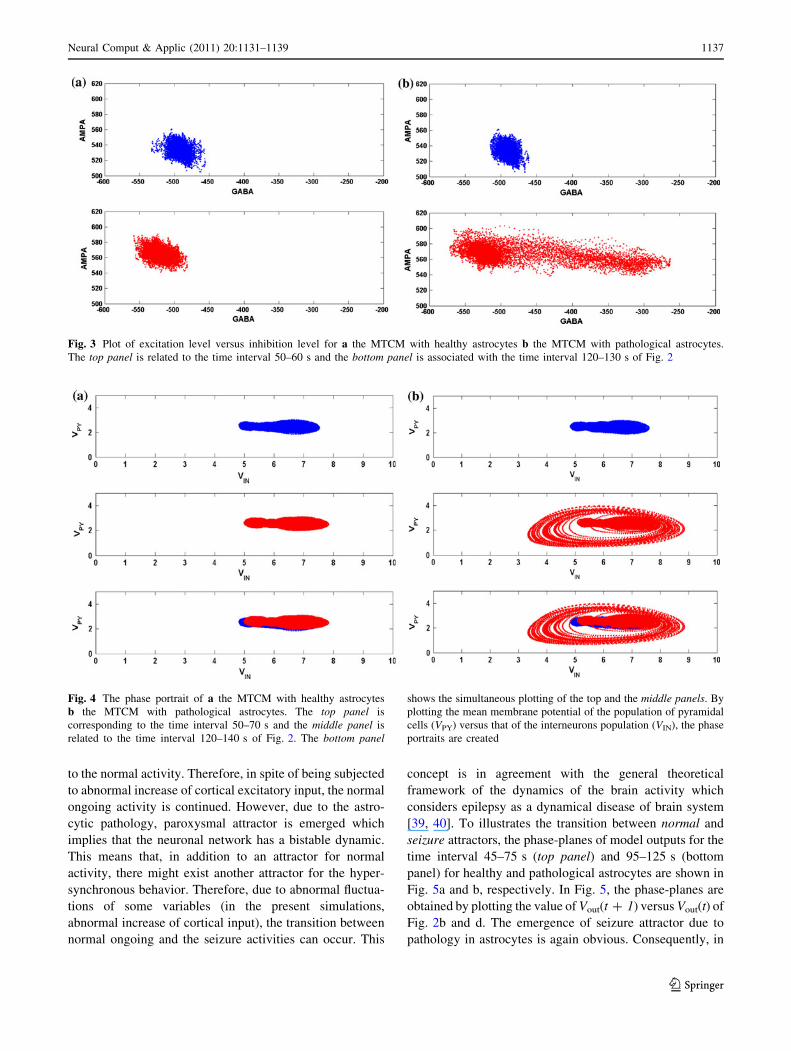
to the normal activity. Therefore, in spite of being subjected
to abnormal increase of cortical excitatory input, the normal
ongoing activity is continued. However, due to the astro-
cytic pathology, paroxysmal attractor is emerged which
implies that the neuronal network has a bistable dynamic.
This means that, in addition to an attractor for normal
activity, there might exist another attractor for the hyper-
synchronous behavior. Therefore, due to abnormal fluctua-
tions of some variables (in the present simulations,
abnormal increase of cortical input), the transition between
normal ongoing and the seizure activities can occur. This
concept is in agreement with the general theoretical
framework of the dynamics of the brain activity which
considers epilepsy as a dynamical disease of brain system
[39, 40]. To illustrates the transition between normal and
seizure attractors, the phase-planes of model outputs for the
time interval 45–75 s (top panel) and 95–125 s (bottom
panel) for healthy and pathological astrocytes are shown in
Fig. 5a and b, respectively. In Fig. 5, the phase-planes are
obtained by plotting the value of Vout(t ? 1) versus Vout(t) of
Fig. 2b and d. The emergence of seizure attractor due to
pathology in astrocytes is again obvious. Consequently, in
Fig. 3 Plot of excitation level versus inhibition level for a the MTCM with healthy astrocytes b the MTCM with pathological astrocytes.
The top panel is related to the time interval 50–60 s and the bottom panel is associated with the time interval 120–130 s of Fig. 2
Fig. 4 The phase portrait of a the MTCM with healthy astrocytes
b the MTCM with pathological astrocytes. The top panel is
corresponding to the time interval 50–70 s and the middle panel is
related to the time interval 120–140 s of Fig. 2. The bottom panel
shows the simultaneous plotting of the top and the middle panels. By
plotting the mean membrane potential of the population of pyramidal
cells (VPY) versus that of the interneurons population (VIN), the phase
portraits are created
Neural Comput & Applic (2011) 20:1131–1139 1137
123
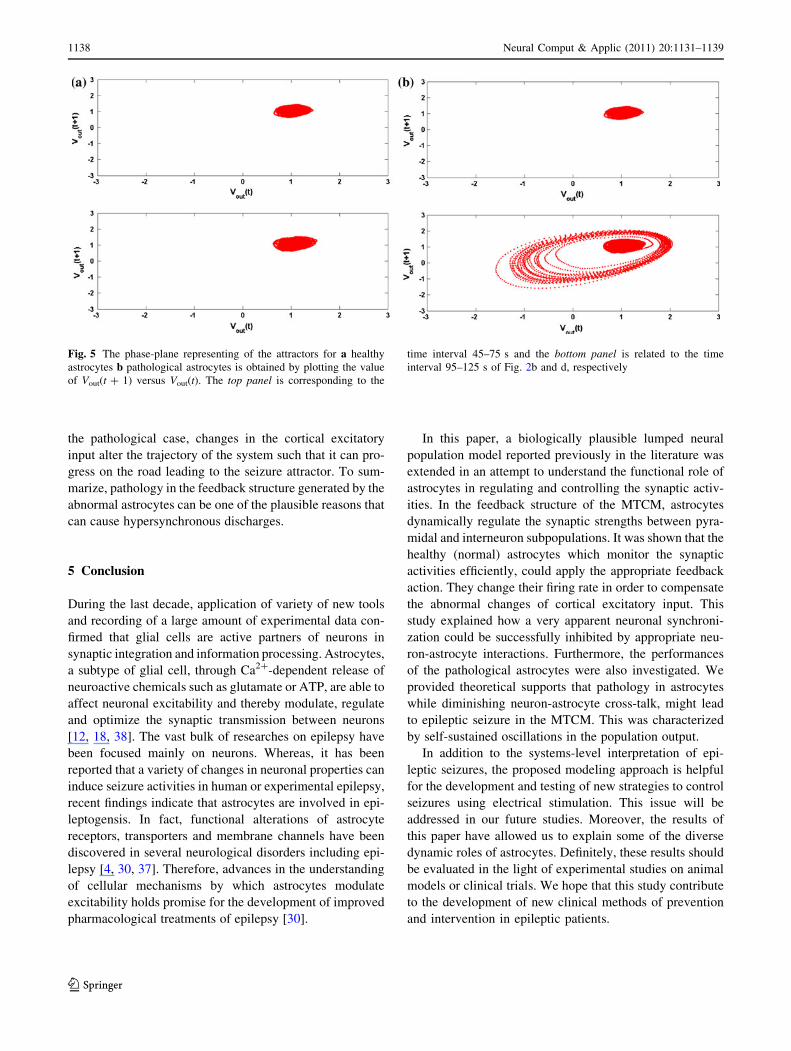
the pathological case, changes in the cortical excitatory
input alter the trajectory of the system such that it can pro-
gress on the road leading to the seizure attractor. To sum-
marize, pathology in the feedback structure generated by the
abnormal astrocytes can be one of the plausible reasons that
can cause hypersynchronous discharges.
5 Conclusion
During the last decade, application of variety of new tools
and recording of a large amount of experimental data con-
firmed that glial cells are active partners of neurons in
synaptic integration and information processing. Astrocytes,
a subtype of glial cell, through Ca2?-dependent release of
neuroactive chemicals such as glutamate or ATP, are able to
affect neuronal excitability and thereby modulate, regulate
and optimize the synaptic transmission between neurons
[12, 18, 38]. The vast bulk of researches on epilepsy have
been focused mainly on neurons. Whereas, it has been
reported that a variety of changes in neuronal properties can
induce seizure activities in human or experimental epilepsy,
recent findings indicate that astrocytes are involved in epi-
leptogensis. In fact, functional alterations of astrocyte
receptors, transporters and membrane channels have been
discovered in several neurological disorders including epi-
lepsy [4, 30, 37]. Therefore, advances in the understanding
of cellular mechanisms by which astrocytes modulate
excitability holds promise for the development of improved
pharmacological treatments of epilepsy [30].
In this paper, a biologically plausible lumped neural
population model reported previously in the literature was
extended in an attempt to understand the functional role of
astrocytes in regulating and controlling the synaptic activ-
ities. In the feedback structure of the MTCM, astrocytes
dynamically regulate the synaptic strengths between pyra-
midal and interneuron subpopulations. It was shown that the
healthy (normal) astrocytes which monitor the synaptic
activities efficiently, could apply the appropriate feedback
action. They change their firing rate in order to compensate
the abnormal changes of cortical excitatory input. This
study explained how a very apparent neuronal synchroni-
zation could be successfully inhibited by appropriate neu-
ron-astrocyte interactions. Furthermore, the performances
of the pathological astrocytes were also investigated. We
provided theoretical supports that pathology in astrocytes
while diminishing neuron-astrocyte cross-talk, might lead
to epileptic seizure in the MTCM. This was characterized
by self-sustained oscillations in the population output.
In addition to the systems-level interpretation of epi-
leptic seizures, the proposed modeling approach is helpful
for the development and testing of new strategies to control
seizures using electrical stimulation. This issue will be
addressed in our future studies. Moreover, the results of
this paper have allowed us to explain some of the diverse
dynamic roles of astrocytes. Definitely, these results should
be evaluated in the light of experimental studies on animal
models or clinical trials. We hope that this study contribute
to the development of new clinical methods of prevention
and intervention in epileptic patients.
Fig. 5 The phase-plane representing of the attractors for a healthy
astrocytes b pathological astrocytes is obtained by plotting the value
of Vout(t ? 1) versus Vout(t). The top panel is corresponding to the
time interval 45–75 s and the bottom panel is related to the time
interval 95–125 s of Fig. 2b and d, respectively
1138 Neural Comput & Applic (2011) 20:1131–1139
123

References
1. Lehnertz K, Bialonski S, Horstmann MT, Krug D, Rothkegel A,
Staniek M, Wagner T (2009) Synchronization phenomena in
human epileptic brain networks. J Neurosci Meth 183(1):42–48
2. Demont-Guignard S, Benquet P, Gerber U, Wendling F (2009)
Analysis of intracerebral EEG recordings of epileptic spikes:
insights from a neural network model. IEEE Trans Biomed Eng
56(12):2782–2795
3. Ricci G, Volpi L, Pasquali L, Petrozzi L, Siciliano G (2009)
Astrocyte-neuron interactions in neurological disorders. J Biol
Phys 35:317–336
4. Seifert G, Carmignoto G, Steinhauser C (in press) Astrocyte
dysfunction in epilepsy. Brian Res Rev
5. Gertrudis P, Araque A (2005) Properties of synaptically evoked
astrocyte calcium signal reveal synaptic information processing
by astrocytes. J Neurosci 25(9):2192–2203
6. Newman EA (2003) New roles for astrocytes: regulation of
synaptic transmission. Trends Neurosci 26:536–542
7. Voltarra A, Steinhauser C (2004) Glial modulation of synaptic
transmission in the hippocampus. GLIA 47:249–257
8. Hertz L, Zielke HR (2004) Astrocytic control of glutamatergic
activity: astrocytes as stars of the show. Trends Neurosci 27(12):
735–743
9. Araque A, Parpura V, Sanzgiri RP, Haydon PG (1999) Tripartite
synapses: glia, the unacknowledged partner. Trends Neurosci
22:208–215
10. Halassa MM, Fellin T, Haydon PG (2007) The tripartite synapse:
roles for gliotransmission in health and disease. Trends Mol Med
13(2):54–63
11. Halassa MM, Fellin T, Haydon PG (2009) Tripartite synapses:
roles for astrocytic purines in the control of synaptic physiology
and behavior. Neuropharmacol 57(4):343–346
12. Fellin T, Pascual O, Haydon PG (2006) Astrocytes coordinate
synaptic networks: balanced excitation and inhibition. Physiology
21:208–215
13. Fellin T, Carmignoto G (2004) Neuron-to-astrocyte signaling in
the brain represents a distinct multifunctional unit. J Physiol 559:
3–15
14. Chakravarthy N, Tsakalis K, Sabesan S, Iasemidis LD (2009)
Homeostasis of brain dynamics in epilepsy: a feedback control
systems perspective of seizures. Ann Biomed Eng 37(3):565–585
15. Suffczynski P, Wendling F, Bellanger JJ, Lopes da Silva FH
(2006) Some insights into computational models of (patho)
physiological brain activity. Proc IEEE 94(4):784–804
16. Nadkarni S, Jung P (2004) Dressed neurons: modeling neural-
glial interactions. Phys Biol 1:35–41
17. Nadkarni S, Jung P (2007) Modeling synaptic transmission of the
tripartite synapse. Phys Biol 4:1–9
18. Nadkarni S, Jung P, Levine H (2008) Astrocytes optimize the
synaptic transmission of information. PLoS Comput Biol 4(5):
1–11
19. Postnove DE, Ryazanova LS, Sosnovtseva OV (2007) Functional
modeling of neural-glial interaction. BioSystems 89:84–91
20. Postnov DE, Koreshkov RN, Brazhe NA, Brazhe AR, Sos-
novtseva OV (2009) Dynamical patterns of calcium signaling in a
functional model of neuron-astrocyte networks. J Biol Phys
35:425–445
21. Garbo AD (2009) Dynamics of a minimal neural model con-
sisting of an astrocyte, a neuron, and an interneuron. J Biol Phys
35:361–382
22. Ullah G, Jung P, Cornell-Bell AH (2006) Anti-phase calcium
oscillations in astrocytes via inositol (1, 4, 5)-trisphosphate
regeneration. Cell Calcium 39:197–208
23. Garbo AD, Barbi M, Chillemi S, Alloisio S, Nobile M (2007)
Calcium signaling in astrocytes and modulation of neural activity.
BioSystems 89:74–83
24. Lavrentovich M, Hemkin SA (2008) Mathematical model of
spontaneous calcium (II) oscillations in astrocytes. J Theoretic
Biol 251(4):553–560
25. Suffczynski P, Kalitzin S, Lopes da Silva FH (2004) Dynamics of
non-convulsive epileptic phenomena modeled by a bistable
neuronal network. Neuroscience 126(2):467–484
26. Hung J, Colicos MA (2008) Astrocytic Ca2? waves guide CNS
growth cones to remote regions of neuronal activity. PLoS ONE
3(11):e3692
27. Suffczynski P, Kalitzin S, Lopes da Silva FH (2008) A neuronal
network model of corticothalamic oscillations: the emergence of
epileptiform absence seizures. In: Soltesz I, Staley K (eds)
Computational neuroscience in epilepsy. Elsevier, Amsterdam,
pp 403–418
28. Destexhe A (2008) Corticothalamic feedback: a key to explain
absence seizures. In: Soltesz I, Staley K (eds) Computational
neuroscience in epilepsy. Elsevier, Amsterdam, pp 184–211
29. Chakravarthy N, Sabesan S, Iasemidis LD, Tsakalis K (2007)
Modeling and controlling synchronization in a neuron level
population model. Int J Neural Syst 17(2):123–138
30. D’Ambrosio R (2004) The role of glial membrane ion channels in
seizures and epileptogenesis. Pharmacol Therapeut 103:95–108
31. Silchenko AN, Tass PA (2008) Computational modeling of par-
oxysmal depolarization shifts in neurons induced by the gluta-
mate release from astrocytes. Biol Cybern 98:61–74
32. Wetherington J, Serrano G, Dingledine R (2008) Astrocytes in
the epileptic brain. Neuron 58:168–178
33. De Keyser J, Mostert JP, Koch MW (2008) Dysfunctional
astrocytes as key players in the pathogenesis of central nervous
system disorders. J Neurol Sci 267:3–16
34. Rogawski MA (2005) Astrocytes get in the act in epilepsy. Nat
Med 11(9):919–920
35. Wendling F, Bellanger JJ, Bartolomei F, Chauvel P (2000) Rel-
evance of nonlinear lumped-parameter models in the analysis of
depth-EEG epileptic signals. Biol Cybern 83(4):367–378
36. Wendling F, Chauvel P (2008) Transition to ictal activity in
temporal lobe epilepsy: insights from macroscopic models. In:
Soltesz I, Staley K (eds) Computational neuroscience in epilepsy.
Elsevier, Amsterdam, pp 356–386
37. Binder DK, Steinhauser C (2009) Role of astrocytes in epilepsy.
astrocytes in (Patho)physiology of the nervous system. Springer,
Berlin, pp 649–671
38. Santello M, Volterra A (2009) Synaptic modulation by astrocytes
via Ca2?-dependent glutamate release. Neuroscience 158(1):
253–259
39. Lopes da Silva FH, Blanes W, Kalitzin SN, Parra J, Suffczynski
P, Velis DN (2003) Epilepsies as dynamical diseases of brain
systems: basic models of the transition between normal and
epileptic activity. Epilepsia 44(12):72–83
40. Lopes da Silva FH (2008) The impact of EEG/MEG signal pro-
cessing and modeling in the diagnostic and management of epi-
lepsy. IEEE Rev Biomed Eng 1:143–156
Neural Comput & Applic (2011) 20:1131–1139 1139
123