contents lists available at sciencedirect
TRANSCRIPT

R
PPi
AC
a
ARRA
KPAIHH
1
go
0d
Veterinary Immunology and Immunopathology 142 (2011) 207– 218
Contents lists available at ScienceDirect
Veterinary Immunology and Immunopathology
j ourna l ho me pag e: www.elsev ier .com/ locate /vet imm
esearch paper
athophysiology of experimental Aeromonas hydrophila infection inuntius sarana: Early changes in blood and aspects of the innatemmune-related gene expression in survivors
. Das, P.K. Sahoo ∗, B.R. Mohanty, J.K. Jenaentral Institute of Freshwater Aquaculture, Kausalyaganga, Bhubaneswar 751 002, India
r t i c l e i n f o
rticle history:eceived 18 February 2011eceived in revised form 30 April 2011ccepted 8 May 2011
eywords:untius saranaeromonas hydrophila
nnate immune responseaematologyistopathology
a b s t r a c t
A decline in production of Puntius sarana worldwide and subsequent intensification ofits culture as a measure for conservation and increase food fish production have led toemergence of diseases. Investigations on innate immunological changes in fish that sur-vives an acute bacterial infection are a few or lacking. An attempt was made to study thedetailed changes in innate immune responses in P. sarana following an intraperitoneal chal-lenge with Aeromonas hydrophila at 2.24 × 107 colony-forming unit (CFU)/fish. Sequentialchanges in transcription of immune related genes in liver and kidney, innate immune status,haematological profile and histopathology of major organs were studied after different timeintervals post-challenge in the survivors. A significant increase in serum myeloperoxidase,ceruloplasmin activities and total leucocyte count, and decline in alternative complementactivity were observed at early infection periods. A significant decline in plasma glucoselevel, total erythrocyte count, haemoglobin content and packed cell volume in blood wasnoticed after challenge. Superoxide radical production and serum antiprotease activityincreased significantly at later period of challenge. In addition, few immune related genespertinent to acute phase reactants and inflammatory response viz., complement component3 (C3), lysozyme G, lysozyme C, �2-microglobulin (B2M), transferrin, interleukin 1�, inter-leukin 8, manganese superoxide dismutase (MnSOD), natural killer cell enhancing factor-Band chemokine CXCa were modulated after bacterial challenge. In liver, a significant up-regulation of C3 and transferrin, and down-regulation of lysozyme G, interleukin 8, MnSODand B2M were noticed in survivors. On the contrary, lysozyme G, lysozyme C, interleukin
1�, interleukin 8, CXCa, and MnSOD were found to be significantly up-regulated and C3was down-regulated in the kidney. All these data suggest that the innate immune factorswere highly modulated during infection process to render protection of P. sarana to thisbacterium. This study also generated the primary information on many innate immunemolecules/genes for this important medium carp species for the first time.. Introduction
Olive barb, Puntius sarana (Hamilton), is an endan-ered medium sized commercially important carp speciesf south-eastern regions of Asia. This fish needs urgent
∗ Corresponding author. Tel.: +91 674 2465407; fax: +91 674 2465407.E-mail address: [email protected] (P.K. Sahoo).
165-2427/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.vetimm.2011.05.017
© 2011 Elsevier B.V. All rights reserved.
attention from conservation viewpoint, as its populationis reported to be on decline (Ghosh et al., 2005). Althoughefforts have been made for breeding, stocking and cultureof this fish, disease is a major setback in its farming andproduction (Jena et al., 2008).
Study of the fish immune system and response ofteleosts to bacterial infections has been greatly expandedin last few years. However, there is no information avail-able on immune system or status of P. sarana. Aeromonas

and Im
208 A. Das et al. / Veterinary Immunologyhydrophila, a Gram-negative bacterial pathogen, is thecausative agent of haemorrhagic septicemia and ulcer-ative syndrome in freshwater fishes including Indiancarps causing high mortalities in hatcheries and culturesystems leading to severe economic loss (Sahoo et al.,2008). Knowledge about immune defence of olive barbagainst A. hydrophila infection is important in terms ofdeveloping any preventive and control measures as thisspecies suffer mostly from secondary bacterial ulcers. Ini-tial reactions to bacterial pathogens depend mainly onnon-specific defence mechanisms. The specific immuneresponses against A. hydrophila bacterins have been studiedby a number of researchers (Vivas et al., 2004; Poobalaneet al., 2010) but the role of innate immune factors dur-ing the progress of A. hydrophila infection in fish is poorlystudied (Rodrıguez et al., 2008; Harikrishnan et al., 2010).Furthermore, the knowledge on interactions between thisbacterium and its host has been obtained mainly fromvaccination studies. However, it is important for futureimprove of vaccines to address basic immunity mecha-nisms conferring protection to a host surviving a primaryinfection (Raida and Buchmann, 2009).
Acute phase response proteins such as comple-ment components, transferrin, ceruloplasmin, lysozyme,antiprotease, etc. are the major players of innate immunityin response to tissue infection or injury (Peatman et al.,2007). Complement system plays a vital role in activationof innate and acquired immunity by phagocytosis, respi-ratory burst, chemotaxis, cell lysis and B-cell proliferation(Boshra et al., 2006). Complement component 3 (C3) isthe central component of the complement cascade thathelps in activation of all the pathways leading to inflam-matory reactions and lysis of the pathogens (Mishra et al.,2009). Transferrin, an iron binding multi-task globularglycoprotein, plays an important role in the nutritionalimmunity, by restricting the iron availability for replicatingpathogens and serves as a primary activating moleculein macrophage antimicrobial response (Stafford andBelosevic, 2003; Sahoo et al., 2009). It is also associatedwith a heterogenous group of serum proteins called ceru-loplasmin released from the hepatocytes in response tohomeostatic disturbances. It helps transferrin in transportof iron and modulates the innate immune system (Siwickiet al., 2006). Lysozyme is strongly bactericidal in nature,primarily associated with defence against Gram-positivebacteria as well as Gram-negative bacteria causing lysis ofthe pathogen. It is also known to be an opsonin and activatethe complement system and phagocytes. Among the fivetypes of lysozymes, C and G-types are found in fishes(Saurabh and Sahoo, 2008). Being an essential componentof the MHC class I molecules �2-microglobulin (B2M)alerts the immune system against bacterial-infected cellsin antigen presentation pathway and is necessary for heavychain folding and peptide binding (Christie et al., 2007).Like mammals, fish have also a well-organised network ofsignalling molecules like cytokines and chemokines whichhelps in controlling the acquired immune system. These
are small, secreted peptides that have the ability to attracta wide range of immune cells to sites of infection anddisease (Liu et al., 2007a,b). Interleukin-1� is a lymphocyte-activating cytokine released from phagocytes, which ismunopathology 142 (2011) 207– 218
vital for initiating specific immune responses (Low et al.,2003), while interleukin 8 is a type of chemokine, whichinduces the neutrophils to release hydrolytic enzymes andprovokes respiratory burst activity and helps in angiogen-esis (Inoue et al., 2003). CXCa belongs to a superfamily ofchemotactic cytokines identified in fish and carps reportedto be induced after bacterial infection. It has a regulatoryeffect on inflammation and angiogenesis (Peatman et al.,2007). In order to prevent the destructive effect of reactiveoxygen species, an antioxidant defence enzyme calledsuperoxide dismutase (SOD) is released in relation tooxidative stress (Ni et al., 2007). It is a metalloenzyme thatconverts superoxide radical and reactive oxygen speciesinto molecular oxygen and hydrogen peroxide.
The present work elucidates the changes in variousinnate immune parameters and transcriptional activitiesof a range of genes involved in the innate defence andcorrelate these parameters with the damage caused by A.hydrophila in the muscle, liver and kidney tissues duringinfection and further getting rid of primary infection by thefish. In the present paper, we used intraperitonial injec-tion to ensure an identical pathogen dose in all animalsand we analyzed the expression of various innate immunefactors including inflammatory cytokines and a series ofacute phase reactants in liver and anterior kidney tissuesof survivors. Furthermore, the ultimate reflections of thesereactions are monitored through studying haematologi-cal picture in a sequential manner after single challengewith bacteria to draw a comprehensive role played bythis bacterium. In our challenge experiments, barbs weresusceptible to infection by intraperitoneal injection of A.hydrophila and revealed evidences for major changes ininnate immune response to get rid of the infection pro-cess. To our knowledge, this is the first investigation to lookin to any immune mechanism in this endangered speciesand a comprehensive study for understanding A. hydrophilapathogenesis.
2. Materials and methods
2.1. Fish
P. sarana (average weight, 85.6 ± 8.2 g), obtained froma mono-culture pond of the Central Institute of Freshwa-ter Aquaculture, Bhubaneswar, India, were maintained inthe wet laboratory for two weeks prior to the challengeexperiment and fed with standard pellet feed at 3% ofbody weight. The fish were kept in fibre-reinforced plastictanks of 500 L capacity under continuous aeration. Aboutone-tenth of the water was siphoned daily to remove thefaecal materials and unused food particles with subsequentwater level adjustment using dechlorinated tap water. Thewater quality parameters viz., temperature 29–31◦C, dis-solved oxygen: 5.65 ± 0.72 mg L−1; pH: 8.2 ± 0.82; nitrites:0.015 ± 0.009 mg L−1; ammonia: 0.109 ± 0.024 mg L−1
were measured during the study.
2.2. Preparation of bacteria
The pathogenic strain of A. hydrophila (Ah # 04/2003)obtained from the Fish Health Management Division of the

and Imm
C1hcsiioLsfit(
2
t2ttctmciwftLlebutpgscuoapAfpaaigt
2
creoew
A. Das et al. / Veterinary Immunology
entral Institute of Freshwater Aquaculture, was grown for8 h in tryptone soy broth in a shaker incubator at 30 ◦C. Thearvested cells were washed, resuspended in PBS and theoncentration was checked by measuring the optical den-ity at 540 nm. The lethal dose (LD50) value of A. hydrophilan this fish species was previously determined by follow-ng Reed and Muench (1938) and the obtained LD50 dosef 2.24 × 107 CFU was used for the challenge study. ForD50 determination, 10-fold dilutions of bacterial suspen-ion were injected intraperitoneally to triplicate groups ofve fish per each dilution. The fish were observed for mor-ality for a period of ten days and the cumulative mortality%) determined at the end to calculate the value.
.3. Experimental design
Thirty-six 500 L capacity of fibre-reinforced plasticanks with three tanks for each time point (0, 1, 3, 6, 12,4, 48 h and 4, 7, 10, and 14 days along with positive con-rol) containing twelve fish in each tank were utilized inhis study. As it was not possible to calculate the per-ent mortality of challenged fish from the experimentalanks, a positive control was taken. The fish in experi-
ental and positive control tanks were intraperitoneallyhallenged with A. hydrophila at the above mentioned dosen 0.1 mL PBS. The negative control group fish were injected
ith 0.1 mL of PBS only. Fifteen fish were bled non-lethallyrom caudal vein (five from each tank at the mentionedime point, after anaesthetizing with MS 222, Sigma, St.ouis, MO, USA) at 0, 2, 4, 7, 10 and 14 days post chal-enge (dpc) and about 1 mL of blood was collected fromach fish using a plastic syringe. An aliquot of 200 �Llood was heparinised (50 IU mL−1) in a separate tube andtilized for analyses of haematological parameters viz.,otal erythrocyte count (TEC), total leucocyte count (TLC),acked cell volume (PCV), haemoglobin (Hb) and plasmalucose level, whereas a portion of non-heparinised bloodample was allowed to clot and serum was separated byentrifuging at 1500 × g at 4 ◦C. The collected serum wassed for studying immunological parameters viz., super-xide production assay, myeloperoxidase (MPO) activity,lternative complement haemolytic activity (ACH50), totalrotein level, antiprotease and ceruloplasmin activities.nterior kidney and liver tissues were aseptically collected
rom three non-bled fish (one from each tank for one timeoint) at each time point (0, 1, 3, 6, 12, 24, 48 hpc and 4, 7nd 14 dpc) and stored in RNAlater for further expressionnalysis of immune related genes. Furthermore, three clin-cally affected moribund fish from A. hydrophila challengedroups were utilized to collect kidney, liver and muscleissues in 10% neutral buffered formalin for histopathology.
.4. Haematological parameters
TEC and TLC were calculated manually using a haemo-ytometer after dilution of blood (200 and 50 times,espectively) in Hendrick’s solution, and values were
xpressed as number of cells mm−3 of blood. The haemat-crit was measured by microhaematocrit method (Sahoot al., 1998) and haemoglobin concentration (g dL−1)as estimated by cyanomethaemoglobin method usingunopathology 142 (2011) 207– 218 209
Drabkin’s reagent (Blaxhall and Daisley, 1973). The plasmaglucose content was quantified by enzymatic colorimetricmethod using GLUCOSE FL kit (Chema Diagnostica, Italy).
2.5. Non-specific immune parameters
2.5.1. Nitroblue tetrazolium assayThe reactive oxygen species superoxide (O2
–), hydrogenperoxides (H2O2) and hydroxyl radicals (OH−) producedby neutrophils during oxidative burst activity was mea-sured by reduction of a dye nitroblue tetrazolium (NBT), ina method described by Kumari et al. (2006). Briefly, 100 �Lof heparinised blood was mixed with equal proportion of0.2% NBT and incubated for 30 min at 25 ◦C. Furthermore,1 mL of dimethyl formamide was added to 50 �L of reactionmixture to solubilise the insoluble formazan granule. Theproduct was centrifuged at 2000 × g and reduced NBT wasmeasured at 540 nm using a UV–Vis spectrophotometer(Jenway, USA).
2.5.2. Myeloperoxidase activityThe total myeloperoxidase content present in serum
was measured according to Quade and Roth (1997) usinga partially modified technique (Sahoo et al., 2005). Briefly,in a 96-well microtitre plate, 10 �L of serum was dilutedwith 90 �L of HBSS without Ca2+ or Mg2+ and to that35 �L of 20 mM 3,3′,5,5′-tetramethyl benzidine hydrochlo-ride (TMB) (Genei, India) and 5 mM H2O2 were added. Thereaction was stopped by adding 35 �L of 4 M sulphuric acidafter 2 min of incubation. The optical density was read at450 nm in a microtitre plate reader (Anthos 2010, Austria).
2.5.3. Alternative complement haemolytic activity(ACH50)
The alternative complement haemolytic activity(ACH50) assay was carried out using Veronal bufferedsaline and rabbit RBC as target according to Matsuyamaet al. (1988). The value y/(1 − y) and the reciprocal of theserum dilution were plotted on log–log graph paper andthe ACH50 (units mL−1), the reciprocal dilution giving 50%haemolysis [y/(1 − y) = 1], was read from the graph.
2.5.4. Serum antiprotease assayThe anti-trypsin activity of fish sera was determined
according to Zuo and Woo (1997). Briefly, 10 �L of testserum was mixed with 100 �L of trypsin (bovine pancreastype I, Sigma) and incubated at 25 ◦C for 30 min along with110 �L PBS as control. It was further incubated with 1 mLof casein dissolved in PBS (2.5 mg/mL) for 15 min at 25 ◦C.The reaction was terminated with the addition of 500 �L of10% trichloroacetic acid (TCA). The sample was centrifugedat 10,000 × g for 5 min to remove protein precipitates. TheOD of the supernatant was measured at 280 nm and thepercentage trypsin inhibition was calculated comparing itto the control and reference samples.
2.5.5. Ceruloplasmin activity
Ceruloplasmin in serum sample was measured spec-trophotometrically as p-phenylene diamine (PPD) oxidaseactivity (Sigma) according to Pelgrom et al. (1995). Briefly,50 �L of serum or standard ceruloplasmin was incubated
and Im
210 A. Das et al. / Veterinary Immunologywith acetate buffer (1.2 M, pH 5.0) containing 0.1% PPD. Fur-thermore, each sample was mixed with 1 mL of 0.5% NaN3(sodium azide) and incubated at 37 ◦C for 30 min. Again,the reaction was stopped by adding 1 mL of NaN3. One unitof ceruloplasmin was defined as the amount of oxidasethat catalysed a decrease in absorbance of 0.001 min−1 at550 nm.
2.5.6. Total protein levelThe total protein concentration in control as well as
infected fish serum was determined by Bradford (1976)method.
2.6. Isolation of total RNA and cDNA synthesis
Total RNA was isolated from kidney and liver tissues(∼50 mg) using TRI reagent (Sigma) according to manufac-turer’s instruction. To minimise the chances of genomicDNA contamination, the RNA was treated with DNAse I(Fermentas, USA) and subsequently inactivated prior toreverse-transcription following instruction from the man-ufacturer. The purity and quantity of RNA was checkedspectrophotometrically using NanoDrop ND-1000 (Nano-Drop Technologies Inc., USA). RT-reactions lacking reversetranscriptase (RT minus) but not RNA were also performedto verify that the samples did not contain genomic DNA.
One microgram of total RNA was reverse-transcribedfor preparation of cDNA, which was kept at 0 ◦C until fur-ther use. RNA used for reverse transcription was incubatedwith RHP for 5 min at 70 ◦C and cooled at 25 ◦C for 10 minto anneal the primer properly to the RNA. To the mixture,
MMLV-RT-buffer, dNTP, RNAse inhibitor and MMLV-RTenzyme were added and incubated for 5 min at 25 ◦C, fol-lowed by 42 ◦C for 1 h and 95 ◦C for 2 min in a thermal cycler(Mastercycler Gradient Eppendorf, USA).Table 1Details of the primers used in the study.
Target genes Sequences Productsize (bp)
�-Actin F GAC TTC GAG CAG GAG ATG G 138
�-Actin R CAA GAA GGA TGG CTG GAA CAComplement 3F CCC TGG ACA GCA TTA TCA CTC 154
Complement 3R GAT GGT CGC CTG TGT GGTLysozyme GF CTT ATG CAG GTT GAC AAA CG 230
Lysozyme GR GGC AAC AAC ATC ACT GGA GTA ATCLysozyme CF CGC TGT GAT GTT GTC CGT ATC TTC 321
Lysozyme CR GTA ACT TCC CCA GGT ATC C�2-Microglobulin F CTC CAG TCC CAA GAT TCA GGT GTA 206
�2-Microglobulin R GAA GGA GAC GCT CTT GGT GAG GTransferrin1 F CCA CCG AGC GAG AGG CAG GAC TA 627
Transferrin1 R CCC TTA CGC ACC ACG GCCACA ACInterleukin 1�F ATC TTG GAG AAT GTG ATC GAA GAG 561
Interleukin 1�R GAT ACG TTT TTG ATC CTC AAG TGT GAA GCXCa F GGG TGT AGA TCC ACG CTG TC 102
CXCa R CTT TAC AGT GTG GGC TTG GAGInterleukin 8F GGG TGT AGA TCC ACG CTG TC 167
Interleukin 8R AGG GTG CAG TAG GGT CCA GMnSODF ACG AGA CCT GTA GTG CCC TGC 178
MnSODR CGG AAG CCA TCA AGC GTGNKEF-BF ACC GAG ATC ATC GCG TTC 285
NKEF-BR CCG CAG CGT AGC CGT AGT A
s.d.: self designed, n.s.: not submitted.
munopathology 142 (2011) 207– 218
2.7. Gene expression analysis
In order to adopt a semi-quantitative approach, theamplified �-actin gene (a house-keeping gene to checkthe integrity of RNA) was normalised with each targetgene. Cycle numbers and other PCR conditions for eachgene was optimised and employed for further analysis.The PCR reaction used to amplify certain innate immunerelated genes such as complement factor 3 (C3), lysozymeG-type (Lyso-G), lysozyme C (Lyso-C), �2-microglobulin(B2M), transferrin, CXCa, interleukin 1� (IL1�), interleukin8 (IL-8), natural killer cell enhancing factor (NKEF-B)and manganese superoxide dismutase (MnSOD) was per-formed using either self-designed or previously publishedheterologous species primer pairs. Each PCR reaction con-sisted of 40.70 �L nuclease-free water, 5 �L 10× PCR buffer,1 �L 10 mM dNTPs, 1 �L (10 pmole) of each forward andreverse primers, followed by addition of 0.3 �L (1.5 U) TaqDNA polymerase and 1 �L cDNA. The amplification profileof gene-specific primers was consists of an initial denatura-tion of 94 ◦C for 3 min, followed by denaturation of 94 ◦C for45 s for 28–32 cycles, at an appropriate annealing temper-ature for different genes for 45 s and extension at 72 ◦C for1 min 30 s, followed by a final extension for 10 min at 72 ◦C.The details of primers used are given in Table 1. The ampli-fied PCR products for each gene were run in 1.0% agarosegel prepared with 1× Tris–boric acid–EDTA buffer for 1 hat a constant voltage of 100 V.
To assess the level of expression of the target genes,densitometric analysis was performed using AlphaEase FCImaging Software (Alpha Innotech Corp., USA). Ratios of
immune-related genes/�-actin product for each gene ofinterest were calculated and used to measure the differ-ences in expression levels between control and infectedfish.Annealingtemperature(◦C)
Accessionnumbers
References
55. 3 n.s. s.d.
60 AM773826 Huttenhuis et al. (2006)
54 JF795589 s.d.
52.7 JF795588 s.d.
59.7 AM690445 s.d.
60 AM690342 Stafford and Belosevic (2003)
57.4 n.s. Engelsma et al. (2003)
56 FN429019 Mastellos and Lambris (2002)
53.5 HM569266 s.d.
57.4 HM569267 s.d.
49 JF795590 Chen et al. (2009)

and Imm
2
pariabstitfi
2
nfaTaGb
2
fotd
rmgt
3
Pahb
THb
A. Das et al. / Veterinary Immunology
.8. Cloning and sequencing of the genes
The PCR product of each gene was purified using Geneiurification kit and cloned into pGEMT vector (Promega)ccording manufacturer’s instruction. Plasmid DNA wasecovered by alkaline lysis method and confirmation ofnsertion of positive clone was done restriction digestionnalysis. Afterwards, three positive clones were purifiedy phenol–chloroform extraction method and used forequencing. The sequences are deposited in GenBank andhe accession numbers are given in Table 1. The sequenc-ng was done to confirm the gene-specific transcripts as allhe gene information for this species was generated for therst time in this study.
.9. Histopathology
Formalin-fixed tissues were washed overnight in run-ing water and then dehydrated in grades of alcohol,
urther embedded in paraffin wax and sectioned at 5 �m in rotary microtome (Weswox Optik, Ambala Cant, India).he sections were stained with haematoxylin and eosinnd observed under Axio Scope A1 microscope (Carl Zeiss,ermany) to study the pathological changes in tissues afteracterial infection.
.10. Statistical analysis
For haematological and immunological parameters, dif-erences between the mean values were assessed usingne-way ANOVA followed by Duncan’s multiple rangeests, with values P < 0.05 being accepted as significantlyifferent. All data were represented as mean ± SE.
The differences in expression pattern of the target geneselative to �-actin in control and infected fish was deter-ined using Mann–Whitney U-test. Mean ± SE of each
ene were calculated from three samples taken for eachime period.
. Results
There was no mortality in negative control tanks (in
BS-injected group) whereas 44% mortality was recordedfter 14 dpc in the positive control tanks in response to A.ydrophila injection. The cause of mortality was confirmedy reisolating the bacteria from kidneys of 10% dead fish.able 2aemato-immunological changes in Puntius sarana after Aeromonas hydrophila chearing different superscript(s) row-wise are significantly different (P < 0.05).
Parameter 0 day 2 days
TEC (×106 cells mm−3 of blood) 1.70 ± 0.10b 1.39 ± 0.10a
TLC (×103 cells mm−3 of blood) 19.27 ± 1.17b 10.45 ± 0.47a
Glucose (mg dL−1 of blood) 60.3 ± 4.37c 27.98 ± 1.12a
Haemoglobin (g dL−1 of blood) 6.55 ± 0.27b 4.49 ± 0.15a
Packed cell volume (%) 30.6 ± 0.88d 25.7 ± 0.47c
Myeloperoxidase (OD at 450 nm) 0.87 ± 0.13a 1.74 ± 0.12c
Nitroblue tetrazolium assay (OD at 540 nm) 0.25 ± 0.01a 0.3 ± 0.02a
Ceruloplasmin (units/25 �L of serum) 0.22 ± 0.01a 0.91 ± 0.14b
Anti-protease (% inhibition) 80.26 ± 0.64ab 77.76 ± 2.29a
ACH50 (units mL−1) 28.19 ± 1.43b 21.38 ± 0.91a
Total protein (g dL−1) 3.56 ± 0.41 3.45 ± 0.62
unopathology 142 (2011) 207– 218 211
The TEC was remarkably lower after bacterial exposureon all the time periods of measurement, whereas a widefluctuation in the TLC was noticed in infected fish. The TLCwas decreased initially on 2 dpc as compared to 0 dpc andsubsequently, it was quite higher in level till 14 dpc. How-ever, the plasma glucose level declined significantly afterinfection and restored to initial level (0 dpc) after 10 dpc.A significant decline in values of the haemoglobin and PCVwas also noticed in A. hydrophila-infected fish at all timeperiods after challenge and these values returned backnearer to pre-infection level on 14 dpc (Table 2).
The myeloperoxidase activity of serum in survivors wassignificantly higher in A. hydrophila-infected fish comparedto the control group fish initially up to 4 dpc and alsoon 10 dpc. Similarly, the superoxide production level ininfected fish increased significantly at 4 and 7 dpc, whereasthe ceruloplasmin activity remained higher at all time peri-ods post-infection. A significant increase in antiproteaseactivity was noticed only after 10 dpc and remained highat 14 dpc also. After a significant decline in ACH50 activ-ity initially at 2 dpc, a significant increase was marked at10 dpc, and it regained to control fish level at 14 dpc. Therewas no variation in total serum protein level at any timepoint of post-infection (Table 2).
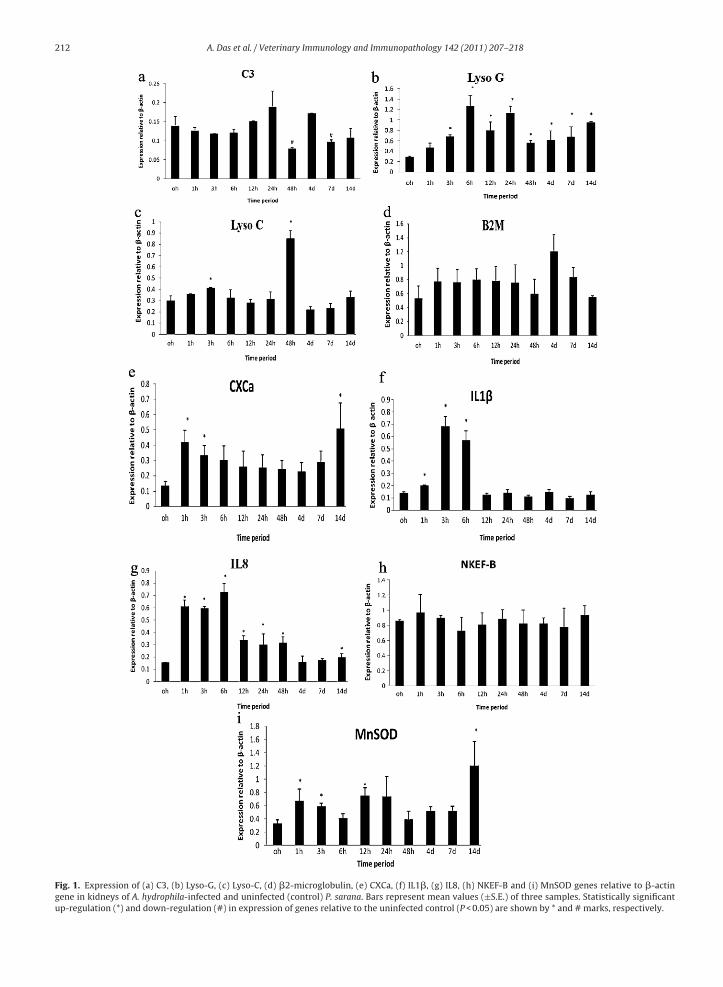
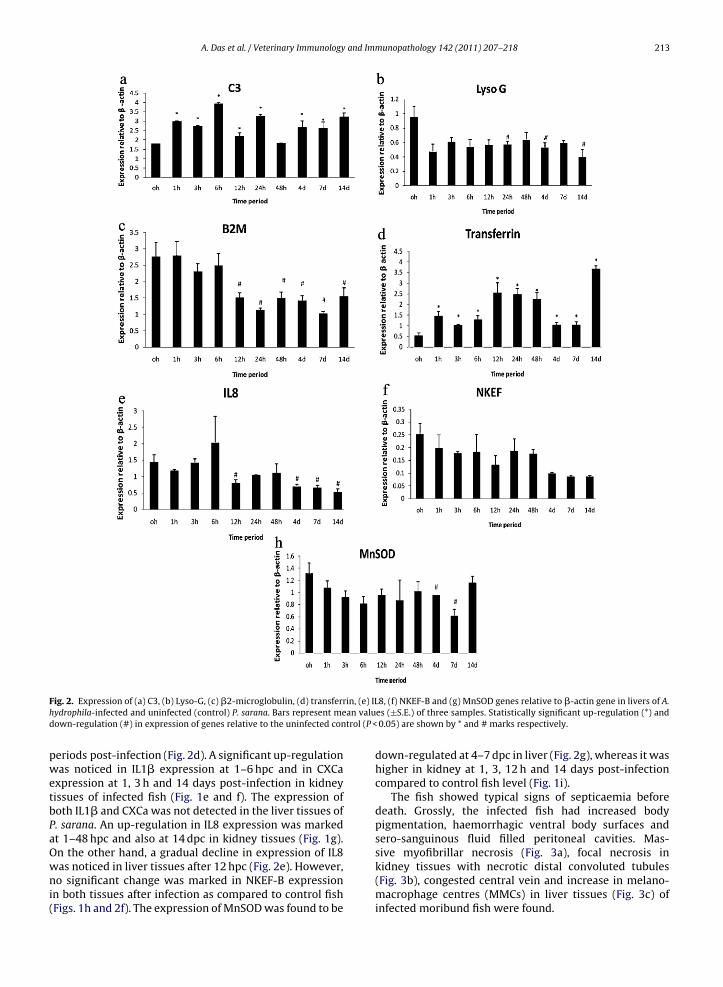
Densitometric analysis showed significant differences(P < 0.05) in expression of C3, Lyso-G, Lyso-C, B2M, trans-ferrin, CXCa, IL1�, IL8, NKEF-B and MnSOD genes in liverand kidney tissues of P. sarana after infection as comparedto control fish level. The expression level of C3 transcriptsin liver tissues was up-regulated significantly at all timeperiods (except at 48 hpc) (Fig. 2a), whereas a significantdown-regulation of the same was evident at 48 hpc and7 dpc in kidney tissues of infected fish (Fig. 1a). Higherexpression levels of lysozyme G was marked from 3 hpc to14 dpc in kidney tissues of infected fish (Fig. 1b). However,a down-regulation in its expression at 24 hpc, and 4 and14 dpc in liver tissues of infected fish were found (Fig. 2b).The expression of lysozyme C was found to be up-regulatedat 3 and 48 h post-challenge in kidney tissues (Fig. 1c),however, its expression was below detection limit inliver tissues either in control or in infected fish. The B2Mexpression showed no significant change in the kidney
(Fig. 1d) whereas it was found to be lower after 12 hpconwards in liver (Fig. 2c). The expression of transferrinwas noticed only in liver tissues and the expressionwas significantly higher in infected fish at all the timeallenge at different time periods. Data are presented as mean ± S.E. Means
4 days 7 days 10 days 14 days
b 1.36 ± 0.18ab 1.18 ± 0.12a 1.15 ± 0.11a 1.39 ± 0.24ab
54.41 ± 0.47e 24.65 ± 0.57c 22.44 ± 3.01bc 31.81 ± 1.87d
30.13 ± 4.33ab 43.24 ± 1.98b 59.39 ± 5.57c 71.99 ± 7.64c
3.81 ± 0.35a 4.13 ± 0.11a 3.81 ± 0.38a 5.63 ± 0.61b
22 ± 0.49b 18.8 ± 1.09a 16.7 ± 1.36a 23.9 ± 1.51bc
1.76 ± 0.17c 1.00 ± 0.08ab 1.27 ± 0.10b 1.05 ± 0.07ab
0.46 ± 0.02b 0.41 ± 0.03b 0.3 ± 0.05a 0.28 ± 0.03a
1.07 ± 0.11b 0.95 ± 0.15b 0.71 ± 0.06b 1.46 ± 0.17c
79.98 ± 1.09ab 81.94 ± 1.62ab 83.70 ± 1.40bc 87.44 ± 0.72c
24.63 ± 1.42ab 30.00 ± 2.72b 36.61 ± 3.07c 29.42 ± 2.11b
3.70 ± 0.71 2.92 ± 0.75 4.48 ± 0.89 4.13 ± 0.44
212 A. Das et al. / Veterinary Immunology and Immunopathology 142 (2011) 207– 218
Fig. 1. Expression of (a) C3, (b) Lyso-G, (c) Lyso-C, (d) �2-microglobulin, (e) CXCa, (f) IL1�, (g) IL8, (h) NKEF-B and (i) MnSOD genes relative to �-actingene in kidneys of A. hydrophila-infected and uninfected (control) P. sarana. Bars represent mean values (±S.E.) of three samples. Statistically significantup-regulation (*) and down-regulation (#) in expression of genes relative to the uninfected control (P < 0.05) are shown by * and # marks, respectively.
A. Das et al. / Veterinary Immunology and Immunopathology 142 (2011) 207– 218 213
F in, (e) ILh an valud trol (P <
pwetbPaOwni(
ig. 2. Expression of (a) C3, (b) Lyso-G, (c) �2-microglobulin, (d) transferrydrophila-infected and uninfected (control) P. sarana. Bars represent meown-regulation (#) in expression of genes relative to the uninfected con
eriods post-infection (Fig. 2d). A significant up-regulationas noticed in IL1� expression at 1–6 hpc and in CXCa
xpression at 1, 3 h and 14 days post-infection in kidneyissues of infected fish (Fig. 1e and f). The expression ofoth IL1� and CXCa was not detected in the liver tissues of. sarana. An up-regulation in IL8 expression was markedt 1–48 hpc and also at 14 dpc in kidney tissues (Fig. 1g).n the other hand, a gradual decline in expression of IL8
as noticed in liver tissues after 12 hpc (Fig. 2e). However,o significant change was marked in NKEF-B expressionn both tissues after infection as compared to control fishFigs. 1h and 2f). The expression of MnSOD was found to be
8, (f) NKEF-B and (g) MnSOD genes relative to �-actin gene in livers of A.es (±S.E.) of three samples. Statistically significant up-regulation (*) and
0.05) are shown by * and # marks respectively.
down-regulated at 4–7 dpc in liver (Fig. 2g), whereas it washigher in kidney at 1, 3, 12 h and 14 days post-infectioncompared to control fish level (Fig. 1i).
The fish showed typical signs of septicaemia beforedeath. Grossly, the infected fish had increased bodypigmentation, haemorrhagic ventral body surfaces andsero-sanguinous fluid filled peritoneal cavities. Mas-sive myofibrillar necrosis (Fig. 3a), focal necrosis in
kidney tissues with necrotic distal convoluted tubules(Fig. 3b), congested central vein and increase in melano-macrophage centres (MMCs) in liver tissues (Fig. 3c) ofinfected moribund fish were found.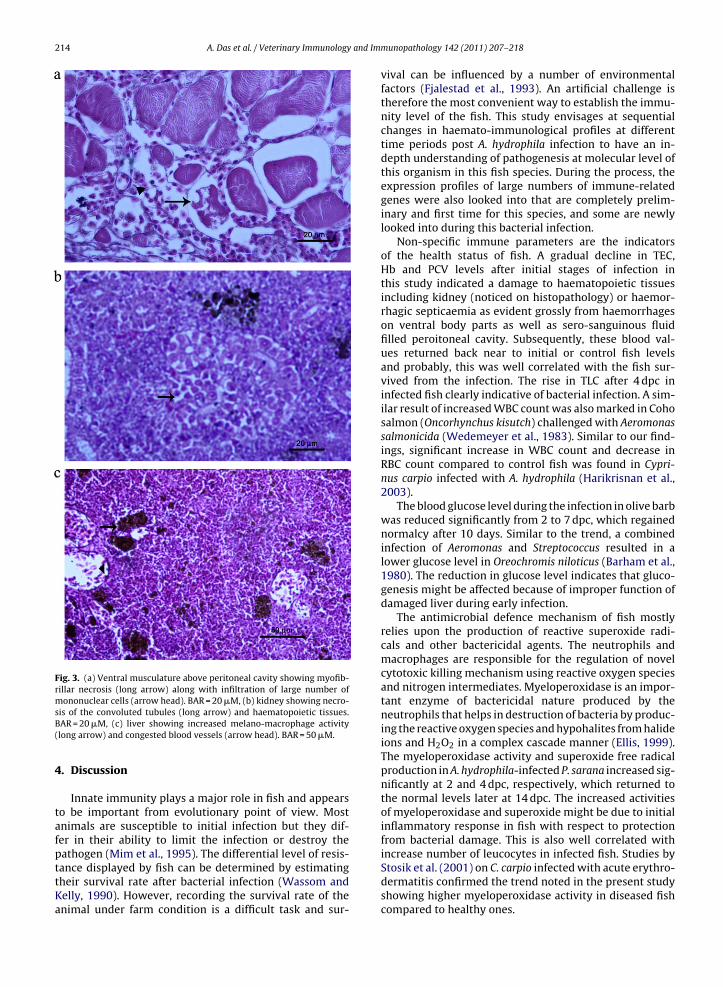
214 A. Das et al. / Veterinary Immunology and Im
Fig. 3. (a) Ventral musculature above peritoneal cavity showing myofib-rillar necrosis (long arrow) along with infiltration of large number ofmononuclear cells (arrow head). BAR = 20 �M, (b) kidney showing necro-
sis of the convoluted tubules (long arrow) and haematopoietic tissues.BAR = 20 �M, (c) liver showing increased melano-macrophage activity(long arrow) and congested blood vessels (arrow head). BAR = 50 �M.4. Discussion
Innate immunity plays a major role in fish and appearsto be important from evolutionary point of view. Mostanimals are susceptible to initial infection but they dif-fer in their ability to limit the infection or destroy thepathogen (Mim et al., 1995). The differential level of resis-
tance displayed by fish can be determined by estimatingtheir survival rate after bacterial infection (Wassom andKelly, 1990). However, recording the survival rate of theanimal under farm condition is a difficult task and sur-munopathology 142 (2011) 207– 218
vival can be influenced by a number of environmentalfactors (Fjalestad et al., 1993). An artificial challenge istherefore the most convenient way to establish the immu-nity level of the fish. This study envisages at sequentialchanges in haemato-immunological profiles at differenttime periods post A. hydrophila infection to have an in-depth understanding of pathogenesis at molecular level ofthis organism in this fish species. During the process, theexpression profiles of large numbers of immune-relatedgenes were also looked into that are completely prelim-inary and first time for this species, and some are newlylooked into during this bacterial infection.
Non-specific immune parameters are the indicatorsof the health status of fish. A gradual decline in TEC,Hb and PCV levels after initial stages of infection inthis study indicated a damage to haematopoietic tissuesincluding kidney (noticed on histopathology) or haemor-rhagic septicaemia as evident grossly from haemorrhageson ventral body parts as well as sero-sanguinous fluidfilled peroitoneal cavity. Subsequently, these blood val-ues returned back near to initial or control fish levelsand probably, this was well correlated with the fish sur-vived from the infection. The rise in TLC after 4 dpc ininfected fish clearly indicative of bacterial infection. A sim-ilar result of increased WBC count was also marked in Cohosalmon (Oncorhynchus kisutch) challenged with Aeromonassalmonicida (Wedemeyer et al., 1983). Similar to our find-ings, significant increase in WBC count and decrease inRBC count compared to control fish was found in Cypri-nus carpio infected with A. hydrophila (Harikrisnan et al.,2003).
The blood glucose level during the infection in olive barbwas reduced significantly from 2 to 7 dpc, which regainednormalcy after 10 days. Similar to the trend, a combinedinfection of Aeromonas and Streptococcus resulted in alower glucose level in Oreochromis niloticus (Barham et al.,1980). The reduction in glucose level indicates that gluco-genesis might be affected because of improper function ofdamaged liver during early infection.
The antimicrobial defence mechanism of fish mostlyrelies upon the production of reactive superoxide radi-cals and other bactericidal agents. The neutrophils andmacrophages are responsible for the regulation of novelcytotoxic killing mechanism using reactive oxygen speciesand nitrogen intermediates. Myeloperoxidase is an impor-tant enzyme of bactericidal nature produced by theneutrophils that helps in destruction of bacteria by produc-ing the reactive oxygen species and hypohalites from halideions and H2O2 in a complex cascade manner (Ellis, 1999).The myeloperoxidase activity and superoxide free radicalproduction in A. hydrophila-infected P. sarana increased sig-nificantly at 2 and 4 dpc, respectively, which returned tothe normal levels later at 14 dpc. The increased activitiesof myeloperoxidase and superoxide might be due to initialinflammatory response in fish with respect to protectionfrom bacterial damage. This is also well correlated withincrease number of leucocytes in infected fish. Studies by
Stosik et al. (2001) on C. carpio infected with acute erythro-dermatitis confirmed the trend noted in the present studyshowing higher myeloperoxidase activity in diseased fishcompared to healthy ones.
and Imm
atodooicowiStrleatwemaanhIpobgsh
bbNcptCmvpintartaqtifiirnfidw
A. Das et al. / Veterinary Immunology
Ceruloplasmin is a copper containing glycoprotein withn inflammatory role that converts reduced ferrous iono oxidised ferric form and facilitates the mobilisationf iron from cells to blood. It also acts like superoxideismutase to protect the body from the effect of super-xide radicals (Leenakunakorn et al., 2005). The releasef acute phase proteins like ceruloplasmin increases dur-ng stress and infection (Yada et al., 2004). In this study,eruloplasmin level increased significantly in all time peri-ds post-infection in the survivors, whereas similar resultas found in channel catfish Ictalurus punctatus following
nfection with Edwardsiella ictaluri (Peatman et al., 2007).ahoo et al. (2008) observed a significant positive correla-ion in families of rohu (Labeo rohita) showing increasedesistance to A. hydrophila infection and ceruloplasminevel. Pathogens secrete several proteases to obtain nutri-nts by degrading host’s proteins as a source of aminocids for their growth and replication. Fish plasma con-ains a number of protease inhibitors or antiproteases,hich inhibit the extracellular enzymes and restrict the
ntry of pathogens (Rao and Chakrabarti, 2004), there byaintaining a homeostasis of body. The activity of these
ntiproteases is seemed to be unaffected during infectionnd immunization (Magnadottir, 2006). Similarly, no sig-ificant difference was observed after the injection of A.ydrophila in P. sarana except for a slight increase at 14 dpc.t is understood that proteolysis cannot be performed if therotease is unable to cleave the bond in the ‘bait’ regionf antiprotease (Freedman, 1991). Investigating the reasonehind the consistency of antiprotease in infected fish sug-ested that the antiprotease secreted by the fish might beufficient to combat the protease activity or the proteaseas little effect on the activation of antiprotease.
The role of complement in phagocytosis, respiratoryurst, chemotaxis and cell lysis during pathogenesis haseen reviewed by various authors (Boshra et al., 2006;ikoskelainen et al., 2006). Furthermore, out of theomplement mediated pathways, alternative complementathway activated by the pathogens are considered ashe most important component of innate immune system.omplement factor 3 is the central and multifunctionalolecules of the complement cascade involved in the acti-
ation of three major immune response pathways. In theresent experiment, a significant fluctuation was noted
n the ACH50 activity as well as C3 expression in kid-ey and liver tissues. Expression of C3 showed oppositerends in liver and kidney with significant up-regulation (atlmost all time periods post-challenge) in liver and down-egulation (at 2 and 7 dpc) in kidney. This partly explainshe abrupt changes in ACH50 activity, which dropped oncet 2 dpc, returning to normal values thereafter and subse-uently rising significantly above normal at 10 dpc. Fromhe above results, it can be predicted that, at 48 hpc, thenfected tissues obtained from three randomly selectedsh might have been damaged severely, and hence it lost
ts capacity to synthesize the enzyme and afterwards itsegeneration might have started to bring its level near to
ormalcy. This result provides an additional support to thendings that C3 plays an important role in the immuneefence in response to bacterial challenge. Similar findingas observed in amphioxus Branchiostoma belcheri whenunopathology 142 (2011) 207– 218 215
challenged with Vibrio anguillarum, Escherichia coli andStaphylococcus aureus (Wang et al., 2009).
For decades, lysozymes have been considered as animportant molecule of innate defence system which acts ashydrolases in breaking the bacterial cell wall, thus causingits lysis. Besides antimicrobial action, it promotes phagocy-tosis by directly activating polymorphonuclear leucocytesand macrophages or indirectly by opsonic effect. Out offive major types of lysozymes, only C and G-types arereported in fish (Savan et al., 2003; Saurabh and Sahoo,2008). Antibacterial property of both the types of lysozymeagainst various bacteria viz., Listonella (Vibrio) anguil-larum, Vibrio alginolyticus, V. vulnificus and A. hydrophilawas well-documented (Larsen et al., 2009). In our exper-iment, we found early (3 hpc) up-regulation of both Lyso-Gand C-types expressions in the kidney. However, in liveronly expression of Lyso-G was noticed, which was down-regulated at 1 and 4 dpc. This acute phase reactants couldbe modulated as a response to degree and time of damageto the organs. A clear time course study on histopatholog-ical picture of these organs could probably be of help toexplain the major modulations in immune related genes.
�2-Microglobulin (B2M) is a cell surface protein non-covalently associated with MHC-I molecule and plays arole in binding, processing and presentation of the endoge-nous antigens (Choi et al., 2006). In the present study, nosignificant alteration in expression of the gene was foundin infected fish in comparison to control in kidney. Basalexpression of B2M is high even in uninfected P. sarana,and therefore it is likely that up-regulations during infec-tion are not necessarily crucial for effective antibacterialresponse against A. hydrophila. In an earlier study from thislaboratory, Mohanty and Sahoo (2008) corroborated thatno change was marked in B2M transcripts in A. hydrophilainfected Cirrhinus mrigala. However, in liver tissue, sig-nificant down-regulation in B2M transcripts was noticedfrom 12 h to 14 days post-challenge in P. sarana. Similarly,a down-regulation in B2M expression, though in kidney,was noted in L. rohita after Edwardsiella tarda exposure(Mohanty and Sahoo, 2010).
Transferrin is a major acute phase protein involved iniron transport and antimicrobial activity. Mostly its expres-sion is confined to liver, however, in some fishes expressionwas also found in brain (Sahoo et al., 2009). In case of P.sarana, although its expression was not detected in kidney;the expression was found to be up-regulated significantlyat almost all time periods post-infection. Similarly, it wasup-regulated up to 7-fold in channel catfish (I. punctatus)after infection with E. ictaluri (Peatman et al., 2007) andin the rainbow trout infected and re-infected with Yersiniaruckeri after 8 hpc and again 3 dpc (primary infection) andafter 7 dpc (secondary infection) (Raida and Buchmann,2009).
Cytokines are a group of signalling molecules that playa significant role in initiation and regulation of inflamma-tory processes. Chemokines and interleukins (IL) belongto a superfamily of small secreted proteins found to have
an ability to attract the leucocytes to the site of injury(Liu et al., 2007a,b). IL8 is a subfamily of CXC chemokine,which help in induction of chemotaxis, stimulating theneutrophils to release lysosomal enzyme to carry out
and Im
216 A. Das et al. / Veterinary Immunologyrespiratory burst activity, phagocytosis and angiogene-sis (Holland et al., 2003; Strieter et al., 1995). IL1� is aproinflammatory cytokine that acts as an inflammatorymediator during the microbial invasion, activates the lym-phocytes and induces other cytokines to release (Tassakkaaand Sakai, 2004). P. sarana challenged with A. hydrophilashowed up-regulation of IL1� and IL8 gene transcriptsat initial few hours of infection thus clearly indicative ofinflammatory response in fish to this bacterium. The sub-sequent decline in these transcripts may be well correlatedwith reduction in inflammatory activity in survived fish.The rise in IL8 transcripts in initial phase also might bestimulating the expression and release of lysozymes asobserved in this study. Zebrafish injected with A. hydrophilaalso showed induction in IL1� expression at 1 hpc and itbegan to decline at 2 hpc, returning to control levels at6 hpc whereas in heat-inactivated bacteria-injected fish itpeaked at 2 hpc with a 195-fold increase over the controllevels and returned to control levels at 6 hpc (Rodrıguezet al., 2008). However, in Atlantic cod the highest expres-sion of IL1� was found at 24 and 48 h following LPSstimulation and IPNV injection (Seppola et al., 2008). On theother hand, IL8 was found to be down-regulated at 12 hpcin liver in this study and the expression of CXCa and IL1�could not be detected in liver. Lack of detectable expressionof IL1� in liver was similar to that in sea bass Dicentrarchuslabrax, where it was weakly detected in liver after treat-ment of LPS (Scapigliati et al., 2001). The down-regulationin IL8 transcripts in later part of infection in liver mightindicate that the fish were gradually getting rid of infectionwith negligible bacterial load or release of other cytokineswhich might be playing role in the healing process.
Natural killer cells enhancing factor-B (NKEF-B) has theability to enhance the cytotoxic activity of natural killercells during uncontrolled cell division or tumor condition(Sauri et al., 1995). In addition to that it also protect thecellular molecules from the effect of superoxide radicals,thus act as an anti-oxidant. The expression of NKEF-B hasalready been studied in various fishes viz., common carp,C. carpio, channel catfish, I. punctatus and ayu, Plecoglossusaltivelis. The expression of this gene in liver and kidney wasfound to be insensitive to the bacterial infection similarto the findings in carp and puffer fish in earlier instances(Huang et al., 2009; Dong et al., 2007; Li and Waldbieser,2006).
All oxygen-respiring animals possess an importantantioxidant enzyme known as superoxide dismutase toprotect the body from the toxic activities of reactive oxygenspecies and oxidative stress (Cheng et al., 2006). Althoughfree radicals implicate their beneficial effect on pathogenclearance, signal transduction, gene regulation and angio-genesis, over production of these radicals may causecytotoxic death of the cell due to oxidative stress (Fanget al., 2002). It catalyzes the formation of O2 from O2
–, thusminimising the lethal effect of reactive species. MnSOD isclosely associated with the innate immune response as evi-dent from studies using bacteria, endotoxins and viruses.
In the present study, MnSOD was up-regulated in theinfected kidney tissues as reported in earlier findings (Choet al., 2009). The host might be utilizing this mechanismto protect its tissues from further damage during inflam-munopathology 142 (2011) 207– 218
matory process. Microscopically, the damage noticed invarious organs of fish was clear indication of damage dueto release of bacterial toxins and septicaemia. The pres-ence of increased melanomacrophage centres (MMCs) inAeromonas-infected liver might be indicative of a protectiveresponse of liver tissue against free radical damage causedby bacteria.
It is interesting to find that up-regulation or down-regulation of immune related genes and few immunemolecules were noticed at different phases of infection inP. sarana with regards to A. hydrophila infection. The tissuesamples were collected from survived fish after challenge.Thus, up- and down-regulation of the above genes or theirproducts studied here might be playing important role inthis bacterial pathogenesis and survival. Our results addto the understanding of the Indian medium carp immuneresponses to A. hydrophila and provide a foundation forfurther genomic characterization of immune-related genesfrom this important culturable fish.
Acknowledgement
Thanks are due to Dr. A.E. Ekanath, Director of theCentral Institute of Freshwater Aquaculture, Bhubaneswar,India for providing necessary facilities during the study.
References
Barham, W.T., Smit, G.L., Schoonbee, H.J., 1980. The hematological assess-ment of bacterial infection in rainbow trout Salmo gairdneri. J. FishBiol. 17, 275–281.
Blaxhall, P.C., Daisley, K.W., 1973. Routine haematological methods for usewith fish blood. J. Fish Biol. 5, 771–781.
Boshra, H., Li, J., Sunyer, J.O., 2006. Recent advances on the complementsystem of teleost fish. Fish Shellfish Immunol. 20, 239–262.
Bradford, M., 1976. A rapid and sensitive method for the quantification ofmicrogram quantities of protein. Anal. Biochem. 72, 248–254.
Chen, J., Wu, H.Q., Niu, H., Shi, Y.H., Li, M.Y., 2009. Increased liver proteinand mRNA expression of natural killer cell-enhancing factor B (NKEF-B) in ayu (Plecoglossus altivelis) after Aeromonas hydrophila infection.Fish Shellfish Immunol. 6, 567–571.
Cheng, W., Tung, Y.H., Liu, C.H., Chen, J.C., 2006. Molecular cloningand characterisation of cytosolic manganese superoxide dismutase(cytMn–SOD) from the giant freshwater prawn Macrobrachium rosen-bergii. Fish Shellfish Immunol. 20, 438–449.
Cho, Y.S., Lee, S.Y., Bang, I.C., Kim, D.S., Nam, Y.K., 2009. Genomic orga-nization and mRNA expression of manganese superoxide dismutase(Mn–SOD) from Hemibarbus mylodon (Teleostei, Cypriniformes). FishShellfish Immunol. 27, 571–576.
Choi, W., Lee, E.U., Choi, T., 2006. Cloning and sequence analysis of the �2-microglobulin transcript from flounder, Paralichthys olivaceous. Mol.Immunol. 43, 1565–1572.
Christie, D., Wei, G., Fujiki, K., Dixon, B., 2007. Cloning and characterizationof a cDNA encoding walleye (Sander vitreum) beta-2 microglobulin.Fish Shellfish Immunol. 22, 727–733.
Dong, W.R., Xiang, L.X., Shao, J.Z., 2007. Cloning and characterisation oftwo natural killer enhancing factor genes (NKEF-A and NKEF-B) inpufferfish, Tetraodon nigroviridis. Fish Shellfish Immunol. 22, 1–15.
Ellis, A.E., 1999. Immunity to bacteria in fish. Fish Shellfish Immunol. 9,291–308.
Engelsma, M.Y., Stet, R.J., Saeij, J.P., Verburg-van Kemenade, B.M., 2003.Differential expression and haplotypic variation of two interleukin-1beta genes in the common carp (Cyprinus carpio L.). Cytokine 22,21–32.
Fang, Y.Z., Yang, S., Wu, G., 2002. Free radicals, antioxidants, and nutrition.Nutrition 18, 872–879.
Fjalestad, K.T., Gjedrem, T., Gjerde, B., 1993. Genetic improvement of dis-ease resistance in fish: an overview. Aquaculture 111, 65–74.
Freedman, S., 1991. The role of alpha 2-macroglobulin in furunculosis: acomparison of rainbow trout and brook trout. Comp. Biochem. Physiol.98B, 549–553.

and Imm
G
H
H
H
H
H
I
J
K
L
L
L
L
L
L
M
M
M
M
M
M
M
N
N
P
A. Das et al. / Veterinary Immunology
hosh, D., Roy, S.K., Bhattarjee, U., 2005. Puntius sarana: an endangeredspecies – a case study on its present status. Fishing Chim. 25, 39–44.
arikrishnan, R., Moon, Y.G., Kim, M.C., Kim, J.S., Heo, M.S., 2010. Phy-totherapy of Aeromonas hydrophila infected goldfish, Carassius auratus.J. World Aqua. Soc. 41, 391–401.
arikrisnan, R., Rani, M.N., Balasundaram, C., 2003. Hematological andbiochemical parameters in common carp, Cyprinus carpio, follow-ing herbal treatment for Aeromonas hydrophila infection. Aquaculture221, 41–50.
olland, J.W., Gould, C.R.W., Jones, C.S., Noble, L.R., Secombes, C.J.,2003. The expression of immune-regulatory genes in rainbow trout,Oncorhynchus mykiss, during a natural outbreak of proliferative kidneydisease (PKD). Parasitology 126 (7), 95–102.
uang, R., Gao, Y.L., Wang, Y.P., Hua, W., Guo, Q.L., 2009. Structure, orga-nization and expression of common carp (Cyprinus carpio L.) NKEF-Bgene. Fish Shellfish Immunol. 26, 220–229.
uttenhuis, H.B.T., Grou, C.P.O., Taverne-Thiele, A.J., Taverne, N., Rombout,J.H.W.M., 2006. Carp (Cyprinus carpio L.) innate immune factors arepresent before hatching. Fish Shellfish Immunol. 20, 586–596.
noue, Y., Endo, M., Harutaa, C., Taniuchi, T., Moritomo, T., Nakanishi, T.,2003. Molecular cloning and sequencing of the silver chimera (Chi-maera phantasma) interleukin-8 cDNA. Fish Shellfish Immunol. 15,269–274.
ena, J.K., Das, P.C., Kar, S., Singh, T.K., 2008. Olive barb, Puntius sarana(Hamilton) is a potential candidate species for introduction into thegrow-out carp polyculture system. Aquaculture 280, 154–157.
umari, J., Sahoo, P.K., Swain, T., Sahoo, S.K., Sahu, A.K., Mohanty, B.R.,2006. Seasonal variation in the innate immune parameters of the Asiancatfish Clarias batrachus. Aquaculture 252, 121–127.
arsen, A.N., Terese, S., Gunbjorg, S., Seppola, M., Jorgensen, T.O., 2009.Molecular characterisation of a goose-type lysozyme gene in Atlanticcod (Gadus morhua L.). Fish Shellfish Immunol. 26, 122–132.
eenakunakorn, W., Sriworthy, R., Soontaros, S., 2005. Ceruloplasmin oxi-dase activity as a biomarker of lead exposure. J. Occup. Health 47,57–60.
i, R.W., Waldbieser, G.C., 2006. Genomic organisation and expressionof the natural killer cell enhancing factor (NKEF-B) gene in channelcatfish, Ictalurus punctatus (Rafinesque). Fish Shellfish Immunol. 20,72–82.
iu, Y., Chen, S., Meng, L., Zhang, Y., 2007a. Cloning, characterization andexpression analysis of a novel CXC chemokine from turbot (Scophthal-mus maximus). Fish Shellfish Immunol. 23, 711–720.
iu, Y., Chen, S., Meng, L., Zhang, Y., 2007b. Cloning, characterization andexpression analysis of a CXCL10-like chemokine from turbot (Scoph-thalmus maximus). Aquaculture 272, 199–207.
ow, C., Wadsworth, S., Burrells, C., Secombes, C.J., 2003. Expression ofimmune genes in turbot (Scophthalmus maximus) fed a nucleotide-supplemented diet C. Aquaculture 221 (1–4), 23–40.
agnadottir, B., 2006. Innate immunity of fish (overview). Fish ShellfishImmunol. 20, 137–151.
astellos, D., Lambris, J.D., 2002. Complement: more than a ‘guard’ againstinvading pathogens? Trends Immunol. 23, 485–490.
atsuyama, H., Tanaka, K., Nakao, M., Yano, T., 1988. Characterization ofthe alternative complement pathway of carp. Dev. Comp. Immunol.12, 403–408.
im, C.A., Dimmock, N.J., Nash, A., Stephen, J., 1995. General principles.In: Mims’ Pathogenesis of Infectious Disease, 4th ed. Academic Press,London, pp. 1–8.
ishra, J., Sahoo, P.K., Mohanty, B.R., Das, A., 2009. Sequence information,ontogeny and tissue-specific expression of complement compo-nent C3 in Indian major carp, Labeo rohita. Indian J. Exp. Biol. 47,672–678.
ohanty, B.R., Sahoo, P.K., 2008. Sequence analysis and tissue-specificexpression of the �2-microglobulin gene in Aeromonas hydrophilainfected Cirrhinus mrigala. Indian J. Anim. Sci. 78, 1426–1430.
ohanty, B.R., Sahoo, P.K., 2010. Immune responses and expression pro-files of some immune-related genes in Indian major carp, Labeorohita to Edwardsiella tarda infection. Fish Shellfish Immunol. 28,613–621.
i, D., Song, L., Gao, Q., Wu, L., Yu, Y., Zhao, J., Limei, Q., Huan, Z., Fangfang,S., 2007. The cDNA cloning and mRNA expression of cytoplasmic Cu,Zn superoxide dismutase (SOD) gene in scallop Chlamys farreri. FishShellfish Immunol. 23, 1032–1042.
ikoskelainen, S., Kjellsen, O., Lilius, E.-M., Schroder, M.B., 2006. Respira-tory burst activity of Atlantic cod (Gadus morhua L.) blood phagocytesdiffers markedly from that of rainbow trout. Fish Shellfish Immunol.
21, 199–208.eatman, E., Baoprasertkul, P., Terhune, J., Xu, P., Nandi, S., Kucuktas, H., Li,P., Wang, S., Somridhivej, B., Dunham, R., Liu, Z., 2007. Expression anal-ysis of the acute phase response in channel catfish (Ictalurus punctatus)
unopathology 142 (2011) 207– 218 217
after infection with a Gram-negative bacterium. Dev. Comp. Immunol.31, 1183–1196.
Pelgrom, S.M.G.J., Lock, R.A.C., Balm, P.H.M., Bonga, S.E.W., 1995. Inte-grated physiological response of tilapia, Oreochromis mossambicus, tosublethal copper exposure. Aquat. Toxicol. 32, 303–320.
Poobalane, S., Thompson, K.D., Ardo, L., Verjan, N., Han, H.J., Jeney, G.,Hirono, I., Aoki, T., Adams, A., 2010. Production and efficacy of anAeromonas hydrophila recombinant S-layer protein vaccine for fish.Vaccine 28, 3540–3547.
Quade, M.J., Roth, J.A., 1997. A rapid, direct assay to measure degranulationof bovine neutrophil primary granules. Vet. Immunol. Immunopathol.58, 239–248.
Raida, M.K., Buchmann, K., 2009. Innate immune response in rainbowtrout (Oncorhynchus mykiss) against primary and secondary infectionswith Yersinia ruckeri O1. Dev. Comp. Immunol. 33, 35–45.
Rao, Y.V., Chakrabarti, R., 2004. Enhanced anti-protease in Labeo rohitafed with diet containing herbal ingredients. Indian J. Clin. Biochem.19, 132–134.
Reed, L.J., Muench, H., 1938. A simple method of estimating fifty percentend points. Am. J. Hyg. 27, 493–497.
Rodrıguez, I., Novoa, B., Figueras, A., 2008. Immune response of zebrafish(Danio rerio) against a newly isolated bacterial pathogen Aeromonashydrophila. Fish Shellfish Immunol. 25, 239–249.
Sahoo, P.K., Kumari, J., Mishra, B.K., 2005. Non-specific immune responsesin juveniles of Indian major carp. J. Appl. Ichthyol. 21, 151–155.
Sahoo, P.K., Mahapatra, K.D., Saha, J.N., Barat, A., Sahoo, M., Mohanty,B.R., Gjerde, B., Odegard, J., Rye, R., Salte, R., 2008. Family asso-ciation between immune parameters and resistance to Aeromonashydrophila infection in the Indian major carp, Labeo rohita. Fish Shell-fish Immunol. 25, 163–169.
Sahoo, P.K., Mohanty, B.R., Kumari, J., Barat, A., Sarangi, N., 2009. Cloning,nucleotide sequence of phylogenetic analysis, and tissue specificexpression of transferrin gene in Cirrhinus mrigala infected withAeromonas hydrophila. Comp. Immunol. Microbiol. Infect. Dis. 32,527–537.
Sahoo, P.K., Mukherjee, S.C., Sahoo, S.K., 1998. Aeromonas hydrophilaversus Edwardsiella tarda: a pathoanatomical study in Clarias batra-chus. J. Aqua. 6, 57–66.
Saurabh, S., Sahoo, P.K., 2008. Lysozyme: an important defence moleculeof fish innate immune system. Aqua. Res. 39, 223–239.
Sauri, H., Butterfield, L., Kim, A., Shau, H., 1995. Antioxidant function ofrecombinant human natural killer enhancing factor. Biochem. Bio-phys. Res. Commun. 208, 964–969.
Savan, R., Aman, A., Sakai, M., 2003. Molecular cloning of G type lysozymecDNA in common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 15,263–268.
Scapigliati, G., Buonocore, F., Bird, S., Zou, J., Pelegrin, P., Falasca, C.,Prugnoli, D., Secombes, C.J., 2001. Phylogeny of cytokines: molecu-lar cloning and expression analysis of sea bass Dicentrarchus labraxinterleukin-1�. Fish Shellfish Immunol. 11, 711–726.
Seppola, M., Larsen, A.N., Steiro, K., Robertsen, B., Jensen, I., 2008. Char-acterisation and expression analysis of the interleukin genes, IL-1b,IL-8 and IL-10 in Atlantic cod (Gadus morhua L.). Mol. Immunol. 45,887–897.
Siwicki, A.K., esŁ, Z., Fuller Jr., J.C., Nissen, S., Trapkowska, S., Gzabski,E., Kowalska, A., KazunŁ, K., Terech-Majewska, E., 2006. Influence of�-hydroxy-�-methylbutyrate on nonspecific humoral defense mech-anisms and protection against furunculosis in pikeperch (Sanderlucioperca). Aqua. Res. 37, 127–131.
Stafford, J.L., Belosevic, M., 2003. Transferrin and the innate immuneresponse of fish: identification of a novel mechanism of macrophageactivation. Dev. Comp. Immunol. 27, 539–554.
Stosik, M., Deptula, W., Travnicek, M., 2001. Resistance in carps (Cyprinuscarpio L.) affected by a natural bacterial infection. Vet. Med. Czech. 46,6–11.
Strieter, R.M., Polverini, P.J., Kunkel, S.L., Arenberg, D.A., Burdick,M.D., Kasper, J., 1995. The functional role of the ELR motifin CXC chemokines-mediated angiogenesis. J. Biol. Chem. 270,27348–27357.
Tassakkaa, A.C.M.A.R., Sakai, M., 2004. Expression of immune-relatedgenes in the common carp (Cyprinus carpio L.) after stimulation byCpG oligodeoxynucleotides. Aquaculture 242, 1–12.
Vivas, J., Riano, J., Carracedo, B., Razquin, B.E., Lopez-Fierro, P., Naharro,G., Villena, J.A., 2004. The auxotrophic aroA mutant of Aeromonashydrophila as a live attenuated vaccine against A. salmonicida infec-tions in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol.
16, 193–206.Wang, G., Zhang, S., Wang, Z., 2009. Responses of alternative complementexpression to challenge with different combinations of Vibrio anguil-larum, Escherichia coli and Staphylococcus aureus: evidence for specific

and Im
218 A. Das et al. / Veterinary Immunologyimmune priming in amphioxus Branchiostoma belcheri. Fish ShellfishImmunol. 26, 33–39.
Wassom, D.L., Kelly, E.A.B., 1990. The role of the major histocompatibility
complex in resistance to parasitic infections. Crit. Rev. Immunol. 10,31–52.Wedemeyer, G.A., Gould, R.W., Yasutake, W.T., 1983. Some potentials andlimits of the leucocrit test as a fish health assessment method. J. FishBiol. 23, 711–716.
munopathology 142 (2011) 207– 218
Yada, T., Muto, K., Azuma, T., Ikuta, K., 2004. Effects of prolactin and growthhormone on plasma levels of lysozyme and ceruloplasmin in rainbowtrout. Comp. Biochem. Physiol. 139, 57–63.
Zuo, X., Woo, P.T.K., 1997. Natural anti-proteases in rainbow trout,Oncorhynchus mykiss and brook charr, Salvelinus fontinalis and thein vitro neutralization of fish �2-macroglobulin by the metallopro-tease from the pathogenic haemoflagellate, Cryptobia salmositica.Parasitology 114, 375–381.