unilateral striatal dopamine depletion: time-dependent effects on cortical function and behavioural...
TRANSCRIPT

Unilateral striatal dopamine depletion: time-dependenteffects on cortical function and behavioural correlates
Heinz Steiner* and Stephen T. KitaiDepartment of Anatomy and Neurobiology, University of Tennessee, College of Medicine, Memphis, TN 38163, USA
Keywords: basal ganglia, c-fos, cortex, nigrostriatal, rat, striatum, zif 268
Abstract
Previously, we showed that unilateral blockade of D1 dopamine receptors in the striatum inhibits immediate-early geneexpression bilaterally throughout large parts of the cortex, including sensory-evoked expression in the barrel cortex. To further
investigate this dopamine regulation of cortical function, we examined the effects of dopamine depletion on cortical gene
regulation and behavioural correlates. Two days after unilateral infusion of 6-hydroxydopamine into the midbrain, rats displayed a(to some degree) bilateral reduction in cortical zif 268 expression that was more pronounced on the lesioned side. This decrease
was found across motor, somatosensory, insular and piriform, but not cingulate, cortex, similar to the effects of blockade of
striatal D1 receptors. Furthermore, whisker stimulation-evoked c-fos and zif 268 expression in the barrel cortex ipsilateral to the
lesion was also attenuated by acute dopamine depletion. These cortical de®cits were accompanied by a breakdown ofspontaneous behaviours in an open-®eld test. In contrast, 21 days after dopamine depletion, both basal and sensory-evoked
gene expression in the cortex were near-normal. This cortical recovery was paralleled by recovery in locomotion and in sensory-
guided behaviour (scanning) related to the hemisphere contralateral to the lesion, but not in scanning by the dopamine-depletedhemisphere. Our results suggest that striatal dopamine exerts a widespread facilitatory in¯uence on cortical function that is
necessary, but not suf®cient, for normal behaviour. Moreover, the mechanisms mediating this cortical facilitation appear to be
subject to substantial neuroplasticity after dopamine perturbation.
Introduction
Interactions between the basal ganglia and the cerebral cortex are
critical for the generation of goal-directed behaviour. Dopamine in
the striatum regulates information ¯ow in cortico-basal ganglia±
cortical circuits. Striatal dopamine has been implicated in functions
as diverse as movement initiation, response selection, habit learning,
cognition and reward, and is associated with various basal ganglia/
cortical disorders, including Parkinson's disease, schizophrenia and
drug addiction (Albin et al., 1989; Alexander et al., 1990; Carlsson &
Carlsson, 1990; DeLong, 1990; Graybiel, 1995; Robbins et al., 1998;
Schultz et al., 1998; Redgrave et al., 1999; Sarter & Bruno, 1999;
Hollerman et al., 2000). Current models of basal ganglia±cortical
circuits propose, for example, that dopamine hypofunction or
hyperfunction in the striatum results in underactive or overactive
thalamocortical inputs and, consequently, in hypokinetic or hyper-
kinetic disorders, respectively (Penney & Young, 1983; Albin et al.,
1989; Alexander et al., 1990; DeLong, 1990). Results of various
functional studies principally support these models. Thus, dopamine
agonist treatments that produce behavioural activation also result in
increased cortical activation, as indicated by functional markers such
as the expression of immediate-early genes in the cortex (Paul et al.,
1992; Dilts et al., 1993; Daunais & McGinty, 1994; Steiner & Gerfen,
1994; Wang & McGinty, 1995; LaHoste et al., 1996; Badiani et al.,
1998). However, in these studies, the observed changes in cortical
gene regulation were generally widespread and not limited to the
frontal/premotor areas emphasized as cortical targets of basal ganglia
output in the models (Alexander et al., 1986; Alexander & Crutcher,
1990; Alexander et al., 1990). Interpretation of these ®ndings is
dif®cult, however, because these studies used systemic drug
treatments, thus precluding conclusions regarding the site of drug
action.
Recently, we showed that local manipulation of basal ganglia
output by intrastriatal drug administration produced similar wide-
spread cortical effects. Thus, blocking striatal dopamine action by
intrastriatal infusion of an opioid agonist attenuated immediate-early
gene expression in the cortex (Steiner & Gerfen, 1995), indicating a
role for striatal dopamine receptors in such cortical gene regulation.
A more detailed investigation of the cortical regions affected and the
dopamine receptor subtypes involved demonstrated that unilateral
blockade of striatal D1-type receptors inhibited immediate-early gene
expression induced by the D1/D2 receptor agonist apomorphine
bilaterally throughout motor, somatosensory, insular and piriform
cortex (Steiner & Kitai, 2000b). Furthermore, blocking striatal D1
receptors also inhibited whisker stimulation-evoked immediate-early
gene expression in the barrel cortex (Steiner & Kitai, 2000b). These
®ndings indicate that stimulation of striatal D1 receptors facilitates
cortical function.
In the present study, we further investigated this dopamine
regulation of cortical function by assessing the effects of dopamine
depletion on gene expression in the cortex and behavioural correlates.
The transcription factors/immediate-early genes zif 268 and c-fos
Correspondence: Dr Heinz Steiner, at *present address belowE-mail: steinerh@®nchcms.edu
*Present address: Department of Cellular and Molecular Pharmacology, FinchUniversity of Health Sciences/The Chicago Medical School, 3333 Green BayRoad, North Chicago, IL 60064, USA
Received 12 March 2001, revised 20 August 2001, accepted 24 August 2001
European Journal of Neuroscience, Vol. 14, pp. 1390±1404, 2001 ã Federation of European Neuroscience Societies

were used as functional markers (e.g. Sharp et al., 1993; Chaudhuri,
1997; Herdegen & Leah, 1998). In the ®rst experiment, we mapped
time-dependent effects of a 6-hydroxydopamine (6-OHDA) lesion on
`basal' expression of zif 268 in various cortical regions. In the second
experiment, 6-OHDA effects on whisker stimulation-evoked c-fos
and zif 268 expression in the barrel cortex were examined. These
cortical gene regulation effects were compared with behavioural
changes after dopamine depletion, especially with an index of
whisker utilization (thigmotactic scanning). Our results show that
acute unilateral dopamine depletion attenuated basal and sensory-
evoked immediate-early gene expression in the cortex, similar to
unilateral blockade of striatal D1 receptors. However, these effects
were transient and as animals regained near-normal cortical gene
expression they recovered from some, but not all, behavioural de®cits
produced by the lesion. A preliminary account of these results has
been presented in abstract form (Steiner & Kitai, 2000a).
Materials and methods
Subjects
A total of 56 male Sprague-Dawley rats (170±210 g; Sasco, St. Louis,
MO, USA) were used in this study, 28 in Experiment 1 and 28 in
Experiment 2. In both experiments, the animals were assigned to one
of four groups (n = 5±8 each) that received either an infusion of
6-OHDA or vehicle into the midbrain, and were tested either 2 or
21 days after the infusion. The rats were housed in groups of two to
three under standard laboratory conditions. They had free access to
food and water, and were maintained under a 12-h light : 12-h dark
cycle (lights on at 07.00 h). The experiments were carried out
between 13.00 and 16.00 h. The procedures used in this study were
approved by the Animal Care and Use Committee of the University
of Tennessee, Memphis.
Dopamine depletion
The rats were anaesthetized with equithesin (3.5 mL/kg) and placed
in a David Kopf stereotaxic frame. The neurotoxin 6-OHDA
(6-OHDA HBr, Sigma, St. Louis, MO, USA; 16 mg in 2 mL of
0.9% NaCl/0.02% ascorbic acid), or vehicle, was infused with a pump
(rate 0.5 mL/min) into the area of the rostral border of the right
substantia nigra. The coordinates used for the tip of the infusion
cannula were (relative to the interaural line): A, +4.5; L, 1.5; V, +2.0
with the incisor bar set at ±2.5 mm (Paxinos & Watson, 1998). All
rats received an injection of desipramine (25 mg/kg, i.p.; Sigma) 30±
45 min before the infusion.
Behavioural test
Two days (`acute') or 21 days after the 6-OHDA or vehicle infusion,
behavioural effects were examined during a 10-min test session in a
novel open ®eld (60 3 60 3 40 cm, with lines dividing the ¯oor into
3 3 3 squares). The behaviour was video-taped and assessed from
the tapes by an experimenter who was unaware of the treatment. The
behavioural analysis started 30 s after the animal was placed into
the centre of the open ®eld. The following behavioural items
were measured: distance traveled (number of lines crossed with
all four feet) and turning towards the left or right side (number of
half turns with a diameter < 20 cm). As a measure of whisker
utilization, we assessed thigmotactic scanning (Steiner et al., 1986)
with the left or right side (measured as the number of lines crossed
while walking along the wall with the snout within 5 cm of the wall)
(see Fig. 2D).
Whisker stimulation
In Experiment 2, we tested whisker stimulation-evoked immediate-
early gene expression in the barrel cortex ipsilateral to the dopamine
depletion at 2 or 21 days after the 6-OHDA infusion. Thus, following
the open-®eld test, one mystacial whisker (C2 on the left side of the
snout) was stimulated using procedures described previously in detail
(Melzer & Steiner, 1997; Steiner & Kitai, 2000b). In short, 30 min
prior to the start of the stimulation a steel ®lament (length 6 mm,
diameter 0.3 mm) was glued to whisker C2 (centre of ®lament
» 8 mm from the skin) under light Metofane anaesthesia. The distal
end of whisker C2 was clipped. All other mystacial whiskers on both
sides of the face were clipped close to the skin. The rat was then
allowed to recover. Whisker C2 was stimulated by exposing the rat to
a pulsating magnetic ®eld (Van der Loos stimulator; Melzer et al.,
1985). The magnetic ®eld (mean strength 10.8 mT, RMS) was
produced by a copper coil powered by a solid-state relay (George
Dold, Research Services Branch, NIMH; based on the design by
Melzer et al., 1985). With the longitudinal axis of the magnetic ®eld
orientated horizontally, the ®eld pulses de¯ect whiskers mostly in
rostrocaudal directions. A pulse rate of » 8 Hz was chosen in order to
mimic the whisking frequency of a whisking (`snif®ng') rat. The
awake rat was placed into the acrylic stimulation chamber (cylinder,
15 cm in length, 14 cm in diameter) inside the coil and stimulated for
15 min.
Tissue preparation
Immediately following the open-®eld test, or 10 min after whisker
stimulation, the rat was killed with CO2. The brain was rapidly
removed, frozen in isopentane cooled on dry ice, and stored at ±20 °C
until cryostat sectioning. Coronal sections (12 mm) through striatum
and midbrain or, for the stimulated brains, tangential sections through
the somatosensory cortex followed by coronal sections through
striatum and midbrain, were collected. The sections were thaw-
mounted onto glass slides twice coated with gelatin, dried on a
hotplate and stored at ±20 °C. In preparation for in situ hybridization
histochemistry, the sections were, at room temperature, ®xed in 4%
paraformaldehyde/0.9% saline for 10 min, incubated in a fresh
solution of 0.25% acetic anhydride in 0.1 M triethanolamine/0.9%
saline (pH 8.0) for 10 min, dehydrated, defatted for 2 3 5 min in
chloroform, rehydrated and air-dried. The slides were then stored at
±20 °C until hybridization. Every sixth tangential section through the
somatosensory cortex was ®xed and stained for cytochrome oxidase
activity to locate barrel C2 in layer IV (Melzer & Steiner, 1997). In
addition, sets of striatal sections were processed for tyrosine
hydroxylase immunohistochemistry (Steiner et al., 1999) to assess
the loss of dopamine terminals.
In situ hybridization histochemistry
Oligonucleotide probes (48-mers; Life Technologies, Baltimore, MD,
USA) were labelled with [35S]-dATP (Steiner & Kitai, 2000b). The
probes had the following sequence: c-fos, complementary to bases
1227±1274, GenBank accession number X06769; zif 268, bases 352±
399, M18416; enkephalin, bases 436±483, M28263; tyrosine
hydroxylase, bases 1441±1488, M10244. Labelled probe (» 3 3
106 c.p.m.) in 100 mL of hybridization buffer was added to each
slide. The sections were coverslipped and incubated at 37 °C
overnight. After incubation, the slides were ®rst rinsed in four
washes of 13 saline citrate (150 mM sodium chloride, 15 mM sodium
citrate). Then they were washed three times 20 min each in 23 saline
citrate/50% formamide at 40 °C, followed by two washes 30 min
each in 13 saline citrate at room temperature. After a brief water
Dopamine depletion and cortical function 1391
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

rinse, the sections were air-dried and then apposed to X-ray ®lm
(BioMax MR-2, Kodak) for 3 days to 3 weeks.
Analysis of autoradiograms
Enkephalin expression was measured in coronal sections from a
rostral striatal level (`rostral', at » 10.5 mm rostral to the interaural
line; Paxinos & Watson, 1998) and a mid-striatal level (`middle',
9.5 mm), and also monitored at two more caudal striatal levels (8.2±
7.6 mm). Basal zif 268 expression was analysed in sections from the
rostral and middle striatal levels, as well as in coronal sections
collected at the level of the barrel ®eld (`caudal', 7.2±6.7 mm).
Stimulation-evoked c-fos and zif 268 expression was investigated in
tangential sections through the right somatosensory cortex that were
adjacent to sections containing barrel C2 (layer IV), as determined by
cytochrome oxidase histochemistry. Tyrosine hydroxylase immunor-
eactivity was assessed in rostral and middle striatal sections, and
tyrosine hydroxylase mRNA expression was determined at several
rostrocaudal levels through the substantia nigra (4.3±3.0 mm).
Gene expression was measured in the following areas (Paxinos &
Watson, 1998). Rostral and middle striatal levels: striatum, nucleus
accumbens, olfactory tubercle, total cortex, cingulate, `sensorimotor'
(including M2, M1, S1, S2, insular regions), piriform cortex. Caudal
level: total cortex, retrosplenial, motor, hindlimb/forelimb, `somato-
sensory' (rest S1, S2), insular, piriform cortex. In stimulated animals,
gene expression was measured in barrel C2 and in the surrounding
barrels (C1, C3, B2, D2).
Levels of gene expression were determined by densitometry on
®lm autoradiograms, using a Macintosh-based image analysis system
(NIH Image, Wayne Rasband, NIMH). The `mean density' values
presented are background-corrected: for coronal sections, mean
density over gray matter minus mean density over white matter; for
tangential sections, mean density in C2 minus averaged mean density
in surrounding barrels. The illustrations of ®lm autoradiograms
displayed in Figs 1, 3 and 6 are computer-generated images, and are
contrast-enhanced where necessary. Maximal hybridization signal is
black.
Statistics
Treatment effects on gene expression and behaviour were determined
with three-factor [main effects: lesion, time, hemisphere/side (with-
in)] or two-factor ANOVAs (main effects: lesion, time) using the
Statistica software package (Statistica 4.1 for Macintosh, StatSoft,
Tulsa, OK, USA). Differences between individual groups were
described with Newman±Keuls post hoc tests.
noninfused6-OHDA
noninfusedvehicle
striatum
2 days 21 daysV 6V 6
0
20
40
60
80
100
mea
n de
nsity
****
**aa
**
6-OHDA6-OHDA
+3.4
+4.0
+4.3
2 days 21 days
6-OHDA
2 days 21 days
S
S
csh
6-OHDA 6-OHDA 6-OHDA
A C
B
rost
ral
mid
dle
tyrosine hydroxylase enkephalin
D
SNc VTA
FIG. 1. Effects of unilateral 6-OHDA lesion on tyrosine hydroxylase and enkephalin expression 2 days (`acute') and 21 days after the lesion. (A) Examples ofcoronal sections from the rostral and middle striatum stained for tyrosine hydroxylase immunoreactivity are shown for animals killed 2 or 21 days after the6-OHDA infusion into the right midbrain. (B) Illustrations of ®lm autoradiograms depict tyrosine hydroxylase mRNA expression in sections from threerostrocaudal levels [4.3±3.4 mm rostral to the interaural line (Paxinos & Watson, 1998)] through the midbrain for such animals. (C) Examples of ®lmautoradiograms show enkephalin expression in sections from the rostral and middle striatum. The arrows point to the lateral striatal zone that displayed thestrongest increase in enkephalin expression 2 days after the lesion. (D) Mean density values (mean 6 SEM, arbitrary units) for enkephalin expressionmeasured across the total striatum at the middle level in the noninfused and infused hemispheres are given for rats that received an infusion of vehicle (V) or6-OHDA (6) into the right midbrain (n = 5±8). **P < 0.01 vs. noninfused or as indicated; aaP < 0.01 vs. acute. S, striatum; c, nucleus accumbens core; sh,nucleus accumbens shell; SNc, substantia nigra pars compacta; VTA, ventral tegmental area.
1392 H. Steiner and S. T. Kitai
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

Results
Dopamine depletion characterized by tyrosine hydroxylaseimmunoreactivity and mRNA, and enkephalin expression:2 days vs. 21 days
Twenty-one days after unilateral 6-OHDA infusion, tyrosine
hydroxylase immunoreactivity in the striatum (Fig. 1A) and mRNA
in the substantia nigra (Fig. 1B) were undetectable on the side of the
infusion in all animals. Enkephalin expression was increased
throughout the striatum ipsilateral to the infusion, as compared to
the contralateral striatum or to vehicle-infused controls (Fig. 1C;
Fig. 1D shows numerical data for the middle striatal level from
animals in Experiment 1). These results indicate a near-total
dopamine depletion at this time point (Li et al., 1990; Nisenbaum
et al., 1996). In contrast, 2 days after 6-OHDA administration, both
tyrosine hydroxylase immunoreactivity and mRNA were only partly
eliminated. In the substantia nigra, tyrosine hydroxylase mRNA was
generally mostly reduced in rostral and medial parts and less or little
in lateral and caudal parts (Fig. 1B). In the striatum, immuno-
reactivity was preferentially reduced in medial areas (Fig. 1A). At
this time point, changes in enkephalin mRNA levels poorly matched
those in tyrosine hydroxylase distribution. Thus, a small but
statistically signi®cant increase in enkephalin expression was seen
across the striatum (Fig. 1D), but this increase was most robust in
lateral parts (Fig. 1C). A dissociation between enkephalin expression
and tyrosine hydroxylase levels was also apparent in the nucleus
accumbens. Thus, minimal or no changes in enkephalin expression
were noted at any time (Fig. 1C; numerical data not shown), despite
an apparently near-total loss of tyrosine hydroxylase immunoreac-
tivity at 2 days (core) and 21 days (core and shell) (Fig. 1A), and of
tyrosine hydroxylase mRNA in the ventral tegmental area at 21 days
(Fig. 1B).
Open-®eld behaviour
Figure 2 depicts lesion effects on open-®eld behaviour in Experiment
1. Locomotor activity (distance traveled) did not differ between the
vehicle-infused groups tested 2 days or 21 days after the infusion
(Fig. 2A). Two days after unilateral 6-OHDA administration, the rats
showed very little forward locomotion, but displayed occasional
turning towards the side of the lesion. This was re¯ected by an almost
complete lack of line crossings (1% of vehicle controls) (Fig. 2A) and
a moderate number of ipsiversive half turns (Fig. 2B). The rats tested
21 days after the lesion still showed a reduced amount of locomotion
(42% of controls), but their crossing rate was statistically signi®cantly
increased compared to the 2-day group (Fig. 2A). The amount of
FIG. 2. Open-®eld behaviour 2 days (acute) and 21 days after the 6-OHDA infusion into the right midbrain. (A) The distance traveled (number of crossings,mean 6 SEM) during the 10-min test, given as the total (left) and for 2-min blocks (right), is shown for rats that received a vehicle (V) or a 6-OHDAinfusion (6) (n = 5±7). (B) Turning behaviour (total number of net half turns) is depicted for these animals. Positive values indicate turning ipsiversive to thelesion. (C) Scanning (number of crossings inside a 5-cm-wide zone along the wall, indicated by the broken line in D) is shown for the sides of the facefunctionally related to the noninfused and vehicle (V)- or 6-OHDA (6)-infused hemispheres. (D) Schematic illustration of the dominant behaviouralasymmetries seen in the open ®eld 21 days after unilateral dopamine depletion, turning ipsiversive to the lesion (left) and scanning with the whisker siderelated to the noninfused hemisphere (i.e. the side ipsilateral to the 6-OHDA lesion) (right). **P < 0.01 as indicated; aaP < 0.01, aP < 0.05, vs. acute.
Dopamine depletion and cortical function 1393
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

turning ipsiversive to the lesion was somewhat higher at 21 days than
at 2 days (Fig. 2B).
The animals that received a vehicle infusion into the midbrain did
not display signi®cant asymmetries in thigmotactic scanning, either
2 days or 21 days after the infusion (Fig. 2C). The behavioural
breakdown 2 days after the 6-OHDA infusion also resulted in an
almost complete elimination of scanning at this time point (Fig. 2C).
The animals tested 21 days after the 6-OHDA lesion showed a
scanning asymmetry. Thus, scanning with the side of the face
functionally related to the dopamine-depleted hemisphere (i.e.
scanning with the left side) was not or minimally increased compared
to 2-day animals. In contrast, scanning with the side associated with
the noninfused hemisphere (i.e. scanning with the right whiskers) had
almost completely recovered (Fig. 2C). Principally, the same time-
dependent effects of dopamine depletion on scanning behaviour were
found in Experiment 2 (Fig. 6C); the other behavioural effects in
these latter animals were also similar (not shown).
Reduced basal zif 268 expression in the cortex after acute,but not prolonged, unilateral dopamine depletion
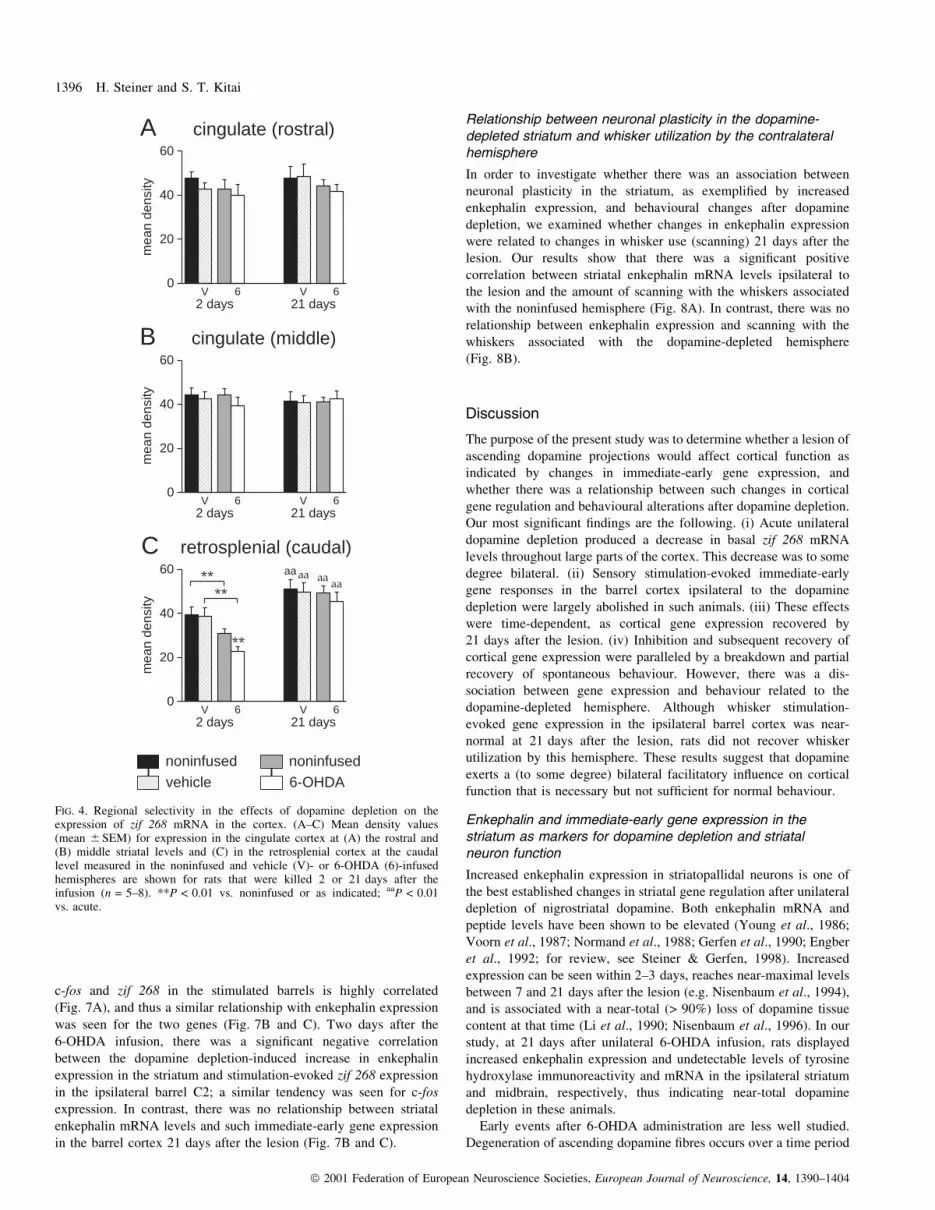
Effects of dopamine depletion on basal zif 268 expression are shown
in Figs 3±5 and Table 1. Acute dopamine depletion (2-day group)
zif 268 expression in total cortex
noninfused6-OHDA
noninfusedvehicle
mid
dle
caud
al
A
B
0
20
40
60
ME
AN
DE
NS
ITY **
aa
**
**
*
aaaa
aa
2 days 21 daysV 6V 6
0
20
40
60
ME
AN
DE
NS
ITY
** aa
**
* aa
** **
*
2 days 21 daysV 6V 6
2 days 21 daysV 6V 6
0
20
40
60
80
100
120
ME
AN
DE
NS
. (%
NIF
)
aa**
2 days 21 days66
0
20
40
60
80
100
120M
EA
N D
EN
S. (
% C
) aa
**cc
aa
c
2 days 21 daysV 6V 6
0
20
40
60
80
100
120
ME
AN
DE
NS
. (%
NIF
)
** aa
2 days 21 days66
0
20
40
60
80
100
120
ME
AN
DE
NS
. (%
C)
aa
**cc
c
zif 268 expression after "acute" dopamine depletion (2 days)rostral middle caudal
6-OHDA6-OHDA6-OHDA
mea
n de
nsity
(%
C)
mea
n de
nsity
(%
C)
mea
n de
nsity
(%
NIF
)m
ean
dens
ity (
% N
IF)
mea
n de
nsity
mea
n de
nsity
FIG. 3. Expression of zif 268 mRNA in the cortex, striatum and nucleus accumbens after unilateral dopamine depletion. (A) Illustrations of ®lmautoradiograms depicting zif 268 expression in coronal sections from rostral (left), middle (middle) and caudal (right) striatal levels are shown for rats killed2 days after the 6-OHDA infusion into the right midbrain. The arrows point to zones with increased zif 268 expression in the lateral striatum ipsilateral to thelesion. (B) Mean density values (mean 6 SEM) for zif 268 expression measured across the total cortex in middle and caudal sections are given for rats thatwere killed 2 or 21 days after the infusion (n = 5±8). The results are shown as absolute values for the noninfused and vehicle (V)- or 6-OHDA (6)-infusedhemispheres (left), and as relative values, either expressed as percentage of the noninfused side (% NIF) (middle) or as percentage of corresponding values invehicle-infused controls (% C) (right). **P < 0.01, *P < 0.05 vs. noninfused or as indicated; aaP < 0.01 vs. acute; ccP < 0.01, cP < 0.05 vs. vehicle-infusedcontrols.
1394 H. Steiner and S. T. Kitai
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

produced a statistically signi®cant decrease in basal zif 268 mRNA
levels in the ipsilateral cortex, as compared to the noninfused side and
to vehicle-infused controls, on all rostrocaudal levels examined
(Fig. 3A and B). Moreover, a statistically signi®cant reduction in zif
268 expression was also found in the cortex contralateral to the
6-OHDA infusion (noninfused side), although this effect was less
robust. In contrast, at 21 days after the lesion, zif 268 mRNA levels in
the dopamine-depleted and noninfused hemispheres were close to the
levels in vehicle-infused control animals (Fig. 3B). However, there
were also statistically signi®cant effects of the infusion per se (lower
values in vehicle-infused hemisphere compared to noninfused
hemisphere) and general time effects (lower levels at 2 days, e.g. in
noninfused hemisphere of 2-day vehicle-infused animals compared to
corresponding 21-day animals). Some of these effects may be related
to damage produced by lowering the infusion cannula to the ventral
midbrain (Dragunow & Robertson, 1988; Jacobs et al., 1994). To
eliminate the contribution of such effects, we also analysed relative
zif 268 mRNA levels (values expressed as percentage of those in the
noninfused hemisphere, or of those in the corresponding vehicle-
infused controls). This analysis con®rmed a signi®cant, to some
degree bilateral, effect of 6-OHDA on zif 268 expression in the cortex
at 2 days after the neurotoxin infusion, and a subsequent recovery of
this effect (Figs 3B and 5, Table 1).
Our regional analysis showed that this 6-OHDA-induced decrease
in zif 268 mRNA levels was, to a variable degree, present in all
cortical areas examined, with the exception of the cingulate cortex
(Table 1, Figs 4 and 5). The ipsilateral decrease was very robust
across retrosplenial, motor, somatosensory, insular and piriform
cortical areas. The more moderate contralateral reduction was most
robust in the retrosplenial and the somatosensory cortex (Table 1,
Fig. 4C). Notably, the distinct bilateral decrease in the retrosplenial
area was contrasted by a lack of an effect in the rostrally adjacent
cingulate cortex (Fig. 4). Thus, zif 268 mRNA levels in the cingulate
cortex at both rostrocaudal levels examined were unchanged 2 and
21 days after dopamine depletion (Figs 4 and 5A).
Effects on basal zif 268 expression in the striatum andnucleus accumbens
Effects of 6-OHDA on zif 268 expression in striatal areas on three
rostrocaudal levels are depicted in Fig. 3A. Figure 5 compares the
effects in the cortex (middle level) (Fig. 5A±C) with those in the
striatum (middle level) (Fig. 5D and E) and in the nucleus accumbens
(rostral level) (Fig. 5F). When measured across the total striatum, zif
268 mRNA levels were statistically signi®cantly decreased in both
hemispheres at 2 days after the 6-OHDA infusion (Fig. 5D).
However, on the lesioned side a more pronounced decrease medially
was contrasted by a statistically signi®cant increase in a restricted
zone in the most lateral striatum (Figs 3A and 5E). At 21 days after
the lesion, zif 268 mRNA levels in the lateral striatum were not
signi®cantly different from controls. Similar to the medial striatum, in
the nucleus accumbens, zif 268 expression was signi®cantly
decreased bilaterally, at both time points (Fig. 5F). Again, this effect
was somewhat more pronounced on the dopamine-depleted side,
especially at 21 days after the lesion (Fig. 5F). Similar effects were
seen for both core and shell of nucleus accumbens (data not shown).
Changes in zif 268 expression in the olfactory tubercle were similar to
those in the nucleus accumbens (data not shown).
Attenuation of whisker stimulation-evoked immediate-earlygene expression in the somatosensory cortex after acutedopamine depletion
In Experiment 2, we assessed sensory stimulation-evoked immediate-
early gene expression in the dopamine-depleted hemisphere follow-
ing the open-®eld test. These rats received 15-min stimulation of
whisker C2 on the side of the face contralateral to the 6-OHDA
lesion, and c-fos and zif 268 mRNA levels were measured in the
stimulated barrel C2. Acute dopamine depletion produced a pro-
nounced attenuation of whisker stimulation-evoked gene expression
in the barrel cortex ipsilateral to the lesion (Figs 6A and B, and 7).
Thus, for both immediate-early genes, stimulation-evoked mRNA
levels in barrel C2 were signi®cantly reduced compared to those in
the vehicle-infused animals, at 2 days after 6-OHDA infusion.
Especially, c-fos mRNA levels were hardly above background in
most of these animals (Fig. 7B). In contrast, 21 days after the lesion,
whisker stimulation-evoked c-fos and zif 268 expression in barrel C2
did not differ from that in the vehicle controls (Figs 6B and 7A).
Therefore, whisker stimulation-evoked immediate-early gene expres-
sion in the barrel cortex had recovered at a time when the same rats
failed to show signi®cantly improved utilization of these whiskers
(i.e. scanning with the side functionally related to the dopamine-
depleted hemisphere; Fig. 6C).
Using enkephalin expression as a marker, we examined whether
there was a relationship between dopamine depletion-induced
changes in gene expression in the striatum and changes in whisker
stimulation-evoked immediate-early gene expression in the barrel
cortex after the 6-OHDA lesion. Stimulation-evoked expression of
TABLE 1. Expression of zif 268 mRNA in the cortex after dopamine depletion
Cortical area
Expression of zif 268 mRNA
At 2 days At 21 days
Noninfused 6-OHDA Noninfused 6-OHDA
Retrosplenial 78.3 6 5.6³³ 58.5 6 5.3**³³ 96.6 6 6.5²² 91.8 6 8.0²²
Motor 82.2 6 3.7 49.8 6 7.1**³³ 94.6 6 4.6 94.7 6 6.2²²
Hindlimb/forelimb 90.2 6 8.0 47.2 6 4.9**³³ 100.4 6 7.1 90.6 6 7.3²²
Somatosensory 84.1 6 7.0³ 61.0 6 3.7**³³ 103.1 6 8.3² 99.4 6 8.5²²
Insular 84.8 6 2.9 58.3 6 4.9**³³ 91.0 6 7.8 104.0 6 7.9²²
Piriform 93.0 6 4.1 67.7 6 6.4**³³ 96.1 6 9.3 118.8 6 10.4²²
Total cortex 85.2 6 5.1³ 61.0 6 4.0**³³ 98.5 6 7.6²² 99.6 6 7.8²²
Mean density values (mean 6 SEM, in percentage of values in the vehicle-infused controls) measured in various cortical regions on the noninfused and 6-OHDA-infused sides in coronal sections collected at the level of the barrel ®eld (`caudal' level) are given for animals that were killed 2 days or 21 days after 6-OHDAinfusion into the right midbrain (n = 5±8). **P < 0.01 vs. noninfused side; ²²P < 0.01, ²P < 0.05, vs. acute (2 days); ³³P < 0.01, ³P < 0.05 vs. vehicle-infusedcontrols.
Dopamine depletion and cortical function 1395
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404
c-fos and zif 268 in the stimulated barrels is highly correlated
(Fig. 7A), and thus a similar relationship with enkephalin expression
was seen for the two genes (Fig. 7B and C). Two days after the
6-OHDA infusion, there was a signi®cant negative correlation
between the dopamine depletion-induced increase in enkephalin
expression in the striatum and stimulation-evoked zif 268 expression
in the ipsilateral barrel C2; a similar tendency was seen for c-fos
expression. In contrast, there was no relationship between striatal
enkephalin mRNA levels and such immediate-early gene expression
in the barrel cortex 21 days after the lesion (Fig. 7B and C).
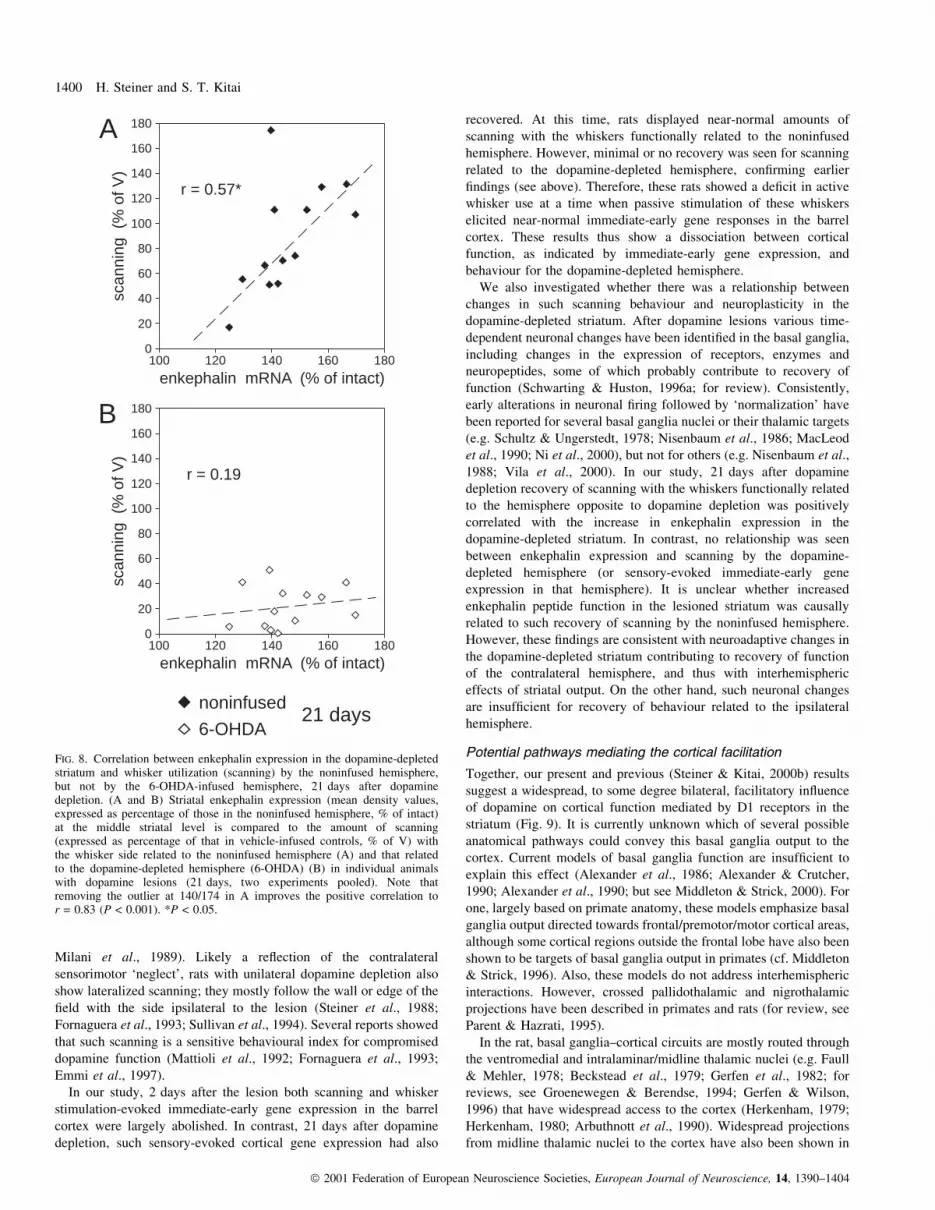
Relationship between neuronal plasticity in the dopamine-depleted striatum and whisker utilization by the contralateralhemisphere
In order to investigate whether there was an association between
neuronal plasticity in the striatum, as exempli®ed by increased
enkephalin expression, and behavioural changes after dopamine
depletion, we examined whether changes in enkephalin expression
were related to changes in whisker use (scanning) 21 days after the
lesion. Our results show that there was a signi®cant positive
correlation between striatal enkephalin mRNA levels ipsilateral to
the lesion and the amount of scanning with the whiskers associated
with the noninfused hemisphere (Fig. 8A). In contrast, there was no
relationship between enkephalin expression and scanning with the
whiskers associated with the dopamine-depleted hemisphere
(Fig. 8B).
Discussion
The purpose of the present study was to determine whether a lesion of
ascending dopamine projections would affect cortical function as
indicated by changes in immediate-early gene expression, and
whether there was a relationship between such changes in cortical
gene regulation and behavioural alterations after dopamine depletion.
Our most signi®cant ®ndings are the following. (i) Acute unilateral
dopamine depletion produced a decrease in basal zif 268 mRNA
levels throughout large parts of the cortex. This decrease was to some
degree bilateral. (ii) Sensory stimulation-evoked immediate-early
gene responses in the barrel cortex ipsilateral to the dopamine
depletion were largely abolished in such animals. (iii) These effects
were time-dependent, as cortical gene expression recovered by
21 days after the lesion. (iv) Inhibition and subsequent recovery of
cortical gene expression were paralleled by a breakdown and partial
recovery of spontaneous behaviour. However, there was a dis-
sociation between gene expression and behaviour related to the
dopamine-depleted hemisphere. Although whisker stimulation-
evoked gene expression in the ipsilateral barrel cortex was near-
normal at 21 days after the lesion, rats did not recover whisker
utilization by this hemisphere. These results suggest that dopamine
exerts a (to some degree) bilateral facilitatory in¯uence on cortical
function that is necessary but not suf®cient for normal behaviour.
Enkephalin and immediate-early gene expression in thestriatum as markers for dopamine depletion and striatalneuron function
Increased enkephalin expression in striatopallidal neurons is one of
the best established changes in striatal gene regulation after unilateral
depletion of nigrostriatal dopamine. Both enkephalin mRNA and
peptide levels have been shown to be elevated (Young et al., 1986;
Voorn et al., 1987; Normand et al., 1988; Gerfen et al., 1990; Engber
et al., 1992; for review, see Steiner & Gerfen, 1998). Increased
expression can be seen within 2±3 days, reaches near-maximal levels
between 7 and 21 days after the lesion (e.g. Nisenbaum et al., 1994),
and is associated with a near-total (> 90%) loss of dopamine tissue
content at that time (Li et al., 1990; Nisenbaum et al., 1996). In our
study, at 21 days after unilateral 6-OHDA infusion, rats displayed
increased enkephalin expression and undetectable levels of tyrosine
hydroxylase immunoreactivity and mRNA in the ipsilateral striatum
and midbrain, respectively, thus indicating near-total dopamine
depletion in these animals.
Early events after 6-OHDA administration are less well studied.
Degeneration of ascending dopamine ®bres occurs over a time period
noninfusedvehicle
noninfused6-OHDA
B cingulate (middle)
0
20
40
60
ME
AN
DE
NS
ITY
2 days 21 daysV 6V 6
A cingulate (rostral)
0
20
40
60M
EA
N D
EN
SIT
Y
2 days 21 daysV 6V 6
C retrosplenial (caudal)
0
20
40
60
ME
AN
DE
NS
ITY
**
aa
**
aa**aaaa
2 days 21 daysV 6V 6
mea
n de
nsity
mea
n de
nsity
mea
n de
nsity
FIG. 4. Regional selectivity in the effects of dopamine depletion on theexpression of zif 268 mRNA in the cortex. (A±C) Mean density values(mean 6 SEM) for expression in the cingulate cortex at (A) the rostral and(B) middle striatal levels and (C) in the retrosplenial cortex at the caudallevel measured in the noninfused and vehicle (V)- or 6-OHDA (6)-infusedhemispheres are shown for rats that were killed 2 or 21 days after theinfusion (n = 5±8). **P < 0.01 vs. noninfused or as indicated; aaP < 0.01vs. acute.
1396 H. Steiner and S. T. Kitai
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

of several hours to several days after the neurotoxin infusion during
which some tyrosine hydroxylase immunoreactivity is still present
(e.g. Zahm, 1991; for review, see Schwarting & Huston, 1996a). In
contrast, impulse ¯ow-dependent dopamine release in the striatum
appears to cease within 20±40 min of 6-OHDA infusion
(Svenningsson et al., 1999). This early loss of dopamine release is
followed closely by increased expression of c-fos and zif 268 in the
striatum (Cole & Di Figlia, 1994; Brog & Zahm, 1995; Svenningsson
et al., 1999; Schuller & Marshall, 2000), which occurs predominantly
in enkephalin/D2 receptor-expressing (striatopallidal) neurons
(Svenningsson et al., 1999; Schuller & Marshall, 2000). As shown
here and before (Svenningsson et al., 1999; Schuller & Marshall,
2000), this immediate-early gene response is most robust in the lateral
striatum, similar to gene induction after acute D2 receptor blockade
(e.g. Steiner & Gerfen, 1999). Such increased immediate-early gene
expression is thought to re¯ect increased activity in these neurons due
to a loss of stimulation of inhibitory D2 receptors. Current models of
basal ganglia function predict that increased activity in striatopallidal
neurons together with reduced activity in striatonigral neurons after
dopamine depletion produce diminished thalamocortical activation
and bradykinesia (e.g. Albin et al., 1989; DeLong, 1990).
Consistently, in our study, basal and sensory-evoked immediate-
early gene expression in the cortex were attenuated at this early time
point.
The early strong immediate-early gene response in the striatum
subsequently subsides to some degree (Schuller & Marshall, 2000),
and 3 weeks after a 6-OHDA lesion, net zif 268 mRNA levels in the
striatum re¯ect a minor increase in expression in striatopallidal
neurons in conjunction with a more pronounced decrease in
expression in striatonigral neurons (Gerfen et al., 1995). Together,
these dopamine release and gene regulation studies indicate that
striatal neuron function is altered within a short time period after
6-OHDA infusion and that these initial effects are subsequently
modi®ed (see also below).
Acute dopamine depletion results in attenuated immediate-early gene expression in the cortex
Earlier studies demonstrated that acute treatments with various
dopamine agonists produce increased expression of immediate-early
genes throughout most of the cortex (Paul et al., 1992; Dilts et al.,
1993; Daunais & McGinty, 1994; Steiner & Gerfen, 1994; Wang &
McGinty, 1995; LaHoste et al., 1996; Badiani et al., 1998; Berke
et al., 1998; Steiner & Kitai, 2000b). Consistently, our present
®ndings show that acute dopamine depletion results in decreased zif
268 mRNA levels in the cortex. The unilateral 6-OHDA lesion
produced robust decreases throughout large parts of the ipsilateral
cortex that reached 40±50% in some areas. In addition, smaller
reductions (10±20%) were seen in some contralateral cortical areas.
Although widespread, this lesion effect was regionally selective, as
attenuated zif 268 expression was found across retrosplenial, motor,
somatosensory, insular and piriform cortex, but not in the cingulate
cortex. Furthermore, sensory-evoked responses (i.e. whisker stimu-
A cingulate cortex B sensorimotor cortex C piriform cortex
D total striatum E lateral striatum F nucl. accumbens
0
20
40
60
80
100
120
ME
AN
DE
NS
. (%
C)
aa
*
cc
2 days 21 days66
0
20
40
60
80
100
120
ME
AN
DE
NS
. (%
C)
aa
**
cc
2 days 21 days66
0
20
40
60
80
100
120
ME
AN
DE
NS
. (%
C)
2 days 21 days66
0
20
40
60
80
100
120
140
ME
AN
DE
NS
. (%
C)
cccc cc
2 days 21 days66
noninfused6-OHDA
0
20
40
60
80
100
120
140M
EA
N D
EN
S. (
% C
)
**
c
2 days 21 days66
0
20
40
60
80
100
120
140
ME
AN
DE
NS
. (%
C)
cc
cccc
*
cc
2 days 21 days66
mea
n de
nsity
(% C
)m
ean
dens
ity (%
C)
mea
n de
nsity
(% C
)m
ean
dens
ity (%
C)
mea
n de
nsity
(% C
)m
ean
dens
ity (%
C)
FIG. 5. Differential effects of dopamine depletion on zif 268 expression in cortical and striatal regions. (A±F) Mean density values (mean 6 SEM) expressedas percentage of corresponding values in vehicle-infused controls (% C) for (A) cingulate, (B) sensorimotor and (C) piriform cortex, and for (D) the total and(E) lateral striatum, all at the middle striatal level, and (F) the nucleus accumbens at the rostral level measured in the noninfused and 6-OHDA (6)-infusedhemispheres are given for rats that were killed 2 or 21 days after the 6-OHDA infusion (n = 5±8). **P < 0.01, *P < 0.05 vs. noninfused; aaP < 0.01 vs.acute; ccP < 0.01, cP < 0.05 vs. vehicle-infused controls.
Dopamine depletion and cortical function 1397
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

lation-evoked c-fos and zif 268 expression) in the ipsilateral barrel
cortex were also greatly reduced in such animals.
Our ®ndings of to some degree bilateral effects of unilateral
dopamine depletion on cortical gene expression are consistent with
results of previous studies. Unilateral 6-OHDA lesions have been
found to produce changes in the expression of neuropeptides,
glutamic acid decarboxylase and glutamate receptors in fronto-
parietal cortical areas that were partly bilateral (Lindefors et al.,
1989; Lindefors et al., 1990; Rodriguez-Puertas et al., 1999). For
example, metabotropic glutamate receptor subtype 3 mRNA levels
were increased ipsilaterally, whereas subtype 4 mRNA levels were
increased bilaterally 4 weeks after a unilateral 6-OHDA lesion
(Rodriguez-Puertas et al., 1999). Moreover, D1 receptor agonist
treatments after unilateral dopamine depletion produced widespread
induction of immediate-early genes in the cortex of both hemispheres
(Steiner & Gerfen, 1996; Berke et al., 1998; Rodriguez-Puertas et al.,
1999).
Regional and laminar patterns of changes in cortical gene
expression after dopamine agonist (see above) or antagonist
(Steiner & Kitai, 2000b) treatments, as well as after dopamine
depletion (see above; present results) poorly match dopamine
innervation (Bjorklund & Lindvall, 1984; Berger et al., 1991) or
dopamine receptor localization (Mansour et al., 1990; Bouthenet
et al., 1991; Mengod et al., 1991; Gaspar et al., 1995; Defagot et al.,
1997; Ciliax et al., 2000) in the rat cortex, suggesting that such gene
regulation is the effect of altered cortical inputs rather than local
dopamine action (LaHoste et al., 1996). Our previous results showed
that acute blockade of striatal D1 receptors by intrastriatal infusion of
a D1 receptor antagonist attenuates basal and apomorphine-induced
immediate-early gene expression in the cortex, indicating a
facilitatory role for striatal D1 receptors in cortical gene regulation
(Steiner & Kitai, 2000b). The present results are consistent with such
a role for striatal D1 receptors. Notably, the present lesion effects
shared several characteristics with those of blockade of striatal D1
receptors (Steiner & Kitai, 2000b). For example, in both studies,
`basal' and sensory-evoked gene expression were inhibited; all
cortical areas examined were affected with the exception of the
cingulate cortex (see also LaHoste et al., 1996, for similar effects of
dopamine agonists); the effects were to various degrees bilateral, but
were more robust ipsilaterally. We thus speculate that, in the present
study, the attenuated cortical immediate-early gene expression after
acute dopamine depletion re¯ects a loss of dopamine action in the
striatum and ensuing functional alterations in basal ganglia±cortical
circuits. The correlation between increased striatal enkephalin
expression (marker for dopamine depletion, see above) and reduced
whisker stimulation-evoked gene expression in the barrel cortex
observed 2 days after 6-OHDA infusion is consistent with this notion.
However, despite these similarities between the effects of striatal
D1 receptor blockade and those of the 6-OHDA infusion, we can not
exclude the possibility that other effects of the 6-OHDA lesion
contributed to the observed neuronal and behavioural changes. For
example, the neurotoxin infusion may also have produced nonspeci®c
injury, the effects of which dissipated over time. Moreover, although
the vast majority of mesotelencephalic dopamine projections termin-
ates in the striatum, several other forebrain structures also receive
dopamine inputs from the midbrain (Fallon & Moore, 1978; Fallon
et al., 1978; Bjorklund & Lindvall, 1984). A loss of such extrastriatal
dopamine terminals may thus have contributed to the lesion effects.
Regarding the effects on gene expression in the contralateral cortex, it
can be argued that they could be due to lesion of the small crossed
nigrostriatal projection (Fass & Butcher, 1981; Fallon et al., 1983;
Pritzel et al., 1983), or to diffusion of the neurotoxin across the
midline and resulting injury to dopamine neurons in the contralateral
hemisphere. Although we did not observe evidence for a contralateral
lesion by inspecting tyrosine hydroxylase immunoreactivity in the
c-fosB zif 268
0
20
40
60M
EA
N D
EN
SIT
Y aa
**
2 days 21 daysV 6V 6
0
10
20
30
40
50
ME
AN
DE
NS
ITY
aa
**
2 days 21 daysV 6V 6
noninfusedvehicle
noninfused6-OHDA
2 days 21 daysV 6V 6
0
20
40
60
SC
AN
NIN
G
**
**
*
aa
*
scanningC
A
C1
C3
B2
D2
CO activityzif 268c-fos
"acute" dopamine depletion (2 days)
vehi
cle
6-O
HD
Acr
ossi
ngs
mea
n de
nsity
mea
n de
nsity
FIG. 6. Whisker stimulation-evoked expression of c-fos and zif 268 mRNAsin the barrel cortex after dopamine depletion. (A) Illustrations of ®lmautoradiograms show c-fos (left) and zif 268 expression (middle) in barrel C2in tangential sections through layer IV of the barrel cortex ipsilateral to thevehicle or 6-OHDA infusion in animals that had the contralateral whisker C2stimulated 2 days after the infusion (acute). Barrel C2 was located bycytochrome oxidase (CO) activity in adjacent sections (right); surroundingbarrels C1, C3, B2 and D2 are outlined. (B) Mean density values (mean6 SEM) for stimulation-evoked c-fos and zif 268 expression in barrel C2 areshown for animals that received an infusion of vehicle (V) or 6-OHDA (6)into the ipsilateral midbrain and were stimulated 2 or 21 days after theinfusion and killed thereafter (n = 5±8). Values presented are the differencebetween the C2 value and averaged values measured in the surroundingbarrels. (C) Scanning (mean 6 SEM) during a 10-min open-®eld test prior towhisker stimulation is shown for the sides of the face functionally related tothe noninfused and vehicle (V)- or 6-OHDA (6)-infused hemispheres in theseanimals. Note the dissociation between stimulation-evoked gene expressionin the barrel cortex ipsilateral to the dopamine depletion (recovered) andactive utilization (scanning) of the whiskers functionally related to thedopamine-depleted hemisphere (not recovered) 21 days after the lesion.**P < 0.01, *P < 0.05 as indicated; aaP < 0.01 vs. acute.
1398 H. Steiner and S. T. Kitai
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

striatum and mRNA in the midbrain, these markers may not be
sensitive enough for more subtle effects. However, zif 268 expression
in the lateral striatum (the most sensitive marker we assessed, see
above) tended to be decreased in the contralateral striatum, and not
increased as seen after dopamine depletion. Future studies will have
to ascertain the role of striatal dopamine.
Parallel changes in cortical immediate-early gene expressionand some behaviours after dopamine depletion
The decrease in cortical gene expression after dopamine depletion
was transient. Both basal zif 268 expression and sensory-evoked c-fos
and zif 268 expression were back to control levels 21 days after the
lesion. Parallel functional recovery was seen for some, but not all,
behaviours when rats were tested in a novel open ®eld. Numerous
studies have shown that unilateral dopamine depletion produces early
bradykinesia/akinesia followed by some recovery, and persisting
behavioural asymmetries including ipsiversive turning and a loss of
responsiveness to sensory stimulation on the contralateral body
surface (sensorimotor `neglect') (Ungerstedt, 1971; Marshall et al.,
1974; Ljungberg & Ungerstedt, 1976; Carli et al., 1985; Brown &
Robbins, 1989; for review, see Schwarting & Huston, 1996b). The
degree of early bradykinesia is directly related to the extent of
dopamine depletion in the striatum (Fornaguera et al., 1994). In the
present study, rats tested 2 days after 6-OHDA infusion showed very
little forward locomotion and scanning, but some turning. In contrast,
rats tested 21 days after the lesion displayed signi®cantly more
locomotion, but retained turning and scanning asymmetries. Thus, the
recovery in cortical immediate-early gene expression was paralleled
by a partial recovery from akinesia.
An important question raised by these parallel changes in
immediate-early gene expression and behaviour is to what degree
the observed effects on gene expression were in¯uenced by changes
in behavioural activity (sensorimotor feedback). Thus, did the acute
decrease and subsequent recovery in cortical zif 268 mRNA levels
merely re¯ect acute akinesia and subsequent behavioural recovery in
the open-®eld test? An in¯uence of behavioural activities on some
cortical gene expression is likely. For example, sensory input
produced by whisking seems to affect gene expression in the
somatosensory cortex (Steiner & Gerfen, 1994). However, several
observations argue against behavioural activation, sensorimotor
feedback and/or nonspeci®c arousing inputs being the principal
determinants of changes in cortical gene expression in our studies.
First, while possible, it is not obvious how such effects could account
for the distinct regional differences found. For example, after acute
dopamine depletion, the insular cortex was affected to a similar
degree as sensory and motor areas, whereas the cingulate cortex was
not affected. Also, in the contralateral cortex, in contrast to the
unaffected cingulate regions, the most robust decrease was observed
in the caudally adjacent retrosplenial cortex. Second, rats displayed
signi®cantly lower zif 268 mRNA levels ipsilateral than contralateral
to the lesion across all affected cortical areas, despite an almost
complete cessation of behaviour. Third, a dissociation between
cortical gene regulation and behavioural activity was also found in
our previous study. Thus, intrastriatal infusion of a low dose of the
D1 antagonist SCH-23390 produced a bilateral partial attenuation of
apomorphine-induced gene expression across the cortex without
causing noticeable behavioural inhibition (Steiner & Kitai, 2000b).
These ®ndings suggest that at least part of the present changes in
cortical gene expression were a direct consequence of dopamine
depletion and ensuing neuronal changes rather than the result of
behavioural changes.
Barrel cortex function and whisker use after dopaminedepletion
In order to directly compare whisker-evoked responses in the barrel
cortex with whisker-guided behaviour after the 6-OHDA lesion, we
assessed the amount of scanning during exploration of the open ®eld.
When rats travel along the wall of such a ®eld, they generally
maintain a certain distance to the wall that allows them to touch the
wall with their mystacial whiskers (`scanning'). The importance of
whisker sensory input for this behaviour is demonstrated by the
®nding that scanning is lateralized after clipping the mystacial
whiskers on one side; rats then follow the wall preferentially with the
whisker-intact side orientated towards the wall (Steiner et al., 1986;
J
J
J
JJ
J
O
O
OO
O
O
FF
F
F
F
F
â
â
ââ â
â
â
â
-20
0
20
40
60
80
100
120
140
160
-20 0 20 40 60 80 100 120 140 160
O
O
OO
O
O
â
â
â ââ
â
â
â
-20
0
20
40
60
80
100
120
140
160
100 120 140 160 180
O
O
O
O
O
O
â
â
â
â
â
â
â
â
-20
0
20
40
60
80
100
120
140
160
100 120 140 160 180
zif 268 mRNA (% of V)
c-fo
s m
RN
A (
% o
f V)
enkephalin mRNA (% of intact) enkephalin mRNA (% of intact)c-
fos
mR
NA
(%
of V
)
zif 2
68 m
RN
A (
% o
f V)
A CB
r = 0.00
r = -0.88*
r = 0.16
r = -0.60
r = 0.93***
2 daysV
621 days
V
6
FIG. 7. Correlation between enkephalin expression in the dopamine-depleted striatum and whisker stimulation-evoked immediate-early gene expression in theipsilateral barrel cortex 2 days, but not 21 days, after dopamine depletion. (A) Correlation between stimulation-evoked expression of c-fos and zif 268 (meandensity values, expressed as percentage of those in vehicle controls, % of V) in barrel C2 in individual animals that received an infusion of vehicle (V) or6-OHDA (6) and were stimulated either 2 or 21 days after the infusion. (B,C) Relationship between mid-striatal enkephalin mRNA levels (expressed aspercentage of those in the noninfused hemisphere, % of intact) and stimulation-evoked c-fos (B) and zif 268 expression (C) in the ipsilateral barrel C2 isshown for 6-OHDA-infused rats (6) stimulated 2 or 21 days after the lesion. ***P < 0.001, *P < 0.05.
Dopamine depletion and cortical function 1399
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404
Milani et al., 1989). Likely a re¯ection of the contralateral
sensorimotor `neglect', rats with unilateral dopamine depletion also
show lateralized scanning; they mostly follow the wall or edge of the
®eld with the side ipsilateral to the lesion (Steiner et al., 1988;
Fornaguera et al., 1993; Sullivan et al., 1994). Several reports showed
that such scanning is a sensitive behavioural index for compromised
dopamine function (Mattioli et al., 1992; Fornaguera et al., 1993;
Emmi et al., 1997).
In our study, 2 days after the lesion both scanning and whisker
stimulation-evoked immediate-early gene expression in the barrel
cortex were largely abolished. In contrast, 21 days after dopamine
depletion, such sensory-evoked cortical gene expression had also
recovered. At this time, rats displayed near-normal amounts of
scanning with the whiskers functionally related to the noninfused
hemisphere. However, minimal or no recovery was seen for scanning
related to the dopamine-depleted hemisphere, con®rming earlier
®ndings (see above). Therefore, these rats showed a de®cit in active
whisker use at a time when passive stimulation of these whiskers
elicited near-normal immediate-early gene responses in the barrel
cortex. These results thus show a dissociation between cortical
function, as indicated by immediate-early gene expression, and
behaviour for the dopamine-depleted hemisphere.
We also investigated whether there was a relationship between
changes in such scanning behaviour and neuroplasticity in the
dopamine-depleted striatum. After dopamine lesions various time-
dependent neuronal changes have been identi®ed in the basal ganglia,
including changes in the expression of receptors, enzymes and
neuropeptides, some of which probably contribute to recovery of
function (Schwarting & Huston, 1996a; for review). Consistently,
early alterations in neuronal ®ring followed by `normalization' have
been reported for several basal ganglia nuclei or their thalamic targets
(e.g. Schultz & Ungerstedt, 1978; Nisenbaum et al., 1986; MacLeod
et al., 1990; Ni et al., 2000), but not for others (e.g. Nisenbaum et al.,
1988; Vila et al., 2000). In our study, 21 days after dopamine
depletion recovery of scanning with the whiskers functionally related
to the hemisphere opposite to dopamine depletion was positively
correlated with the increase in enkephalin expression in the
dopamine-depleted striatum. In contrast, no relationship was seen
between enkephalin expression and scanning by the dopamine-
depleted hemisphere (or sensory-evoked immediate-early gene
expression in that hemisphere). It is unclear whether increased
enkephalin peptide function in the lesioned striatum was causally
related to such recovery of scanning by the noninfused hemisphere.
However, these ®ndings are consistent with neuroadaptive changes in
the dopamine-depleted striatum contributing to recovery of function
of the contralateral hemisphere, and thus with interhemispheric
effects of striatal output. On the other hand, such neuronal changes
are insuf®cient for recovery of behaviour related to the ipsilateral
hemisphere.
Potential pathways mediating the cortical facilitation
Together, our present and previous (Steiner & Kitai, 2000b) results
suggest a widespread, to some degree bilateral, facilitatory in¯uence
of dopamine on cortical function mediated by D1 receptors in the
striatum (Fig. 9). It is currently unknown which of several possible
anatomical pathways could convey this basal ganglia output to the
cortex. Current models of basal ganglia function are insuf®cient to
explain this effect (Alexander et al., 1986; Alexander & Crutcher,
1990; Alexander et al., 1990; but see Middleton & Strick, 2000). For
one, largely based on primate anatomy, these models emphasize basal
ganglia output directed towards frontal/premotor/motor cortical areas,
although some cortical regions outside the frontal lobe have also been
shown to be targets of basal ganglia output in primates (cf. Middleton
& Strick, 1996). Also, these models do not address interhemispheric
interactions. However, crossed pallidothalamic and nigrothalamic
projections have been described in primates and rats (for review, see
Parent & Hazrati, 1995).
In the rat, basal ganglia±cortical circuits are mostly routed through
the ventromedial and intralaminar/midline thalamic nuclei (e.g. Faull
& Mehler, 1978; Beckstead et al., 1979; Gerfen et al., 1982; for
reviews, see Groenewegen & Berendse, 1994; Gerfen & Wilson,
1996) that have widespread access to the cortex (Herkenham, 1979;
Herkenham, 1980; Arbuthnott et al., 1990). Widespread projections
from midline thalamic nuclei to the cortex have also been shown in
ϑ
ϑϑ
ϑϑ
ϑ
ϑ
Ο
Ο
Ο
Ο
Ο
Ο
0
20
40
60
80
100
120
140
160
180
100 120 140 160 180
ϑ
ϑ
ϑ
ϑ ϑ
ϑ
ϑ
ΟΟ
ΟΟ
Ο
Ο
0
20
40
60
80
100
120
140
160
180
100 120 140 160 180
B
A
r = 0.57*
r = 0.19
scan
ning
(%
of V
)
enkephalin mRNA (% of intact)
enkephalin mRNA (% of intact)
scan
ning
(%
of V
)
21 daysnoninfused
6-OHDA
FIG. 8. Correlation between enkephalin expression in the dopamine-depletedstriatum and whisker utilization (scanning) by the noninfused hemisphere,but not by the 6-OHDA-infused hemisphere, 21 days after dopaminedepletion. (A and B) Striatal enkephalin expression (mean density values,expressed as percentage of those in the noninfused hemisphere, % of intact)at the middle striatal level is compared to the amount of scanning(expressed as percentage of that in vehicle-infused controls, % of V) withthe whisker side related to the noninfused hemisphere (A) and that relatedto the dopamine-depleted hemisphere (6-OHDA) (B) in individual animalswith dopamine lesions (21 days, two experiments pooled). Note thatremoving the outlier at 140/174 in A improves the positive correlation tor = 0.83 (P < 0.001). *P < 0.05.
1400 H. Steiner and S. T. Kitai
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

the monkey (e.g. Bentivoglio et al., 2000). Interestingly, a recent
tracing study in the rat showed that neurons of the ventromedial
thalamic nucleus send axon collaterals to both the whisker motor and
the barrel cortex, thus providing a potential anatomical route by
which basal ganglia output could directly in¯uence both of these
cortical areas (Jones et al., 2000).
However, it is also possible that pathways not classically
considered part of basal ganglia±cortical circuits contribute to such
basal ganglia regulation of cortical function. For example, it has been
shown that unilateral dopamine depletion produces bilateral changes
in gene expression in the reticular nucleus of the thalamus (Delfs
et al., 1996), a structure that also receives inputs from the basal
ganglia and is of central importance for the regulation of cortical
activation. Therefore, several candidate anatomical pathways exist
that could mediate the widespread facilitatory in¯uence of striatal
dopamine on cortical function suggested by our studies. Future
experimentation will have to elucidate the possible contributions of
these pathways.
Functional signi®cance: a hypothesis
Regardless of the speci®c connections involved, the present time-
dependent effects on cortical gene regulation in comparison with the
associated behavioural effects may offer some indications as to the
functional organization of this basal ganglia±cortical input. Our
results suggest that this input provides rather general, bilateral
cortical facilitation and is likely superimposed upon and acts in
concert with the speci®c (e.g. motor) basal ganglia±cortical circuitry
(Fig. 9). For example, after unilateral dopamine depletion, the early
bilateral behavioural breakdown was followed by complete recovery
in behaviour (scanning) dependent on speci®c sensorimotor functions
of the contralateral hemisphere, but not in scanning related to the
hemisphere ipsilateral to the lesion. Similarly, early bilateral de®cits
and some subsequent recovery for the `intact' side were found for
skilled forelimb reaching movements after unilateral 6-OHDA lesions
(e.g. Miklyaeva et al., 1994; Whishaw et al., 1997). Whilst it is clear
that activity in sensorimotor basal ganglia-cortical circuits is
extensively altered in the dopamine-depleted hemisphere (e.g.
Albin et al., 1989; Levy et al., 1997; Obeso et al., 1997), our
®ndings suggest that the bilateral facilitatory cortical input is only
transiently suppressed in both hemispheres, perhaps because of
remaining cortical input from the unlesioned basal ganglia and/or
neuroadaptive changes following the lesion (see above). Thus, after
an initial period of general cortical failure re¯ected by the general
behavioural breakdown, recovery of the widespread cortical input
seems to allow recovery of contralateral cortical function and
dependent behaviours, and probably `mixed' behaviours to some
degree (e.g. locomotion), but not (or considerably less) of `ipsilateral'
behaviours that are still affected by compromised function in the
ipsilateral speci®c sensorimotor circuits.
Our results thus suggest dual basal ganglia±cortical inputs
(unilateral speci®c and bilateral general, facilitatory) that seem to
arise in the same striatal areas (Fig. 9; Steiner & Kitai, 2000b). Both
seem to be necessary for normal cortical function and behaviour. This
concept of dual basal ganglia±cortical inputs may provide an
improved framework for investigating and better understanding
how the basal ganglia in general, and dopamine in particular, regulate
cortical function.
Conclusions
The results of our studies suggest that stimulation of D1 dopamine
receptors in the striatum provides bilateral facilitation of cortical
function that is necessary for normal behaviour. This facilitation
affects many cortical regions, which may explain in part the diversity
of functions and disorders associated with dopamine.
Acknowledgements
This work was supported by the National Parkinson Foundation and USPHSGrants DA11261 (H.S.) and NS26473 (S.T.K.). We thank Weiwen Sun forexcellent technical assistance and Pastor Couceyro for helpful discussions.
Abbreviation
6-OHDA, 6-hydroxydopamine.
whiskerC2
barrelC2
D1
specific (e.g., motor)
widespread facilitatory
striatum
++
++
+
+
++
++
FIG. 9. Schematic illustration of thewidespread, to some degree bilateral,facilitatory input to the cortex arising in thestriatum, as indicated by our ®ndings. Thiscortical facilitation is dependent on D1receptors in the striatum (Steiner & Kitai,2000b). It is presently unknown which ofseveral possible anatomical circuits from thestriatum to the cortex mediates this facilitation(see Discussion). However, our results suggestthat the facilitatory circuits (solid arrows) are,partly or completely, separate from andsuperimposed upon the speci®c (e.g. motor;broken arrows) basal ganglia±cortical circuitsthat target the frontal lobe. The whisker-to-barrel pathway from the left whisker C2 to theright barrel C2 is indicated by a dotted line.
Dopamine depletion and cortical function 1401
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

References
Albin, R.L., Young, A.B. & Penney, J.B. (1989) The functional anatomy ofbasal ganglia disorders. Trends Neurosci., 12, 366±375.
Alexander, G.E. & Crutcher, M.D. (1990) Functional architecture of basalganglia circuits: neural substrates of parallel processing. Trends Neurosci.,13, 266±271.
Alexander, G.E., Crutcher, M.D. & DeLong, M.R. (1990) Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor,`prefrontal' and `limbic' functions. Prog. Brain Res., 85, 119±146.
Alexander, G.E., DeLong, M.R. & Strick, P.L. (1986) Parallel organization offunctionally segregated circuits linking basal ganglia and cortex. Annu. Rev.Neurosci., 9, 357±381.
Arbuthnott, G.W., MacLeod, N.K., Maxwell, D.J. & Wright, A.K. (1990)Distribution and synaptic contacts of the cortical terminals arising fromneurons in the rat ventromedial thalamic nucleus. Neuroscience, 38, 47±60.
Badiani, A., Oates, M.M., Day, H.E., Watson, S.J., Akil, H. & Robinson, T.E.(1998) Amphetamine-induced behavior, dopamine release, and c-fosmRNA expression: modulation by environmental novelty. J. Neurosci.,18, 10579±10593.
Beckstead, R.M., Domesick, V.B. & Nauta, W.J. (1979) Efferent connectionsof the substantia nigra and ventral tegmental area in the rat. Brain Res., 175,191±217.
Bentivoglio, M., Galkin, T., Ungerleider, L.G., Friedman, D. & Mishkin, M.(2000) Widespread projections from the medial thalamus to cortical layer Iin the monkey. Soc. Neurosci. Abstr., 26, 1710.
Berger, B., Gaspar, P. & Verney, C. (1991) Dopaminergic innervation of thecerebral cortex: unexpected differences between rodents and primates.Trends Neurosci., 14, 21±27.
Berke, J.D., Paletzki, R.F., Aronson, G.J., Hyman, S.E. & Gerfen, C.R. (1998)A complex program of striatal gene expression induced by dopaminergicstimulation. J. Neurosci., 18, 5301±5310.
BjoÈrklund, A. & Lindvall, O. (1984) Dopamine containing systems in theCNS. In BjoÈrklund, A. & HoÈkfelt, T. (eds), Handbook of ChemicalNeuroanatomy (Vol. 2, Part 1). Elsevier, London, pp. 55±122.
Bouthenet, M.-L., Souil, E., Martres, M.-P., Sokoloff, P., Giros, B. &Schwartz, J.-C. (1991) Localization of dopamine D3 receptor mRNA in therat brain using in situ hybridization histochemistry: comparison withdopamine D2 receptor mRNA. Brain Res., 564, 203±219.
Brog, J.S. & Zahm, D.S. (1995) Morphology and Fos immunoreactivity revealtwo subpopulations of striatal neurotensin neurons following acute 6-hydroxydopamine lesions and reserpine administration. Neuroscience, 65,71±86.
Brown, V.J. & Robbins, T.W. (1989) De®cits in response space followingunilateral striatal dopamine depletion in the rat. J. Neurosci., 9, 983±989.
Carli, M., Evenden, J.L. & Robbins, T.W. (1985) Depletion of unilateralstriatal dopamine impairs initiation of contralateral actions and not sensoryattention. Nature, 313, 679±682.
Carlsson, M. & Carlsson, A. (1990) Interactions between glutamatergic andmonoaminergic systems within the basal ganglia ± implications forschizophrenia and Parkinson's disease. Trends Neurosci., 13, 272±276.
Chaudhuri, A. (1997) Neural activity mapping with inducible transcriptionfactors. Neuroreport, 8, v±ix.
Ciliax, B.J., Nash, N., Heilman, C., Sunahara, R., Hartney, A., Tiberi, M., Rye,D.B., Caron, M.G., Niznik, H.B. & Levey, A.I. (2000) Dopamine D5receptor immunolocalization in rat and monkey brain. Synapse, 37, 125±145.
Cole, D.G. & Di Figlia, M. (1994) Reserpine increases Fos activity in the ratbasal ganglia via a quinpirole-sensitive mechanism. Neuroscience, 60, 115±123.
Daunais, J.B. & McGinty, J.F. (1994) Acute and chronic cocaineadministration differentially alters striatal opioid and nuclear transcriptionfactor mRNAs. Synapse, 18, 35±45.
Defagot, M.C., Malchiodi, E.L., Villar, M.J. & Antonelli, M.C. (1997)Distribution of D4 dopamine receptor in rat brain with sequence-speci®cantibodies. Mol. Brain Res., 45, 1±12.
Delfs, J.M., Ciaramitaro, V.M., Soghomonian, J.J. & Chesselet, M.F. (1996)Unilateral nigrostriatal lesions induce a bilateral increase in glutamatedecarboxylase messenger RNA in the reticular thalamic nucleus.Neuroscience, 71, 383±395.
DeLong, M.R. (1990) Primate models of movement disorders of basal gangliaorigin. Trends Neurosci., 13, 281±285.
Dilts, R.P.J., Helton, T.E. & McGinty, J.F. (1993) Selective induction of Fosand FRA immunoreactivity within the mesolimbic and mesostriataldopamine terminal ®elds. Synapse, 13, 251±263.
Dragunow, M. & Robertson, H.A. (1988) Brain injury induces c-fos protein(s)
in nerve and glial-like cells in adult mammalian brain. Brain Res., 455, 295±299.
Emmi, A., Rajabi, H. & Stewart, J. (1997) Behavioral and neurochemicalrecovery from partial 6-hydroxydopamine lesions of the substantia nigra isblocked by daily treatment with D1/D5, but not D2, dopamine receptorantagonists. J. Neurosci., 17, 3840±3846.
Engber, T.M., Boldry, R.C., Kuo, S. & Chase, T.N. (1992) Dopaminergicmodulation of striatal neuropeptides: differential effects of D1 and D2receptor stimulation on somatostatin, neuropeptide Y, neurotensin,dynorphin and enkephalin. Brain Res., 581, 261±268.
Fallon, J.H., Koziell, D.A. & Moore, R.Y. (1978) Catecholamine innervationof the basal forebrain. II. Amygdala, suprarhinal cortex and entorhinalcortex. J. Comp. Neurol., 180, 509±532.
Fallon, J.H. & Moore, R.Y. (1978) Catecholamine innervation of the basalforebrain: IV. Topography of the dopamine projection to the basal forebrainand neostriatum. J. Comp. Neurol., 180, 545±580.
Fallon, J.H., Wang, C., Kim, Y., Canepa, N., Loughlin, S. & Seroogy, K.(1983) Dopamine- and cholecystokinin-containing neurons of the crossedmesostriatal projection. Neurosci. Lett., 40, 233±238.
Fass, B. & Butcher, L.L. (1981) Evidence for a crossed nigrostriatal pathwayin rats. Neurosci. Lett., 22, 109±113.
Faull, R.L. & Mehler, W.R. (1978) The cells of origin of nigrotectal,nigrothalamic and nigrostriatal projections in the rat. Neuroscience, 3, 989±1002.
Fornaguera, J., Carey, R.J., Dai, H., Huston, J.P. & Schwarting, R.K.W. (1994)Differentiation of motor inactivation from movement asymmetry effects inan animal model of hemi-parkinsonism. Neuroreport, 6, 173±176.
Fornaguera, J., Schwarting, R.K.W., Boix, F. & Huston, J.P. (1993)Behavioral indices of moderate nigrostriatal 6-hydroxydopamine lesion: apreclinical Parkinson's model. Synapse, 13, 179±185.
Gaspar, P., Bloch, B. & Le Moine, C. (1995) D1 and D2 receptor geneexpression in the rat frontal cortex: cellular localization in different classesof efferent neurons. Eur. J. Neurosci., 7, 1050±1063.
Gerfen, C.R., Engber, T.M., Mahan, L.C., Susel, Z., Chase, T.N., Monsma,F.J. Jr & Sibley, D.R. (1990) D1 and D2 dopamine receptor-regulated geneexpression of striatonigral and striatopallidal neurons. Science, 250, 1429±1432.
Gerfen, C.R., Keefe, K.A. & Gauda, E.B. (1995) D1 and D2 dopaminereceptor function in the striatum: coactivation of D1- and D2-dopaminereceptors on separate populations of neurons results in potentiatedimmediate-early gene response in D1-containing neurons. J. Neurosci.,15, 8167±8176.
Gerfen, C.R., Staines, W.A., Arbuthnott, G.W. & Fibiger, H.C. (1982) Crossedconnections of the substantia nigra in the rat. J. Comp. Neurol., 207, 283±303.
Gerfen, C.R. & Wilson, C.J. (1996) The basal ganglia. In Swanson, L.W.,BjoÈrklund, A. & HoÈkfelt, T. (eds), Handbook of Chemical Neuroanatomy.Elsevier, London, pp. 371±468.
Graybiel, A.M. (1995) Building action repertoires: memory and learningfunctions of the basal ganglia. Curr. Opin. Neurobiol., 5, 733±741.
Groenewegen, H.J. & Berendse, H.W. (1994) The speci®city of the`nonspeci®c' midline and intralaminar thalamic nuclei. Trends Neurosci.,17, 52±57.
Herdegen, T. & Leah, J.D. (1998) Inducible and constitutive transcriptionfactors in the mammalian nervous system: control of gene expression byJun, Fos and Krox, and CREB/ATF proteins. Brain Res. Rev., 28, 370±490.
Herkenham, M. (1979) The afferent and efferent connections of theventromedial thalamic nucleus in the rat. J. Comp. Neurol., 183, 487±517.
Herkenham, M. (1980) Laminar organization of thalamic projections to the ratneocortex. Science, 207, 532±535.
Hollerman, J.R., Tremblay, L. & Schultz, W. (2000) Involvement of basalganglia and orbitofrontal cortex in goal-directed behavior. Prog. Brain Res.,126, 193±215.
Jacobs, O., Van Bree, L., Mailleux, P., Zhang, F., Schiffmann, S.N., Halleux,P., Albala, N. & Vanderhaeghen, J.J. (1994) Homolateral cerebrocorticalincrease of immediate early gene and neurotransmitter messenger RNAsafter minimal cortical lesion: blockade by N-methyl-D-aspartate antagonist.Neuroscience, 59, 827±836.
Jones, M.O., Whiteley, S.J.O., Mayhew, J.E. & Redgrave, P. (2000) Returnloop from basal ganglia to barrel cortex: An anatomical tracing study in rat.Soc. Neurosci. Abstr., 26, 964.
LaHoste, G.J., Ruskin, D.N. & Marshall, J.F. (1996) Cerebrocortical Fosexpression following dopaminergic stimulation: D1/D2 synergism and itsbreakdown. Brain Res., 728, 97±104.
Levy, R., Hazrati, L.N., Herrero, M.T., Vila, M., Hassani, O.K., Mouroux, M.,
1402 H. Steiner and S. T. Kitai
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

Ruberg, M., Asensi, H., Agid, Y., Feger, J., Obeso, J.A., Parent, A. &Hirsch, E.C. (1997) Re-evaluation of the functional anatomy of the basalganglia in normal and Parkinsonian states. Neuroscience, 76, 335±343.
Li, S.J., Jiang, H.K., Stachowiak, M.S., Hudson, P.M., Owyang, V., Nanry, K.,Tilson, H.A. & Hong, J.S. (1990) In¯uence of nigrostriatal dopaminergictone on the biosynthesis of dynorphin and enkephalin in rat striatum. Mol.Brain Res., 8, 219±225.
Lindefors, N., Brene, S., Herrera-Marschitz, M. & Persson, H. (1989) Regionspeci®c regulation of glutamic acid decarboxylase mRNA expression bydopamine neurons in rat brain. Exp. Brain Res., 77, 611±620.
Lindefors, N., Brene, S., Herrera-Marschitz, M. & Persson, H. (1990)Neuropeptide gene expression in brain is differentially regulated bymidbrain dopamine neurons. Exp. Brain Res., 80, 489±500.
Ljungberg, T. & Ungerstedt, U. (1976) Sensory inattention produced by 6-hydroxydopamine-induced degeneration of ascending dopamine neurons inthe brain. Exp. Neurol., 53, 585±600.
MacLeod, N.K., Ryman, A. & Arbuthnott, G.W. (1990) Electrophysiologicalproperties of nigrothalamic neurons after 6-hydroxydopamine lesions in therat. Neuroscience, 38, 447±456.
Mansour, A., Meador-Woodruff, J.H., Bunzow, J.R., Civelli, O., Akil, H. &Watson, S.J. (1990) Localization of dopamine D2 receptor mRNA and D1and D2 receptor binding in the rat brain and pituitary: an in situhybridization-receptor autoradiographic analysis. J. Neurosci., 10, 2587±2600.
Marshall, J.F., Richardson, J.S. & Teitelbaum, P. (1974) Nigrostriatal bundledamage and the lateral hypothalamic syndrome. J. Comp. Physiol. Psychol.,87, 808±830.
Mattioli, R., Schwarting, R.K.W. & Huston, J.P. (1992) Recovery fromunilateral 6-hydroxydopamine lesion of substantia nigra promoted by theneurotachykinin substance P 1±11. Neuroscience, 48, 595±605.
Melzer, P. & Steiner, H. (1997) Stimulus-dependent expression of immediate-early genes in rat somatosensory cortex. J. Comp. Neurol., 380, 145±153.
Melzer, P., Van der Loos, H., DoÈr¯, J., Welker, E., Robert, P., Emery, D. &Berrini, J.-C. (1985) A magnetic device to stimulate selected whiskers offreely moving or restrained small rodents: its application in a deoxyglucosestudy. Brain Res., 348, 229±240.
Mengod, G., Vilaro, M.T., Niznik, H.B., Sunahara, R.K., Seeman, P.,O'Dowd, B.F. & Palacios, J.M. (1991) Visualization of a dopamine D1receptor mRNA in human and rat brain. Mol. Brain Res., 10, 185±191.
Middleton, F.A. & Strick, P.L. (1996) The temporal lobe is a target of outputfrom the basal ganglia. Proc. Natl Acad. Sci. USA, 93, 8683±8687.
Middleton, F.A. & Strick, P.L. (2000) Basal ganglia and cerebellar loops:motor and cognitive circuits. Brain Res. Rev., 31, 236±250.
Miklyaeva, E.I., Castaneda, E. & Whishaw, I.Q. (1994) Skilled reachingde®cits in unilateral dopamine-depleted rats: impairments in movement andposture and compensatory adjustments. J. Neurosci., 14, 7148±7158.
Milani, H., Steiner, H. & Huston, J.P. (1989) Analysis of recovery frombehavioral asymmetries induced by unilateral removal of vibrissae in therat. Behav. Neurosci., 103, 1067±1074.
Ni, Z.G., Gao, D.M., Benabid, A.L. & Benazzouz, A. (2000) Unilateral lesionof the nigrostriatal pathway induces a transient decrease of ®ring rate withno change in the ®ring pattern of neurons of the parafascicular nucleus inthe rat. Neuroscience, 101, 993±999.
Nisenbaum, L.K., Crowley, W.R. & Kitai, S.T. (1996) Partial striataldopamine depletion differentially affects striatal substance P andenkephalin messenger RNA expression. Mol. Brain Res., 37, 209±216.
Nisenbaum, L.K., Kitai, S.T., Crowley, W.R. & Gerfen, C.R. (1994) Temporaldissociation between changes in striatal enkephalin and substance Pmessenger RNAs following striatal dopamine depletion. Neuroscience,60, 927±937.
Nisenbaum, E.S., Stricker, E.M., Zigmond, M.J. & Berger, T.W. (1986) Long-term effects of dopamine-depleting brain lesions on spontaneous activity oftype II striatal neurons: relation to behavioral recovery. Brain Res., 398,221±230.
Nisenbaum, E.S., Stricker, E.M., Zigmond, M.J. & Berger, T.W. (1988)Spontaneous activity of type II but not type I striatal neurons is correlatedwith recovery of behavioral function after dopamine-depleting brain lesions.Brain Res., 473, 389±393.
Normand, E., Popovici, T., Onteniente, B., Fellmann, D., Piatier-Tonneau, D.,Auffray, C. & Bloch, B. (1988) Dopaminergic neurons of the substantianigra modulate preproenkephalin A gene expression in rat striatal neurons.Brain Res., 439, 39±46.
Obeso, J.A., Rodriguez, M.C. & DeLong, M.R. (1997) Basal gangliapathophysiology. A critical review. Adv. Neurol., 74, 3±18.
Parent, A. & Hazrati, L.N. (1995) Functional anatomy of the basal ganglia. I.
The cortico-basal ganglia-thalamo-cortical loop. Brain Res. Rev., 20, 91±127.
Paul, M.L., Graybiel, A.M., David, J.-C. & Robertson, H.A. (1992) D1-likeand D2-like dopamine receptors synergistically activate rotation and c-fosexpression in the dopamine-depleted striatum in a rat model of Parkinson'sdisease. J. Neurosci., 12, 3729±3742.
Paxinos, G. & Watson, C. (1998) The Rat Brain in Stereotaxic Coordinates.Academic Press, New York.
Penney, J.B.Jr. & Young, A.B. (1983) Speculations on the functional anatomyof basal ganglia disorders. Annu. Rev. Neurosci., 6, 73±94.
Pritzel, M., Sarter, M., Morgan, S. & Huston, J.P. (1983) Interhemisphericnigrostriatal projections in the rat: bifurcating nigral projections and loci ofcrossing in the diencephalon. Brain Res. Bull., 10, 385±390.
Redgrave, P., Prescott, T.J. & Gurney, K. (1999) The basal ganglia: avertebrate solution to the selection problem? Neuroscience, 89, 1009±1023.
Robbins, T.W., Granon, S., Muir, J.L., Durantou, F., Harrison, A. & Everitt,B.J. (1998) Neural systems underlying arousal and attention. Implicationsfor drug abuse. Ann. N. Y. Acad. Sci., 846, 222±237.
Rodriguez-Puertas, R., Herrera-Marschitz, M., Koistinaho, J. & Hokfelt, T.(1999) Dopamine D1 receptor modulation of glutamate receptor messengerRNA levels in the neocortex and neostriatum of unilaterally 6-hydroxydopamine-lesioned rats. Neuroscience, 89, 781±797.
Sarter, M. & Bruno, J.P. (1999) Abnormal regulation of corticopetalcholinergic neurons and impaired information processing inneuropsychiatric disorders. Trends Neurosci., 22, 67±74.
Schuller, J.J. & Marshall, J.F. (2000) Acute immediate-early gene response to6-hydroxydopamine infusions into the medial forebrain bundle.Neuroscience, 96, 51±58.
Schultz, W., Tremblay, L. & Hollerman, J.R. (1998) Reward prediction inprimate basal ganglia and frontal cortex. Neuropharmacology, 37, 421±429.
Schultz, W. & Ungerstedt, U. (1978) Short-term increase and long-termreversion of striatal cell activity after degeneration of the nigrostriataldopamine system. Exp. Brain Res., 33, 159±171.
Schwarting, R.K.W. & Huston, J.P. (1996a) Unilateral 6-hydroxydopaminelesions of meso-striatal dopamine neurons and their physiological sequelae.Prog. Neurobiol., 49, 215±266.
Schwarting, R.K.W. & Huston, J.P. (1996b) The unilateral 6-hydroxydopamine lesion model in behavioral brain research. Analysis offunctional de®cits, recovery and treatments. Prog. Neurobiol., 50, 275±331.
Sharp, F.R., Sagar, S.M. & Swanson, R.A. (1993) Metabolic mapping withcellular resolution: c-fos vs. 2-deoxyglucose. Crit. Rev. Neurobiol., 7, 205±228.
Steiner, H., Blum, M., Kitai, S.T. & Fedi, P. (1999) Differential expression ofErbB3 and ErbB4 neuregulin receptors in dopamine neurons and forebrainareas of the adult rat. Exp. Neurol., 159, 494±503.
Steiner, H., Bonatz, A.E., Huston, J.P. & Schwarting, R. (1988) Lateralizedwall-facing versus turning as measures of behavioral asymmetries andrecovery of function after injection of 6-hydroxydopamine into thesubstantia nigra. Exp. Neurol., 99, 556±566.
Steiner, H. & Gerfen, C.R. (1994) Tactile sensory input regulates basal andapomorphine-induced immediate-early gene expression in rat barrel cortex.J. Comp. Neurol., 344, 297±304.
Steiner, H. & Gerfen, C.R. (1995) Dynorphin opioid inhibition of cocaine-induced, D1 dopamine receptor-mediated immediate-early gene expressionin the striatum. J. Comp. Neurol., 353, 200±212.
Steiner, H. & Gerfen, C.R. (1996) Dynorphin regulates D1 dopamine receptor-mediated responses in the striatum: relative contributions of pre- andpostsynaptic mechanisms in dorsal and ventral striatum demonstrated byaltered immediate-early gene induction. J. Comp. Neurol., 376, 530±541.
Steiner, H. & Gerfen, C.R. (1998) Role of dynorphin and enkephalin in theregulation of striatal output pathways and behavior. Exp. Brain Res., 123,60±76.
Steiner, H. & Gerfen, C.R. (1999) Enkephalin regulates acute D2 dopaminereceptor antagonist-induced immediate-early gene expression in striatalneurons. Neuroscience, 88, 795±810.
Steiner, H., Huston, J.P. & Morgan, S. (1986) Apomorphine reverses directionof asymmetry in facial scanning after 10 days of unilateral vibrissaeremoval in rat: vibrissotomy-induced denervation supersensitivity? Behav.Brain Res., 22, 283±287.
Steiner, H. & Kitai, S.T. (2000a) Basal ganglia±cortex interactions: Effects ofdopamine depletion on cortical function and behavioral correlates. Soc.Neurosci. Abstr., 26, 1711.
Steiner, H. & Kitai, S.T. (2000b) Regulation of rat cortex function by D1dopamine receptors in the striatum. J. Neurosci., 20, 5449±5460.
Sullivan, R.M., Fraser, A. & Szechtman, H. (1994) Asymmetrical orientation
Dopamine depletion and cortical function 1403
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404

to edges of an open®eld: modulation by striatal dopamine and relationshipto motor asymmetries in the rat. Brain Res., 637, 114±118.
Svenningsson, P., Fourreau, L., Bloch, B., Fredholm, B.B., Gonon, F. &Le Moine, C. (1999) Opposite tonic modulation of dopamine andadenosine on c-fos gene expression in striatopallidal neurons.Neuroscience, 89, 827±837.
Ungerstedt, U. (1971) Striatal dopamine release after amphetamine or nervedegeneration revealed by rotational behaviour. Acta Physiol. Scand.(Suppl.), 367, 49±68.
Vila, M., PeÂrier, C., FeÂger, J., Yelnik, J., Faucheux, B., Ruberg, M., Raisman-Vozari, R., Agid, Y. & Hirsch, E.C. (2000) Evolution of changes inneuronal activity in the subthalamic nucleus of rats with unilateral lesion ofthe substantia nigra assessed by metabolic and electrophysiologicalmeasurements. Eur. J. Neurosci., 12, 337±344.
Voorn, P., Roest, G. & Groenewegen, H.J. (1987) Increase of enkephalin anddecrease of substance P immunoreactivity in the dorsal and ventral striatum
of the rat after midbrain 6-hydroxydopamine lesions. Brain Res., 412, 391±396.
Wang, J.Q. & McGinty, J.F. (1995) Differential effects of D1 and D2dopamine receptor antagonists on acute amphetamine- ormethamphetamine-induced up-regulation of zif/268 mRNA expression inrat forebrain. J. Neurochem., 65, 2706±2715.
Whishaw, I.Q., Woodward, N.C., Miklyaeva, E. & Pellis, S.M. (1997)Analysis of limb use by control rats and unilateral DA-depleted rats in theMontoya staircase test: movements, impairments and compensatorystrategies. Behav. Brain Res., 89, 167±177.
Young, W.S. III, Bonner, T.I. & Brann, M.R. (1986) Mesencephalic dopamineneurons regulate the expression of neuropeptide mRNAs in the ratforebrain. Proc. Natl. Acad. Sci. USA, 83, 9827±9831.
Zahm, D.S. (1991) Compartments in rat dorsal and ventral striatum revealedfollowing injection of 6-hydroxydopamine into the ventral mesencephalon.Brain Res., 552, 164±169.
1404 H. Steiner and S. T. Kitai
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1390±1404