the aging striatal dopamine function
TRANSCRIPT

at SciVerse ScienceDirect
Parkinsonism and Related Disorders 18 (2012) 426e432
Contents lists available
Parkinsonism and Related Disorders
journal homepage: www.elsevier .com/locate/parkreldis
Review
The aging striatal dopamine function
Olivier Darbin a,b,*
aDepartment of Neurology, University South Alabama, 307 University Blvd., Mobile, AL 36688, USAbDivision of System Neurophysiology, National Institute for Physiological Sciences, Okazaki, Japan
a r t i c l e i n f o
Article history:Received 27 August 2011Received in revised form23 November 2011Accepted 27 November 2011
Keywords:ElderlyCatecholamineNigro-striatal pathwayMotor activityDopamine depletion
* Corresponding author. Department of Neurology,College of Medicine, 3401 Medical Park Dr., Bldg 3,USA. Tel.: þ1 770 329 8773; fax: þ1 251 660 5924.
E-mail address: [email protected].
1353-8020/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.parkreldis.2011.11.025
a b s t r a c t
Movement disorders are prevalent in the elderly and may have both central and peripheral origins. Age-related parkinsonism often results in movement disorders identical to some of the cardinal symptoms oftypical Parkinson’s disease (TPD). Nevertheless, there may be limited similarity in the underlyingdysfunction of the sensory-motor circuitry since these two conditions exhibit different changes in thenigro-striatal pathway. In this short review, we highlight some of the key distinctions between aging andTPD regarding striatal dopaminergic activity and discuss them in the context of therapeutic strategies toalleviate motor decline in the elderly.
� 2011 Elsevier Ltd. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4262. Aging and striatal dopaminergic neurotransmission . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .427
2.1. Striatal dopamine metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4272.2. Striatal dopamine receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4272.3. Age-related response to dopaminergic treatments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4272.4. Aged-related striatal dopamine alterations: what are the possible consequences on basal ganglia circuitry activity? . . . . . . . . . . . . . . . . . . . . . 4282.5. Striatal dopaminergic system, aging and Parkinson’s disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 429
3. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 430Acknowledgment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 430Full financial disclosure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 430References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 430
1. Introduction
Motor signs of parkinsonism increase with aging and affectmore than 50% of people over the age of 85 years [1]. They arepredictive of lifespan [1e3], contribute to the perception of dete-rioration in quality of life [4] and have an economical impact. In theelderly, motor symptoms with the highest prevalence include:bradykinesia (37%), gait disturbance (51%) and rigidity (43%) but
University of South AlabamaSuite 205, Mobile, AL 36608,
All rights reserved.
resting tremor, a cardinal symptom of typical Parkinson’s disease(TPD), has a low prevalence (5%) in the elderly [1]. In contrast toTPD, dopaminergic replacement strategies are ineffective atrelieving the burden associated with age-related parkinsonism.Though this lack of benefit may indicate complex and diffusealterations along the motor efferent pathway [5], it may also be dueto the fact that TPD and age-related parkinsonism exhibit differentneuropathological hallmarks in the central motor circuitry.Understanding the specificities of age-related parkinsonism, incomparison toTPD, is an important step for efficient cross disciplineresearch between these two conditions. Ultimately, the identifica-tion of reversible central dysfunctions in the aging central motorcircuitry could facilitate our ability to develop new therapeutic

O. Darbin / Parkinsonism and Related Disorders 18 (2012) 426e432 427
strategies to reduce the severity of or delay the onset of parkin-sonism related to aging.
Numerous structural studies have pointed to age-relatedchanges in the basal ganglia. The basal ganglia, as part of thecortico-cortical loops within the sensory-motor circuitry, areinvolved in the planning, initiation and control of voluntarymovement [6e9]. Specifically, dopamine depletion in the majorinput nucleus of the basal ganglia, e.g. the striatum, is manifestedby the inability to initiate and cease movement, the inability tosuppress involuntary movement, an abnormality in the velocityand amount of movement, and abnormal muscle tone [9e11]. Intypical PD, the depletion in striatal dopamine is considered to bea contributing factor to akinesia which is responsive to dopaminereplacement therapy. Markers for striatal dopaminergic activities,such as enzymes [12e15] and receptors [16e19], have consistentlybeen reported as being altered [12,20e24] in the aging basalganglia. However, Lewy body pathologies and loss of dopaminergiccells in the nigro-striatal pathway, which are hallmarks for TPD (i.e.see Ref. [25]), are not reliable markers for aging [20,23,24]. Age-related parkinsonism-like symptoms have been identified inexperimental animal models [2,26]. Here, we review evidence thatage-related alterations in striatal dopamine function are differentfrom those observed in TPD and could contribute, at least partially,to the poor response of age-related parkinsonism to dopaminereplacement therapies.
2. Aging and striatal dopaminergic neurotransmission
2.1. Striatal dopamine metabolism
Aging results in morphological and neurochemical changes inthe basal ganglia that may contribute to the decline in motor,cognitive and affective functions [27e32]. Parkinsonism related toaging may occur without major decrease in the number of nigraldopaminergic cells [23,33e38], and with only moderate degener-ation of the nigro-striatal pathway (15e45%) [23,25,26,39e46].This is an important distinction from TPD for which cardinal motorsymptoms occur when nigral dopaminergic cell loss exceeds 80%[47]. When it occurs in the aging brain, dopaminergic cell loss doesnot correlate in time with the development of motor symptoms. Infact, dopamine cell loss has been shown to be more prevalent inyoung and adult subjects than middle-aged and old subjects [23].
However, aging results in decreased striatal dopaminergicactivity in rodents [24,48e50], monkeys [51], primates [52] andhumans [13,14]. Early studies showed decreased levels in dopaminetissue content in the rat striatum [53]. In aged rodents [54e58] andaged non-human primates [59], in-vivo monitoring of the extra-cellular content of dopamine (and its first metabolite) by micro-dialysis confirmed decreased base line levels and evoked responseto a high potassium challenge. From this, one can infer that dopa-mine depletion, but not dopaminergic cell loss, is a hallmark foraging and correlates with the occurrence of motor decline [36](Fig. 1).
A decline in synthesis (rather than increased degradation)appears to be the main contributor to age-related dopaminedepletion. Clinical studies using positron emission tomographywith 11C-labeled L-DOPA have established an age-related decreasein striatal L-DOPA utilization and dopamine synthesis in the stria-tum of the neurologically normal elderly [21]. This was alsoconfirmed in aged non-human primates that exhibit a reducedincrease in striatal dopamine levels following local administrationof a DAT inhibitor [16]. In adult mammals, the dopamine synthesisis limited by the activity of tyrosine hydroxylase (TH) [49,60] and L-DOPA decarboxylase (DDC, or AADC for aromatic amino aciddecarboxylase). In aged subjects, the activity for both TH [12e15]
and DDC [16,61,62] is decreased but their relative contribution tothe depletion in striatal dopamine remains a matter of debate[12,13] (Fig. 2). In comparison to aging, typical PD is associated withan increased striatal dopaminergic metabolism in the survivingdopaminergic terminals as the loss of DA neurons progresses in thesubstantia nigra compacta [63,64] (Fig. 2).
Aging affects not only the striatal synthesis of dopamine butalso the local activity and expression of the dopamine transporter(DAT) [65e72]. At least two mechanisms appear to be involved inage-related decline in striatal DAT function. The first mechanisminvolves a DAT redistribution away from the plasma membrane[65] consecutive to a deficit in glycosylation [38]. The secondmechanism involves a decrease in DAT binding and DAT-mRNAwhich may be a down regulation in response to the decrease inDA levels [69].
As a consequence of this, it is remarkable that both aging andTPD result in striatal dopamine depletion. However, the origins ofthe structural and metabolic alterations of this depletion in dopa-mine differ greatly between these two conditions.
2.2. Striatal dopamine receptors
The age-related decrease in striatal dopamine metabolism isalso associated with changes in local dopamine receptors. Inrodents, monkeys and humans, studies have reported an age-related decline in the binding of selective agents to either D2-[16,17] or D1-dopamine receptors [16e19]. Therefore, the expres-sion for two DA receptor families is reduced in the aged striatum(Fig. 2). This is in contrast to TPD which, in fact, exhibits anincreased expression in striatal dopamine D2 receptors (but not D1)[73e76].
Age-related alteration in the striatal dopaminergic terminalsincludes functional impairments at both pre- and post-synapticlevels. In the next paragraph, we discuss the impact of thesealterations on the effects of dopaminergic treatment for age-relatedmotor decline.
2.3. Age-related response to dopaminergic treatments
In patients with typical PD and in the first half decade oftreatment, brain dopamine replacement is beneficial for somespecific motor symptoms including bradykinesia and hypokinesia[77]. In contrast to TPD, clinical studies performed in healthyelderly humans have reported poor or no benefit from dopaminereplacement therapy (L-DOPA) on age-related motor decline. Ina double-blind crossover study in normal elderly humans, theeffects of carbidopa/levodopa have been investigated onmovementvelocity, reaction time, tremor and visual evoked responses (VER).This treatment was found to have no benefit on both motor func-tions and VER [78]. More recently, the lack of efficacy of anti-parkinsonian dopaminergic medication was also reported onevent-related potentials (ERPs) during a stimulus-response (S-R)compatibility task in elderly humans [79]. It is probable thatdecreased DDC activity in the aged striatum [12] may contribute toa poor neuronal utilization of L-DOPA and the limited benefit of thistreatment for age-related motor decline [80e82]. In addition toa weak bio-transformation of L-DOPA, DA may also have a limitedaction since aging is associated with a decreased expression instriatal post-synaptic DA receptors. In aged rhesus monkeys,systemic administration of apomorphine weakly increases motoractivities in comparatively to its effect in young primates [83].Interestingly, experimental studies in rodents have shown that age-related decline in motor function is not reversed by D2R genetransfer in the striatum [84], suggesting that other mechanisms,downstream to post-synaptic dopaminergic receptors, may also
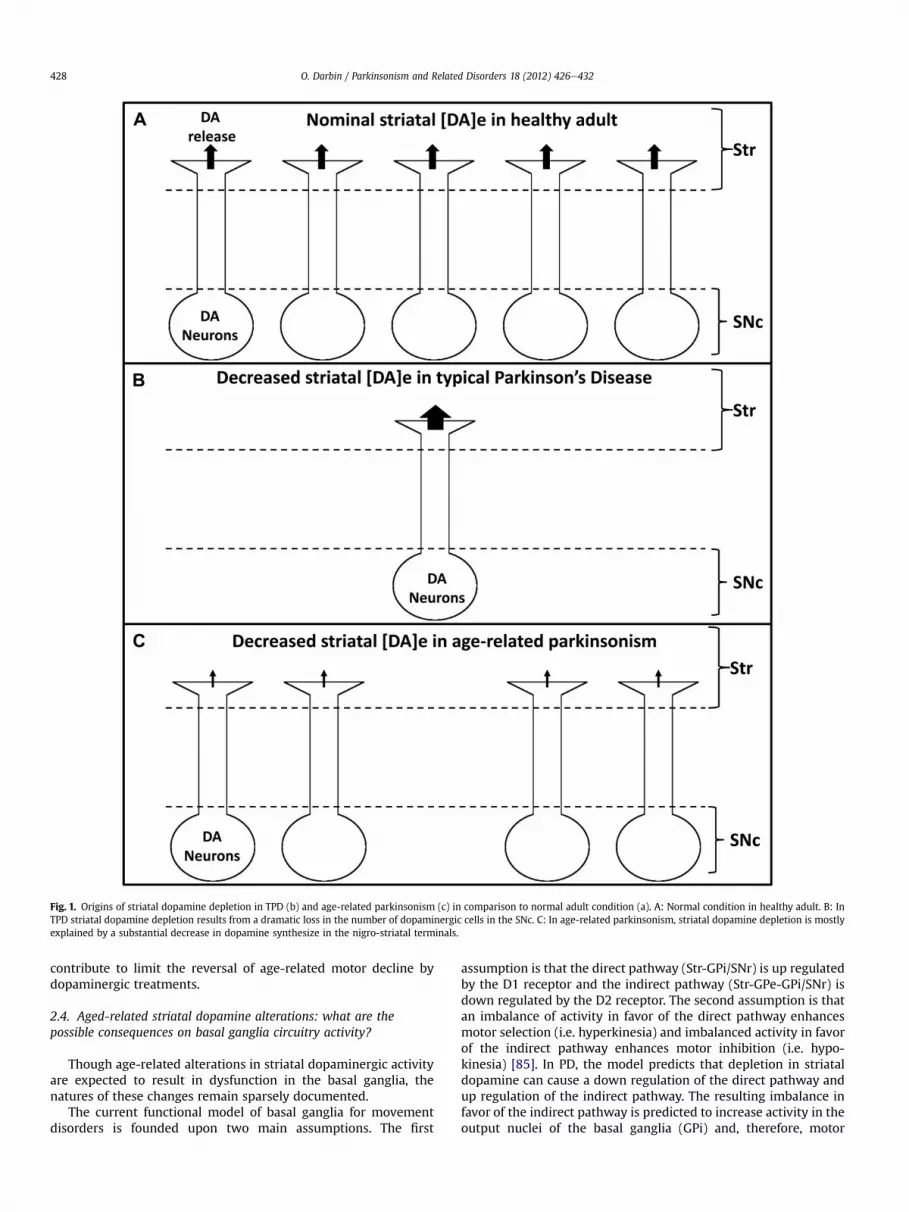
Fig. 1. Origins of striatal dopamine depletion in TPD (b) and age-related parkinsonism (c) in comparison to normal adult condition (a). A: Normal condition in healthy adult. B: InTPD striatal dopamine depletion results from a dramatic loss in the number of dopaminergic cells in the SNc. C: In age-related parkinsonism, striatal dopamine depletion is mostlyexplained by a substantial decrease in dopamine synthesize in the nigro-striatal terminals.
O. Darbin / Parkinsonism and Related Disorders 18 (2012) 426e432428
contribute to limit the reversal of age-related motor decline bydopaminergic treatments.
2.4. Aged-related striatal dopamine alterations: what are thepossible consequences on basal ganglia circuitry activity?
Though age-related alterations in striatal dopaminergic activityare expected to result in dysfunction in the basal ganglia, thenatures of these changes remain sparsely documented.
The current functional model of basal ganglia for movementdisorders is founded upon two main assumptions. The first
assumption is that the direct pathway (Str-GPi/SNr) is up regulatedby the D1 receptor and the indirect pathway (Str-GPe-GPi/SNr) isdown regulated by the D2 receptor. The second assumption is thatan imbalance of activity in favor of the direct pathway enhancesmotor selection (i.e. hyperkinesia) and imbalanced activity in favorof the indirect pathway enhances motor inhibition (i.e. hypo-kinesia) [85]. In PD, the model predicts that depletion in striataldopamine can cause a down regulation of the direct pathway andup regulation of the indirect pathway. The resulting imbalance infavor of the indirect pathway is predicted to increase activity in theoutput nuclei of the basal ganglia (GPi) and, therefore, motor

Fig. 2. Age-related changes in striatal dopamine neurotransmission (b) comparative to normal adult condition (a). A: In normal adult, Tyrosine (Tyr) is metabolized in L-DOPA bythe Tyrosine Hydroxylase (TH) which is transformed in Dopamine (DA) by the L-DOPA decarboxylase (DDC). Intracellular dopamine ([DA]i) is released by transporter-dependent orexocytotic mechanisms in the extra-cellular space ([DA]e). When in the extra-cellular space, dopamine modulates the medium spiny neurons through D1-like receptors for thedirect pathway or through D2-like receptors for the indirect pathway. B: In age-related parkinsonism, expression of both TH and DDC decreases the synthesis of dopamine resultingin striatal dopamine depletion. In addition, and at the post-synaptic levels, the expressions of both D1 and D2 family receptors are reduced.
O. Darbin / Parkinsonism and Related Disorders 18 (2012) 426e432 429
inhibition [86e89]. Though this model is rigorously debated, it maybe used as a reference for discussing basal ganglia dysfunction inregard to movement disorders.
In the aging striatum, depletion in both dopamine and dopa-minergic receptors has the potential to decrease the D2-mediatedinhibition on the indirect pathway and the D1-mediated excita-tion on the direct pathway. Concerning the model previously cited,it is conceivable that an increased activity in the GPi couldcontribute to age-related motor decline. A few fMRI studies haveattempted to determine age-related changes in basal gangliaactivity with a special focus on the output nuclei [90e92]. PreviousfMRI studies in the elderly have reported an aging-relatedincreased activity in the GP [93]. Due to technical limitations, theGPe and GPi were not distinguished in this last study and therefore,the study failed to identify the effects of aging specifically on theoutput nuclei of the basal ganglia (namely the GPi and the SN). Inanother study, the effects of the D1/D2 agonist apomorphine werespecifically investigated on the output nuclei of the basal ganglia[83]. In young non-human primates, systemic administration ofapomorphine increases the local measurement of blood oxygenlevel-dependency (BOLD response) in the GPi and SN [83],a response consistent with a decreased activity in the output nucleiand hyperkinesias, as predicted by the model. In aged primates, the
same pharmacological challenge failed to affect the activities in GPiand SN [83]. This finding reinforces the hypothesis that aging isassociated with decreased dopaminergic control of balancedactivity between the direct and indirect pathways and thereforefailed to report direct evidence for an imbalance between the directand indirect pathway.
From studies on the striatal dopaminergic synapse and theresponses of motor decline to dopaminergic treatment, there is anaccumulation of evidence that raises questions and controversiesabout whether aging and PD [52] exhibit similar dysfunctions in thebasal ganglia circuitry.
2.5. Striatal dopaminergic system, aging and Parkinson’s disease
Experimental studies in primates showed that even in theabsence of an overall neuronal loss, changes in the characteristics ofdopaminergic cells reflect functional deficits and increasedvulnerability to injury with age [23]. In addition, the overall age-related decrease in striatal dopamine neurotransmission maylimit some compensatory responses to the decrement in DA cellloss in PD [94] and therefore, in line with Collier et al. [95], have thepotential to decrease the threshold for the expression of parkin-sonian symptoms [95,96]. However, Lewy body pathology,

O. Darbin / Parkinsonism and Related Disorders 18 (2012) 426e432430
hallmarks of PD, is less prevalent [97] than signs for parkinsonism[1] in the elderly population. Therefore, there is neither evidence tosupport age-dependent neurodegenerative processes as primarycauses of TPD [14], nor to associate automatically, age-relatedparkinsonism to early stage TPD. Nevertheless, age-relatedparkinsonism likely contributes to age-dependent occurrences ofcomplications in patients with TPD [96] such as changes in clinicalsymptoms [98] and response to treatments [99,100].
3. Conclusion
Normal aging is associated with circuitry alterations in the basalganglia. The effects of aging on the striatal dopaminergic synapsediffer from those resulting from TPD. The specific effects of aging ondopaminergic neurotransmission in the striatum likely contribute toboth age-related parkinsonism and the poor improvement inmobility with dopaminergic treatment. These differences raisequestions about whether age-related parkinsonism and TPD sharesimilar basal ganglia dysfunctions. It is expected that the identifica-tion of electrophysiological hallmarks of aging in the basal gangliawill aid in developing a functionalmodel of the agingmotor circuitry.
The extent of age-related dysfunction at the level of the striataldopaminergic synapse suggests a shift of focus toward non-dopaminergic anti-parkinsonian medications [101] and morecomparisons with atypical parkinsonian conditions. Ultimately,identification of treatments that alleviate age-related parkinsonismmay also benefit the long-termmanagement of aging patients withTPD.
Acknowledgment
Dr O. Darbin is principally supported by the Department ofNeurology University of South Alabama College of Medicine,Mobile, AL and by the Division of System Neurophysiology at theNational Institute of Japan for Physiological Sciences (NIJPS-Oka-saki). The author would like to thank Dr D.E. Salter and Susan Calnefor the editing of the manuscript. This manuscript is dedicated tothe memory of Pr R. Joly.
Full financial disclosure
Nothing to report.
References
[1] Bennett DA, Beckett LA, Murray AM, Shannon KM, Goetz CG, Pilgrim DM,et al. Prevalence of parkinsonian signs and associated mortality ina community population of older people. N Engl J Med 1996;334:71e6.
[2] Ingram DK. Age-related decline in physical activity: generalization to non-humans. Med Sci Sports Exerc 2000;32:1623e9.
[3] Wilson RS, Schneider JA, Beckett LA, Evans DA, Bennett DA. Progression ofgait disorder and rigidity and risk of death in older persons. Neurology 2002;58:1815e9.
[4] Larsson L, Ramamurthy B. Aging-related changes in skeletal muscle. Mech-anisms and interventions. Drugs Aging 2000;17:303e16.
[5] Dickstein DL, Kabaso D, Rocher AB, Luebke JI, Wearne SL, Hof PR. Changes inthe structural complexity of the aged brain. Aging Cell 2007;6:275e84.
[6] Middleton FA, Strick PL. Basal ganglia and cerebellar loops: motor andcognitive circuits. Brain Res Brain Res Rev 2000;31:236e50.
[7] Alexander GE, Crutcher MD. Functional architecture of basal ganglia circuits:neural substrates of parallel processing. Trends Neurosci 1990;13:266e71.
[8] Delong MR. Primate models of movement-disorders of basal ganglia origin.Trends Neurosci 1990;13:281e5.
[9] Turner RS, Grafton ST, Votaw JR, Delong MR, Hoffman JM. Motor subcircuitsmediating the control of movement velocity: a PET study. J Neurophysiol1998;80:2162e76.
[10] Obeso JA, Rodriguez MC, DeLong MR. Basal ganglia pathophysiology. Acritical review. Adv Neurol 1997;74:3e18.
[11] Saint-Cyr JA. Basal ganglia: functional perspectives and behavioral domains.Adv Neurol 2005;96:1e16.
[12] Cruz-Muros I, Afonso-Oramas D, Abreu P, Barroso-Chinea P, Rodriguez M,Gonzalez MC, et al. Aging of the rat mesostriatal system: differences betweenthe nigrostriatal and the mesolimbic compartments. Exp Neurol 2007;204:147e61.
[13] Haycock JW, Becker L, Ang L, Furukawa Y, Hornykiewicz O, Kish SJ.Marked disparity between age-related changes in dopamine and otherpresynaptic dopaminergic markers in human striatum. J Neurochem2003;87:574e85.
[14] Kish SJ, Shannak K, Rajput A, Deck JH, Hornykiewicz O. Aging producesa specific pattern of striatal dopamine loss: implications for the etiology ofidiopathic Parkinson’s disease. J Neurochem 1992;58:642e8.
[15] Lloyd KG, Hornykiewicz O. Occurrence and distribution of aromatic L-aminoacid (L-DOPA) decarboxylase in the human brain. J Neurochem 1972;19:1549e59.
[16] Harada N, Nishiyama S, Satoh K, Fukumoto D, Kakiuchi T, Tsukada H. Age-related changes in the striatal dopaminergic system in the living brain:a multiparametric PET study in conscious monkeys. Synapse 2002;45:38e45.
[17] Suzuki M, Hatano K, Sakiyama Y, Kawasumi Y, Kato T, Ito K. Age-relatedchanges of dopamine D1-like and D2-like receptor binding in the F344/N ratstriatum revealed by positron emission tomography and in vitro receptorautoradiography. Synapse 2001;41:285e93.
[18] Wang Y, Chan GL, Holden JE, Dobko T, Mak E, Schulzer M, et al. Age-dependent decline of dopamine D1 receptors in human brain: a PET study.Synapse 1998;30:56e61.
[19] Giorgi O, De Montis G, Porceddu ML, Mele S, Calderini G, Toffano G, et al.Developmental and age-related changes in D1-dopamine receptors anddopamine content in the rat striatum. Brain Res 1987;432:283e90.
[20] Stark AK, Pakkenberg B. Histological changes of the dopaminergic nigros-triatal system in aging. Cell Tissue Res 2004;318:81e92.
[21] Ota M, Yasuno F, Ito H, Seki C, Nozaki S, Asada T, et al. Age-related decline ofdopamine synthesis in the living human brain measured by positron emis-sion tomography with L-[beta-11C]DOPA. Life Sci 2006;79:730e6.
[22] Ishida Y, Okawa Y, Ito S, Shirokawa T, Isobe K. Age-dependent changes indopaminergic projections from the substantia nigra pars compacta to theneostriatum. Neurosci Lett 2007;418:257e61.
[23] McCormack AL, Di Monte DA, Delfani K, Irwin I, DeLanney LE, Langston WJ,et al. Aging of the nigrostriatal system in the squirrel monkey. J Comp Neurol2004;471:387e95.
[24] Colebrooke RE, Humby T, Lynch PJ, McGowan DP, Xia J, Emson PC. Age-related decline in striatal dopamine content and motor performance occursin the absence of nigral cell loss in a genetic mouse model of Parkinson’sdisease. Eur J Neurosci 2006;24:2622e30.
[25] Fearnley JM, Lees AJ. Ageing and Parkinson’s disease: substantia nigraregional selectivity. Brain 1991;114(Pt 5):2283e301.
[26] Emborg ME, Ma SY, Mufson EJ, Levey AI, Taylor MD, Brown WD, et al. Age-related declines in nigral neuronal function correlate with motor impair-ments in rhesus monkeys. J Comp Neurol 1998;401:253e65.
[27] Barili P, De Carolis G, Zaccheo D, Amenta F. Sensitivity to ageing of the limbicdopaminergic system: a review. Mech Ageing Dev 1998;106:57e92.
[28] Reeves S, Bench C, Howard R. Ageing and the nigrostriatal dopaminergicsystem. Int J Geriatr Psychiatry 2002;17:359e70.
[29] Smith CD, Chebrolu H, Wekstein DR, Schmitt FA, Markesbery WR. Age andgender effects on human brain anatomy: a voxel-based morphometric studyin healthy elderly. Neurobiol Aging 2007;28:1075e87.
[30] Gunning-Dixon FM, Head D, McQuain J, Acker JD, Raz N. Differential aging ofthe human striatum: a prospective MR imaging study. AJNR Am J Neuro-radiol 1998;19:1501e7.
[31] Good CD, Johnsrude IS, Ashburner J, Henson RN, Friston KJ, Frackowiak RS.A voxel-based morphometric study of ageing in 465 normal adult humanbrains. Neuroimage 2001;14:21e36.
[32] Raz N, Gunning-Dixon FM, Head D, Dupuis JH, Acker JD. Neuroanatomicalcorrelates of cognitive aging: evidence from structural magnetic resonanceimaging. Neuropsychology 1998;12:95e114.
[33] Pakkenberg H, Andersen BB, Burns RS, Pakkenberg B. A stereological study ofsubstantia nigra in youngandold rhesusmonkeys. BrainRes 1995;693:201e6.
[34] Kubis N, Faucheux BA, Ransmayr G, Damier P, Duyckaerts C, Henin D, et al.Preservation of midbrain catecholaminergic neurons in very old humansubjects. Brain 2000;123:366e73.
[35] McNeill TH, Koek LL. Differential effects of advancing age on neurotrans-mitter cell loss in the substantia nigra and striatum of C57BL/6N mice. BrainRes 1990;521:107e17.
[36] Emerich DF, McDermott P, Krueger P, Banks M, Zhao J, Marszalkowski J, et al.Locomotion of aged rats: relationship to neurochemical but not morpho-logical changes in nigrostriatal dopaminergic neurons. Brain Res Bull 1993;32:477e86.
[37] Irwin I, DeLanney LE, McNeill T, Chan P, Forno LS, Murphy Jr GM, et al. Agingand the nigrostriatal dopamine system: a non-human primate study. Neu-rodegeneration 1994;3:251e65.
[38] Salvatore MF, Apparsundaram S, Gerhardt GA. Decreased plasma membraneexpression of striatal dopamine transporter in aging. Neurobiol Aging 2003;24:1147e54.
[39] McGeer PL, McGeer EG, Suzuki JS. Aging and extrapyramidal function. ArchNeurol 1977;34:33e5.
[40] Mann DM, Yates PO. The effects of ageing on the pigmented nerve cells of thehuman locus caeruleous and substantia nigra. ActaNeuropathol 1979;47:93e7.

O. Darbin / Parkinsonism and Related Disorders 18 (2012) 426e432 431
[41] Thiessen B, Rajput AH, Laverty W, Desai H. Age, environments, and thenumber of substantia nigra neurons. Adv Neurol 1990;53:201e6.
[42] Ma SY, Roytt M, Collan Y, Rinne JO. Unbiased morphometrical measurementsshow loss of pigmented nigral neurones with ageing. Neuropathol ApplNeurobiol 1999;25:394e9.
[43] Siddiqi Z, Kemper TL, Killiany R. Age-related neuronal loss from the sub-stantia nigra-pars compacta and ventral tegmental area of the rhesusmonkey. J Neuropathol Exp Neurol 1999;58:959e71.
[44] Cabello CR, Thune JJ, Pakkenberg H, Pakkenberg B. Ageing of substantia nigrain humans: cell loss may be compensated by hypertrophy. Neuropathol ApplNeurobiol 2002;28:283e91.
[45] Chu Y, Kompoliti K, Cochran EJ, Mufson EJ, Kordower JH. Age-relateddecreases in Nurr1 immunoreactivity in the human substantia nigra. J CompNeurol 2002;450:203e14.
[46] Vaillancourt DE, Spraker MB, Prodoehl J, Zhou XJ, Little DM. Effects of agingon the ventral and dorsal substantia nigra using diffusion tensor imaging.Neurobiol Aging; 2010.
[47] Bernheimer H, Birkmayer W, Hornykiewicz O, Jellinger K, Seitelberger F.Brain dopamine and the syndromes of Parkinson and Huntington. J NeurolSci 1973;20:415e55.
[48] Miguez JM, Aldegunde M, Paz-Valinas L, Recio J, Sanchez-Barcelo E. Selectivechanges in the contents of noradrenaline, dopamine and serotonin in ratbrain areas during aging. J Neural Transm 1999;106:1089e98.
[49] Udenfriend S. Tyrosine hydroxylase. Pharmacol Rev 1966;18:43e51.[50] Yurek DM, Hipkens SB, Hebert MA, Gash DM, Gerhardt GA. Age-related
decline in striatal dopamine release and motoric function in brown Norway/Fischer 344 hybrid rats. Brain Res 1998;791:246e56.
[51] Gerhardt GA, Cass WA, Yi A, Zhang Z, Gash DM. Changes in somatodendriticbut not terminal dopamine regulation in aged rhesus monkeys. J Neurochem2002;80:168e77.
[52] Hurley PJ, Elsworth JD, Whittaker MC, Roth RH, Redmond Jr DE. Agedmonkeys as a partial model for Parkinson’s disease. Pharmacol BiochemBehav 2011;99:324e32.
[53] Gozlan H, Daval G, Verge D, Spampinato U, Fattaccini CM, Gallissot MC, et al.Aging associated changes in serotoninergic and dopaminergic pre- andpostsynaptic neurochemical markers in the rat brain. Neurobiol Aging 1990;11:437e49.
[54] Dluzen DE, McDermott JL, Ramirez VD. Changes in dopamine release in vitrofrom the corpus striatum of young versus aged rats as a function of infusionmodes of L-dopa, potassium, and amphetamine. Exp Neurol 1991;112:153e60.
[55] Joseph JA, Dalton TK, Hunt WA. Age-related decrements in the muscarinicenhancement of Kþ-evoked release of endogenous striatal dopamine: anindicator of altered cholinergic-dopaminergic reciprocal inhibitory control insenescence. Brain Res 1988;454:140e8.
[56] Nakano M, Mizuno T. Age-related changes in the metabolism of neuro-transmitters in rat striatum: a microdialysis study. Mech Ageing Dev 1996;86:95e104.
[57] Rose GM, Gerhardt GA, Conboy GL, Hoffer BJ. Age-related alterations inmonoamine release from rat striatum: an in vivo electrochemical study.Neurobiol Aging 1986;7:77e82.
[58] Santiago M, Machado A, Cano J. Effects of age and dopamine agonists andantagonists on striatal dopamine release in the rat: an in vivo microdialysisstudy. Mech Ageing Dev 1993;67:261e7.
[59] Gerhardt GA, Cass WA, Henson M, Zhang Z, Ovadia A, Hoffer BJ, et al. Age-related changes in potassium-evoked overflow of dopamine in the striatumof the Rhesus monkey. Neurobiol Aging 1995;16:939e46.
[60] Levitt M, Spector S, Sjoerdsma A, Udenfriend S. Elucidation of the rate-limiting step in norepinephrine biosynthesis in the perfused guinea-pigheart. J Pharmacol Exp Ther 1965;148:1e8.
[61] Martin WRW, Palmer MR, Patlak CS, Calne DB. Nigrostriatal function inhumans studied with positron emission tomography. Ann Neurol 1989;26:535e42.
[62] Cordes M, Snow BJ, Cooper S, Schulzer M, Pate BD, Ruth TJ, et al. Age-dependent decline of nigrostriatal dopaminergic function: a positron emis-sion tomographic study of grandparents and their grandchildren. Ann Neurol1994;36:667e70.
[63] Sossi V, de la Fuente-Fernandez R, Schulzer M, Troiano AR, Ruth TJ, Stoessl AJ.Dopamine transporter relation to dopamine turnover in Parkinson’s disease:a positron emission tomography study. Ann Neurol 2007;62:468e74.
[64] Biju G, de la Fuente-Fernandez R. Dopaminergic function and progression ofParkinson’s disease: PET findings. Parkinsonism Relat Disord 2009;15(Suppl.4):S38e40.
[65] Hebert MA, Larson GA, Zahniser NR, Gerhardt GA. Age-related reductions in[3H]WIN 35,428 binding to the dopamine transporter in nigrostriatal andmesolimbic brain regions of the fischer 344 rat. J Pharmacol Exp Ther 1999;288:1334e9.
[66] Shimizu I, Prasad C. Relationship between [3H]mazindol binding to dopa-mine uptake sites and [3H]dopamine uptake in rat striatum during aging.J Neurochem 1991;56:575e9.
[67] Erixon-Lindroth N, Farde L, Wahlin TB, Sovago J, Halldin C, Backman L. Therole of the striatal dopamine transporter in cognitive aging. Psychiatry Res2005;138:1e12.
[68] van Dyck CH, Seibyl JP, Malison RT, Laruelle M, Zoghbi SS, Baldwin RM, et al.Age-related decline in dopamine transporters: analysis of striatal subregions,
nonlinear effects, and hemispheric asymmetries. Am J Geriatr Psychiatry2002;10:36e43.
[69] Cruz-Muros I, Afonso-Oramas D, Abreu P, Perez-Delgado MM, Rodriguez M,Gonzalez-Hernandez T. Aging effects on the dopamine transporter expres-sion and compensatory mechanisms. Neurobiol Aging; 2007.
[70] Mozley LH, Gur RC, Mozley PD, Gur RE. Striatal dopamine transporters andcognitive functioning in healthy men and women. Am J Psychiatry 2001;158:1492e9.
[71] Friedemann MN, Gerhardt GA. Regional effects of aging on dopami-nergic function in the Fischer-344 rat. Neurobiol Aging 1992;13:325e32.
[72] Hebert MA, Gerhardt GA. Age-related changes in the capacity, rate, andmodulation of dopamine uptake within the striatum and nucleus accumbensof Fischer 344 rats: an in vivo electrochemical study. J Pharmacol Exp Ther1999;288:879e87.
[73] Elsworth JD, Brittan MS, Taylor JR, Sladek Jr JR, Redmond Jr DE, Innis RB, et al.Upregulation of striatal D2 receptors in the MPTP-treated vervet monkey isreversed by grafts of fetal ventral mesencephalon: an autoradiographicstudy. Brain Res 1998;795:55e62.
[74] Morissette M, Goulet M, Calon F, Falardeau P, Blanchet PJ, Bedard PJ, et al.Changes of D1 and D2 dopamine receptor mRNA in the brains of monkeyslesioned with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine: correctionwith chronic administration of L-3,4-dihydroxyphenylalanine. Mol Phar-macol 1996;50:1073e9.
[75] Rinne JO, Laihinen A, Ruottinen H, Ruotsalainen U, Nagren K, Lehikoinen P,et al. Increased density of dopamine D2 receptors in the putamen, but not inthe caudate nucleus in early Parkinson’s disease: a PET study with [11C]raclopride. J Neurol Sci 1995;132:156e61.
[76] Gagnon C, Gomez-Mancilla B, Markstein R, Bedard PJ, Di Paolo T. Effect ofadding the D-1 agonist CY 208-243 to chronic bromocriptine treatment ofMPTP-monkeys: regional changes of brain dopamine receptors. Prog Neu-ropsychopharmacol Biol Psychiatry 1995;19:667e76.
[77] Yokochi M. Reevaluation of levodopa therapy for the treatment ofadvanced Parkinson’s disease. Parkinsonism Relat Disord 2009;15(Suppl.1):S25e30.
[78] Newman RP, LeWitt PA, Jaffe M, Calne DB, Larsen TA. Motor function in thenormal aging population: treatment with levodopa. Neurology 1985;35:571e3.
[79] Willemssen R, Falkenstein M, Schwarz M, Muller T, Beste C. Effects of aging,Parkinson’s disease, and dopaminergic medication on response selection andcontrol. Neurobiol Aging 2011;32:327e35.
[80] White NJ, Barnes TR. Senile parkinsonism, a study of current treatment. AgeAgeing 1981;10:81e6.
[81] Diederich NJ, Moore CG, Leurgans SE, Chmura TA, Goetz CG. Parkinsondisease with old-age onset: a comparative study with subjects with middle-age onset. Arch Neurol 2003;60:529e33.
[82] Frank C, Pari G, Rossiter JP. Approach to diagnosis of Parkinson disease. CanFam Physician 2006;52:862e8.
[83] Zhang Z, Andersen A, Grondin R, Barber T, Avison R, Gerhardt G, et al.Pharmacological MRI mapping of age-associated changes in basal gangliacircuitry of awake Rhesus monkeys. NeuroImage 2001;14:1159e67.
[84] Ingram DK, Ikari H, Umegaki H, Chernak JM, Roth GS. Application of genetherapy to treat age-related loss of dopamine D2 receptor. Exp Gerontol1998;33:793e804.
[85] Gerfen CR, Surmeier DJ. Modulation of striatal projection systems by dopa-mine. Annu Rev Neurosci; 2010.
[86] Galvan A, Wichmann T. Pathophysiology of parkinsonism. Clin Neurophysiol2008;119:1459e74.
[87] Nambu A. A new dynamic model of the cortico-basal ganglia loop. Prog BrainRes 2004;143:461e6.
[88] Obeso JA, Marin C, Rodriguez-Oroz C, Blesa J, Benitez-Temino B, Mena-Segovia J, et al. The basal ganglia in Parkinson’s disease: current concepts andunexplained observations. Ann Neurol 2008;64(Suppl. 2):S30e46.
[89] Tang JK, Moro E, Mahant N, Hutchison WD, Lang AE, Lozano AM, et al.Neuronal firing rates and patterns in the globus pallidus internus of patientswith cervical dystonia differ from those with Parkinson’s disease.J Neurophysiol 2007;98:720e9.
[90] Ward NS, Frackowiak RS. Age-related changes in the neural correlates ofmotor performance. Brain 2003;126:873e88.
[91] Mattay VS, Fera F, Tessitore A, Hariri AR, Das S, Callicott JH, et al. Neuro-physiological correlates of age-related changes in human motor function.Neurology 2002;58:630e5.
[92] Calautti C, Serrati C, Baron JC. Effects of age on brain activation duringauditory-cued thumb-to-index opposition: a positron emission tomographystudy. Stroke 2001;32:139e46.
[93] Wu T, Zang Y, Wang L, Long X, Li K, Chan P. Normal aging decreases regionalhomogeneity of the motor areas in the resting state. Neurosci Lett 2007;423:189e93.
[94] Ohashi S, Mori A, Kurihara N, Mitsumoto Y, Nakai M. Age-related severity ofdopaminergic neurodegeneration to MPTP neurotoxicity causes motordysfunction in C57BL/6 mice. Neurosci Lett 2006;401:183e7.
[95] Collier TJ, Lipton J, Daley BF, Palfi S, Chu Y, Sortwell C, et al. Aging-relatedchanges in the nigrostriatal dopamine system and the response to MPTP innonhuman primates: diminished compensatory mechanisms as a prelude toparkinsonism. Neurobiol Dis 2007;26:56e65.

O. Darbin / Parkinsonism and Related Disorders 18 (2012) 426e432432
[96] Adler CH, Hentz JG, Joyce JN, Beach T, Caviness JN. Motor impairment innormal aging, clinically possible Parkinson’s disease, and clinically probableParkinson’s disease: longitudinal evaluation of a cohort of prospective braindonors. Parkinsonism Relat Disord 2002;9:103e10.
[97] Markesbery WR, Jicha GA, Liu H, Schmitt FA. Lewy body pathology in normalelderly subjects. J Neuropathol Exp Neurol 2009;68:816e22.
[98] Kostic V, Przedborski S, Flaster E, Sternic N. Early development of levodopa-induced dyskinesias and response fluctuations in young-onset Parkinson’sdisease. Neurology 1991;41:202e5.
[99] Sossi V, de la Fuente-Fernandez R, Schulzer M, Adams J, Stoessl AJ. Age-related differences in levodopa dynamics in Parkinson’s: implications formotor complications. Brain 2006;129:1050e8.
[100] Nagayama H, Ueda M, Kumagai T, Tsukamoto K, Nishiyama Y, Nishimura S,et al. Influence of ageing on the pharmacokinetics of levodopa in elderlypatients with Parkinson’s disease. Parkinsonism Relat Disord 2011;17:150e2.
[101] Emborg ME, Moirano J, Raschke J, Bondarenko V, Zufferey R, Peng S, et al.Response of aged parkinsonian monkeys to in vivo gene transfer of GDNF.Neurobiol Dis 2009;36:303e11.