dopamine-mediated regulation of striatal neuronal and network interactions
TRANSCRIPT

REVIEW
Dopamine-mediated regulation of striatal neuronal and network interactions S h a o - P i i O n n , A n t h o n y R. W e s t a n d A n t h o n y A. G r a c e
The dopaminergic system exerts dynamic modulation of glutamatergic afferent drive that is dependent on the temporal pattern of the dopaminergic input and the subtypes of striatal neurons affected. The differences in feedforward inhibition between striatal neurons comprising the direct and indirect output
pathway confer distinct response-pattern differences in their respective targets, supporting brief bursts
of activity inType-I neurons but attenuating repetitive activity inType-II cells. This temporal patterning is further modulated by NO-mediated signaling, and by tonic and phasic dopamine-mediated stimulation, which exerts preferential actions on indirect and direct output neurons, respectively.
As a result, the striatal network is forced into state-dependent patterns of activity that differentially regulate muscle tone and voluntary motor activity via distinct output projections from the striatum. Trends Neurosci. (2000) 23 (Suppl.), $48-$56
Shao-Pii Onn is at
the Dept of Neumbiology and
Anatomy, MCP Hahnemann
University, Philadelphia,
PA 19129, USA and
Anthony R. West and
Anthony A. Grace are at the Depts of Neuroscience and
Psychiatry, University of
Pittsburgh, Pittsburgh,
PA 15260, USA.
T HE STRIATUM is considered to be the center of sensorimotor integration within the basal ganglia. It
receives widespread excitatory input from all regions of cortex that converge with extensive dopaminergic afferents from the midbrain. The primary recipients of glutamatergic-dopaminergic inputs are the spiny neurons, the predominant cell type in the striatum. Evidence indicates that two distinct output pathways originate from two separate populations of striatal spiny neurons, and they appear to act in a functionally oppo- site manner to integrate sensorimotor information and regulate the basal-ganglia output nuclei 1. In rodents, one population of spiny neurons projects directly to basal- ganglia output neurons within the entopeduncular nucleus (EPN) and the substantia nigra zona reticulata (SNr). The other pathway is termed the indirect path- way and projects via the globus pallidus (GP) and sub- thalamus (STN) to influence these basal-ganglia output regions. This article focuses on recent findings concern- ing dopamine (DA)-mediated modulation of the activ- ities of single striatal cells and their network interactions. A disruption of the balance in tonic and phasic DA release 2 in the striatum in neurological diseases such as Parkinson's disease (PD) might alter striatal network activities to create an imbalance of information out- flow between the direct and indirect output pathways, leading to abnormal motor control.
Regulation of striatal neurons by dopamine and glutamate
The physiological action of DA is historically thought to be inhibitory in the striatum, based on its effects on the firing frequency of dopamine-receptive neurons. Thus, DA applied iontophoretically in vivo or bath- applied DA in vitro predominantly decrease sponta- neous or intracellular current-elicited firing frequency of striatal spiny neurons 3-8. This inhibition is presum- ably mediated postsynaptically and occurs indepen- dently of membrane depolarization or hyperpolariz- ation by D2- and Dl-receptor agonists, respectively 7'9. In addition, DA receptors exert a regulatory influence over glutamate-mediated ' tone' in the striatum. Thus,
activation of D2 receptors on cortical afferent termi- nals l°'n reduces cortically elicited EPSPs recorded in spiny neurons 4,s,12. This presynaptic role of D2 receptors in modulating cortical glutamate-mediated transmission in striatum is observed in approximately 60% of cells in both the dorsal and ventral striatum s (S-P. Onn, A.R. West and A.A. Grace, unpublished observations). This is consistent with the anatomical localization of D2-receptor immunoreactivity to some but not all terminals of cortical o r i g i n 1°'~1. Despite the fact that Dl-receptor-immunoreactive axons are extremely rare in the striatum m'~,14, some studies suggest that D1 receptors also affect glutamate-mediated EPSPs elicited by intrastriatal stimulation ls-17. However, intrastriatal stimulation activates both extrinsic (glutamatergic fibers from cortex and thalamus) and intrinsic (GABAergic and cholinergic) afferents; thus, it is unclear whether the response elicited is indeed dependent on cortical affer- ents. There is also evidence that Dl-receptor agonists can attenuate glutamate-mediated transmission indirectly via the release of adenosine from postsynaptic neurons, resulting in trans-synaptic feedback suppression of glutamate-mediated transmission ~s.
Recent in vitro studies using whole-cell recordings have provided additional data regarding the modulatory role of DA in the regulation of glutamate-mediated trans- mission. DA exerts a bimodal influence on glutamate- mediated synaptic transmission, in that the low concen- trations of DA that preferentially activate D1 receptors cause an increase in NMDA-receptor-mediated currents, whereas high concentrations of DA that activate D2 receptors suppress AMP~-receptor-mediated currents 18,~9. This is consistent with extracellular studies using ionto- phoretic application of DA in anesthetized 2° and awake 2~ rats, where low versus high ejection currents will enhance or suppress glutamate-mediated transmission, respectively. Such Dl-receptor-mediated facilitation of NMDA currents appears to involve cAMP, protein kinase A and dopamine-and-cAMP-regulated phospho- protein (DARPP)-32 signal-transduction cascades 22-24. This permissive role of D1 receptors on NMDA-receptor- mediated synaptic function is further reflected in the
$ 4 8 "FINS Vol. 23, No. 10 (Suppl.), 2000 0166-2236/00/$ - see front matter © 2000 Elsevier Science Ltd. All rights reserved. PIh 51471-1931(00)00020-3

S-P. Onn et al. - Regulation of striatal neuronal and network interactions RE- -V~W
(a) (b)
_ _ ®
DA
Q
Glutamate
(c)
DA
Basal ganglia, Parkinson's disease and levodopa therapy: TINS supplement
Fig. 1. The tonic-phasic model of dopamine system regulation can account for a number of phenomena related to dopamine (DA)-mediated modulation of motor control within the striatum. (a) Phasic stimulation. DAergic terminals in the striatum (1) synapse primarily on the side or the neck of dendritic spines (2) that arise from striatal medium spiny neurons. Action potentials generated by dopaminergic neuron discharges arrive into the axon terminal within the striatum (1) to cause spike-dependent DA release (red circles) into the synaptic deft, where it stimulates postsynaptic DA receptors (squares). The DA released in this manner is high in amplitude [up to 1 mM (Ref. 29)] but very brief in duration, in that it is rapidly removed from the synaptic cleft by the DA transporter and accumulated in the presynaptic terminal before it can escape the synaptic cleft. This high-amplitude but brief-duration DA release is termed the phasic DA input, and is proposed to mediate behaviorally relevant events. (b) Tonic stimulation. Tonic DA release is proposed to be mediated by at least two events. Action potentials arising from glutamatergic afferents (1) cause the release of glutamate (blue circles) into the synaptic cleft (2) at the spine-head of the medium spiny neuron. The glutamate is of sufficient concentration to diffuse from the synaptic cleft and stimulate [either directly via presynaptic glutamate receptors (3) or indirectly via NO] the dopaminergic terminal to release DA. These presynaptic signals might trigger the release of DA from the terminal at sites distal to the synaptic cleft (4). Studies have shown that the DA that escapes reuptake (5) from the synaptic cleft after sustained spike actMty also contributes to this tonic DA pool (6). This pool is constantly present in the extrasynaptic space, and is tightly regulated by multiple feedback systems. Tonic extrasynaptic DA levels are very much lower in concentration (that is, tens of nanomolar) compared with those found in the phasic DA response, and thus are not capable of effectively stimulating intrasynaptic DA receptors (squares). Tonic DA is also in dynamic equilibrium with glutamatergic afferents, as studies have shown that such levels of DA are sufficient to provide baseline stimulation of presynaptic D2 receptors (squares) on corticostriatal afferents s (7), thereby limiting glutamate-stimulated DA release as well. (c) Tonic DA will presynaptically inhibit spike-dependent phasic DA release. Although the tonic levels of DA present in the extrasynaptic space (1) are very low in concentration, they are nonetheless sufficient to provide constant baseline stimulation of presynaptic D2 receptors. In particular, they provide stimulation of the synthesis- and release-modulating autoreceptors present on the dopaminergic terminal (squares). In this way, tonic DA provides a feedback inhibition of spike-dependent DA release (2), resulting in a lower concentration of DA released phasically in response to action potentials in the DA terminal (3). Adapted, with permission, from Ref. 2.
expression of immediate-early genes (lEGs) such as l o s in the Dl-receptor-substance P-containing spiny neurons. Expression of IEGs is known to be contingent upon co-activation of both D1 and NMDA receptors 2s,z6.
It is known that Dl-receptor or D2-receptor agonists, applied alone or simultaneously, typically produce a reduction in membrane excitability of striatal spiny n e u r o n s 3,7-9,z7. However, it appears that the D 1-receptor- agonist-induced decrease in excitability can be reversed to a facilitation of spiking upon subsequent D2-receptor- agonist administration 9. Thus, the response produced by combined D1- and D2-receptor-agonist stimulation is dependent on the temporal sequence of agonist administration. This type of Dl-receptor-dependent modulation of D2-receptor-mediated excitation is absent in slices taken from the DARPP-32 knockout mice, which suggests an involvement of a G-protein-mediated response via the cAMP-protein-kinase cascade 23,zs. One explanation for this observation might be that D2-receptor activation can reverse the phosphoryl- ation of DARPP-32 proteins caused by Dl-receptor activation, which decreases the inhibitory influence of this signaling pathway on spiny-neuron activity.
This temporal dependence of D1- and D2-receptor activation could have significant functional implications when interpreted in the context of the tonic-phasic model of dopaminergic-system regulation (Fig. 1). Thus, during sustained activity such as the burst firing of dopaminergic neurons as reward-associated stimuli 3° or high-frequency stimulation of dopaminergic fibers 3~, the resultant massive phasic DA release 2 would be expected
to stimulate both D1 and D2 receptors located within the synapse. DA would also diffuse from the synapse to stimulate the extrasynaptic D1 receptors 1°'11'I3'14'3z in the form of tonic DA stimulation 2. Such simultaneous D1- and D2-receptor activation should produce addi- tive inhibitory effects. If the activity is maintained for an extended period of time, the extrasynaptic D1 receptors would continue to be tonically stimulated. As a consequence, any subsequent phasic activation of dopaminergic-cell f r ing would lead to an attenuated DA-mediated inhibitory action on D2 receptors, which are known to be more concentrated within the synapse 1°'11. Therefore, the system appears to be designed to attenuate phasic DA-mediated responses under con- ditions in which the dopaminergic system is being tonically overdriven. Thus, the phasic DA-mediated response would be strongly dependent on the history of activity that occurs within specific striatal sub- regions. Indeed, such an arrangement would selec- tively regulate the aforementioned modulation of glutamate-mediated transmission by DA that occurs within 'hot spots' of high DA-mediated transmission within the striatum. By contrast, a lack of such tempo- ral dynamics of D1- and D2-receptor activation in sur- rounding modules (cold spots; owing to a lack of sustained DA-mediated activity) would lead to a gen- eralized reduction of membrane excitability in these neurons. This decrease in neuronal activities outside the active module or modules might be important for enhancing the signal-noise ratio zl within parallel striatal circuits.
TINS Vol. 23, No. 10 (Suppl.), 2000 S49

R g g ~ f ~ s-P. O n n e t a L - Regulation of striatal neuronal and network interactions
Electrophysiological identification of striatal cell subtypes
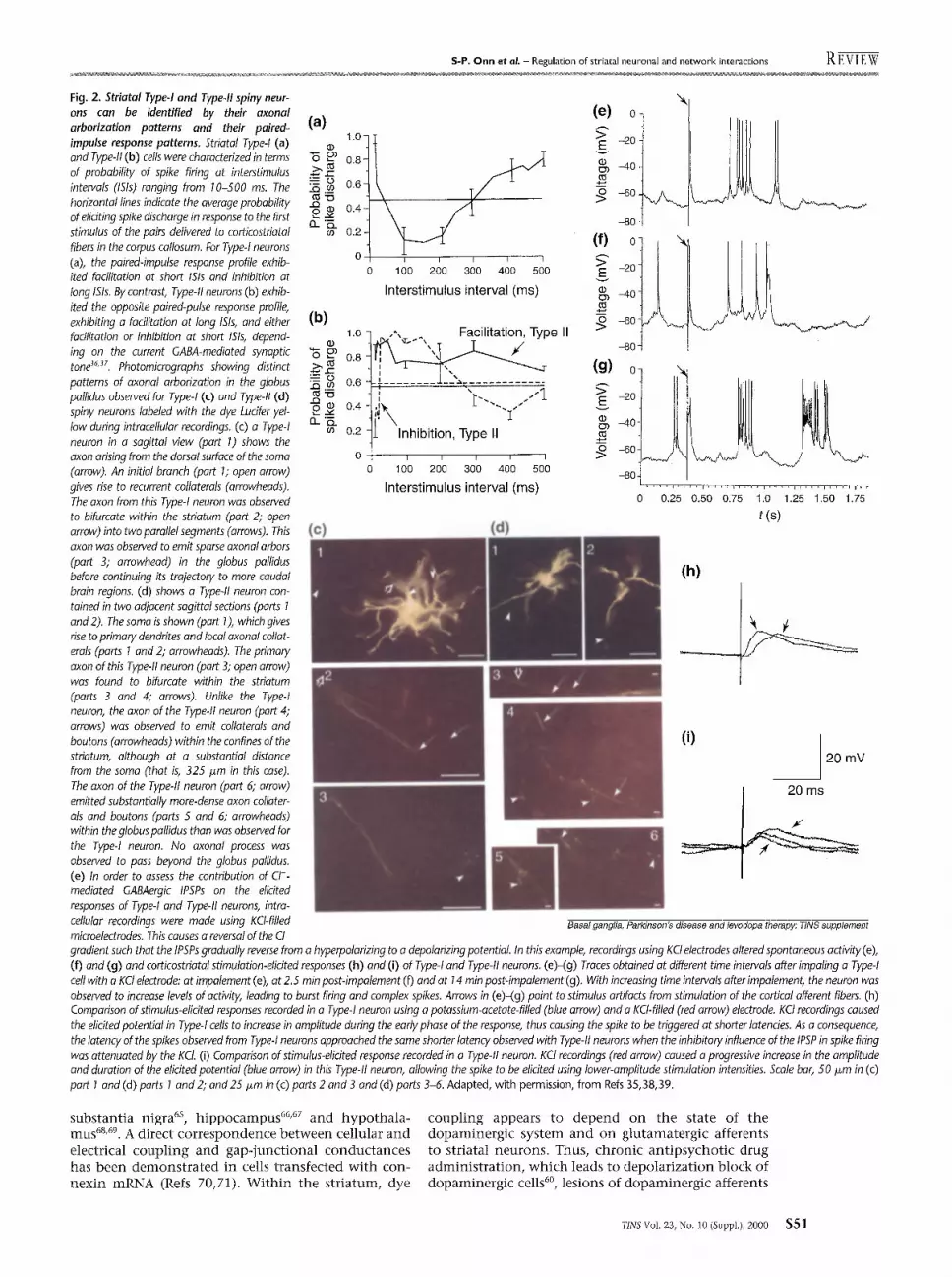
One impor tant determinant in defining the actions of DA in the striatum is whether the cell type involved comprises the proiection neuron of the direct or the indirect output pathway. Al though the ceils of origin of these two pathways have not been clearly identified, studies have defined two classes of striatal neurons based on functional characteristics that correlate with the properties predicted for these distinct output sys- tems. The first electrophysiological study suggesting that two striatal cell types are present was based primarily on differences in their extracellular action, potential waveform and response to chronic haloperidol admin- istration s3. However, as extracellular waveform is no t a reliable measure for cell identification, owing to its dependence on electrode characteristics s4, a more- reproducible me thod was developed for defining these two cell types based on their response patterns to paired- pulse st imulation of corticostriatal afferents s4. Thus, in response to presentat ion of two stimuli delivered at threshold levels for eliciting spike discharge, the prob- ability that the second impulse would elicit a spike was found to be dependent on the interstimulus interval (ISI) 34'35 (Fig. 2). Two distinct response patterns were recorded from striatal cells: the Type-I cells showed facilitation of spike discharge in response to the second pulse when the interpulse interval was 10 ms, but a decreased probabili ty of spiking when the interval was 100 ms; the Type-II cells showed the opposite pattern, exhibiting inhibi t ion at 10 ms 1Sis and facilitation at 100 ms ISis. In addition, the latency of the cortically elicited spike observed in Type-I neurons was signifi- cantly longer than that of the Type-II neurons. Further studies confirmed that the two cell types distinguished by this method corresponded to the same two groups of neurons previously identified by extracellular waveform analysis 34, with Type-I neurons exhibiting monophas ic spikes and a facil i tat ion-inhibit ion response pattern, and Type-II neurons having biphasic spikes and an inhibi t ion-faci l i tat ion response pattern with a greater sensitivity to DA-mediated inhibi t ion a3.
Type-I and Type-II striatal neurons
There are a number of properties of the Type-I and Type-II neurons that suggest they might be the cells of origin of the direct and indirect output projections, respectively: Type-I neurons are preferentially sensitive to D 1-receptor agonists 3~, whereas the Type-II neurons are potent ly inhibited by D2-receptor agonists a3,39. Further- more, a l though activity of bo th Type-I and Type-II neurons increases following lesions of dopaminergic neurons, only the firing rate of Type-II neurons returns to control levels upon behavioral recovery from gross motor deficits 4°,41. This is consistent with electrophysio- logical and metabolic studies, as lesions that are suffi- cient to induce profound motor disruption also alter the firing pattern 4z-44 and metabolic activity 4s'46 of neurons in the GP-STN pathway. In addition, a correspondence between Type-II neurons and the striatopallidal projec- t ion is supported by correlated anatomical data f rom intracellular staining of electrophysiologically charac- terized neurons, showing that axons of Type-II neurons terminate exclusively in the GP whereas axons of Type-I neurons project beyond this structure s6. This evidence thus provides substantial support for a model in which Type-I neurons are the Dl-receptor-sensitive
cells conta ining GABA and substance P or dynorphin . These neurons project to the EPN and SNr, whereas the Type-II neurons might be the GABA- and enkephalin- conta in ing neurons that are preferentially sensitive to D2-receptor agonists and project to the GP (Fig. 2).
Type-I and Type-II cells appear to be med ium spiny neurons with similar electrophysiological properties that differ in their projection sites and elicited response profile. These differences in spike latencies and paired- impulse response profiles appear to be dependent on t iming of their GABAergic inhibition at short intervals: a GABA-mediated conductance change occurs simul- taneously with the EPSPs in Type-I cells, which leads to a delay in triggering the elicited spike, whereas a later-developing GABA-mediated conductance change in Type-II cells results in an inhibi t ion of spiking at short intervals aT. The increase in st imulat ion strength required to activate Type-II cells in this case contributes to the longqatency facilitation in this cell type. Studies have described two populat ions of striatal cells that differ with respect to the distribution of inhibi tory or parvalbumin- immunoreact ive 47 synapses on their proximal dendritic segments. Indeed, such a differential distribution of GABAergic inputs could account for the latency difference in IPSPs that distinguish the paired- pulse response pattern of these cell types. These GABAergic synapses are likely to arise f rom striatal GABAergic interneurons, rather than fi'om collaterals of the projection neurons 48, as studies show that (1) the response patterns of the two cell types can occur in iso- lation, thereby precluding an interdependence in the paired-pulse response profile; (2) the onset of the IPSP is too rapid to be triggered by spiking in neighboring Type-I or Type-II cells (in fact, it often is triggered before the spike), and therefore must arise f rom a third GABAergic neuron class to mediate this response; and (3) a short- axon parvalbumin- immunoreact ive GABAergic inter- neuron has been observed in the striatum of rats 49's° that directly inhibits med ium spiny neurons.
Neuron synchronization and cellular coupling: DARPP-32 and NO
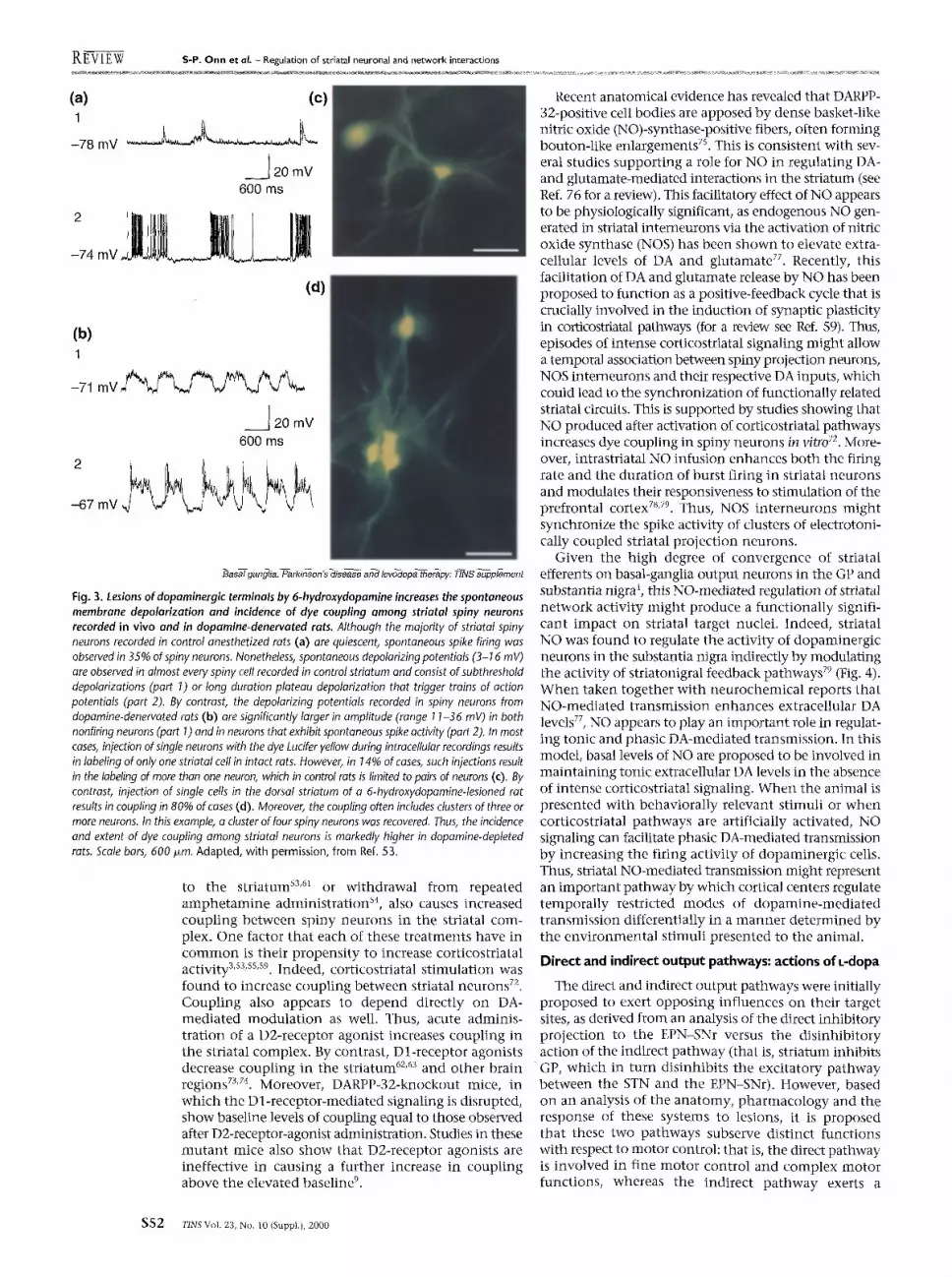
W h e n recorded intracellularly in vivo, spiny neurons often exhibit a bistable activity pattern, consisting of long-duration depolarized membrane plateau potentials occurring at -1 Hz (Refs 51-57), al ternating with a hyperpolarized membrane potential. These slow mem- brane oscillations allow the cell membrane to shift between two membrane potentials (bistable neurons) and are believed to reflect the state of glutamate-mediated tone from cortical afferents s'ss's4's6'sS. The level of bistable activity is likely to be higher in the awake rat, given that the activity level of cortical neurons is depressed in the anesthetized rat, and bistable membrane activity is known to be sensitive to the levels of anesthesia used during recordings sl'szs6. Moreover, treatments that appear to cause an increase in corticostriatal glutamate- mediated tone, such as 6-hydroxydopamine- induced lesions sa and chronic antipsychotic drug t reatment s9,6° also cause an increase in the bistable state within striatal neurons, as well as an increase in cellular coupling (Fig. 3). Over the past decade, studies have shown that different manipulat ions of dopaminergic systems can modulate cellular coupling in a regionally selective manner . Cellular coupling is a morphological index of electrical coupling and has been demonstrated in several brain regions, including the striatum ss,61-63, septum 64,
$ 5 0 T I N S Vol. 23, No. 10 (Suppl.), 2000
S-P. Onn et aL - Regulation of striatal neuronal and network interactions R ~-v] E W
Fig. 2. Striatal Type-I and Type-II spiny neur- ons can be identified by their axonal arborization patterns and their paired- impulse response patterns. Striatal Type-I (a) and Type-II (b) cells were characterized in terms of probability of spike firing at interstimulus intervals (ISis) ranging from 10-500 ms. The horizontal lines indicate the average probability of eliciting spike discharge in response to the first stimulus of the pairs delivered to corticostriatal fibers in the corpus callosum. For Type-I neurons (a), the paired-impulse response profile exhib- ited facilitation at short ISis and inhibition at long ISis. By contrast, Type-II neurons (b) exhib- ited the opposite paired-pulse response profile, exhibiting a facilitation at long ISis, and either facilitation or inhibition at short ISis, depend- ing on the current GABA-mediated synaptic tone 36y. Photomicrographs showing distinct patterns of axonal arborization in the globus pallidus observed for Type-I (c) and Type-II (d) spiny neurons labeled with the dye Lucifer yel- low during intracellular recordings. (c) a Type-I neuron in a sagittal view (part 1) shows the axon arising from the dorsal surface of the soma (arrow). An initial branch (part 1; open arrow) gives rise to recurrent collaterals (arrowheads). The axon from this Type-I neuron was observed to bifurcate within the striatum (part 2; open arrow) into two parallel segments (arrows). This axon was observed to emit sparse axonal arbors (part 3; arrowhead) in the globus pallidus before continuing its trajectory to more caudal brain regions. (d) shows a Type-II neuron con- tained in two adjacent sagittal sections (parts 1 and 2). The soma is shown (part 1), which gives rise to primary dendrites and local axonal collat- erals (parts 1 and 2; arrowheads). The primary axon of this Type-II neuron (part 3; open arrow) was found to bifurcate within the striatum (parts 3 and 4; arrows). Unlike the Type-I neuron, the axon of the Type-II neuron (part 4; arrows) was observed to emit collaterals and boutons (arrowheads) within the confines of the striatum, although at a substantial distance from the soma (that is, 325 i~m in this case). The axon of the Type-II neuron (part 6; arrow) emitted substantially more-dense axon collater- als and boutons (parts 5 and 6; arrowheads) within the globus pallidus than was observed for the Type-I neuron. No axonal process was observed to pass beyond the globus pallidus. (e) In order to assess the contribution of CI - mediated GABAergic IPSPs on the elicited responses of Type-I and Type-II neurons, intra- cellular recordings were made using KCI-filled microelectrodes. This causes a reversal of the CI
(a) (e)~ o-. 1.o- > g g -20-
~ 0 0.6- :-~ ,~ -60 .
0.4- '- -80- a. "~_ 0.2- (f) 0 ~
0 I t i i I ~ " 0 100 200 300 400 500 F: -20"
Interstimulus interval (ms) ~ -4o- cd
(b) ~ -60- 1.o- ,*, . Facilitation, Type II
. -7I ~ -80- ,-g o 8 - ~
0 ~ 0.4 I X E -20- if- • 5_ " I 0") -40-
0.2 Inhibition, Type II ~ 0 ~ i ~ -60-
0 10'0 20; 30; 400 500 -80- Interst imulus interval (ms)
(c) (d)
i
~ , , , , J . . . . i . . . . , . . . . J . . . . i . . . . . i . . . . J , ,
0,25 0.50 0.75 1.0 1.25 1.50 1.75
t (s)
4 ~.
? i~ i i ? ?;̧ 5̧ /i;il i i;i; ; ; ; ~i@i; ;̧ ̧ ¸̧
3
(h)
(i) 20 mV
Basal ganglia, Parkinson's disease and levodopa therapy: TINS supplement
gradient such that the IPSPs gradually reverse from a hyperpolarizing to a depolarizing potentiaL In this example, recordings using KCI electrodes altered spontaneous actMty (e), (f) and (g) and corticostriatal stimulation-elicited responses (h) and (i) of Type-I and Type-II neurons. (e)-(g) Traces obtained at different time intervals after impaling a Type-I cell with a KCI electrode: at impalement (e), at 2.5 min post-impalement (f) and at 14 rain post-impalement (g). With increasing time intervals after impalement, the neuron was observed to increase levels of actMty, leading to burst firing and complex spikes. Arrows in (e)-(g) point to stimulus artifacts from stimulation of the corticai afferent fibers. (h) Comparison of stimulus-elicited responses recorded in a Type-I neuron using a potassium-acetate-filled (blue arrow) and a KCl-filled (red arrow) electrode. KCI recordings caused the elicited potential in Type-I cells to increase in amplitude during the early phase of the response, thus causing the spike to be triggered at shorter latencies. As a consequence, the latency of the spikes observed from Type-I neurons approached the same shorter latency observed with Type-II neurons when the inhibitory influence of the IPSP in spike firing was attenuated by the KCI. (i) Comparison of stimulus-elicited response recorded in a Typeql neuron. KCI recordings (red arrow) caused a progressive increase in the amplitude and duration of the elicited potential (blue arrow) in this Type-II neuron, allowing the spike to be elicited using lower-amplitude stimulation intensities. Scale bar, 50 ixm in (c) part I and (d) parts 1 and 2; and 25 ffm in (c) parts 2 and 3 and (d) parts 3-6. Adapted, with permission, from Refs 35,38,39.
substantia nigra 6s, hippocampus 66'67 and hypothala- mus 6a'69. A direct correspondence between cellular and electrical coupling and gap-junctional conductances has been demonstrated in cells transfected with con- nexin mRNA (Refs 70,71). Within the striatum, dye
coupling appears to depend on the state of the dopaminergic system and on glutamatergic afferents to striatal neurons. Thus, chronic antipsychotic drug administration, which leads to depolarization block of dopaminergic cells 6°, lesions of dopaminergic afferents
TINS Vol. 23, No. 10 (Suppl.), 2000 $ 5 1
REVigW S-P. O n n et al. - Regulation of striatal neuronal and network interactions
(a) (c) 1
mV
_ _ l 20 mV 600 ms
2
-74 rnV
(d)
(b) 1
__1 20 mV 600 ms
-67 mV
Basal ganglia, Parkinson's disease and levodopa therapy: TINS supplement
Fig. 3. Lesions of dopaminergic terminals by 6-hydroxydopamine increases the spontaneous membrane depolarization and incidence of dye coupling among striatal spiny neurons recorded in vivo and in dopamine-denervated rats. Although the majority of striatal spiny neurons recorded in control anesthetized rats (a) are quiescent, spontaneous spike firing was observed in 35% of spiny neurons. Nonetheless, spontaneous depolarizing potentials (3-16 mV) are observed in almost every spiny cell recorded in control striatum and consist of subthreshold depolarizations (part 1) or long duration plateau depolarization that trigger trains of action potentials (part 2). By contrast, the depolarizing potentials recorded in spiny neurons from dopamine-denervated rats (b) are significantly larger in amplitude (range 11-36 mV) in both nonfiring neurons (part 1) and in neurons that exhibit spontaneous spike activity (part 2). In most cases, injection of single neurons with the dye Lucifer yellow during intracellular recordings results in labeling of only one striatal cell in intact rats. However, in 14% of cases, such injections result in the labeling of more than one neuron, which in control rats is limited to pairs of neurons (c). By contrast, injection of single cells in the dorsal striatum of a 6-hydroxydopamine-lesioned rat results in coupling in 80% of cases (d). Moreover, the coupling often includes dusters of three or more neurons. In this example, a duster of four spiny neurons was recovered. Thus, the incidence and extent of dye coupling among striatal neurons is markedly higher in dopamine-depleted rats. Scale bars, 600 I~m. Adapted, with permission, from Ref. 53.
to the striatum s3,61 or withdrawal from repeated amphetamine administration s4, also causes increased coupling between spiny neurons in the striatal com- plex. One factor that each of these treatments have in common is their propensity to increase corticostriatal activity 3's3'ss'sg. Indeed, corticostriatal stimulation was found to increase coupling between striatal neurons 72. Coupling also appears to depend directly on DA- mediated modulation as well. Thus, acute adminis- tration of a D2-receptor agonist increases coupling in the striatal complex. By contrast, Dl-receptor agonists decrease coupling in the striatum 62'63 and other brain regions 7~'74. Moreover, DARPP-32-knockout mice, in which the Dl-receptor-mediated signaling is disrupted, show baseline levels of coupling equal to those obselwed after D2-receptor-agonist administration. Studies in these mutant mice also show that D2-receptor agonists are ineffective in causing a further increase in coupling above the elevated baseline 9.
Recent anatomical evidence has revealed that DARPP- 32-positive cell bodies are apposed by dense basket-like nitric oxide (NO)-synthase-positive fibers, often forming bouton-like enlargements 7s. This is consistent with sev- eral studies supporting a role for NO in regulating DA- and glutamate-mediated interactions in the striatum (see Ref. 76 for a review). This facilitatol T effect of NO appears to be physiologically significant, as endogenous NO gen- erated in striatal intemeurons via the activation of nitric oxide synthase (NOS) has been shown to elevate extra- cellular levels of DA and glutamate 77. Recently, this facilitation of DA and glutamate release by NO has been proposed to function as a positive-feedback cycle that is crucially involved in the induction of synaptic plasticity in corticostriatal pathways (for a review see Ref. $9). Thus, episodes of intense corticostriatal signaling might allow a temporal association between spiny projection neurons, NOS interneurons and their respective DA inputs, which could lead to the synchronization of functionally related striatal circuits. This is supported by studies showing that NO produced after activation of corticostriatal pathways increases dye coupling in spiny neurons in vitro 72. More- over, intrastriatal NO infusion enhances both the firing rate and the duration of burst firing in striatal neurons and modulates their responsiveness to stimulation of the prefrontal cortex 78'79. Thus, NOS interneurons might synchronize the spike activity of clusters of electrotoni- cally coupled striatal projection neurons.
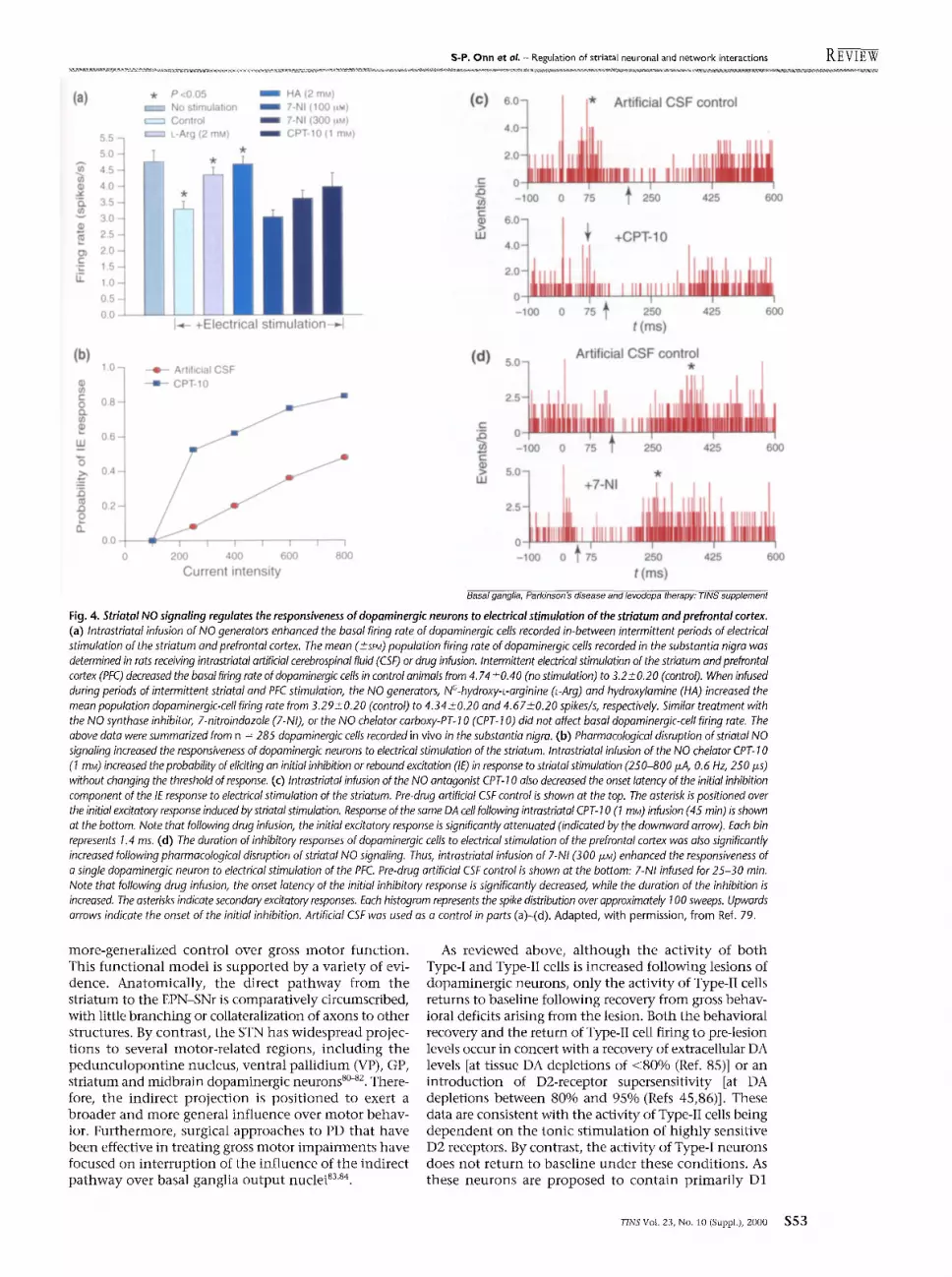
Given the high degree of convergence of striatal efferents on basal-ganglia output neurons in the GP and substantia nigra 1, this NO-mediated regulation of striatal network activity might produce a functionally signifi- cant impact on striatal target nuclei. Indeed, striatal NO was found to regulate the activity of dopaminergic neurons in the substantia nigra indirectly by modulating the activity of striatonigral feedback pathways 79 (Fig. 4). When taken together with neurochemical reports that NO-mediated transmission enhances extracellular DA levels 77, NO appears to play an important role in regulat- ing tonic and phasic DA-mediated transmission. In this model, basal levels of NO are proposed to be involved in maintaining tonic extracellular DA levels in the absence of intense corticostriatal signaling. When the animal is presented with behaviorally relevant stimuli or when corticostriatal pathways are artificially activated, NO signaling can facilitate phasic DA-mediated transmission by increasing the firing activity of dopaminergic cells. Thus, striatal NO-mediated transmission might represent an important pathway by which cortical centers regulate temporally restricted modes of dopamine-mediated transmission differentially in a manner determined by the environmental stimuli presented to the animal.
Direct and indirect output pathways: actions of L-doRa
The direct and indirect output pathways were initially proposed to exert opposing influences on their target sites, as derived from an analysis of the direct inhibitory projection to the EPN-SNr versus the disinhibitory action of the indirect pathway (that is, striatum inhibits
GP, which in turn disinhibits the excitatory pathway between the STN and the EPN-SNr). However, based on an analysis of the anatomy, pharmacology and the response of these systems to lesions, it is proposed that these two pathways subserve distinct functions with respect to motor control: that is, the direct pathway is involved in fine motor control and complex motor functions, whereas the indirect pathway exerts a
S S 2 TINS Vol. 23, No. 10 (Suppl.), 2000
S-P. O n n et aL - Regulation of striatal neuronal and network interactions REviEW
( a )
~9 v
t-
L!~
(b)
(1) c/) c- O r.,9 .e LIJ "6 =>, 25
o 13..
5.5q 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 0.5 0.0
1.0-
0.8-
0.6-
0.4-
0.2-
0.0 o
"k P<0.05 i HA(2mM) No stimulation i 7-NI (100 ~,M) Control i 7-NI (300 F~M) L-Arg (2 rnM) i CPT-10 (t raM)
I~-- +Electrical s t i m u l a t i o n - ~
Artificial CSF --m-- CPT-10
'1= I I I I I I I
200 400 600 800 Current intensity
(0) 6.0~ -k Artificial CSF control
2.04"0!U I'~'11 ~[i1~11111|' M i l l ' II IIIli I1 ' " [1t~"1 ~"~ ~ o .~ I j, t i I
-100 0 75 T 250 425 600 r-
I+ LU +CPT-10
lll dl o I IIIIImlll IIIII I I I I I I I I I I I l l I II I I - 1 0 0 0 5 250 425 600
t (ms)
( d )
r- 25
e-
A r t i f i c i a l CSF control
0 I I I I I , , ,
- 1 0 0 0 75 T 250 425 600
tu +7-NI
2.5
o I IIIIltlll L I I
-100 0 ~ 75 250 425 600 t (ms)
Basal gangfia, Parkinson's disease and levodopa therapy." TINS supplement
Fig. 4. Striatal NO signaling regulates the responsiveness of dopaminergk neurons to electrical stimulation of the striatum and prefrontal cortex. (a) Intrastriatal infusion of NO generators enhanced the basal firing rate of dopaminergic cells recorded in-between intermittent periods of electrical stimulation of the striatum and prefrontal cortex. The mean ( +-- SEM) population firing rate of dopaminergic cells recorded in the substantia nigra was determined in rats receiving intrastriatal artificial cerebrospinal fluid (CSF) or drug infusion. Intermittent electrical stimulation of the striatum and prefrontal cortex (PFC) decreased the basal firing rate of dopaminergic cells in control animals from 4.74 +0.40 (no stimulation) to 3.2+_0.20 (control). When infused during periods of intermittent striatal and PFC stimulation, the NO generators, NC-hydroxy-L-arginine O-Arg) and hydroxylamine (HA) increased the mean population dopaminergic-cell firing rate from 3.29+_0.20 (control) to 4.34+_0.20 and 4.67±0.20 spikes/s, respectively. Similar treatment with the NO synthase inhibitor, 7-nitroindazole (7-NI), or the NO chelator carboxy-PT- 10 (CPT- 10) did not affect basal dopaminergic-cell firing rate. The above data were summarized from n = 285 dopaminergic cells recorded in vivo in the substantia nigra. (b) Pharmacological disruption of striatal NO signaling increased the responsiveness of dopaminergic neurons to electrical stimulation of the striatum. Intrastriatal infusion of the NO chelator CPT- 10 (1 mM) increased the probability of eliciting an initial inhibition or rebound excitation (IE) in response to striatal stimulation (250-800 ixA, 0.6 Hz, 250 ffs) without changing the threshold of response. (c) Intrastriatal infusion of the NO antagonist CPT- 10 also decreased the onset latency of the initial inhibition component of the IE response to electrical stimulation of the striatum. Pre-drug artificial CSF control is shown at the top. The asterisk is positioned over the initial excitatory response induced by striatal stimulation. Response of the same DA cell following intrastriatal CPT- 10 (1 mM) infusion (45 min) is shown at the bottom. Note that following drug infusion, the initial excitatory response is significantly attenuated (indicated by the downward arrow). Each bin represents 1.4 ms. (d) The duration of inhibitory responses of dopaminergic cells to electrical stimulation of the prefrontal cortex was also significantly increased following pharmacological disruption of striatal NO signaling. Thus, intrastriatal infusion of 7-NI (300 I~M) enhanced the responsiveness of a single dopaminergic neuron to electrical stimulation of the PFC Pre-drug artificial CSF control is shown at the bottom: 7-NI infused for 25-30 min. Note that following drug infusion, the onset latency of the initial inhibitory response is significantly decreased, while the duration of the inhibition is increased. The asterisks indicate secondary excitatory responses. Each histogram represents the spike distribution over approximately 100 sweeps. Upwards arrows indicate the onset of the initial inhibition. Artificial CSF was used as a control in parts (a)-(d). Adapted, with permission, from Ref. 79.
more-generalized control over gross moto r function. This functional model is supported by a variety of evi- dence. Anatomically, the direct pa thway from the striatum to the EPN-SNr is comparatively circumscribed, with little branching or collateralization of axons to other structures. By contrast, the STN has widespread projec- tions to several motor-related regions, including the pedunculopont ine nucleus, ventral pallidium (VP), GP, striatum and midbrain dopaminergic neurons 8°-8z. There- fore, the indirect projection is posi t ioned to exert a broader and more general influence over motor behav- ior. Furthermore, surgical approaches to PD that have been effective in treating gross motor impairments have focused on interruption of the influence of the indirect pa thway over basal ganglia output nuclei 83'84.
As reviewed above, a l though the activity of bo th Type-I and Type-II cells is increased following lesions of dopaminergic neurons, only the activity of Type-II cells returns to baseline following recovery from gross behav- ioral deficits arising from the lesion. Both the behavioral recovery and the return of Type-II cell firing to pre-lesion levels occur in concert with a recovery of extracellular DA levels [at tissue DA depletions of <80% (Ref. 85)] or an introduction of D2-receptor supersensitivity [at DA depletions between 80% and 95% (Refs 45,86)]. These data are consistent with the activity of Type-II cells being dependent on the tonic stimulation of highly sensitive D2 receptors. By contrast, the activity of Typed neurons does not return to baseline under these conditions. As these neurons are proposed to contain primarily D1
TINS VoI. 23, No. 10 (Suppl.), 2000 S53

RE~EW S-P. Onn et o l . - Regulation of striatal neuronal and network interactions
receptors and are less sensitive to DA, the activity of Type-I neurons is proposed to be dependent more on the actions of spike-dependent DA released into the synapse (where it reaches very high concentrations) than on the level of extracellular DA. This system would not recover spontaneously, because the synapses lost to the lesion are not replaced. Such a condition implies that there are behaviors that are not restored upon recovery from gross behavioral deficits such as akinesia. Indeed, such behavioral deficits are reported to occur with smaller lesion sizes, and are not ameliorated by acute c-dopa treatment or transplants 42'87'88. This model is also consistent with studies showing that lesion sizes as small as 60% depletion (which do not induce gross behavioral deficits 89'9°) alter substance P and dynorphin levels in the striatum, whereas much larger lesions are required to affect enkephalinergic systems 91. This cor- respondence can be extended to studies of the direct and indirect projection systems. Accordingly, metabolic studies have demonstrated that DA depletion sufficient to result in akinesia [that is, >95% (Refs 89,90)] is required to observe changes in 2-deoxyglucose accumu- lation in the GP and the STN (Refs 45,46). By contrast, in rats with smaller unilateral depletions of striatal DA, the circling behavior produced by administration of a DA-receptor agonist is accompanied by increased 2-deoxyglucose accumulation in the ipsilateral SNr
(Refs 80,92). These two systems thereby work in coordi- nation to regulate distinct but interdependent aspects of motor function, with each being affected differently by its DA innervation.
For further discussion on this topic see Box 1.
S e l e c t e d r e f e r e n c e s 1 Smith, Y. et al. (1998) Microcircuitry of the direct and indirect
pathways of the basal ganglia. Neuroscience 86, 353-387 2 Grace, A.A. (1991) Phasic versus tonic dopamine release and the
modulation of dopamine system responsivity: a hypothesis for the etiology of schizophrenia. Neuroscience 41, 1-24
3 Calabresi, P. etal.(1990)Synapticandintrinsiccontrolofmembrane excitability of neostriatal neurons II. An in vitro analysis. J. NeurophysioL 63, 654-662
4 Levine, M.S. et al. (1996) Neuromodulatory actions of dopamine on synaptically-evoked neostriatal responses in slices. Synapse 24, 65-78
50'Donnell , P. and Grace, A.A. (1994) "Conic D2-mediated attenuation of cortical excitation in nucleus accumbens neurons recorded in vitro. Brain Res. 634, 105-112
6 Twery, M.J. (1994) Intracellularly recorded response of rat striatal neurons in vitro to fenoldopam and SKF 38393 following lesions of midbrain doparnine cells. Synapse 18, 67-78
7 Uchimura, N. et al. (1986) Hyperpolarizing and depolarizing actions of dopamine via D1 and D2 receptors on nucleus accumbens neurons. Brain Res. 374, 368-372
8 White, F.J. and Wang, R.Y. (1986) Electrophysiological evidence for the existence of both D-1 and D-2 dopamine receptors in the rat nucleus accumbens. J. NeuroscL 6, 274-280
9 0 n n , S-P. etal. (1996) Dopamine modulates membrane excitability of striatal spiny cells via DARPP-32 signal transduction pathway. Soc. Neurosci. Abstr. 26, 1089
$54 rms Vol. 23, No. 10 (Suppl.), 2000

$-P. Onn et a l . - Regulation of striatal neuronal and network interactions RE-VI E W
10 An, X. etal. (1999) Tyrosine hydroxylase-immunoreactive terminals in the monkey accumbens core and shell: synaptic relationship to dopamine D1 and D2 receptors. Soc. Neurosci. Abstr. 25, 1470
11 Sesack S.R. etal. (1994) Ultrastructural localization of D2 receptor- like immunoreactivi ty in midbrain dopamine neurons and their striatal targets. J. Neurosci. 14, 88-106
12 Brown, J.R. and Arbuthnott, G.W. (1983) The electrophysiology of dopamine (D2) receptors: a study of the actions of dopamine on corticostriatal transmission. Neuroscience 10, 349-355
13 Hersch, S.M. et al. (1995) Electron microscopic analysis of D1 and D2 dopamine receptor proteins in the dorsal striatum and their synaptic relationships with motor corticostriatal afferents. J. NeuroscL 15, 5222-5237
14 Levey, A.L. et al. (1993) Localization of D1 and D2 dopamine receptors in brain with subtype-specific antibodies. Proc. Natl. Acad. SCi. U. S. A. 90, 8861-8865
15 Harvey, J. and Lacey, M.G. (1997) A postsynaptic interaction between dopamine D1 and NMDA receptors promotes presynaptic inhibition in the rat nucleus accumbens via adenosine release. J. Neurosci. 17, 5271-5280
16 Nicola, S.M. et al. (1996) Psychostimulants depress excitatory synaptic transmission in the nucleus accumbens via presynaptic Dl-like dopamine receptors. J. Neurosci. 16, 1591-1604
17 Pennartz, C.M.A. et al. (1992) Presynaptic dopamine D1 receptors at tenuate excitatory and inhibitory limbic inputs to the shell region of the rat nucleus accumbens studied in vitro. J. Neurophysiol. 67, 1325-1334
18 Cepeda, C. et al. (1993) Neuromodulatory actions of dopamine in the neostriatum are dependent upon the excitatory amino acid receptor subtypes activated. Proc. Natl. Acad. SO. U. S. A. 90, 9576-9580
19 Zheng, P. et al. (1999) Opposing modulat ion of cortical NMDA receptor-mediated responses by low and high concentrations of dopamine. Neuroscience 91,527-535
20 Hu, X.T. and Wang, R.Y. (1988) Comparison of effects of D-1 and D-2 dopamine receptor agonists on neurons in the rat caudate putamen: an electmphysiologicaI study. J. Neurosci. 8, 4340-4348
21 Kiyatkin, E.A. and Rebec, G.V. (1996) Dopaminergic modulat ion of glutamate-induced excitations of neurons in the neostriatum and nucleus accumbens of awake, unrestrained rats. J. Neurophysiol. 75, 142-152
22 Colwell, C.S. and Levine, M.S. (1995) Excitatory synaptic transmission in neostriatal neurons: regulation by cyclic AMP-dependent mechanisms. J. Neurosci. 15, 1704-1713
23 Fienberger, A.A. et al. (1998) DARPP-32: regulator of the efficacy of dopaminergic neurotransmission. Science 281, 838-842
24 Snyder, G.L. et al. (1998) A dopamine/D1 receptor/protein kinase A/ dopamine- and cAMP-regulated phosphoprotein (Mr. 32 kDa)/protein phosphatase-1 pathway regulates dephosphorylation of the NMDA receptor. 1. Neurosci. 18, 10297-10303
25 Konradi, C. et al. (1996) Amphetamine and dopamine-induced immediate early gene expression in striatal neurons depends on postsynaptic NMDA receptors and calcium. J. NeuroscL 16, 4231~t239
26 Wang, J.Q. et al. (1994) NMDA receptors mediate amphetamine- induced regulation of zif/268 and preprodynorphin mRNA expression in rat striatum. Synapse 18, 343-353
27 Calabresi, P. et al. (1987) Intracellular studies on the dopamine- induced firing inhibition of neostriatal neurons in vitro: evidence for D1 receptor involvement. Neuroscience 20, 757-771
28 Nishi, A. et al. (1997) Bidirectional regulation of DARPP-32 phosphorylat ion by dopamine. J. Neurosci. 17, 8147-8155
29 May, LJ. et al. (1988) Differentiation of dopamine overflow and uptake progress in the extracellular fluid of rat caudate nucleus with fast-scan in vivo voltammetry. J. Neurochem. 51, 1060-1069
30 Schultz, W. (1998) Predictive reward signal of dopamine neurons. J. Neurosci. 80, 1-27
31 Gonon, F. (1997) Prolonged and extrasynaptic excitatory action of dopamine mediated by D1 receptors in the rat striatum in vivo. J. Neurosci. 17, 5972-5978
32 Smiley, J.F. et al. (1994) D1 dopamine receptor immunoreactivity in h u m a n and monkey cerebral cortex: predominent and extra- synaptic localization in dendritic spines. Proc. Natl. Acad. SCi. U. S. A. 91, 5720-5724
TINS Vol. 23, No. 10 (Suppl.), 2000 S 5 5

RE-VIEW s-P. O n n et ol. - Regulation of striatal neuronal and network interactions
33 Skirboll, L.R. and Bunney, B.S. (1979) The effects of acute and chronic haloperidol treatment on spontaneous firing neurons in caudate nucleus of the rat. Life ScL 25, 1419-1434
34 Nisenbaum, E.S. etal. (1988) Evidence for two funcfionally distinct subpopulations of neurons within the rat striatum. ]. Neurosci. 8, 4138-4150
35 Onn, S-P. etaL (1994) Identification and characterization of striatal cell types us ing in vivo intracellular recordings in rats. I. Basic physiology and response patterns to corticostriatal fiber stimulation. Synapse 16, 161-180
36 Nisenbaum, E.S. et al. (1992) Functionally distinct subpopulations of striatal neurons are differentially regulated by GABAergic and dopaminergic inputs. 11. In vitro analysis. Neuroscience 48, 579-593
37 Nisenbaum, E.S. et al. (1992) Functionally distinct subpopulations of striatal neurons are differentially regulated by GABAergic and dopaminergic inputs. I. In vivo analysis. Neuroscience 48, 561-578
38 Onn, S-P. etal. (1994) Identification and characterization of striatal cell subtypes using in vivo intracellular recording and dye labeling in rats. Ill. Morphological correlates and compartmental organization. Synapse 16, 231-254
39 Onn, S-P. et al. (1994) Identification and characterization of striatal cell types using in vivo intracellular recording in rats: II. Membrane factors underlying paired-pulse response profiles. Synapse 16, 195-210
40 Nisenbaum, E.S. et al. (1986) Long-term effects of dopamine- depleting brain lesions on spontaneous activity of Type II striatal neurons: relation to behavioral recovery. Brain Res. 398, 221-230
41 Nisenbaum, E.S. etal. (1988) Spontaneous activity of Type I1 but not Type 1 striatal neurons is correlated with recovery of behavioral funct ion after brain lesion. Brain Res. 473, 389-393
42 Miller, W.C. and DeLong, M.R. (1987) Altered tonic activity of neurons in the globus pallidus and subthalamic nucleus in the primate MPTP model of parkinsonism. In The Basal Ganglia II (Carpenter, M.B. and Jayaraman, A., eds), pp. 415M~27, Plenum Press
43 Filion, M. et al. (1988) Abnormal influences of passive limb movement on the activity of globus pallidus neurons in parkinsonian monkeys. Brain Res. 444, 165-176
44 Wichman, T. et al. (1990) Increased neuronal activity in the subthalamic nucleus of MPTP treated monkeys. Mov. Disord. (Suppl. 1), 78
45 Kozlowski, M.R. and Marshall, J.F. (1980) Plasticity of C ~4 2-deoxy- d-glucose incorporation into neostriatum and related structures in response to dopamine neuron damage and apomorphine replacement. Brain Res. 197, 167-183
46 Mitchell, I.J. et aL (1989~ Neuronal mechanisms underlying parkinsonian symptoms based upon regional uptake of 2- deoxyglucose in monkeys exposed to 1-methyl-4-phenyl-l,2,3,6- tetrahydropyridine. Neumscience 32, 213-226
47 Bennett, B.D. and Bolam, J.p. (1994) Synaptic input and output of parvalbumin-immunoreactive neurons in the neostriatum of the rat. Neuroscience 62, 707-719
48 Jaeger, D. el al. (1994) Surround inhibition among projection neurons is weak or nonexis tent in the rat neostriatum. J. Neurophysiol. 72, 2555-2558
49 Cowan, R.L. et al. (1990) Parvalbumin-containing GABAergic interneurons in the rat neostriatum. J. Comp. Neurol. 302, 197-205
50 Kita, H. etaL (1990) Parvalbumin-immunoreactive neurons in the rat neostriatum: a light and electron microscopic study. Brain Res. 536, 1-15
51 Leung, L.S. and Yim, C.Y.C. (1993) Rhythmic delta-frequency activities in the nucleus accumbens of anesthetized freely moving rats. Can. J. Physiol. Pharmacol. 71,311-320
52 O'Donnell, P. and Grace, A.A. (1995) Synaptic interactions anrong excitatory afferents to nucleus accumbens neurons: hippocampal gating of prefrontal cortical input. J. Neurosci. 15, 3622-3639
53 Onn, S-P. and Grace, A.A. (1999) Alterations in electrophysiological activity and dye coupling of striatal spiny and aspiny neurons in dopamine-denervated rat striatum recorded in vivo. Synapse 31, 1-15
54 Onn, S-P. and Grace, A.A. (2000) Amphetamine withdrawal alters bistable states and cellular coupling in rat prefrontal cortex and nucleus accumbens recorded in viw). ]. Neurosci. 20, 2332-2345
55 Stern, E.A. et al. (1998) Membrane potential synchrony of simulta- neously recorded striatal spiny neurons in vivo. Nature 394, 475-478
56 Wilson, C.J. and Kawaguchi, Y. (1996) The origins of two-state spontaneous membrane potential fluctuations of neostriatal spiny neurons. J. Neurosci. 16, 2397-2410
57 Yim, C.Y. and Mogenson, G.J. (1988) Neuromodulatory action of dopamine in the nucleus accumbens: an in vivo intracellular study. Neuroscience 26, 403-415
58 Calabresi, P. et al. (1999) Glutamate-triggered events inducing corticostriatal long-term depression. J. NeuroscL 19, 2489-2499
59 Calabresi, P. et al. (2000) Synaptic transmission in the striatum: from plasticity to neurodegeneration. Prog. Neurobiol. 61,231-265
60 Onn, S-P. and Grace, A.A. (1995) Repeated treatment with haloperidol and clozapine exerts differential effects on dye coupling between neurons in subregions of striatum and nucleus accumbens. J. NeuroscL 15, 7024-7030
61 Cepeda, C. et al. (1989) Dye coupling in the neostriatum of the rat. I. Modulation by dopamine lesions. Synapse 4, 229-237
62 O'Donnell, P. and Grace, A.A. (1993) Dopaminergic modulat ion of dye coupling between neurons in the core and shell regions of
the nucleus accumbens. J. NeuroscL 13, 3486-3471 63 Onn, S-P. and Grace, A.A. (1994) Dye coupling between rat striatal
neurons recorded in vivo: compartmental organization and modulation by dopamine. ]. Neurophysiol. 71, 1917-1934
64 Phelan, K.D. et aL (1993) Morphological and electrophysiological evidence for electrotonic coupling of rat dorsolateral septal nucleus neurons in vitro. Synapse 13, 39-49
65 Grace, A.A. and Bunney, B.S. (1983) Intracellular and extracellular electrophysiology of nigral dopaminergic neurons L Identification and characterization. Neuroscience 10, 317-331
66 Andrew, R.D. et al. (1982) Coupling in rat hippocampal slices: dye transfer between CA1 pyramidal cells. Brain Res. Bull. 8, 211-222
67 Nunez, A. et al. (1990) In vivo electrophysiological analysis of Lucifer yellow-coupled hippocampal pyramids. Exp. Neurol. 108, 76-82
68 Hatton, G.I. and Yang, Q.Z. (1990) Activation of excitatory amino acid inputs to supraopfic neurons. I. Induced increases in dye-coupling in lactating, but not virgin or male rats. Brain Res. 513, 264-269
69 Yang, Q.Z. and Hatton, G.I. (1999) Nitric oxide via cGMP- dependent mechanisms increases dye coupling and excitability of rat supraoptic nucleus neurons. I. Neurosci. 19, 4270-4279
70 Eghbali, B. et al. (1990) Expression of gap junction channels in communication-incompetent cells after stable h'ansfection with cDNA encoding connexin 32. Proc. Natl. Acad. Sci. U. S. A. 87, 1328-1331
71 Moreno, A.P. et al. (1992) Connexin 32 gap junction channels in stably transfected cells: unitary conductance. Biophys. J. 62, 51-53
72 O'Donnell, P. and Grace, A.A. (1997) Cortical afferents modulate striatal gap junction permeability via nitric oxide. Neuroscience 76, 1-5
73 Harsanyi, K. and Mangel, S.C. (1992) Activation of a D2 receptor increases electrical coupling between retinal horizontal cells by inhibit ing dopamine release. Proc. Natl. Acad. Sci. U. S. A. 89, 9220-9224
74 Rorig, B. et al. (1995) Dye coupling between pyramidal neurons in developing rat prefrontal and frontal cortex is reduced by protein kinase A activation and dopamine. J. Neurosci. 15, 7386-7400
75 Gomez-Urquijo, S.M. et al. (1999) Neurocircuits of the basal ganglia studied in organotypic cultures: focus on tyrosine hydroxylase, nitric oxide synthase and neuropeptide immunocytochemistry. Neuroscience 94, 1133-1151
76 Moore, H. et al. (1999) The regulation of forebrain dopamine transmission: relevance to the pathophysiology and psycho- pathology of schizophrenia. BioL Psychiatry 46, 42-55
77 West, A.R. and Galloway, M.R. (1997) Endogenous nitric oxide facilitates striatal dopamine and glutamate efflux in vivo: role of ionotropic glutamate receptor-dependent mechanisms. Neuropharrnacology 36, 1571-1581
78 West, A.R. and Grace, A.A. (1999) Intrastriatal infusion of nitric oxide increases the firing rate and duration of burst firing of striatal neurons recorded in viw). Soc. NeuroscL Abstr. 25, 173
79 West, A.R. and Grace, A.A. (2000) Striatal nitric oxide signaling regulates the neuronal activity of midbrain dopamine neurons in vivo. J. NeurophysioL 83, 1796-1808
80 Groenwegen, H.J. and Berendse, H.W. (1990) Connections of the subthalamic nucleus with ventral striatopallidal parts of the basal ganglia in the rat. J. Comp. Neurol. 294, 607-622
81 Smith, LD. and Grace, A.A. (1992) Role of the subthalamus nucleus in the regulation of nigral dopamine neuron activity. Synapse 12, 287-303
82 Takada, M. etaL (1988) Two separate populations of the subthalamic nucleus project to the basal ganglia and to the pedunculopontine tegmental region. Brain Res. 442, 72-80
83 Delong, M.R. and Wichmann, T. (1993) Basal ganglia-thalamocor tical circuits in parkinsonian signs. Clin. NeuroscL 1, 18-26
84 Svennilson, E. etal. (1960) Treatment of par~nsonism by stereotactic thermolesions in the pallidal region. Acta Psychiatr. Neurol. Scand. 35, 358-377
85 Robinson, T.E. and Whishaw, I.Q. (1988) Normalization of extrace/lular dopamine in striatum following recovery from a partial unilateral 6-OHDA lesion of the substantia nigra: a microdialysis study in freely moving rats. Brain Res. 450, 209-224
86 Wooten, G.F. and Collins, R.C. (1983) Effects of dopaminergic stimulation on functional brain metabolism in rats with unilateral substantia nigra lesions. Brain Res. 263, 267-275
87 Lindner, M.D. et aL (1995) Implantation of encapsulated catechol- amine and GDNF-producing cells in rats with unilateral dopamine depletions and parkinsonian symptoms. Exp. Neurol. 132, 62-76
88 Spirduso, W.W. etal. (1985) Reactive capacity: a sensitive behavioral marker of movenrent initiation and nigrostriatal dopamine function. Brain Res 335, 45-54
89 Zigmond, M.J. et al. (1984) Neurochemical compensat ion after nigrostriatal bundle injury in an animal model of preclinical Parkinsonism. Arch. Neurol. 41,856-861
90 Zigmond, M.J. et al. (1990) Compensat ions after lesions of central dopaminergic neurons: some clinical and basic implications. Trends NeuroscL 13, 290-296
91 Nisenbaum, L.K. et al. (1994) Partial lesions of the nigrostriatal dopamine pathway alter substance P but not enkephalin mRNAs in the rat striatum. Soc. Neurosci. Abstr. 20, 922
92 Brown, L.L. and Wolfson, L.I. (1983) A dopamine-sensitive striatal efferent system mapped with ~C-deoxyglucose in the rat. Brain Res. 261,213-229
$ 5 6 71NS Vol. 23, No. l0 (Suppl.), 2000