ringkasan - perpustakaan pertanian
TRANSCRIPT

Vll
Balai Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian (Balitbiogen) adalahsalah satu lembaga penelitian di bawah Badan Penelitian dan Pengembangan Pertanianyang mempunyai tugas melaksanakan penelitian bioteknologi dan rintisan tanamanpangan. Penelitian di Balitbiogen terdiri atas tujuh program yang dilaksanakan oleh parapeneliti yang berasal dari lima kelomppk, yaitu (1) Sumber Daya Genetika, (2) BiologiMolekuler, (3) Reproduksi dan Pertumbuhan, (4) Rekayasa Protein dan Imunologi, serta (5)Mikrobiologi dan Teknologi Proses.
Balitbiogen dalam menjalankan fungsinya didukung oleh 361 orang pegawai negerisipil (PNS) yang terdiri dari 60 orang pejabat fungsional peneliti, 43 orang litkayasa, tigaorang pranata komputer, dua orang pustakawan, satu orang statistisi, dan 252 orang sebagaitenaga penunjang (pembantu penelitian dan administrasi). Selain didukung PNS,Balitbiogen juga didukung oleh 78 orang tenaga honorer yang terdiri dari satu orangPascasarjana, tiga orang Sarjana, 36 orang SLTA, tujuh orang SLTP, dan 31 orang SD.
Pada tahun anggaran (TA) 2002 kegiatan penelitian di Balitbiogen ditopang danasebesar Rp 13.389.989.000 yang terdiri atas anggaran rutin (Rp 7.298.139.000), anggaranpembangunan (Rp 5.827.020.000), dan kerja sama penelitian (Rp 264.830.000). Anggaranrutin digunakan untuk belanja pegawai (gaji PNS), barang, pemeliharaan, dan perjalanan.Sedangkan anggaran pembangunan digunakan antara lain untuk penelitian, tunjanganpelaksana penelitian (TPP), honor pelaksana penelitian (HPP), penyebarluasan hasil penelitian, pengadaan alat laboratorium serta pengadaan buku dan jumal ilmiah. Sumber danalain diperoleh dari proposal yang diusulkan oleh peneliti Balitbiogen ke berbagai sumberpendanaan.
Pada TA 2002 adalah telah diperbaharui dan dikonservasi plasma nutfah tanamanpangan yang meliputi 750 aksesi padi sawah dan padi gogo, 80 aksesi padi liar, 400 aksesijagung, 500 aksesi kedelai, 550 aksesi ubi kayu, 75 aksesi terigu, 208 aksesi sorgum, 600aksesi kacang tanah, 400 aksesi kacang hijau, 140 aksesi kacang-kacangan minor, 1.739aksesi ubi jalar, 158 aksesi ubi-ubian minor dan 115 aksesi talas. Selain itu, telahdikonservasi secara in vitro 10 aksesi plasma nutfah ubi kayu, 70 aksesi ubi jalar, dan 10aksesi talas.
Hasil karakterisasi sifat morfologi menunjukkan 24 varietas padi memiliki malaipanjang (>30 cm) dan biji berat G>30 g/1000 biji); 17 varietas jagung dengan bobot 300 biji>80 g; 9 aksesi kedelai berbiji besar (10,5 g/100 biji); 7 aksesi ubi kayu mempunyai hasilumbi per pohon >3,0 kg; 2 aksesi terigu mempunyai hasil tinggi (>2,5 kg/petak); 6 aksesisorgum ber biji berat (>80 g/malai); 33 aksesi kacang tanah yang mempunyai jumlahpolong >20 polong per tanaman dimana 40 aksesi diantaranya memiliki 3-4 biji /polong; 13plasma nutfah kacang hijau mempunyai bobot biji per tanaman tinggi (12,7-22,3 g) danbobot 1000 biji tinggi (57,7-67,3 g), 3 aksesi toleran terhadap penyakit bercak dauncercospora; 310 aksesi ubi jalar berumbi besar dengan variasi jumlah umbi per tanamanantara 2-10 umbi, 5 aksesi diduga tahan terhadap hama lanas; umur berbunga dan umurpanen kacang tunggak antara 39-60 hari dan 73-88 hari; sifat morfologi plasma nutfah talasberagam kecuali bentuk daun; tiga aksesi garut dengan hasil 1,0-1,2 kg/rumpun, 4 aksesi ubi
Ringkasan

viii
kelapa dengan hasil 4,7-13,0 kg/rumpun, 5 aksesi gembiii dengan hasil 1,2-2,25 kg/rumpun,dan 4 aksesi ganyong dengan hasil 2,5-2,84 kg/rumpun.
Hasil evaluasi menunjukkan 24 aksesi padi dan 14 aksesi kedelai toleran Al; 10 aksesijagung sedang sampai toleran Al; dan 107 aksesi padi toleran Fe. Delapan aksesi padi agaktahan sampai tahan terhadap penyakit hawar daun bakteri (HDB) kelompok Viii; 77 aksesipadi agak tahan sampai tahan terhadap penyakit HDB kelompok IV; 10 aksesi padi agaktahan terhadap penyakit bias; dan 20 aksesi plasma nutfah jagung tahan terhadap penyakitbulai dengan persentase serangan 0-5%. Satu aksesi ubi jalar tahan terhadap hama Ianas.
Hasil karakterisasi mutu gizi plasma nutfah diperoleh 7 aksesi padi ketan dengan kadaramilosa < 10%, 18 aksesi bukan padi ketan mempunyai kadar amilosa rendah dan sedang(10-23%); 5 aksesi jagung termasuk jagung ketan dengan kadar amilosa rendah (<20%); 7aksesi kedelai mengandung kadar protein tinggi (+39%); 5 aksesi kacang tanah dengankadar protein sedang (27,3-38,0%) dan 2 aksesi dengan kadar lemak cukup tinggi (35 dan38%); kadar pati 44 aksesi ubi jalar dan 10 aksesi ubi kelapa mempunyai kadar pati >25%;12 aksesi ubi kayu mengandung HCN rendah (<22 ppm); 14 aksesi sorgum memilikikandungan tanin rendah (<0,25%) dengan warna biji putih dan kuning.
Hasil karakterisasi molekuler menunjukkan primer A01, A04, dan B01 dapatmengamplifikasi DNA dari 97 plasma nutfah ubi jalar yang berasal dari Irian dengan jumlahpita DNA bervariasi antara 1 -9 pita.
Database plasma nutfah tanaman pangan telah menampung 7.988 record yang meliputi3.563 aksesi padi, 705 aksesi jagung, 174 aksesi sorgum, 771 aksesi kedelai, 270 aksesikacang tanah, 1.029 aksesi kacang hijau, 434 aksesi ubi kayu, 912 aksesi ubi jalar, dan 130aksesi kacang minor (kacang tunggak).
Hingga tahun 2002, jenis mikroba yang dikoleksi Balitbiogen, yaitu mikroba penyuburtanah (Rhizobium tipe liar, Rhizobium mutan antibiotik, Rhizobium mutan EMS, danRhizobium fusan, Azospirillum, Azotobacter, pelarut-P, filosfer, mikoriza, dan endofitik padidan jagung), mikroba perombak bahan organik (kapang dan bakteri), mikrobabioremediasi, mikroba patogen [Erwinia carotovora pv. carotouora, Ralstoniasolanacearum, Pseudomonas syringae pv. glycinea, Xanthomonas campestris pv.campestris, X. campestris pv. glycines, X. oryzae pv. oryzae, Rhizoctonia solani, Fusariumoxysporum var. cubensis, F. oxysporum, F. solani, Alternaria porri, Elsinoe iwatae,
Cercospora bataticola, dan Alternaria colocasiae), mikroba antagonis [Bacillus subtilis, B.polymixa, Pseudomonas flurescens, Trichoderma harzianum, dan Saccharomyces spp.).
Potensi sebagian isolat mikroba penyubur tanah telah digali dan dikembangkan antara lainsebagai biofertilizer, inokulan benih matriconditioning, dan tetua untuk perbaikan kualitasgenetik inokulan.
Hasil isolasi mikroba perombak bahan organik diperoleh 23 isolat yang terdiri dari 13isolat bakteri dan 10 isolat kapang. Isolat mikroba perombak bahan organik unggul M10memiliki aktivitas enzim tertinggi (0,1298 U/ml).
Hasil uji toleransi galur F7 (IR64 x Cabacu) terhadap kekeringan di lapang pada duatahap perkembangan tanaman yang berbeda diperoleh beberapa galur konsisten toleran,
. konsisten peka, dan tidak konsisten pada kedua pengamatan.

ix
Hasil uji daya tembus akar populasi F7 (IR64 x IRAT112) di rumah kaca menunjukkanjumlah akar yang mampu menembus Win bervariasi (1-11).
Hasil uji genotipe pada 30 lokus markah mikrosatelit dan 33 lokus markah AFLPmenunjukkan bahwa populasi F7 (IR64 x Cabacu) bias ke arah salah satu tetua (IR64) danmemiliki tingkat heterosigositas yang masih cukup tinggi. Diperoleh data genotipe pada 68lokus markah mikrosatelit dari 94 galur F2 (Dupa x ITA131). Hasil analisis keterpautanmenunjukkan 12 primer mikrosatelit yang terpaut dengan sifat ketahanan tanaman padivarietas Danau Tempe terhadap bias. Hasil pengujian ketahanan terhadap penyakit biaspada populasi F8 (Danau Tempe x Kencana Bali) di rumah kaca menunjukan bahwa 103galur tahan, 68 galur agak tahan, dan 7 galur tidak tahan terhadap ras 173. Hasil pengujianterhadap ras 033 menunjukan bahwa pada 74 galur tahan, 48 galur agak tahan, 31 galurtidak tahan, dan 43 galur sangat peka.
Empat primer mikrosatelit, yaitu RM 197 dan RM 276 dari kromosom 6, RM 201 darikromosom 9 dan RM 332 dari kromosom 11 telah digunakan dalam analisis sidik ragam danmenunjukkan polimorfisme antara aksesi spesies padi liar.
Hasil evaluasi ketahanan spesies padi liar terhadap HDB menunjukkan O. minuta me-rupakan spesies yang paling tahan terhadap penyakit HDB strain IV dan VIII. Pada umumnyaO. officinalis merupakan spesies yang sangat baik sebagai sumber gen ketahanan terhadaphama wereng coklat, dan O. officinalis (106520 dan 105220) menunjukkan reaksi palingtahan terhadap penyakit bias ras 041, 073, dan 173. Padi liar O. glaberrima 101297 baik digunakan sebagai tetua dalam persilangan karena mempunyai sifat tahan terhadap kekering-an, umur genjah, tanaman pendek, dan tahan terhadap kerebahan.
Transformasi padi dengan gen crylA(b) untuk ketahanan terhadap penggerek batangpadi Scirphophaga innotata menghasilkan 280 tanaman putatif transgenik Taipei-309 dan 1tanaman putatif transgenik Cisadane. Analisis PCR menghasilkan 4 galur (19 tanaman)Taipei-309 generasi To dan 10 tanaman (2 tanaman Tl, 7 tanaman T2, 1 tanaman T3) yangpositif mengandung gen crylA(b) dan pada pengujian bioasai menunjukkan tingkat ketahanan agak tahan sampai sangat tahan. Bioasai terhadap sundep menghasilkan 13 tanamansangat tahan, 5 tanaman tahan, 9 tanaman agak tahan, sedangkan terhadap beluk menghasilkan 19 tanaman sangat tahan, 2 tanaman tahan, dan 5 tanaman agak tahan.
Analisis PCR untuk gen pin\\ pada tanaman kedelai untuk ketahanan terhadap penggerek polong Etiella zinckenella, menghasilkan 1 tanaman Wilis generasi R2 dan I tanamanTidar generasi R3 yang positif mengandung gen pinll. Bioasai pada tanaman Wilis R3menunjukkan tingkat ketahanan peka sampai sangat peka. Transformasi kedelai dengangen oyIA(b) menghasilkan 2 planlet varietas Tidar.
Analisis PCR tanaman putatif transgenik kacang tanah Rl menghasilkan 3 tanamanyang positif mengandung gen CP-PStV dan menunjukkan tingkat ketahanan tahan (gejalatidak muncul dan intensitas serangan 0%). Bioasai terhadap PStV pada 18 tanaman menghasilkan 8 tanaman tahan, 5 tanaman agak tahan, dan 5 tanaman tidak tahan.
Transformasi ubi jalar untuk ketahanan terhadap hama boleng Cylas formicarius ataupenyakit virus SPFMV melalui Agrobacterium tumefaciens menghasilkan 21 tanaman putatiftransgenik Jewel yang ditransformasi dengan gen pintt dan 5 tanaman putatif transgenikyang ditransformasi dengan gen CP-SPFMV.

Uji pola restriksi dengan enzim Hindlll menghasilkan 2 koloni yang mengandung 2fragmen berukuran 11,837 kb dan 4,887 kb, yang diduga telah mengandung gen a-amilaseinhibitor (a-ai). Uji selanjutnya dengan enzim BamHl dan Xba\ menghasilkan 2 arah gen a-ai, yaitu arah ke kanan (pCambia-aail) dan ke kiri (pCambia-ccai2), yang mempunyai polarestriksi seperti yang diharapkan. Kedua plasmid tersebut telah ditransformasi ke dalamAgrobacterium tumefaciens LBA4404.
Uji gus terhadap 20 eksplan tomat menunjukkan 3 eksplan positif mengekspresikangen gus. Setiap varietas pepaya yang ditransformasi dengan gen gus dan beberapa kalusmenunjukkan ekspresi sementara yang ditandai dengan berubahnya warna kalus dari
kuning menjadi biru akibat bereaksi dengan larutan X-gluc. Larutan ini berfungsi sebagaipendeteksi adanya gen pelapor. Eksplan daun muda krisan dengan gen gus melalui A.
tumefaciens menunjukkan ekspresi positifHasil pengujian morfologi jagung transgenik menunjukkan tidak ditemukan gejala
pertumbuhan yang menyimpang dari morfologi jagung normal. Pada pengujian dayatumbuh, jagung transgenik RR NK603 memiliki daya tumbuh yang lebih baik apabiladibandingkan dengan jagung kontrol. Uji inuasiveness menunjukkan jagung transgenik RRNK 603 memiliki kemampuan kompetisi yang sama dengan jagung kontrol (nilai rata-rataagresivitas -0,247).
Hasil pengujian morfologi kapas transgenik DP 5690 Bt tidak menunjukkan kelainanatau abnormalitas sifat-sifat fenotipik. Uji inuasiveness menunjukkan kapas DP5690 Btmemiliki kemampuan kompetisi yang sama dengan kapas kontrol (nilai rata-rata agresivitas-0,23).
Hasil pengujian morfologi kapas transgenik Nu Pearl Bl/RR tidak menunjukkan kelainanatau abnormalitas sifat-sifat fenotipik. Pada pengujian invasiveness, menunjukkan kapas NuPearl memiliki kemampuan kompetisi yang sama dengan kapas kontrol (nilai rata-rataagresivitas 0,1). Pengujian yang telah dilakukan pada semua tanaman transgenik menunjukkan tidak ditemukannya adanya sifat weedy dan invasive yang berbeda dengan tanamannon transgenik.
Studi regenerasi dan penyimpanan tanaman bernilai ekonomi tinggi secara in vitromenunjukkan formulasi media terbaik untuk merangsang pembentukan tunas melinjoadalah WPM + 0,1 mg/1 thidiazuron + 100 mg/1 glutamine + BA 0,5 mg/1. Perendamanplanlet dalam larutan IBA 500 mg/1 selama 1 jam dalam proses aklimatisasi dapat meng-induksi perakaran tanaman melinjo. Media yang digunakan untuk mengakarkan tanamaniles-iles dalam kultur in vitro adalah media dasar V^ MS, tanpa zat pengatur tumbuh.
Sedangkan media tumbuh terbaik untuk aklimatisasi iles-iles di rumah kaca adalah campur-an tanah dan casting atau campuran tanah dan pupuk kandang dengan perbandingan 1:1.Persentase perakaran manggis paling tinggi (66,7%) pada media WPM berasal dari media %WPM + IBA 10 mg/1 sedangkan persentase paling tinggi (50%) pada media MS berasal darimedia ]A MS + IBA 5 mg/1. Pada umur 3 bulan, akar sudah mulai terbentuk pada 5 mediaWPM. Durasi prakultur yang optimal untuk kultur ubi kayu adalah 4 had. Kombinasi perlaku-
an prakultur 4 hari dan loading selama 20 menit merupakan perlakuan terbaik untuk semuajenis krioprotektan yang diuji. Larutan PVS3 (50% gliserol + 50% sukrosa dalam media MS)mempunyai daya hambat yang paling rendah terhadap pertumbuhan kultur. Kultur yammempunyai toleransi yang cukup tinggi terhadap taraf sukrosa tinggi hingga 0,7 M pada

^jijisads is^Bajaq uep jSSuq ASHH[isBqSuad Biuopuqiq [as nies iuo|o>( qaiojadtuaui >piun Suiuoi^ jaquins uc^jipefip
pdBp SubA (gOH 3-(S)-HH UBp ^^oo -(3)-HH) iSSup q\p^ UBJHiSBqSuaui pisuapd SubABiuopuqiq iuo|O>[ jouiou Bnp qa|ojadtuauj AS^M W^ HSBqSuad Biuopuqiq ps•[ui/ps ,,01 redBDusui iiunui UB^piq isuadsns uiB|Bp sg isjppp Jjrqun u^p ^oiiBdBDUdiu ipBd urpp jpqs^p uBp ASHH !SJR13P ^nlun VSI13 ie>lSuBJ3d
UB|iSBqj3q3>i UBjpBJfSuiusui i/Sui i VSUBp qBqiUBDSjp^q ^d^p (S-HA ubSusp PIBA\ bjbiub uBSuB[isj3d iisblj BUiBiruaj) ouquisBqooip SubA iBSBp Bipdui Bnuiss BpB^ nSSuiui z u^p i Bnj qiqaj SubA uiBjiq Subdbjj ubSuspUB^SuipuBqip BfiqsdB iSSup qiqaj SubA UBqBquiBDd^Jdd UB|isBqj3q3^j uB^uaquiaoi (nSSuiuig) Bni qiqaj SubA uiB^iq Subdb>j usSuap (p) 13(ba^ UBSuB^isisd jisbij ouquis jnuinSubA Suopd >{BAuBq idBjsj (%z^'6^) i^Sup SubA UBjisBqisqa^j sssjuasiad
1/61 N UBP (6) 13RM nl!B^ 'bAujbij ]BqBi35| usSusp n^hq Subdb^j ubSubjisj^j
(UBn^|BIi3d |7 UBp) % g^ UBSusp IBdUJBS 0 BJBJUB>jBAuBq qiqai isbipbj usSuap idBjaj usqB] SubA (UBn^Bpsd u^p) %gg jbji>|3S BAusq
ojjio ui is>|3[3s UBp [BSBiaq SubA ubujbubi 'isbjpbj BduBX wruodsfoco j dBpsqjai UBqBiSubA ubujbubj ^^AuBq q;q3| UB>n;sBqSu3ui ^dBp luruods^^co j iBiqij Subjis is^aps u^SuapUB>pn[uB[!p jBJBsnj uibsb is^sps usSusp iSBUiquio^ip SubA sri^^ BpBd pBJJf [ iSBipsi sisop
uB^nfunusm wruods^^co j dBpBqjaj is^(3[3s (isBq B^BqB p|UB[d uBiJnSuad'iSBisusSdi d^qB] ts^pps undnsui sn|B>( dBqsj is^siss ^rqun isbjsusSsj
UB^fuainuaui uidd 00S usSusp iBduiBS |y iSBqussuo^ jb^SuiusoiisBJ3U3S3i uBnduiBuis^ UB^uamusui psj^ g u^Suap iBduiss iSBipBi sisop
(ISBJ3U3S3J undnBui sn|B>|) is^sps dBqBj BpBd ^9HI UBP 3I3I!H uB>fSuipuBqip iSSuqM'q3| 60-l iSBi3U383J UBndujBuis^ isBisuaSsj undnBUi sn|B>| dsqBi BpBd is^japs ubSusoSn|B>| ISBJ3U3S3J BABp UB^(UnjnU3UI JBdBp pBi}{ IBdlUBS ISBipBJ SISOp UB)B>fSuiU3d UB>|
-j^nfunusui qspusj Hd usp {V uBSuap is^spsip qBpps ipBd ^luouquis snp^i isBJSuaSay
I/Sui i vvi + (3/1) SIAI UBp [BSBJsq jjpq SubAUBJB^(Bi3d UB|isBqi3q3Jf uB^SuBps^ (%0l'0Z UBp 03'IZ) |SSuij jnqnq i)Bf sbpuba isBisusSsjUB|isBqj3q3Ji [/Sui ^'o uijbsz + i/Sui To WI + I/Sui g va + SMI UBp i/Sui z'o uojnzBjpiqi+ j/Suj g va + SVi eipaui BpBd [/Sui g'o uojnzBipiqj + j/Sui g va + SVi Bip^ujuB^jni|r|!p srqB^ B(iqBdB %og redBDuaui SBurq i3d jn^Suni^ qBfBQ sbpuba|Bmb Bipsuj UBSuap blubs SubA Bipsui Bp^d jnqn^qnsip snp^) B|iqBdB %og UBp oz ipefuaui
%OS"St' !JBP JBJfSuiuauj ops ubmbSu^^ u^p suBp^s^ sbpuba isbjsusSsj UBjisBqjsqsJi ub>|-^fnfunuauj Baipu^ iped snp^f isBjauaSaJ dBpsqas) Bipaiu isbplluoj pSBqjsq qaiBSusd
(puiBjs sbpuba)99 UBp '(si|!M sbpuba) ^g '(ojopuis sbpuba) 23 qBpps %os UBp g^ 93d u^Susp is>i3|3sBipsui BpBd 5JJ1BLUOS ouquia ib]ox ubSuu3>I3>j dBpBqjai is^aps >jrqun UBJjBunSip pdBp %oguep 92 93d (^^8'3) lauiBis sbpuba UBp (t'S'g) sijiav sbpuba usSuap UB^SuipuBqip B^iqBdB(81'^) ^B^UBq Suipd ^qBuios ouqiua ^niuaqtuatu ojopuis sbpuba 9t^^ B!p^ui BpBd BPB<1uB^^nfunuaiu 93d UBSuap is^apsip SubA ppps^ ^iuouqois ps bssblu is^jauaSaj |isbh
j/Sui ['1 dva iSBjpsuo^iBpBd qapiadip ainuJisjjBui ssunj BAu>[BAuBg SBuni UBuainuad UB^qBqaAuaoi j/SiuI ubp qiqaj isbjju3Suo>| UBSuap dVa usqEquiBuad UBJjSuspas 'uiBqq Subiusjj UBp !|iqiuaSSBunj BAu>(BAuBq UB^iBJjSuiuaiu pdBp i/Sui 1 pduiBS dV3 UBuaqoiad u
IX

xii
Skrining sel hibridoma penghasil McAb RS memperoleh tiga nomor koloni hibridomapotensial menghasilkan McAb tinggi dan stabil (RS-(l)-l CIO, RS-1(1)-1 F08, dan RS-(6)-2F09) yang dapat dijadikan sumber kloning untuk memperoleh koloni satu sel hibridomapenghasil McAb RS tinggi dan bereaksi spesifik. Telah diperoleh teknik baku untuk produksiantibodi monoklonal untuk deteksi dan identiflkasi RRSV dan RS.
Berdasarkan pola RIA bakteri hasil fusi terbagi 6 kelompok. Hasil karakterisasimenggunakan pola RIA menunjukkan bahwa bakteri hasil fusi memiliki karakter yangberbeda dengan parentalnya. Sampai saat ini telah diperoleh 142 isolat bakteri endofitikyang terdiri dari 95 isolat bakteri endofitik padi dan 47 isolat bakteri endofitik jagung. Hasilseleksi terhadap isolat bakteri endofitik diperoleh 5 isolat unggul bakteri endofitik padi dan 5isolat unggui bakteri endofitik jagung.
Pada percobaan lapang di Citayam, dosis inokulan jamur MA hanya berpengaruh terhadap tinggi tanaman pada minggu ke-10 dan ke-12. Tinggi tanaman cenderung menurundengan brertambahnya dosis inokulan yang diberikan. Dosis inokulan berpengaruh terhadap % kolonisasi akar, BK brangkasan, jumlah spora dalam tanah, tapi tidak berpengaruhterhadap BK pipilan dan BK tongkol. Baik pada pertumbuhan vegetatif maupun pada waktupanen, % kolonisasi akar meningkat dengan bertambahnya dosis inokulan yang diberikan.Demikian pula dengan jumlah spora, meningkat dengan bertambahnya dosis inokulan yangdiberikan.
Pada percobaan kamar kaca dosis inokulan berpengaruh terhadap % kolonisasi akar,jumlah spora dalam tanah, tinggi tanaman (hanya pada minggu ke-4), tapi tidak ber-
^pengaruh terhadap konsentrasi N, P, Mg, Mn daun tanaman jagung.Hasil penelitian biokonversi kacang-kacangan melalui teknologi fermentasi dan
enzimatik diperoleh 141 isolat bakteri asam laktat, 31 isolat di antaranya memiliki kemam-puan unggul dalam menghasilkan asam laktat. Selain itu, diperoleh lima isolat BAL yangmampu secara efektif memfermentasi susu kacang-kacangan. Hasil uji organoleptik, susukacang merah yang difermentasi oleh BAL KBB 3 lebih disukai dibandingkan denganyoghurt komersial pembanding.
Hasil ekstraksi xilan dari kulit kedelai diperoleh sekitar 3,2%, yaitu untuk 1 kg kulit arikedelai diperoleh 32 g xilan. Formulasi media terbaik untuk produksi xilanase dari isolatbakteri AIII-5 dengan sumber xilan dari kulit kedelai ialah polipepton 0,1%, ekstrak khamir0,3%, dan xilan 1%.
Kegiatan pemasyarakatan hasil penelitian memegang peranan penting dalam menjem-batani antara penghasil teknologi, dalam hal ini Balitbiogen dengan pengguna. Pada TA2002, Balitbiogen menerbitkan Buletin Agrobio Vol. 5 No. 1 dan No. 2, Warta Balitbio nomor18, 19, dan 20, Laporan Tahunan 2001, Prosiding Seminar Hasil Penelitian Rintisan danBioteknologi Tanaman, dan Leaflet. Balitbiogen juga menyelenggarakan Apresiasi ProgramPenelitian Bioteknologi dan Sumber Daya Genetik Pertanian, Pameran dan Debat PendapatBioteknologi, Presentasi Rutin, serta Komersialisasi Teknologi.

logi untuk mewujudkan pertanian yangtangguh. Hasil penelitian bioteknologi dinegara-negara maju dan berbagai lembaga
penelitian intemasional menunjukkan bah-wa kendaia yang selama ini sulit atau bahkan mustahil dipecahkan secara konven-sional dapat diatasi menggunakan bioteknologi. Dengan memanfaatkan bioteknologi, program pemuliaan untuk mendapat-kan jenis unggul dapat dipercepat danlebih terarah. Selain itu, melalui rekayasagenetik dapat diperoleh jenis tanaman ataukomoditas pertanian lain yang mempunyaipotensi genetik lebih baik. Teknik kulturjaringan dapat dimanfaatkan untuk meng-hasilkan bibit tanaman dalam jumlah yangbanyak dalam waktu yang cepat dan bebaspenyakit. Pemanfaatan mikroba dapatmenghasilkan pupuk dan pestisida biologiyang lebih ramah lingkungan, probiotik,vaksin, dan hormon. Selain itu, mikroba da
pat dimanfaatkan sebagi perombak limbahpertanian (dekomposer), agen bioremedia-si, untuk bioproses yang dapat meningkat-kan nilai tambah, dan dapat digunakan un-.tuk memanfaatkan limbah menjadi produkyang bermanfaat sekaligus dapat meng-atasi masalah pencemaran lingkunganoleh limbah. Keuntungan ekonominasional Amerika Serikat bagi petani kapastransgenik pada tahun 1999 mencapai99 juta dolar Amerika Serikat.
Pertanian tangguh yang menjadi sasar-an pembangunan pertanian di Indonesiadapat dijabarkan sebagai pertanian yangmampu memenuhi permintaan konsumen,
dapat menjamin pendapatan dan kesejah-teraan petani secara berkelanjutan dan ti-dak merusak lingkungan hidup. Untuk me-wujudkan pertanian tangguh tersebut, tan-tangan dan kendaia yang dihadapi semakinberat dan kompleks. Hal ini disebabkan an-tara lain (1) permintaan produk pertaniansebagai bahan pangan, pakan, dan bahanbaku industri terus meningkat sejalan de-ngan peningkatan jumlah penduduk danpengembangan berbagai macam industri,(2) terjadi pergeseran area pertanian danlahan yang subur ke lahan yang kurang su-bur dan marjinal sebagai alih fungsi lahanpertanian subur dan pembukaan area pertanian baru pada lahan yang kurang suburdan marjinal, (3) tuntutan konsumen ter-hadap mutu hasil pertanian terns meningkat, bahkan beberapa komoditas tanaman,temak, dan ikan menghendaki beberapamacam mutu yang spesifik penggunaan-nya, (4) sistem pasar yang semakin terbukamengharuskan komoditas yang bersang-kutan mempunyai daya saing yang kuat,yang berarti menghendaki sistem usahata-ni yang efisien, dan (5) globalisasi yangberpengaruh antara lain meningkatnya per-
saingan, sehingga kualitas hasil pertanianyang dipersyaratkan oleh pasar menjadisemakin kompleks dan mendasar.
Tantangan tersebut tidak dapat diatasidengan teknologi kovensional saja. Biotek-nologi yang sedang berkembang denganpesat menjanjikan dukungan yang sangatdiperlukan dalam pengembangan tekno
I. Pendahuluan
Laporan Tahunan Balitbiogen 2002© Copyright 2003, Balitbiogen

kondusif untuk pengembangan bioteknologi merupakan salah satu acuan dalampenyusunan program pengembangan dan
penelitian bioteknologi.Balitbiogen menetapkan program
utama sebagai berikut:1.Biologi Molekuler yang mencakup ke-
giatan rekayasa genetika, identifikasimarkah molekuler untuk sifat-sifat pen-ting dan pemanfaatannya untuk pemu-
liaan dan deteksi dini hama dan penya-kit, karakterisasi molekuler tanamandan hewan.
2.Kultur in vitro yang mencakup kegiatanseleksi in vitro untuk ketahanan terha-dap cekaman biotik, konservasi in vitroplasma nutfah, mikro propagasi, penye-lamatan embrio, kultur anter, embriotransfer, inseminasi buatan.
3.Bioproses yang mencakup kegiatan penelitian biofertilizer, biopestisida, biode-komposer, vaksin, probiotik, dan biore-midiasi.
4.Bioprospeksi yang mencakup kegiatankoleksi, penelitian dan pemanfaatansumberdaya genetik dan biologi gunamendapatkan sumber-sumber baru se-nyawa kimia, gen, organisme, dan pro
duk alamiah lain untuk tujuan ilmiahdan atau komersial.
5.Pengelolaan dan pemanfaatan plasma
nutfah.
6.Bioinformatika.
7.Analisis risiko keamanan hayati produkrekayasa genetika.
Matriks penetapan prioritas programBalitbiogen 2002-2010 disajikan pada TabelII. 1. Sedangkan untuk menunjang keberha-
• Balai Penelitian Bioteknologi dan Sum-berdaya Genetik Pertanian (Balitbiogen)berdasarkan Surat Keputusan MenteriPertanian Nomor 78/Kpts/OT.210/1/2002,mempunyai fungsi melaksanakan penelitian konservasi, karakterisasi, dan biologi
molekuler sumberdaya genetik pertanian;bioteknologi sel, jaringan, rekayasa genetik, dan teknologi bioproses sumberdayagenetik pertanian; penelitian analisis risikolingkungan serta keamanan hayati dan pa-
ngan produk bioteknologi; mengembang-kan sistem bioinformatika sumberdaya ge-
•netik pertanian; dan melaksanakan penelitian komponen teknologi sistem, dan usahaagribisnis produk bioteknologi pertanian.
Berdasarkan mandat tersebut, dalammerumuskan prioritas penelitian bioteknologi pertanian harus mempertimbangkankeungguian dan daya saing produk di pasarintemasional dan nasional dibandingkandengan teknologi yang sedang berjalan.Penelitian akan diprioritaskan pada keter-sediaan teknologi, fasilitas kerja sama ke-
lembagaan di tingkat nasional dan intemasional.
Program bioteknologi sampai dengantahun 2010 disusun berdasarkan tantanganyang diperkirakan akan muncul, peluangyang tersedia bagi pembangunan pertanian, sumberdaya, dan fasilitas yang ada diBalitbiogen serta perkembangan ilmu danteknologi yang ada. Secara garis besar, fak-tor yang mempengaruhi penyusunan priori
tas program dapat dikelompokkan menjadifaktor eksternal dan internal.
Berbagai masalah yang dihadapidalam pembangunan pertanian, kebijakanpemerintah, dan faktor lingkungan yang
II. Program Penelitian
Laporan Tahunan Balitbiogen 2002© Copyright 2003, Balitbiogen

Pangan, sayuran, dan pakan ternakPangan dan sayuranPangan dan sayuran
Pangan dan sayuranPangan dan sayuran
Pangan dan sayuranPangan dan sayuran
1.Tanaman transgenik tahan hama atau penyakit2.Markah untuk sifat penting dan kit diagnostik untuk penyakit, galur
harapan tahan cekaman biotik atau abiotik3.Tanaman tahan hasil seleksi in vitro, kultur anter, dan penyelamatan Padi, kedelai, dan kacang hijau
embrio4.Biofertilizer, biodekomposer, dan biopestisida5.Gen dan sumber baru senyawa kimia6.Tambahan koleksi plasma nutfah dan plasma nutfah yang sudah
terkarakterisasi serta bank gen7.Database plasma nutfah dan genom8.Teknik analisis risiko keamanan hayati dan produk bioteknologi yang
dinyatakan aman
KomoditasNo. Rencana output
Tabel 11.1. Matriks penetapan prioritas pogram Balitbiogen 2002-2010
efisiensi dan keberhasilan pencapaiantujuan.
6.Memanfaatkan fasilitas laboratoriumseefisien mungkin dengan prioritas untuk penelitian unggulan, serta mengko-
mersialkan fasilitas yang ada.7.Meningkatkan keterampilan dan penge-
tahuan operator generator manual se-
hingga apabila aliran listrik PLN terputusdapat segera menghidupkan generator.Pasokan listrik dari generator dipriori-taskan untuk bank gen dan penelitianunggulan.
8.Dalam keadaan dana perawatan per-alatan yang sangat terbatas, maka akan
disusun prioritas peralatan yang harusmendapat perawatan atau perbaikan.
Penyusunan prioritas perawatan peralatan akan ditentukan berdasarkanpertemuan antara Pimpinan, KetuaKelti, Kepala Laboratorium, dan KepalaSeksi Pelayanan Teknis.
9.Meningkatkan motivasi peneliti melaluipemberian penghargaan kepada peneliti yang berprestasi. Penghargaan dapatberupa kesempatan mengikuti pelatih-an jangka pendek atau panjang, memberikan kesempatan mengikuti seminar
silan pelaksanaan program tersebut Balit-biogen akan melakukan langkah-langkahsebagai berikut:1.Memberikan prioritas penelitian pada
komoditas yang mempunyai nilai stra-tegis dan nilai komersial tinggi dan padabidang yang sumberdaya manusianyapaling kuat dan hasilnya akan memberikan manfaat yang tinggi.
2.Memaksimalkan pemanfatan FasilitasUji Terbatas untuk kegiatan penelitiantanaman transgenik, analisis risiko ke-amanan hayati produk bioteknologi,dan untuk komersialisasi dalam rangkapenggalangan dana penelitian.
3.Menjalin kerja sama dengan lembagainternasional yang memiliki keahlianyang dapat kita manfaatkan, memilikigen dan markah untuk kegiatan penelitian biologi molekuler.
4.Menjaring dana penelitian dari lembagapenelitian intemasional yang memilikiprogram yang sejenis dan memanfaat-kannya untuk meningkatkan pengeta-huan peneliti.
5.Menjalin kerja sama dan melakukankoordinasi dengan lembaga penelitiannasional dalam rangka meningkatkan
Laporan Tahunan Balitbio 2002

C.Pengembangan teknik produksi danaplikasi antibodi monoklonal untukdeteksi dan identifikasi patogen tum-buhan (3 kegiatan)1.Evaluasi lapangan perangkat ELISA
dengan antibodi poliklonal (PAb)untuk deteksi dini dan identifikasiRRSV, PStV, dan Ralstonia solanace-arum (/?5)
2.Teknik produksi antibodi monoklonal (McAb) untuk deteksi dan identifikasi virus kerdil hampa padi (RRSV)
3.Teknik produksi antibodi monoklonal (McAb) untuk deteksi dan identifikasi bakteri Ralstonia solanace-arum (RS)
D.Biokonversi kacang-kacangan melaluiteknologi fermentasi dan enzimatik (3kegiatan)1.Seleksi dan karakterisasi bakteri
asam laktat (BAL) Indigenus2.Efektifitas bakteri asam laktat (BAL)
dalam pembuatan produk fermentasi berbasis protein/susu nabati
3.Produksi xilanase untuk biokonversilimbah kacang-kacangan
E.Karakterisasi molekuler dan kloning gencry dari isolat bacillus thuringiensis lokaluntuk pengendalian hama (3 kegiatan)1.Konfirmasi virulensi beberapa isolat
Bt lokal yang mengandung gen cryterhadap hama tanaman
2.Ekstraksi DNA Bacillus thuringiensisisolat lokal yang mengandung gencry dan pembuatan pustaka plasmidBt
3.Isolasi dan karakterisasi Bacillusthuringiensis isolat lokal untuk bio-insektisida lalat Chrysomya bezzianapenyebab myiasis
sepanjang tersedia sponsor atau berupamateri. Selain itu, akan diberikan pem-
binaan khusus pada peneliti yang moti-vasinya rendah.
"Pada tahun anggaran 2002, penelitianyang dibiayai oleh APBN adalah 14 proposal penelitian yang dalam pelaksanaannyaterdiri dari 67 kegiatan penelitian, yaitu
A.Teknologi pengembangan pupuk hayatidan perombak bahan organik untuk la-han kering dan lahan sawah (4 kegiatan)1.Fusi protoplas intraspesies antar
Bradyrhizobium japonicum2.Peningkatan daya kompetisi rhizo-
bium yang diberi penginduksi gennod pada berbagai varietas kedelaidi lahan masam
3.Isolasi dan seleksi mikroba diazotrof,endofitik dan penghasil zat pemacu
tumbuh pada tanaman padi danjagung
4.Evaluasi produksi inokulan mikofos-fat skala pilot dan uji keefektifannya
B.Peningkatan efektifitas biopestisida untuk pengendalian hama (4 kegiatan)1.Pengembangan teknik multiplikasi
nematoda patogen serangga padamedia cair dalam fermentor skala 10liter
2.Pengembangan media selektif untukstabilisasi fase primer bakteri sim-bion nematoda patogen serangga(NPS)
3.Puriflkasi dan karakterisasi toksinbakteri simbion nematoda patogenserangga (NPS)
4.Pengaruh sinar ultra violet terhadapvirulensi isolat NPV yang menginfek-si ulatgrayak pada kedelai
Program Penelitian

5. Induksi dan multiplikasi tunas ken-tang hitam dan gembili untuk penyimpanan secara kultur in vitro
H. Konservasi sumber daya genetik mikro-
ba pertanian (6 kegiatan)1.Koleksi, karakterisasi dan preservasi
mikroba penyubur tanah dan pe-
rombak bahan organik2.Koleksi dan isolasi mikroba hidrolitik
untuk perbaikan mutu pangan3.Koleksi dan penyimpanan mikroba
bioremediasi4.Seleksi dan karakterisasi mikroba
patogen5.Seleksi dan karakterisasi mikroba
pengendali hama dan penyakit6.Penyimpanan, pemeliharaan, dan
penyusunan database sumberdayagenetik mikroba pertanian
I. Pemanfaatan teknik kultur jaringan danmolekuler untuk perbaikan genetik padidengan menggunakan spesies liar (3kegiatan)1.Evaluasi dan karakterisasi ketahanan
spesies padi liar terhadap cekamanbiotik dan abiotik dengan menggunakan kultur iri vitro dan markahmolekuler
2.Pembentukan populasi interspesifikpadi melalui kultur embrio secara invitro
3.Perbaikan galur mandul jantan dangalur pemulih kesuburan melaluikultur anter
J. Perakitan tanaman transgenik tahan hama atau penyakit tanaman (12 kegiatan)1.Transformasi padi Japonica (T-309)
dan Indica dengan gen crylA2.Analisis molekuler lanjutan tanaman
putatif transgenik padi gen crylAgenerasi Tl dan T2
F.Pelestarian, karakterisasi, dan evaluasiplasma nutfah tanaman (7 kegiatan)1.Rejuvenasi, karakterisasi morfologi,
dan mutu gizi plasma nutfah tanaman pangan
2.Pengembangan database plasmanutfah tanaman pangan
3.Eksplorasi plasma nutfah tanamanpangan
4.Evaluasi toleransi plasma nutfah pa-di, jagung, dan kedelai terhadap la-han bermasalah (kekeringan, lahanmasam, keracunan Al dan Fe)
5.Evaluasi ketahanan plasma nutfahtanaman pangan terhadap penyakithawar daun bakteri, bias, dan tungropada padi; bulai pada jagung; em-bun tepung pada kacang hijau; sapusetan pada kacang tanah; dan viruskerdil(SSV) pada kedelai
6.Evaluasi ketahanan plasma nutfahtanaman pangan terhadap hamawereng coklat pada padi, lalat bibitpada jagung, dan hama lanas padaubi jalar
7.Karakterisasi plasma nutfah tanamanpangan secara molekuler
G.Studi regenerasi dan penyimpanan tanaman bernilai ekonomi tinggi secarakultur in vitro (5 kegiatan)1.Inisiasi akar pada tanaman melinjo
melalui kultur in vitro2.Optimasi sistem perakaran dan akli-
matisasi iles-iles (Amorpophalus sp.)3.Inisiasi akar manggis dari tunas in
vitro4.Pengaruh beberapa krioprotektan
terhadap keberhasilan penyimpananubi-ubian secara kriopreservasi
Laporan Tahunan Balitbio 2002

5. Penyelamatan embrio hasil persi-langan kacang hijau dengan kerabatliamya
L. Identiflkasi dan aplikasi markah molekuler untuk pemuliaan tanaman (9 kegiatan)1.Pengujian sifat toleransi tanaman pa
di terhadap kekeringan pada popu-lasiF7(IR64xIRAT112)
2.Uji rumah kaca untuk toleransi terhadap kekeringan pada populasi F7(IR64xlRAT112)
3.Analisis AFLP dan mikrosatelit untukpemetaan markah molekuler untuksifat toleransi terhadap kekeringan
4.Analisis segregasi populasi F2 (Dupax ITA131) dan uji fenotipik populasiF3
5.Identiflkasi markah molekuler untukketahanan terhadap bias
6.Analisis populasi inbrida rekombinandengan beberapa isolat bias di rumah kaca (monosiklik dan polisiklik)
7.Analisis segregasi populasi inbridarekombinan di lapang
8.Markah DNA untuk deteksi dinipenyakit bulai pada tanaman jagung
9.Markah DNA untuk deteksi dinipenyakit bulai pada benih yang sakit
M. Rekayasa genetik untuk perakitantanaman hortikultura tahan terhadaphama utama dan peningkatan kualitashasil (3 kegiatan)1.Penyisipan gen GUS pada bawang
merah dan tomat2.Perbaikan regenerasi pasca transfor
masi dengan gen pelapor GUS(penyisipan gen GUS pada tomat)
3.Transformasi cDNA gene delayedripening ke vektor ekspresi Agrobacterium tumefaciens LBA-4404
3.Bioasai lanjutan tanaman putatiftransgenik padi crylA generasi TldanT2
4.Analisis molekuler lanjutan gen pinllpada tanaman kedelai transgenik R3dan R4
5.Bioasai tanaman kedelai transgenikpinll terhadap hama penggerek po-long (Etiella zinckenella, Treitschke)
6.Transformasi kedelai dengan gencrylA melalui metode particlebombardment
7.Analisis molekuler gen CP-PStV padatanaman kacang tanah transgenik
8.Bioasai tanaman kacang tanah transgenik terhadap virus bilur kacangtanah (PStV)
9.Transformasi ubi jalar dengan genpinll dan gen CP-SPFMV melaluiAgrobacterium tumefaciens
10.Analisis molekuler integrasi ^en pinllpada ubi jalar
ll.Pengujian tanaman ubi jalar pinllsecara bioasai
12.Rekonstruksi gen a-amilase inhibitorpada plasmid biner
K. Pengembangan metode seleksi in vitrodan penyelamatan embrio untuk me-
ningkatkan toleransi tanaman terhadapfaktor abiotik dan biotik (5 kegiatan)1.Regenerasi massa sel embrionik
kedelai yang diseleksi dengan PEG2.Pengaruh berbagai formulasi media
terhadap regenerasi kalus padiIndica
3.Regenerasi kalus embriogenik padisetelah seleksi in vitro dengan Al danpH rendah
4.Pengujian planlet abaka hasil seleksisilang terhadap F. oxysporum
Program Penelitian

N. Analisis dan pendugaan risiko peman-faatan tanaman transgenik
Oi samping proposed penelitian yangdidanai oleh APBN, di Balitbiogen ada tigaproposal yang didanai oleh RUT, yaitu
1.Pembentukan buah partenokarpi padatanaman salak melalui rekayasagenetika.
2.Peningkatan ketahanan terhadap aluminium pada tanaman kedelai melaluikultur in vitro
3.Reduksi pencemaran lahan pertanianoleh senyawa logam berat denganteknologi bioremediasi
Laporan Tahunan Balitbio 2002

tif akan mengefektifkan serta mengefisien-kan pemanfaatannya.
Rejuvenasi, Karakterisasi Morfologi, danMutu Gizi Plasma Nutfah Tanaman
Pangan
Padi Budi Daya
Hasil pengamatan 500 aksesi padi diPusakanegara dan 200 aksesi di Muaramenunjukkan bahwa sifat tanaman sangatbervariasi. Tinggi tanaman berkisar antara64,2-210,4 cm; anakan antara 4-34; panjangmalai antara 16,5-51,2 cm; dan bobot 1000butir antara 10,4-42,0 g. Beberapa varietasmempunyai jumlah anakan sedikit sepertivarietas Rantai Mas (Reg. 5348), Tunjung(Reg. 5660), Genjah Pare (Reg. 20610), danMelawai (Reg. 30380), sedangkan varietasyang mempunyai anakan banyak (>30anakan), yaitu varietas Betonan (Reg.6059).
Sebanyak 175 aksesi mempunyai panjang malai lebih dari 30 cm dan 96 aksesimempunyai bobot 1000 butir >30 g. Varietas yang mempunyai malai panjang danbutir banyak umumnya mempunyai bobot1000 butir rendah dan sebaliknya varietasdengan bobot 1000 butir lebih berat padaumumnya mempunyai butir isi <200 butir.Beberapa varietas dengan malai >30 cmdan bobot butir >.3O g disajikan pada TabelIII.l. Varietas yang mempunyai malai ter-panjang adalah Hawara Kaos (Reg. 5324),sedangkan varietas yang memiliki bobot1000 butir terberat adalah Kalimutu(R. 20628).
' PELESTARIAN, KARAKTERISASI, DANEVALUASI PLASMA NUTFAH TANAMAN
Ida H. Somantri, Tiur S. Silitonga, Sri G. Budiarti,Sri A. Rais, Nani Zuraida, Minantyorini,
Tintin Suhartini, Hadiatmi, Lukman Hakim,Mamik Setyowati, Sutoro, Asadi, Nurwita Dewi,Hakim Kurniawan, Dodin Koswanudin, Suyono,
M. Machmud, Trijoko Santoso,A. Dinar Ambarwati, Aniversari Apriana, dan
Atmitri Sisharmini
Dengan semakin berkembangnyapenggunaan varietas baru oleh petani, ma-ka varietas lokal Qandraces) akan terdesakdan tidak mustahii akan musnah. Sebelumteriambat, landraces dan varietas liarnyaperlu diselamatkan melalui eksplorasi dandilestarikan dalam bank gen.
Pelestarian dengan cara rejuvenasidan penyimpanan dengan baik dan benarsangat perlu dilakukan dan harus menda-patkan perhatian. Namun demikian, pelestarian tanpa diberdayakan dengan caraidentifikasi sifat-sifatnya tidak banyak man-faatnya. Oleh sebab itu, perlu ada identifikasi sifat-sifat yang dimiliki oleh plasmanutfah tersebut. Identifikasi yang berupakarakterisasi molekuler atau konvensionalseperti sifat morfologi, agronomi, dan mutugizi serta evaluasi terhadap cekaman biotikdan abiotik perlu dilakukan. Informasi tersebut dapat digunakan untuk mengetahuiprofil keragaman dan koleksi yang kitamiliki. Di samping itu, perlu melakukaneksplorasi plasma nutfah untuk memper-kaya koleksi dan menyelamatkan sumberdaya genetik yang semakin berkurang aki-bat pembangunan atau bencana alam.Selanjutnya sistem pengelolaan data plasma nutfah yang baik, lengkap, dan informa-
III. Hasil Penelitian 2002
Laporan Tahunan Balitfoiogen 2002© Copyright 2003, Balitbiogen
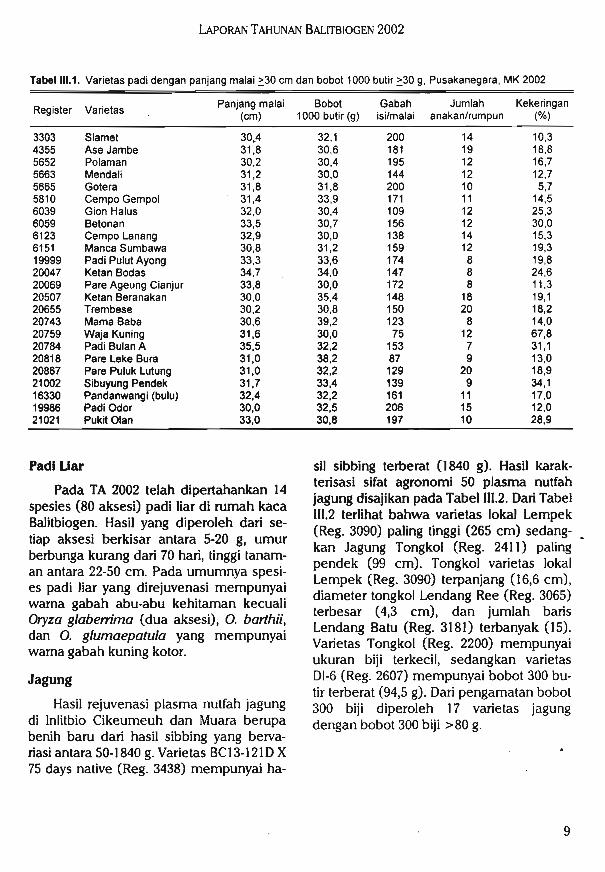
sil sibbing terberat (1840 g). Hasil karak-terisasi sifat agronomi 50 plasma nutfahjagung disajikan pada Tabel III.2. Dari TabelIII.2 terlihat bahwa varietas lokal Lempek(Reg. 3090) paling tinggi (265 cm) sedang-kan Jagung Tongkol (Reg. 2411) palingpendek (99 cm). Tongkol varietas lokalLempek (Reg. 3090) terpanjang (16,6 cm),diameter tongkol Lendang Ree (Reg. 3065)terbesar (4,3 cm), dan jumlah barisLendang Batu (Reg. 3181) terbanyak (15).Varietas Tongkol (Reg. 2200) mempunyaiukuran biji terkecil, sedangkan varietasDI-6 (Reg. 2607) mempunyai bobot 300 butir terberat (94,5 g). Dari pengamatan bobot300 biji diperoleh 17 varietas jagungdengan bobot 300 biji >80 g.
Padi Liar
Pada TA 2002 telah dipertahankan 14spesies (80 aksesi) padi liar di rumah kacaBalitbiogen. Hasil yang diperoleh dari se-tiap aksesi berkisar antara 5-20 g, umurberbunga kurang dari 70 hari, tinggi tanam-an antara 22-50 cm. Pada umumnya spesi
es padi liar yang direjuvenasi mempunyaiwarna gabah abu-abu kehitaman kecuali
Oryza glaberrima (dua aksesi), O. barthii,dan O. glumaepatula yang mempunyaiwama gabah kuning kotor.
Jagung
Hasil rejuvenasi plasma nutfah jagungdi Inlitbio Cikeumeuh dan Muara berupabenih baru dari hasil sibbing yang berva-riasi antara 50-1840 g. Varietas BC13-121D X75 days native (Reg. 3438) mempunyai ha
28,912,017,034,118,913,031,167,814,018,219,111,324,619,819,315,330,025,314,55,7
12,716,718,810,3
(%)Kekeringan
1015119
2097
128
2018
888
12141212111012121914
anakan/rumpunJumlah
19720616113912987
15375
123150148172147174159138156109171200144195181200
isi/malaiGabah
30,832,532,233,432,238,232,230,039,230,835,430,034,033,631,230,030,730,433,931,830,030,430,632,1
1000 butir (g)Bobot
33,030,032,431,731,031,035,531,630,630,230,033,834,733,330,832,933,532,031,431,831,230,231,830,4
(cm)Panjang malai
Pukit OlanPadi OdorPandanwangi (bulu)Sibuyung PendekPare Puluk LutungPare Leke BuraPadi Bulan AWaja KuningMama BabaTrembeseKetan BeranakanPare Ageung CianjurKetan BodasPadi Pulut AyongManca SumbawaCempo LanangBetonanGion HalusCempo GempolGoteraMendaliPolamanAse JambeSlamet
Varietas
21021199861633021002208672081820784207592074320655205072006920047199996151612360596039581056655663565243553303
Register
Tabel III.1. Varietas padi dengan panjang malai >30 cm dan bobot 1000 butir >30 g, Pusakanegara, MK 2002
Laporan Tahunan Balitbiogen 2002

10
Coklat (0,9%), putih (99,1%)LB 72 (77)Ryocoh (28)Kuning, hitam, hijau, coklat
Lok Ngawi (29), Sindoro (30), Sumbawa (30)Ungu (6,8%), putih (93,2%)G.2120 (83), Otak (83), Mlg 3017 (83), Clark (83)
Umur berbunga (HST)29-57Warna bungaUmur masak (HST)83-92Tinggi tanaman (cm)17,8-69,4Banyaknya cabang1-5Warna buluPolong isi/tanaman9-77Bobot 100 biji (g)5,1-28Warna biji
Kisaran KeteranganKarakter
Tabel III.3. Karakterisasi plasma nutfah kedelai, Balitbiogen MT 2002
2411 (99) - 3090 (265)2124 (39,5)-3090 (113)Hijau muda, hijau tua, hijau keunguanPutih, keunguanKuning, krem, kuning kemerahan, coklat, unguKuning, kuning kemerahan, putih, campurMutiara, semi mutiara, semi gigi kuda2200, 2473 (9,4) - 3090 (16,6)2473 (2,9) - 3188, 3181, 3065 (4,3)1991,2006, 2022 (10)-3181 (15)3057 (50) - 3223 (96)2411 (6)-3188 (9,8)1991, 2022 (4) - 2435, 3065 (6)2124 (19,6)-3199 (4,1)3209 (2,4)-3323 (10,5)2124 (7)-3247 (19)2200 (43,4) - 2607 (94,5)43,3-94,5
7-192,4-10,519,6-41
4-66-9,850-9610-15
2,9-4,39,4-16,6
--
--
39,5-11399-265
Bobot 300 butir (g)Banyaknya cabang malaiPanjang tangkai malaiPanjang malai (cm)Banyaknya daunLebar daunPanjang daun (cm)Banyaknya barisDiameter tongkolPanjang tongkolTipe bijiWarna bijiWarna sekam dan rambutWarna urat pusatWarna daun, batang, dan pelepahTinggi tongkol (cm)Tinggi tanaman (cm)
Kisaran KeteranganKarakter
Tabel III.2. Keragaman 50 plasma nutfah jagung, Inlitbio Cikeumeuh 2002
Ubi Kayu
Hasil karakterisasi menunjukkan keragaman pada beberapa sifat morfologi se-perti warna urat daun, tangkai daun, pupus
daun, jumlah lobus daun maupun panjangdan lebar daun. Helai daun didominasioleh warna hijau. Pupus daun berwarna hijau, coklat muda, coklat, coklat tua, dan hijau kecoklatan. Urat daun atas dan bawahberwarna merah, merah muda, hijau, hijaumuda, dan kombinasi hijau dan merah, se-dangkan pusat urat daun berwarna merah,
hijau, dan merah muda. Tangkai daun atasdan bawah berwarna merah, hijau, merahmuda, merah tua, dan kombinasi merah
dan hijau. Panjang tangkai daun berkisar
Kedelai
Hasil karakterisasi 500 aksesi kedelaimenunjukkan keragaman sifat morfologiyang diamati. Terdapat sembilan aksesiberbiji besar dengan bobot 100 biji >10,5 g,di antaranya adalah Ryocoh, Manchuria,17/9/3/8/0, Kipas Putih, Slamet, AVRDC-G2120, GM 409, Kerinci/Dempo, dan Orba.Pada umumnya plasma nutfah kedelai ber-umur sedang sampai dalam. Umur berbu-nga berkisar antara 29-57 hari, umur masak
90% antara 83-92 hari, tinggi tanaman berkisar antara 17,8-69,4 cm, polong isi/tanam-
an 9-77, dan bobot 100 biji antara 5,1-28 g,warna biji bervariasi, yaitu kuning, hitam,hijau, dan coklat (Tabel 1II.3).
Hasil Penelitian 2002

11
C40danC33(1), V132(149)C40 (12), V167 (2624,5)C37 (4,8), C25 (42,9)C37 (3,8), C33 (35)C37 (4,9), C28 (24)C3 (53,5), C10(88,7)Thasos(99), C46(118)
H80(102), V182(115)H71 (591,7), H23 (1016,8)H1 (3,4), H 18 (4,4)H87 (27,4), ina (34,9)V170(6,9), H99(8,1)H85 (6), H99(9)V236 (62,3), H71 (77.5)H40(87), H99(107)H40 (48), H99 (64)
Keterangan
1-14912-2624,54,8-42,93,8-354,9-24
53,5-88,799-118
102-115591,7-1016,8
3,4-4,427,4-34,96,9-8,1
6-962-77
87-10748-64
Kisaran
Banyaknya rumpun panenHasil panen/petak (g)Hasil biji/rumpun (g)Banyaknya malai/rumpun.Banyaknya anakan produktifTinggi tanaman (cm)Umur panen (hari)
Koleksi Balitbiogen dan IPBBanyaknya rumpun panen/petakHasil panen/petak (3 m2) (g)Bobot 100 butir(g)Banyaknya butir/malaiPanjang malai (cm)Banyaknya malai/rumpunTinggi tanaman (cm)Umur panen (hari)Umur berbunga (hari)
Koleksi Balitbiogen
Karakter
Tabel 111.4. Keragaman plasma nutfah terigu, IP Kuningan, MK 2002
Hasil karakterisasi plasma nutfahterigu koleksi Balitbiogen dan IPB memilikiumur panen berkisar antara 99-118 hari,tinggi tanaman antara 53,5-88,7 cm, jumlahrumpun panen per petak berkisar antara 1 -
149, jumlah anakan produktif berkisar antara 4,9-24, banyaknya malai per tanamanantara 3,8-35, dan hasil panen per petakberkisar antara 12-2624,5 g (Tabel III.4).Varietas C3 merupakan varietas terpendek,sedangkan C10 paling tinggi. Varietas Cllmempunyai wama malai kuning keemas-an, sedangkan C8 mempunyai warna malai
kehijauan pendek/kecil. Varietas C46 danC47, mempunyai batang hijau keperakan.Enam aksesi (Sw Triso, V162, V219, VI61,C4, dan C5) mempunyai hasil biji 1 kgsedangkan 2 aksesi (VI92 dan VI67) mempunyai hasil biji >2,5 kg.
Sorgum
Enam aksesi sorgum (1CSR 70, ICSR91026, A.226.72, ICSV 93024, Sorgum Lao,dan Lok. Kaltim No. 14) mempunyai hasilbiji kering tinggi dengan bobot biji permalai >80 g. Hasil biji kering per malai ber-
antara 10,8-24,5 cm. Jumlah lobus daunbervariasi antara 5, 7, dan 9 lobus denganpanjang lobus daun antara 6,5-21,2 cm danlebar lobus daun antara 1,4-4,5 cm. Tinggitanaman bervariasi antara 1,3-2,4 m.Banyaknya umbi antara 1-11 umbi dan bo-bot umbi antara 0,3-3,9 kg. Diperoleh tujuh
aksesi ubi kayu yang mempunyai hasil umbi per pohon >3,0 kg, yaitu Gm-3, Sm 1565-19, Cmc 33-38-4, Kl, No. 726, No. 247, dan
Vandemix.
Terigu
Hasil karakterisasi plasma nutfah terigu koleksi Balitbiogen menunjukkan bah-wa varietas H40 mempunyai umur berbu-nga paling genjah (48 hari) dan umur ma-sak paling cepat (87 hari). Varietas H99mempunyai umur berbunga paling lambat(64 hari) dan umur masak paling dalam(107 hari). Walaupun umurnya paling lambat, varietas H99 mempunyai malai pertanaman terbanyak (8,6) dan malai terpan-jang (8,1 cm). Hasil biji per petak bervariasiantara 591,7-1016,8 g dan jumlah rumpunparien berkisar antara 102-115 (Tabel 111.4).
Laporan Tahunan Balitbiogen 2002
12
Keris,TUB7, CK2, Hegari GenjahIrat 204, ICSV 89034, Demak 3Cantel Abrit Wonogiri, Red OchuliMandau, Lepang, Sil.75, Butter Krek 4
ICSB 31, ICSV 89037K905, Irat 204, ICSV-LM-90501ICSB 11,Sangkur, No. 14Badik, Gadam Human
ICSR 70, Demak 5, KolotCantel A. Wonogiri, KerisNeam Reket, Selayer 3, Red OchuliButter Bebelit 2, Selayer 2, KSB II1,4
17,824,556,3
18,334,144,7
2,9
21,51,0
68,88,7
33751
117
3871936
452
14318
Pahit (skor 4)Tawar (skor 3)Sedang (skor 2)Manis (skor 1)
Rasa cairan batangHitamMerahCoklatKuning
Warna sekamCoklatMerahKuningPutih
Warna biji
Banyaknya aksesi Persentase (%) Contoh aksesiKarakter
Tabel III.6. Distribusi beberapa sifat kualitatif aksesi plasma nutfah sorgum
1 = manis, 2 = sedang, 3 = tawar
KuningMerahKuningCoklatKuningMerahKuningCoklatCoklatMerahCoklatMerah
374226763316264134062903
2,053,163,023,122,153,2
100,680,880,380,789,880,6
26,430,420,432,229,429,9
155156160171228346
10298108110108105
807382828380906 Sorgum Lao
ICSR70ICSR 91026A. 226.72ICSV 93024Lok. Kaitim No. 14
838861740798912
Biji Sekam
WarnaBanyaknya Rasa •
Biji per 100 biji biji per malai batangmalai (g) (g)
BobotPanjangmalai(cm)
Umur (hari)— Tinggi
Berbunga Masak (cm>No. reg. Aksesi
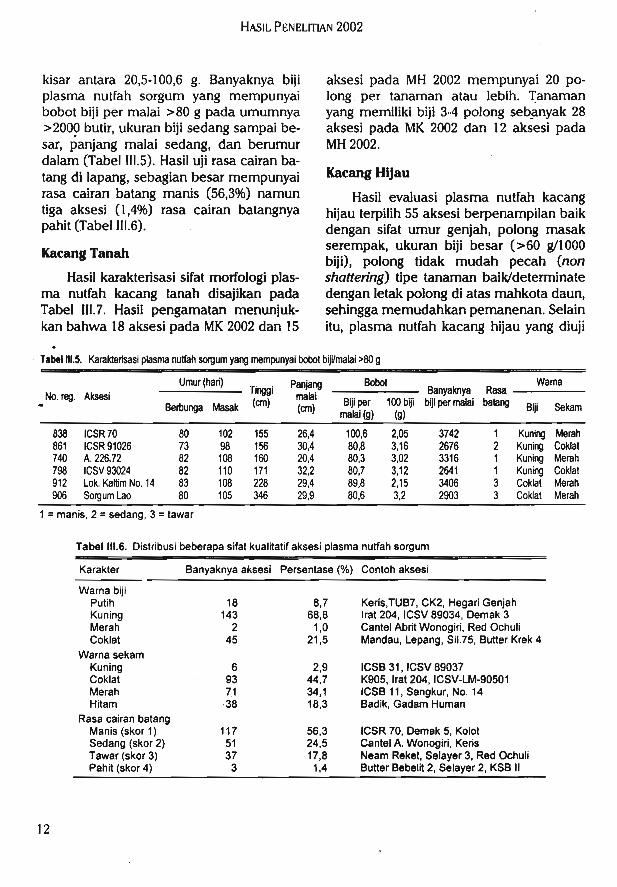
Tabel 111.5. Karakterisasi plasma nutfah sorgum yang mempunyai bobot biji/malai >80 g
aksesi pada MH 2002 mempunyai 20 po-long per tanaman atau lebih. Tanaman
yang memiliki biji 3-4 polong sebanyak 28aksesi pada MK 2002 dan 12 aksesi padaMH 2002.
Kacang Hijau
Hasil evaluasi plasma nutfah kacanghijau terpilih 55 aksesi berpenampilan baikdengan sifat umur genjah, polong masakserempak, ukuran biji besar (>60 g/1000biji), polong tidak mudah pecah {nonshattering) tipe tanaman baik/determinatedengan letak polong di atas mahkota daun,sehingga memudahkan pemanenan. Selainitu, plasma nutfah kacang hijau yang diuji
kisar antara 20,5-100,6 g. Banyaknya bijiplasma nutfah sorgum yang mempunyaibobot biji per malai > 80 g pada umumnya>2000 butir, ukuran biji sedang sampai be-sar, panjang malai sedang, dan berumurdalam (Tabel III.5). Hasil uji rasa cairan ba-tang di lapang, sebagian besar mempunyairasa cairan batang manis (56,3%) namuntiga aksesi (1,4%) rasa cairan batangnyapahit (Tabel III.6).
Kacang Tanah
Hasil karakterisasi sifat morfologi plasma nutfah kacang tanah disajikan padaTabel III.7. Hasil pengamatan menunjuk-kan bahwa 18 aksesi pada MK 2002 dan 15
Hasil Penelitian 2002
13
242,5060,00
9,5410,6446,5018,00604,026,4434,00
7,00233,5088,00
2,4039,002,674,268,303,005,005,891,003,00
18,0073,00
60,0047,00
5,417,55
15,4812,00
201,0010,9810,004,00
96,7783,00
Hasil rejuvenasi (g)Umur bunga (hari)Lebar daun (cm)Panjang daun (cm)Panjang polong (cm)Diameter polong (cm)Banyaknya biji/polongBobot 100 (g)Banyaknya polongBanyaknya cabangTinggi tanaman (cm)Umur masak (hari)
Rata-rata Minimum MaksimumPeubah
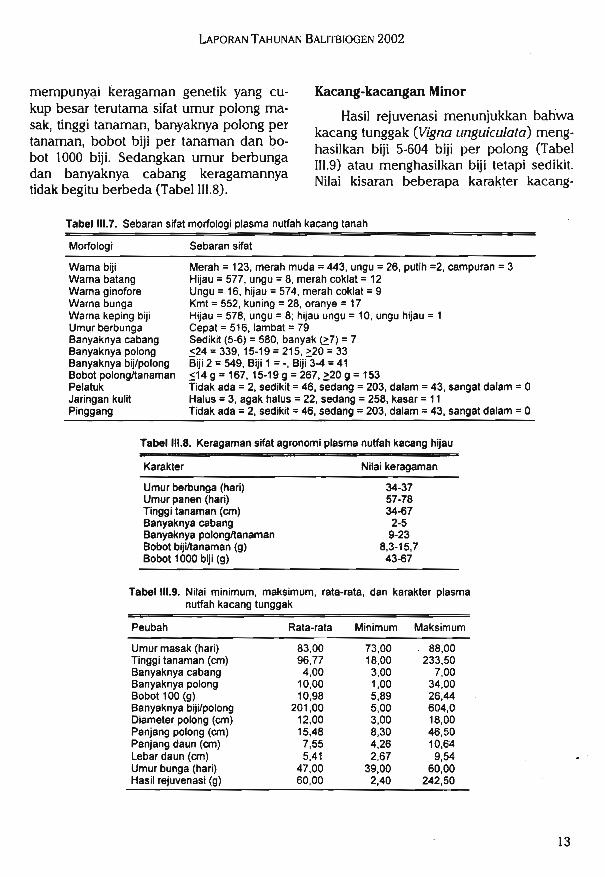
Tabel III.9. Nilai minimum, maksimum, rata-rata, dan karakter plasmanutfah kacang tunggak
34-3757-7834-672-59-23
8,3-15,743-67
Umur berbunga (hah)Umur panen (hah)Tinggi tanaman (cm)Banyaknya cabangBanyaknya polong/tanamanBobot biji/tanaman (g)Bobot 1000 biji (g)
Nilai keragamanKarakter
Tabel 111.8. Keragaman sifat agronomi plasma nutfah kacang hijau
Merah = 123, merah muda = 443, ungu = 26, putih =2, campuran = 3Hijau = 577, ungu = 8, merah coklat = 12Ungu = 16, hijau = 574, merah coklat = 9Kmt = 552, kuning = 28, oranye = 17Hijau = 578, ungu = 8; hijau ungu = 10, ungu hijau = 1Cepat = 516, lambat = 79Sedikit (5-6) = 580, banyak (>7) = 7<24 = 339, 15-19 = 215, >20 = 33Biji 2 = 549, Biji 1 = -, Biji 3-4 = 41<14 g = 167, 15-19 g = 267, >20 g = 153Tidak ada = 2, sedikit = 46, sedang = 203, dalam = 43, sangat dalam = 0Halus = 3, agak halus = 22, sedang = 258, kasar = 11Tidak ada = 2, sedikit = 46, sedang = 203, dalam = 43, sangat dalam = 0
Warna bijiWarna batangWarna ginoforeWarna bungaWarna keplng bijiUmur berbungaBanyaknya cabangBanyaknya polongBanyaknya bij/polongBobot polong/tanamanPelatukJaringan kulitPinggang
Sebaran sifatMorfologi
Tabel III.7. Sebaran slfat morfologi plasma nutfah kacang tanah
Kacang-kacangan Minor
Hasil rejuvenasi menunjukkan baliwakacang tunggak (V7gna unguiculata) meng-hasilkan biji 5-604 biji per polong (Tabel1II.9) atau menghasilkan biji tetapi sedikit.Nilai kisaran beberapa karakter kacang-
mempunyai keragaman genetik yang cu-kup besar terutama sifat umur polong ma-
sak, tinggi tanaman, banyaknya polong pertanaman, bobot biji per tanaman dan bo-bot 1000 biji. Sedangkan umur berbungadan banyaknya cabang keragamannyatidak begitu berbeda (Tabel III.8).
Laporan Tahunan Balitbiogen 2002

14
6,900,641,06
Maksimum
MK 2002
76,006,000,891,77
18,80
Maksimum
5,400,490,78
Minimum
Balitbiogen,
71,005,100,641,48
16,16
Minimum
5,900,540,87
Rata-rata
Panjang polong (cm)Diameter polong (cm)Lebar polong (cm)
Karakter
Tabel 111.11. Karakteristik kacang gude di
73,855,600,701,66
17,50
Rata-rata
Umur bunga (hah)Panjang polong (cm)Diameter polong (cm)Lebar polong (cm)Bobot 100 butir (g)
Karakter
Tabel 111.10. Karakteristik plasma nutfah kacang komak
daun bawah bervariasi dari warna kuning,hijau, putih, ungu muda sampai ungu gelap, merah, hitam serta hijau bercak ungu.Variasi lebar daun antara 20-42 cm, pan-
jang daun antara 29-62 cm, panjang tangkaidaun berpelepah antara 27-68 cm, dan panjang total tangkai daun antara 45-116 cm.
Umbi-umbian Minor
Sebanyak 158 aksesi plasma nutfahumbi-umbian minor terdiri dari 29 aksesiganyong, 18 aksesi garut, 16 aksesi gadung,30 aksesi gembili, 52 aksesi ubi kelapa, dan13 aksesi suweg telah dikonservasi dilapang.
Keragaman Plasma nutfah garut yangdikoleksi di Balitbiogen berasal dari JawaBarat, Jawa Tengah, Jawa Timur, dan
Sulawesi Tengah. Variasi karakter mor-fologi plasma nutfah garut dan ganyong disajikan pada Tabel 111.12. Tiga aksesi garutmempunyai potensi hasil paling tinggi,banyaknya anakan, panjang umbi, dandiameter umbi besar dibandingkan denganaksesi lain (Tabel III. 13). Dari tabel tersebuttampak bahwa aksesi yang menunjukkanhasil tinggi ada hubungan dengan banyaknya daun (>10 helai/tanaman) dan bobotumbi terberat diperoleh dari tanaman yang
kacangan minor disajikan pada Tabel III. 10,danIII.ll.
Ubi Jalar
Hasil rejuvenasi 347 aksesi plasmanutfah ubi jalar koleksi dari Bali, NusaTenggara Barat, dan Sumba diperoleh 310aksesi berumbi besar dan banyaknya umbibervariasi antara 2-10 umbi/aksesi, karenatidak semua tanaman dalam setiap aksesiberumbi. Sebanyak 89 aksesi mempunyaiketahanan sedang sampai tahan terhadaphama lanas.
Talas
Hasil karakterisasi plasma nutfah talasmenunjukkan keragaman pada beberapakarakter kecuali bentuk daun. Pada umum-
nya permukaan daun tidak mengkilat, ha-nya beberapa aksesi yang mempunyai permukaan daun mengkilat. Warna daun ber
variasi antara hijau gelap dan kehitaman.Warna tulang dan pinggiran daun bervariasi antara warna putih, kuning, hijau, ungu
muda sampai ungu gelap. Tangkai daunatas bervariasi dari warna kuning, merah,
ungu muda sampai ungu gelap, hitam, dan
hijau. Tangkai daun tengah berwama kuning, hijau, dan hitam, sedangkan tangkai
Hasil Penelitian 2002
15
- •--
-
-
8,28,08,28,5
181612
daun/tanamanBanyaknya
--
_
-
81,787,2
112,2117,6
82,089,289,1
i tanaman (cm)Tinggi
3,75,13,93,12,8
10,05,85,06,6
.--
3,33,63,0
13,510,59,7
11,19,0
•
--
--
21,526,626,1
umbi (cm) umbi (cm)Panjang Diameter
2,31,21,21,31,6
13,05,85,04,7
2,5 62,8 72,7 72,7 8
1,0 6,01,2 7,91,1 7,6
umbi/rumpun (kg) anakan/rumpunBobot Banyaknya
665570568567562
Gembili63660152536
Ubi kelapa477551615
Ganyong40338729
Garut
asalJenis/no. reg.
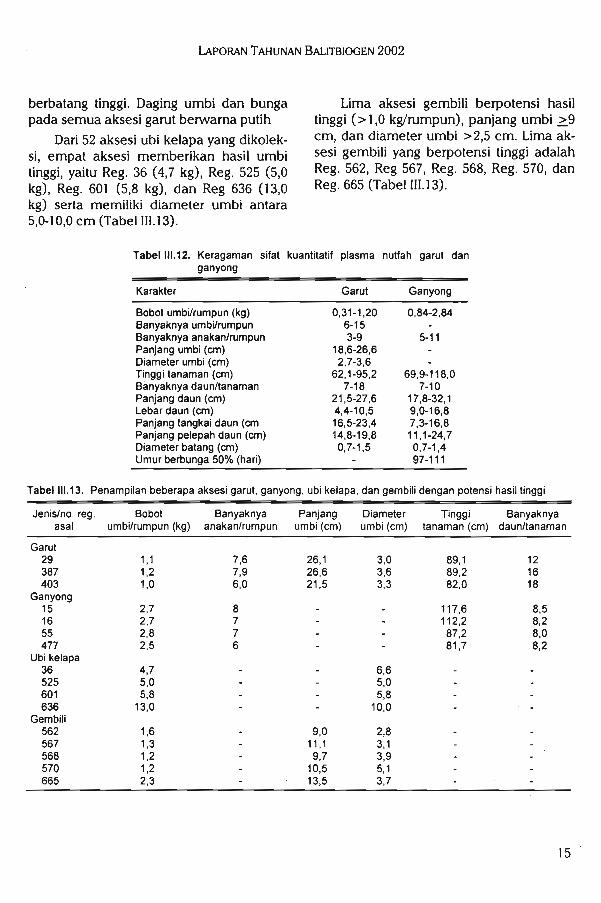
Tabel 111.13. Penampilan beberapa aksesi garut, ganyong, ubi kelapa, dan gembili dengan potensi hasil tinggi
97-1110,7-1,4
11,1-24,77,3-16,89,0-16,8
17,8-32,17-10
59,9-118,0
-
5-11-
0,84-2,84
Ganyong
-
0,7-1,514,8-19,816,5-23,44,4-10,521,5-27,6
7-1862,1-95,2 (
2,7-3,618,6-26,6
3-96-15
0,31-1,20
Garut
Umur berbunga 50% (hari)Diameter batang (cm)Panjang pelepah daun (cm)Panjang tangkai daun (cmLebar daun (cm)Panjang daun (cm)Banyaknya daun/tanamanTinggi tanaman (cm)Diameter umbi (cm)Panjang umbi (cm)Banyaknya anakan/rumpunBanyaknya umbi/rumpunBobot umbi/rumpun (kg)
Karakter
berbatang tinggi. Daging umbi dan bungaLima aksesi gembili berpotensi hasilpada semua aksesi garut berwarna putihtinggi (> 1,0 kg/rumpun), panjang umbi >9
Dari 52 aksesi ubi kelapa yang dikolek-cm- dan diameter umbi >2,5 cm. Lima ak-si, empat aksesi memberikan hasil umbisesi gembili yang berpotensi tinggi adalahtinggi, yaitu Reg. 36 (4,7 kg), Reg. 525 (5,0Re- 562> ReS 567. Reg- 568. Reg- 570> dankg), Reg. 601 (5,8 kg), dan Reg 636 (13,0Reg. 665 (Tabel 111.13).kg) serta memiliki diameter umbi antara5,0-10,0 cm (Tabel III. 13).
Tabel 111.12. Keragaman sifat kuantitatif plasma nutfah garut danganyong
Laporan Tahunan Balitbiogen 2002

16
Kandungan protein pada 30 aksesi ka-cang tanah bervariasi antara 23,1-28,0% dengan kandungan lemak antara 23,4-38,8%.
Empat aksesi kacang tanah yang mempunyai kadar protein tinggi (27,3-28,0%), yaituLok. Bima B (Reg. 1402), Lok SindangBarang (Reg. 986), Rapuk Sumbawa A(Reg. 1436), dan Rapuk Sumbawa C (Reg.1438). Kadar lemak yang tinggi terdapat pa-da Rapuk Ongko C (Reg. 1411) dan RapukSumbawa C (Reg. 1440), masing-masing38,8 dan 35,0%. Sedangkan aksesi RapukOngko (Reg. 1408) mempunyai kadarlemak paling rendah (23,4%).
Analisis kandungan pati 50 aksesi ubijalar dan 10 aksesi ubi kelapa (Dioscoreaalata) memperlihatkan bahwa 88% aksesiubi jalar dan semua aksesi ubi kelapamempunyai kadar pati >25%. Hasil analisiskandungan pati plasma nutfah ubi jalarberkisar antara 17,8-59,7% dan pada ubikelapa antara 41,9-50,4%. Tujuh aksesi ubijalar dan dua aksesi ubi kelapa mempunyaikandungan pati >0%.
Kandungan HCN plasma nutfah ubikayu bervariasi antara 6,3-150,5 ppm. Seba
nyak 12 aksesi ubi kayu memiliki kandungan HCN rendah (<22 ppm).
Hasil analisis kadar tanin pada 35aksesi plasma nutfah sorgum bervariasi
antara 0,16-0,35% dan 40% (14 aksesi) diantaranya mempunyai kadar tanin rendah(<0,25%). Pada umumnya biji aksesi sorgum yang mempunyai kadar tanin rendahberwarna putih dan kuning sedangkan bijiaksesi sorgum yang mempunyai kandungan tanin j>0,30% berwarna coklat dan
merah.
Konservasi In Vitro Plasma Nutfah UbiJalar, Ubi Kayu, dan Taias
Sebanyak 70 aksesi ubi jalar, 10 aksesiubi kayu, dan 10 aksesi taias telah dikon-servasi secara in vitro.
Karakterisasi Mutu Gizi Plasma NutfahTanaman Pangan
Keragaman kadar amilosa 100 aksesiplasma nutfah padi yang dianalisis berkisarantara 8,8-28,5%, enam aksesi di antaranya
adalah padi ketan dengan kadar amilosa<10%. Tujuh aksesi padi bukan ketan mem-punyai kadar amilosa rendah (10-19%), 36aksesi mempunyai kadar amilosa sedang(20-24%), dan 51 aksesi mempunyai kadaramilosa tinggi (25-35%).
Hasil analisis 99 aksesi plasma nutfahjagung bervariasi antara 10,2-28,1% dan lima di antaranya mempunyai kadar amilosarendah (10,2-18,2%), yaitu Bulareget (Reg.3550), Reg. 3282, Putik (Reg. 3075), Lok.Nata (Reg. 3082), dan Lok. Nggeru (Reg.3268). Aksesi lain mempunyai kadar amilosa sedang dan tinggi. Jagung dengan kadaramilosa rendah dikenal sebagai jagung ketan banyak ditemukan di Sulawesi Selatan,Nusa Tenggara Barat, dan Nusa TenggaraTimur, warna biji putih dengan rasa enak
dan pulen.
Hasil analisis kandungan protein danlemak 40 aksesi kedelai bervariasi antara29,7-39,8% dan 9,9-17,4%. Lok. Kediri (Reg.3508) mempunyai kadar protein yang paling rendah dan Lok. Ongko-2 (Reg. 4194)mempunyai kadar lemak paling rendah(9,9%) tetapi kadar proteinnya tinggi(39,7%). Di antara 40 aksesi kedelai yangdianalisis, tujuh di antaranya mengandungkadar protein tinggi (.+39%). Sedangkankandungan lemak plasma nutfah kedelaiyang diuji rendah.
Hasil Penelitian 2002

17
'Meliputi kacang tunggak (V. unguiculata), kacang gude (Cajanus cajan),komak, dan koro; "meliputi talas (Colocasia esculenta (L)), ganyong {Cannaedulis (Ker.)), patat/garut (Maranta arundinacea (L.)), gembili (Dioscoreaesculenta (L.)), gadung (Dioscorea hispida (Dent.)), ubi kelapa (Dioscoreaalata (L.)), dan suweg/iles-iles (Amorphophalus campanulatus); '"kacangtunggak (V. unguiculata)
Beium dientry130***912434
1029270771174705
3563
Dalam database
150184
1232434
10291194900200800
3801
Dalam bank gen
Jumlah aksesi
Ubi minor**Kacang minor*Ubi jalarUbi kayuKacang hijauKacang tanahKedelaiSorgumJagungPadi
Komoditas
Tabet 111.14. Banyaknya aksesi plasma nutfah tanaman pangan tahun 2002
aksesi ubi kayu, dan 130 aksesi kacangminor) dan 35 deskriptor baru (13 deskrip-tor kacang tanah dan 22 deskriptor kacangminor). Penambahan dan perbaikan datajuga telah dilakukan pada entry padi,kacang tanah, dan ubi kayu. Rekapitulasikegiatan entry data plasma nutfah tanamanpangan selama tahun 2002 disajikan padaTabel III. 15.
Pengembangan Slstem Database PlasmaNutfah Tanaman Pangan
Data plasma nutfah tanaman panganselanjutnya disusun dalam bentuk KatalogPlasma Nutfah Tanaman Pangan, yang se-
tiap tahun akan selalu disusun dan diper-baharui seiring dengan status pengelolaandatabase saat itu. Katalog ini memuat infor-
masi mengenai data paspor, data karakteri-sasi serta data evaluasi. Format penyusun-
an katalog tidak mengalami perubahan se-bagaimana katalog tahun sebelumnya.Dalam hal ini, nama varietas/galur/kultivardisusun menurut urutan abjad (sortingascending) untuk memudahkan penggunadalam menelusuri informasi dari suatu va-
Pengembangan Database Plasma NutfahTanaman Pangan
Dokumentasi Data Karakteristik PlasmaNutfah Tanaman Pangan
Penambahan data terjadi baik berupapenambahan data baru, pembaharuan{update) data maupun perbaikan (validasi)data. Sampai Januari 2003, database plasma nutfah tanaman pangan telah menam-pung 7.988 record yang meliputi 3.563aksesi padi, 705 aksesi jagung, 174 aksesisorgum, 771 aksesi kedelai, 270 aksesi ka-cang tanah, 1029 aksesi kacang hijau, 434aksesi ubi kayu, 912 aksesi ubi jalar, dan130 aksesi kacang minor (kacang tunggak).V. unguiculata (kacang tunggak) meru-pakan komoditas baru yang masuk dalamdatabase pada kegiatan tahun ini. Perinci-an banyaknya aksesi dari masing-masingkomoditas beserta banyaknya karakteryang telah berhasil dientry dalam databasedisajikan dalam Tabel III. 14.
Selama tahun 2002 terjadi penambahan data sebanyak 864 aksesi baru (305aksesi padi, 105 aksesi kacang tanah, 324
Laporan Tahunan Balitbiogen 2002

18
Evaluasi Toleransi Plasma Nutfah Padi,Jagung, dan Kedelai terhadap
Lahan Bermasalah (Lahan Masam,Keracunan Al, dan Fe)
Evaluasi Plasma Nutfah Padi terhadapKeracunan Al
Dari 200 aksesi, satu aksesi (KetanRingo) mati dan hampir seluruh aksesimempunyai daya tumbuh baik. Pada mula-nya pertanaman cukup baik, namun sete-lah berumur 30 hari tanaman mulai me-
nunjukkan keracunan. Pada umur tersebut
beberapa aksesi telah menunjukkan skor3-5 dan 7.
Hasil pengamatan pada umur 70 hariterlihat daun tanaman semakin mengu-ning. Banyak tanaman menunjukkan skor 7
dan 9. Beberapa aksesi mempunyai skor1-3 (Tabel III. 16).
Dari hasil pengamatan diketahui bah-wa keadaan tanah tidak seragam. Hal iniditunjukkan oleh skor tanaman pemban-ding peka yang bervariasi antara 1-9. Untukitu, seluruh aksesi pada Tabel 111.16 yangdikategorikan toleran perlu diuji ulang pada lokasi yang berbeda kadar aluminium-nya.
* Kacang tunggak (V. unguiculata)
rietas tertentu. Diharapkan katalog tersebutdapat dimanfaatkan oleh pengguna baikinternal maupun eksternal.
Guna lebih memudahkan dalam pe-ngelolaan data, maka sejak beberapa tahun terakhir telah disusun sistem databaseplasma nutfah tanaman pangan berbasis
Microsoft Access 2000.
Eksplorasi Plasma Nutfah TanamanPangan di Provinsi Kalimantan Barat
Pengumpulan dan Perolehan PlasmaNutfah Tanaman Pangan
Hasil eksplorasi diperoleh 191 aksesiplasma nutfah tanaman pangan, yaitu 129aksesi padi terdiri dari 69 aksesi padi sa-wah, 31 aksesi padi ladang, dan 29 aksesipadi pasang surut. Sebanyak 64 aksesi pa-lawija terdiri dari tiga aksesi jagung, 11aksesi ubi kayu, dan 17 aksesi ubi jalar,enam aksesi kacang tanah, satu aksesi ka
cang hijau, delapan aksesi ubi kelapa/krim-bang, 15 aksesi talas, dua aksesi kacangtunggak, dan satu aksesi jewawut.
-
Data baruValidasi dataUpdate data
-
Data baru
--
Update data
Status
-
223523-
13---
20
Deskriptor
status entry data
-
Data baru*Validasi data
Data baru-
update dataData baru dan
-
Data baru
Status
Banyaknya dan
-130912324
-
270---
305
Aksesi
Ubi minorKacang minorUbi jalarUbi kayuKacang hijau
Kacang tanahKedelaiSorgumJagungPadi
Komoditas
Tabel 111.15. Rekapitulasi kegiatan entry data plasma nutfah tanaman pangan selamatahun 2002
Hash Penelitian 2002
19
Evaluasi Plasma Nutfah Kedelai terhadapKeracunan Al
Dari hasil pengamatan diperoleh 14galur harapan toleran terhadap Al denganskor pengamatan 1-2, yaitu Reg. 1368
(317/882), B902A (Bali A), B3193 (GadekA), B3556 (M 2581), B3567 (Mlg 2593),B3665 (Lokal Pasuruan), B3677 (G 2120Mb), B3780 (M 3002), B3794 (M 3020),B4194 (Lok. Ongko 2), 1569 (Gm-388 Si),2812 (C73-01-113A-C-Pop), 3233 (KedelaiKecipir Putih), 3268 (C7301-1278-Pop),umur berbunga 37-50 had. Plasma nutfah
kedelai toleran terhadap keracunan Al disajikan pada Tabel 111.18.
Evaluasi Plasma Nutfah Jagung terhadapKeracunan Al
Dari 100 aksesi jagung yang diuji, di-peroleh 96 aksesi mempunyai daya tum-
buh baik, sedangkan lima aksesi (galurSukamandi) mempunyai daya tumbuh50-<20%. Dari hasil uji toleransi terhadapkeracunan Al, diperoleh 10 aksesi plasmanutfah jagung yang sedang sampai tolerandengan skor 2-3. Di antara 10 aksesi, tigaaksesi dengan skor Al 2,2-2,3, yaitu varietas
Kelinci (Reg. 2006), Leha-leha (Reg. 2026),dan Lokal Anyar (Reg. 3176). Plasma nutfah jagung terpilih yang toleran sampai sedang (skor 2-3) keracunan Al beserta pem-banding tahan dan peka disajikan padaTabel III.l7.
333
1-31-31-31-333
1-31-33
1-31-31-3
11
1-31-31-33
1-33
1-3
SkorAI
Raden RataLemo KwatikLakatan PutihRingkaSibauBuntut ItamPandanwangiKetumbarRingka KadirBuntut KasarRingka SinanPanceKetupatBilakPelangiSegeretTayanPadi DewiPadi CalonPemangkatLawiUmbang PulaiUmbang TopanTalun Undang
Varietas
KS56KS14KS13212862127121270212692126821267212662126521262212612126021235212302122821225212202119121190205232051920408
No. aksesi
Tabel 111.16. Plasma nutfah padi toleran terhadap keracunanAl, Tamanbogo, Lampung, MH 2002
Laporan Tahunan Balitbiogen 2002
20
Pengamatan pada umur 8 MST menunjukkan penurunan toleransi pada plasma nutfah padi yang diuji. Banyaknya aksesi yang tahan (skor 1-3 dan 3) menjadi 107nomor dan banyaknya aksesi yang sedang(skor 5) menjadi 49 nomor, sedangkanbanyaknya aksesi yang peka (skor 7) masihsama dengan pengamatan pada umur 4MST, yaitu 38 nomor. Beberapa aksesi yangtoleran (skor 1-3) pada pengamatan 8 MSTadalah Randah Padang K, Getik, Mendalet,Menta, Ritgen, Jedah, Ketan, Sampang K,
Evaluasi Plasma Nutfah Pad! terhadapKeracunan Fe
Hasil evaluasi terhadap 200 aksesiplasma nutfah tanaman padi umur 4 ming-gu setelah tanam (MST) atau 55 hari sejaktabur, sudah menunjukkan gejala keracunan Fe. Terdapat 37 aksesi peka terhadapkeracunan Fe (skor 7), 112 aksesi toleranterhadap keracunan Fe (skor 1-3 dan 3)serta 46 aksesi bereaksi sedang (skor 5).
317/882Bali AGM-388 SiC73-01-113A-C-PopGadekAKedelai Kecipir PutihM2581Mlg 2593Lokal PasuruanG 2120 MbM3002M3020C7301-1278-PopLok. Ongko 2
1368B902A15692812B31933233B3556B3567B3665B3677B3780B37943268B4194
VarietasNo. reg.
' 1 = sangat toleran, 2 = toleran, 3 = sedang, 4 = peka, 5 = sangat peka/mati
Tabel 111.18. Plasma nutfah kedelai toleran terhadap keracunan Al(skor 1-2) Tamanbogo, Lampung, MH 2002
745787964587
Silk delay(hari)
8562758883828288
100898365
Persentase daya tumbuh(%)
3,02,52,32,22,22,22,52,02,02,22,02,5
Aspek tanaman70 hari (1-5)
3,02,22,52,32,52,52,32,52,22,52,52,5
Skor keracunan Al(1-5)*
ArjunaAntasenaLokal Lendang BatuLokal AnyarLokal SepapanEEW-DMR-CoS,Leha-lehaImproved TiniquibKelinciSW90L-206-17Galur 124SiGalur111Si
Varietas
31813176312633232026333020063336
No. reg.
Tabel 111.17. Plasma nutfah jagung toleran sampai sedang terhadap keracunan Al (skor 2-3) pada umur 42 HST,Tamanbogo, Lampung, MH 2002
Hasil Penelitian 2002

21
Evaluasi Ketahanan Plasma Nutfah Paditerhadap Bias
Sebanyak 200 aksesi plasma nutfahpadi telah ditanam pada akhir November2002 di desa Cimenteng, Sukabumi. Peru-
bahan cuaca yang dicirikan dengan curahhujan yang tidak merata ternyata kurangmenunjang penelitian. Selama bulan per-
tama dan kedua curah hujan sangat kurang. Selama bulan Januari hanya terjadi 4hari hujan 5-15 mm. Tidak meratanya harihujan menyebabkan pertumbuhan tanaman terhambat dan perkembangan penyakitterganggu.
Curah hujan pada bulan Februari cu-kup merata dan selama 11 hari berkisarantara 5-86 mm/hari atau 241 mm sebulan(data pengamatan curah hujan, Sukamaju,
Februari 2003). Temperatur minimum 17Cdan maksimum 32C, kelembaban 59-96%,tampak perkembangan penyakit cukup pe-
sat. Hasil pengamatan diperoleh 10 aksesi(Loneng, Pembagi, Padi Jangkong, Kayok,Pandan, Bungung Inih, Pulut Timun, Pulut
Timau, Pulut Alui, dan Pulut Namang) agaktahan terhadap serangan penyakit (skor 3).
Evaluasi Ketahanan Plasma NutfahJagung terhadap Penyakit Bulai
Dari hasil uji penyakit bulai diperoleh20 aksesi plasma nutfah jagung termasuksangat tahan dengan persentase seranganpenyakit bulai 0-5%. Sebagai varietas kon-
trol tahan (Lagaligo) terserang 25,4%, se-dangkan sebagai varietas kontrol rentan(Antasena) terserang 88,6%. Aksesi yangsangat tahan dan laju serangannya padaumur 3, 4, dan 5 MST disajikan pada Tabel111.19.
Komas A, Komas B, dan Sipulo. Varietas
Pulu Denni B, Genjah Mada, Pudak Kuning,Cere Bandung, dan Rojolele memiliki tole-ransi sama (skor 1-3) seperti pada peng-amatan umur 45 MST.
Evaluasi Ketahanan Plasma Nutfah
Tanaman Pangan terhadap PenyakitHawar Daun Bakteri dan Bias pada Padi,Bulai pada Jagung, dan Sapu Setan pada
Kacang Tanah
Evaluasi Ketahanan Plasma Nutfah Paditerhadap Penyakit Hawar Daun Bakteri
Evaluasi ketahanan plasma nutfahpadi terhadap penyakit hawar daun bakteri(HDB) telah dilakukan di rumah kaca,Balitbiogen pada MH 2002. Sebanyak 250aksesi plasma nutfah padi telah diuji terhadap isolat kelompok IV dan VIII. Pertum-buhan tanaman pada umumnya baik. Selu-
ruh tanaman diinokulasi pada saat primor-
dia dan pengamatan dilakukan dua minggukemudian.
Dari 250 aksesi yang diuji ketahanan-nya terhadap penyakit hawar daun bakterikelompok VIII diperoleh delapan aksesiyang menunjukkan reaksi tahan sampaiagak tahan dengan skor 1-3, yaitu varietasCina (Reg. 3291), 1082/185 (Reg. 3445),Mendalet (Reg. 5657), Tholo (Reg. 5720),Ritgen (Reg. 6550), Jedah (Reg. 6601),Bokongan (Reg. 6859), Angkong (Reg.7237), Kayok (Reg. KBP 57), Bumbung Inih(KBP 66), Jala (KBP 80), Kapor (KBP 96),Panglima (KBP 97), dan Ringka (KBP 118).
Pengujian terhadap kelompok IV diperoleh 25 aksesi dengan skor 1, 52 aksesiskor 3, 28 aksesi skor 5, 12 aksesi skor 7,
dan 25 aksesi skor 9.
Laporan Tahunan Balitbiogen 2002

22
rentan, dan 15 aksesi (5%) agak tahan terhadap wereng coklat IR42.
Dari 17 dan 15 aksesi plasma nutfahpadi yang agak tahan terhadap werengcoklat populasi IR42 dan populasi IR64, ter-nyata yang hanya dua aksesi yang mempu-nyai respon stabil terhadap kedua populasiwereng coklat, yaitu varietas Ase Botongdan Ase Pute.
Evaluasi Ketahanan Plasma Nutfah UbiJalar terhadap Hama Lanas
Dari 50 aksesi plasma nutfah yang diujiketahanannya terhadap hama lanas (Cylasformicarius), diperoleh satu aksesi tahan(varietas Yoban) yang mempunyai skorrata-rata 0,3. Varietas lain (sembilan aksesi) mempunyai reaksi agak tahan (skor1,2), 24 aksesi agak rentan (skor 2,3), dan16 aksesi rentan (skor >3,2).
Evaluasi Ketahanan Plasma NutfahTanaman terhadap Hama Wereng Coklat
pada Padi dan Hama Lanas padaUbi Jalar
Evaluasi Ketahanan Plasma Nutfah Paditerhadap Hama Wereng Coklat
Dari 300 aksesi plasma nutfah padiyang diuji tingkat ketahanannya terhadapwereng coklat populasi IR42 atau biotipeSU dengan metode seedling test menun-jukkan respon tingkat ketahanan yang ber-variasi. Respon yang paling dominanadalah 182 aksesi (60,7%) rentan, 95 aksesi(31,7%) agak rentan, dan 17 aksesi (5,7%)agak tahan terhadap wereng coklat IR42.
Demikian juga ke-300 aksesi plasmanutfah padi yang diuji tidak ada yang tahanterhadap wereng coklat populasi IR64.Respon yang paling dominan adalah 171aksesi (57%) rentan, 108 aksesi (36%) agak
88,625,4
4,34,23,84,92,93,512,22,32,54,14,724,63,33,04,654,702,854,152,452,3
(%)Total serangan pada 5 MST
16,65,0000000,7100000,72001,0000000
5MST
penyakit bulai (%)
50,012,44,34,23,84,92,91,42,22,32,54,14,03,13,31,04,654,72,854,152,452,3
4MST
laju serangan
228000001,4000001,501,0000000
3MST
Persentase
Antasena (pekaLagaligo (tahan)SGP/IVTI/36/22P5G18(S)C3-28-3-1-2P4G19(S)C2-126-1-1-2Arc-S5-Pop 2-1P5g18(S)C2-56-3-1-1P5/k/T1/544/12-2P4G19(S)C2-110-2-1-1Arc 178-1-4-1-3-2-1-1-xb3-3P4G19(S)C2-17-2-2-1Arc 178-1-3-1-4-2-2-xb3-3SGPD/675/5-1P4G19(S)C2-90-1-2-2P4G19(S)C2-84-1-1-1P4G19(S)C2-114-3-1-1P5K/T1/523/12-3Arc 178-1-3-1-1-4-1-1-xb3-2SGPD/675/15-2P4G19(S)C2-47-2-1-1P4G19(S)C2-126-1-1-4Arc 83-2-3-1-1-1-2-1-xb3-2
Nama aalur
Tabel 111.19. Hasil evaluasi plasma nutfah jagung terhadap penyakit bulai
Hasil Penelitian 2002

23
jukkan tingkat polimorfisme yang relatif cukup tinggi. Pita DNA hasil PCR yang dihasil-kan untuk masing-masing primer, jumlahdan ukurannya ^angat bervariasi. Apabila
dilihat dari ukurannya, maka pita DNA yangdihasilkan dari ketiga primer tersebut ber-ukuran tidak lebih dari 2000 bp.
Perbandingan di antara tiga primeryang digunakan untuk menghasilkan pitaDNA disajikan pada Tabel 111.20.
KONSERVASI SUMBER DAYA GENETIKMIKROBA PERTANIAN
Dwi N. Susilowati, Arief Indrasumunar,Rosmimik, Rasti Saraswati, Erni Yuniarti,
M. Machmud, Sutrisno, R.D.M. Simanungkalit,Lukman Gunarto, Jumanto, M. Sudjadi, dan
Yadi Suryadi,
Mikroba menyumbang potensi ekono-mi terbesar dibandingkan dengan kelom-pok organisme lain karena keragamannyayang sangat tinggi, kemampuan untukhidup dan bertahan pada berbagai kondisiekstrim, perkembangbiakan yang cepat,kemampuan untuk menghasilkan produkdalam laju konsentrasi tinggi, serta sifatfisiologi yang relatif mudah dikendalikandan dimanipulasi.
Kelompok mikroba penyubur tanahberperan sangat penting bagi sistem pro-duksi pertanian yang berkelanjutan. Pe-
manfaatan mikroba penyubur tanah yangsesuai dengan kondisi tanah merupakanaltematif yang murah untuk meningkatkankesuburan tanah, efisiensi pemupukan,
dan mengurangi bahaya pencemaran.
Pemanfaatan mikroba tanah yang dapat mengakumulasi logam berat dapat di-upayakan untuk melindungi tanah sebagaisarana produksi pangan terhadap pencemaran. Microbial remediation seperti, bak-teri, fungi, alga, dan yeast mampu meng-
Plasma nutfah ubi jalar yang agak ta-han (skor rata-rata antara 1,0-1,3) antara
lain Mantang Biru, Retok, dan Ubi Gendola.Apabila dilihat dari banyaknya larva, pupa,dan imago yang terdapat pada umbi, ter-nyata pada varietas yang tahan menunjuk-kan populasi larva, pupa, dan imago yang
paling rendah, selanjutnya pada varietasagak tahan, populasi larva, pupa, dan imago lebih rendah dari varietas agak rentandan rentan, sedangkan populasi larva, pu
pa, dan imago paling tinggi diperoleh padavarietas yang rentan.
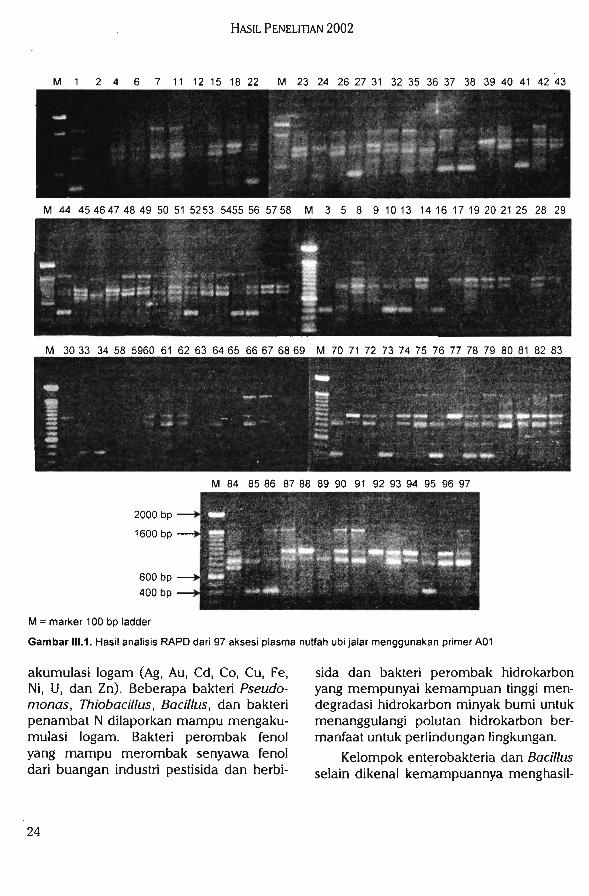
Karakterisasi Molekuler Plasma NutfahTanaman Pangan
Polimorfisme yang terjadi dari analisisRAPD ditunjukkan dengan terbentuknya pita DNA pada individu yang satu sementaraindividu yang lain tidak terbentuk pita DNApada posisi atau ukuran yang sama. Dengan
adanya polimorfisme ini dapat menggam-barkan tingkat keragaman genetik plasmanutfah yang dikarakterisasi. Semakin tinggitingkat polimorfisme maka tingkat keragaman genetik di antara individu plasmanutfah juga akan semakin tinggi. Primeryang digunakan pada penelitian ini diha-rapkan akan memberikan hasil analisisdengan tingkat polimorfisme yang cukuptinggi.
Tiga buah primer oligonukleoida acak,yaitu primer A01, A04, dan B01 telah digunakan untuk mengamplifikasi 97 aksesiplasma nutfah ubi jalar. Hasil amplifikasiPCR dengan menggunakan tiga primeroligonukleotida acak dapat dilihat padaGambar III. 1, II1.2, dan II1.3. Dari ketiga
gambar tersebut dapat dilihat bahwa seca-ra umum, primer yang digunakan dapatmengamplifikasi DNA dari aksesi yang di-analisis. Dari tingkat polimorfisme juga dapat dilihat bahwa primer tersebut menun-
Laporan Tahunan Balitbiogen 2002
24
sida dan bakteri perombak hidrokarbonyang mempunyai kemampuan tinggi men-degradasi hidrokarbon minyak bumi untukmenanggulangi polutan hidrokarbon ber-manfaat untuk perlindungan lingkungan.
Kelompok enterobakteria dan Bacillusselain dikenal kemampuannya menghasil-
akumulasi logam (Ag, Au, Cd, Co, Cu, Fe,Ni, U, dan Zn). Beberapa bakteri Pseudo-
monas, Thiobacillus, Bacillus, dan bakteripenambat N dilaporkan mampu mengaku-
mulasi logam. Bakteri perombak fenolyang mampu merombak senyawa fenol
dari buangan industri pestisida dan herbi-
M 84 85 86 87 88 89 90 91 92 93 94 95 96 97
2000 bp1600 bp
600 bp
400 bp
M = marker 100 bp ladder
Gambar III.t. Hasil analisis RAPD dari 97 aksesi plasma nutfah ubi jalar menggunakan primer A01
Ji
ii3
8382
t ,^ r^,
81
i ^
807978
-
77
,-••
7675747372
I'
71
ff '<n
f70
•4
M696867666564636261596058343330M
M 44 45 4647 48 49 50 51 5253 5455 56 57 58 M 3 5 8 9 10 13 14 16 17 19 20 21 25 28 29
M 1 2 4 6 7 11 12 15 18 22 M 23 24 26 27 31 32 35 36 37 38 39 40 41 42 43
Hasil Penelitian 2002
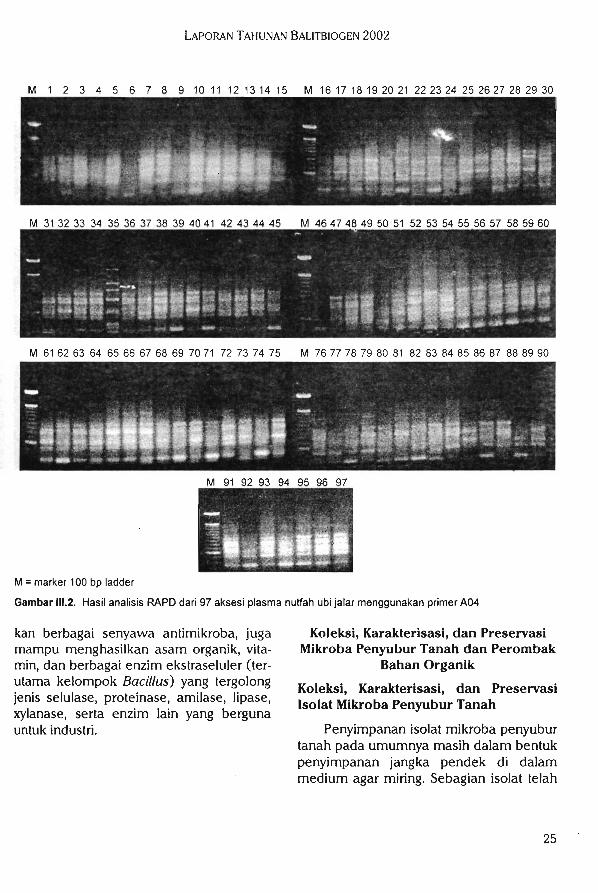
25
Koleksi, Karakterisasi, dan Preservasi
Mikroba Penyubur Tanah dan PerombakBahan Organik
Koleksi, Karakterisasi, dan Preservasi
Isolat Mikroba Penyubur Tanah
Penyimpanan isolat mikroba penyuburtanah pada umumnya masih dalam bentukpenyimpanan jangka pendek di dalammedium agar miring. Sebagian isolat telah
kan berbagai senyawa antimikroba, jugamampu menghasilkan asam organik, vitamin, dan berbagai enzim ekstraseluler (ter-
utama kelompok Bacillus) yang tergolongjenis selulase, proteinase, amilase, lipase,
xylanase, serta enzim lain yang bergunauntuk industri.
M = marker 100 bp ladder
Gambar III.2. Hasil analisis RAPD dari 97 aksesi plasma nutfah ubi jalar menggunakan primer A04
M 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 M 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90
^ ,^L
•t s I 1 f f • "9^ 9 •"" r •^J [Mi*
M 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 M 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 M 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Laporan Tahunan Balitbiogen 2002
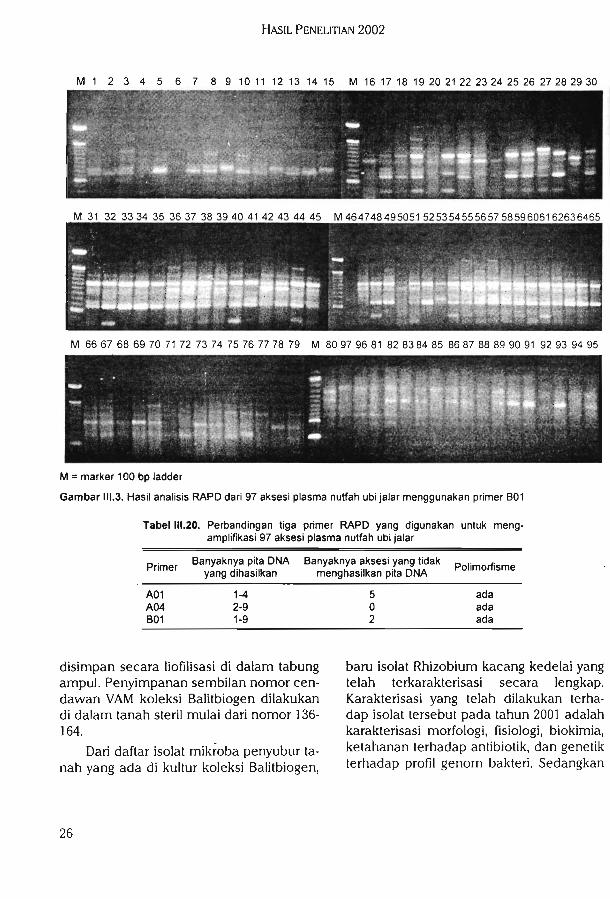
26
bam isolat Rhizobium kacang kedelai yangtelah terkarakterisasi secara lengkap.Karakterisasi yang telah dilakukan terha-
dap isolat tersebut pada tahun 2001 adalahkarakterisasi morfologi, fisiologi, biokimia,ketahanan terhadap antibiotik, dan genetikterhadap profil genom bakteri. Sedangkan
disimpan secara liofilisasi di dalam tabungampul. Penyimpanan sembilan nomor cen-
dawan VAM koleksi Balitbiogen dilakukandi dalam tanah steril mulai dari nomor 136-164.
Dari daftar isolat mikroba penyubur tanah yang ada di kultur koleksi Balitbiogen,
adaadaada
205
1-92-91-4
B01A04A01
M = marker 100 bp ladder
Gambar III.3. Hasil analisis RAPD dari 97 aksesi plasma nutfah ubi jalar menggunakan primer B01
Tabel 111.20. Perbandingan tiga primer RAPD yang digunakan untuk meng-amplifikasi 97 aksesi plasma nutfah ubi jalar
Primer Banyaknya pita DNA Banyaknya aksesi yang tidak Po|imorfismePnmer yang dihasilkanmenghasilkan pita DNA Polimortisme
f. ^f. 4.V -t *^ ;••'. I %. 4
M 66 67 68 69 70 71 72 73 74 75 76 77 78 79 M 80 97 96 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95
"#^^ ^#^ *• I, f
M 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 M 464748495051 525354555657 5859606162636465
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 M 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Hasil Penelitian 2002

27
(BCr 1.2, BCr 2.1, BCr 2.3, BCbd 1.3, dan
APK 2.4) sebagai inokulan tanaman padidan dua isolat unggul bakteri endofitik daritanaman jagung (JCbd 2.1 dan JLk-CN 2.5)sebagai inokulan tanaman jagung.
Koleksi, Karakterisasi, dan Preservasi
Isolat Mikroba Perombak Bahan Organik
Hasil isolasi mikroba perombak bahanorganik telah diperoleh 23 isolat yang ter-diri dari 13 isolat jenis bakteri dan 10 isolatjenis kapang. Tumbuhnya koloni bakteri di-tandai dengan terbentuknya zona beningpada media spesifik untuk selulolitik(media CMC) yang menunjukkan terjadi-nya hidrolisis substrat oleh enzim selulase.
Aktivitas enzim tertinggi (0,1298 U/ml)dihasilkan oleh kapang M10. Enzim selulase dapat dihasilkan oleh beberapa jeniskapang dan bakteri sebagai respon terhadap selulosa pada lingkungan hidupnya.Kemampuan memproduksi enzim men-
jadikan mikroba tersebut mampu meng-hidrolisis selulosa menjadi glukosa ataugula lain yang larut dan dapat dijadikansumber karbon bagi pertumbuhannya.
Filtrat enzim selulase yang diperolehdari proses ekstraksi masih berupa enzimkasar, sehingga masih perlu diproses lebihlanjut dengan cara pengendapan dengangaram netral konsentrasi tinggi (amoniumsulfat) yang bersifat mudah larut, tidaktoksik, dan dapat menstabilkan enzim.Pengendapan enzim dilakukan pada konsentrasi amonium sulfat 30-90%. Pengen
dapan protein dengan amonium sulfat dengan menggunakan metode sentrifugasiakan memisahkan protein dari kontaminanprotein, seperti karbohidrat, dan lipid. Aktivitas CMC-ase mempunyai nilai tertinggipada konsentrasi amonium sulfat 60%(Gambar III.4). Hal ini menunjukkan bah-wa penambahan amonium sulfat pada kon-
pada tahun 2002 penekanan pada karak-terisasi fisiologi dan biokimia isolat Rhizo-bium hasil fusi protoplas intergenerik danintraspesies (Tabel 111.21). Karakter morfo-logi koloni dari Rhizobium hasil fusi protoplas baik secara intergenerik maupun intraspesies ternyata tidak mengalami perubah-
an dari karakter morfologi tetuanya, yaituberbentuk bulat, cembung, tepian rata, ber-warna pink, dan berlendir.
Berdasarkan uji keefektifan isolatRhizobium hasil fusi protoplas dalam halpenambatan N2 udara, diketahui empat iso
lat hasil fusi protoplas intergenerik (INTER-7, INTER-9, INTER-10, dan INTER-13) sangatefektif menambat N2, sedangkan untuk
hasil fusi protoplas intraspesies diperolehlima isolat (INTRA-4, INTRA-8, INTRA-10,INTRA-15, dan INTRA-23) sangat efektif menambat N2. Isolat INTRA-8 yang memilikinilai keefektifan simbiotik dan kapasitassimbiotik lebih tinggi dari keempat isolatlain telah digunakan sebagai salah satuinokulan untuk benih matriconditioning.
Di samping karakterisasi morfologi,fisiologi, dan biokimia terhadap isolatRhizobium hasil fusi protoplas, juga dilaku-kan karakterisasi terhadap isolat bakteridiazotrof endofitik dan filosfer. Pada pene-litian tahun 2001 hanya berhasil diisolasi 58isolat bakteri diazotrof endofitik dari ta-naman padi dan jagung, sedangkan padatahun 2002 telah berhasil diisolasi 90 isolat,sehingga jumlah keseluruhan isolat bakteridiazotrof endofitik 148 isolat. Beberapaisolat tersebut telah diuji kemampuannyadalam menghasilkan zat pemacu tumbuhauksin secara kolorimetri menggunakanspektrofotometer dan kemampuannya dalam menambat nitrogen udara menggunakan gas kromatografi. Setelah dilakukanpenyeleksian lebih lanjut, dipilih lima isolatunggul bakteri endofitik dari tanaman padi
Laporan Tahunan Balitbiogen 2002
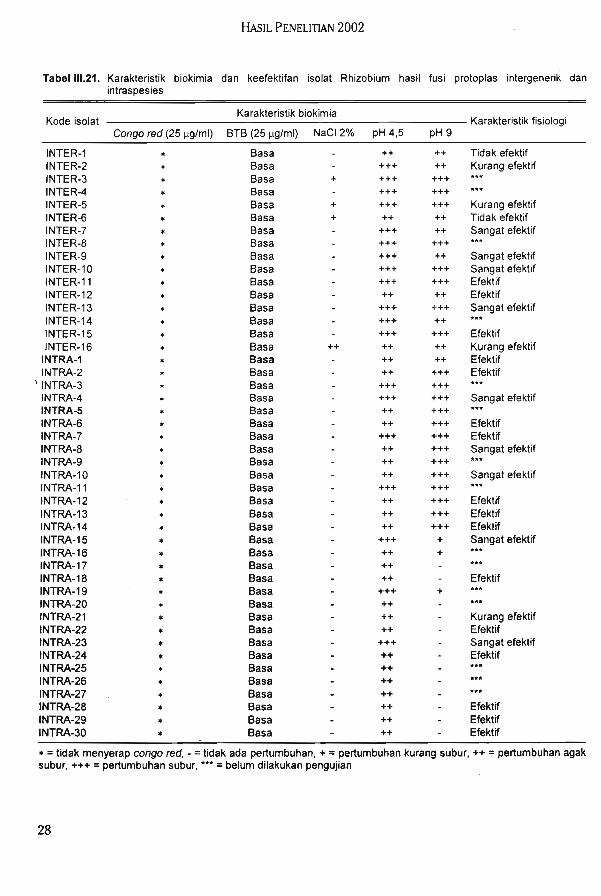
28
? = tidak menyerap congo red, - = tidak ada pertumbuhan, + = pertumbuhan kurang subur, ++ = pertumbuhan agaksubur, +++ = pertumbuhan subur, *** = belum dilakukan pengujian
EfektifEfektifEfektif
Kurang efektifEfektifSangat efektifEfektif
Efektif
Kurang efektifTidak efektifSangat efektif
Sangat efektifSangat efektifEfektifEfektifSangat efektif***EfektifKurang efektifEfektifEfektif
Sangat efektif
EfektifEfektifSangat efektif
Sangat efektif***EfektifEfektifEfektifSangat efektif
Tidak efektifKurang efektif
BasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasaBasa
INTRA-30 *INTRA-29 *INTRA-28 *INTRA-27 *INTRA-26 *INTRA-25 *INTRA-24 *INTRA-23 ?INTRA-22 *INTRA-21 ?INTRA-20 *INTRA-19 ?INTRA-18 *INTRA-17 *INTRA-16 *INTRA-15 ?INTRA-14 *INTRA-13 *INTRA-12 *INTRA-11 *INTRA-10 *INTRA-9 *INTRA-8 INTRA-7 ?INTRA-6 *INTRA-5 *INTRA-4 *INTRA-3 *INTRA-2 *INTRA-1 ?
INTER-16 *INTER-15 *INTER-14 *INTER-13 ?INTER-12 *INTER-11 *INTER-10 *INTER-9 *INTER-8 *INTER-7 *INTER-6 ?INTER-5 *INTER-4 •INTER-3 *INTER-2 *INTER-1 *
• Karakteristik fisiologiCongo red (25 pg/ml) BTB (25 pg/ml) NaCI 2% pH 4,5 pH 9
Karakteristik biokimiaKode isolat
1.21. Karakteristik biokimia dan keefektifan isolat Rhizobium hasil fusi protoplas intergenerik danintraspesies
Hasil Penelitian 2002
Tabel I
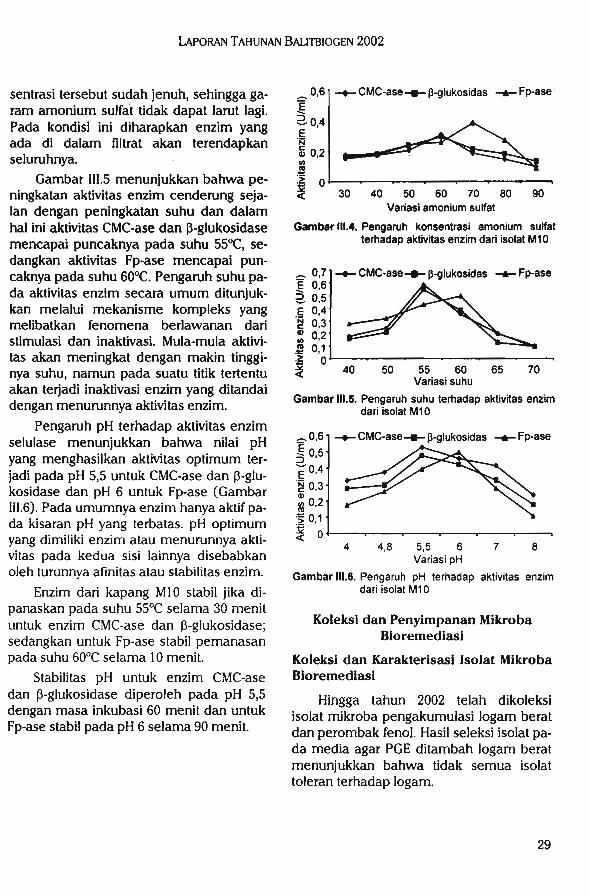
29
5,5 67Variasi pH
Gambar III.6. Pengaruh pH terhadap aktivitas enzimdari isolat M10
Koleksi dan Penyimpanan MikrobaBioremediasi
Koleksi dan Karakterisasi Isolat MikrobaBioremediasi
Hingga tahun 2002 telah dikoleksiisolat mikroba pengakumulasi logam beratdan perombak fenol. Hasil seleksi isolat pada media agar PGE ditambah logam beratmenunjukkan bahwa tidak semua isolattoleran terhadap logam.
84,8
- CMC-ase-•-p-glukosidas -*-Fp-ase| 0,5^ 0.4I 0,3J 0,2
;f 0,1
I 0
Gambar III.5. Pengaruh suhu terhadap aktivitas enzimdari isolat M10
7055 60 65Variasi suhu
5040
-CMC-ase-a- p-glukosidas -*- Fp-ase0,70,60,50,40,30,20,1
Gambar 111.4. Pengaruh konsentrasi amonium sulfatterhadap aktivitas enzim dari isolat M10
9030 40 50 60 70 80Variasi amonium sulfat
- CMC-ase -#- p-glukosidas -*- Fp-ase
ES 0,2
sentrasi tersebut sudah jenuh, sehingga ga-ram amonium sulfat tidak dapat larut lagi.Pada kondisi ini diharapkan enzim yangada di dalam flltrat akan terendapkanseluruhnya.
Gambar III.5 menunjukkan bahwa pe-ningkatan aktivitas enzim cenderung seja-lan dengan peningkatan suhu dan dalamhal ini aktivitas CMC-ase dan p-glukosidasemencapai puncaknya pada suhu 55C, se-dangkan aktivitas Fp-ase mencapai puncaknya pada suhu 60C. Pengaruh suhu pada aktivitas enzim secara umum ditunjuk-kan melalui mekanisme kompleks yangmelibatkan fenomena berlawanan daristimuiasi dan inaktivasi. Mula-mula aktivitas akan meningkat dengan makin tinggi-nya suhu, namun pada suatu titik tertentu
akan terjadi inaktivasi enzim yang ditandaidengan menurunnya aktivitas enzim.
Pengaruh pH terhadap aktivitas enzimselulase menunjukkan bahwa nilai pHyang menghasilkan aktivitas optimum terjadi pada pH 5,5 untuk CMC-ase dan p-glu-kosidase dan pH 6 untuk Fp-ase (GambarHI.6). Pada umumnya enzim hanya aktif pada kisaran pH yang terbatas. pH optimumyang dimiliki enzim atau menurunnya aktivitas pada kedua sisi lainnya disebabkanoleh turunnya afinitas atau stabilitas enzim.
Enzim dari kapang M10 stabil jika di-panaskan pada suhu 55C selama 30 menituntuk enzim CMC-ase dan p-glukosidase;sedangkan untuk Fp-ase stabi! pemanasanpada suhu 60C selama 10 menit.
Stabilitas pH untuk enzim CMC-asedan p-glukosidase diperoleh pada pH 5,5dengan masa inkubasi 60 menit dan untukFp-ase stabil pada pH 6 selama 90 menit.
Laporan Tahunan Balitbiogen 2002

30
97-01, PG 97-02, MR 1.12-05, PB 138, Al,
dan B 2.2) mampu tumbuh subur (++) pada logam Pb dari konsentrasi 397,4-662,4ppm. Isolat MT 77-09, MT 77-10, MT 77-19,
MR 24-02, MR 25-02, BB 288, dan OM 343mampu tumbuh sampai konsentrasi 662,4ppm tetapi kurang subur (+).
Sebanyak 10 isolat (MT 53-02, MT 77-08, PG 65-06, PG 97-01, PG 97-02, PG 101-26, MR 1.12-05, PB 138, Al, dan B 2.2)mampu tumbuh subur (+ +) pada mediumyang ditambah logam Cd dari konsentrasi5-7 ppm (Tabel 111.23).
Sebanyak 31 isolat telah dikarakterisasimorfologi sel yang meliputi reaksi pewar-naan Gram, pembentukan endospora, dan
bentuk sel (Tabel 111.24). Sebanyak 27 isolattermasuk kelompok Gram positif, empatisolat (MT53-01, MP12-05, Bd-3, dan Bd-4)merupakan kelompok Gram negatif. Dari
16 isolat Gram positif yang diuji, 15 isolatmembentuk endospora, sedangkan empat
isolat Gram negatif belum dikarakterisasiadanya bentuk endospora. Bentuk sel iso
lat yang diamati menunjukkan bentuk selbatang dan teramati dengan mikroskopmulai perbesaran 400 kali sedangkan bentuk sel keempat isolat bakteri Gram negatifbaru bisa diamati pada perbesaran 1000kali (batang pendek). Satu isolat Gram positif dan tidak membentuk endospora (OM343) berbentuk bulat agak lonjong menunjukkan ciri bentuk sel khamir (Saccharo-myces), sudah tampak jelas pada perbesaran lensa mikroskop 100 kali. Lima belasisolat Gram positif yang membentuk endospora dan selnya berbentuk batang merupakan genus Bacillus spp.
Dari hasil identifikasi secara morfologisel, biokimia, dan fisiologi terhadap enamisolat terpilih, empat isolat (MT77-08,MR1.12-05, PG65-06, dan PG97-02) menunjukkan ciri Bacillus breuis, isolat PG97-01
Di antara 77 isolat yang mamp<u meng-akumulasi logam berat, 59 isolat berasal
dari tanah gambut Ekosistern Ahr HitamKalimantan Tengah, 10 isolat berasal daritanah savvah tercemar limbah industri diBekasi dan Karawang, empat isolatt berasal
dari daerah industri di sekitar alirajn sungaiBengawan Solo, dan empat isolat berasal
dari tanah sawah di sekitar daerah industriBandung Selatan.
Sebanyak 27 isolat marnpu tumbuhpada media PGE yang mengandung logamMn dengan konsentrasi tertinggi 1375 ppm;11 isolat mampu tumbuh pad.a media PGEyang mengandung Fe dengan konsentrasi
tertinggi 1116,9 ppm; 15 isolat manupu tumbuh pada media PGE yang mengandung Zndengan konsentrasi tertinggi 1 76,2 ppm; 40isolat mampu tumbuh pada media PGEyang mengandung Pb dengan konsentrasitertinggi 662,4 ppm; 46 isolat rnampu tumbuh pada media PGE yang mengandungCd dengan konsentrasi tertinggi 10 ppm.Dua isolat (PG 65-6 dan MR25-01)) toleranterhadap logam Mn, Fe, Zn, Pb, dan Cd
sedangkan isolat lain toleran terhadapkurang dari kelima logam tersebut.
Ketahanan hidup mikrofoa p>engaku-mulasi logam berat pada media yang mengandung logam Zn lebih rendah daripadalogam Mn, yaitu hanya sampai pada kon
sentrasi 176,2 ppm, sedangkan kemampu-
an hidup pada media yang mengandung Fedapat mencapai 1116,9 ppm.
Berdasarkan uji kemampuan tumbuhpada logam Pb (397,4-662,4 ppm) dan Cd(5-7 ppm), ke-28 isolat mikroba pengaku-mulasi logam berat menunjukkan kemampuan tumbuh yang berbeda. Pengamatan
pertumbuhan mikroba pengakumulasi logam berat pada medium yang ditambah logam Pb (Tabel 111.22) menunjukkan sembi-lan isolat (MT 53-02, MT 77-08. PG 65-06, PG
H.ASIL PENELITIAN 2002
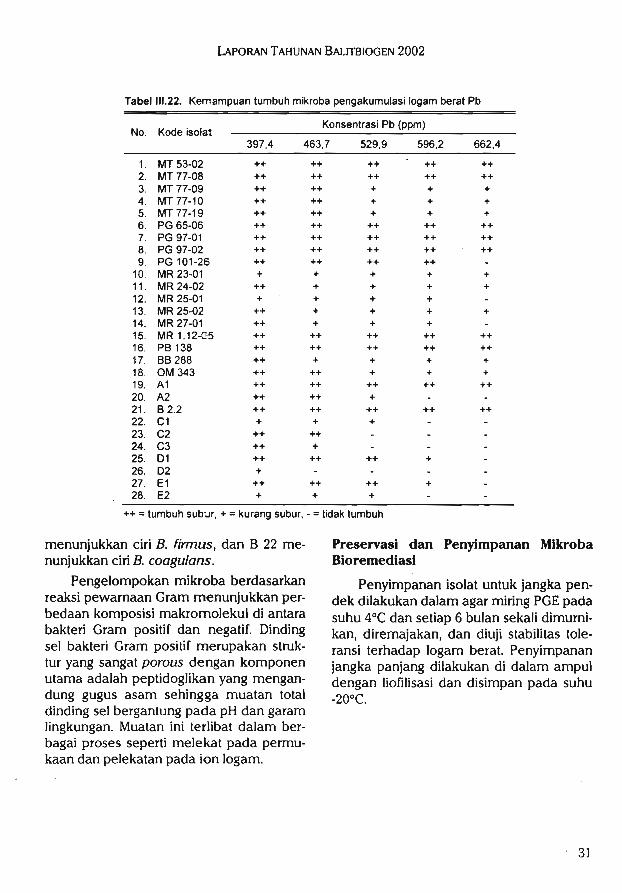
31
Preservasi dan Penyimpanan MikrobaBioremediasi
Penyimpanan isolat untuk jangka pen-dek dilakukan dalam agar miring PGE padasuhu 4C dan setiap 6 bulan sekali dimurni-kan, diremajakan, dan diuji stabilitas tole-ransi terhadap logam berat. Penyimpanan
jangka panjang dilakukan di dalam ampuldengan liofilisasi dan disimpan pada suhu-20C.
menunjukkan ciri B. firmus, dan B 22 me-
nunjukkan ciri B. coagulans.
Pengelompokan mikroba berdasarkanreaksi pewarnaan Gram menunjukkan per-
bedaan komposisi makromolekul di antarabakteri Gram positif dan negatif. Dindingsel bakteri Gram positif merupakan struk-tur yang sangat porous dengan komponen
utama adalah peptidoglikan yang mengan-dung gugus asam sehingga muatan totaldinding sel bergantung pada pH dan garamlingkungan. Muatan ini terlibat dalam ber-
bagai proses seperti melekat pada permu-
kaan dan pelekatan pada ion logam.
++ = tumbuh subur, + = kurang subur, - = tidak tumbuh
1.MT 53-022.MT 77-083.MT 77-094.MT 77-105.MT77-196.PG 65-067.PG 97-018.PG 97-029.PG 101-26
10.MR 23-0111.MR 24-0212.MR 25-0113.MR 25-0214.MR 27-0115.MR 1.12-0516.PB13817.BB28818.OM34319.A120.A221.B2.222.C123.C224.C325.D126.D227.E128.E2
662,4596,2529,9463,7397,4
Konsentrasi Pb (ppm)No. Kode isolat
Tabel 111.22. Kemampuan tumbuh mikroba pengakumulasi logam berat Pb
Laporan Tahunan Balitbiogen 2002
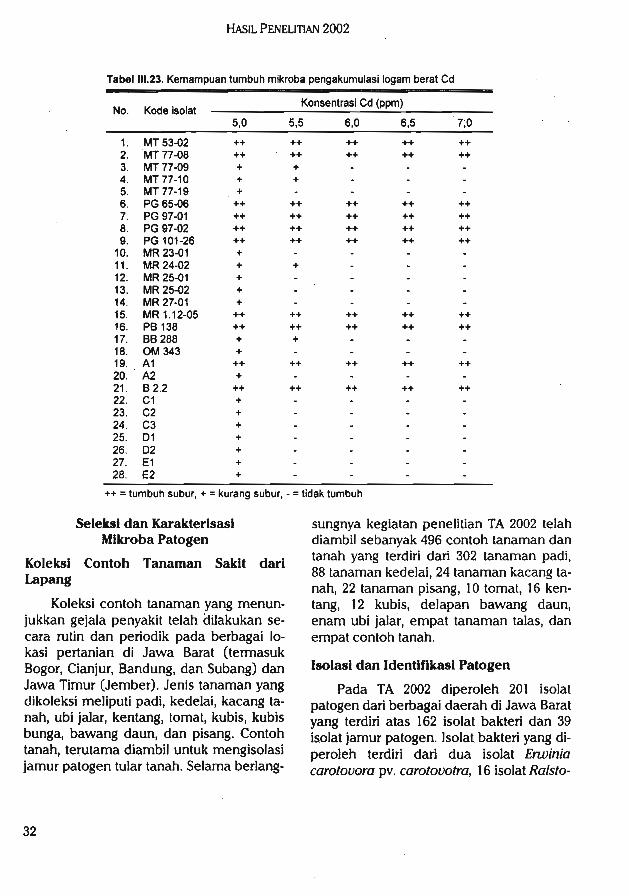
32
sungnya kegiatan penelitian TA 2002 telahdiambil sebanyak 496 contoh tanaman dantanah yang terdiri dari 302 tanaman padi,88 tanaman kedelai, 24 tanaman kacang tanah, 22 tanaman pisang, 10 tomat, 16 kentang, 12 kubis, delapan bawang daun,enam ubi jalar, empat tanaman talas, danempat contoh tanah.
Isolasi dan Identifikasi Patogen
Pada TA 2002 diperoleh 201 isolatpatogen dari berbagai daerah di Jawa Baratyang terdiri atas 162 isolat bakteri dan 39isolat jamur patogen. Isolat bakteri yang diperoleh terdiri dari dua isolat Erwiniacarotovora pv. carotovotra, 16 isolat Ralsto-
Seleksi dan KarakterisasiMikroba Patogen
Koleksi Contoh Tanaman Sakit dariLapang
Koleksi contoh tanaman yang menun-jukkan gejala penyakit telah dilakukan se-cara rutin dan periodik pada berbagai lo-kasi pertanian di Jawa Barat (termasukBogor, Cianjur, Bandung, dan Subang) danJawa Timur (Jember). Jenis tanaman yangdikoleksi meliputi padi, kedelai, kacang ta-nah, ubi jalar, kentang, tomat, kubis, kubisbunga, bawang daun, dan pisang. Contohtanah, terutama diambil untuk mengisolasijamur patogen tular tanah. Selama berlang-
++ = tumbuh subur, + = kurang subur, - = tidak tumbuh
28. E2E1D2D1C3C2C1B2.2A2A1OM343BB288PB138MR 1.12-05MR 27-01MR 25-02MR 25-01MR 24-02MR 23-01PG 101-26PG 97-02PG 97-01PG 65-06MT 77-19MT 77-10MT 77-09MT 77-08MT 53-02
27.26.25.24.23.22.21.20.19.18.17.16.15.14.13.12.11.10.9.8.7.6.5.4.3.2.1.
7,06,56,05,55,0
Konsentrasi Cd (ppm)No. Kode isolat
Tabel 111.23. Kemampuan tumbuh mikroba pengakumulasi logam berat Cd
Hasil Penelitian 2002

33
Preservasi
Isolat bakteri patogen umumnya di-simpan di dalam ampul berukuran 1 mldalam kondisi kering beku di kulkas ataufreezer bersuhu -20C. Tiap ampul diberi label dan diisi 0,5 ml suspensi biakan bakteriumur 48 jam dengan kerapatan 108 CFU/ml. Selain disimpan dalam bentuk keringbeku,' khusus isolat R. solanacearum jugadisimpan dalam tabung Eppendorf berisiakuades steril. Tiap isolat patogen disimpan rangkap lima.
* Belum dlkarakterisasi
nia solanacearum, dua isolat Pseudomo-
nas syringae pv. glycinea, 10 isolat Xantho-monas campestris pv. campestris, 10 isolat
X. campestris pv. glycines, dan 122 isolat X.oryzae pv. oryzae. Jamur patogen yang di-koleksi 18 isolat Rhizoctonia solani, duaisolat Fusarium oxysporum var. cubensis,dua isolat F. oxysporum, dua isolat F.
solani, enam isolat Alternaria porri, limaisolat Elsinoe iwatae, dua isolat Cercosporabataticola, dan dua isolat Altemaria coloca-siae (Tabel IH.25).
* Batang pendek* Batang* Batang pendek* Batang pendek* Batang* Batang+ Batang+ Batang+ Batang
Sel khamir* Batang pendek+ Batang
Batang+ Batang+ Batang+ Batang+ Batang
Batang* Batang+ Batang+ Batang
Batang+ Batang+ Batang* Batang
BatangBatang
+ Batang+ Batang+- Batang
Batang
Endospora Bentuk sel
++--++++++-+++++++++++++
++++++
Pewarnaan Gram
PD10ABAB5.1Bd-4Bd-3A1C2PB138B22BB288OM343MP1.2-05MR1.12-05PG121-02PG101-26PG97-02PG97-01PG65-06PG65-01PG38-01MR27-01MR25-02MR24-01MR23-01MT53-02MT53-01MT54-01MT77-19MT77-10MT77-09MT77-08MT77-07
Isolat
31.30.29.28.27.26.25.24.23.22.21.20.19.18.17.16.15.14.13.12.11.10.9.8.7.6.5.4.3.2.1.
No.
Tabel HI.24. Hasil karakteristik beberapa isolat bioremediasi
Laporan Tahunan Balitbiogen 2002

34
isolat B. polymixa, 17 isolat Pseudomonasfluorescens, 25 isolat khamir Saccharomy-ces spp., dan 16 isolat Trichoderma spp.
Preservasi Isolat
Sebagian isolat mikroba antagonisyang telah dikarakterisasi disimpan untukjangka menengah menggunakan teknikpenyimpanan di dalam minyak mineral.
Penyimpanan jangka panjang di dalamampul menggunakan teknik kering beku.Bakteri B. subtilis, B. polymixa, B. thuringi-
ensis, dan P. fluorescens disimpan di dalamampul. Setiap kemasan isolat disimpanrangkap lima. Isolat yang disimpan untukjangka menengah ditempatkan pada suhuruang, sedangkan untuk jangka panjang disimpan di dalam freezer dengan suhu
-20C.
Uji Viabilitas Mikroba
Belum semua koleksi isolat mikrobaantagonis yang telah dikoleksi pada tahun
Uji Viabilitas
Seluruh isolat patogen yang sudah diujiviabilitasnya, baik yang disimpan dalamampul maupun dalam akuades steril (/?.solanacearum) masih hidup dan ciri-ciri-nya masih tetap dipertahankan. Hal ini me-
nunjukkan bahwa teknik preservasi dancara penyimpanannya cukup baik.
Seleksi dan KarakterisasiMikroba Antagonis
Koleksi, Isolasi, dan Identifikasi
Selama tahun 2002 telah dikoleksi 350contoh tanah, air, dan tanaman dari berba-
gai daerah di Jawa Barat. Isolasi dan identifikasi ciri-ciri mikroba antagonis serta ke-mampuan antagonismenya terhadap X.oryzae pv. oryzae (Xoo) dan Rhizoctonia
solani (RhS) memperoleh 76 isolat mikroba antagonis yang memiliki potensi untukpengendalian kedua jenis patogen utamatersebut. Ke-76 isolat mikroba antagonis
terdiri dari 13 isolat Bacillus subtilis, lima
201
182225262
12210101622
Banyaknya isolat
Jumlah
Rhizoctonia solaniF. solaniF. oxysporumFusarium oxysporum var. cubensisElsinoe iwataeCercospora bataticolaA. porriAlternaria colocaciae
JamurX. oryzae pv. oryzaeX. campestris pv. glycinesXanthomonas campesths pv. campestrisRalstonia solanacearumPseudomonas symingae pv. glycineaErwinia carotovora pv. carotovora
Bakteri
Jenis patogen
Tabel 111.25. Jenis dan banyaknya isolat bakteri dan jamur patogentanaman pangan yang telah dikoleksi dan dikarakterisasipada tahun 2002
Hasil Penelitian 2002

35
(kabupaten dan provinsi), inang isolat, danwaktu isolasi. Data lain yang lebih spesifikpada masing-masing mikroba adalaha.Mikroba penyubur tanah
Bentuk koloni (BK), tipe koloni (TK),reaksi metabolisme (RM), reaksi terha-dap antibiotik (RA), nilai keefektifan(NE), kemampuan kompetisi (KK), danketahanan terhadap kemasaman tanah
(TA).b.Mikroba perombak bahan organik
Bentuk koloni (BK), jari-jari koloni (JJ),reaksi metabolisme (RM), dan keefektifan dalam merombak bahan organik(BO).
c.Mikroba bioremediasiBentuk koloni (BK), kemampuan meng-
akumulasi mangan (Mn), besi (Fe), seng(Zn), timbal (Pb), kadmium (Cd), dankemampuan merombak fenol (FOH).
d.Mikroba hidrolitikBentuk koloni (BK), bentuk sel (BS),reaksi gram (RG), endospora (E), motili-tas (M), dan katalase (K).
e.Mikroba pengendali hama dan penyakittanamanBentuk koloni (BK), nilai keefektifan(NE), dan antagonis terhadap patogen(AP).
f.Mikroba patogen tanamanBentuk koloni (BK), nilai keefektifan(NE), dan tanaman inang.
Banyaknya isolat yang telah disusun kedalam database pada setiap jenis mikrobadisajikan pada Tabel III.26.
Dari database yang telah direkam, di-peroleh banyak isolat yang memiliki sifatpenting seperti efektif dalam menambatnitrogen, merombak bahan organik, me-ningkatkan mutu pangan, mengakumulasi
logam berat, serta tahan terhadap kema
saman tanah dan sangat kompetitif terhadap strain yang lain (Tabel 111.27).
2001 dapat diuji viabilitasnya karena jum-lahnya yang cukup banyak, sehingga perludilakukan secara rutin dan bertahap, seba-gai upaya pemeliharaan. Banyaknya mikro-
ba yang telah diuji viabilitasnya 61 isolatyang terdiri dari dua isolat B. subtilis, 40isolat B. thuringiensis, 12 isolat P. fluores-cens, lima isolat T. harzianum, dan dua
isolat 5. cereviciae. Viabilitas seluruh isolatyang telah diuji masih baik, koloni mikrobamasih hidup dan tumbuh dengan baik. Halini menunjukkan bahwa teknik penyim-panan yang digunakan cukup baik.
Penyimpanan, Pemeliharaan, dan
Pengembangan Database Plasma NutfahMikroba Pertanian
Penyimpanan Mikroba dalam KondisiKering Beku
Sebanyak 61 isolat Rhizobium, 660isolat B. japonicum, 10 isolat Azospirillum,tujuh isolat bakteri endofitik, 23 isolatmikroba remediasi, dua isolat mikrobapelarut phosphate, 13 isolat virus patogentanaman, 25 isolat P. syringae, 10 isolatbakteri hawar daun kedelai (X. campestris
pv. glycines), dan lima isolat bakteriErwinia carotovora pv. carotovora telah
disimpan dalam kondisi kering beku.Masing-masing isolat disimpan dalamampul sebanyak 3-5 ulangan.
Pengembangan Database Plasma NutfahMikroba Pertanian
Data karakter mikroba pertanian yang
disusun ke dalam database meliputi ber-bagai koleksi mikroba penyubur tanah,mikroba perombak bahan organik, mikroba bioremediasi, mikroba hidrolitik, mikroba pengendali hama dan penyakit tanaman, serta mikroba patogen tanaman.
Data paspor dari setiap mikroba pertanian yang dikoleksi meliputi asal isolat
Laporan Tahunan Balitbiogen 2002

36
B. coagulans El.4A
FCB 7/2, FCB 63/2, FCB 63/3, FCB 152/2, FCB179, FCB 189/1, FCB 189/2, FCB 190/1, FCB193/2, FCB 193/3, FCB 206/3, 16D, R2. R 61, L17-1FCB 26, FCB 44, FCB 152/2, FCB 178, FCB189/2, FCB 206, FCB 227, FCB 229, FCB 230,TKG4BFCB 152, FCB 152/1, FCB 152/2, FCB 152/3,FCB 179, FCB 179/1, FCB 179/2, FCB 179/355.13,96.12,76.22
PG65-06, MR25-01, MT53-02, MT77-08, MT77-09, MT77-10, MT77-19, PG97-01, PG97-02,PG101-26, MR27-0PB138, BB288, OM343B. drculans 9b3
Tahan terhadap faktorkemasaman tanah
Sangat kompetitifterhadap strain lainAktivitas seiulase tinggi
Efektif mengakumulasi4-5 macam logam berat(Mn, Fe, Zn, Pb, Cd)Efektif merombak fenol
Bacillus Penghasil protease yangefektif menghambatlipase pada bekatulPenghasil fitase yangefektif menghidrolisisfitat pada bekatul
4. Hidrolitik
Perombak bahanorganikBioremediasi3.
2.
Rhizobium Sangat efektif dalammenambat nitrogen
1. Penyubur tanah
IsolatSifat pentingNo. Mikroba pertanian
Tabel 111.27. Isolat yang mempunyai sifat penting
1333Jumlah
51025761325
1881228
5111
751329
94
205
1319
68661
Erwinia carotovora pv. carotovoraXanthomonas campestris pv. glycinesP. syringae pv. glycineaRalstonia solanacearumVirusSaccharomyces spp.Trichoderma spp.B. thuringiensisPseudomonas fluorescensB. subtilisVirusBakteri asam laktatB. coagulansBacillus drculans
Bakteri lignoselulolitikTrichodermaMycorrhizaBakteri pelarut PBakteri diazotrop endofitikAzotobacterAzospirillumShinorhizobiumBradyrhizobium japonicumRhizobium
Patogen
Pengendali hayati
HidrolitikBioremediasi
Perombak bahan organik
Penyubur tanah
6.
5.
4.3.
2.
1.
Banyaknya isolatNama mikrobaNo. Mikroba pertanian
Tabel 111.26. Isolat mikroba pertanian yang telah dimasukkan ke dalam komputer
Hasil Penelitian 2002

37
Pengujian Sifat Toleransi Tanaman Paditerhadap Kekeringan pada Populasi F7
(IR64 x Cabacu)
Hasil skoring 200 galur F7 (IR64 xCabacu) pada dua tahap perkembangantanaman yang berbeda, diketahui bahwabeberapa galur toleran pada kedua peng-amatan skoring, peka pada kedua peng-
amatan skoring, dan toleransinya tidakkonsisten pada kedua pengamatan. Datapengamatan skor kekeringan daun padapopulasi F7 turunan persilangan IR64 xCabacu disajikan pada Tabel 111.28.
Uji Rumah Kaca untuk Toleransiterhadap Kekeringan pada Populasi F7
(IR64xIRAT112)Daya tembus akar dicerminkan oleh
data banyaknya akar yang mampu menem-bus lilin. Dari 117 galur F7 (IR64 x IRAT112)yang ditanam dalam pot berlapis lilin diketahui bahwa banyaknya akar yang mampumenembus lilin berkisar antara 1-11. Hal inimencerminkan bahwa kemampuan akar
menembus lilin yang berbeda di antaragalur F7. Data hasil skrining daya tembusakar di rumah kaca disajikan pada Tabel111.29.
Analisis AFLP dan Mikrosatelit untukPemetaan Markah Molekuler untuk Sifat
Toleransi terhadap Kekeringan
Hasil analisis mikrosatelit menggunakan 256 pasang primer diperoleh 157 pa-sang primer yang menghasilkan polimorfis-me antara tetua toleran (Cabacu) dan tetuapeka (IR64) terhadap kekeringan.
Dari 157 pasang primer, dipilih 30 pa-sang primer yang menghasilkan jarak pitadari kedua tetua minimal 7 bp, dan digunakan untuk menguji genotipe markah dari192 galur F7 (IR64 x Cabacu).
Berdasarkan variabel plasma nutfahmikroba yang telah disusun ke dalam database, sebagian besar isolat belum dikarak-terisasi dan dievaluasi. Hanya 43 isolatRhizobium yang sudah lengkap, sehinggakarakterisasi dan evaluasi terhadap isolatlain perlu dilanjutkan untuk melengkapidatabase yang telah dikembangkan.
IDENTIFIKASI DAN APLIKASI MARKAHMOLEKULER DALAM PEMULIAAN
TANAMANMasdiar Bustamam, Sugiono Moeljopawiro,Didi Suardi, Erwina Lubis, Haeni Purwanti,Kumiawan R. Trijatmiko, Joko Prasetiyono,
Reflinur, Tasliah, Ahmad Dadang,Alina Akhdiya, dan Sri G. Budiarti
Perkembangan ilmu-ilmu dasar, me-
nyediakan suatu cara pendeteksian genetikdengan menggunakan markah molekuler.
Penggunaan markah molekuler dapatmemberikan suatu gambaran dengan ting-
kat resolusi yang tinggi dari perbedaan genetik di antara individu tanaman dan dapatdigunakan sebagai alat dalam pemuliaantanaman untuk seleksi dengan penandaan
gen tertentu.
Gen pengendali sifat kuantitatif sulitdiidentifikasi dan diintrogresi ke tanamanlain karena gen ini terletak di berbagai lo-kus pada beberapa kromosom. Penggunaan markah molekuler telah membuka ke-
sempatan untuk mengidentifikasi lokuspengatur sifat kuantitatif dan memungkin-kan para pemulia untuk memindahkan sifat kuantitatif tertentu ke tanaman lain dengan bantuan markah molekuler yang ter-
paut dengan sifat tersebut yang dikenaldengan MAS.
Laporan Tahunan Balitbiogen 2002
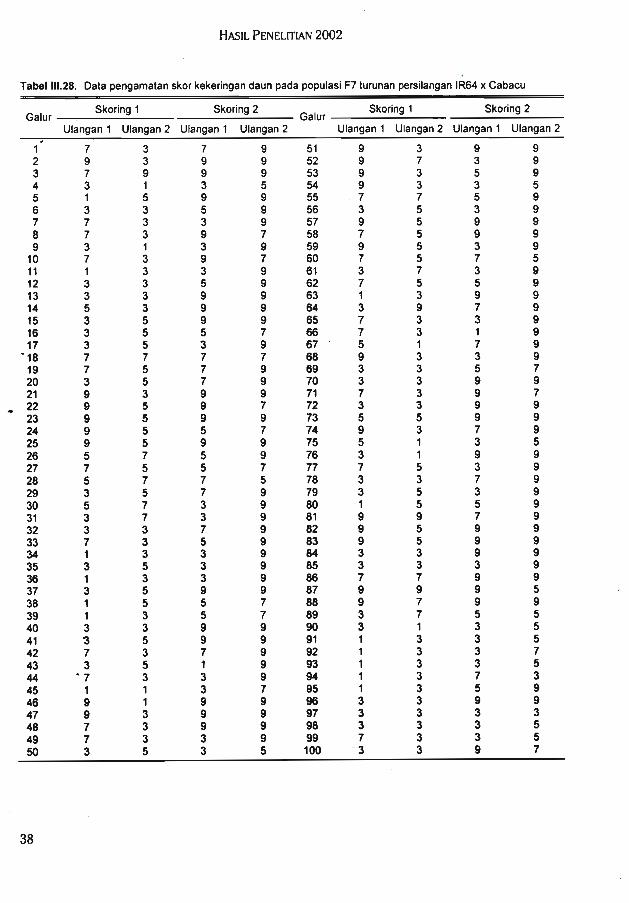
38
99959999959999999979799959999999999959555753993557
93535399373597317359999739373579993999533337593339
373375555575393313333353115355955337977133333333333
73331111133997339991337359ut3733957731737979379999
10099989796959493929190898887868584838281807978777675747372717069686766656463626160595857565554535251
5999979999977999999999at7997979997979999979799959997
9939539393599953777999595577337533395599713399933
339153331333335557553555575757733353553353531133353
77991
'7373311313173353ot7599999377333(it3317377313797
50494847464544434241403938373635343332313029282726252423222120191817161514131211109876ot4321
Ulangan 1 Ulangan 2 Ulangan 1 Ulangan 2
Skoring 2Skoring 1Galur
Ulangan 1 Ulangan 2 Ulangan 1 Ulangan 2
Skoring 2Skoring 1Galur
Tabel 111.28. Data pengamatan skor kekeringan daun pada populasi F7 turunan persilangan IR64 x Cabacu
Hasil Penelitian 2002
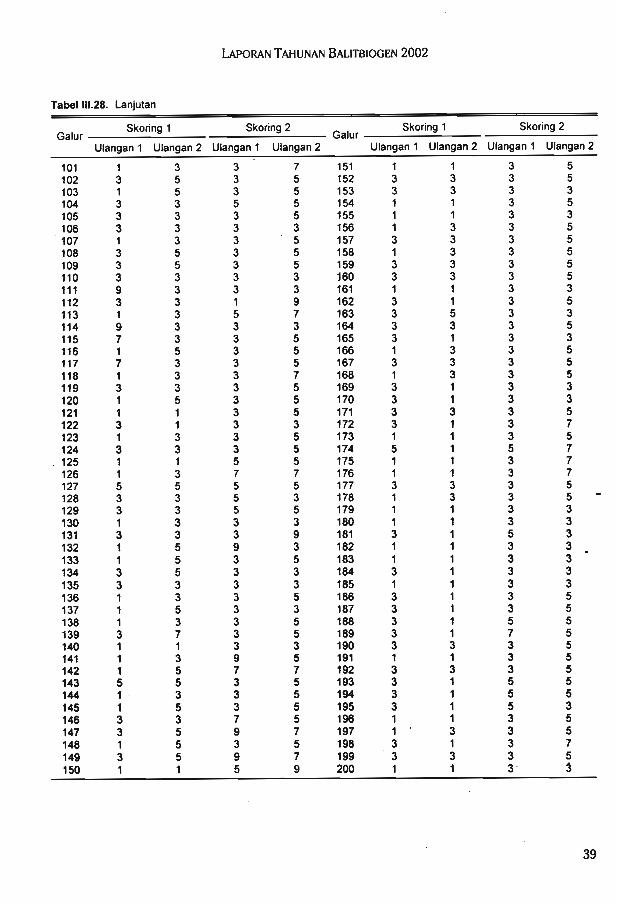
39
35755355555555533333335577757533555353535555535355
Ulangan 2
33333555333753333335333333533333333333333333333333
Ulangan 1
Skoring 2
13131111313111111111113311111311333135113333311331
Ulangan 2
1331 •1333313333313113111311513333131333313313111331
Ulangan 1
Skoring 1
200199198197196195194193192191190189188187186185184183182181180179178177176175174173172171170169168167166165164163162161160159158157156155154153152151
Gslur
97575555753553533539353575553555755537933555355557
Ulangan 2
59397333793333333393355575333333333335133333335333
Ulangan 1
Skoring 2
1 1) 5
5) 5J 3
53
) 5531
J 7353
J 3
•:
I
•111;3 5
55
J 33
! 3> 3> 5
3133
! 115
11j1I;
1
•1;113 3
3r 3
5' 3
1;1•9 3
3J 3) 31 35 5i 5
3J 35 3J 3
5) 5
3
1;i
^;
;;•*;
Ulangan 1 Uiangan 2
Skoring 1
150149148147146145144143142141140139138137136135134133132131130129128127126
. 125124123122121120119118117116115114113112111110109108107106105104103102101
OdlUl
Tabel 111.28. Lanjutan
Laporan Tahunan Balitbiogen 2002
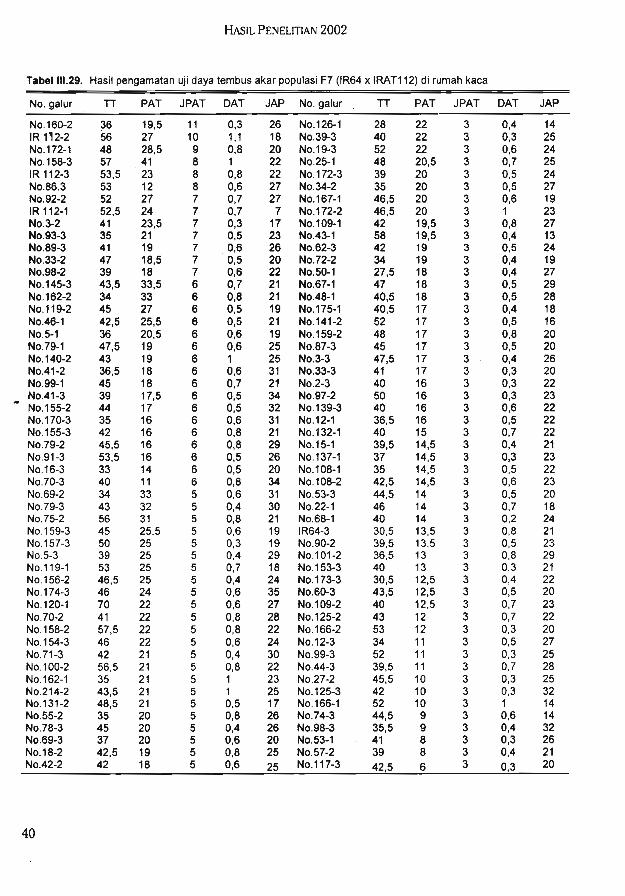
40
2021263214143225282527202223202221292321241820232223212222222322202620201618282927192413272319272425242514
JAP
0,30,40,30,40,610,30,30,70,30,50,30,70,70,50,40,30,80,50,80,20,70,50,60,50,30,40,70,50,60,30,30,30,40,50,80,50,40,50,50,40,40,50,40,810,60,50,50,70,60,30,4
DAT
33333333333333333333333333333333333333333333333333333
JPAT
68899
101010111111121212,512,512,5131313,513,514141414,514,514,514,51516161616171717171717181818191919,519,52020202020,5222222
PAT
42,5394135,544,5524245,539,5523453434043,530,54036,539,530,5404644,542,5353739,54036,54050404147,545485240,540,54727,53442584246,546,5353948524028
TT
No. 117-3No.57-2No.53-1No.98-3No.74-3No. 166-1No. 125-3No.27-2No.44-3No.99-3No. 12-3No. 166-2No. 125-2No. 109-2No.60-3No. 173-3No. 153-3No.101-2No.90-2IR64-3No.68-1No.22-1No. 53-3No. 108-2No. 108-1No. 137-1No. 15-1No. 132-1No. 12-1No. 139-3No.97-2No.2-3No.33-3No.3-3No.87-3No. 159-2No.141-2No. 175-1No.48-1No.67-1No.50-1No.72-2No.62-3No.43-1No. 109-1No. 172-2No. 167-1No.34-2No. 172-3No.25-1No. 19-3No.39-3No. 126-1
No. galur
252520262617252322302422282735241829191921303134202629213132342131252519211921212220262317
727272222201826
JAP
0,60,80,60,40,80,5110,80,40,60,80,80,60,60,40,70,40,30,60,80,40,60,80,50,50,80,80,60,50,50,70,610,60,60,50,50,80,70,60,50,60,50,30,70,70,60,810,81,10,3
DAT
555555555555555555555556666666666666666677777778889
1011
JPAT
1819202020212121212122222222242525252525,53132331114161616161717,51818191920,525,5273333,51818,5192123,5242712234128,52719,5
PAT
4242,537453548,543,53556,5424657,541704646,553395045564334403353,545,5423544394536,54347,53642,5453443,5394741354152,5525353,557485636
TT
No.42-2No. 18-2No.69-3No.78-3No. 55-2No.131-2No.214-2No.162-1No. 100-2No.71-3No. 154-3No. 158-2No.70-2No. 120-1No. 174-3No. 156-2No.119-1No.5-3No. 157-3No. 159-3No.75-2No.79-3No.69-2No.70-3No. 16-3No.91-3No.79-2No. 155-3No. 170-3No. 155-2No.41-3No.99-1No.41-2No. 140-2No.79-1No. 5-1No.46-1No. 119-2No. 162-2No. 145-3No.98-2No.33-2No 89-3No.93-3No.3-2IR 112-1No.92-2No.86.3IR 112-3No. 158-3No. 172-1IR 112-2No. 160-2
No. galur
Tabel 111.29. Hasil pengamatan uji daya tembus akar populasi F7 (IR64 x IRAT112) di rumah kaca
Hasil Penelitian 2002
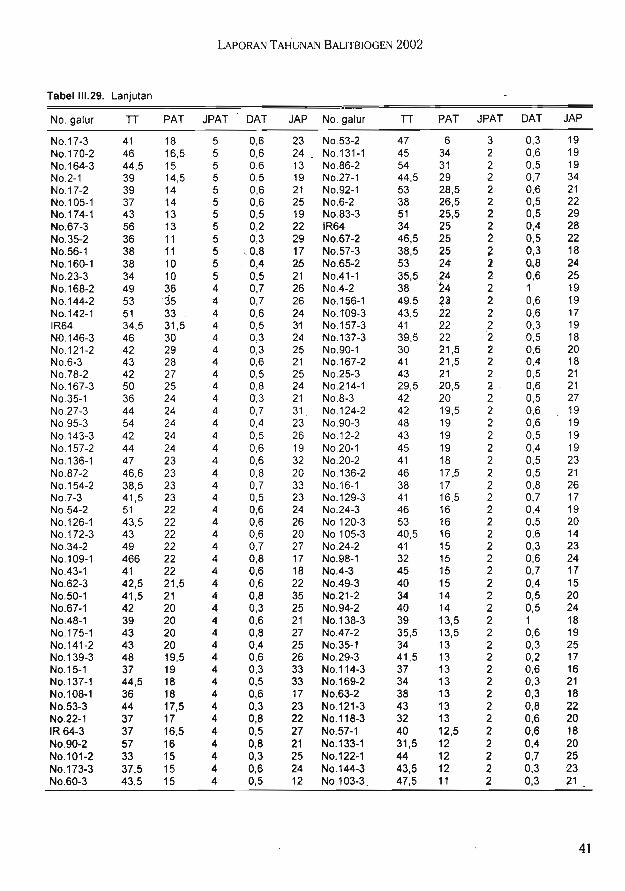
41
2123252018202218211617251918242015172423142019172621231919191927212118201819171919252418222829222134191919
JAP
0,30,30,70,40,60,60,80,30,30,60,20,30,610,50,50,40,70,60,30,60,50,40,70,80,50,50,40,50,60,60,50,60,50,40,60,50,30,60,610,60,80,30,50,40,50,50,60,70,50,60,3
DAT
22222222222222222222222222222222222222222222222222223
JPAT
1112121212,51313131313131313,513,514141515151516161616,51717^51819191919,52020,52121,521,5222222?3^4
242425252525,526,528,52931346
PAT
47,543,54431,540324338343741,53435,53940344045324140,5534641384641454348424229,543413039,54143,549,53835,55338,546,53451385344,5544547
TT
No. 103-3No. 144-3No. 122-1No. 133-1No.57-1No. 118-3No. 121-3No.63-2No. 169-2No.114-3No.29-3No.35-1No.47-2No. 138-3No.94-2No.21-2No.49-3No.4-3No.98-1No.24-2No. 105-3No. 120-3No.24-3No. 129-3No.16-1No. 136-2No.20-2No.20-1No. 12-2No.90-3No. 124-2No.8-3No.214-1No.25-3No. 167-2No.90-1No. 137-3No. 157-3No. 109-3No. 156-1No.4-2No.41-1No.65-2No.57-3No.67-2IR64No.83-3No.6-2No.92-1No.27-1No.86-2No.131-1No.53-2
No. galur
1224252127222317333326252721253522181727202624233320321926233121242521252431242626212517292219252119132423
JAP
0,50,60,30,80,50,80,30,60,50,30,60,40,80,60,30,80,60,60,80,70,60,60,60,50,70,80,60,60,50,40,70,30,80,50,60,30,30,50,60,70,70,50,4
,0,80,30,20,50,60,60,50,60,60,6
DAT
44444444444444444444444444444444444444444555555555555
JPAT
1515151616,51717,518181919,5202020202121,5222222222222232323232424242424252728293031,5333536101011111313141414,51516,518
PAT
43,537,533573737443644,537484343394241,542,541466494343,55141,538,546,6474442544436504243424634,551534934383836564337393944,54641
TT
No.60-3No. 173-3No. 101-2No.90-2IR 64-3No.22-1No.53-3No. 108-1No. 137-1No.15-1No. 139-3No. 141-2No. 175-1No.48-1No.67-1No.50-1No.62-3No.43-1No. 109-1No.34-2No. 172-3No.126-1No. 54-2No.7-3No. 154-2No.87-2No.136-1No. 157-2No. 143-3No.95-3No.27-3No.35-1No. 167-3No.78-2No.6-3No.121-2NO. 146-3IR64No.142-1No. 144-2No. 168-2No.23-3No. 160-1No.56-1No.35-2No.67-3No. 174-1No.105-1No. 17-2No.2-1No. 164-3No. 170-2No. 17-3
No. galur
Tabel 111.29. Lanjutan
Laporan Tahunan Balitbiogen 2002
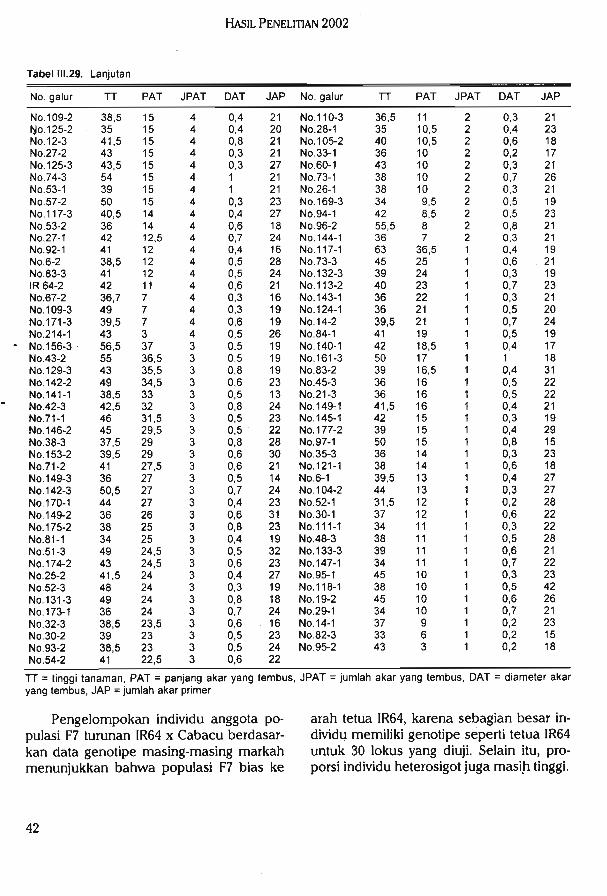
42
arah tetua IR64, karena sebagian besar individu memiliki genotipe seperti tetua IR64untuk 30 lokus yang diuji. Selain itu, pro-porsi individu heterosigot juga masih tinggi.
Pengelompokan individu anggota po-pulasi F7 turunan IR64 x Cabacu berdasar-kan data genotipe masing-masing markahmenunjukkan bahwa populasi F7 bias ke
TT = tinggi tanaman, PAT = panjang akar yang tembus, JPAT = jumlah akar yang tembus, DAT = diameter akaryang tembus, JAP = jumlah akar primer
181523212642232221282222282727182315291921222231181719242021231921192121231921262117182321
JAP
0,20,20,20,70,60,50,30,70,60,50,30,60,20,30,40,60,30,80,40,30,40,50,50,410,40,50,70,50,30,70,30,60,40,30,80,50,50,30,70,30,20,60,40,3
DAT
111111111111111111111111111111111122222222222
JPAT
369
101010101111111112121313141415151516161616,51718,51921212223242536,5
788,59,5
1010101010,510,511
PAT
43333734453845343938343731,54439,5383650394241,536363950424139,53636403945633655,5423438384336403536,5
TT
No.95-2No.82-3No.14-1No.29-1No. 19-2No.118-1No.95-1No. 147-1No. 133-3No.48-3No.111-1No.30-1No.52-1No. 104-2No.6-1No.121-1No. 35-3No.97-1No. 177-2No. 145-1No. 149-1No.21-3No.45-3No.83-2No. 161-3No. 140-1No.84-1No. 14-2No. 124-1No. 143-1No. 113-2No. 132-3No.73-3No.117-1No.144-1No.96-2No.94-1No. 169-3No.26-1No.73-1No.60-1No.33-1No. 105-2No.28-1No. 110-3
No. galur
222423
. 16241819272332192331232414213028222324132319191926191916212428162418272321212721212021
JAP
0,60,50,50,60,70,80,30,40,60,50,40,80,60,40,70,50,60,60,80,50,50,80,50,60,80,50,50,50,60,30,30,60,50,50,40,70,60,40,3110,30,30,80,40,4
DAT
3333333333333333333333333334444444444444444444
JPAT
22,5232323,52424242424,524,525252627272727,5292929,531,5323334,535,536,53737771112121212,514141515151515151515
PAT
4138,53938,536494841,543493438364450,5364139,537,5454642,538,549435556,54339,54936,7424138,541423640,550395443,54341,53538,5
TT
No.54-2No.93-2No.30-2No.32-3No. 173-1No.131-3No.52-3No.25-2No. 174-2No.51-3No.81-1No. 175-2No. 149-2No. 170-1No. 142-3No. 149-3No.71-2No. 153-2No.38-3No. 146-2No.71-1No.42-3No.141-1No. 142-2No. 129-3No.43-2No. 156-3No.214-1No.171-3No. 109-3No.67-2IR 64-2No.83-3No.6-2No.92-1No.27-1No.53-2No. 117-3No.57-2No.53-1No.74-3No. 125-3No.27-2No. 12-3No. 125-2 -No. 109-2
No. galur
Tabel 111.29. Lanjutan
Hasil Penelitian 2002

43
Identifikasi Markah Molekuler untukKetahanan terhadap Bias
Pada tahun 2002 telah dilakukan analisis keterpautan markah mikrosatelit terhadap sifat ketahanan penyakit bias menggunakan program MAPMAKER. Dari 15 markah mikrosatelit yang dianalisis, diperoleh12 primer mikrosatelit yang terpaut dengansifat ketahanan terhadap bias, yaitu RM 151dan RM 104 pada kromosom 1; RM 130 danRM 251 pada kromosom 3; RM 197, RM 190,RM 276 dan RM 225 pada kromosom 6;serta RM 21, RM 254, RM 224, dan RM 187pada kromosom 11.
Analisis Populasi Inbrida Rekombinandengan Beberapa Isolat Bias di Rumah
Kaca (Monosiklik dan Polisiklik)
Pada tahun anggaran 2002 telah dilakukan pengujian ketahanan 200 galurinbrida rekombinan hasil persilanganDanau Tempe x Kencana Bali terhadapdua ras bias, yaitu ras 173 dan ras 033. Hasilpengujian terhadap ras 173 menunjukanbahwa 103 galur (53,9%) bereaksi tahan(T), 68 galur (35,6%) bereaksi agak tahan(AT), tujuh galur (3,6%) bereaksi tidak tahan (P), dan 22 galur (11%) tidak tumbuh.Hasil pengujian terhadap ras 033 menunjukkan bahwa 74 galur (37%) bereaksitahan, 48 galur (24%) bereaksi agak tahan,31 galur (15,5%) bereaksi tidak tahan, 43galur (21,5%) bereaksi sangat rentan, danempat galur (2%) tidak tumbuh.
Markah DNA untuk Deteksi Dini PenyakitBulai pada Tanaman Jagung yang Sakit
Hasil isolasi DNA genom cendawanPeronosclerospora maydis menggunakanmetode CTAB/NaCl dari IRRI belum me-muaskan karena sebagian besar terdegra-
dasi (Gambar III.8). Oleh karena itu, perludilakukan isolasi DNA lagi menggunakan
RM 566 menghasilkan pita polimorfik yang dapat diskor, sedangkan GT 167 menghasilkan pita monomor-fis yang tidak bisa diskor
Gambar III.7. Pola pita 94 galur F2 dengan menggunakan primer (A) RM 566 dan B (GT 167)
IITA131
Hasil analisis AFLP menggunakan duapasang primer selektif (E45-M59 dan E45-M62) diperoleh 33 markah AFLP yang poli-morfik pada kedua tetua (IR64 dan Caba-cu). Ke-33 markah ini diskoring pada 186individu F7 turunannya. Hasil analisis de-ngan software Genescan dan Genotyper
menghasilkan data genotipe markah darikedua tetua dan 186 individu F7 turunannya. Dari pengelompokan banyaknya individu berdasarkan genotipenya dapat dike-tahui adanya kecenderungan yang samaseperti data markah mikrosatelit, yaitu ter-
jadinya bias ke arah tetua IR64 (sebagianbesar individu F7 memiliki genotipe sepertiIR64).
Analisis Segregasi F2 PersilanganDupa x ITA131 dan Uji Fenotipik pada
Populasi F3
Dari 94 galur F2 yang diamplifikasimenggunakan 74 primer mikrosatelit, diperoleh 68 primer menghasilkan pola pitayang heterosigot (dapat diskor), 2 primermenghasilkan pola pita monorfis (tidakdapat diskor), dan 4 primer menghasilkanpola pita yang banyak (tidak dapat diskor).Gambar III.7 menunjukkan hasil amplikasiyang telah diperoleh.
Laporan Tahunan Balitbiogen 2002

44
Gambar III.9. Hasil isolasi DNA dari seedling jagung
PEMANFAATAN TEKNIK KULTURJARINGAN DAN MOLEKULER UNTUKPERBAIKAN GENETIK PADI DENGAN
MENGGUNAKAN SPESIES LIARTiur S. Silitonga, Buang Abdullah,
Tintin Suhartini, Ida H, Somantri, Didi Suardi,Masdiar Bustamam, Bahagiawati,
A. Dinar Ambarwati, Iswari S. Dewi,Anggiani Nasution, Mukelar Amir, dan Tasliah
Perkembangan bioteknologi yang pe-sat menghasilkan teknik yang memungkin-kan introgresi gen dari kerabat jauh padidan cara seleksi yang lebih cepat dan tepatdiibandingkan dengan cara konvensional.Teknik kultur jaringan dapat digunakan untuk menyelamatkan embrio muda dari per-
silangan interspesifik yang dikenal dengankultur embrio dan teknik markah moleku-
1 = ulangan 1,2 = ulangan 2, 3 = ulangan 3,4 = ulangan 4<
Gambar III.8. Hasil isolasi DNA P. maydis menggunakan metode CTAB/NaCl dari IRRI
metode lain sehingga diperlukan sporacendawan P. maydis dalam jumlah yangcukup. Tetapi karena sifatnya parasit obli-
gat dan pengaruh cuaca dominan terhadapsporulasi cendawan ini, maka pengumpul-
an spora menjadi masalah apabila cuacatidak mendukung.
Analisis untuk menentukan lokasi DNAspesiflk cendawan P. maydis yang dilakukan dengan menggunakan markah RFLPdari CYMMIT (pCLY83) belum berhasil, sehingga ^nalisis PCR tidak dapat dilakukan.
Markah DNA untuk Deteksi Dini PenyakitBulai pada Benih Jagung yang Sakit
Isolasi DNA dari seedling jagung ter-infeksi cendawan P. maydis dengan meng
gunakan metode CYMMIT telah berhasil dilakukan. Gambar III.9 menunjukkan hasil
isolasi DNA seedling jagung yang dilakukandengan menggunakan metode SanghaiMaroof. Metode ini memberikan hasil yang
sangat baik karena DNA yang diperolehcukup banyak dan tidak terdegradasi.Sedangkan isolasi DNA genom dari sporacendawan dengan menggunakan metode
CTAB/NaCl dari IRRI memberikan hasilyang belum memuaskan karena sebagianbesar terdegradasi. Oleh karena itu, perlu
dilakukan isolasi DNA lagi dengan menggunakan metode yang lain. Penelitian untuk mendapatkan metode isolasi DNA darispora cendawan Peronosclerospora
maydis penting dilakukan sebagai langkahawal untuk mendesain markah spesifikcendawan ini guna deteksi benih maupuntanaman.
Hasil Penelitian 2002

45
1 Padi Afrika
22120012080031004
Rentan terhadapStrain IV dan VIII
25012000303
1060000
IV dan VIII
0000000000000000
VIII
32101914000920140
IV
Tahan terhadap strain
79233926383
1911
1144
Aksesi diuji -
AAAACCDDCCDDCCDDCCDDBBCCEECCCCBBAAAAAAAA
Genom
Jumlah
0. sativa (cultivar)0. glaberrima*0. malampuzhaensis0. latifolia0. grandiglumis0. alta0. minuta0. australiensis0. rhizomatis0. officmalis0. punctata0. glumaepatula0. barthii0. rufipogon0. nivara
Spesies
Table 111.30. Jumlah aksesi spesies padi liar yang diuji dan tahan terhadap penyakit hawar daunbakteri
Sebanyak 74 aksesi dari 13 spesies liardan tiga aksesi padi Afrika (O. glaberrima)yang diuji terhadap dua strain HDB menunjukkan 25 aksesi tahan terhadap HDB strainIV dan VIII, 32 aksesi tahan terhadap strainIV, dan 22 aksesi rentan terhadap strain IVdan VIII. Dua padi budi daya yang digunakan sebagai kontrol, yaitu IR64 hanya tahanterhadap strain IV dan BP364B-MR-33-3-PN-
5-1 rentan terhadap strain IV dan VIII(Tabel 111.30).
Dari 15 spesies yang diuji, enam spesies (O. officmalis, O. minuta, O. rhizoma-tis, O. punctala, O. malampuzhaensis, dan
O. glaberrima) merupakan sumber gen ketahanan terh^dap HDB strain IV dan VIII,enam spesies (O. rufipogon, O. barthii, O.alta, O. grandiglumis, O. latlfolia, dan O.malampuzhaensis) merupakan sumberketahanan terhadap HDB strain IV, dan tigaspesies (O. nivara, O. glumaepatula, dan
ter-Evaluasi Ketahanan Spesies Liarhadap Penyakit Hawar Daun Bakteri
ler dapat digunakan sebagai penanda padaseleksi suatu sifat yang disebut markeraided selection (MAS).
Evaluasi dan Karakterisasi Ketahanan
Spesies Padi Liar terhadap CekamanBiotik dan Abiotik dengan Menggunakan
Kultur In Vitro dan Markah Molekuler
Analisis Sidik Jari DNA Spesies Padi Liardengan Menggunakan Markah Mikro-satelit
Uji kualitas DNA hasil isolasi 85 aksesipadi liar diperoleh 96 ekstrak DNA yang cu-kup baik sebagai bahan penelitian molekuler. Empat primer mikrosatelit, yaitu RM197dan RM276 dari kromosom 6, RM201 darikromosom 9, dan RM332 dari kromosom11 telah digunakan dalam analisis sidik jari.Hasil sementara dari keempat primer ter-sebut menunjukkan adanya polimorfismeantara aksesi spesies liar.
Laporan Tahunan Bautbiogen 2002

46
Satu aksesi padi Afrika (O. glaberrima)menunjukkan ketahanan terhadap HDBstrain IV dan VIII, dua aksesi rentan terha
dap HDB. Seperti halnya dengan O. rufipogon dan O. nivara, O. glaberrima merupa
kan sumber ketahanan yang mudah digunakan dalam program perbaikan varietaspadi budi daya karena merupakan keluar-ga dekat dari O. satiua.
Evaluasi Padi Liar terhadap PenyakitBias
Sebanyak 76 aksesi padi liar diuji dirumah kaca terhadap tiga macam ras, yaituras 041 (Pantura Jawa Barat), ras 073 (Ci-kembar-Sukabumi), dan ras 173 (Cikem-bar, Sukabumi). Ketahanan padi liar sangatberbeda terhadap ketiga ras tersebut. Padiliar lebih tahan terhadap ras 041 dibanding-kan dengan ras 073 dan 173. Spesies yangsama dengan aksesi yang berbeda menunjukkan ketahanan yang bervariasi. O.
australiensis menunjukkan reaksi yangberagam antara 14,44-100%, demikian pula19 aksesi O. officinalis bervariasi antara 0-100%. Secara umum O. officinalis lebih tahan dibandingkan dengan padi liar lainnya(Tabel 111.32).
Hasil pengujian terhadap ras 041 diper-oleh delapan aksesi tahan, enam aksesiagak tahan, dan sisanya agak rentan dansangat rentan. Seluruh spesies padi liar
yang diuji tidak tahan terhadap ras 073 dan173.
Evaluasi Ketahanan Spesies Liar terhadap Hama Wereng Coklat
Sebanyak 76 varietas padi liar dari ber-bagai spesies diuji ketahanannya terhadapwereng coklat biotipe 3 (SU). Berdasarkanreaksi terhadap wereng coklat, padi liaryang diskrining dibagi dalam tiga kelompok(Tabel 111.33), yaitu kelompok rentan terdiri
O. australiensis) rentan terhadap HDB. O.officinalis merupakan sumber ketahanan
HDB yang baik, karena dari 19 aksesi, 10 ta-han terhadap strain IV dan VIII, dan sembi-lan tahan terhadap strain IV saja. Demikianjuga O. minuta dan O. rhizomatis tahan
terhadap kedua strain HDB. Dari 11 aksesiO. punctata, enam aksesi tahan terhadap
dua strain dan dua aksesi tahan terhadapstrain IV. Sedangkan dua aksesi 0. malam-
puzhaensis tahan terhadap kedua straindan satu aksesi tahan strain IV saja.
O. minuta merupakan spesies yang
paling tahan terhadap kedua strain HDBIndonesia karena panjang gejala seranganpaling pendek, yaitu 0-0,1 cm untuk strainIV dan 0,1-0,3 cm untuk strain VIII (Tabel111.31). Spesies ini juga tahan terhadap HDBFilipina dan Malaysia, dan sifat ketahanan-nya berhasil diintrogresikan ke padi budidaya.
Empat spesies liar (O. niuara, O.
rufipogon, O. barthii, dan O. glumaepatula)dan padi Afrika (O. glaberrima) merupakankerabat dekat padi budi daya karena mem-punyai genom yang sama (AA) namun ha-nya spesies pertama dan terakhir yangmempunyai gen ketahanan terhadap strainIV dan VIII. O. rufipogon dan barthii hanyamempunyai gen ketahanan terhadap strainIV. O. rufipogon dan O. nivara merupakan
nenek moyang padi budi daya Asia, sedangkan O. barthii nenek moyang padiAfrika. Oleh karena itu, kedua spesies ter-
sebut merupakan sumber ketahanan yanglebih mudah digunakan untuk perbaikanketahanan terhadap HDB padi budi daya.Sifat ketahanan terhadap bias dari O.rufipogon telah berhasil diintrogresikan kedalam padi budi daya demikian juga sifatketahanan terhadap penyakit kerdil rumputdari O. niuara.
Hasil Penelitian 2002
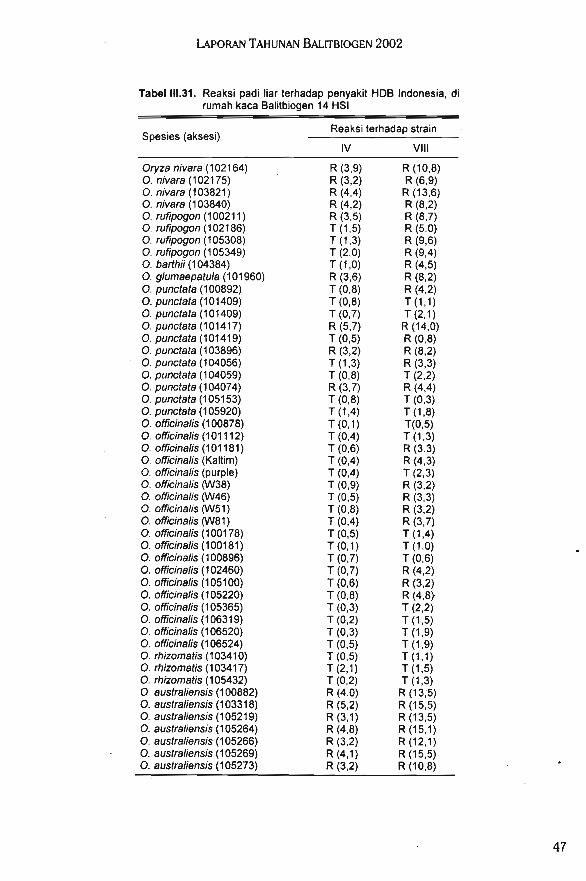
47
R(10,8)R(15,5)R (12,1)R(15,1)R(13,5)R(15,5)R(13,5)TC^3)
T(1,1)T(1,9)T(1,9)
T (2,2)R (4,8>R(3,2)R (4,2)T^^)
R(3,7)R (3,2)R (3,3)R (3,2)T (2,3)R (4,3)R (3.3)T(1,3)T(0,5)
T (0,3)R (4,4)T(2,2)R (3,3)R (8,2)R(0,8)R(14,0)T(2,1)
R (4,2)R (8,2)R (4,5)R (9,4)R (9,6)R (5.0)R (8,7)R (8,2)R(13,6)R (6,9)R(10,8)
VIII
R (3,2)R (4.1)R(3,2)R (4,8)R (3,1)R (5,2)R (4.0)T(0i2)T (0,5)T (0,5)T (0,3)T (0,2)T (0,3)T(0,8)T (0,6)T(0,7)T (0,7)
T(0,5)T (0,4)T (0,8)T (0,5)T (0,9)T (0,4)T (0,4)T (0,6)T (0A)T(0,'i)
T(0,8)R (3,7)T (0,8)T(1,3)R (3,2)T(0,5)R(5,7)T (0,7)T (0,8)T (0,8)R (3,6)T(1,0)T(2,0)T(1,3)T(1,5)R(3,5)R (4,2)R (4,4)R (3,2)R (3,9)
IVOryza nivara (102164)0. nivara (102175)O. nivara (103821)O. nivara (103840)O. rufipogon (100211)O. rufipogon (102186)O. rufipogon (105308)O. rufipogon (105349)O. barthiii (104384)O. glumaepatula (101960)O. punctata (100892)O. punctata (101409)O. punctata (101409)O. punctata (101417)O. punctata (101419)O. punctata (103896)O. punctata (104056)O. punctata {104059)O. punctata {104074)O. punctata (105153)O. punctata (105920)O. officinalis (100878)O. officinalis (101112)O. officinalis (101181)O. officinalis (Kaltim)O. officinalis (purple)O. officinalis (W38)O. officinalis (W46)O. officinalis (W51)O. officinalis (W81)O. officinalis (100178)O. officinalis (100181)O. officinalis (100896)O. officinalis (102460)O. officinalis (105100)O. officinalis (105220)O. officinalis (105365)O. officinalis (106319)O. officinalis (106520)O. officinalis (106524)O. rhizomatis (103410)O. rhizomatis (103417)O. rhizomatis (105432)O. australiensis (100882)O. australiensis (103318)O. australiensis (105219)O. australiensis (105264)O. australiensis (105266)O. australiensis (105269)O. australiensis (105273)
Tabel 111.31. Reaksi padi liar terhadap penyakit HDB Indonesia, dirumah kaca Balitbiogen 14 HSI
Reaksi terhadap strain
Laporan Tahunan Balitbiogen 2002

48
alta (CCDD), dan O. ridleyi (HHJJ). Spesiestersebut merupakan kerabat jauh padi budidaya sehingga untuk mengintrogresikangen ke dalam padi budi daya (AA) tidakmudah. Gen ketahanan terhadap werengcoklat telah berhasil diintrogresikan dari O.officinalis ke dalam padi budi daya denganpenyelamatan embrio muda secara in vitro
dan silang balik dengan tetua rekaren padibudidayanya. Dengan menggunakan mar-
kah molekuler restriction fragment lengthpolymorphism (RFLP) segmen introgresitersebut telah dipetakan, yang terletak pa-da kromosom 11 dan 12. Gen ketahananterhadap wereng coklat dari O. australiensis
dari spesies O. nivara, O. glumaepatula, O.
grandiglumis, O. rufipogon, O. punctata,
dan O. rhizomatis; kelompok gabungan(sebagian aksesi menunjukkan ketahanandan sebagian lagi menunjukkan kerentan-an) terdiri dari O. latifolia, O. australiensis,O. punctata, dan O. malamphuzaensis, dan
kelompok ketiga adalah kelompok yangrelatif menunjukkan reaksi tahan terdiridari O. officinalis, O. alta, dan O. minuta.
Dari 19 aksesi O. officinalis (CC) yangdiuji, 17 aksesi menunjukkan reaksi tahan.Spesies lain yang dapat digunakan sebagaisumber ketahanan adalah O. minuta
(BBCC), O. malampuzhaensis (CCDD), O.
T = tahan (<3,0 cm), R = rentan (>3,0 cm)
R(15,5)R(14,6)
R(3,3)T(1,3)R (4,9)R (3,2)T(1,3)T (0,2)R (3,5)R (3,9)R (3,8)R (8,7)R (4.0)R (4,5)R (3,7)R (3,5)R (3,4)R (4,2)R(3,5)R(4,1)R(3,7)R(3,5)R (3,7)R(10,5)R (3,7)T (0,3)T(0,1)T(0,2)
R(14,7)
VIII
R(12,3)T (2,5)
R (3,5)T (0,9)R (3,8)T (0,4)T(1,2)T (0,6)T (0,7)T (0,3)T (0,6)R (5,6)T (0,8)T(0,9)T (0,6)T (0,8)T (0,8)R(1,8)T(1,5)T(1,4)T(1,7)T(1,0)R (2,2)R(4,3)T(1,7)T(0,1)T (0,0)T (0,0)R (4,0)
IV
Reaksi terhadap strain
- BP364B-MR-33-3-PN-5-1-IR640. sativa:0. glaberrima (101914)0. glaberrima (101297)0. glaberrima (100156)0. malampuzhaensis (105329)0. malampuzhaensis (105223)0. malampuzhaensis (100957)0. latifolia (105141)0. latifolia (102164)0. latifolia (101392)0. latifolia (100966)0. latifolia (100914)0. latifolia (100168)0. latifolia (100885)0. /af/fb//a (100170)0. latifolia (100165)0. grandiglumis (105156)0. grandiglumis (105560)0. a/fa (120952)0. a/fa (105222)0. a/fa (105143)0. a/fa (105138)0. a/fa (100925)O.alta (100888)0. minuta (101386)0. minuta (101141)0. minuta (101089)0. australiensis (105623)
Spesies (aksesi)
Tabel 111.31. Lanjutan
Hasil Penelitian 2002
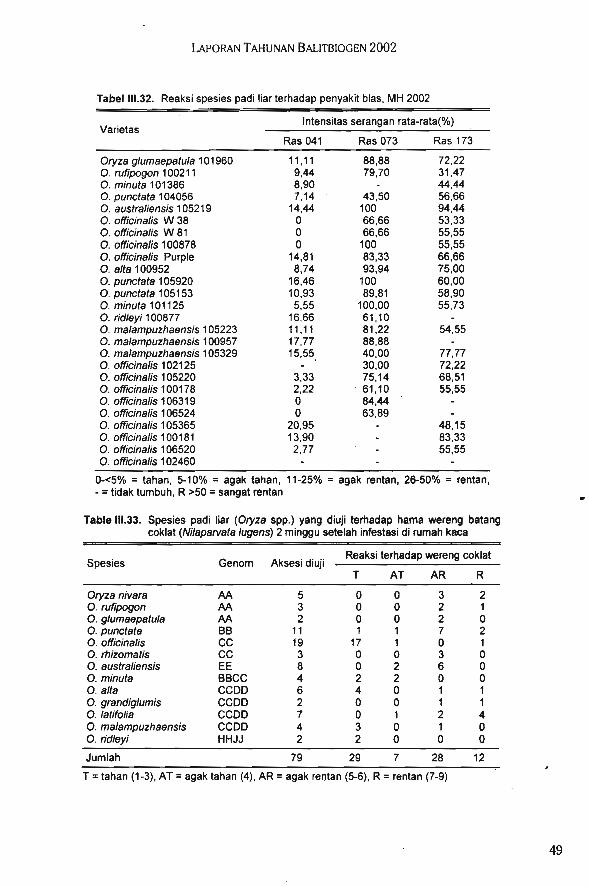
49
T = tahan (1-3), AT = agak tahan (4), AR = agak rentan (5-6), R = rentan (7-9)
120041100012012
R
280121106307223
AR
70010022011000
AT
2923004200
171000
T
Reaksi terhadap wereng coklat
7924726483
1911
235
Aksesi diuii
HHJJCCDDCCDDCCDDCCDDBBCCEECCCCBBAAAAAA
^^V< P\^> 1 1Genom
Jumlah
0. ridleyi0. malampuzhaensis0. latifolia0. grandiglumis0. alta0. minuta0. australiensisO. rhizomatis0. officinalis0. punctata0. glumaepatula0. rufipogonOryza nivara
WwwvlVvSoesies
Table III.33. Spesies padi liar (Oryza spp.) yang diuji terhadap hama wereng batangcoklat (Nilaparvata lugens) 2 minggu setelah infestasi di rumah kaca
agak rentan, 26-50% = rentan,0-<5% = tahan, 5-10% = agak tahan, 11-25%- = tidak tumbuh, R >50 = sangat rentan
-55,5583,3348,15
-
55,5568,5172,2277,77
-
54,55-
55,7358,9060,0075,0066,6655,5555,5553,3394,4456,6644,4431,4772,22
Ras173
-
.
63,8984,4461,1075,1430,0040,0088,8881,2261,10
100,0089,81
10093,9483,33
10066,6666,66
10043,50
-
?9,7088,88
Ras 073
-2,77
13,9020,95
002,223,33
-15,5517,7711,1116,665,55
10,9316,468,74
14,81000
14,447,148,909,44
11,11
Ras 041
Intensitas serangan rata-rata(%)
0. officinalis 1024600. officinalis 1065200. officinalis 1001810. officinalis 1053650. officinalis 1065240. officinalis 1063190. officinalis 1001780. officinalis 1052200. officinalis 1021250. malampuzhaensis 1053290. malampuzhaensis 1009570. malampuzhaansis 1052230. ridfey/1008770. minuta 1011250. punctata 1051530. punctata 1059200. alta 1009520. officinalis Purple0. officinalis 1008780. officinalis W 810. officinalis W 380. australiensis 1052190. punctata 1040560. minuta 1013860. rufipogon 100211Oryza glumaepatula 101960
Vs rist^ s
Tabel 111.32. Reaksi spesies padi liar terhadap penyakit bias, MH 2002
Laporan Tahunan Balitbiogen 2002

50
bandingkan dengan persilangan O. minutadan O. officinalis karena kekerabatan O.
punctata dan O. rufifogon lebih dekatdengan padi budi daya.
Kegiatan silang balik pertama (BC1)sudah dilakukan pada tanaman Fl, yaitupada kombinasi persilangan Way Rarem2/O. punctata 101409, IR64 2/O. punctata101409, IR64 70. punctata 100892, IR64 70.minuta 101141, IR64 70. officinalis 100878,Pelita 1-1 70. punctata 101409, Pelita 1-170. punctata 100892, IR64 70. officinalis100896, dan Pelita 1-1 70. minuta 10114.
Perbaikan Galur Mandul Jantan danGalur Pemulih Kesuburan melalui
Kultur Anter
Persilangan Galur Pelestari denganVarietas Unggul
Persilangan galur pelestari dengan varietas unggul menghasilkan benih, walau-
pun keberhasilannya tidak sama. Dari 16persilangan, enam persilangan menghasilkan benih sedikit (<100 butir) dan sepuluhpersilangan menghasilkan benih antara165-643 butir sedangkan jumlah benih ter-banyak (643 butir) dicapai oleh persilanganIR62829B x Ciherang (Tabel 111.34). Selan-jutnya persilangan IR58025B x Sintanur danIR62829B x Ciherang dipilih untuk pembentukan galur pelestari, serta persilangan
IR53942R x Ciherang dan BR827-35R xSintanur untuk pembentukan galur pemulih kesuburan.
Kultur Anter Fl Hasil Silangan GalurPelestari atau Galur Pemulih Kesuburandengan Varietas Unggul
Pada penelitian ini, kalus yang diper-oleh tidak semuanya menghasilkan tanaman hijau, sebagian kalus menghasilkantanaman yang albino (Tabel III.35). Kalusyang menghasilkan tanaman terbanyak di-
juga telah berhasil diintrogresikan dan di-tandai dengan markah molekuler. Gen ter-
sebut terletak di kromosom 12 dan bertautdengan markah RG457. Beberapa galurgenerasi tiga dari silang balik keempat(F3BC4) antara padi tipe baru IRRI(IR65600-81-5-3-2-1) dengan O. minuta
aksesi 101089 menunjukkan reaksi tahanterhadap wereng coklat.
Evaluasi Ketahanan/Toleransi SpesiesLiar terhadap Kekeringan
Perakaran padi liar yang mampu me-
nembus lapisan lilin bervariasi dari 0-14akar. Padi liar yang menunjukkan dayatembus akar tinggi di antaranya adalah O.glumaepatula 101960, O. glabetrima101297, O. niuara 103860, O. punctata105153, O. alta 105222, dan 0. rufipogon100211.
Dibandingkan dengan kontrol IRAT 112(Gajah Mungkur yang merupakan varietaspadi tahan kekeringan), daya tembus akarpadi liar O. glaberrima hampir sama danjauh lebih tinggi daripada IR64.
Pembentukan Populasi InterspesifikPadi melalui Kultur Embrio secara
In Vitro
Telah dilakukan 24 kombinasi per-silangan interspesifik padi budi daya dengan spesies padi liar. Padi budi daya yangdigunakan adalah Pelita 1-1,1R64, dan WayRarem sedangkan padj liar yang digunakanadalah O. minuta 101141, O. punctata101409, O. punctata 100892, O. australien-sis 105219, O. australiensis 105623, O,officinalis 100896, O. officinalis 100878, danO. rufifogon 102186.
Dari 24 kombinasi persilangan diper-oleh tanaman Fl antara 1-29 tanaman.
Tanaman Fl pada persilangan dengan O.punctata dan O. rufifogon lebih banyak di
Hasil Penelitian 2002
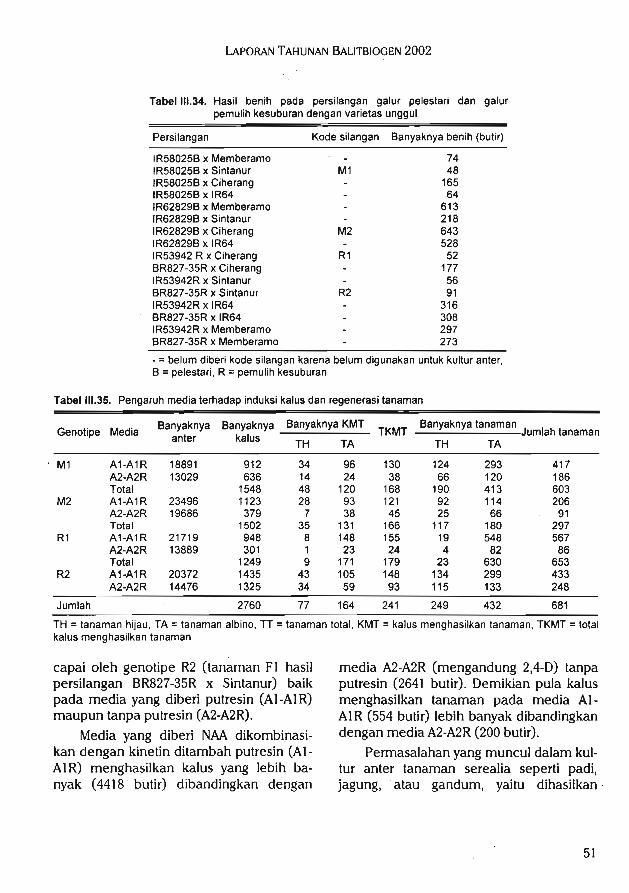
51
media A2-A2R (mengandung 2,4-D) tanpaputresin (2641 butir). Demikian pula kalusmenghasilkan tanaman pada media Al-Al R (554 butir) lebih banyak dibandingkandengan media A2-A2R (200 butir).
Permasalahan yang muncul dalam kultur anter tanaman serealia seperti padi,
jagung, atau gandum, yaitu dihasilkan
capai oleh genotipe R2 (tanaman Fl hasilpersilangan BR827-35R x Sintanur) baikpada media yang diberi putresin (A1-A1R)maupun tanpa putresin (A2-A2R).
Media yang diberi NAA dikombinasi-kan dengan kinetin ditambah putresin (Al-A1R) menghasilkan kalus yang lebih ba-nyak (4418 butir) dibandingkan dengan
TH = tanaman hijau, TA = tanaman albino, TT = tanaman total, KMT = kalus menghasilkan tanaman, TKMT = totalkalus menghasilkan tanaman
681248433653
86567297
91206603186417
•Jumlah tanaman
432133299630
8254818066
114413120293
TA
tanaman
24911513423
419
1172592
19066
124
TH
Banyaknya
24193
14817924
15516645
12116838
130
TKMT
16459
10517123
148131
3893
1202496
TA
773443
918
357
28481434
TH
Banyaknya KMT
2760132514351249
301948
1502379
11231548636912
Banyaknyakalus
1447620372
1388921719
1968623496
1302918891
Banyaknyaanter
A2-A2RA1-A1RTotalA2-A2RA1-A1RTotalA2-A2RA1-A1RTotalA2-A2RA1-A1R
Media
Jumlah
R2
R1
M2
M1
Genotipe
- = belum diberi kode silangan karena belum digunakan untuk kultur anter,B = pelestari, R = pemulih kesuburan
Tabel III.35. Pengaruh media terhadap induksi kalus dan regenerasi tanaman
273297308316
9156
17752
528643218613
641654874
Banyaknya benih (butir)
--
-R2
--
R1-
M2-
-
M1-
Kode silangan
BR827-35R x MemberamoIR53942R x MemberamoBR827-35R x IR64IR53942R x IR64BR827-35R x SintanurIR53942R x SintanurBR827-35R x CiherangIR53942 R x CiherangIR62829B x IR64IR62829B x CiherangIR62829B x SintanurIR62829B x MemberamoIR58025B x IR64IR58025B x CiherangIR58025B x SintanurIR58025B x Memberamo
Persilangan
Tabel 111.34. Hasil benih pada persilangan galur pelestari dan galurpemulih kesuburan dengan varietas unggul
Laporan Tahunan Balitbiogen 2002
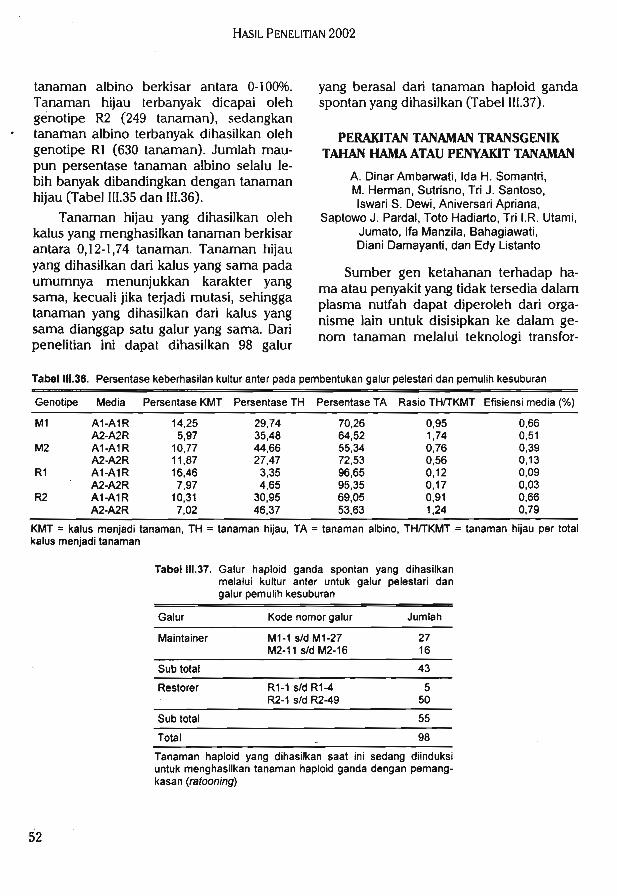
52
Tanaman haploid yang dihasilkan saat ini sedang diinduksiuntuk menghasilkan tanaman haploid ganda dengan pemang-kasan (ratooning)
9855
550
43
2716
Jumlah
R1-1 s/dR1-4R2-1 s/d R2-49
M1-1 s/dM1-27M2-11 s/dM2-16
Kode nomor galur
Total
Sub total
Restorer
Sub total
Maintainer
Galur
tanaman hijau per totalKMT = kalus menjadi tanaman, TH = tanaman hijau, TA = tanaman albino, TH/TKMT =kalus menjadi tanaman
Tabel III.37. Galur haploid ganda spontan yang dihasilkanmelalui kultur anter untuk galur pelestari dangalur pemulih kesuburan
0,790,660,030,090,130,390,510,66
Efisiensi media (%)
1,240,910,170,120,560,761,740,95
Rasio TH/TKMT
53,6369,0595,3596,6572,5355,3464,5270,26
Persentase TA
46,3730,954,653,35
27,4744,6635,4829,74
Persentase TH
7,0210,317,97
16,4611,8710,775,97
14,25
Persentase KMT
A2-A2RA1-A1RA2-A2RA1-A1RA2-A2RA1-A1RA2-A2RA1-A1R
Media
R2
R1
M2
M1
Genotipe
Tabel IH.36. Persentase keberhasilan kultur anter pada pembentukan galur pelestarl dan pemulih kesuburan
yang berasal dari tanaman haploid gandaspontan yang dihasilkan (Tabel 111.37).
PERAKITAN TANAMAN TRANSGENIKTAHAN HAMA ATAU PENYAKIT TANAMAN
A. Dinar Ambarwati, Ida M. Somantri,M. Herman, Sutrisno, Tri J. Santoso,
Iswari S. Dewi, Aniversari Apriana,Saptowo J. Pardal, Toto Hadiarto, Tri I.R. Utami,
Jumato, Ifa Manzila, Bahagiawati,Diani Damayanti, dan Edy Listanto
Sumber gen ketahanan terhadap ha-ma atau penyakit yang tidak tersedia dalamplasma nutfah dapat diperoleh dari orga-nisme lain untuk disisipkan ke dalam ge-nom tanaman melalui teknologi transfor-
tanaman albino berkisar antara 0-100%.
Tanaman hijau terbanyak dicapai olehgenotipe R2 (249 tanaman), sedangkantanaman albino terbanyak dihasilkan olehgenotipe Rl (630 tanaman). Jumlah mau-pun persentase tanaman albino selalu le-
bih banyak dibandingkan dengan tanamanhijau (Tabel 111.35 dan 111.36).
Tanaman hijau yang dihasilkan olehkalus yang menghasilkan tanaman berkisarantara 0,12-1,74 tanaman. Tanaman hijau
yang dihasilkan dari kalus yang sama padaumumnya menunjukkan karakter yangsama, kecuali jika terjadi mutasi, sehinggatanaman yang dihasilkan dari kalus yangsama dianggap satu galur yang sama. Daripenelitian ini dapat dihasilkan 98 galur
Hasil Penelitian 2002

53
analisis molekuler lanjutan gen crylA danbioasai lanjutan tanaman putatif transgenikgenerasi Tl, T2, dan T3; (2) transformasikedelai dengan gen crylA melalui penem-bakan partikel, analisis molekuler lanjutan,dan bioasai lanjutan tanaman kedelaigenerasi R3 dan R4; (3) analisis molekulerdan bioasai tanaman kacang tanah PStV;(4) melakukan transformasi ubi jalar dengan gen pinll atau gen CP-SPFMV melaluiA. tumefaciens, analisis molekuler, dan bio
asai tanaman ubi jalar pinll; dan (5) melakukan rekonstruksi gen a-amilase inhibitorpada plasmid biner dalam Agrobacterium.
Transformasi Tanaman Padi untukKetahanan terhadap Hama
Penggerek Batang
Hasil transformasi pada Taipei-309(Japonica) dan Cisadane (Indica) disajikanpadaTabelIII.38.
Dari 200 kalus embriogenik padiTaipei-309 yang ditembak, hanya 26 (13%)yang mampu beregenerasi pada mediaNK4N2 dan menghasilkan 280 tanaman hi-jau (putatif transgenik) sedangkan dari 100kalus embriogenik padi Cisadane baru diperoleh satu tanaman hijau (putatif transgenik) menggunakan media MSP. Hal inimenunjukkan bahwa sumber eksplan(genotip) dan tingkat embriogenik kalussangat berpengaruh dalam proses regene-
rasi tanaman hijau. Sampai saat ini, kalusyang belum dapat beregenerasi masih te-rus berproliferasi dan sebagian kalus ber-
ada dalam tahap green spots pada mediaNK4N2 maupun media MSP untuk melan-jutkan pertumbuhannya.
Tanaman putatif transgenik (TO, Tl, T2,dan T3) diteliti lebih lanjut melalui analisismolekuler (PCR) untuk mengetahui inte-grasi gen crylA(b) dalam genom tanaman.
masi. Teknologi transformasi melalui pe-
nembakan partikel digunakan di Balitbiogen, Bogor dalam perakitan tanaman paditransgenik tahan terhadap hama pengge-rek batang {Scirpophaga sp. Wlk) dan tanaman kedelai tahan terhadap hama peng-gerek polong {Etiella zinckenella), sedang-kan transformasi melalui Agrobacteriumtumefaciens digunakan untuk perakitan tanaman ubi jalar tahan terhadap hama bo-leng {Cylas formicarius) dan penyakit vimsSPFMV {Sweet Potato Feathery MottleVirus). Transformasi tanaman kacang ta-
nah dengan gen CP-PStV {Coat Protein-Peanut Stripe Virus) telah dilakukan diUniversitas Queensland, Australia, yang se-
lanjutnya dilakukan pengujian molekulerdan bioasai di Balitbiogen.
Sumber gen ketahanan yang digunakan adalah gen crylA yang diisolasi dariBacillus thuringiensis untuk tanaman padidan kedelai, gen proteinase inhibitor (pin)
II dari tanaman kentang, dan gen CP-SPFMV {Coat Protein-SPPMV) untuk tanaman ubi jalar.
Penelitian transformasi dilakukan untuk penelitian konstruksi gen. Gen a-ami-
lase inhibitor (pTA3) yang diperoleh dariDr. T.J. Higgins, telah dimanfaatkan untuk
penelitian awal konstruksi gen ke dalamplasmid biner, untuk selanjutnya dapat digunakan dalam studi transformasi.
Perakitan tanaman padi transgenikpada tahun 2001 menghasilkan tanamanTaipei-309 (T-309) generasi To, Tl, maupunT2 telah positif mengandung gen crylA(b)dan beberapa tanaman sangat tahan dan
tahan pada pengujian bioasai. Transformasi tanaman kedelai menghasilkan tanamanWilis dan Tidar generasi Rl atau R2 yangpositif mengandung gen pinll.
Kegiatan penelitian tahun 2002 adalah(1) transformasi padi dengan gen crylA,
Laporan Tahunan Balitbiogen 2002

54
M = 1 Kb ladder, + = kontrol positif, - = kontrol negatif, A = air
Gambar 111.10. Hasil analisis PCR VII pada tanaman putatif transgenik padi Taipei-309
pUBB = crylA(b); pRQ6 = gus, hpt; NH40 = NB + 35 atau 40 mg/l higromisin; PRNH40 = NB + 2 mg/l BAP + 1 mg/lNAA + 5 mg/l ABA + 35 atau 40 mg/l higromisin; NK4N2 = NB + 4 mg/l kinetin + 2 mg/l NAA; MSP = MS + 2 mg/lkinetin + 0,5 mg/l NAA + 10~3 M spermidin; * = penelitian masih berlangsung
M 2 5 6 11 13 15 17 23 28 49 51 53 54 58 59 60 + M 68 69 78 81 84 - A +
1
1
MSP*
pada media
280
280
NK4N2
Jumlah tanaman hijauregenerasi
27 (9%)
26(13%)1 (1%)
Kalus
85 (28,3)
71 (35,5)14(14)
PRNH40
media seleksi
96 (32)
71 (35,5)25 (25)
NH40
Kalus hidup pada
300
200100
Jumlaheksplan
Total
Taipei-309Cisadane
Varietas
Tabel 111.38. Transformasi tanaman padi dengan plasmid pUBB dan pRQ6 pada molaritas 4 : 1 menggunakanmikroprojektil
jukkan kategori agak tahan sampai sangattahan pada pengujian bioasai (sundep danbeluk). Dari 90 tanaman yang sudah diujibioasai pada tahun 2002 diperoleh 21 tanaman agak tahan, tahan, dan sangat ta
han, yang kemudian dianalisis denganPCR. Hasil pengujian PCR ditampilkanpadaGambar III. 10.
Dari Gambar III. 10 terlihat bahwa 10sampel tanaman yang terdiri dari duatanaman Tl, tujuh tanaman T2, dan satu
tanaman T3 positif mengandung gencrylA(b). Apabila dikombinasikan dengandata hasil bioasai, kesepuluh tanaman ter-
sebut menunjukkan kategori agak tahan,tahan, atau sangat tahan pada pengujiansundep maupun beluk (Tabel 111.39).Dengan demikian, sifat ketahanan tersebutmungkin berasal dari ekspresi gen crylA(b)yang telah terintegrasi ke dalam genomtanaman padi.
Analisis PCR dilakukan menggunakandua primer nukleotida spesifik untukmengamplifikasi daerah spesifik genoylA(b) dan fragmen DNA yang dihasilkanmempunyai ukuran 1000 pasangan basa.
Pada tahun 2002 telah dilakukan PCR VIpada 12 sampel tanaman padi Taipei-309generasi TO, tetapi hasil analisis menunjuk-
kan tidak ada sampel yang positif mengan-dung gen crylA(b). Sedangkan pada PCRVIII telah diuji 32 galur tanaman putatiftransgenik Taipei-309 generasi TO danmenghasilkan empat galur (19 tanaman)yang positif mengandung gen crylA(b).Tanaman positif dipelihara untuk penguji-an kestabilan integrasi gen pada generasiselanjutnya maupun untuk pengujian bio-asai. Untuk mengkonfirmasi data bioasaidengan data molekuler, dilakukan analisisPCR VII untuk tanaman putatif transgenikgenerasi Tl, T2, dan T3 yang sudah menun-
Hasil Penelitian 2002

55
ekspresikan protein crylA(b) dalam jumlahcukup sehingga bersifat insektisidal terhadap hama penggerek yang menyerang.
Untuk verifikasi lebih lanjut perlu dilakukananalisis molekuler pada beberapa tanaman
terpilih dengan tingkat ketahanan agaktahan sampai sangat tahan. Hasil analisismolekuler disajikan pada Tabel III.39.
Transformasi Tanaman Kedelai untukKetahanan terhadap Hama Penggerek
Polong (Etiella zinckenelld)
Dari penelitian tahun 2001 telah diper-oleh tanaman kedelai yang positif me-
ngandung genp/nll, yaitu delapan tanamanTidar generasi kedua hasil transformasimelalui A. tumefaciens (8 ATI R2) danempat tanaman Wilis generasi pertama hasil transformasi melalui penembakan parti-
kel (4 WP2 Rl). Penelitian dilanjutkan pada tahun 2002 untuk melihat kestabilan
Bioasai dilakukan untuk mengetahuitingkat ketahanan tanaman putatif transgenik terhadap penggerek batang kuning.Hasil bioasai untuk tingkat ketahanantanaman disajikan pada Tabel 111.40.
Dari 46 tanaman yang diuji menunjuk-kan tingkat ketahanan yang bervariasi darisangat tahan sampai sangat rentan. Peng-
amatan gejala sundep menghasilkan 13 tanaman dengan kategori sangat tahan, limatanaman tahan, dan sembilan tanaman
agak tahan. Sedangkan pengamatan gejalabeluk menghasilkan 19 tanaman dengankategori sangat tahan, dua tanaman tahan,lima tanaman agak tahan, dan nomor-
nomor tanaman lain termasuk dalam kategori agak rentan sampai rentan. Sementaraitu, bioasai pada lima tanaman kontrol
(Taipei-309 non transgenik) semuanya me-nunjukkan kategori sangat rentan. Didugatanaman yang sangat tahan dapat meng-
ST = sangat tahan, T = tahan, AT = agak tahan, AP = agak peka
81 +54176
84696860 +59 +534923 +1558 +51 +13 +5 +
7828 +11 +
2
1 ^ \^l 1 1 \^ I ^^ 1 ^/ \ 1 lbl^^l 1 1 ^^l Nomor DNA Hasil PCR
STSTT
APSTSTSTATSTSTSTSTSTSTATT
ATSTSTSTST
Beluk
ATT
ATATSTSTSTSTSTSTAPSTSTSTSTT
STTT
ATT
Sundep
Tingkat ketahanan
A2-1BA1-3BA3-7A2-5B24-4BB24-4B24-5B24-6B24-2B24-3B24-7B24-3BB24-4B5-5B5-2
B5-4B5-2B5H-5-15H-5-1B5H-75B-2
i ^^^i i i^^i ^^i mi i i^^iiNomor tanaman
T3
T2
T1
^Jwl Iwl Cl^7lGenera^
Tabel 111.39. Konfirmasi hasil bioasai dengan hasi! analisis PCR pada tanamanputatif transgenik padi Taipei-309 generasi T1, T2, dan T3
Laporan Tahunan Balitbiogen 2002

56
sampel DNA kedelai Wiiis ditampilkanpadaGambarIII.il.
Biji dari tanaman yang positif mengandung gen pinll ditanam kembali untukmendapatkan generasi selanjutnya (R4 untuk Tidar dan R3 untuk Wilis) dan dilihatkestabilan keberadaan gen p/nllnya. Dari36 tanaman yang diuji (15 tanaman dariTidar dan 21 tanaman dari Wilis), tidak adatanaman yang terdeteksi mengandung genpinll pada generasi R4 atau R3. Hasil initidak sesuai dengan yang diharapkan kare-na secara teori, jumlah tanaman yang me
ngandung gen pinll pada generasi selanjutnya memiliki perbandingan 3:1. Ketidakkonsistenan gen pinll ini menunjukkanbahwa gen tersebut tidak terintegrasi dengan stabil pada genom tanaman tersebut.
Kemungkinan lain adalah terjadi error rateyang mengakibatkan false-positif, di manatanaman yang sebenamya negatif tetapi
integrasi gen pinll pada generasi selanjut-nya, yaitu R2, R3, atau R4.
Dari 208 biji yang disemai, tumbuh 202tanaman yang kemudian diambil daunnyauntuk diekstraksi DNAnya. Dari dua teknikekstraksi DNA yang digunakan, yaitu modi-fikasi metode Murray dan Thompson danminiprep CTAB, ternyata metode minipreplebih efektif. Tingkat kemurnian DNA darihasil pengukuran dengan alat spektrofoto-meter berkisar 1-2,78. Untuk mendeteksikeberadaan gen pinll, digunakan primerspesiflk yang digunakan akan mengamplifi-kasi 600 bp fragmen gen pinll. Dari 167sampel DNA yang diuji menunjukkan duasampel positif mengandung gen pinll, yaitutanaman Wilis generasi kedua (R2) hasiltransformasi melalui penembakan partikel(WP2-38-13) dan Tidar generasi ketiga (R3)hasil transformasi melalui A. tumefaciens(ATI-11.8.15). Hasil PCR pada beberapa
ST = sangat tahan, T = tahan, AT = agak tahan, AP = agak peka, P = peka, SP = sangat peka,S = sundep, B = beluk
52211-1-----
---11
SP
552--41---
11---1
P
1012
124221--
113-32
AP
kategori ketahanan
59-222-112.
1-11•
1
AT
251----1----11.1-2
T
1913
121-23652-224-11
ST
Jumlah tanaman dengan
BSBSBSBSBSB(ftBSBSBS
pada gejalaPengamatan
46
7
8
6
7
2
6
5
5
yang diujiJumlah tanaman
Total
T3 (A3)
T3 (A2)
T3(A1)
T2 (B-24)
T2(B-6)
T2 (B-5)
T1 (T-5H)
T1 (T-5B)
9 ^WI ^^4I l^ll 1 IV • 1Asal tanaman
Tabel 111.40. Tingkat ketahanan tanaman putatif transgenik padi Taipei-309 generasi T1, T2, T3 terhadappenggerek batang padi kuning fase vegetatif (gejala sundep) dan generatif (gejala beluk)
Hasil Penelitian 2002

57
terdeteksi negatif atau tidak mengandunggenp/7?II.
Di samping menggunakan gen p/nll,transformasi kedelai juga dilakukan dengan gen crylA yang efektif untuk ketahanan terhadap hama golongan Lepidopteraterutama penggerek, sehingga diharapkanakan dihasilkan tanaman kedelai yangmempunyai ketahanan terhadap penggerek polong. Transformasi dilakukan padakotiledon muda varietas Tidar dan Wilisdengan metode penembakan (Sanford eta/.) yang dimodifikasi oleh (Sawant et al.)menggunakan plasmid pSBB (oyLA(b))yang dikotransformasikan dengan pRQ6{gus, hpt) dalam molaritas 4:1. Hasil transformasi disajikan pada Tabel 111.42.
Banyaknya eksplan yang bisa berta-han/tumbuh pada media seleksi sangatmenurun dibandingkan dengan banyaknyaeksplan awal yang ditembak. Setelah satuhari penembakan, banyak eksplan yang
terdeteksi positif pada generasi sebelum-
nya.Pada tahun 2002 dilakukan bioasai
untuk 10 tanaman Wilis generasi R3, yaituempat tanaman dari event penembakan
pertama (WP1) dan enam tanaman darievent penembakan kedua (WP2). Kriteriaketahanan tanaman dikelompokkan berda-sarkan persentase serangan polong, yaitu
tahan (0-10), agak tahan (11-30), agakrentan (31-50), rentan (51-70), dan sangatpeka (71-100). Persentase serangan teren-dah pada polong 51,8% dan biji 44,3% di-jumpai pada tanaman dengan kode WP1-1-1, sedangkan pada tanaman kontrol,
polong 95,4% dan biji 96,7%. Dari 10 tanaman yang diuji menunjukkan tingkat ketahanan rentan sampai sangat rentan dengan
rata-rata persentase polong terserang
81,3% dan persentase biji terserang 79,7(Tabel 111.41). Apabila dikonfirmasi denganhasil analisis molekuler, tanaman tersebut
Primer spesifik memperbanyak fragmen sepanjang 600 bp dari genp/nll. Lane M = X Hind\\\, lane 1 = kontrol negatif (tanpa templateDNA), lane 2 = kontrol positif (pTWa), lane 3 sampai 11 = WP1-9,WP1-9, WP1-11, WP1-19, WP2-46, WP2-13, WP2-19, WP2-28,dan WP2-38.13 secara berurutan. Ukuran fragmen pada lane 1sama dengan fragmen hasil amplifikasi kontrol positif (600 bp) yangdiduga sebagai bagian dari gen p/nll
Gambar 111.11. Hasil PCR DNA kedelal varietas Wilis (WP2) dengan menggunakan primer spesifik untuk gen p/nll
M1 23456789 10 11
Laporan Tahunan Balitbiogen 2002

58
lam genom tanaman. Isolasi DNA dari dauntanaman putatif transgenik dan nontrans-genik dilakukan dengan prosedur Scott danBendich dan modifikasi metode CTAB dariTanaka dan Nakatani.
Dari 18 sampel DNA yang diuji, rata-rata menunjukkan kemurnian DNA yangsudah sesuai digunakan untuk analisis PCR(1,8-2,0). Untuk tahap optimasi, analisisPCR baru dilakukan pada lima nomor tanaman kacang tanah generasi Rl. HasilPCR menunjukkan bahwa tiga nomor tanaman, yaitu RII.Ddl.2, RII.Ddl.4, dan Cp4
Dd2 positif mengandung gen CP-PStV dengan munculnya pita DNA yang berukuran300 bp(Gambar 111.12).
Pada pengujian biasanya digunakankriteria tahan (gejala tidak muncul, intensi-tas serangan 0%), agak tahan (gejala muncul pada hari ke-12 sampai 13 setelah ino-kulasi, intensitas serangan 33,3%), dan tidak tahan (gejala muncul pada hari ke-3
berubah warna menjadi putih dan tidakdapat melanjutkan pertumbuhan. VarietasWilis lebih responsif dalam menghasilkaneksplan tahan (10%) maupun jumlah em-brio somatik (124) daripada varietas Tidar,tetapi belum bisa beregenerasi. Dari 35embrio somatik varietas Tidar dihasilkandua planlet yang tumbuh normal, sedang-kan pada embrio somatik lain dijumpaibentuk terompet atau berdaun satu. Sebe-
lum diaklimatisasi, planlet masih perlupendewasaan untuk pertumbuhan daundan akar yang lebih banyak.
Transformasi Tanaman Kacang Tanahuntuk Ketahanan terhadap PStV
Pada penelitian tahun 2001 telah diaklimatisasi beberapa tanaman putatiftransgenik kacang tanah dan dilakukanperbanyakan benih. Benih tersebut dita-nam kembali dan digunakan sebagai mated dalam pengujian molekuler (PCR) untuk mengetahui integrasi gen CP-PStV da
pSBB = crylA(b), pRQ6 = gus, hpt, seleksi = higromisin 30 mg/l
2
Banyaknyaplanlet
35124
Banyaknyaembrio somatik
melalui penembakan partikel
Sangat rentan
Sangat rentan
96,7
79,7
55(4,15%)34(10,0%)
i Eksplan tahan padamedia seleksi
1324340
Banyaknyaeksplan
Transformasi kedelai dengan gen c/ylA(b)
95,4
81,3
TidarWilis
Varietas
Tabel 111.42.
Kontrol
Rata-rataRentanSangat rentanSangat rentanSangat rentanSangat rentanSangat rentanSangat rentanSangat rentanSangat rentanRentan
59,396,193,169,696,277,7
100,086,973,544,3
66,498,891,270,694,976,9
100,081,680,351,8
WP2-46-12WP2-46-11WP2-46-4WP2-46-3WP2-46-2WP2-46-1WP1-1-4WP1-1-3WP1-1-2WP1-1-1
Biji terserang (%) Kriteria ketahananPolong terserang (%)Kode tanaman
Tabel 111.41. Bioasai tanaman kedelai Wilis generasi R3 terhadap penggerek polong E. zinckenella
Hasil Penelitian 2002

59
Tidak tahanTidak tahanTidak tahanTidak tahanTidak tahanAgak tahanAgak tahanAgak tahanAgak tahanAgak tahanTahanTahanTahanTahanTahanTahanTahanTahan
Klasifikasi
66,766,766,766,766,733,333,333,333,333,3
00000000
Persentase tanaman terserang
33356
1313131212----
-
_
Waktu gejala muncul
KontrolEel 3Eel 1R2Dd2(1)R2Dd1.2R2Dd1.1R2 Dd 2 (3)R2 Dd 2 (4)Eel 5Eel 6Cp4 Dd 2.3Cp4 Dd2.2Eel 2R2 Dd2 (5)R2 Dd2 (2)R2 Dd1 (4)R2 Dd1 (3)R2Dd1 (1)
No. tanaman uji
M = marker, 1 = DNA No. 5, 2 = DNA No. 7, 3 = DNA No. 12, 4 = DNA No. 17, 5 = DNANo. 19 (No. 2 PStV sudah di PCR), 6 = DNA No. 20 PCP I, 7 = DNA No. 21 (No. 1 PStVsudah di PCR, 8 = DNA No. 18 kontrol dan 9 = DNA No. 10 ditambah pUBC wereng
Gambar 111.12. Hasil amplifikasi PCR tanaman putatif transgenik kacang tanahmenggunakan primer spesifik gen CP-PStV
Tabel III.43. Tingkat ketahanan tanaman putatif transgenik kacang tanah terhadap PStV
•4-490 pb
M1 23456789
Transformasi Tanaman Ubi Jalar untuk
Ketahanan terhadap Hama Boleng(Cylas formicarius) dan Penyakit Virus
SPFMVTransformasi tanaman ubi jalar hanya
dilakukan melalui Agrobacterium tumefa-ciens karena berdasarkan hasil penelitiansebelumnya, teknik ini lebih efektif di-
sampai 6 setelah inokulasi, intensitas se-rangan 66,7%). Dari 18 tanaman yang diujidiperoleh delapan tanaman tahan, lima tanaman agak tahan, dan lima tanaman tidaktahan (Tabel 111.43). Di samping tingkat ke-tahanan, dilakukan pula pengamatan mor-fologi tanaman, seperti tinggi batang, ba-
nyaknya polong, bobot polong, dan ba-nyaknya polong bernas.
Laporan Tahunan Balitbiogen 2002

60
Tiga tanaman (0,72%) berhasil diperoleh dari 414 eksplan petiol Jewel menggunakan media Rl maupun R2, sedangkanpada Bis 182-81 masih belum bisa beregenerasi.
Transformasi dengan gen pinll hanyadilakukan pada varietas Jewel menggunakan eksplan daun dan petiol. Setelahditransformasi, eksplan diseleksi dan di-regenerasikan secara organogenesis maupun embriogenesis. Hasil transformasi ubi
jalar dengan gen pinll melalui jalur regenerasi secara organogenesis disajikan pada
Tabel 111.45. Dengan media Rl dan R2 diperoleh 14 tunas (25%) dari 56 eksplan petiol dan tujuh tunas (26,9%) dari 26 eksplandaun menggunakan media Rl. Penelitian
tahun 2000 menggunakan media Rl hanyamenghasilkan 2,4% kalus daun yang bisaberegenerasi. Jalur regenerasi lain, yaitu
embriogenesis belum dapat menghasilkantunas, baik dengan perlakuan karborundum maupun tanpa karborundum. Untuk
eksplan petiol perlakuan pelukaan dengankarborundum dapat meningkatkan banyaknya eksplan terseleksi (84,4%), sedangkanpada eksplan daun perlakuan tanpa karborundum ternyata lebih efektif (72,4%).
Lima tanaman hasil transformasi de
ngan gen CP-SPFMV dan 21 tanaman hasiltransformasi dengan gen pinll telah diakli-matisasi dalam pot/ember dan sebagian di-tanam di tanah guludan dalam rumah kacaFasilitas Uji Terbatas untuk pengujian ana-lisis molekuler dan bioasai.
Analisis molekuler PCR dilakukanuntuk mengkonfirmasi integrasi gen pinllmaupun gen CP-SPFMV dalam genomtanaman putatif transgenik ubi jalar (21 tanaman dengan gen pinll dan lima tanamandengan gen CP-SPFMV hasil transformasiTA 2002 dan lima tanaman hasil transformasi dengan gen pinll serta empat tanam-
bandingkan melalui penembakan partikel.Transformasi menggunakan AgrobacteriumLBA4404 yang berisi pGApin {pinll, npt\\)untuk ketahanan terhadap hama danpMON10574 (CP-SPFMV, gus, nptll) untukketahanan terhadap virus. Sebagai jaringantarget untuk ditransformasi digunakan eks-
plan daun/petiol varietas Jewel dan Bis182-81. Beberapa perlakuan yang dicobaadalah konsentrasi asetosiringon (0, 100,200 \M), lama inokulasi eksplan dalam sus-pensi bakteri (30 dan 60 menit), jalur rege-nerasi (organogenesis, embriogenesis),
dan pelukaan (karborundum, tanpa karbo-rundum). Uji ekspresi gen gus secara histo-
kimia dilakukan satu bulan setelah trans-fprmasi dan menunjukkan bahwa penam-
bahan asetosiringon 100 fiM dan lama inokulasi 60 menit dapat meningkatkan efi-siensi transformasi (banyaknya eksplan
' positif gus dan banyaknya spot biru).Pada tranformasi dengan gen CP-
SPFMV, jalur regenerasi yang digunakanadalah secara organogenesis dengan
media Rl (MS + 0,2 mg/1 kinetin) di manaregenerasi dimulai dengan terbentuknyaakar dan tunas yang muncul dari bagiantengah kalus. Pada perlakuan konsentrasi
asetosiringon 100 nM maupun lama inokulasi 60 menit, dari total 1440 eksplan petiolBis 182-81 yang ditransformasi dan direge-nerasikan dengan seleksi kanamisin 50mg/1 menghasilkan 44 (3%) eksplan ber-akar tetapi belum dapat beregenerasimembentuk tunas (tanaman). Sedangkanpada Jewel diperoleh dua tanaman dari 80(5,5%) eksplan berakar. Untuk meningkatkan tanaman yang diperoleh, di sampingmedia regenerasi Rl (MS + 0,2 mg/1 kinetin) dicoba media regenerasi lain, yaitu R2(MS + 0,2 mg/1 2ip). Hasil transformasimenggunakan media regenerasi Rl dan R2disajikan pada Tabel III.44.
Hasil Penelitian 2002
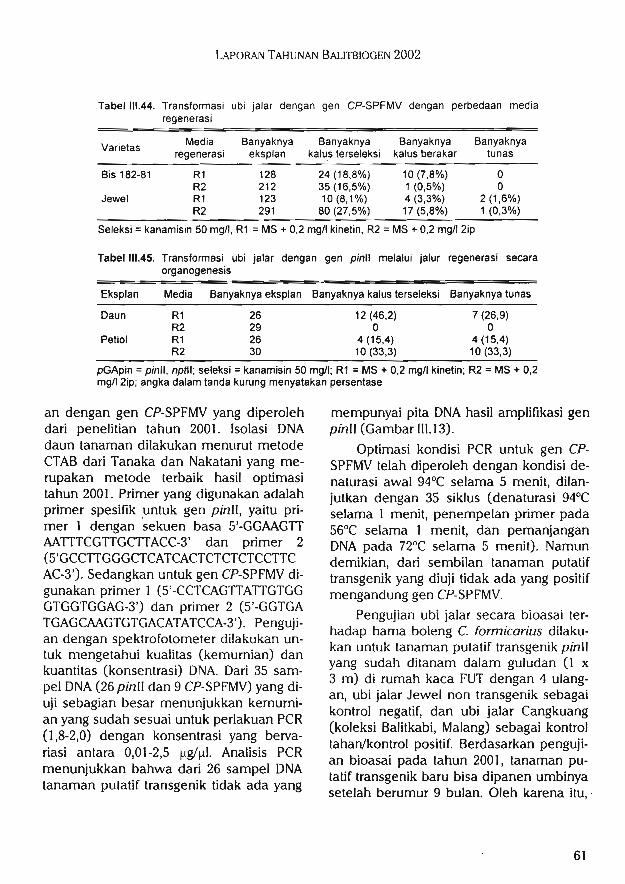
61
mempunyai pita DNA hasil amplifikasi genp/nll (Gambar 111.13).
Optimasi kondisi PCR untuk gen CP-SPFMV telah diperoleh dengan kondisi de-naturasi awal 94C selama 5 menit, dilan-
jutkan dengan 35 siklus (denaturasi 94Cselama 1 menit, penempelan primer pada
56C selama 1 menit, dan pemanjanganDNA pada 72C selama 5 menit). Namundemikian, dari sembilan tanaman putatif
transgenik yang diuji tidak ada yang positifmengandung gen CP-SPFMV.
Pengujian ubi jalar secara bioasai ter-hadap hama boleng C. formicarius dilaku
kan untuk tanaman putatif transgenik pinttyang sudah ditanam dalam guludan (1 x3 m) di rumah kaca FUT dengan 4 ulang-an, ubi jalar Jewel non transgenik sebagai
kontrol negatif, dan ubi jalar Cangkuang(koleksi Balitkabi, Malang) sebagai kontroltahan/kontrol positif. Berdasarkan pengujian bioasai pada tahun 2001, tanaman putatif transgenik baru bisa dipanen umbinyasetelah berumur 9 bulan. Oleh karena itu, •
an dengan gen CP-SPFMV yang diperolehdari penelitian tahun 2001. Isolasi DNAdaun tanaman dilakukan menurut metode
CTAB dari Tanaka dan Nakatani yang me-rupakan metode terbaik hasil optimasitahun 2001. Primer yang digunakan adalahprimer spesifik untuk gen pin\\, yaitu primer 1 dengan sekuen basa 5'-GGAAGTT
AATTTCGTTGCTTACC-3' dan primer 2(5'GCCTTGGGCTCATCACTCTCTCTCCTTCAC-3'). Sedangkan untuk gen CP-SPFMV digunakan primer 1 (5'-CCTCAGTTATTGTGGGTGGTGGAG-3') dan primer 2 (5'-GGTGATGAGCAAGTGTGACATATCCA-3'). Penguji-an dengan spektrofotometer dilakukan untuk mengetahui kualitas (kemurnian) dankuantitas (konsentrasi) DNA. Dari 35 sam-
pel DNA (26 pintt dan 9 CP-SPFMV) yang di-uji sebagian besar menunjukkan kemurnian yang sudah sesuai untuk perlakuan PCR(1,8-2,0) dengan konsentrasi yang berva-
riasi antara 0,01-2,5 ^g/^l. Analisis PCRmenunjukkan bahwa dari 26 sampel DNAtanaman putatif transgenik tidak ada yang
pGApin = pintt, npfll; seleksi = kanamisin 50 mg/l; R1 = MS + 0,2 mg/l kinetin; R2 = MS + 0,2mg/l 2ip; angka dalam tanda kurung menyatakan persentase
7 (26,9)0
4(15,4)10(33,3)
12(46,2)0
4(15,4)10 (33,3)
26292630R2
R1R2R1
Petiol
Daun
Seleksi = kanamisin 50 mg/l, R1 = MS + 0,2 mg/l kinetin, R2 = MS + 0,2 mg/l 2ip
Tabel III.45. Transformasi ubi jalar dengan gen p/'nll melalui jalur regenerasi secara
organogenesis
Eksplan Media Banyaknya eksplan Banyaknya kalus terseleksi Banyaknya tunas
00
2 (1,6%)1 (0,3%)
10(7,8%)1 (0,5%)4 (3,3%)17(5,8%)
24(18,8%)35(16,5%)
10(8,1%)80 (27,5%)
128212123291
R1R2R1R2
Jewel
Bis 182-81
Banyaknya Banyaknya Banyaknyakalus terseleksi kalus berakar tunas
Banyaknyaeksplan
MediaregenerasiVarietas
Tabel 111.44. Transformasi ubi jalar dengan gen CP-SPFMV dengan perbedaan media
regenerasi
Laporan Tahunan Balitbiogen 2002

62
aai). Dari uji restriksi dengan enzim BamHI dan Xbal diperoleh dua arah (kanan dankiri). Arah ke kanan (pCambia-aail darikoloni 43) menunjukkan dua fragmen berukuran 13,485 kb dan 3,219 kb denganenzim BomHI, sedangkan dengan enzim
Xbal diperoleh fragmen berukuran 15,421kb dan 1,303 kb. Arah ke kiri (pCambia-aai2 dari koloni 58) menunjukkan dua fragmen berukuran 15,026 kb dan 1,698 kbdengan enzim BamHI, sedangkan denganenzim Xbal diperoleh fragmen berukuran13,082 kb dan 3,642 kb. Hal ini menunjukkan bahwa plasmid pCambia-aail danpCambia-aai2 mempunyai pola restriksi se-
perti yang diharapkan. Kedua plasmid tersebut selanjutnya ditransformasikan ke dalam Agrobacterium tumefaciens LBA4404dan menunjukkan bahwa beberapa kolonidapat tumbuh pada media LB dengankanamisin dan rifampisin. A. tumefaciens
yang sudah mengandung pCambial 301 dengan gen a-ai selanjutnya dapat digunakanuntuk transformasi gen pada tanaman.
bioasai dilakukan secara in planta, yaitudengan menginfestasikan tiga pasanghama boleng per tanaman secara langsungpada tanaman putatif transgenik yang di-tanam dalam guludan maupun dalamember.
Rekonstruksi Gen a-amilase Inhibitor
pada Plasmid Biner
Penelitian untuk mendapatkan kon-struksi gen a-amilase inhibitor (aai) padaplasmid biner pCambia!301 (gus, npt\\,hpt), meliputi beberapa tahap kegiatan,yaitu isolasi plasmid pCambia 1301 danpTA3 (gen aai) serta uji pola restriksinya,ligasi dan transformasi ke dalam Eschericiacoli DH5a.
Dari hasil proses ligasi yang sudahditransformasi ke dalam E. coli diperoleh182 koloni yang tumbuh pada media YEP +kanamisin dan diharapkan koloni tersebutsudah mengandung pCambial301-aai.Pengujian awal untuk membuktikan insersifragmen gen aai pada pCambial301 dilakukan dengan memotong DNA plasmidmenggunakan enzim Hindlll. Dari 60sampel DNA plasmid, diperoleh dua koloni(nomer 43 dan 58) yang menunjukkanfragmen berukuran 11,837 kb (fragmenpCambial301) dan 4,887 kb (fragmen gen
M = DNA X H/ncflll, 4-35 = tanaman ubi jalar putatif transgenik, - = ubi jaiar non transforman, A = ddH2O, + = kontrolpositif (600 bp)
Gam bar 111.13. Hasil amplifikasi PCR tanaman putatif transgenik ubi jalar menggunakan primer spesifik gen pin\\
' i
4 5 6 7 11 12 16 17 18 19 20 21 22 23 24 25 26 +M 27 28 29 30 3132 33 34 35 - A + + + M
Hasil Penelitian 2002

63
Penyisipan Gen Gus pada TanamanBawang Merah dan Tomat
Bawang Merah
Hasil uji gus pada bawang merah me-nunjukkan bahwa antara kalus transfor-man dan kontrol tidak berbeda, semuamenunjukkan warna biru. Hal tersebut ke-
mungkinan disebabkan oleh suatu senya-wa tertentu yang terdapat dalam kalusbawang apabila diberi X-gluc menunjukkanreaksi berwarna biru.
Kalus yang ditanam dalam media se-leksi setelah ditransformasi dengan A.tumefaciens memperlihatkan bahwa bak-teri cefotaxine media yang mengandungantibiotik (250 dan 500 mg/1) tidak tumbuh(Tabel 111.46).
Tabel 111.46 menunjukkan bahwa kalusyang mati bervariasi antara 10-70% tergan-tung kombinasi perlakuan. Kombinasi per-lakuan terbaik adalah cefotaxime 250 mg/1,masa inkubasi 3 hari, dan konsentrasi glu-kosa 40 g/1 pada media MS, sedangkankombinasi perlakuan cefotaxime 500 mg/1,masa inkubasi 3 hari, dan konsentrasiglukosa 20 g/1 tidak menunjukkan hasilyang memuaskan.
Tabel 111.47 menunjukkan bahwa konsentrasi hygromycine 100 mg/1 sudah dapatmematikan kalus bawang merah (berwarna putih) setelah diinkubasi dalam ruangkultur selama 14 hari, sedangkan untukkonsentrasi 50 mg/1 kalus bawang merahmasih hidup (berwarna kuning) samadengan kontrol (0 mg/1 hygromycine)
Kalus bawang merah yang masihhidup pada media cefotaxime 250 mg/1 dan500 mg/1 sebanyak 19 petri. Masing-masing10 unit kalus ditanam pada media regene-rasi 1 dan setelah diinkubasi selama 3-4minggu, dipindah ke media regenerasi 2.
TRANSFORMASI GENETIK UNTUKPENCIPTAAN VARIETAS TAHAN OPT DAN
TAHAN PENYIMPANAN TANAMANHORTIKULTURA
Eri Sofiari, Sutrisno, S. Purnomo, Budi Marwoto,Saptowo J. Pardal, Budi Santosa, A. Sutanto,
Makful, Lia Sanjaya, Diani Damayanti,Tri Indraini R. Utami, dan Slamet
Teknologi transgenik telah membantupemuliaan dalam perbaikan karaktertanaman yang sulit dilakukan dengan carapersilangan konvensional. Rekayasa gene-tik mengalami kemajuan pesat dalam duapuluh tahun terakhir. Negara-negara yangmenolak produk transgenik di Eropa se-perti Inggris, Perancis, dan Jerman ternyatapenelitiannya berjalan terus dan mendapatdukungan dana yang besar. Terlepas dariadanya pihak yang belum menerima produk tanaman transgenik di Indonesia, risetrekayasa genetik perlu karena banyak per-soalan crop improvement seperti ketahan-an terhadap organisme pengganggu tanaman (OPT) perlu didukung dengan tek-noiogi ini. Penggunaan pestisida yang me-lebihi ambang batas telah menjadi kekha-watiran bersama sehingga, perekayasaanvarietas baru sebagai komponen teknologiproduksi yang ramah lingkungan agar di-peroleh produk pertanian yang bebas pestisida perlu dilakukan. Pemanfaatan protein kristalin yang bersifat insektisida biologis yang dikode oleh gen famili cry adalahteknologi masa depan yang akan dikem-bangkan untuk menanggulangi seranggahama termasuk di antaranya ulatgrayak[Spodoptera spp.) pada bawang merahdan krisan serta Helicovewa armigera padatomat.
Laporan Tahunan Balitbiogen 2002
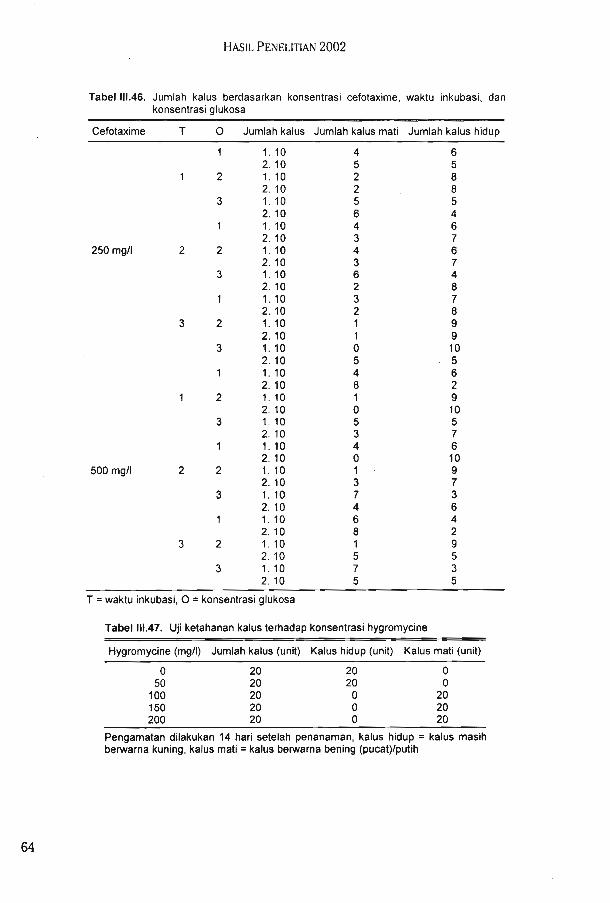
64
Pengamatan dilakukan 14 hari setelah penanaman, kalus hidup = kalus masihberwarna kuning, kalus mati = kalus berwarna bening (pucat)/putih
202020
00
Kalus mati (unit)
000
2020
Kalus hidup (unit)
2020202020
Jumlah kalus (unit)
200150100
500
Hygromycine (mg/l)
T = waktu inkubasi, O = konsentrasi glukosa
Tabel III.47. Uji ketahanan kalus terhadap konsentrasi hygromycine
6588546767487899
105629
105761097364295355
75186473104350184501123263434652254
2. 101. 102.101.102. 101. 102.101.102. 101.102. 101.102. 101. 102.101. 102. 101. 102. 101. 102. 101.102. 101.102. 101.102.101. 102.101. 102. 101. 102. 101.102. 101. 10
3
2
1
3
2
1
3
2
1
3
2
1
3
2
1
3
2
1
500 mg/l
250 mg/l
O Jumlah kalus Jumlah kalus mati Jumlah kalus hidupCefotaxime
Tabel 111.46. Jumlah kalus berdasarkan konsentrasi cefotaxime, waktu inkubasi, dankonsentrasi glukosa
Hasil Penelitian 2002

65
masi ke dalam sel kalus. Hasil transformasidisajikan pada Tabel III.50.
Krisan
Tiga varietas krisan, yaitu Dewi Sartika(DS), Retno Dumilah (RD), dan Larasati(LS) telah digunakan dalam transformasimelalui vektor A. tumefaciens yang mengandung gen reporter gus (p. glucuroni-dase). Dua macam eksplan, yaitu interno-
dus (ruas batang) dan potongan daun mu-da telah diinokulasi dengan A. tumefaciensstrain EHA 105 yang mengandung plasmidpCambia 1301 (gen gus dan hph) denganperlakuan kerapatan bakteri (OD) : 0,5 : 1,2, lama perendaman 5, 10, 15 menit, ko-
kultivasi 5 dan 7 hari.
TATBTCTDTE
OD1.0OD0.5Perlakuan
HSK = hari setelah ko-kultivasi, TA = tomat kultivar LV6117, TB = tomat kultivar LV 6118, TC = tomat kultivarLV 6119, TD = tomat varietas Intan, TE = tomat kultivarCln 294, B1 dan B2 = OD bakteri 0,5 dan 1,0
Tabel 111.49. Banyaknya spot biru pada eksplan tomattransforman hasil uji gus
TA-BITA-B2TB-BITB-B2TC-BITC-132TD-131TD-B2TE-BITE-B2
1 HSK 2 HSK 3 HSK 5 HSKPerlakuan
Tabel 111.48. Pertumbuhan bakteri pada 1, 2, 3, dan 5HSK
Uji gus menunjukkan bahwa antaraeksplan transforman dengan kontrol (nontransformasi) memberikan respon yangsama, yaitu semuanya menunjukkan spot
biru.
Hasil uji gus: P8S = 14 petri masing-masing 10 unit kalus hasil biru, kontrol(P8S) 2 petri hasil biru, sedangkan NS = 13petri masing-masing 10 unit kalus hasilbiru, kontrol (P4S) 2 petri hasil biru.
Kalus yang sudah diseleksi dalammedia seleksi, sampai saat ini masih hidupkurang lebih tiga minggu. Diduga kalustersebut mengandung gen hph.
Tomat
Lima kultivar tomat (LV 6117, LV 6118,LV 6119, Intan, dan Cln 294) koleksi Balitsa,Lembang digunakan dalam percobaan ini.Tabel 111.48 menunjukkan bahwa setelah 5had inkubasi bakteri tumbuh. Uji gus ter-hadap 20 eksplan menggunakan larutan X-gluc menunjukkan tiga eksplan positifmengekspresikan gen gus, yaitu tomat kultivar LV6118 (TB) dan Cln 294 (TE) padaOD 1,0, serta tomat kultivar LV6119 padaOD 0,5 seperti yang disajikan pada Tabel111.49.
Pepaya
Sebanyak 113 kalus pepaya varietasDampit dan 195 kalus varietas Sari Rona te-lah berhasil ditransformasi dengan metodepenembakan partikel. Pada perlakuan per-tama sebanyak 120 kalus dan setiap varietas ditransformasi dengan gen gus (pRQ-6)dan beberapa kalus menunjukkan ekspresisementara yang ditandai dengan berubah-nya warna kalus dari kuning menjadi biruakibat bereaksi dengan larutan X-gluc.Larutan ini berfungsi sebagai pendeteksiadanya gen pelapor (gen gus). Hal ini menunjukkan gen pelapor telah tertransfor-
Laporan Tahunan Balitbiogen 2002
66
000
7hr
000
5hr
15 menit
(%)
0 012,5 025 0
5 hr 7 hr
10 menit
012,537,5
7hr
0.0
0
5hr
5 menit
Larasati (rendam)
0 00 00 0
5 hr 7 hr
15 menit
037,5
0
7hr
0 50 00 75 250 62,5 0
5 hr 7 hr 5 hr
5 menit 10 menit
(rendam) (%)
krisan hasil transformasi dengan gen gus melalui A.
000
7hr
000
5hr
15 menit
(%)
0 00 00 0
5 hr 7 hr
10 menit
000
7hr
000
5hr
5 menit
Larasati (rendam)
0 00 00 0
5hr 7hr
15 menit
000
7hr
Retno Dumilah
0 0 037,5 0 050 0 0
7 hr 5 hr 7 hr
62,5 012,5 2562,5 37^5
7hr 5hr
000
5hr
5 menit 10 menit 15 menit
(rendam) (%)Dewi Sartika
0,51,02,0
OD
Jumlah eksplan 8 ruas/perlakuan ODeoo = 1 x 108sel/ml
Tabel III.52. Persentase gus positif pada eksplan daun mudatumefaciens
0 0 00 0 00 0 0
5hr 7hr 5 hr
5 menit 10 menit
(rendam) (%)Retno Dumilah
0 0 00 0 00 0 0
7 hr 5 hr 7 hr
0 00 00 0
7 hr 5 hr
000
5 hr
5 menit 10 menit 15 menit
(rendam) (%)Dewi Sartika
0,51,02,0
OD
'jarak tembak terhadap kalus target
Tabel 111.51. Persentase gus positif pada eksplan ruas batang krisan hasil transformasi dengan gen gus melalui A.tumefaciens
124
9 cm*
11 2
5 cm* 7cun*
Sari Rona
771
9 cm*
512
7cun*
Dampit
Jumlah sampel positif uji gus
332
5 cm*
303060
Sari Rona -
o o oCO CO CD
Dampit
Jumlah sampel
123
Ulangan
Tabel 111.50. Jumlah sampel varietas Dampit dan Sari Rona positif uji gus pada berbagaijarak tembak
Dengan demikian, eksplan daun mudaChrysantenum sesuai untuk transformasi.
Dari ketiga varietas yang digunakan,temyata eksplan daun muda krisan DewiSartika menunjukkan hasil yang paling baikdibandingkan dengan dua varietas lain.Perlakuan inokulasi terbaik adalah dengankerapatan bakteri (2 x 108 sel/ml), lamaperendaman 5 atau 10 menit dan lama ko-
kultivasi 7 hari.
Tabel 111.51 menunjukkan hasil ujiekspresi gus pada eksplan mas batang(intemodus). Di mana semua eksplan padasetiap perlakuan menunjukkan hasil nega-tif (tidak ada spot biru). Pada Tabel 111.52terlihat bahwa eksplan daun muda lebihsesuai untuk transformasi gen gus melalui
Agrobacterium. Sebagian besar eksplan pada semua perlakuan menunjukkan per-tumbuhan koloni bakteri A. tumefaciens.
Gus menunjukkan hasil uji gus positif.
Hasil Penelitian 2002

67
akan dimanfaatkan di Indonesia harus di-evaluasi dan dilakukan pengujian secarabertahap di Fasilitas Uji Terbatas (FUT)hingga lapangan terbatas.
Jagung Transgenik
Pengujian dilakukan pada jagungtransgenik hasil penelitian Balitbiogen dengan kode A, B, C, D, dan E yang berasal
dari kalus yang sama, jagung Antasena di-gunakan sebagai kontrol. Hasil pengamat-
an terhadap peubah pertumbuhan tanaman selama delapan minggu (fase vegetatif)disajikan pada Tabel 111.53.
Tinggi tanaman jagung transgenik Adan C lebih rendah dibandingkan dengankontrol sejak minggu kedua hingga minggukedelapan pengamatan. Hasil analisis sta-
tistik data pada minggu kedelapan menun-jukkan perbedaan tersebut tidak nyata me-nurut uji t pada taraf 5% (P jagung A =0,184; P jagung C = 0,176).
Kelima jagung transgenik yang diujiberasal dari sumber kalus yang sama. Be-
berapa perbedaan penampilan morfologiyang terlihat dari kelima jagung transgenikini menunjukkan indikasi adanya variasigenetik dari tanaman transgenik yang di-hasilkan. Analisis dengan uji t pada saatpertumbuhan vegetatif maksimum (8minggu) menunjukkan tinggi tanaman danpanjang daun jagung transgenik D lebih ke-cil dibandingkan jagung Antasena sebagaikontrol dan daun jagung transgenik A lebihlebar dibandingkan dengan kontrol, se-dangkan peubah pertumbuhan lain padajagung transgenik A, B, C, dan E tidak ber-beda dibandingkan dengan kontrol.
Pengamatan morfologi yang dilakukantidak menunjukkan adanya gejala pertumbuhan yang ekstrim berupa abnormalitas/kelainan fenotipik yang mengarah padasifat weedy dan invasive.
ANALISIS DAN PENDUGAAN RISIKOPEMANFAATAN TANAMAN TRANSGENIK
M. Herman, Sutrisno, Ida H. Somantri,A. Dinar Ambarwati, Saptowo J. Pardal,Diani Damayanti, Tri Indraini R. Utami,Miftakhurohmah, dan Retno Pangestuti
Ada kekhawatiran sekelompok masya-rakat terhadap tanaman transgenik. Kekhawatiran tersebut bisa menjadi anggapanbahwa tanaman hasil rekayasa genetik da-pat memindahkan gen ke kerabat liar,
menjadi gulma super yang bersifat weedydan invasive, menimbulkan dampak nega-
tif bagi organisme bukan sasaran sepertiserangga berguna, menyebabkan alergi,
menyebabkan timbulnya hama super, ke-racunan, atau bakteri di dalam perut men
jadi resisten terhadap antibiotik akibatpenggunaan markah tahan antibiotik dalam tanaman transgenik. Oleh karena itu,sebelum tanaman transgenik dikomersiali-
sasikan perlu dilakukan evaluasi dan kajianteknis aspek keamanan hayati.
Sehubungan dengan hal tersebut telahdikeluarkan Keputusan Menteri PertanianNo. 856/Kpts/HK.330/9/1997 tentang Keten-tuan Keamanan Hayati Produk Bioteknolo-gi Pertanian Hasil Rekayasa Genetik. Karena di dalam Keputusan Menteri Pertaniantersebut belum mencakup aspek keaman
an pangan maka telah ditetapkan Keputusan Bersama Menteri Pertanian, Menteri Ke-hutanan dan Perkebunan, Menteri Kesehat-
an, dan Menteri Negara Pangan dan Horti-kultura No. 998. l/Kpts/OT.210/9/99; 79O.a/KptsIX/1999; 1145A/MENKES/SKB/IX/1999;015A/Nmeneg PHOR/09/1999 tentangKeamanan Hayati dan Keamanan PanganProduk Pertanian Hasil Rekayasa Genetikyang telah ditandatangani pada tanggal 29September 1999. Dalam Keputusan Bersama Empat Menteri tersebut ditentukanbahwa setiap tanaman transgenik yang
Laporan Tahunan Balitbiogen 2002
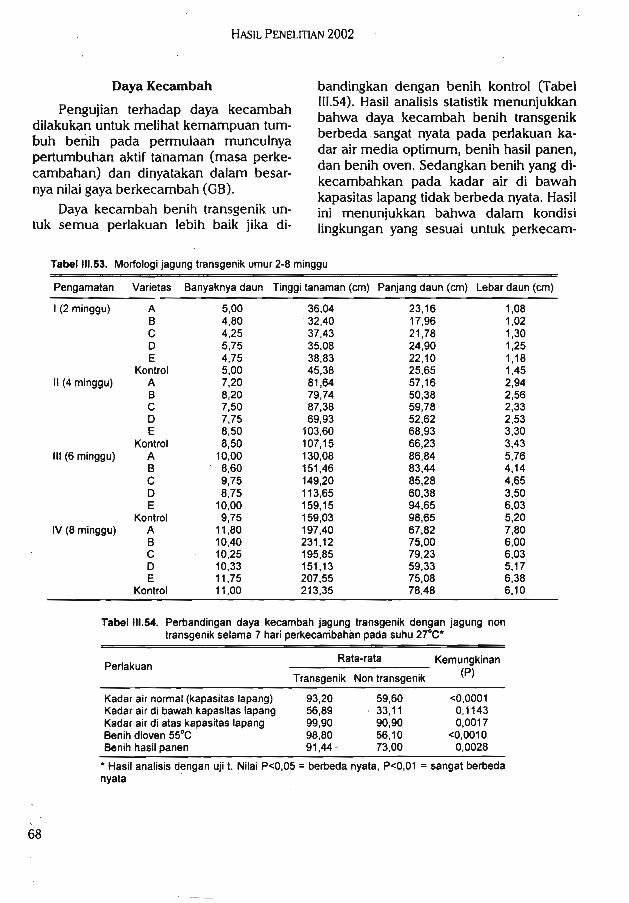
68
Kadar air normal (kapasitas lapang)93,2059,60<0,0001Kadar air di bawah kapasitas lapang56,89• 33,110,1143Kadar air di atas kapasitas lapang99,9090,900,0017Benih dioven 55C98,8056,10<0,0010Benih hasil panen91,4473,000,0028
* Hasil analisis dengan uji t. Nilai P<0,05 = berbeda nyata, P<0,01 = sangat berbeda
nyata
Kemungkinan(P)Transgenik Non transgenik
Rata-rataPerlakuan
Tabel 111.54. Perbandingan daya kecambah jagung transgenik dengan jagung nontransgenik selama 7 hari perkecambahan pada suhu 27C*
6,106,385,176,036,007,805,206,033,504,654,145,763,433,302,532,332,562,941,451,181,251,301,021,08
78,4875,0859,3379,2375,0067,8298,6594,6560,3885,2883,4486,8466,2368,9352,6259,7850,3857,1625,6522,1024,9021,7817,9623,16
213,35207,55151,13195,85231,12197,40159,03159,15113,65149,20151,46130,08107,15103,6069,9387,3879,7481,6445,3838,8335,0837,4332,4036,04
11,0011,7510,3310,2510,4011,809,75
10,008,759,758,60
10,008,508,507,757,508,207,205,004,755,754,254,805,00
KontrolEDCBA
KontrolEDCBA
KontrolEDCBA
KontrolEDCBA
IV (8 minggu)
III (6 minggu)
II (4 minggu)
1 (2 minggu)
Pengamatan Varietas Banyaknya daun Tinggi tanaman (cm) Panjang daun (cm) Lebar daun (cm)
Tabel 111.53. Morfologi jagung transgenik umur 2-8 minggu
bandingkan dengan benih kontrol (Tabel111.54). Hasil analisis statistik menunjukkanbahwa daya kecambah benih transgenikberbeda sangat nyata pada perlakuan ka-dar air media optimum, benih hasil panen,dan benih oven. Sedangkan benih yang di-kecambahkan pada kadar air di bawahkapasitas lapang tidak berbeda nyata. Hasilini menunjukkan bahwa dalam kondisilingkungan yang sesuai untuk perkecam-
Daya Kecambah
Pengujian terhadap daya kecambahdilakukan untuk melihat kemampuan tum-buh benih pada permulaan munculnyapertumbuhan aktif tanaman (masa perke-cambahan) dan dinyatakan dalam besar-nya nilai gaya berkecambah (GB).
Daya kecambah benih transgenik untuk semua perlakuan lebih baik jika di-
Hasil Penelitian 2002

69
hasil relatif jagung transgenik,3 - total hasilNT = non transgenik, T = transgenik,1 = hasil relatif jagung kontrol,2 •relatif,4 = nilai agresivitas
-0,247
-1
-0,019
-0,066
-0,656
1
Aggresive*
1
1.467
1,323
0,903
1
RYT3
1
1,165
0,705
0.330
0
Rb2
0
0,302
0,618
0,573
1
Ra1
15,133
17,633
10,667
5,000
0,000
Rata-rataBM T (g)
0,000
5,800
11,867
11,000
19,200
Rata-rataBM NT (g)
302623263026242628282631262829
Totaltanaman
302623202418121414878000
a BanyaknyaIT tanaman T
Rata-rata nilai Agresivitas
000668
121214201923262829
Banyakny
ONT: 100 T (3)0NT:100T(2)0NT:100T(1)25NT : 75 T (3)25NT : 75 T (2)25NT:75T(1)50NT : 50 T (3)5ONT:5OT(2)50NT:50T(1)75NT: 25 T (3)75NT : 25 T (2)75NT:25T(1)100NT: OT (3)100NT: 0T(2)100NT: 0T(1)
Komposisi benih/UI
Tabel 111.55. Hasil pengujian invasiveness jagung transgenik RR dan jagung non transgenik
maupun faktor dalam benih itu sendiri(kandungan air benih/dormansi).
Invasiveness
Pengujian invasiveness dilakukan pada jagung transgenik RR NK 603 var C7603.Hasil pengujian menunjukkan rata-ratanilai agresivitas dari perbandingan jagungnon transgenik (kontrol) dengan jagungtransgenik adalah -0,247. Ini menunjukkanbahwa jagung transgenik memiliki kemampuan kompetisi yang sama dengan jagungnon transgenik dan tidak mempunyaipotensi yang lebih besar untuk menjadiinvasive dibandingkan dengan kontrol. Hasil perhitungan disajikan pada Tabel III.55.
Kapas Transgenik
Pengujian terhadap tanaman kapastransgenik dilakukan dengan mengguna-kan dua varietas, yaitu kapas transgenik DP5690 dan Bt Nu Pearl Bt/RR, sebagai kontroldigunakan jagung non transgenik DP 5690non Bt.
bahan, yaitu pada suhu 27C dengan kadarair media optimum, benih jagung transge-nik memiliki daya tumbuh yang lebih baikjika dibandingkan benih kontrol. Demikianjuga pada kondisi kadar air tinggi (di ataskapasitas lapang) dan kandungan air benihyang rendah (benih kering mutlak setelahdioven pada suhu 55C selama tujuh hari)benih jagung transgenik lebih mampumengatasi cekaman yang ada sehingga te-tap dapat berkecambah dengan baik (dayatumbuh di atas 90%), namun pada kondisikandungan air dalam media perkecambah-an rendah, benih transgenik maupun kontrol tidak dapat berkecambah dengan baik(daya tumbuh di bawah 90%).
Hasil yang diperoleh dari pengamatanterhadap daya kecambah mengindikasikanbahwa benih jagung transgenik RR NK 603var C7603 memiliki kemampuan yang lebihbaik dibandingkan dengan benih kontroldalam mengatasi kendala yang ada padaproses perkecambahannya baik kendalaakibat faktor luar (cekaman kelembaban)
Laporan Tahunan Balitbiogen 2002

70
DP 5690 Bt lebih pendek dibandingkan dengan kontrol. Perbedaan ini berbeda nyatamenurut uji t pada taraf 5% (Tabel 111.56).
Daun tanaman kapas transgenik DP5690 Bt sejak awal pengamatan lebih sem-pit dibandingkan dengan kontrol. Perbedaan lebar daun semakin signifikan sejalandengan bertarnbahnya umur tanaman.
Pada akhir fase vegetatif (8 MST) uji t menunjukkan perbedaan yang sangat nyata(P<0,0001, Tabel 111.56).
Diameter batang tanaman kapas transgenik DP 5690 Bt hingga minggu keempatpengamatan tidak berbeda dengan kontrol,namun dalam proses pertumbuhan selan-
jutnya lebih besar dibandingkan dengankontrol. Tabel III.56 menunjukkan, per
bedaan tersebut nyata menurut uji t padataraf 5%.
Umur tanaman kapas transgenik DP5690 Bt saat muncul bunga pertama lebihcepat satu had (63 had) dibandingkan dengan kontrol (64 had). Munculnya bollpertama pada kapas transgenik DP 5690 Btlebih cepat dua had (68 had) jika dibandingkan dengan kontrol (70 had). Bollkapas transgenik DP 5690 Bt lebih banyak(7,32,6) dibandingkan dengan kontrol(3,40,8) dan berbeda sangat nyata menurut uji t (P = 0,003). Diameter boll kapastransgenik DP 5690 Bt sedikit lebih besar(2,00,l) dibandingkan dengan kontrol(2,0+.0,l), tidak berbeda nyata menurut ujit (P = 0,357).
Meskipun terdapat beberapa perbedaan pada morfologi tanaman kapas transgenik DP 5690 Bt dibandingkan dengan kontrol namun tidak ditemukan perbedaanyang ekstrim berupa abnormalitas/kelainanfenotipik yang mengarah pada sifat weedydan invasive.
Morfologi Kapas
Kapas Transgenik DP 5690 Bt
Hasil pengamatan terhadap peubahmorfologi kapas transgenik DP 5690 Bt di-bandingkan dengan kapas kontrol selamadelapan minggu (fase vegetatif) disajikanpada Gambar III. 14. Dari Gambar III. 14 ter-
lihat bahwa tinggi tanaman kapas transgenik DP 5690 Bt tidak berbeda dengan kapaskontrol. Pada saat dicapai pertumbuhan
vegetatif maksimum (8 MST) analisis dengan uji t tidak menunjukkan perbedaanyang nyata (Tabel 111.56).
Banyaknya daun tanaman kapas trans
genik DP 5690 Bt pada minggu kedua hing-ga keempat dan kedelapan tidak berbedadengan kontrol namun pada minggu ke-enam, jumlah daun tanaman kapas trans
genik DP 5690 Bt lebih banyak dibanding-kan dengan kontrol.
Banyaknya cabang vegetatif tanamankapas transgenik DP 5690 Bt tidak berbedadengan kontrol sejak minggu kedua hinggakedelapan pengamatan (P = 0,256, Tabel111.56). Banyaknya cabang generatif tanaman kapas transgenik DP 5690 Bt pada minggu keempat tidak berbeda dengan kontrolnamun pada minggu keenam, cabang generatif tanaman kapas transgenik DP 5690Bt lebih banyak dibandingkan dengan kontrol. Selama minggu keenam sampai minggu kedelapan laju pembentukan cabanggeneratif tanaman kapas transgenik DP5690 Bt lebih rendah dibandingkan dengankontrol, sehingga cabang pada minggu kedelapan relatif sama dengan kontrol (P =0,741, Tabel 111.56).
Panjang daun tanaman kapas transgenik DP 5690 Bt sampai minggu keempatpengamatan tidak berbeda dengan kontrol,namun selama minggu keempat sampaikedelapan daun tanaman kapas transgenik
Hasil Penelitian 2002
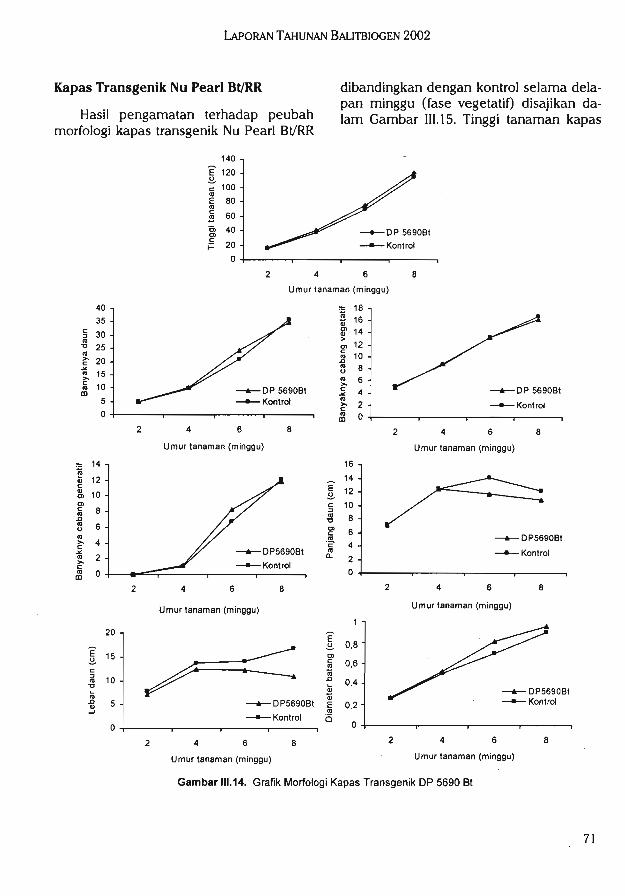
71
Gambar 111.14. Grafik Morfologi Kapas Transgenik DP 5690 Bt
46Umur tanaman (minggu)Umur tanaman (minggu)
46
Umur tanaman (minggu)Umur tanaman (minggu)
-DP5690Bt- Kontrol
46Umur tanaman (minggu)
161412 -10 -
86420
46Umur tanaman(minggu)
518 n1516S"14 •a12ra10
46
Umur tanaman (minggu)
140 n? 120oIT 100 •I 80a 60 -S 40
j! 20
0
4035302520 -15
50
dibandingkan dengan kontrol selama dela-pan minggu (fase vegetatiO disajikan da-lam Gambar III. 15. Tinggi tanaman kapas
Kapas Transgenik Nu Pearl Bt/RR
Hasil pengamatan terhadap peubahmorfologi kapas transgenik Nu Pearl Bt/RR
Laporan Tahunan Balitbiogen 2002
72
'Hasil analisis dengan uji t, P<0,05 - berbeda nyata, P<0,01 = sangatberbeda nyata
<0,0001<0,00010,0070,0111,0000,0520,051
16,912,20,9
12,116,735,8
115,3
Kontrol
10,910,3
1,013,916,740,1
121,1
Nu Pearl
Lebar daun (cm)Panjang daun (cm)Diameter batang (cm)Cabang generatifCabang vegetatifBanyaknya daunTinggi tanaman (cm)
Tabel 111.57. Peubah morfologi kapas transgenik Nu Pearl dengankontrol pada 8 MST *
berbeda nyata, P<0,01 - sangat'Hasil analisis dengan uji t, P<0,05 =berbeda nyata
Tinggi tanaman (cm)120,5115,30,122Banyaknya daun34,735,80,620Cabang vegetatif16,316,70,256Cabang generatif11,812,10,741Diameter batang (cm)1,00,90,043Panjang daun (cm)10,912,20,001Lebar daun (cm)13,016,9<0,0001
• KemungkinanDP 5690 Bt Kontrol
Jenis kapasPeubah
Tabel 111.56. Peubah morfologi kapas transgenik DP 5690 Bt dengankontrol pada 8 MST *
keempat lebih banyak dibandingkan dengan kontrol. Hasil uji t pada minggu kedelapan menunjukkan beda nyata pada taraf5%(P = 0,011, Tabel 111.57).
Daun tanaman kapas transgenik Nu
Pearl sejak awal pengamatan lebih pendekdibandingkan dengan kontrol, perbedaanini sangat nyata menurut uji t pada minggukedelapan (Tabel 111.57).
Lebar daun tanaman kapas transgenikNu Pearl hingga minggu keenam tidak berbeda dibandingkan dengan kontrol, namunpada minggu kedelapan daun kapas transgenik Nu Pearl menjadi lebih sempit dibandingkan dengan kontrol dan berbeda sangat nyata menurut uji t (P<0,0001, Tabel111.57).
transgenik Nu Pearl tidak berbeda dengankontrol (Gambar III. 15). Pada saat dicapaipertumbuhan vegetatif maksimum (8 MST)analisis dengan uji t tidak menunjukkanperbedaan yang nyata (label 111.57).
Daun tanaman kapas transgenik Nu
Pearl terlihat lebih banyak dibandingkandengan kontrol sejak minggu kedua hinggakedelapan. Analisis dengan uji t pada minggu kedelapan tidak menunjukkan perbedaan yang nyata (P = 0,052, Tabel 111.57).Cabang vegetatif kapas transgenik Nu Pearltidak berbeda dengan kontrol sejak minggukedua hingga kedelapan. Pada minggu kedelapan cabang vegetatif kapas transgenikNu Pearl sama banyak dengan kontrol (P =1, Tabel 111.57). Cabang generatif tanamankapas transgenik Nu Pearl sejak minggu
Hash Penelitian 2002
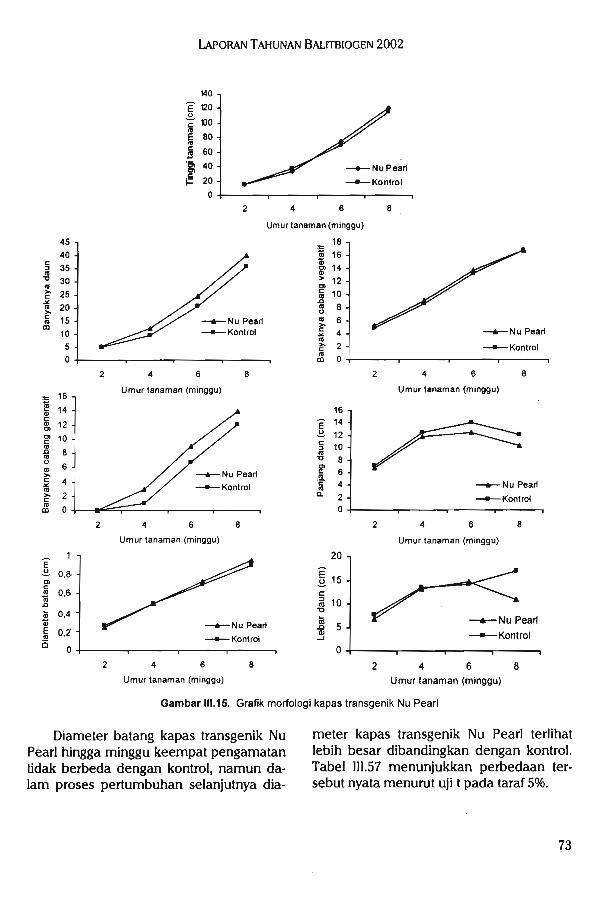
73
meter kapas transgenik Nu Pearl terlihatlebih besar dibandingkan dengan kontrol.Tabel III.57 menunjukkan perbedaan ter-sebut nyata menurut uji t pada taraf 5%.
Diameter batang kapas transgenik NuPearl hingga minggu keempat pengamatantidak berbeda dengan kontrol, namun da-lam proses pertumbuhan selanjutnya dia
Gambar 111.15. Grafik morfologi kapas transgenik Nu Pearl
468Umur tanaman (minggu)
246Umur tanaman (minggu)
earlKontral
20I 15-
I 10 -T3
_l0
246Umur tanaman (minggu)Umur tanaman (minggu)
-Nu Pearl-Kontral
1614121086420
Umur tanaman (minggu)Umur tanamanminu
1
0.&
20,6S^^0,4IS
.2:50
01
I
Laporan Tahunan Balitbiogen 2002
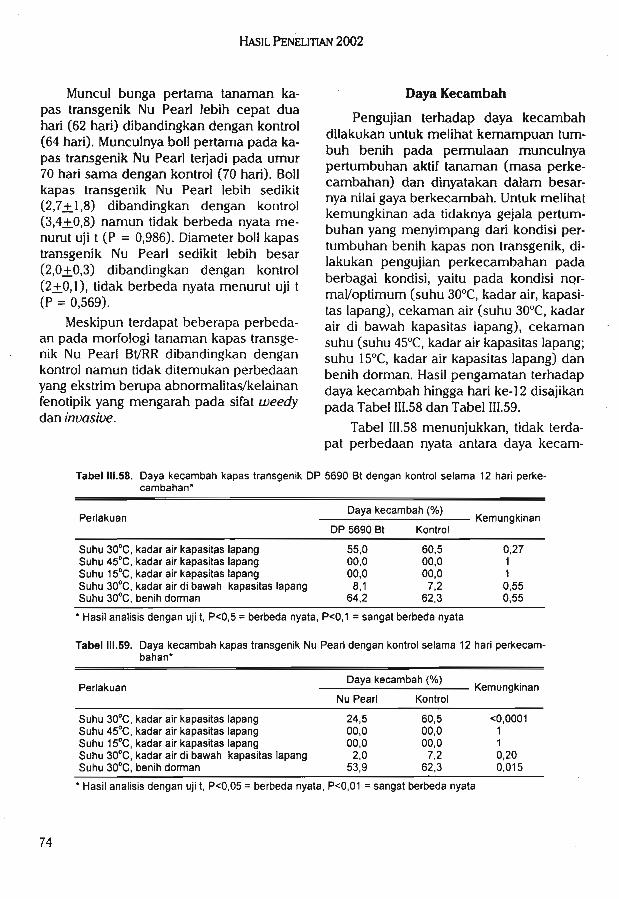
74
' Hasil analisis dengan uji t, P<0,05 = berbeda nyata, P<0,01 = sangat berbeda nyata
0,0150,2011
<0,0001
62,37,2
00,000,060,5Suhu 30C, kadar air kapasitas lapang24,5
Suhu 45C, kadar air kapasitas lapang00,0Suhu 15C, kadar air kapasitas lapang00,0Suhu 30C, kadar air di bawah kapasitas lapang2,0Suhu 30C, benih dorman53,9
KemungkinanKontrolNu Pearl
Daya kecambah (%)Perlakuan
* Hasil analisis dengan uji t, P<0,5 = berbeda nyata, P<0,1 = sangat berbeda nyata
Tabel III.59. Daya kecambah kapas transgenik Nu Pearl dengan kontrol selama 12 hari perkecambahan*
Suhu 30C, kadar air kapasitas lapangSuhu 45C, kadar air kapasitas lapangSuhu 15C, kadar air kapasitas lapangSuhu 30C, kadar air di bawah kapasitas lapangSuhu 30C, benih dorman0,55
0,55110,27
62,37,2
00,000,060,5
Kontrol
64,28,1
00,000,055,0
DP 5690 Bt
Daya kecambah (%)Perlakuan
Tabel 111.58. Daya kecambah kapas transgenik DP 5690 Bt dengan kontrol selama 12 hari perkecambahan*
Daya Kecambah
Pengujian terhadap daya kecambahdilakukan untuk melihat kemampuan tum-buh benih pada permulaan munculnyapertumbuhan aktif tanaman (masa perke-
cambahan) dan dinyatakan dalam besar-nya nilai gaya berkecambah. Untuk melihatkemungkinan ada tidaknya gejala pertumbuhan yang menyimpang dari kondisi pertumbuhan benih kapas non transgenik, dilakukan pengujian perkecambahan padaberbagai kondisi, yaitu pada kondisi nqr-mal/optimum (suhu 30C, kadar air, kapasi-tas lapang), cekaman air (suhu 30C, kadarair di bawah kapasitas lapang), cekamansuhu (suhu 45C, kadar air kapasitas lapang;suhu 15C, kadar air kapasitas lapang) danbenih dorman. Hasil pengamatan terhadap
daya kecambah hingga hari ke-12 disajikanpada Tabel 111.58 dan Tabel 111.59.
Tabel 111.58 menunjukkan, tidak terdapat perbedaan nyata antara daya kecam-
Muncul bunga pertama tanaman ka-
pas transgenik Nu Pearl lebih cepat duahari (62 hari) dibandingkan dengan kontrol(64 hari). Munculnya boll pertama pada ka-pas transgenik Nu Pearl terjadi pada umur70 hari sama dengan kontrol (70 hari). Bollkapas transgenik Nu Pearl lebih sedikit(2,71,8) dibandingkan dengan kontrol(3,40,8) namun tidak berbeda nyata me-
nurut uji t (P = 0,986). Diameter boll kapastransgenik Nu Pearl sedikit lebih besar(2,00,3) dibandingkan dengan kontrol(20,l), tidak berbeda nyata menurut uji t(P = 0,569).
Meskipun terdapat beberapa perbeda-an pada morfologi tanaman kapas transge
nik Nu Pearl Bt/RR dibandingkan dengankontrol namun tidak ditemukan perbedaanyang ekstrim berupa abnormalitas/kelainanfenotipik yang mengarah pada sifat weedydan invasive.
Hasil Penelitian 2002
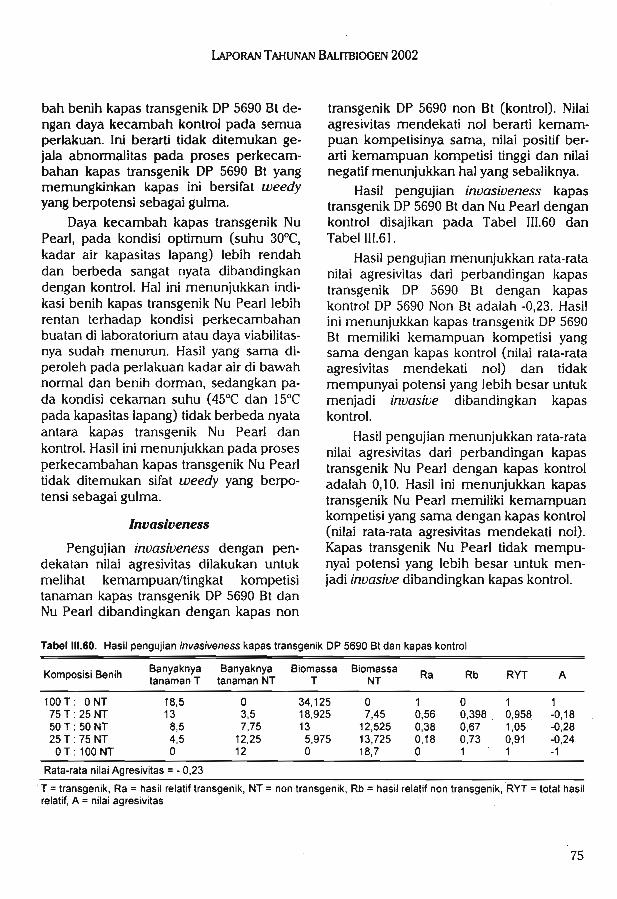
75
T = transgenik, Ra = hasil relatif transgenik, NT = non transgenik, Rb = hasil relatif non transgenik, RYT = total hasilrelatif, A = nilai agresivitas
1-0,18-0,28-0,24-1
A
10,9581,050,911
RYT
00,3980,670,731
Rb
10,560,380,180
Ra
07,45
12,52513,72518,7
BiomassaNT
34,12518,92513
5,9750
BiomassaT
03,57,75
12,2512
Banyaknyatanaman NT
Rata-rata nilai Agresivitas = - 0,23
18,5138,54,50
Banyaknyatanaman T
100 T: 0NT75 T : 25 NT50 T : 50 NT25 T : 75 NT0T:100NT
Komposisi Benih
Tabel 111.60. Hasil pengujian invasiveness kapas transgenik DP 5690 Bt dan kapas kontrol
transgenik DP 5690 non Bt (kontrol). Nilaiagresivitas mendekati nol berarti kemam-puan kompetisinya sama, nilai positif berarti kemampuan kompetisi tinggi dan nilainegatif menunjukkan hal yang sebaliknya.
Hasil pengujian invasiveness kapastransgenik DP 5690 Bt dan Nu Pearl dengankontrol disajikan pada Tabel 111.60 danTabel 111.61.
Hasil pengujian menunjukkan rata-ratanilai agresivitas dari perbandingan kapastransgenik DP 5690 Bt dengan kapaskontrol DP 5690 Non Bt adalah -0,23. Hasilini menunjukkan kapas transgenik DP 5690Bt memiliki kemampuan kompetisi yangsama dengan kapas kontrol (nilai rata-rata
agresivitas mendekati nol) dan tidakmempunyai potensi yang lebih besar untukmenjadi invasive dibandingkan kapaskontrol.
Hasil pengujian menunjukkan rata-ratanilai agresivitas dari perbandingan kapastransgenik Nu Pearl dengan kapas kontroladalah 0,10. Hasil ini menunjukkan kapastransgenik Nu Pearl memiliki kemampuankompetisi yang sama dengan kapas kontrol(nilai rata-rata agresivitas mendekati nol).Kapas transgenik Nu Pearl tidak mempunyai potensi yang lebih besar untuk menjadi invasive dibandingkan kapas kontrol.
bah benih kapas transgenik DP 5690 Bt de-ngan daya kecambah kontrol pada semuaperlakuan. Ini berarti tidak ditemukan ge-jala abnormalitas pada proses perkecam-bahan kapas transgenik DP 5690 Bt yangmemungkinkan kapas ini bersifat weedyyang berpotensi sebagai gulma.
Daya kecambah kapas transgenik NuPearl, pada kondisi optimum (suhu 30C,kadar air kapasitas lapang) lebih rendahdan berbeda sangat nyata dibandingkandengan kontrol. Hal ini menunjukkan indi-kasi benih kapas transgenik Nu Pearl lebihrentan terhadap kondisi perkecambahanbuatan di laboratorium atau daya viabilitas-nya sudah menurun. Hasil yang sama di-
peroleh pada perlakuan kadar air di bawahnormal dan benih dorman, sedangkan pada kondisi cekaman suhu (45C dan 15Cpada kapasitas lapang) tidak berbeda nyataantara kapas transgenik Nu Pearl dankontrol. Hasil ini menunjukkan pada prosesperkecambahan kapas transgenik Nu Pearltidak ditemukan sifat weedy yang berpotensi sebagai gulma.
Invasiveness
Pengujian invasiveness dengan pen-dekatan nilai agresivitas dilakukan untukmelihat kemampuan/tingkat kompetisitanaman kapas transgenik DP 5690 Bt danNu Pearl dibandingkan dengan kapas non
Laporan Tahunan Bautbiogen 2002

76
tanaman melinjo dan manggis secara kultur in vitro, formulasi media perakaran danmetode aklimatisasi untuk iles-iles, metode
prakultur yang optimal, dan formulasi krio-protektan yang efektif bagi ubi kayu danyam untuk penyimpanan dengan teknikkriopreservasi dan formulasi media pertu-
nasan dan multiplikasi tanaman kentanghitam dan gembili.
Inisiasi Akar pada Tanaman Melinjomelalui Kultur In Vitro
Pada umur biakan 12 minggu setelahtanam tidak ada pengaruh media dasardan thidiazuron serta interaksinya terhadaptinggi tunas (Tabel 111.62). Pada dua mediayang diuji, peningkatan konsentrasi thidiazuron sampai 0,3 mg/1 tinggi tunasnya lebihpendek daripada thidiazuron kontrol danpeningkatan konsehtrasi hingga 0,1 mg/1.
Jumlah tunas paling banyak 1,6 berasaldari media WPM + thidiazuron 0,1 dan 0,3mg/1, yaitu tidak berbeda nyata dengankombinasi perlakuan lainnya.
Penampakan biakan pada beberapaperlakuan berbeda terutama apabila di-tinjau dari kondisi tunas dan daun. Ukurandaun pada media WPM pada umumnyalebih lebar, dibandingkan dengan ulangandan kadar media Anderson. Terhambatnya
pelebaran daun ini diperkirakan karena pe-nambahan thidiazuron ke dalam media
STUDI REGEIMERASI DAN PENYIMPANANTANAMAN BERNILAI EKONOMI TINGGI
SECARA KULTUR IN VITRONovianti Sunarlim, Yadi Rusyadi, Yati Supriati,
Ika Roostika, Arief V. Noviati, Ika Mariska,Ragapadmi Purnamaningsih, Murtado,
Widiati H. Adil, dan Hadiatmi
Kultur in vitro dapat digunakan untukperbanyakan tanaman yang sukar diperba-nyak secara konvensional dan penyimpan-
an tanaman yang berkembang biak secaravegetatif misalnya umbi-umbian.
Selain digunakan untuk perbanyakantanaman. Biasanya tanaman ubi-ubian di-
simpan dalam bentuk tanaman di kebunkoleksi, namun dengan cara tersebut se-
ringkali terjadi kehilangan genotipe daritanaman karena gangguan hama penyakitdan cekaman lingkungan. Selain itu, diper-
lukan tempat yang luas, tenaga, dan biayayang besar untuk mempertahankan tanam
an di lapang.
Teknik penyimpanan melalui pem-
bekuan dapat dilakukan melalui beberaparhetode antara lain pembekuan konvensio
nal, pembekuan sederhana, dan vitrifikasi(pembekuan vitra cepat). Untuk metodevitrifikasi tidak diperlukan alat khusus untuk penurunan suhu secara bertahap.
Tujuan dari penelitian ini ialah untukmendapatkan metode inisiasi akar pada
T = transgenik, Ra = hasil relatif transgenik, NT = non transgenik, Rb = hasil relatif non transgenik, RYT = total hasil
relatif, A = nilai agresivitas
Rata-rata nilai Agresivitas = - 0,10
-10,0740,0730,1651
10,9151,2511,0151
10,5690,5800,2760
00,3460,6710,7361-
46,8726,6727,1713,070
011,321,924,0332,63
20,714,711
5,30
05,7
11,315,722,3
100 NT75 NT50 NT25 NT
ONT
0T:25 T:50 T:75 T:
100T:
RYTRbRaBiomassa Biomassa.TNT
Banyaknyatanaman NT
Banyaknyatanaman TKomposisl Benih
Tabel 111.61. Hasil pengujian invasiveness kapas transgenik Nu Pearl dan kapas kontrol
Hasil Penelitian 2002

•77
0(0/15)0(0/15)
20(3/15)0(0/15)
13(2/15)20(3/15)33(5/15)26(4/15)
0250500750
Angka-angka dalam satu lajur yang diikuti huruf sama tidak berbeda pada tarafnyata 5% menurut uji DMRT
Tabel III.63. Persentase tunas yang hidup dan berakar setelah 4 bulan diaklimatisasi pada lingkunganeksternal
Konsentrasi IBA (mg/l) Persentase tanaman yang hidup (%) Persentase tanaman yang berakar (%)
1,0"1,2"1,2"
Anderson
0,9"1,6""1,6""
WPM
Jumlah tunas (cm)
1,0"0,9"0,8"
Anderson
1,1"1,1"0,8"
WPM
Tinggi tunas (cm)
kontrol0,10,3
Konsentrasi thidiazuron (mg/l) -
Tabel 111.62. Pengaruh media dasar dan thidiazuron terhadap tinggi dan jumlahtunas pada umur 12 minggu
33%). Persentase tertinggi tanaman yanghidup setelah aklimatisasi (33%) diperolehpada perlakuan perendaman tunas sebe-
lum diaklimatisasi, dalam larutan IBA 500mg/1.
Tabel 111.63 menunjukkan bahwa jum-lah eksplan yang mampu bertahan setelahdiaklimatisasi cukup tinggi tetapi persentase eksplan yang mampu mengeluarkan
akar masih sangat rendah. Perendaman
eksplan dalam larutan IBA pada konsen-tfasi 500 mg/1 selama satu jam mampumenginduksi tumbuhnya perakaran.
Optimasi Sistem Perakaran danAklimatisasi Iles-iles (Amorpophalus sp.)
Pemberian IAA pada media dasar VtMS atau ^.-MS tidak menunjukkan penga-
ruh yang nyata terhadap peningkatan jum-lah akar iles-iles pada umur 12 minggu(Tabel 111.64). Media dasar MS yang diturun-kan kandungan makronya dari V^^ menjadi%, ternyata tidak berpengaruh nyata terhadap jumlah maupun panjang akar, bah-kan cenderung memperpendek panjang
biakan karena thidiazuron memiliki ke-mampuan untuk menginduksi terbeiituk-nya etilen yang dapat menghambat per-luasan daun. Dengan kandungan nitrat le-
bih rendah maka keefektifan etilen dalammerombak auksin lebih tinggi.
Pembentukan akar pada biakan invitro sangat bervariasi tergantung pada spe-sies tanaman itu sendiri. Tanaman dari go-
longan gymnospermae seperti melinjo sangat sulit menginduksi perakarannya, pe-nelitian perakaran melinjo secara in vivopada daun mulai terlihat dua minggu sete-lah tanaman diaklimatisasi. Beberapa ta-namcm menjadi layu dan akhirnya mati,
akibat perbedaan kelembaban udara karena pada kondisi kultur in vitro kelembabansangat tinggi dan daun in vivo umumnya ti-
dak mempunyai kutikula, stomata belumberperan secara sempurna sehingga pro
ses transpirasi yang terjadi sangat tinggi.Persentase tanaman yang hidup setelah 4bulan diaklimatisasi disajikan pada Tabel111.63.
Persentase tanaman yang hidup sete
lah diaklimatisasi relatif kecil (kurang dari
Laporan Tahunan Balitbiogen 2002

78
3,02,22,02,0
Jumlah anakan
7,917,620,412,6
_
Tinggi tanaman (cm)
8080
10010020
Keberhasilan tumbuh (%)
Tanah + komposTanah + sekamTanah + kastingTanah + pupuk kandangTanah
Media aklimatisasi
Angka-angka dalam satu lajur yang diikuti huruf sama tidak berbedapada taraf nyata 5% menurut uji DMRT
Tabel 111.65. Pengaruh media tumbuh terhadap persentase tumbuh kultur in vitro serta tinggitanaman dan jumlah anakan pada umur 13 minggu
3,2"2,1"3,3'4,4"3,4'2,3"2,3"4,1'
Panjang akar (cm)
8,3"6,0'7,4"6,8"9,2'7,4'7,2'5,8'
Jumlah akar
Vi MS, IAA 1,5 mg/I1/2 MS, IAA 1,0 mg/I1/2 MS, IAA 0,5 mg/I1/i MS, tanpa IAA% MS, IAA 1,5 mg/I% MS, IAA 1,0 mg/IV. MS, IAA 0,5 mg/1V. MS, tanpa IAA
Perlakuan
Tabel 111.64. Pengaruh pemberian IAA terhadap jumlah dan panjangakar iles-iles pada umur 12 minggu dalam media dasar MS
ma, yaitu hijau berbercak putih atau belangtetapi permukaan batang agak bervariasiantara iicin dan agak kasar.
Inisiasi Akar Manggis dari Tunas In Vitro
Pemberian charcoal pada media ta-
nam menyebabkan tinggi biakan dan jumlah daun/biakan bertambah. Apabila dilihatsecara visual, biakan tanpa penambahan
charcoal kecil-kecil dan kerdil, dengan di-tambahkannya charcoal sebanyak 0,15%,biakan tumbuh normal dan lebih hijau.Perbedaan media MS dan WPM tidakmemberikan perbedaan tinggi dan jumlahdaun, tinggi biakan sekitar 0,8 cm dan jumlah daun sekitar 6 daun/biakan. Penambahan BAP dari 3 mg/I menjadi 5 mg/1 tidakmempengaruhi tinggi biakan dan jumlahdaun/biakan (Tabel 111.66).
Inisiasi akar awal terbentuk pada umurbiakan 6 bulan pada perlakuan }A MS + IBA5 mg/1. Persentase biakan yang berakar
akar, walaupun secara statistik tidak ber-beda nyata.
Berdasarkan hasil percobaan dapat di-simpulkan bahwa untuk mengakarkan iles-
iles dengan kultur in vitro cukup digunakanmedia dasar MS pada taraf lA formula, tan-pa diberi zat pengatur tumbuh. Tidak ber-pengaruhnya IAA terhadap perakaranmungkin disebabkan kandungan auksinendogen yang dibutuhkan untuk mengini-siasi perakaran sudah cukup tersedia didalam jaringan tanaman.
Tanaman iles-iles hanya bertahan 6minggu pada media tunas kemudian selu-ruh tanaman mati (Tabel 111.65). Tanaman
yang ditumbuhkan pada media campurantanah + kasting rata-rata mencapai 20,4cm pada minggu ke-13. Pada parameter
jumlah anakan tampak tidak ada perbeda-an yang berarti dengan rata-rata jumlahanakan 1 -2 anakan per tanaman.
Umumnya batang iles-iles yang diakli-matisasi mempunyai motif yang hampir sa-
Hasil Penelitian 2002

79
1,23,22,0000
(cm)panjang akar
Rerata
16,750,033,3
000
berakarbiakan yangPersentase
03,01,2000(cm)
panjang akarRerata
050,033,3
000
berakarbiakan yangPersentase
02,60000
(cm)panjang akar
Rerata
050,0
0000
berakarbiakan yangPersentase
V^MS + IBAIOmg/l% MS + IBA 5 mg/l% MS + IBA 10 mg/lV2 MS + IBA 5 mg/lMS + IBA 10 mg/lMS + IBA 5 mg/l
Perlakuan
12 bulan9 bulan6 bulan
Tabel 111.67. Pengaruh beberapa formula MS dan konsentrasi IBA terhadap persentase biakan yang berakar danpanjang akar pada umur 6, 9, dan 12 bulan
Normal, biasaNormal, biasaKerdil, kecilKerdil, kecil
Normal, biasaNormal, biasaKerdil, kecilKerdil, kecil
6,06,04,04,5
5,56,34,34,4
0,820,880.380,35
0,880,780,690,59
WPM + BAP 5 mg/l + charcoal 0,15%WPM + BAP 3 mg/l + charcoal 0,15%WPM + BAP 5 mg/lWPM + BAP 3 mg/l
MS + BAP 5 mg/l + charcoal 0,15%MS + BAP 3 mg/1 + charcoal 0,15%MS + BAP 5 mg/1MS + BAP 3 mg/1
Tinggi biakan (cm) Jumlah daun Visual biakanPerlakuan
Tabel 111.66. Pengaruh berbagai media terhadap tinggi biakan, jumlah daun, dan visual biakanpada umur 5 bulan
Pengaruh Beberapa Krioprotektanterhadap Keberhasilan Penyimpanan
Ubi-ubian secara Kriopreservasi
Berdasarkan data yang diperoleh da-pat diketahui bahwa durasi prakultur hing-ga 4 hari tidak mempengaruhi daya hidupkultur ubi kayu, bahkan persentase kulturyang bertunas tetap tinggi (75-100%). Persentase kalus terendah (28,5%) dihasilkandari perlakuan prakultur selama 4 hari.
Hasil percobaan juga menunjukkan bahwasemaidn lama durasi prakultur cenderungmenyebabkan semakin menurunnya tinggikultur dan jumlah daun (Tabel 111.69).
Hasil percobaan perendaman dalamlarutan vitrifikasi menunjukkan bahwa kultur ubi kayu mempunyai daya hidup yangcukup tinggi (75-100%) pada semua jeniskrioprotektan, durasi prakultur, dan durasiloading yang diujikan. Tinggi kultur dan
sebanyak 50% dan tidak berubah sampaiumur 12 bulan, sedangkan panjang akarbertambah walaupun tidak terlalu banyak,pada umur 12 bulan panjang akar 3,2 cm.Pada perlakuan V2 MS + IBA 10 mg/1 biakanmulai berakar pada umur 9 bulan denganrerata panjang 1,2 cm dan persentase biakan yang berakar 33,3% sampai umur 12bulan. Pada perlakuan Vi MS + IBA 10 mg/1mulai berakar pada umur 12 bulan sebanyak 16,7% (Tabel 111.67).
Panjang akar tidak berbeda di antaraperlakuan yang dicoba, yaitu sekitar 2 cmpada umur 6 bulan (Tabel 111.68). Jumlahakar untuk semua perlakuan hanya satu
biakan.
Laporan Tahunan Balitbiogen 2002
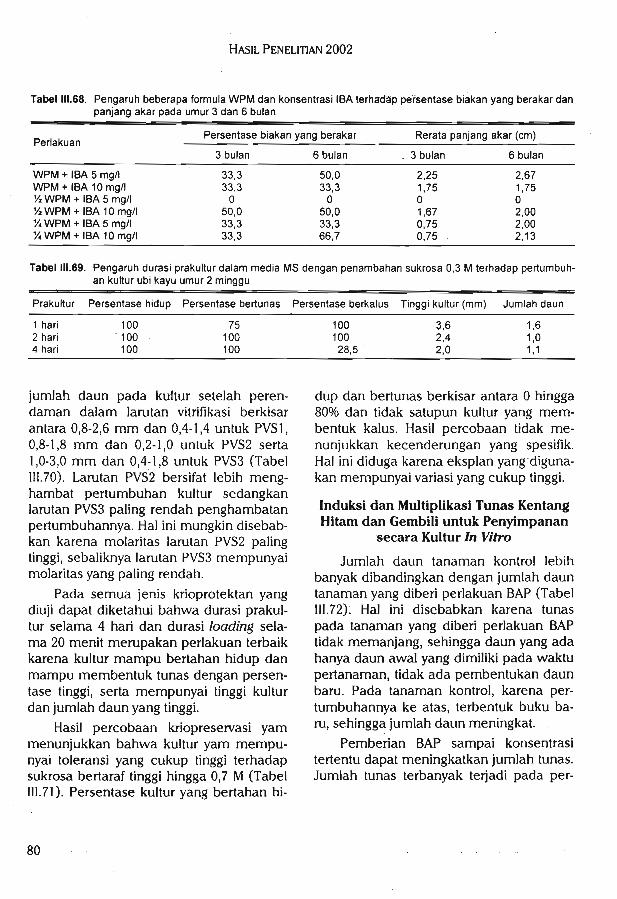
80
dup dan bertunas berkisar antara 0 hingga80% dan tidak satupun kultur yang membentuk kalus. Hasil percobaan tidak menunjukkan kecenderungan yang spesifik.Hal ini diduga karena eksplan yang diguna-kan mempunyai variasi yang cukup tinggi.
Induksi dan Multiplikasi Tunas KentangHitam dan Gembili untuk Penyimpanan
secara Kultur In Vitro
Jumlah daun tanaman kontrol lebihbanyak dibandingkan dengan jumlah dauntanaman yang diberi perlakuan BAP (Tabel111.72). Hal ini disebabkan karena tunaspada tanaman yang diberi perlakuan BAPtidak memanjang, sehingga daun yang adahanya daun awal yang dimiliki pada waktupertanaman, tidak ada pembentukan daunbam. Pada tanaman kontrol, karena per
tumbuhannya ke atas, terbentuk buku barn, sehingga jumlah daun meningkat.
Pemberian BAP sampai konsentrasitertentu dapat meningkatkan jumlah tunas.Jumlah tunas terbanyak terjadi pada per-
jumlah daun pada kultur setelah peren-daman dalam larutan vitrifikasi berkisarantara 0,8-2,6 mm dan 0,4-1,4 untuk PVS1,
0,8-1,8 mm dan 0,2-1,0 untuk PVS2 serta1,0-3,0 mm dan 0,4-1,8 untuk PVS3 (Tabel
111.70). Larutan PVS2 bersifat lebih meng-hambat pertumbuhan kultur sedangkan
larutan PVS3 paling rendah penghambatanpertumbuhannya. Hal ini mungkin disebab-
kan karena molaritas larutan PVS2 palingtinggi, sebaliknya larutan PVS3 mempunyaimolaritas yang paling rendah.
Pada semua jenis krioprotektan yangdiuji dapat diketahui bahwa durasi prakultur selama 4 hari dan durasi loading sela-ma 20 menit merupakan perlakuan terbaikkarena kultur mampu bertahan hidup danmampu membentuk tunas dengan persen
tase tinggi, serta mempunyai tinggi kulturdan jumlah daun yang tinggi.
Hasil percobaan kriopreservasi yam
menunjukkan bahwa kultur yam mempunyai toleransi yang cukup tinggi terhadapsukrosa bertaraf tinggi hingga 0,7 M (Tabel111.71). Persentase kultur yang bertahan hi
1,61,01,1
(mm) Jumlah daun
M terhadap pertumbuh-
3,62,42,0
Tinggi kultur
100100
28,5
Persentase berkalus
MS dengan penambahan sukrosa 0,3
75100100
hidup Persentase bertunas
100' 100
100
Persentase
Pengaruh durasi prakultur dalam mediaan kultur ubi kayu umur 2 minggu
1 hari2 hari4 hari
Prakultur
Tabel 111.69.
2,132,002,0001,752,67
6 bulan
panjang akar (cm)
0,750,751,6701,752,25
. 3 bulan
Rerata
66,733,350,0
033,350,0
6 bulan
yang berakar
33,333,350,0
033,333,3
3 bulan
Persentase biakan
%WPM + IBA10mg/l% WPM + IBA 5 mg/lY2WPM + IBA10mg/lV2 WPM + IBA 5 mg/lWPM + IBA10mg/lWPM + IBA 5 mg/l
Perlakuan
panjang akar pada umur 3 dan 6 bulanTabel 111.68. Pengaruh beberapa formula WPWI dan konsentrasi IBA terhadap persentase biakan yang berakar dan
Hasil Penelitian 2002
804060
04020804060804060
804060
04020804060804060
753175317531
+ asam sitrat 100 ppmMS + sukrosa 0,7 M
+ asam sitrat 100 ppmMS + sukrosa 0,5 M
+ asam sitrat 100 ppmMS + sukrosa 0,3 M
0,81,60,60,41,20,80,81,81,20,61,01,00,20,80,80,60,80,61,40,81,30,60,40,81,00,61,4
1,43,02,40,61,42,01,02,02,21,01,81,60,81,20,81,01,41,21,62,62,00,81,21,61,21,41,8
806060
10010010060
1006020208080
100100100
8010060607580
10010060
100100
1001008060
10010060
10010060
100loo608040808080
100100
758080806080
100
10010010010010010010010010010010010080
100100100100100100100
7580
100100100100100
30'201030201030201030'20'10'30'20'10'30'20'10'30'20'10'30'20'10'30'20'10'
.81
lakuan BAP 1,0 nrtg/1 dan menurun kembaliDari Tabel III.73 dan hasil analisis sta-pada perlakuan BAP 2,0 mg/1 (Tabel III.72). tistik terlihat bahwa tidak ada interaksi an-
PVS1 = 22% gliserol + 13% propilen gtikol + 13% etilen glikol dalam media MS dengan taraf sukrosa 3%, PVS2 =30% gliserol + 15% etilen glikol + 15% DMSO dalam media MS dengan taraf sukrosa 0,4 M, PVS3 = media MS +50% gliserol + 50% sukrosa, Loading = media MS + gliserol 2 M + sukrosa 0,4 M
Tabel 111.71. Pengaruh perlakuan prakultur pada berbagai taraf sukrosa dan durasi prakultur
Media prakulturDurasi prakultur (hari) Persentase hidup Persentase bertunas Persentase berkalus
000000000000
Laporan Tahunan Balitbiogen 2002
Tabel 111.70. Pengaruh berbagai macam krioprotektan terhadap pertumbuhan kultur ubi kayu umur 2 minggu
Jenis larutanDurasi Durasi loading Persentase Persentase Persentase Tinggi kultur Jumlahkrioprotektan prakultur(menit)hidupbertunas berkalus(mm)daun
PVS11 hari
2hari
4 hari
PVS21 hari
2 hari
4 hari
PVS31 hari
2 hari
4 hari

82
ngan perlakuan. Selain itu, juga diketahuitidak ada perlakuan yang berbeda.
Jumlah daun tanaman yang diberi perlakuan kinetin lebih banyak dibandingkandengan tanaman kontrol. Hal ini disebab-
kan karena tanaman yang diperlakukan dengan kinetin memiliki tunas lebih banyakdibandingkan dengan tanaman kontrol(Tabel 111.73). Jumlah tunas pada tanamanyang diperlakukan dengan kinetin cenderung bertambah sesuai dengan peningkat-an konsentrasi kinetin sampai taraf tertentudan kemudian menjadi stabil (Tabel III.74).
tara nomor dan perlakuan. Kedua nomor
kentang hitam yang diuji memberikanrespon yang sama setiap perlakuan.
Rata-rata jumlah daun pada perlakuan
BAP lebih banyak daripada kontrol, karenajumlah tunas lebih banyak (Tabel 111.73).Pertambahan jumlah buku juga lebih besarpada kontrol daripada tanaman yang diper-lakukan dengan BAP. Hal ini terjadi sehu-bungan dengan kecenderungan tanaman
kontrol untuk tumbuh ke atas, sehinggalebih tinggi dan memiliki jumlah buku yanglebih banyak (Tabel 111.73).
Pada percobaan perlakuan kinetin, ter
jadi pola pertumbuhan yang sama denganpercobaan pemberian BAP. Tanaman kontrol cenderung tumbuh ke atas, sedangkan
tanaman yang diperlakukan dengan kinetincenderung membentuk tunas. Dari hasil
analisis statistik diketahui bahwa tidak adainteraksi antara nomor kentang hitam de
1,2+1,84,4+1,12,8+1,53,0+1,6
1,2+1,13,0+1,03,4+1,12,4+1,1
12,0+6,219,0+2,516,4+5,412,4+3,2
11,4+3,016,2+5,415,0+8,29,6+5,9
KH4100,00,51,02,0
KH4110,00,51,02,0
Jumlah Daun Jumlah TunasKonsentrasi BAP (mg/l)
Pengaruh beberapa taraf konsentrasi BAP terhadapjumlah daun dan banyaknya tunas dua nomor kentanghitam
0,20,46,41,78,24,83,63,9
Jumlah tunas
6,21,32,2+0,82,00,72,21,6
Jumlah daun
Tabel III.73.
BAP 0,0 mg/lBAP 0,5 mg/lBAP 1,0 mg/lBAP 2,0 mg/l
Perlakuan
Tabel 111.72. Pengaruh beberapa taraf BAP terhadap beberapa karaktervegetatif gembili
Hasil Penelitian 2002

. 83
vitro pada tanaman padi (toleran terhadaplahan masam), kedelai (toleran terhadaplahan kering), abaka (ketahanan terhadappenyakit layu), dan penyelamatan embriokacang hijau (ketahanan terhadap penyakitkudis).
Regenerasi Massa Sel Embrionik Kedelaiyang Diseleksi dengan PEG
Sebelum seleksi in vitro, dilakukan pe-nanaman embrio zigotik muda pada mediaM4C untuk menghasilkan kalus embrioge-nik dan struktur embrio somatik. Dari 395embrio zigotik varietas Wilis, Slamet, danSindoro yang dikulturkan tidak semua dapat diinduksi membentuk kalus karenakontaminasi oleh patogen internal. Seba-
nyak 395 embrio zigotik Wilis, 174 Slamet,dan 138 Sindoro yang dapat membentukkalus embriogenik masing-masing hanya37, 55, dan 31 (Tabel 111.75). Banyaknya embrio somatik yang dapat terbentuk berkisarantara 128-156. Rata-rata embrio somatik
per eksplan paling banyak berasal dari varietas Sindoro (4,13) diikuti Wilis (3,54) danSlamet (2,84).
Embrio somatik yang muncul dari kalus embriogenik berbentuk globular, hati,dan seperti terompet. Tahap selanjutnya,
PENGEMBANGAN METODE SELEKSIIN VITRO DAN PENYELAMATAN EMBRIO
UNTUK MENINGKATKAN TOLERANSITANAMAN TERHADAP FAKTOR BIOTIK
DAN ABIOTIKIka Mariska, Sri Hutami, Deden Sukmadjaja,
Mesak Tombe, Endang G. Lestari,Ragapadmi Purnamaningsih, Ali Husni,
Mia Kosmiatin, Yadi Rusyadi, Widiati H. Adil
Keragaman genetik yang tinggi meru-pakan salah satu faktor utama dalam per-baikan sifat tanaman secara konvensional.
Dengan berkembangnya teknologi kultur invitro, keragaman genetik dapat ditingkat-kan antara lain melalui keragaman soma-klonal. Salah satu metode keragaman
somakional yang lebih efektif dan efisienadalah seleksi in vitro. Melalui metode ter-sebut penyaringan sifat genetik lebih ter-arah kepada perubahan yang diinginkan.Dengan kultur in vitro berbagai sel variandihasilkan dan diseleksi dengan komponenseleksi tertentu. Frekuensi diperolehnya so-
maklon yang diinginkan dapat meningkatkarena intensitas seleksi yang efektif danhomogen terhadap massa sel dapat diberi-kan. Pada penelitian ini dilakukan pening-katan keragaman genetik melalui seleksi in
2,2+0,82,2+0,82,0+0,01,4+1,31,2+1,3
2,2+1,62,2+0,42,3+1,31,8+1,11,4+1,3
13,6+5,615,8+3,817,8+3,012,2+9,212,8+4,4
15,6+8,215,0+5,012,8+4,615,2+6,910,4+2,1
Tabel 111.74. Pengaruh beberapa taraf konsentrasi kinetin terhadapbeberapa karakter vegetatif dua nomor kentang hitam
Konsentrasi kinetin (mg/l) Jumiah Daun Jumlah Tunas
KH4100,00.51,02,04,0
KH4110,00,51,02,04,0
Laporan Tahunan Balitbiogen 2002
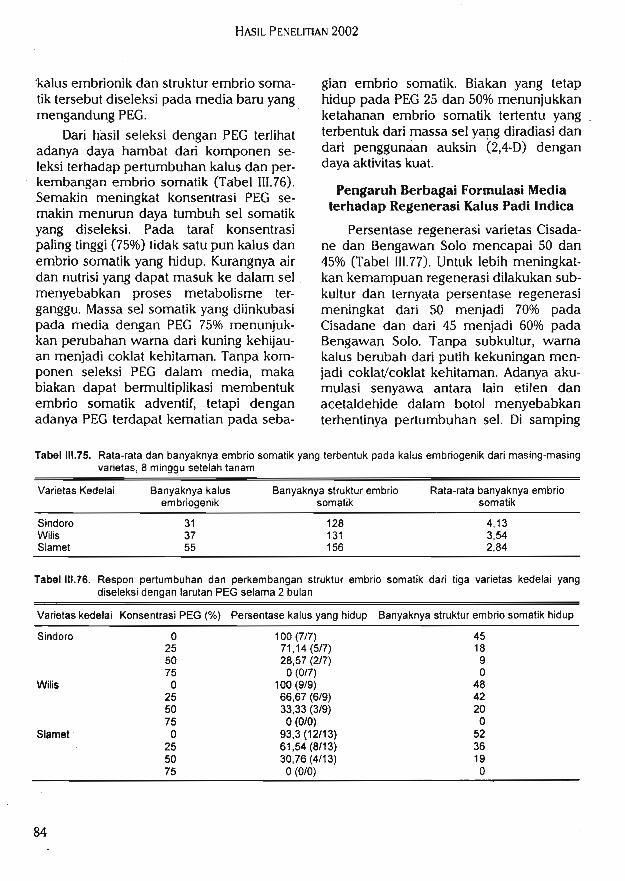
84
451890
484220
052361900 (0/0)
30,76(4/13)61,54(8/13)93,3(12/13)0 (0/0)
33,33 (3/9)66,67 (6/9)
100 (9/9)0 (0/7)
28,57 (2/7)71,14(5/7)
100(7/7)
755025
0755025
0755025
0
Slamet
Wilis
Sindoro
Tabel 111.76. Respon pertumbuhan dan perkembangan struktur embrio somatik dari tiga varietas kedelai yangdiseleksi dengan larutan PEG selama 2 bulan
Varietas kedelai Konsentrasi PEG (%) Persentase kalus yang hidup Banyaknya struktur embrio somatik hidup
4,133,542,84
128131156
313755
SindoroWillsSlamet
Rata-rata banyaknya embriosomatik
Banyaknya struktur embriosomatik
Banyaknya kalusembriogenik
Varietas Kedelai
Tabel 111.75. Rata-rata dan banyaknya embrio somatik yang terbentuk pada kalus embriogenik dari masing-masingvarietas, 8 minggu setelah tanam
gian embrio somatik. Biakan yang tetaphidup pada PEG 25 dan 50% menunjukkanketahanan embrio somatik tertentu yangterbentuk dari massa sel yang diradiasi dandari penggunaan auksin (2,4-D) dengandaya aktivitas kuat.
Pengaruh Berbagai Formulasi Mediaterhadap Regenerasi Kalus Padi Indica
Persentase regenerasi varietas Cisada-
ne dan Bengawan Solo mencapai 50 dan45% (Tabel 111.77). Untuk lebih meningkat-kan kemampuan regenerasi dilakukan sub-kultur dan ternyata persentase regenerasi
meningkat dari 50 menjadi 70% padaCisadane dan dari 45 menjadi 60% padaBengawan Solo. Tanpa subkultur, warna
kalus berubah dari putih kekuningan menjadi coklat/coklat kehitaman. Adanya aku-mulasi senyawa antara lain etilen dan
acetaldehide dalam botol menyebabkanterhentinya pertumbuhan sel. Di samping
kalus embrionik dan struktur embrio soma-
tik tersebut diseleksi pada media baru yangmengandung PEG.
Dari hasil seleksi dengan PEG terlihatadanya daya hambat dari komponen seleksi terhadap pertumbuhan kalus dan per-kembangan embrio somatik (Tabel HI.76).Semakin meningkat konsentrasi PEG se-makin menurun daya tumbuh sel somatikyang diseleksi. Pada taraf konsentrasipaling tinggi (75%) tidak satu pun kalus danembrio somatik yang hidup. Kurangnya airdan nutrisi yang dapat masuk ke dalam selmenyebabkan proses metabolisme ter-
ganggu. Massa sel somatik yang diinkubasipada media dengan PEG 75% menunjuk-kan perubahan warna dari kuning kehijau-
an menjadi coklat kehitaman. Tanpa komponen seleksi PEG dalam media, makabiakan dapat bermultiplikasi membentukembrio somatik adventif, tetapi denganadanya PEG terdapat kematian pada seba-
Hasil Penelitian 2002

85
GM1 = MS + BA 3 mg/l + thidiazuron 0,2 mg/l; GM2 = MS + BA 3 mg/| +thidiazuron 0,5 mg/l; GM3 = MS + BA 3 mg/l + IAA 0,1 mg/l + zeatin 0,2 mg/l;GM4 = MS + BA 3 mg/l + IAA 0,3 mg/l + zeatin 0,1 mg/l
0302010
00
2020
25302050
GM1GM2GM3GM4
bertunasdengan spot hijau berwarna hitamPersentase kalus Persentase kalus Persentase kalus
Formulasi media
1 = MS + BA 5 mg/l + IAA 0,1 mg/l + zeatin 0,1 mg/l; 2 = MS + BA 3 mg/l + IAA 0,1 mg/l +zeatin 0,1 mg/l
Tabel 111.78. Pengaruh formulasi media terhadap regenerasi kalus membentuktunas pada varietas Gajah Mungkur
7060
5045
Formulasi media Persentase Persentase regenerasi subkultur(mg/1)regenerasipada media yang samaVarietas
CisadaneBengawan Solo
Tabel 111.77. Pengaruh formulasi media dan subkultur terhadap kemampuan regenerasikalus
generasi membentuk tunas. Kemampuan
regenerasi Gajah Mungkur pada formulasimedia yang dicoba masih rendah (10-30%),sehingga pada percobaan selanjutnya dicoba formulasi media lain.
Dengan formulasi media bam, yaituMS + BA (1 dan 5 mg/l) + IAA 0,1 mg/1 +prolin 100 mg/1 persentase keberhasilanpembentukan spot/nodul hijau dapatmeningkat (60 dan 80%).
Kemampuan regenerasi kalus Jati-luhur pada empat formulasi media cukuptinggi antara 60,50-71,20% (Tabel 111.79).Pada media MS + BA 3 mg/1 + thidiazuron0,2 mg/1 dan MS + BA 3 mg/1 + IAA 0,1 mg/1+ zeatin 0,2 mg/1 keberhasilan membentuktunas mencapai 71,20 dan 70,10%. Padaberbagai tanaman, kombinasi 2 zat penga-tur tumbuh yang mempunyai peran fisio-logi yang sama dapat memberikan hasilyang lebih baik dibandingkan dengan kombinasi 2 jenis zat pengatur tumbuh yangberbeda.
itu, kalus yang hidup akan terus melakukanpembelahan tanpa proses diferensiasi.Auksin 2,4-D yang digunakan pada tahapawal tampaknya masih tetap berperan me-macu pertumbuhan kalus walaupun biak-
an sudah disubkultur pada media regene-rasi. Dengan demikian, sel akan terus melakukan proses mitosis tanpa melakukanperubahan dalam spesialisasi dan fungsi-nya menjadi jaringan maupun organ.
Pada proses diferensiasi adanya rege-nerasi ditandai dengan timbulnya spot/nodul hijau dari permukaan kalus. Namunspot hijau tersebut tidak semuanya dapatmembentuk tunas adventif seperti varietas
Gajah Mungkur. Keberhasilan pertumbuhan spot hijau paling tinggi (50%) berasaldari media MS + BA 3 mg/1 + IAA 0,3 mg/1+ zeatin 0,1 mg/1, tetapi persentase kalusyang bertunas hanya 10% (Tabel 111.78).Berbeda dengan kalus yang berasal darimedia MS + BA 3 mg/1 + thidiazuron 0,5mg/1 yang dapat membentuk spot hijaumencapai 30% dan semuanya dapat bere-
Laporan Tahunan Balitbiogen 2002
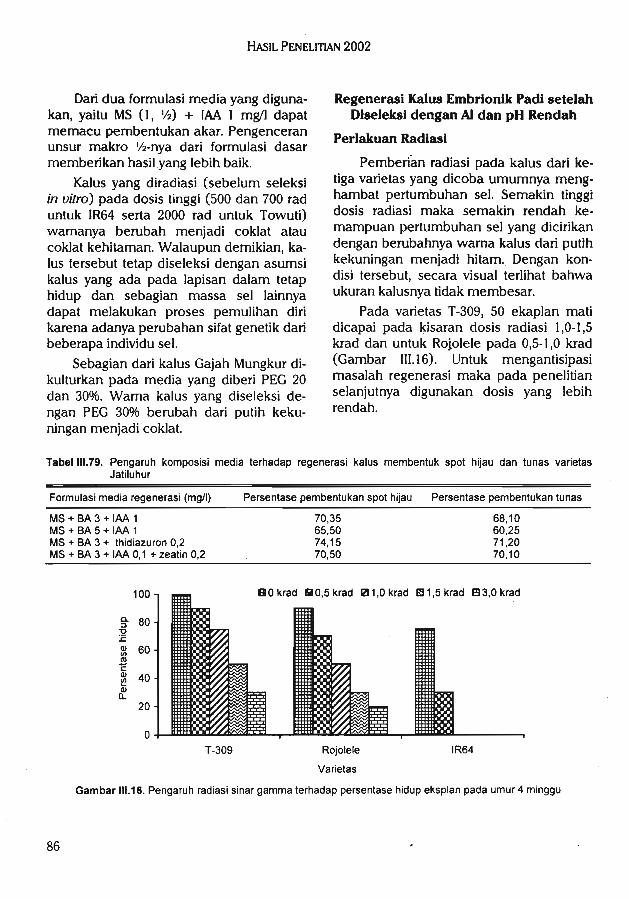
86
Rojolele
Varietas
Gambar 111.16. Pengaruh radiasi sinar gamma terhadap persentase hidup eksplan pada umur 4 minggu
IR64T-309
0 krad Q 0,5 krad 01,0 krad 01,5 krad 03,0 krad100
• 80
1 6CH| 40-a>a.
20-
0
68,1060,2571,2070,10
70,3565,5074,1570,50
MS + BA 3 + IAA 1MS + BA 5 + IAA 1MS + BA 3 + thidiazuron 0,2MS + BA 3 + IAA 0,1 + zeatin 0,2
Persentase pembentukan spot hijau Persentase pembentukan tunasFormulasi media regenerasi (mg/l)
Tabel 111.79. Pengaruh komposisi media terhadap regenerasi kalus membentuk spot hijau dan tunas varietasJatiluhur
Regenerasi Kalus Embrionik Padi setelahDiseleksi dengan Al dan pH Rendah
Perlakuan Radiasi
Pemberian radiasi pada kalus dari ke-tiga varietas yang dicoba umumnya meng-hambat pertumbuhan sel. Semakin tinggidosis radiasi maka semakin rendah ke-mampuan pertumbuhan sel yang dicirikandengan berubahnya warna kalus dari putihkekuningan menjadi hitam. Dengan kon-
disi tersebut, secara visual terlihat bahwaukuran kalusnya tidak membesar.
Pada varietas T-309, 50 ekaplan matidicapai pada kisaran dosis radiasi 1,0-1,5krad dan untuk Rojolele pada 0,5-1,0 krad(Gambar 111.16). Untuk mengantisipasimasalah regenerasi maka pada penelitianselanjutnya digunakan dosis yang lebihrendah.
Dari dua formulasi media yang diguna-kan, yaitu MS (1, V2) + IAA 1 mg/1 dapatmemacu pembentukan akar. Pengenceranunsur makro !/2-nya dari formulasi dasar
memberikan hasil yang lebih baik.Kalus yang diradiasi (sebelum seleksi
in vitro) pada dosis tinggi (500 dan 700 raduntuk 1R64 serta 2000 rad untuk Towuti)wamanya berubah menjadi coklat ataucoklat kehitaman. Walaupun demikian, kalus tersebut tetap diseleksi dengan asumsikalus yang ada pada lapisan dalam tetaphidup dan sebagian massa sel lainnyadapat melakukan proses pemulihan dirikarena adanya perubahan sifat genetik daribeberapa individu sel.
Sebagian dari kalus Gajah Mungkur di-kulturkan pada media yang diberi PEG 20dan 30%. Warna kalus yang diseleksi dengan PEG 30% berubah dari putih keku-ningan menjadi coklat.
Hasil Penelitian 2002
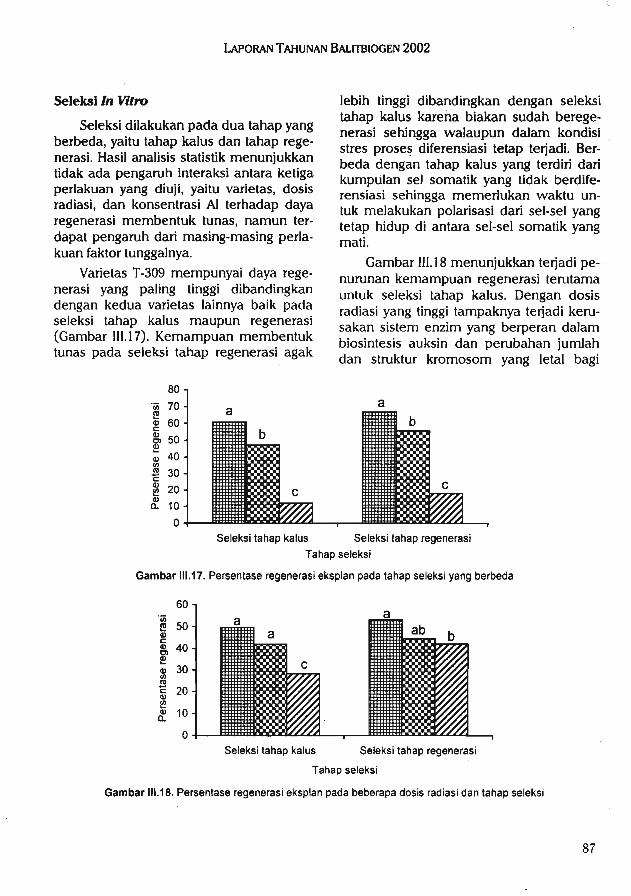
Seleksi tahap kalusSeleksi tahap regenerasi
Tahap seleksi
Gambar 111.18. Persentase regenerasi eksplan pada beberapa dosis radiasi dan tahap seleksi
Seleksi tahap kalusSeleksi tahap regenerasi
Tahap seleksi
Gambar 111.17. Persentase regenerasi eksplan pada tahap seleksi yang berbeda
60
2 50-<u| 4"u 30-(ATO1 20-22. 10"
0
10-
20-30-
40-
50-
60-
70-
2rseisiS)a>c25)
lebih tinggi dibandingkan dengan seleksitahap kalus karena biakan sudah berege-nerasi sehingga walaupun dalam kondisistres proses diferensiasi tetap terjadi. Berbeda dengan tahap kalus yang terdiri darikumpulan sel somatik yang tidak berdife-rensiasi sehingga memerlukan waktu un-
tuk melakukan polarisasi dari sel-sel yangtetap hidup di antara sel-sel somatik yangmati.
Gambar III. 18 menunjukkan terjadi pe-nurunan kemampuan regenerasi terutama
untuk seleksi tahap kalus. Dengan dosisradiasi yang tinggi tampaknya terjadi keru-sakan sistem enzim yang berperan dalam
biosintesis auksin dan perubahan jumlahdan struktur kromosom yang letal bagi
80
Seleksi In Vitro
Seleksi dilakukan pada dua tahap yangberbeda, yaitu tahap kalus dan tahap rege-nerasi. Hasil analisis statistik menunjukkantidak ada pengaruh interaksi antara ketigaperlakuan yang diuji, yaitu varietas, dosisradiasi, dan konsentrasi Al terhadap dayaregenerasi membentuk tunas, namun ter-
dapat pengaruh dari masing-masing perlakuan faktor tunggalnya.
Varietas T-309 mempunyai daya regenerasi yang paling tinggi dibandingkandengan kedua varietas lainnya baik padaseleksi tahap kalus maupun regenerasi(Gambar III.17). Kemampuan membentuktunas pada seleksi tahap regenerasi agak
Laporan Tahunan Balitbiogen 2002

88
Angka-angka dalam satu lajur yang diikuti huruf sama tidak berbeda pada taraf
nyata 5%
18,00"32,00"34,0052,00*58,00"*64,00**70,00"
Tahap regenerasi
22,0034,00*32,00*40,00*48,00*"44,00"*60,00"
Tahap kalus
Tahap seleksi
Al 500 + pH 4,0Al 400 + pH 4,0Al 300 + pH 4,0Al 200 + pH 4,0AI100 + pH4,0Kontrol pH 4,0Kontrol pH 5,8
KomDonen sfileksi
Banyaknya tunas dan jumlah akar serta pe-
nampilan dari masing-masing individu yangdiperbanyak bervariasi.
Hasil pengujian dengan konidia padamasing-masing populasi yang diperlakukandengan radiasi (0, 1 krad) dan komponenseleksi (asam fusarat dan filtrat F.oxysporum) disajikan pada Tabel 111.81.
Tanpa radiasi tampaknya hanya ta-naman yang berasal dari seleksi awal ber-
tingkat dengan konsentrasi asam fusarat 15sampai 30 ppm dan filtrat 50% yang tetaphidup (25%) setelah diuji dengan konidiakerapatan 103, 10", maupun 10s. Berbeda
dengan perlakuan kombinasi radiasi padaseleksi in vitro lebih bariyak tanaman yanghidup. Paling banyak (75%) berasal dariradiasi 1 krad dengan seleksi in vitro asamfusarat 30-45 ppm dan filtrat 50%. Tanpadiseleksi dengan filtrat (0%) tetapi langsungdengan asam fusarat konsentrasi tinggi wa-
laupun diradiasi 1 krad, persentase hidup-nya paling rendah, yaitu 30%. Keadaan ter-sebut menunjukkan bahwa kombinasi perlakuan antara pemberian mutagen dengan
seleksi in vitro memberikan hasil yanglebih tinggi.
Tabel 111.80. Persentase regenerasi eksplan pada beberapa konsentrasi Al dantahap seleksi
metabolisme sel-sel somatik. Namun di
lain pihak, perubahan jumlah dan strukturkromosom yang tidak letal diharapkan da-pat memunculkan genotipe baru dengansifat yang lebih tahan terhadap Al dan pHrendah.
Penganih faktor tungggal lainnya, yaituAl dan pH rendah disajikan pada Tabel111.80. Tanpa Al dalam media serta pH 5,8kemampuan regenerasi eksplan palingtinggi dibandingkan dengan perlakuan laindan tidak berbeda nyata dengan kontrolpada pH 4 serta Al 100 ppm + pH 4. Sema-kin tinggi konsentrasi Al, yaitu 500 ppm +pH 4 maka kemampuan regenerasinyapaling rendah.
Penurunan kemampuan regenerasi
terjadi dengan semakin meningkatnya do-sis Al pada seleksi tahap kalus maupunregenerasi.
Pengujian Planlet Abaka Hasil Seleksiterhadap Fusarium oxysporum
Tunas in vitro hasil seleksi silangantara asam fusarat dengan filtrat Fusarium
oxysporum sebelum diaklimatisasi diper-banyak terlebih dahulu pada media MS +BA 5 mg/1 + thidiazuron 0,4 mg/1 + asamsitrat 100 mg/1. Untuk perakaran digunakanMS yang diberi auksin (IBA atau NAA).
Hasil Penelitian 2002
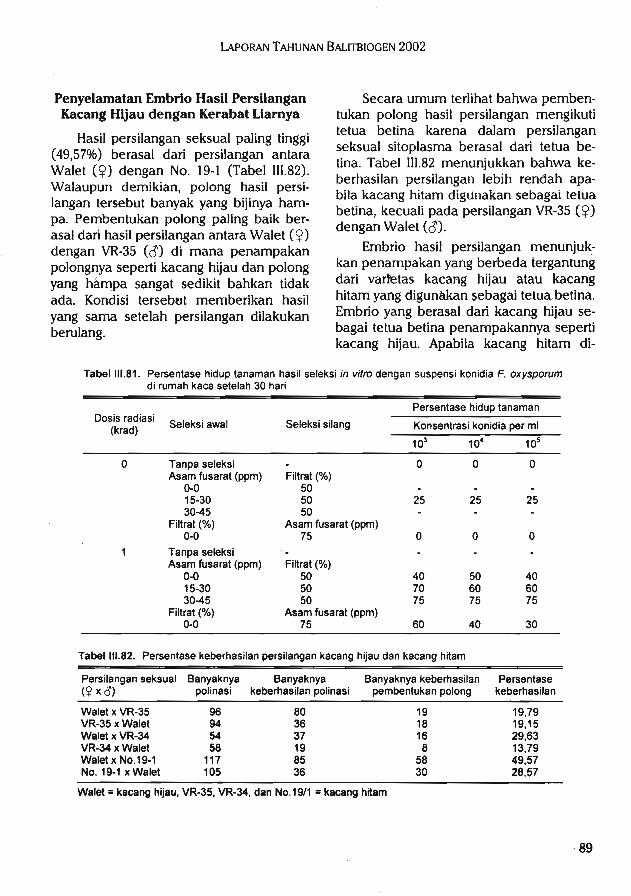
89
Walet = kacang hijau, VR-35, VR-34, dan No. 19/1 = kacang hitam
28,5749,5713,7929,6319,1519,79
keberhasilanPersentase
30
756040
.
0
.25
0
105
polong
3058
8161819
368519373680
105117
58549496
polinasi keberhasilan polinasi pembentukanBanyaknya Banyaknya keberhasilan
hitam
40
756050
.
0
.25.
0
104
No. 19-1 x WaletWalet x No. 19-1VR-34 x WaletWalet x VR-34VR-35 x WaletWalet x VR-35
(?X(J)Persilangan seksual Banyaknya
Tabel 111.82. Persentase keberhasilan persilangan kacang hijau dan kacang
60
757040
.
0
-25.-
0
103
Konsentrasi konidia per ml
Persentase hidup tanaman
75Asam fusarat (ppm)
505050
Filtrat (%)
75Asam fusarat (ppm)
505050
Filtrat (%)
Seleksi silang
0-0Filtrat (%)
30-4515-300-0
Asam fusarat (ppm)Tanpa seleksi
0-0Filtrat (%)
30-4515-300-0
Asam fusarat (ppm)Tanpa seleksi
Seleksi awal
1
0
Dosis radiasi(krad)
Tabel 111.81. Persentase hidup tanaman hasil seleksi in vitro dengan suspensi konidia F. oxysporumdi rumah kaca setelah 30 hari
Secara umum terlihat bahwa pemben
tukan polong hasil persilangan mengikutitetua betina karena dalam persilanganseksual sitoplasma berasal dari tetua betina. Tabel 111.82 menunjukkan bahwa ke-berhasilan persilangan lebih rendah apa-bila kacang hitam digunakan sebagai tetuabetina, kecuaii pada persilangan VR-35 (?)dengan Walet (61).
Embrio hasil persilangan menunjukkan penampakan yang berbeda tergantungdari vartetas kacang hijau atau kacanghitam yang digun&kan sebagai tetua betina.Embrio yang berasal dari kacang hijau sebagai tetua betina penampakannya sepertikacang hijau. Apabila kacang hitam di-
Penyelamatan Embrio Hasil PersilanganKacang Hijau dengan Kerabat Liarnya
Hasil persilangan seksual paling tinggi(49,57%) berasal dari persilangan antaraWalet (?) dengan No. 19-1 (Tabel 111.82).Walaupun demikian, polong hasil persilangan tersebut banyak yang bijinya ham-pa. Pembentukan polong paling baik berasal dari hasil persilangan antara Walet ($)dengan VR-35 (c?) di mana penampakanpolongnya seperti kacang hijau dan polongyang hampa sangat sedikit bahkan tidakada. Kondisi tersebut memberikan hasilyang sama setelah persilangan dilakukanberulang.
Laporan Tahunan Balitbiogen 2002
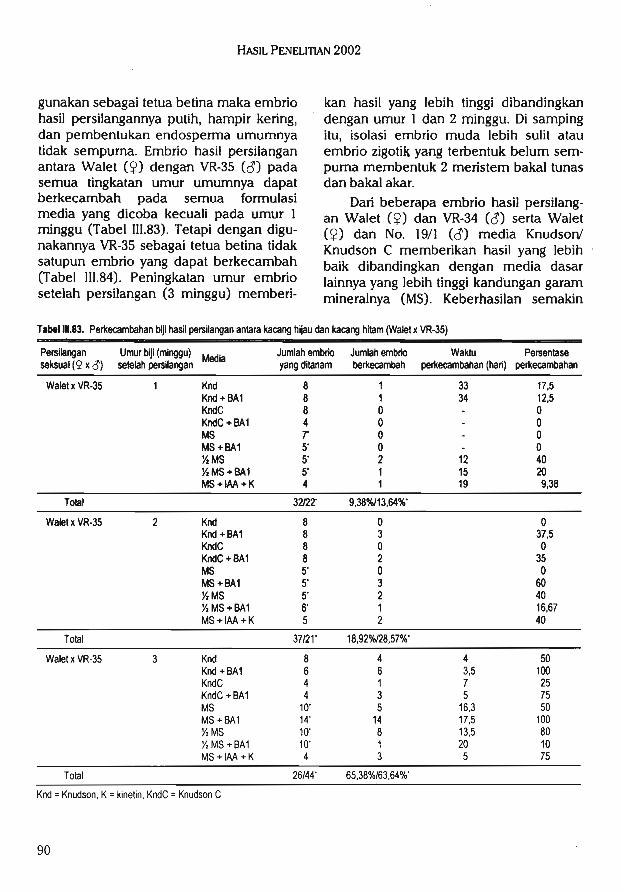
90
Knd = Knudson, K = kinetin, KndC = Knudson C
751080
100507525
10050
4016,674060
035
037,5
0
9,382040000012,517,5
perkecambahanPersentase
52013,517,516,3573,54
191512
3433
perkecambahan (hari)Waktu
65,38%/63,64%-
318
1453164
18,92%/28,57%-
212302030
9,38%/13,64%-
112000011
berkecambahJumiah embrio
26/44"
410-10'14-10-4468
37/2V5615'5-518888
32/221
4515-5'r4888
yang ditanamJumiah embrio
MS+IAA+K'/iMS+BAI^MSMS + BA1MSKndC + BA1KndCKnd+BA1
3 Knd
MS + IAA+KJ4MS + BA1ViMSMS + BA1MSKndC+BA1KndCKnd+BA1
2 Knd
MS + 1AA+K'^MS + BA1ViMSMS + BA1MSKndC+BA1KndCKnd + BA1
1 Knd
setelah persilangan MeaiaUmur biji (minggu) ^..
Total
Walet x VR-35
Total
Walet x VR-35
Total
Walet x VR-35
seksual (? x <J)Persilangan
Tabel 111.83. Perkecambahan biji hasil persilangan antara kacang hijau dan kacang hitam (Walet x VR-35)
kan hasil yang lebih tinggi dibandingkandengan umur 1 dan 2 minggu. Di sampingitu, isolasi embrio muda lebih sulit atauembrio zigotik yang terbentuk belum sempurna membentuk 2 meristem bakal tunasdan bakal akar.
Dari beberapa embrio hasil persilangan Walet (^) dan VR-34 (<$) serta Walet(?) dan No. 19/1 (<^) media Knudson/Knudson C memberikan hasil yang lebihbaik dibandingkan dengan media dasarlainnya yang lebih tinggi kandungan garammineralnya (MS). Keberhasilan semakin
gunakan sebagai tetua betina maka embriohasil persilangannya putih, hampir kering,dan pembentukan endosperma umumnya
tidak sempurna. Embrio hasil persilanganantara Walet (?) dengan VR-35 (c^) padasemua tingkatan umur umumnya dapatberkecambah pada semua formulasi
media yang dicoba kecuali pada umur 1minggu (Tabel 111.83). Tetapi dengan digu-nakannya VR-35 sebagai tetua betina tidaksatupun embrio yang dapat berkecambah(Tabel 111.84). Peningkatan umur embriosetelah persilangan (3 minggu) memberi-
Hasil Penelitian 2002
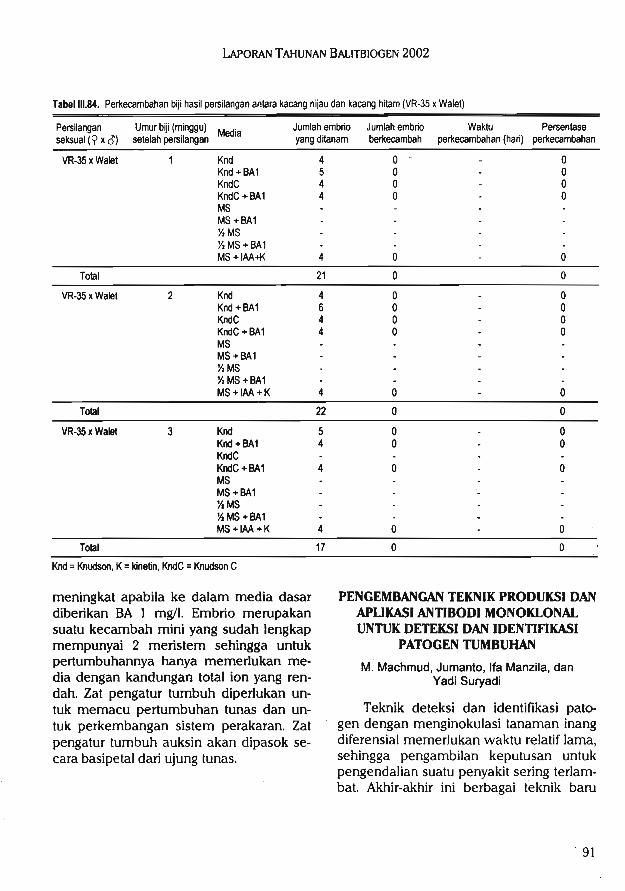
' 91
PENGEMBANGAN TEKNIK PRODUKSI DANAPLIKASIANTIBODI MONOKLONALUNTUK DETEKSI DAN IDENTIFIKASI
PATOGEN TUMBUHANM. Machmud, Jumanto, Ifa Manzila, dan
Yadi Suryadi
Teknik deteksi dan identifikasi pato-gen dengan menginokulasi tanaman inangdiferensial memerlukan waktu relatif lama,sehingga pengambilan keputusan untukpengendalian suatu penyakit sering terlam-bat. Akhir-akhir ini berbagai teknik bam
Knd = Knudson, K = kinetin, KndC = Knudson C
meningkat apabila ke dalam media dasardiberikan BA 1 mg/1. Embrio merupakansuatu kecambah mini yang sudah lengkapmempunyai 2 meristem sehingga untukpertumbuhannya hanya memerlukan me
dia dengan kandungan total ion yang ren-dah. Zat pengatur tumbuh diperlukan untuk memacu pertumbuhan tunas dan untuk perkembangan sistem perakaran. Zat
pengatur tumbuh auksin akan dipasok se-cara basipetal dari ujung tunas.
00
0
00
00
0000
00
'. !-
0000
perkecambahan (hari) perkecambahanWaktu Persentase
00
-0.00
00.
_.0000
00
-.0000 '
berkecambahJumlah embrio
174
4.45
224
4464214!-.4454
yang ditanamJumlah embrio
MS + IAA + Ky.MS + BA1^/iMSMS + BA1MSKndC + BA1KndCKnd + BA1
3 Knd
MS + IAA+Ky>MS + BA11/iMSMS+BA1MSKndC+BA1KndCKnd + BA1
2 Knd
MS + IAA+Ky,MS + BA1
MS + BA1MSKndC + BA1KndCKnd+BA1
1 Knd
setelah persilangan mealdUmurbiji(minggu) M ..
Total
VR-35 x Walet
Total
VR-35 x Walet
Total
VR-35 x Walet
seksual(?xc?)Persilangan
Tabel 111.84. Perkecambahan bijl hasll persilangan antara kacang hijau dan kacang hitam (VR-35 x Walet)
Laporan Tahunan Balitbiogen 2002

92
siap pakai, tiap paket dibungkus dengankantong plastik. Larutan bufer dibuat dengan menambahkan akuades sebelum digunakan. Pada masing-masing kemasandiberi label sehingga pengguna mudahmembedakan masing-masing komponen
perangkat ELISA. Protokol teknik ELISA untuk masing-masing patogen juga disedia-kan, sehingga para pengguna dapat mela-kukan pengujian secara langsung, berda-
sarkan petunjuk dalam protokol tersebut.
Evaluasi Keefektifan Perangkat ELISAuntuk Deteksi RRSV
Hasil pengujian menunjukkan bahwaperangkat ELISA yang telah dirakit cukupefektif untuk deteksi RRSV. Dengan peng-enceran PAb RRSV 1:10 dapat mendeteksienceran ekstrak daun terinfeksi RRSV hing-ga 10"4, suatu konsentrasi RRSV yang cukuprendah (Tabel 111.85). Keefektifan PAb RRSVyang digunakan masih cukup baik hinggaenceran 400 kali, sehingga penggunaan antibodi dapat dihemat (Tabel 111.85). Keefektifan PAb masih dapat ditingkatkan palingtidak hingga enceran 1000 kali dengan carameningkatkan titer antiserum RRSV padawaktu produksinya melalui imunisasi kelin-ci atau mencit, diikuti dengan proses padawaktu pemurnian antiserum menjadi PAb.
Deteksi RRSV dari contoh tanaman padi bergejala penyakit kerdil hampa yang di-ambil dari beberapa lokasi di Jawa Barat,menunjukkan bahwa perangkat ELISAyang telah dirakit dapat digunakan secaralangsung untuk mendeteksi RRSV dari ekstrak daun dengan menggunakan enceranPAb 1 : 200 dan 1 : 400 (Tabel 111.86). Padaumumnya, hasil pengujian lebih baik apa-bila PAb yang digunakan mempunyai kepekatan lebih tinggi (enceran 1 : 200). Per-beedaan warna reaksi pada lubang cawanELISA menunjukkan bahwa titer atau kan-dungan partikel RRSV dalam ekstrak con-
yang peka dan akurat untuk deteksi danidentifikasi patogen tanaman telah dikem-bangkan, termasuk teknik serologi dan mo-lekuler. Teknik Enzyme-linked Immunosor-
bent Assay (ELISA) merupakan salah satuteknik serologi yang efektif dan efisien.
Sejak tahun 1994, di Balitbiogen telahdikembangkan teknik produksi antibodipolikional (PAb) untuk deteksi virus tungropadi (rice tungro virus/RTV), virus kerdil ke-delai (soybean stunt virus/SSV), dan virusbilur kacang tanah (peanut stripe virus/PStV), bakteri layu (Ralstonia solanace-arum/RS), serta bakteri hawar kedelai(Pseudomonas syringae pv. glycinea/PSG),dan bisul kedelai (Xanthomonas axonopo-dis pv. glycines/XAG; sin: X. campestris pv.glycines). PAb diproduksi dalam rangkaproduksi antibodi berbagai patogen tum-buhan yang akan digunakan untuk deteksidini dan identifikasi patogen menggunakanteknik ELISA.
Evaluasi Lapang Keefektifan PerangkatELISA dengan PAb untuk Deteksi dan
Identifikasi RRSV dan RS
Perakitan Perangkat ELISA
Pada tahap ini komponen perangkatELISA yang dibuat belum siap dikemas se-cara komersial, walaupun ukuran dari tiap
komponen perangkat telah ditetapkan dandiperkirakan cukup untuk menguji sekitar250-300 contoh tanaman. Perangkat ELISAtersebut terdiri atas antigen RRSV dan RSyang dikemas dengan tabung eppendorf1,5 ml dengan volume 1,0 ml/tabung dan
,sediaan murni RRSV dengan kepekatan,100 ug/ml atau antigen RS dengan kepekatan 100 sel/ml. Konjugat untuk setiap perangkat dikemas dalam tabung eppendorf1,5 ml dengan volume 1,0 ml. Pada peng-gunaannya, konjugat perlu diencerkan 100kali dengan PBS-T. Bufer substrat dikemasdalam bentuk campuran bahan kimia yang
Hasil Penelitian 2002
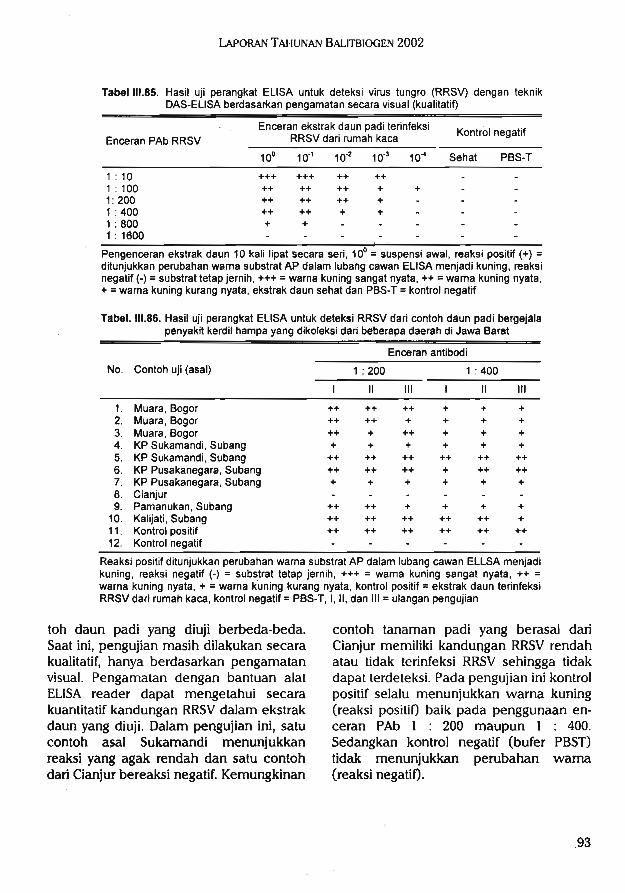
.93
contoh tanaman padi yang berasal dariCianjur memiliki kandungan RRSV rendahatau tidak terinfeksi RRSV sehingga tidakdapat terdeteksi. Pada pengujian ini kontrolpositif selalu menunjukkan warna kuning(reaksi positif) baik pada penggunaan enceran PAb 1 : 200 maupun 1 : 400.Sedangkan kontrol negatif (bufer PBST)tidak menunjukkan perubahan warna(reaksi negatif).
toh daun padi yang diuji berbeda-beda.Saat ini, pengujian masih dilakukan secarakualitatif, hanya berdasarkan pengamatanvisual. Pengamatan dengan bantuan alat
ELISA reader dapat mengetahui secarakuantitatif kandungan RRSV dalam ekstrakdaun yang diuji. Dalam pengujian ini, satucontoh asal Sukamandi menunjukkanreaksi yang agak rendah dan satu contohdari Cianjur bereaksi negatif. Kemungkinan
Reaksi positif ditunjukkan perubahan warna substrat AP dalam lubang cawan ELLSA menjadikuning, reaksi negatif (-) = substrat tetap jernih, +++ = warna kuning sangat nyata, ++ =warna kuning nyata, + = warna kuning kurang nyata, kontrol positif = ekstrak daun terinfeksiRRSV dari rumah kaca, kontrol negatif = PBS-T, I, II, dan III = ulangan pengujian
1.Muara, Bogor2.Muara, Bogor3.Muara, Bogor4.KP Sukamandi, Subang5.KP Sukamandi, Subang6.KP Pusakanegara, Subang7.KP Pusakanegara, Subang8.Cianjur9.Pamanukan, Subang
10.Kalijati, Subang11.Kontrol positif12.Kontrol negatif
II III
:400
I1
III
Enceran antibodi
daun padi bergejaladi Jawa Barat
I II
Contoh uji (asal) 1 : 200
111.86. Hasil uji perangkat ELISA untuk deteksi RRSV dari contohpenyakit kerdii hampa yang dikoleksi dari beberapa daerah
No.
Tabel.
Pengenceran ekstrak daun 10 kali lipat secara seri, 10 = suspensi awal, reaksi positif (+) =ditunjukkan perubahan warna substrat AP dalam lubang cawan ELISA menjadi kuning, reaksinegatif (-) = substrat tetap jernih, +++ = warna kuning sangat nyata, ++ = warna kuning nyata,+ = warna kuning kurang nyata, ekstrak daun sehat dan PBS-T = kontrol negatif
1 :101 : 1001:2001 :4001 :8001 : 1600
Sehat PBS-T10-10"210'10
Kontrol negatifEnceran ekstrak daun padi terinfeksiRRSV dari rumah kacaEnceran PAb RRSV
Tabel 111.85. Hasil uji perangkat ELISA untuk deteksi virus tungro (RRSV) dengan teknikDAS-ELISA berdasarkan pengamatan secara visual (kualitatif)
Laporan Tahunan Balitbiogen 2002
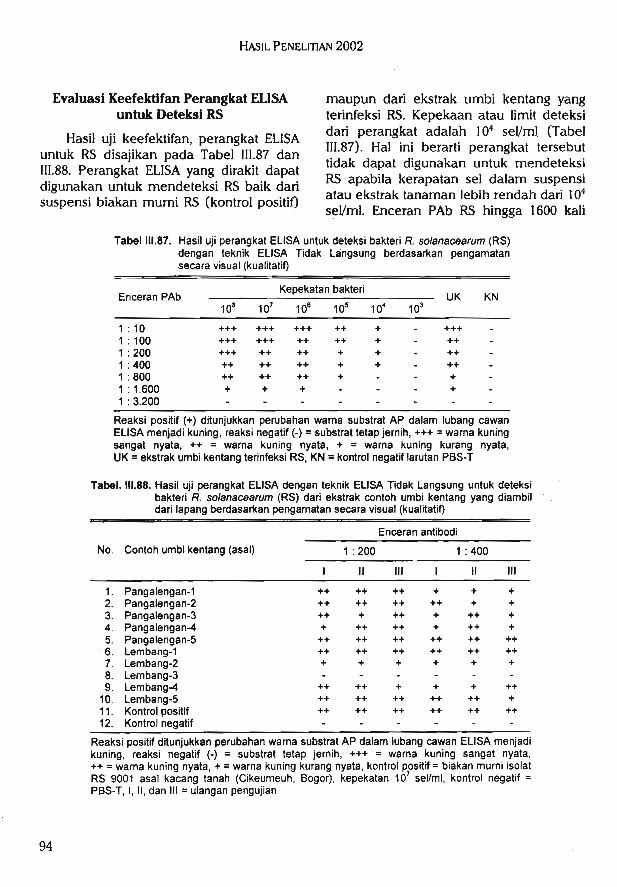
94
Reaksi positif ditunjukkan perubahan warna substrat AP dalam lubang cawan ELISA menjadikuning, reaksi negatif (-) = substrat tetap jernih, +++ = warna kuning sangat nyata,++ = warna kuning nyata, + = warna kuning kurang nyata, kontrol positif = biakan murni isolatRS 9001 asal kacang tanah (Cikeumeuh, Bogor), kepekatan 107 sel/ml, kontrol negatif =PBS-T, I, II, dan III = ulangan pengujian
1.Pangalengan-12.Pangalengan-23.Pangalengan-34.Pangalengan-45.Pangalengan-56.Lembang-17.Lembang-28.Lembang-39.Lembang-4
10.Lembang-511.Kontrol positif12.Kontrol negatif
No. Contoh umbi kentang (asal)1 : 2001 : 400
Reaksi positif (+) ditunjukkan perubahan warna substrat AP dalam lubang cawanELISA menjadi kuning, reaksi negatif (-) = substrat tetap jernih, +++ = warna kuningsangat nyata, ++ = warna kuning nyata, + = warna kuning kurang nyata,UK = ekstrak umbi kentang terinfeksi RS, KN = kontrol negatif larutan PBS-T
Tabel. III.88. Hasil uji perangkat ELISA dengan teknik ELISA Tidak Langsung untuk deteksibakteri R. solanacearum (RS) dari ekstrak contoh umbi kentang yang diambildari lapang berdasarkan pengamatan secara visual (kualitatif)
Enceran antibodi
10+++ +++ +++ ++ + - +++100+++ +++ ++ ++ + - ++200+++ ++ ++ + + ++400++++++ + + - ++800+++++++ - - +1.600+ + + ...+3.200
Evaluasi Keefektifan Perangkat ELISA maupun dari ekstrak umbi kentang yanguntuk Deteksi RSterinfeksi RS. Kepekaan atau limit deteksi
Hasil uji keefektifan, perangkat ELISA^?J7 Pe^kat Kadalah l0* sfml (Tabeluntuk RS disajikan pada Tabel 111.87 danmfj^al '"! berf Perankat te;sel?u!111.88. Perangkat ELISA yang dirakit dapatl'^k dapat d.gunakan untuk mendeteks,digunakan untuk mendeteksi RS baik dariRS apab.la kerapatan sel dalam suspenssuspensi biakan murni RS (kontrol positif) at^U fT tana2^" d-w^ i^^' i ,
Ksel/ml. Enceran PAb RS hmgga 1600 kali
Tabel 111.87. Hasil uji perangkat ELISA untuk deteksi bakteri R. solanacearum (RS)dengan teknik ELISA Tidak Langsung berdasarkan pengamatansecara visual (kualitatif)
^... Kepekatan bakteriEnceran PAb - UK KN
10' 107 106 105 104 103
Hasil Penelitian 2002

95
Penyediaan dan Fusi Sel Mieloma danLimposit
Sel mieloma yang dibiakkan padamedium DMEM + NBS + L glycine tumbuhbaik berupa koloni sel yang terdapat di da-sar medium. Biakan sel mieloma yang ber-
umur 72 jam menghasilkan kerapatan +.107sel/ml. Sel limposit diperoleh dalam bentukcairan/suspensi dalam medium DMEM-NBS. Setiap mencit menghasilkan 108 limposit sehingga dari 5 ekor mencit diperoleh5x 108 limposit.
Fusi sel mieloma dengan sel limpositmencit yang telah diimunisasi denganRRSV menghasilkan fusan sel hibridoma.Hal ini ditunjukkan dengan pertumbuhanbiakan sel pada dasar medium. Hasil fusihanya mencapai 29,7% dan sel hibridomayang diperoleh tidak semuanya menghasilkan McAb, sehingga perlu dilakukanskrining untuk memperoleh sel hibridomapenghasil McAb.
Skrining Sel Hibridoma
Pada skrining I dari 297 lubang cawanmikro yang berisi hibridoma diperoleh 14biakan sel hibridoma yang mampu menghasilkan McAb RRSV (Tabel 111.89). Ke-mampuan menghasilkan McAb dari setiapkoloni sel hibridoma berkisar antara 0,03-1,29. Hibridoma yang menghasilkan McAbrata-rata tertinggi adalah nomor RR-(3)-2H03, RR-(3)-l HI 1, RR-(I)-3 C07, dan RR-(2)-3 C04. Keempat hibridoma tersebut diujidengan teknik AAI-ELISA menunjukkanreaksi positif dengan angka rata-rata OD415berturut-turut 0,68; 0,64; 0,40; dan 0,55, sedangkan menggunakan teknik IDAS diperoleh angka masing-masing 1,22; 0,84; 0,88;dan 0,64.
Skrining II dilakukan untuk memperoleh biakan hibridoma yang potensialmenghasilkan McAb tinggi dan stabil untuk
(1 : 1600) masih dapat digunakan untukmendeteksi RS, tetapi yang efisien adalahenceran 800. Penggunaan PAb dengan
enceran 800 kali cukup efisien dan sangatmenghemat biaya, karena harga antibodi,terutama apabila harus diimpor sangatmahal.
Hasil pengujian perangkat EL1SA untukmendeteksi RS dari umbi kentang menun-jukkan bahwa dari 10 contoh umbi kentangyang diambil dari Lembang dan Pangaleng-an, sembilan contoh di antaranya bereaksi
positif, baik yang diuji menggunakan enceran PAb 400 kali maupun 800 kali. Satucontoh umbi asal Lembang bereaksi nega-tif. Kontrol positif yang berupa suspensibakteri PS 9001 bereaksi positif sedangkankontrol (larutan bufer PBS-T) bereaksi ne-gatif. Hal ini berarti sembilan contoh umbikentang yang diuji mengandung atau ter-infeksi RS, sedangkan satu contoh umbi(Lembang-3) kemungkinan tidak terinfeksiRS atau populasi RS dalam umbi rendah(<104sel/ml).
Produksi McAb untuk Deteksi danIdentifikasi RRSV
Imunisasi Tikus Putih
Contoh darah diambil empat hari se-telah imunisasi terakhir dan diproses untukmemperoleh PAb RRSV dan diuji titernyamenggunakan teknik mikropresipitasi.Hasilnya menunjukkan darah bereaksipositif berdasarkan uji mikropresipitasi. Halini menunjukkan adanya respon antibodidari mencit terhadap antigen RRSV yang di-suntikkan. Setelah diketahui bahwa mencittelah memproduksi PAb, pada hari yang sa-ma, mencit dimatikan untuk diambil limpa-nya.
Laporan Tahunan Balitbiogen 2002
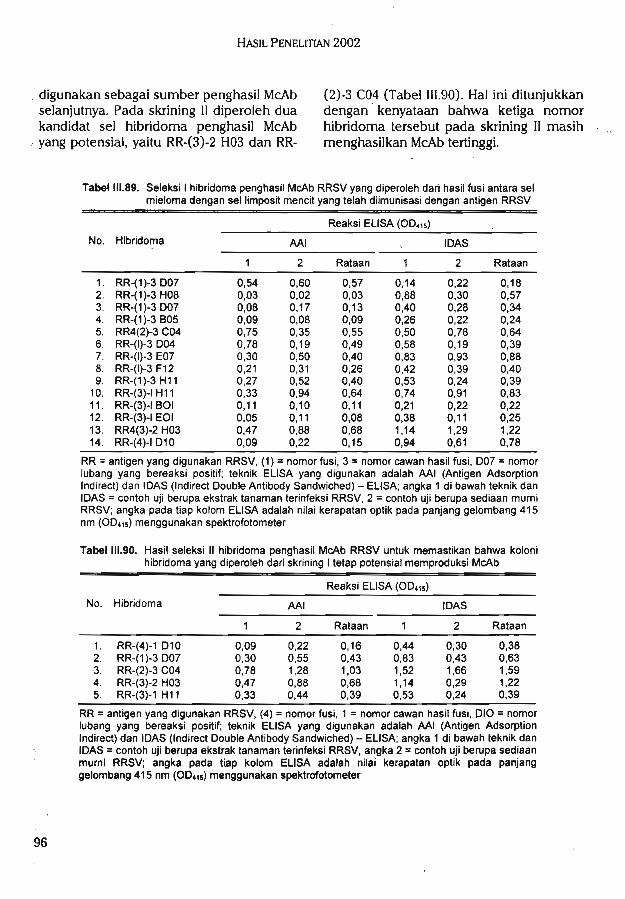
96
RR = antigen yang digunakan RRSV, (4) = nomor fusi, 1 = nomor cawan hasil fusi, DIO = nomorlubang yang bereaksi positif; teknik ELISA yang digunakan adalah AAI (Antigen AdsorptionIndirect) dan IDAS (Indirect Double Antibody Sandwiched) - ELISA; angka 1 di bawah teknik danIDAS = contoh uji berupa ekstrak tanaman terinfeksi RRSV, angka 2 = contoh uji berupa sediaanmurni RRSV; angka pada tiap kolom ELISA adalah nilai kerapatan optik pada panjanggelombang 415 nm (OD4,5) menggunakan spektrofotometer
0,391,221,590,630,38
0,240,291,660,430,30
0,531,141,520,830,44
0,390,681,030,430,16
0,440,881,280,550,22
0,330,470,780,300,09RR-(4)-1 D10
RR-(1)-3D07RR-(2)-3 C04RR-(3)-2 H03RR-(3)-1 H11
1.2.3.4.5.
RataanRataan
IDASAAINo. Hibridoma
Reaksi ELISA (OD4i5)
RR = antigen yang digunakan RRSV, (1) = nomor fusi, 3 = nomor cawan hasil fusi, D07 = nomorlubang yang bereaksi positif; teknik ELISA yang digunakan adalah AAI (Antigen AdsorptionIndirect) dan IDAS (Indirect Double Antibody Sandwiched) - ELISA; angka 1 di bawah teknik danIDAS = contoh uji berupa ekstrak tanaman terinfeksi RRSV, 2 = contoh uji berupa sediaan murniRRSV; angka pada tiap kolom ELISA adalah nilai kerapatan optik pada panjang gelombang 415nm (OD415) menggunakan spektrofotometer
Tabel 111.90. Hasil seleksi II hibridoma penghasil McAb RRSV untuk memastikan bahwa kolonihibridoma yang diperoleh dari skrining I tetap potensial memproduksi McAb
0,781,220,250,220,830,390,400,880,390,640,240,340,570,18
Rataan
0,611,290,110,220,910,240,390,930,190,780,220,280,300,22
2
IDAS
0,941,140,380,210,740,530,420,830,580,500,260,400,880,14
1
0,150,680,080,110,640,400,260,400,490,550,090,130,030,57
Rataan
Reaksi ELISA (OD15)
0,220,880,110,100,940,520,310,500,190,350,080,170,020,60
2AAI
0,090,470,050,110,330,270,210,300,780,750,090,080,030,54
1
RR-(4)-ID10RR4(3)-2 H03RR-(3)-l EOIRR-(3)-l BOIRR-(3)-IH11RR-(1)-3H11RR-(I)-3F12RR-(l)-3 E07RR-(l)-3 D04RR4(2)-3 C04RR-(1)-3B05RR-(1)-3D07RR-(1)-3H08RR-(1)-3D07
Hibridoma
14.13.12.11.10.9.8.7.6.5.4.3.2.1.
No.
Tabel 111.89. Seleksi I hibridoma penghasil McAb RRSV yang diperoleh dari hasil fusi antara selmieloma dengan sel limposit mencit yang telah diimunisasi dengan antigen RRSV
(2)-3 C04 (Tabel III.90). Hal ini ditunjukkandengan kenyataan bahwa ketiga nomorhibridoma tersebut pada skrining II masihmenghasilkan McAb tertinggi.
digunakan sebagai sumber penghasil McAbselanjutnya. Pada skrining II diperoleh duakandidat sel hibridoma penghasil McAbyang potensial, yaitu RR-(3)-2 H03 dan RR-
Hasil Penelitian 2002

97
Nomor hibridoma RS-(1)-01 C10, RS = antigen yang digunakan RS, (1) =nomor fusi, 01 = nomor cawan hasil fusi, C10 = nomor lubang yang bereaksipositif; angka pada kolom reaksi ELISA merupakan hasil pengukurankerapatan optik (OD45o) menggunakan spektrofotometer; nilai di bawahangka 1 = contoh uji berupa ekstrak umbi kentang terinfeksi RS, 2 = contohuji berupa sediaan murni RS
0,530,360,270,350,270,290,120,440,350,47
Rataan
iso)
0,610,370,320,320,280,260,130,470,520,45
2
Reaksi ELISA (OD,
0,450,350,220,370,260,320,110,410,170,48
1
RS-(6)-02 F09RS-(5)-0l E03RS-(4)-02 A09RS-(3)-0l G04RS-(3)-0l E05RS-(3)-0l B05RS-(2)-03C12RS-(1)-0IF08RS-(1)-0IF03RS-(1)-0IC10
1 IIUIIvlvIIIQHibridoma
10.9.8.7.6.5.4.3.2.1.
No.
Tabel 111.91. Nomor koloni hibridoma penghasil McAb RS yang diperolehdari skrining I hasil fusi sel mieloma dengan sel limposit mencityang diimunisasi dengan antigen RS
ditunjukkan adanya pertumbuhan biakansel pada dasar medium. Hasil fusi menca-
pai 30,5% Sel hibridoma yang diperolehtidak semuanya menghasilkan McAb, se-hingga perlu dilakukan skrining untukmemperoleh sel hibridoma penghasilMcAb.
Skrining Sel Hibridoma
Pada skrining I diperoleh 10 biakanhibridoma yang menghasilkan McAb ber-dasarkan reaksi positif dengan teknikELISA yang dibaca dengan spektrofoto-meter dengan OD415 (Tabel 111.91). Kemam-
puan menghasilkan McAb dari setiap ko-loni sel hibridoma berkisar antara 0,12-0,53.
Skrining II dilakukan untuk memperolehbiakan hibridoma yang potensial menghasilkan McAb tinggi sebagai sumber penghasil McAb selanjutnya.
Pada skrining II diperoleh tiga kandidatsel hibridoma penghasil McAb yang potensial, berdasarkan kemampuan menghasilkan McAb yang tinggi, yaitu RS-(1)-1 C10,
Produksi McAb untuk Deteksi danIdentifikasi RS
Imunisasi Tikus Putih
Contoh darah diambil seminggu sete-lah imunisasi terakhir dan dimurnikan se-rumnya. Hasil pengujian dengan teknikmikropresipitasi positif, berarti bahwamencit menunjukkan respon antibodi ter-hadap antigen RS yang disuntikkan.
Penyediaan dan Fusi Sel Mieloma danLimposit
Biakan sel mieioma yang berupa lapis-an film sel mieloma tumbuh di dasar lu-bang cawan medium. Kerapatan sel mielo
ma yang diperoleh pada saat akan dilaku-kan fusi dengan sel limposit mencit adalah107 sel/ml. Sel limposit dari lima mencit di-hasilkan dengan kerapatan 1,02 x 108sel/ml.
Fusi sel mieloma dengan sel limpositmencit yang telah diimunisasi dengan RSmenghasilkan fusan sel hibridoma. Hal ini
Laporan Tahunan Balitbiogen 2002

98
TEKNOLOGI PENGEMBANGANPUPUK HAYATI DAN PEROMBAK
BAHAN ORGANIK UNTUK LAHAN KERINGDAN LAHAN SAWAH
Rasti Saraswati, Dwi Ningsih Susilowati,Arief Indrasumunar, Misgiyarta,
R.D.M. Simanungkalit
Pengembangan teknologi pupukhayati untuk kedelai di lahan kering danlahan kering masam-Al telah memperoleh
strain bakteri bintil akar dari beberapadaerah sentra produksi kedelai di Indonesia. Untuk meningkatkan kemampuan
penambatan N2 Rhizobium telah dilakukanbeberapa upaya seperti peningkatan sis-tem pengiriman melalui teknologi mikro-enkapsulasi dan perbaikan mutu genetikmelalui transfer gen nif dan mutasi kimiadengan ethyl methyl sulfonat. Upaya perbaikan mutu genetik Rhizobium melaluirekayasa genetik seperti transformasi, kon-
jugasi, transduksi, dan transfeksi hanyaberlangsung satu arah, transfer DNA hanyaterjadi dari bakteri donor ke bakteri resi-pien, sedangkan transformasi DNA secara
timbal balik pada beberapa mikroorganis-me dapat dilakukan melalui teknologi fusi
RS-(1)-1 F08, dan RS-(6)-2 F09 dengan rataan angka OD415 masing-masing 0,57; 0,54,
dan 0,64 (Tabel IH.92). Hasil skrining me-nunjukkan bahwa hibridoma yang meng-hasilkan McAb tertinggi pada skrining I jugatetap menghasilkan McAb tertinggi padaskrining II.
Penyimpanan Hibridoma dan McAb
Biakan hibridoma nomor RS-(1)-1 C10,RS-(1)-1 F08, dan RS-(6)-2 F09 yang potensial menghasilkan McAb RS tinggi ditem-patkan dalam botol kriogenik yang disim-pan dalam tabung berisi nitrogen cair.
Koloni hibridoma ini selanjutnya diklonuntuk memperoleh koloni hibridoma satusel (single sel colony hybridoma) yangmenghasilkan McAb spesifik. McAb yangdiperoleh pada kegiatan ini disimpan dalam ampul dalam keadaan kering bekupada suhu -20C.
Nomor hibridoma RS-(1)-0l C10: RS = antigen yang digunakan RS,(1) = nomor fusi, 01 = nomor cawan hasil fusi, C10 = nomor lubang yangbereaksi positif; angka pada kolom reaksi ELISA merupakan hasil peng-ukuran kerapatan optik (OD450) menggunakan spektrofotometer; nilai dibawah angka 1 = contoli uji berupa ekstrak umbi kentang terinfeksi RS,2 = contoh uji berupa sediaan murni RS
0,640,540,570,250,330,31
Rataan
0,690,570,620,240,300,28
2 .
0,580,500,510,360,35
' 0,35
1
RS-(6)-02 F09RS-(1)-0IF08RS-(1)-0IC10RS-(5)-0l E03RS-(3)-0l G04RS-(1)-0IF03
6.5.4.3.2.1.
Tabel 111.92. Hasil skrining II enam nomor koloni hibridoma penghasil McAbR. solanacearum (RS) yang potensial tinggi dan stabil
„,_ L,!^...JReaksi ELISA (ODo)
Hasil Penelitian 2002

99
Pembentukan Protoplas
Berdasarkan perhitungan mengguna-kan haemositometer, jumlah sel awal B.japonicum -L17-lWoifarn 4,05 x 108 sel/ml;
sedangkan jumlah protoplas B. japonicumL17_iki< yang terbentuk 2,95 x 108 proto-
plas/ml, sehingga persentase pembentukan protoplas B. japonicum L17-lWoram
72,84%. Jumlah sel awal B. japonicumPdl0ABkan 4,35 x 108 sel/ml; sedangkanjumlah protoplas B. japonicum Pdl0ABkanyang terbentuk 3,05 x 108 protoplas/ml, sehingga persentase pembentukan protoplasB. japonicum Pd 10ABkan 70,11 %.
Regenerasi Protoplas
Bentuk koloni hasil regenerasi protoplas tidak berbeda dengan bentuk kolonisel awal. Waktu regenerasi protoplas B.japonicum L17-1Woram dan B. japonicum
Pdl0ABkan lebih lama, yaitu 15-18 hari di-bandingkan pertumbuhan sel awal, yaitu 5-
7 hari. Hal tersebut mungkin disebabkanprotoplas memerlukan waktu untuk men-
sintesis dinding sel dan membelah.Jumlah protoplas B. japonicum L17-
jkioram ^,95 x \^ protoplas/ml; sedangkan
jumlah protoplas B. japonicum L17-lWoram
yang tumbuh pada medium regenerasi 1,58x 106 sel/ml, sehingga diperoleh frekuensiregenerasi protoplas B. japonicum L17-lWoram 5,35 x 103. Jumlah protoplas B. japo
nicum Pdl0ABkan 3,05 x 108 protoplas/ml;sedangkan jumlah protoplas B. japonicumPdl0ABkan yang tumbuh pada mediumregenerasi 2,42 x 105 sel/ml, sehingga diperoleh frekuensi regenerasi protoplas B.japonicum Pdl0ABkan 7,93 x 10"4.
Fusi Protoplas
Pengamatan mikroskopis fusi protoplas intraspesies antara B. japonicum L17-jkioram dengan B japonicum Pdl0ABkan
protoplas. Melalui teknologi fusi protoplasdiharapkan gen-gen yang berhubungan de-ngan penambatan nitrogen (gen nod, nif,dan fix) dapat tertransfer secara intra- dan
interspesifik dan membiarkan gen tersebutberekombinasi dan bersegregasi menjadiRhizobium yang lebih unggul daripadastrain asalnya.
Peningkatan kemampuan penambatan N2 selain dapat dilakukan melalui per-baikan mutu genetika Rhizobium itu sen-diri, dapat juga dilakukan melalui peningkatan peran isoflavonoid di mana isoflavo-noid mempunyai peran yang sangat pen-
ting dalam pembentukan bintil akarkedelai.
Fusi Protoplas Intraspesies antar
Bradyrhizobium japonicum
Seleksi Mutan Resisten Antibiotik
Seleksi mutan resisten antibiotik dilakukan untuk mendapatkan penanda gene-
tik mikroba yang nantinya akan digunakanpada proses seleksi hasil fusi protoplasintraspesies. Berdasarkan hasil seleksi ter-
hadap kedua tetua, Bradyrhizobium japonicum LI 7-1 dan B. japonicum PdlOAB ter-hadap antibiotik ampisilin, kanamisin, klo-ramfenikol, dan tetrasiklin dengan kisarankonsentrasi 50-1.000 pg/ml diperoleh bah-wa mutan B. japonicum LI 7-1 resisten ter-hadap kloramfenikol (100 pg/ml) dan mutan B. japonicum PdlOAB resisten terhadapkanamisin (200 pg/ml).
Kurva Pertumbuhan B. japonicum
Berdasarkan pengamatan kurva pertumbuhan B. japonicum L17-lWoram dan
Pdl0ABkan pertengahan fase eksponensialdicapai pada jam ke-72 dengan jumlah selB. japonicum L17-lWoram sebanyak 3,29 x 107
sel/ml dan B. japonicum Pdl0ABkan sebanyak 8,2 x 107 sel/ml.
Laporan Tahunan Balitbiogen 2002

100
INTRA-12-1NTRA-14, INTRA-16-INTRA-l 8,INTRA-20-INTRA-22, INTRA-24-INTRA-30tumbuh subur pada pH 4,5. Pada pH 9 ha-nya INTRA-2-INTRA-14 yang tumbuh sangatsubur, INTRA-1 tumbuh subur, INTRA-15,INTRA-16, dan INTRA-19 tumbuh tidak subur, serta INTRA-17, INTRA-18, dan INTRA-
20-INTRA-30 tidak dapat tumbuh. B. japonicum PdlOAB"3" tumbuh sangat subur pada pH 4,5 dan 9, sedangkan B. japonicumL17-lMoram tumbuh tidak subur pada pH 4,5dan 9.
Berdasarkan hasil pengamatan, pertumbuhan pada medium YEMA dengan pH4,5 dan 9 menunjukkan bahwa INTRA-15-INTRA-30 memiliki sifat gabungan dari ke-dua strain parental, yaitu tahan hidup padapH 4,5 tetapi tidak tahan pada pH 9, sedangkan INTRA-1-INTRA-14 menunjukkanketahanan hidup yang sama dengan B.japonicum PdlOAB1""1.
Berdasarkan pola RIA bakteri hasil fusiterbagi dalam enam kelompok. Kelompok1 terdiri atas INTRA-2, INTRA-5, INTRA-26.Kelompok 2 terdiri atas INTRA-3, INTRA-12,INTRA-20. Kelompok 3 terdiri atas INTRA-1,INTRA-6, INTRA-13, INTRA-19, INTRA-25,INTRA-29. Kelompok 4 terdiri atas INTRA-4,INTRA-8, INTRA-9, INTRA-11, INTRA-16,INTRA-18, INTRA-23, INTRA-27, INTRA-28.Kelompok 5 terdiri atas INTRA-7, INTRA-10,INTRA-14, INTRA-17, INTRA-21, INTRA-22.Kelompok 6 terdiri atas INTRA-15, INTRA-24, INTRA-30. Hasil karakterisasi menggunakan pola RIA menunjukkan bahwa bakteri hasil fusi memiliki karakter yang ber-beda dengan parentalnya.
Pengujian Kemampuan Penambatan N2
Berdasarkan hasil pengamatan tam-pak bahwa bakteri hasil fusi mampu mem-bentuk bintil akar pada tanaman kedelai.Selain itu, rata-rata bobot kering tajuk dari
menggunakan mikroskop fase kontras(pada perbesaran 600x) dilakukan padacampuran protoplas yang ditambah PEGsetelah inkubasi 30 menit. Hasil pengamat-an menunjukkan terjadi fusi antara duaprotoplas, tiga sampai dengan 10 protoplas,dan lebih dari 10 protoplas.
Regenerasi Protoplas Hasil Fusi
Bentuk koloni regenerasi protoplas hasil fusi pada medium nonselektif tidak ber-beda dengan bentuk koloni sel parental.Waktu regenerasi protoplas hasil fusi lebihlama (15-8) had dibandingkan pertumbuh-an sel parental (5-7) hari.
Jumlah sel yang tumbuh pada mediumnonselektif 1,23 x 103 sel/ml sedangkanpada medium selektif 1,85 x 102 sel/ml.Jumlah sel regenerasi terbanyak ialah 1,58x 106 sel/ml, sehingga diperoleh frekuensifusi protoplas intraspesies antara B.japonicum L17-lkloram dengan B. japonicumPdl0ABkan sebesar 1,17 x 104.
Karakterisasi Bakteri Hasil Fusi
Hasil pengamatan menunjukkan bah-wa 1NTRA-1-INTRA-30 merupakan bakteriGram negatif dan berbentuk batang samadengan parentalnya. Bakteri hasil fusiINTRA-1-1NTRA-30 tidak dapat hidup padakonsentrasi NaCl 2%. Ketahanan hidupsemua hasil fusi pada konsentrasi NaCl 2%sama dengan parentalnya. Bakteri hasil fusiINTRA-l-INTRA-30 dan strain parental menunjukkan reaksi basa pada medium BTBdan tidak dapat menyerap warna congored.
Pertumbuhan pada medium YEMAdengan pH 4,5 dan 9 menunjukkan bahwa1NTRA-3, 1NTRA-4, INTRA-7, 1NTRA-11,INTRA-15, INTRA-19, dan INTRA-23 tumbuhsangat subur pada pH 4,5, INTRA-1, INTRA-2, INTRA-5, INTRA-6, 1NTRA-8-1NTRA-10,
Hasil Penelitian 2002
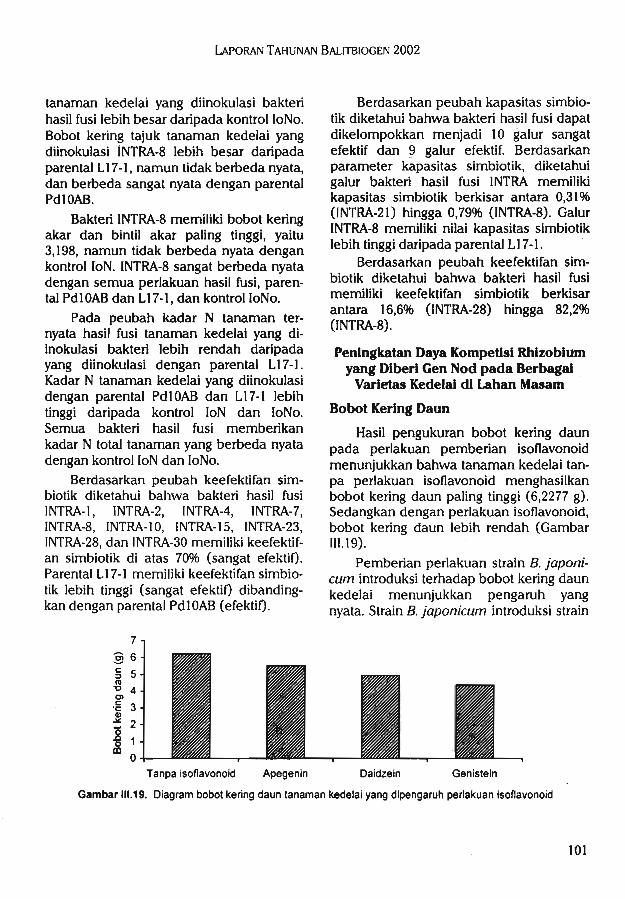
101
Tanpa isoflavonoid ApegeninDaidzeinGenistein
Gambar 111.19. Diagram bobot kering daun tanaman kedelai yang dipengaruh perlakuan isoflavonoid
Berdasarkan peubah kapasitas simbiotik diketahui bahwa bakteri hasil fusi dapatdikelompokkan menjadi 10 galur sangatefektif dan 9 galur efektif. Berdasarkanparameter kapasitas simbiotik, diketahuigalur bakteri hasil fusi INTRA memilikikapasitas simbiotik berkisar antara 0,31%(INTRA-21) hingga 0,79% (INTRA-8). GalurINTRA-8 memiliki nilai kapasitas simbiotiklebih tinggi daripada parental LI 7-1.
Berdasarkan peubah keefektifan simbiotik diketahui bahwa bakteri hasil fusimemiliki keefektifan simbiotik berkisarantara 16,6% (INTRA-28) hingga 82,2%(INTRA-8).
Peningkatan Daya Kompetisi Rhizobiumyang Diberi Gen Nod pada Berbagai
Varietas Kedelai di Lahan Masam
Bobot Kering Daun
Hasil pengukuran bobot kering daunpada perlakuan pemberian isoflavonoidmenunjukkan bahwa tanaman kedelai tan-pa perlakuan isoflavonoid menghasilkanbobot kering daun paling tinggi (6,2277 g).Sedangkan dengan perlakuan isoflavonoid,bobot kering daun lebih rendah (Gambar111.19).
Pemberian perlakuan strain B. japoni-
cum introduksi terhadap bobot kering daunkedelai menunjukkan pengaruh yangnyata. Strain B. japonicum introduksi strain
3(0•o
ht 2
0
7654
tanaman kedelai yang diinokulasi bakterihasil fusi lebih besar daripada kontrol IoNo.Bobot kering tajuk tanaman kedelai yangdiinokulasi INTRA-8 lebih besar daripadaparental LI 7-1, namun tidak berbeda nyata,
dan berbeda sangat nyata dengan parentalPdlOAB.
Bakteri INTRA-8 memiliki bobot keringakar dan bintil akar paling tinggi, yaitu3,198, namun tidak berbeda nyata dengankontrol IoN. INTRA-8 sangat berbeda nyatadengan semua perlakuan hasil fusi, parental Pdl OAB dan LI 7-1, dan kontrol IoNo.
Pada peubah kadar N tanaman ter-
nyata hasil fusi tanaman kedelai yang diinokulasi bakteri lebih rendah daripadayang diinokulasi dengan parental LI 7-1.Kadar N tanaman kedelai yang diinokulasidengan parental PdlOAB dan LI7-1 lebihtinggi daripada kontrol IoN dan IoNo.Semua bakteri hasil fusi memberikankadar N total tanaman yang berbeda nyatadengan kontrol IoN dan IoNo.
Berdasarkan peubah keefektifan sim-biotik diketahui bahwa bakteri hasil fusiINTRA-1, INTRA-2, INTRA-4, INTRA-7,
INTRA-8, INTRA-10, INTRA-15, INTRA-23,INTRA-28, dan INTRA-30 memiliki keefektifan simbiotik di atas 70% (sangat efektif).Parental LI 7-1 memiliki keefektifan simbiotik lebih tinggi (sangat efektif) dibanding-kan dengan parental PdlOAB (efektif).
Laporan Tahunan Balitbiogen 2002
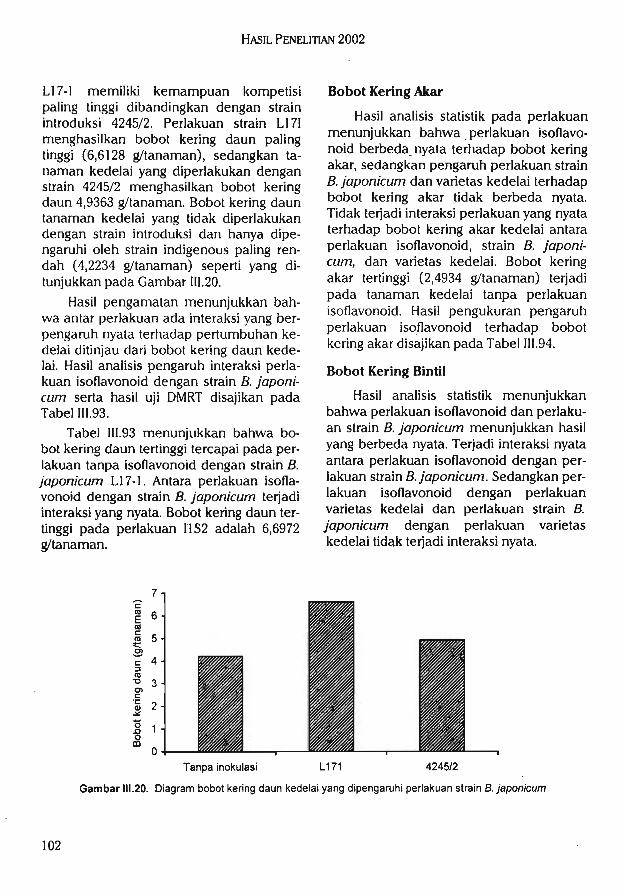
102
Tanpa inokulasiL1714245/2
Gambar III.20. Diagram bobot kering daun kedelai yang dipengaruhi perlakuan strain 8. japonicum
m1
I 2
ID eE 6IDI
Bobot Kering Akar
Hasil analisis statistik pada perlakuanmenunjukkan bahwa perlakuan isoflavonoid berbeda nyata terhadap bobot keringakar, sedangkan pengaruh perlakuan strain
B. japonicum dan varietas kedelai terhadapbobot kering akar tidak berbeda nyata.Tidak terjadi interaksi perlakuan yang nyataterhadap bobot kering akar kedelai antaraperlakuan isoflavonoid, strain B. japonicum, dan varietas kedelai. Bobot kering
akar tertinggi (2,4934 g/tanaman) terjadipada tanaman kedelai tanpa perlakuanisoflavonoid. Hasil pengukuran pengaruhperlakuan isoflavonoid terhadap bobotkering akar disajikan pada Tabel 111.94.
Bobot Kering Bintil
Hasil analisis statistik menunjukkanbahwa perlakuan isoflavonoid dan perlakuan strain B. japonicum menunjukkan hasilyang berbeda nyata. Terjadi interaksi nyataantara perlakuan isoflavonoid dengan perlakuan strain B. japonicum. Sedangkan per
lakuan isoflavonoid dengan perlakuanvarietas kedelai dan perlakuan strain B.japonicum dengan perlakuan varietaskedelai tidak terjadi interaksi nyata.
LI 7-1 memiliki kemampuan kompetisipaling tinggi dibandingkan dengan strainintroduksi 4245/2. Perlakuan strain LI 71menghasilkan bobot kering daun palingtinggi (6,6128 g/tanaman), sedangkan ta-naman kedelai yang diperlakukan denganstrain 4245/2 menghasilkan bobot keringdaun 4,9363 g/tanaman. Bobot kering daun
tanaman kedelai yang tidak diperlakukandengan strain introduksi dan hanya dipe-ngaruhi oleh strain indigenous paling ren-dah (4,2234 g/tanaman) seperti yang di-tunjukkan pada Gambar III.20.
Hasil pengamatan menunjukkan bah-wa antar perlakuan ada interaksi yang ber-
pengaruh nyata terhadap pertumbuhan kedelai ditinjau dari bobot kering daun kedelai. Hasil analisis pengaruh interaksi perlakuan isoflavonoid dengan strain B. japonicum serta hasil uji DMRT disajikan padaTabel 111.93.
Tabel 111.93 menunjukkan bahwa bobot kering daun tertinggi tercapai pada perlakuan tanpa isoflavonoid dengan strain B.japonicum LI 7-1. Antara perlakuan isoflavonoid dengan strain B. japonicum terjadiinteraksi yang nyata. Bobot kering daun tertinggi pada perlakuan I1S2 adalah 6,6972g/tanaman.
Hasil Penelitian 2002

103
dap bobot kering bintil akar kedelai ditun-jukkan pada Gambar 111.22. Bobot keringbintil tanaman kedelai yang diinokulasi dengan isolat introduksi B. japonicum strainLI 71 paling besar (0,3885 g/tanaman), di-ikuti bobot kering bintil tanaman kedelaiyang diinokulasi dengan B. japonicumstrain 4245/2 (0,1642 g/tanaman). Tanamankedelai yang tidak diinokulasi denganstrain introduksi pembentukan bintil akandipengaruhi oleh strain indigenous yangberada dalam tanah masam. Dari data
yang diperoleh, ternyata pengaruh strainindigenous terhadap bobot kering bintilakar kedelai lebih rendah apabila diban-dingkan dengan pengaruh strain introduksi.Hal ini berarti kedua isolat B. japonicumintroduksi strain LI 71 dan 4245/2 memilikikemampuan kompetisi yang lebih baik terhadap bakteri pembentuk bintil indigenous.
Dari perlakuan isoflavonoid, strain B.japonicum, dan varietas kedelai temyataterjadi interaksi perlakuan yang berpenga-ruh nyata terhadap bobot kering bintil akarkedelai, yaitu interaksi antara perlakuanisoflavonoid dengan strain B. japonicum.Pengaruh interaksi perlakuan isoflavonoiddengan strain B. japonicum terhadap bobotkering bintil kedelai yang paling besar adalah interaksi antara tanpa perlakuan isoflavonoid dengan inokulasi B. japonicumstrain LI 71, yaitu sebesar 0,6727 g/tanaman,diikuti pengaruh interaksi perlakuan isoflavonoid daidzein dengan B. japonicumstrain LI 71 sebesar 0,3915 g/tanaman.
Kadar Nitrogen
Hasil analisis statistik menunjukkanpengaruh perlakuan isoflavonoid, strain B.japonicum, dan varietas kedelai menunjukkan kadar nitrogen yang berbeda nyata.Pada percobaan ini terjadi pengaruh inter-
Bobot kering bintil kedelai paling tinggi(0,3095 g/tanaman) diperoleh tanpa perlakuan isoflavonoid. Gambar 111.21 menun-
jukkan bobot kering bintil kedelai yang di-perlakukan dengan isoflavonoid lebih ren-dah daripada tanpa perlakuan isoflavonoid.Dari ketiga isoflavonoid yang digunakan,pengaruh terhadap bobot kering bintil kedelai yang ditimbulkan secara berurutanadalah daidzein memberikan pengaruhyang paling besar terhadap bobot keringbintil kedelai, yaitu 0,236 g/tanaman,apigenin 0,1737 g/tanaman, dan genestein0,1665 g/tanaman.
Pengaruh pemberian isolat B. japonicum dan kemampuan kompetisi isolatintroduksi dengan isolat indigenous terha
Angka-angka dalam satu lajur yang diikutl oleh hurufsama tidak berbeda pada taraf nyata menurut ujiDMRT
2,49341,5616"1,9639"1,5541"
1.Tanpa isoflavonoid2.Apigenin3.Daidzein4.Genistein
Bobot kering akarNo. Perlakuan
Angka-angka dalam satu lajur yang diikutl huruf samatidak berbeda pada taraf nyata menurut uji DMRT
Tabel III.94. Pengaruh perlakuan pemberian isoflavo-noid terhadap bobot kering akar kedelai
4,8743"4,0361"4,146"5,0544""6,4662bc3,3082"5,1525""6,6972b4,6924"4,6641"9,2519a4,7671cd
Bobot kering daun (g)
I4S3I4S2I4S1I3S3I3S2I3S1I2S3I2S2I2S111 S3I1S2I1S1
Perlakuan
12.11.10.9.8.7.6.5.4.3.2.1.
No.
Tabel 111.93. Pengaruh interaksi perlakuan isoflavo-noid dengan perlakuan strain 8. japoni-cum terhadap bobot kering daun kedelai
Laporan Tahunan Balitbiogen 2002
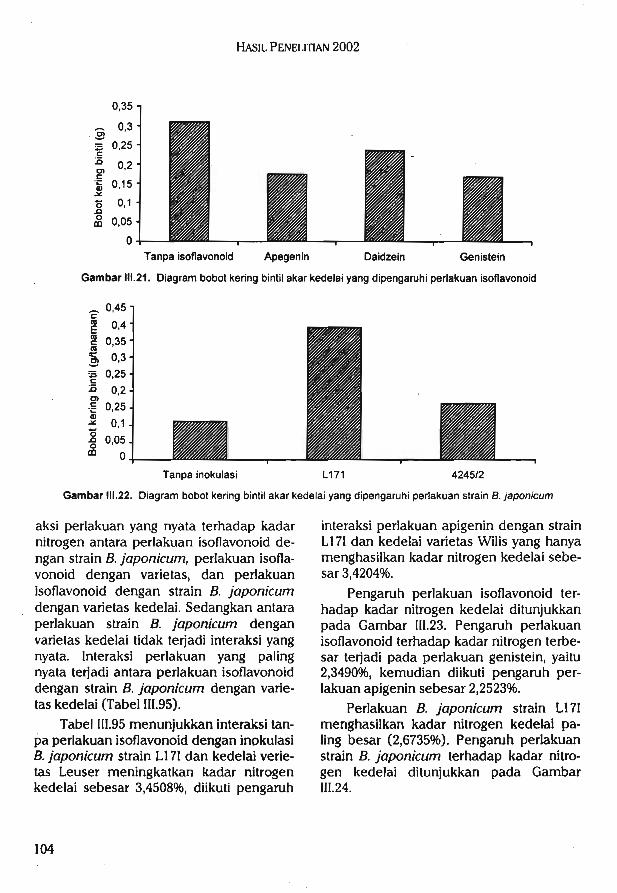
104
interaksi perlakuan apigenin dengan strainLI 71 dan kedelai varietas Wilis yang hanyamenghasilkan kadar nitrogen kedelai sebesar 3,4204%.
Pengaruh perlakuan isoflavonoid terhadap kadar nitrogen kedelai ditunjukkanpada Gambar 111.23. Pengaruh perlakuanisoflavonoid terhadap kadar nitrogen terbe-sar terjadi pada perlakuan genistein, yaitu2,3490%, kemudian diikuti pengaruh perlakuan apigenin sebesar 2,2523%.
Perlakuan B. japonicum strain LI 71menghasilkan kadar nitrogen kedelai paling besar (2,6735%). Pengaruh perlakuanstrain B. japonicum terhadap kadar nitrogen kedelai ditunjukkan pada GambarHI.24.
aksi perlakuan yang nyata terhadap kadarnitrogen antara perlakuan isoflavonoid de-
ngan strain B. japonicum, periakuan isoflavonoid dengan varietas, dan perlakuanisoflavonoid dengan strain B. japonicumdengan varietas kedelai. Sedangkan antaraperlakuan strain B. japonicum denganvarietas kedelai tidak terjadi interaksi yangnyata. Interaksi perlakuan yang palingnyata terjadi antara perlakuan isoflavonoiddengan strain B. japonicum dengan varietas kedelai (Tabel 111.95).
Tabel 111.95 menunjukkan interaksi tanpa perlakuan isoflavonoid dengan inokulasiB. japonicum strain LI 71 dan kedelai verie-tas Leuser meningkatkan kadar nitrogenkedelai sebesar 3,4508%, diikuti pengaruh
Tanpa inokulasiL1714245/2
Gambar III.22. Diagram bobot kering bintil akar kedelai yang dipengaruhi perlakuan strain 6. japonicum
0,45
0,4-0,35
0,3
0,250,2
0,25-
0,1
0,05.
0
Tanpa isoflavonoid ApegeninDaidzeinGenistein
Gambar 111.21. Diagram bobot kering bintil akar kedelai yang dipengaruhi perlakuan isoflavonoid
A
0,35
^ 0,335 0,25
0,2"
0,15
0,1
0,05
Hasil Penelitian 2002
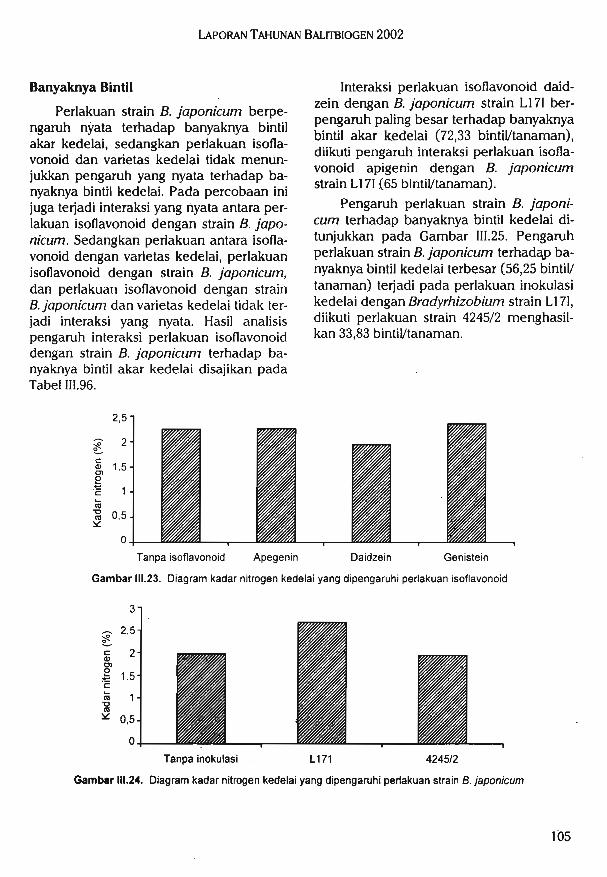
105
Tanpa inokulasiL1714245/2
Gambar 111.24. Diagram kadar nitrogen kedelai yang dipengaruhi perlakuan strain B. japonicum
1-
0,5-
0
Tanpa isoflavonoid ApegeninDaidzeinGenistein
Gambar 111.23. Diagram kadar nitrogen kedelai yang dipengaruhi perlakuan isoflavonoid
3
^2,5
I 1-1
• 0.5-|
2,5 n
g 2I 15
Interaksi perlakuan isoflavonoid daid-zein dengan B. japonicum strain LI 71 ber-pengaruh paling besar terhadap banyaknyabintil akar kedelai (72,33 bintil/tanaman),diikuti pengaruh interaksi perlakuan isoflavonoid apigenin dengan B. japonicumstrain LI 71 (65 bintil/tanaman).
Pengaruh perlakuan strain B. japonicum terhadap banyaknya bintil kedelai di-tunjukkan pada Gambar 111.25. Pengaruhperlakuan strain B. japonicum terhadc> banyaknya bintil kedelai terbesar (56,25 bintil/tanaman) terjadi pada perlakuan inokulasikedelai dengan Bradyrhizobium strain LI 71,diikuti perlakuan strain 4245/2 menghasil-kan 33,83 bintil/tanaman.
Banyaknya Bintil
Perlakuan strain B. japonicum berpe-
ngaruh nyata terhadap banyaknya bintilakar kedelai, sedangkan perlakuan isofla-vonoid dan varietas kedelai tidak menun-
jukkan pengaruh yang nyata terhadap banyaknya bintil kedelai. Pada percobaan inijuga terjadi interaksi yang nyata antara perlakuan isoflavonoid dengan strain B. japonicum. Sedangkan perlakuan antara isoflavonoid dengan varietas kedelai, perlakuanisoflavonoid dengan strain B. japonicum,dan perlakuan isoflavonoid dengan strainB. japonicum dan varietas kedelai tidak terjadi interaksi yang nyata. Hasil analisispengaruh interaksi perlakuan isoflavonoiddengan strain B. japonicum terhadap banyaknya bintil akar kedelai disajikan padaTabei 111.96.
Laporan Tahunan Balitbiogen 2002
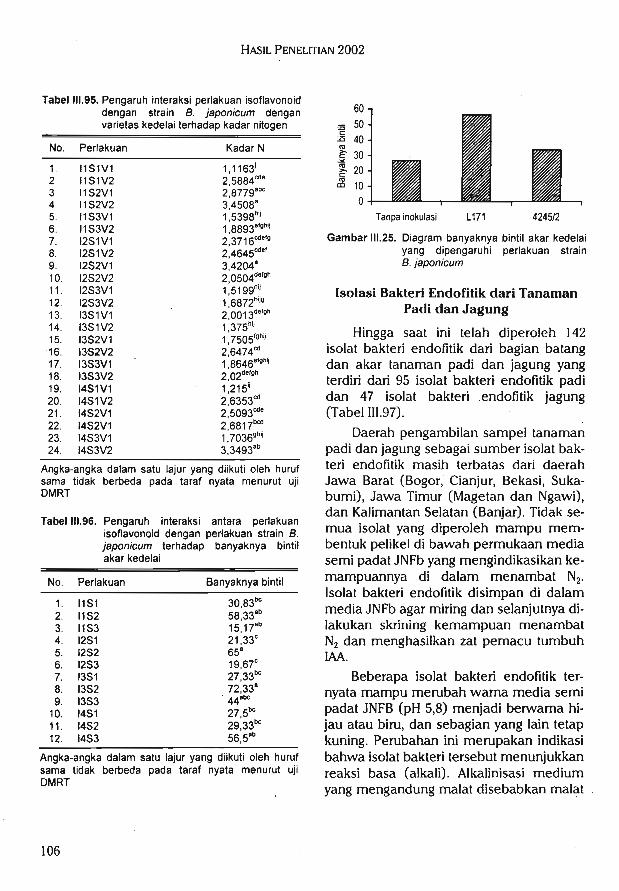
106
Gambar 111.25. Diagram banyaknya bintil akar kedelaiyang dipengaruhi perlakuan strainB. japonicum
Isolasi Bakteri Endofitik dari TanamanPadi dan Jagung
Hingga saat ini telah diperoleh 142isolat bakteri endofitik dari bagian batangdan akar tanaman padi dan jagung yangterdiri dari 95 isolat bakteri endofitik padidan 47 isolat bakteri .endofitik jagung(Tabel 111.97).
Daerah pengambilan sampel tanaman
padi dan jagung sebagai sumber isolat bakteri endofitik masih terbatas dari daerahJawa Barat (Bogor, Cianjur, Bekasi, Suka-
bumi), Jawa Timur (Magetan dan Ngawi),dan Kalimantan Selatan (Banjar). Tidak se-mua isolat yang diperoleh mampu mem-
bentuk pelikel di bawah permukaan mediasemi padat JNFb yang mengindikasikan ke-mampuannya di dalam menambat N2.Isolat bakteri endofitik disimpan di dalammedia JNFb agar miring dan selanjutnya di-lakukan skrining kemampuan menambatN2 dan menghasilkan zat pemacu tumbuhIAA.
Beberapa isolat bakteri endofitik ter-nyata mampu merubah warna media semi
padat JNFB (pH 5,8) menjadi berwarna hi-jau atau biru, dan sebagian yang lain tetapkuning. Perubahan ini merupakan indikasibahwa isolat bakteri tersebut menunjukkanreaksi basa (alkali). Alkalinisasi mediumyang mengandung malat disebabkan malat
4245/2L171Tanpa inokulasi
60-50-40-30-20 -10-
0
Angka-angka dalam satu lajur yang diikuti oleh hurufsama tidak berbeda pada taraf nyata menurut ujiDMRT
56,5'"29,33""27,5""44'""72,33'27,33""19,67"65'21,33"15,17"58,33'"30,83""
Banyaknya bintil
I4S3I4S2I4S1I3S3I3S2I3S1I2S3I2S2I2S1I1S3I1S2I1S1
Perlakuan
12.11.10.9.8.7.6.5.4.3.2.1.
No.
Tabel 111.96. Pengaruh interaksi antara perlakuanisoflavonoid dengan perlakuan strain B.japonicum terhadap banyaknya bintilakar kedelai
Angka-angka dalam satu lajur yang diikuti oleh hurufsama tidak berbeda pada taraf nyata menurut ujiDMRT
1,1163'2,5884""'2,8779'"3,4508'1,5398""1,8893"""2,3716"""2,4645"""3,42042,0504"""1,5199""1,6872""2,0013"""1,375""1,7505'""2,6474""1,8646"""2,02"""1,215*2,6353""2,5093""'2,6817"""1,7036""3,3493'"I4S3V2
I4S3V1I4S2V1I4S2V1I4S1V2I4S1V1I3S3V2I3S3V1I3S2V2I3S2V1I3S1V2I3S1V1I2S3V2I2S3V1I2S2V2I2S2V1I2S1V2I2S1V1I1S3V2I1S3V1I1S2V2I1S2V1I1S1V2I1S1V1
24.23.22.21.20.19.18.17.16.15.14.13.12.11.10.9.8.7.6.5.4.3.2.1.
Kadar NNo. Perlakuan
Tabel 111.95. Pengaruh interaksi perlakuan isoflavonoiddengan strain B. japonicum denganvarietas kedelai terhadap kadar nitogen
Hasil Penelitian 2002

107
Di samping mampu menambat N2,
beberapa isolat bakteri endofitik mampumenghasilkan zat pemacu tumbuh auksin(IAA) dengan kisaran produksi IAA ialah 0-8,295 ppm.
Berdasarkan hasil seleksi terhadapisolat bakteri endofitik diperoleh lima isolatunggul bakteri endofitik padi dan limaisolat unggul bakteri endofitik jagung, yaitua.BCr 1.2 dengan kemampuan produksi
IAA 4,530 ppm, namun kemampuanmenambat N2 belum diamati, sehinggaisolat ini dipilih berdasarkan besarnyakemampuan produksi IAA.
b.BCbd 1.3 dengan kemampuan produksiIAA 5,363 ppm dan kemampuan menambat N2 udara (0,0116 ^u,mol/jam/tabung).
c.APK 2.4 dengan kemampuan produksi
IAA 2,818 ppm dan kemampuan menambat N2 udara 2,683 jumol/jam/ta-bung.
d.BCr 2.3 dengan kemampuan produksi
IAA 1,539 ppm dan kemampuan menambat N2 udara 1,9495 ^umol/jam/ta-bung.
e.BCr 2.1 dengan kemampuan produksi
IAA 1,727 ppm dan kemampuan menambat N21,277 ^imol/jarrVtabung.
Kelima isolat unggul bakteri endofitikpadi selanjutnya diuji kompatibilitasnya terhadap tanaman inang (tanaman padi varie-tas IR64). Kriteria pemilihan kelima isolatunggul didasarkan pada kemampuan menambat N2 dan atau kemampuan menghasilkan zat pemacu tumbuh IAA.
mengalami oksidasi. Pada Azospirillumlipoferum mungkin berkaitan dengan per-kembangan polimorfisme.
Di samping itu, beberapa isolat bakteriendofitik mampu membentuk pelikel dengan jelas dan berada beberapa milimeterdi bawah permukaan medium JNFB semipadat, namun isolat bakteri endofitik yanglain menunjukkan pelikel yang kurang jelaspada permukaan medium. Adanya pelikelyang berupa kabut tipis ini memberikankondisi yang baik untuk aktivitas nitroge-nase (penambatan N2). Hal ini disebabkantidak ada kelebihan oksigen pada mediatersebut, karena laju difusi oksigen ke da-lam media dapat disamakan (korespon)dengan laju respirasi mikroorganisme.
Seleksi Bakteri Endofitik dalam HalKemampuan Menambat N2 dan
Menghasilkan Zat Pemacu Tumbuh IAA
Hasil seleksi bakteri endofitik dalamhal kemampuan menambat N2 dan menghasilkan zat pemacu tumbuh IAA disajikanpada Tabel 111.98.
Isolat yang mampu membentuk pelikel diuji kemampuan menambat N2 ber-dasarkan aktivitas nitrogenasenya (enzimnitrogenase dapat mereduksi N2 menjadiNH3 yang setara dengan kemampuan me
reduksi asetilen menjadi etilen). Etilen hasil reduksi diukur dengan alat gas kromato-grafi. Dari 143 isolat bakteri endofitik padiyang diuji aktivitas nitogenasenya, diper-oleh tiga isolat yang memiliki kemampuanmenambat N2 tinggi, yaitu BCr 2.3 (1,9495/imol/jam/tabung), BCr 2.1 (1,277jam/tabung), dan APK 2.4 (2,683jam/tabung).
Laporan Tahunan Balitbiogen 2002
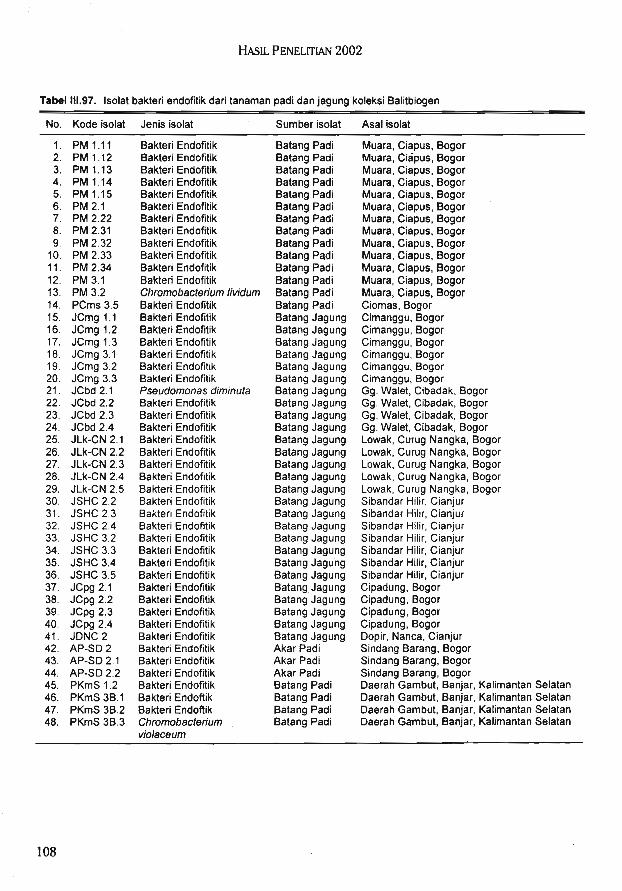
108
Daerah Gambut, Banjar, Kalimantan SelatanDaerah Gambut, Banjar, Kalimantan SelatanDaerah Gambut, Banjar, Kalimantan SelatanDaerah Gambut, Banjar, Kalimantan SelatanSindang Barang, BogorSindang Barang, BogorSindang Barang, BogorDopir, Nanca, CianjurCipadung, BogorCipadung, BogorCipadung, BogorCipadung, BogorSibandar Hilir, CianjurSibandar Hilir, CianjurSibandar Hilir, CianjurSibandar Hilir, CianjurSibandar Hilir, CianjurSibandar Hilir, CianjurSibandar Hilir, CianjurLowak, Curug Nangka, BogorLowak, Curug Nangka, BogorLowak, Curug Nangka, BogorLowak, Curug Nangka, BogorLowak, Curug Nangka, BogorGg. Walet, Cibadak, BogorGg. Walet, Cibadak, BogorGg. Walet, Cibadak, BogorGg. Walet, Cibadak, BogorCimanggu, BogorCimanggu, BogorCimanggu, BogorCimanggu, BogorCimanggu, BogorCimanggu, BogorCiomas, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Ciapus, BogorMuara, Clapus, Bogor
Asal isolat
Batang PadiBatang PadiBatang PadiBatang PadiAkar PadiAkar PadiAkar PadiBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang JagungBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang Padi
Sumber isolat
violaceumChromobacteriumBakteri EndoftikBakteri EndoftikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikPseudomonas diminutaBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikChromobacterium lividumBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri Endofitik
Jenis isolat
PKmS 3B.3PKmS 3B.2PKmS3B1PKmS 1.2AP-SD 2.2AP-SD2.1AP-SD 2JDNC2JCpg 2.4JCpg 2.3JCpg 2.2JCpg 2.1JSHC 3.5JSHC 3.4JSHC 3.3JSHC 3.2JSHC 2.4JSHC 2.3JSHC 2.2JLk-CN 2.5JLk-CN 2.4JLk-CN 2.3JLk-CN 2.2JLk-CN 2.1JCbd 2.4JCbd 2.3JCbd 2.2JCbd 2.1JCmg 3.3JCmg 3.2JCmg 3.1JCmg 1.3JCmg 1.2JCmg 1.1PCms 3.5PM3.2PM3.1PM 2.34PM 2.33PM 2.32PM 2.31PM 2.22PM2.1PM 1.15PM 1.14PM1.13PM1.12PM1.11
Kode isolat
48.47.46.45.44.43.42.41.40.39.38.37.36.35.34.33.32.31.30.29.28.27.26.25.24.23.22.21.20.19.18.17.16.15.14.13.12.11.10.9.8.7.6.5.4.3.2.1.
No.
Tabel 111.97. Isolat bakteri endofitik dari tanaman padi dan jagung koleksi Balitbiogen
Hasil Penelitian 2002

109
Bendo, Magetan, Jawa TimurBendo, Magetan, Jawa TimurBendo, Magetan, Jawa TimurBendo, Magetan, Jawa TimurBendo, Magetan, Jawa TimurCibadak, Sukabumi; Jawa BaratCibadak, Sukabumi, Jawa BaratCibadak, Sukabumi, Jawa BaratCibadak, Sukabumi, Jawa BaratCibadak, Sukabumi, Jawa BaratCibadak, Sukabumi, Jawa BaratCibadak, Sukabumi, Jawa BaratCibadak, Sukabumi, Jawa BaratCibadak, Sukabumi, Jawa BaratLeuwiliang, Bogor, Jawa BaratLeuwiliang, Bogor, Jawa BaratLeuwiliang, Bogor, Jawa BaratLeuwiliang, Bogor, Jawa BaratRabak, Rumpin, Bogor, Jawa BaratRabak, Rumpin, Bogor, Jawa BaratRabak, Rumpin, Bogor, Jawa BaratRabak, Rumpin, Bogor, Jawa BaratRabak, Rumpin, Bogor, Jawa BaratRabak, Rumpin, Bogor, Jawa BaratRabak, Rumpin, Bogor, Jawa BaratRabak, Rumpin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratCaringin, Bogor, Jawa BaratParung, Bogor, Jawa BaratParung, Bogor, Jawa BaratParung, Bogor, Jawa BaratParung, Bogor, Jawa BaratParung, Bogor, Jawa BaratParung, Bogor, Jawa BaratCikarang, BekasiCikarang, BekasiCikarang, BekasiCikarang, BekasiKedung Waringin, CimangguKedung Waringin, CimangguKedung Waringin, CimangguKedung Waringin, CimangguKedung Waringin, CimangguKedung Warlngln, Cimanggu
Asal isolat
Batang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiBatang PadiAkar JagungAkar JagungAkar JagungBatang JagungBatang JagungBatang Jagung
Sumber isolat
Bakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndoftikBakteri EndoftikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndoftikBakteri EndoftikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndoftikBakteri EndoftikBakteri EndofitikBakteri EndofitikBakteri EndofttikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndoftikBakteri EndoftikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndoftikBakteri EndoftikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri EndofitikBakteri Endofitik
Jenis isolat
BBd1.2BBd1ABd3.2ABd 3.1ABd2.1BCbd 3.2BCbd 3.1BCbd 2.4BCbd 2.3BCbd 2.2BCbd 2.1BCbd 1.4BCbd1.3BCbd 1.2BLw 3.2BLw3.1BLw2.2BLw2.1BRb 3.4BRb 3.32BRb 3.31BRb 3.2BRb 3.1BRb 2.3BRb 2.2BRb 2.1BCr 3.4BCr 3.2BCr 3.1BCr 2.3BCr 2.2BCr 2.1BCr 1.3.2BCr 1.3.1BCM.3BCM.2BCM.1BPr 2.3BPr 2.2BPr 1.2.3BPr 1.2.2BPr 1.2.1BPM.1C2.4C2.3C2.2C2.1JKW2.2AJKW2.1AJKW1.3AJKW1.3BJKW 1 2BJKW1.1B
Kode isolat
101.100.99.98.97.96.95.94.93.92.91.90.89.88.87.86.85.84.83.82.81.80.79.78.77.76.75.74.73.72.71.70.69.68.67.66.65.64.63.62.61.60.59.58.57.56.55.54.53.52.51.50.49.
No.
Tabel 111.97. Lanjutan
Laporan Tahunan Balitbiogen 2002
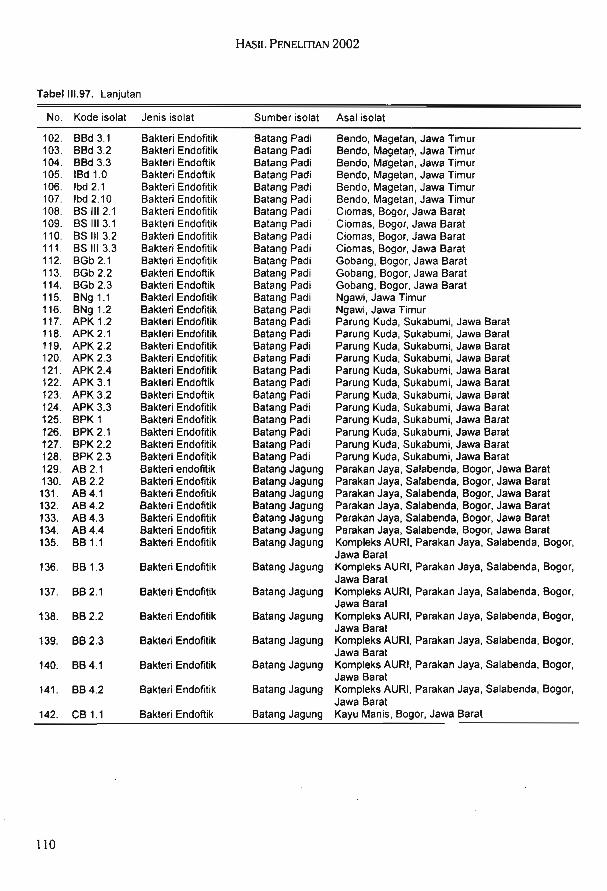
110
102.BBd 3.1Bakteri EndofitikBatang PadiBendo, Magetan, Jawa Timur103.BBd 3.2Bakteri EndofitikBatang PadiBendo, Magetan, Jawa Timur104.BBd 3.3Bakteri EndoftikBatang PadiBendo, Magetan, Jawa Timur105.IBd 1.0Bakteri EndoftikBatang PadiBendo, Magetan, Jawa Timur106.Ibd 2.1Bakteri EndofitikBatang PadiBendo, Magetan, Jawa Timur107.Ibd 2.10Bakteri EndofitikBatang PadiBendo, Magetan, Jawa Timur108.BS III 2.1Bakteri EndofitikBatang PadiCiomas, Bogor, Jawa Barat109.BS III 3.1Bakteri EndofitikBatang PadiCiomas, Bogor, Jawa Barat110.BS III 3.2Bakteri EndofitikBatang PadiCiomas, Bogor, Jawa Barat111.BS III 3.3Bakteri EndofitikBatang PadiCiomas, Bogor, Jawa Barat112.BGb2.1Bakteri EndofitikBatang PadiGobang, Bogor, Jawa Barat113.BGb2.2Bakteri EndoftikBatang PadiGobang, Bogor, Jawa Barat114.BGb2.3Bakteri EndoftikBatang PadiGobang, Bogor, Jawa Barat115.BNg 1.1Bakteri EndofitikBatang PadiNgawi, Jawa Timur116.BNg 1.2Bakteri EndofitikBatang PadiNgawi, Jawa Timur117.APK1.2Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat118.APK 2.1Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat119.APK 2.2Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat120.APK 2.3Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat121.APK 2.4Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat122.APK 3.1Bakteri EndoftikBatang PadiParung Kuda, Sukabumi, Jawa Barat123.APK 3.2Bakteri EndoftikBatang PadiParung Kuda, Sukabumi, Jawa Barat124.APK 3.3Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat125.BPK 1Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat126.BPK 2.1Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat127.BPK 2.2Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat128.BPK 2.3Bakteri EndofitikBatang PadiParung Kuda, Sukabumi, Jawa Barat129.AB2.1Bakteri endofitikBatang JagungParakan Jaya, Salabenda, Bogor, Jawa Barat130.AB2.2Bakteri EndofitikBatang JagungParakan Jaya, Salabenda, Bogor, Jawa Barat
131.AB4.1Bakteri EndofitikBatang JagungParakan Jaya, Salabenda, Bogor, Jawa Barat132.AB4.2Bakteri EndofitikBatang JagungParakan Jaya, Salabenda, Bogor, Jawa Barat133.AB4.3Bakteri EndofitikBatang JagungParakan Jaya, Salabenda, Bogor, Jawa Barat134.AB4.4Bakteri EndofitikBatang JagungParakan Jaya, Salabenda, Bogor, Jawa Barat135.BB1.1Bakteri EndofitikBatang JagungKompleks AURI, Parakan Jaya, Salabenda, Bogor,
Jawa Barat136.BB1.3Bakteri EndofitikBatang JagungKompleks AURI, Parakan Jaya, Salabenda, Bogor,
Jawa Barat137.BB2.1Bakteri EndofitikBatang JagungKompleks AURI, Parakan Jaya, Salabenda, Bogor,
Jawa Barat138.BB2.2Bakteri EndofitikBatang JagungKompleks AURI, Parakan Jaya, Salabenda, Bogor,
Jawa Barat139.BB2.3Bakteri EndofitikBatang JagungKompleks AURI, Parakan Jaya, Salabenda, Bogor,
Jawa Barat140.BB4.1Bakteri EndofitikBatang JagungKompleks AURI, Parakan Jaya, Salabenda, Bogor,
Jawa Barat141.BB4.2Bakteri EndofitikBatang JagungKompleks AURI, Parakan Jaya, Salabenda, Bogor,
Jawa Barat142.CB1.1Bakteri EndoftikBatang JagungKayu Manis, Bogor, Jawa Barat
Sumber isolat Asal isolatNo. Kode isolat Jenis isolat
Tabel 111.97. Lanjutan
Hasil Penelitian 2002
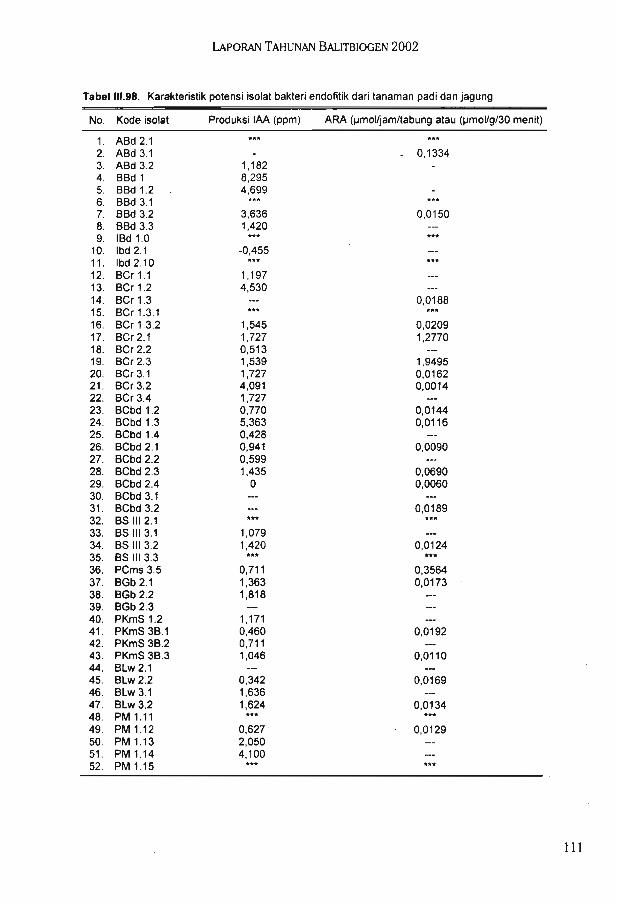
til
0,0192
0,0110
0,0169
0,0134
0,0129
0,0124
0,35640,0173
0,0188***
0,02091,2770
1,94950,01620,0014
0,01440,0116
0,0090
0,06900,0060
0,0189
0,0150
0,1334
***4,1002,0500,627
***1,6241,6360,342
1,0460,7110,4601,171
—1,8181,3630,711
***1,4201,079
***—
01,4350,5990,9410,4285,3630,7701,7274,0911,7271,5390,5131,7271,545
***
4,5301,197
***-0,455
***1,4203,636
***4,6998,2951,182
-
***
PM1.15PM1.14PM1.13PM1.12PM 1.11BLw 3.2BLw3.1BLw 2.2BLw2.1PKmS 3B.3PKmS 3B.2PKmS3B.1PKmS1.2BGb2 3BGb 2.2BGb2.1PCms 3.5BS III 3.3BS III 3.2BS III 3.1BS III 2.1BCbd 3.2BCbd 3.1BCbd 2.4BCbd 2.3BCbd 2.2BCbd 2.1BCbd 1.4BCbd1.3BCbd 1.2BCr 3.4BCr 3.2BCr 3.1BCr 2.3BCr 2.2BCr 2.1BCr 1.3.2BCr 1.3.1BCM.3BCr1.2BCM.1Ibd 2.10Ibd 2.1IBdi.OBBd 3.3BBd 3.2BBd 3.1BBd 1.2 .BBd1ABd 3.2ABd3.1ABd2.1
52.51.50.49.48.47.46.45.44.43.42.41.40.39.38.37.36.35.34.33.32.31.30.29.28.27.26.25.24.23.22.21.20.19.18.17.16.15.14.13.12.11.10.
9.8.7.6.5.4.3.2.1.
Produksi IAA (ppm) ARA (pmol/jam/tabung atau (pmol/g/30 menit)No. Kode isolat
Tabel 111.98. Karakteristik potensi isolat bakterl endofitik dari tanaman padi dan jagung
Laporan Tahunan Balitbiogen 2002
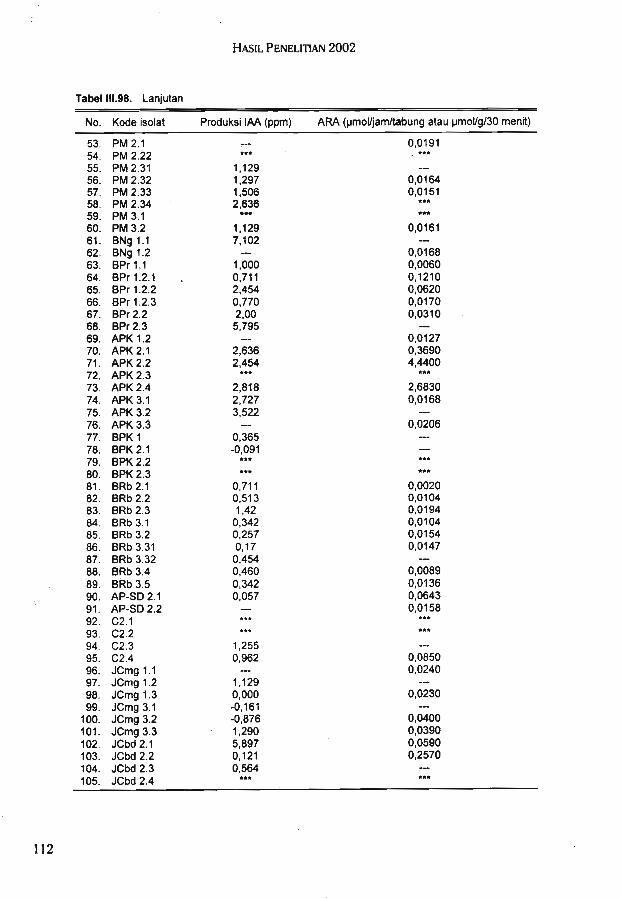
112
0,08500,0240
0,0230
0,04000,03900,05900,2570
0,00200,01040,01940,01040,01540,0147
0,00890,01360,06430,0158
0,0161
0,01680,00600,12100,06200,01700,0310
0,01270,36904,4400
2,68300,0168
0,0206
0,01640,0151
0,0191
***0,5640,1215,8971,290-0,876-0,1610,0001,129
—0,9621,255
******—
0,0570,3420,4600,4540,17
0,2570,3421,42
0,5130,711
******
-0,0910,365
—
3,5222,7272,818
***2,4542,636
5,7952,00
0,7702,4540,7111,000_.
7,1021,129
***2,6361,5061,2971,129
#**•.—
JCbd 2.4JCbd 2.3JCbd 2.2JCbd 2.1JCmg 3.3JCmg 3.2JCmg 3.1JCmg 1.3JCmg1.2JCmg1.1C2.4C2.3C2.2C2.1AP-SD 2.2AP-SD2.1BRb 3.5BRb 3.4BRb 3.32BRb 3.31BRb 3.2BRb 3.1BRb 2.3BRb 2.2BRb 2.1BPK 2.3BPK 2.2BPK2.1BPK1APK 3.3APK 3.2APK 3.1APK 2.4APK 2.3APK 2.2APK 2.1APK1.2BPr 2.3BPr 2.2BPr 1.2.3BPr 1.2.2BPr 1.2.1BPM.1BNg 1.2BNg 1.1PM3.2PM3.1PM 2.34PM 2.33PM 2.32PM 2.31PM 2.22PM2.1
105.104.103.102.101.100.99.98.97.96.95.94.93.92.91.90.89.88.87.86.85.84.83.82.81.80.79.78.77.76.75.74.73.72.71.70.69.68.67.66.65.64.63.62.61.60.59.58.57.56.55.54.53.
Produksi IAA (ppm) ARA (pmol/jam/tabung atau pmol/g/30 menit)No. Kode isolat
Tabel 111.98. Lanjutan
Hasil Penelitian 2002

113
c.JKW 1.1B dengan kemampuan produksi
IAA 1,036 ppm, namun kemampuanmenambat N2 0,0620.
d.JSHC 2.4 dengan kemampuan produksiIAA 0,322 ppm dan kemampuan menambat N2 udara 0,2190 ^,mol/jam/tabung.
e.JLkCN 2.3 dengan kemampuan produksi IAA 0,677 ppm dan kemampuanmenambat N2 udara 0,4690 /umol/jam/tabung.
0,0860
0,10200,03600,08300,04400,0620
***0,05800,0540
0,01500,2190
0,0550
0,46900,0390
*** = belum dilakukan pengamatan
Selain itu, telah diperoleh dan diseleksilima isolat unggul endofitik untuk tanaman
jagung, yaitua.JCbd 2.1 dengan kemampuan produksi
IAA 5,897 ppm dan kemampuan me-nambat N2 0,0590 /xmol/jam/tabung.
b.JCmg 3.3 dengan kemampuan produksiIAA 1,290 ppm dan kemampuan me-nambat N2 udara 0,0390 ^,mol/jam/tabung.
0.2390,120
******
-0,3190,956
***-0,757
***-0,797
******
0,598-0,0402,318
******
.-1,315
***1,0362,298
...6,2103,629
***-0,4380,1601,089
***0,322-0,202
***-0,1213,1450,6770,2790,040
CB1.1BB4.2BB4.1BB2.3BB2.2BB2.1BB 1.3BB1.1AB4.4AB4.3AB4.2AB4.1AB2.2AB2.1JKW2.2AJKW2.1AJKW1.3AJKW2.2BJKW1.3BJKW1.2BJKW1.1BJDNC2JCpg 2.4JCpg 2.3JCpg 2.2JCpg 2.1JSHC 3.5JSHC 3.4JSHC 3.3JSHC 3.2JSHC 2.4JSHC 2.3JSHC 2.2JLk-CN 2.5JLk-CN 2.4JLk-CN 2.3JLk-CN 2.2JLk-CN 2.1
143.142.141.140.139.138.137.136.135.134.133.132.131.130.129.128.127.126.125.124.123.122.121.120.119.118.117.116.115.114.113.112.111.110.109.108.107.106.
Produksi IAA (ppm) ARA (pmol/jam/tabung atau pmol/g/30 menit)No. Kode isolat
Tabel 111.98. Lanjutan
Laporan Tahunan Balitbiogen 2002

114
-
0,0562
0,00730,0244
-
0,01470,00270,0078
-
di batang
0,01380,01210,0209
0,0131
0,0113
-
di akar
ARA (pmol/jam/tabung)
1II
III
III1IIIII
1II
III1II
1II
IIIIIIIII
III
III
Ulangan
BCbd 1.3
BCr 2.3
BCr 2.1
BLw 3.2
BCr 1.2
BRb 2.2
APK 2.1
APK 2.4
Kode Isolat
Tabel 111.99. Hasil analisis reduksi asetilen di bagian akar danbatang pada uji kompatibilitas bakteri endofitik dengantanaman inang padi IR64
Pengaruh inokulasi bakteri endofitikpadi terhadap kemampuan menambat N2memberikan hasil yang positif, dalam artiinokulasi bakteri endofitik memberikan data kemampuan penambatan N2 udara yang
lebih tinggi daripada perlakuan kontroltanpa inokulasi (Tabel III.100).
Pengaruh inokulasi bakteri endofitikpada banyaknya akar lateral tanaman paditernyata berkorelasi positif, yaitu tanaman
padi yang telah diinokulasi kultur bakteriendofitik, baik dari strain BCbd 1.3, APK2.4, BCr 1.2, BCr 2.3, dan BCr 2.1 memiliki
akar lateral lebih banyak daripada tanamanpadi kontrol tanpa inokulasi (Tabel III.101).Demikian juga apabila dibandingkan dengan tanaman padi yang diinokulasi dengan kontrol IAA (2,4-D) dengan konsentra-si 10s, 10"6, 10"r, dan 108 ternyata tanaman
Uji Kompatibilitas Bakteri Endofitikdengan Tanaman Inang
Untuk mengetahui kemampuan eks-presi enzim nitrogenase, maka diuji de-
lapan isolat bakteri endofitik padi untuk ujiautentikasi pada tanaman padi IR64. Isolatyang diuji autentikasi ialah BCr 1.2, BCr 2.3,BCbd 1.3, BLw 3.2, APK 2.1, APK 2.4, BRb2.2, dan BCr 2.1. Hasil analisis reduksi ase-
tilen di bagian akar dan batang tanamanpadi IR64 disajikan pada Tabel 111.99.
Isolat yang mampu mengekspresikanenzim nitrogenase di akar ialah APK 2.1,BCr 1.2, dan BCR 2.1 sedangkan APK 2.4,BRb 2.2, BCr 2.3, dan BCR 2.1 mampu
mengekspresikan enzim nitrogenase di batang dan BCr 2.1 mampu mengekspresikanenzim nitrogenase baik di akar maupun dibatang.
Hasil Penelitian 2002
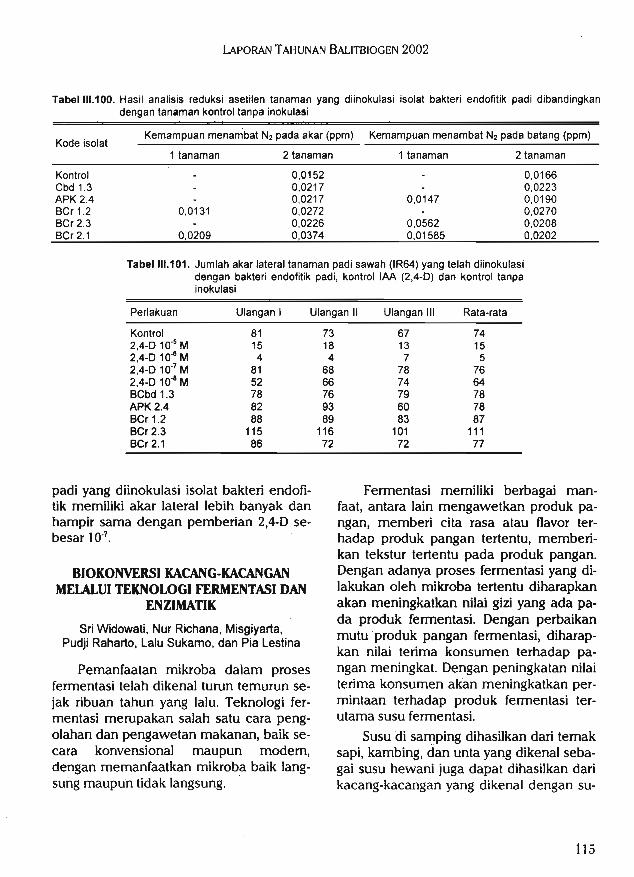
115
Fermentasi memiliki berbagai man-faat, antara lain mengawetkan produk pa-ngan, memberi cita rasa atau flavor ter-hadap produk pangan tertentu, memberi-kan tekstur tertentu pada produk pangan.
Dengan adanya proses fermentasi yang di-lakukan oleh mikroba tertentu diharapkanakan meningkatkan nilai gizi yang ada pada produk fermentasi. Dengan perbaikanmutu produk pangan fermentasi, diharapkan nilai terima konsumen terhadap pangan meningkat. Dengan peningkatan nilaiterima konsumen akan meningkatkan per-mintaan terhadap produk fermentasi ter-utama susu fermentasi.
Susu di samping dihasilkan dari temaksapi, kambing, dan unta yang dikenal seba-gai susu hewani juga dapat dihasilkan darikacang-kacangan yang dikenal dengan su-
padi yang diinokulasi isolat bakteri endofitik memiliki akar lateral lebih banyak danhampir sama dengan pemberian 2,4-D se-besar 10"7.
BIOKONVERSI KACANG-KACANGANMELALUI TEKNOLOGI FERMENTASI DAN
ENZIMATIKSri Widowati, Nur Richana, Misgiyarta,
Pudji Raharto, Lalu Sukarno, dan Pia Lestina
Pemanfaatan mikroba dalam prosesfermentasi telah dikenal turun temurun se-
jak ribuan tahun yang lalu. Teknologi fermentasi merupakan salah satu cara peng-olahan dan pengawetan makanan, baik se-cara konvensional maupun modem,
dengan memanfaatkan mikroba baik lang-sung maupun tidak langsung.
771118778786476
51574
Rata-rata
721018360797478
71367
Ulangan III
721168993766668
41873
Ulangan II
86115
8882785281
41581
Ulangan I
BCr 2.1BCr 2.3BCr 1.2APK 2.4BCbd1.32,4-D 10"8M2,4-D 10"7M2,4-D 10-"M2,4-D 10s MKontrol
Perlakuan
Tabel 111.101. Jumlah akar lateral tanaman padi sawah (IR64) yang telah diinokulasidengan bakteri endofitik padi, kontrol IAA (2,4-D) dan kontrol tanpainokulasi
0,02020,02080,02700,01900,02230,0166
2 tanaman
N2 pada batang (ppm)
0,015850,0562
-0,0147
_
1 tanaman
Kemampuan menambat
0,03740,02260,02720,02170,02170,0152
2 tanaman
pada akar (ppm)
0,0209-
0,0131
_
1 tanaman
Kemampuan menambat N2
BCr 2.1BCr 2.3BCr 1.2APK 2.4Cbd 1.3Kontrol
Kode isolat
Tabel 111.100. Hasil analisis reduksi asetilen tanaman yang diinokulasi isolat bakteri endofitik padi dibandingkandengan tanaman kontrol tanpa inokulasi
Laporan Tahunan Balitbiogen 2002

116
selulosa merupakan komponen terbesar
yang sudah dimanfaatkan untuk industripertanian. Sedangkan untuk hemiselulosa
belum banyak dimanfaatkan atau digalipotensinya untuk industri pertanian.
Xilanase merupakan enzim ekstra se-
luler yang dapat menghidrolisis xilan menjadi xilosa dan xilo-oligosakarida. Xilanasedimanfaatkan untuk proses pemutih ker-tas, campuran pakan temak, penjernihansirup, pembuatan gula xilosa, dan sebagai-
nya. Penggunaan xilanase untuk mengu-
rangi pemakaian khlorin dalam pemutihkertas, telah memberikan peluang untuk
aplikasi bioteknologi dan sekarang telah digunakan pada beberapa pabrik kertas.
Produksi xilanase skala industri mem-butuhkan tersedianya substrat yang murahdan mudah didapat. Untuk mengatasi ma-
salah tersebut, perlu dicari bahan baku berlignoselulosa seperti limbah dari tanamanpangan di antaranya ialah kulit kedelai.Limbah kacang-kacangan tersebut digunakan sebagai sumber karbon pada mediakultivasi bakteri, sehingga diharapkan akanmeningkatkan nilai guna dan ekonominya.
Isolat Mikroba Asam Laktat
Isolasi dilakukan terhadap berbagaisampel yang dianggap sebagai sumberpotensial mikroba yang dimaksud terutamabakteri asam laktat. Sampel yang diperolehsebagai sumber bakteri asam laktat ialahkubis busuk, wortel busuk, sawi busuk, ka-
cang panjang busuk, selada busuk, tomatbusuk, timun busuk, sauerkrout, asinan sawi, asinan lobak, asinan wortel, feses bayi,feses sapi, susu sapi asam, susu kedelai,
pisang busuk, jambu biji busuk, pepaya busuk, nanas busuk, dan sirsak busuk (TabelIII.102).
Hasil isolasi bakteri asam laktat dariberbagai sumber diperoleh 141 isolat
su nabati. Susu nabati yang banyak ditemu-
kan di masyarakat dan di pasaran ialah susu kedelai, sedangkan susu kacang-ka-cangan lain belum banyak ditemukan.Dengan demikian, perlu terobosan untuk
membuat susu nabati dari kacang-kacang-
an lain serta menganalisis nilai gizi yangterdapat pada susu kacang-kacangan ter-
sebut.
Mikroba yang digunakan untuk prosesfermentasi susu ialah bakteri asam laktat.Bakteri asam laktat telah diteliti dan diko-leksi oleh peneliti dan praktisi industri didalam dan luar negeri. Namun demikian,
eksplorasi bakteri asam laktat yang banyakterdapat di alam Indonesia perlu dilakukanuntuk menambah koleksi mikroba sehing-ga dapat dimanfaatkan lebih lanjut. Bakteriasam laktat yang banyak tersebar di Indonesia dapat diisolasi dari berbagai sumber,buah-buahan busuk, sayur busuk, produkasinan tradisional, susu terfermentasi, feses
temak, feses bayi, dan Iain-lain.
Perkembangan dan kemajuan bidangpertanian dan industri pertanian di Indonesia telah menimbulkan peningkatan limbahpertanian yang sebagian besar merupakan
limbah berlignoselulosa. Limbah lignoselu-losa sebagai bahan organik memiliki po-tensi besar sebagai bahan baku berbagaiindustri, antara lain untuk pembuatan ker-tas, kompos, dan fraksinasi menjadi kom-ponen penyusunnya. Pemanfaatan limbah
berlignoselulosa dengan menggunakan ja-sa mikroorganisme dapat menghasilkanenzim ekstraseluler yang mampu mendeg-
radasi bahan berlignoselulosa menjadifraksi penyusunnya.
Sebagian besar limbah tanaman pa-ngan mengandung lignoselulosa atau fraksiseratnya. Ada tiga komponen fraksi serat,yaitu selulosa, hemiselulosa, dan lignin.Dari ketiga komponen fraksi serat tersebut
Hasil Penelitian 2002

117
141
15611
10274
1049
119
1114
.72-
Jumlah isolat
WTBTMBTbSUS, SaSUKSSBSLBSWBSkPSBPPBNNB PR, NbKBBKPBJBBFSSFSB, FASWASSASL
Kode isolat
Jumlah total
WortelTomatTimunSusu sapi asamSusu kedelaiSirsakSeladaSawiSauerkrautPisangPepayaNanasKubisKacang panjangJambu bijiFeses sapiFeses bayiAsinan wortelAsinan sawiAsinan lobak
Nama sampel
20.19.18.17.16.15.14.13.12.11.10.9.8.7.6.5.4.3.2.1.
No.
Tabel 111.103. Jumlah isolat yang diperoleh dari berbagai sumber isolat
BusukBusukBusukAsamKontaminasiBusukBusukBusukAsamBusukBusukBusukBusukBusukBusukBusukBusukSegarSegarSegar
WTBTMBPRTbSUS, SaSUKSSBSLBSWBSkPSBPPBNNB PR, NbKBBKPBJBBFSSFSB, FASWASSASL
Pasar Cempaka Putih, JakartaPasar Ramayana, BogorBogorMangga Dua, JakartaMangga Dua, JakartaPasar Anyar, BogorPasar Anyar, BogorPasar Cempaka Putih, JakartaBogorPasar Cempaka Putih, JakartaPasar Cempaka Putih, JakartaPasar Ramayana, BogorPasar Anyar, BogorPasar Cempaka Putih, JakartaPasar Anyar, BogorMangga Dua, JakartaCempaka Putih, JakartaSuka Sari, BogorSuka Sari, BogorSuka Sari, Bogor
WortelTomatTimunSusu sapi asamSusu kedelaiSirsakSeladaSawiSauerkrautPisangPepayaNanasKubisKacang panjangJambu bijiFeses sapiFeses bayiAsinan wortelAsinan sawiAslnan lobak
20.19.18.17.16.15.14.13.12.11.10.9.8.7.6.5.4.3.2.1.
KeteranganKode isolatAsal sampelNo. Nama sampel
Pengujian kemampuan bakteri asamlaktat dalam menghasilkan asam laktatselama proses fermentasi menggunakan
pembanding Lactobacillus bulgaricus dan
Tabel 102. Sumber isolat dan lokasi asal sumber isolat
(Tabel III. 103). Dari sirsak busuk berhasildiisolasi 27 isolat, sawi busuk 10 isolat, dankacang panjang busuk 14 isolat.
Laporan Tahunan Balitbiogen 2002

118
substrat tersebut dimanfaatkan untuk per-tumbuhan bakteri asam laktat.
Pemanfaatan gula yang ada dalamsubstrat untuk pertumbuhan bakteri asamlaktat akan terlihat dengan meningkatnya
0,800,800,800,800,85
Persentase asam laktat
NNBPR5F3Nb3KBB3SLB22
Kode isolat
fermentasi susu kacang-kacanganBAL yang digunakan dalam
0,500,550,550,550,600,600,600,600,600,600,600,600,600,600,600,600,600,600,700,700,700,700,700,700,700,750,750,800,800,800,800,800,85
Persentase asam laktat
111.105. Lima isolat
L. caseiL bulgaricusSSBA5NNBPR6F2Nb2SUKA1SUK3SSBA8SSB12PSB4PPB9KPB6KPB4KPB12KBB9KBB8FSS3WTB5WTB4SWB4SUK9PSB6PPB7SUK2SWB2TMB5SSBA6NNBPR5F3Nb3KBB3SLB22
Kode isolat
5.4.3.2.1.
No.
Tabel
33.32.31.30.29.28.27.26.25.24.23.22.21.20.19.18.17.16.15.14.13.12.11.10.
9.8.7.6.5.4.3.2.1.
No.
Tabel 111.104. Daftar isolat yang memiliki kemampuanunggul dalam menghasilkan asam laktat
L casei. Bakteri asam laktat hasil isolasi di-
katakan memiliki kemampuan unggul apa-bila mampu menghasilkan asam laktat se-lama proses fermentasi menggunakan me
dia yang sama, yaitu MRS broth dan kondisifermentasi sama. Hasil pengujian kemam
puan bakteri asam laktat dalam menghasilkan asam laktat diperoleh 31 isolat bakteriasam laktat unggul (Tabel III. 104).
Isolat bakteri asam laktat yang berhasildiisolasi dan diidentifikasi, disimpan padamedia MRS agar dapat digunakan untukfermentasi susu, terutama susu kacang-
kacangan. Lima isolat bakteri asam laktatterbaik diuji kemampuan memfermentasisusu kacang hijau, kacang merah, kacangtunggak, kacang tanah, dan kedelai. Ke-
lima isolat bakteri asam laktat terbaik di-sajikan pada Tabel III. 105. Sebagai kontrolpembanding digunakan bakteri asam laktatkomersial, yaitu L. bulgaricus.
Fermentasi utama dilakukan setelahdiperoleh isolat-isolat bakteri asam laktatyang paling efektif untuk fermentasi susukacang-kacangan.
Selama proses fermentasi dapat di-
amati berbagai dinamika fermentasi diantaranya ialah kadar asam laktat, kadargula reduksi, kadar protein, pH, dan kera-
patan sel bakteri asam laktat yang digunakan untuk proses fermentasi. Bakteri asam
laktat memanfaatkan gula glukosa yangada dalam media fermentasi untuk per-
tumbuhannya. Hal ini dapat dilihat dariperubahan gula reduksi dalam substrat fermentasi. Perubahan ini karena gula reduksidigunakan oleh bakteri asam laktat untukpertumbuhannya. Dengan demikian, kadar
gula reduksi dalam substrat akan semakinberkurang atau menurun konsentrasinya.
Pada Gambar III.26, terlihat adanya pe-nurunan kadar gula reduksi dalam mediafermentasi karena gula yang ada dalam
Hasil Penelitian 2002
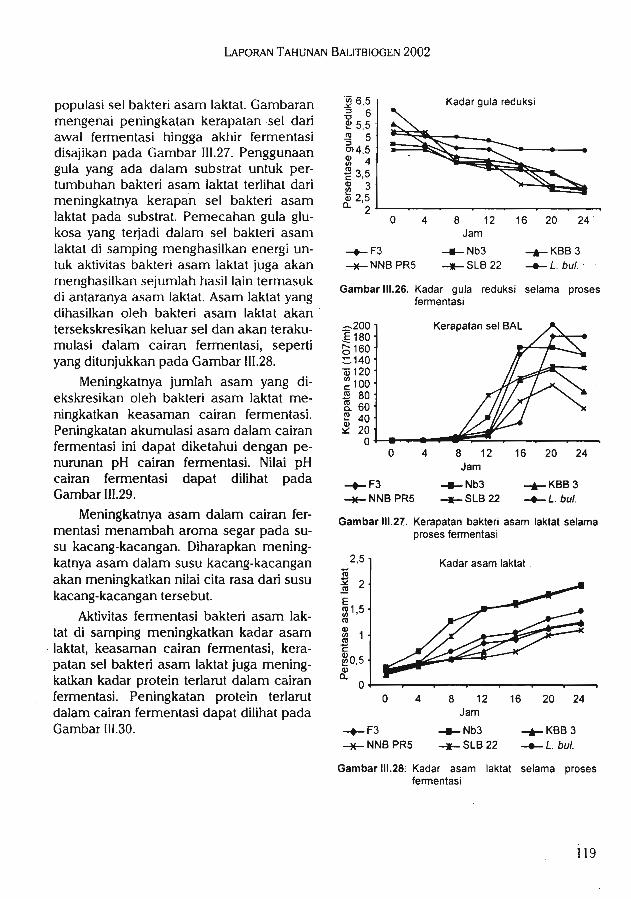
119
Gambar III.28. Kadar asam laktat selama prosesfermentasi
24
-?- L. bul.-4-KBB 3
20 ^16
-X- SLB 22-•-Nb3
Jam8 12
PR5
4
NNBF3
0
Kadar asam laktat
ICDJ'l|o,5
a.
2,5
BS 2
Gambar III.27. Kerapatan bakteri asam laktat selamaproses fermentasi
-K-SLB22 -%-Lbul.__ Nb3 -^- KBB 3
Jam4 8 12 16 20 24m , ^ir w
/sv/J////I ^
Kerapatan sel BAL A.
-X- NNB PR5-?-F3
U H—^—•—0—r\ <
^ 20 'E 402. 60B 80^ 100V20^140
160E180—200
Gambar III.26. Kadar gula reduksi selama prosesfermentasi
16 20 24
-^-KBB3-•- L. bul.
8 12Jam
I-Nb3*-SLB22
F3NNB PR5
Kadarula reduksi•g 6
5,51 5o>4,5
4f 3.5^ 3
2,5a 2
populasi sel bakteri asam laktat. Gambaran
mengenai peningkatan kerapatan sel dariawal fermentasi hingga akhir fermentasidisajikan pada Gambar III.27. Penggunaangula yang ada dalam substrat untuk per-
tumbuhan bakteri asam laktat terlihat darimeningkatnya kerapan sel bakteri asamlaktat pada substrat. Pemecahan gula glu-
kosa yang terjadi dalam sel bakteri asamlaktat di samping menghasilkan energi untuk aktivitas bakteri asam laktat juga akanmenghasilkan sejumlah hasil lain termasukdi antaranya asam laktat. Asam laktat yang
dihasilkan oleh bakteri asam laktat akantersekskresikan keluar sel dan akan teraku-mulasi dalam cairan fermentasi, seperti
yang ditunjukkan pada Gambar III.28.
Meningkatnya jumlah asam yang di-ekskresikan oleh bakteri asam laktat me-ningkatkan keasaman cairan fermentasi.
Peningkatan akumulasi asam dalam cairanfermentasi ini dapat diketahui dengan pe-nurunan pH cairan fermentasi. Nilai pH
cairan fermentasi dapat dilihat padaGambar 111.29.
Meningkatnya asam dalam cairan fermentasi menambah aroma segar pada su-su kacang-kacangan. Diharapkan meningkatnya asam dalam susu kacang-kacangan
akan meningkatkan nilai cita rasa dari susukacang-kacangan tersebut.
Aktivitas fermentasi bakteri asam laktat di samping meningkatkan kadar asamlaktat, keasaman cairan fermentasi, kera
patan sel bakteri asam laktat juga meningkatkan kadar protein terlarut dalam cairanfermentasi. Peningkatan protein terlarut
dalam cairan fermentasi dapat dilihat padaGambar 111.30.
Laporan Tahunan Balitbiogen 2002
120
semakin segar dengan terbentuknya aroma
asam serta memiliki warna yang beragam.Di samping perubahan fisik, susu kacang-kacangan yang telah mengalami fermentasi oleh bakteri asam laktat juga mengalami perubahan kimiawi dengan terbentuknya asam laktat, serta asam lain, me-ningkatnya kadar protein. Dengan demi-kian, proses fermentasi susu kacang-
kacangan juga dapat meningkatkan nilaigizi pada susu asam yang berbahan dasarkacang-kacangan.
Perubahan tekstur, aroma, rasa serta
peningkatan nilai gizi pada susu asam yangberbahan dasar kacang-kacangan diharap-
kan akan menjadi terobosan teknologipengolahan susu kacang-kacangan, me
ningkatkan keragaman cara pengolahansusu kacang-kacangan, meningkatkan pe-
nerimaan produk yang berbahan dasarkacang-kacangan oleh masyarakat, serta
memberikan nilai tambah terhadap susuasam yang berbahan dasar kacang-kacangan. Untuk mengetahui penerimaan masya
rakat terhadap susu asam hasil fermentasisusu kacang-kacangan dengan isolat bak
teri asam laktat, maka dilakukan uji orga-noleptik terhadap susu asam tersebut.
Uji organoleptik dilakukan oleh 15orang panelis. Peubah yang diamati ialahtekstur, aroma, warna, dan rasa. Sebagai
pembanding digunakan yoghurt merekYummy (merupakan produk yoghurtkomersial yang diproduksi oleh industri diJakarta). Hasil uji Organoleptik disajikanpada Tabel III. 107.
Dari uji organoleptik ini diketahui bah-wa kesukaan panelis terhadap susu asam
berasal dari bahan dasar kacang merah,kacang hijau, dan kacang tunggak tidakberbeda nyata terhadap yoghurt komersial,sedangkan susu asam yang berbahan darikacang tanah dan kedelai berbeda nyata
Gambar 111.30. Kadar protein cairan fermentasiselama proses fermentasi
Fermentasi susu kacang-kacangan
menggunakan bakteri asam laktat lokalmampu memberikan cita rasa dan flavoryang khas terhadap susu kacang terfermen-tasi. Di samping itu, fermentasi susu kacang-
kacangan oleh bakteri asam laktat lokalmampu meningkatkan nilai gizi susu terfer-mentasi tersebut. Peningkatan nilai gizi yangterjadi selama proses fermentasi susu kacang-kacangan oleh berbagai isolat bakteriasam laktat disajikan pada Tabel III. 106.
Susu asam hasil fermentasi oleh bakteri asam laktat lokal mengalami perubah-an tekstur menjadi lebih lembut, aroma
t-KBB3\-Lbul.
20 24
I i
MK
^^
-•-Nb3 -iHK-SLB22 -t
8 12 16Jam
Kadar protein ..
pH selama proses fermentasi
^-KBB3^-Lbul.
20 24
^•^^*—t •
-*-Nb3 h-#-SLB22 _,
Jam
8 12 16
pH Fermentasi
-?-F3-*- NNB PR5
0 4
M•*-—•
I 5S 4'
121110
o 8a. 78! 6
Gambar 111.29. Nilai
-?-F3-*- NNB PR5
0 4
N
4A
4,5
X5.5
5
6
6,5
7
Hasil Penelitian 2002
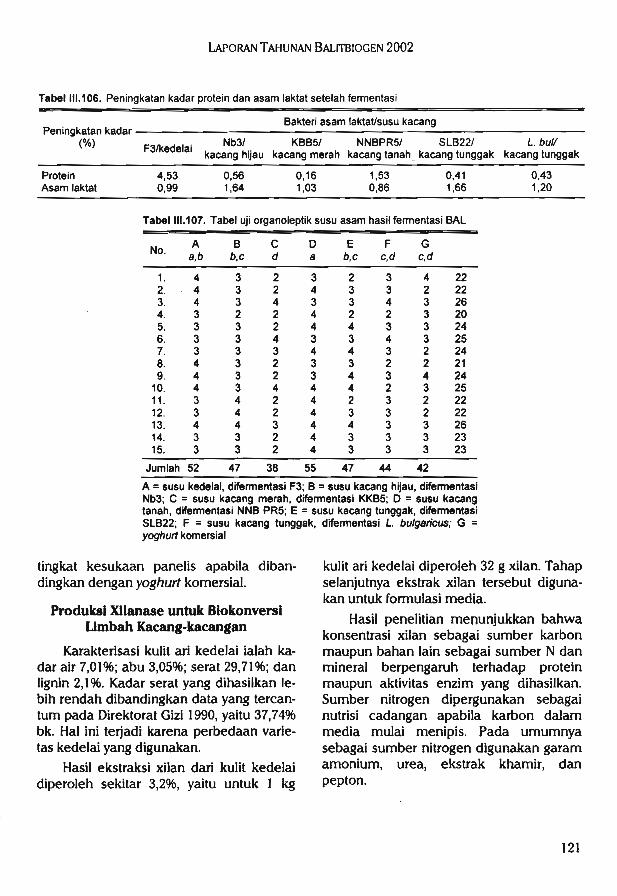
121
kulit ari kedelai diperoieh 32 g xilan. Tahapselanjutnya ekstrak xilan tersebut digunakan untuk formulasi media.
Hasil penelitian menunjukkan bahwakonsentrasi xilan sebagai sumber karbonmaupun bahan lain sebagai sumber N danmineral berpengaruh terhadap proteinmaupun aktivitas enzim yang dihasilkan.Sumber nitrogen dipergunakan sebagainutrisi cadangan apabila karbon dalammedia mulai menipis. Pada umumnyasebagai sumber nitrogen digunakan garamamonium, urea, ekstrak khamir, dan
pepton.
tingkat kesukaan panelis apabila diban-dingkan dengan yoghurt komersial.
Produksi Xilanase untuk BiokonversiUmbah Kacang-kacangan
Karakterisasi kulit ari kedelai ialah kadar air 7,01%; abu 3,05%; serat 29,71%; danlignin 2,1%. Kadar serat yang dihasilkan le-bih rendah dibandingkan data yang tercan-tum pada Direktorat Gizi 1990, yaitu 37,74%bk. Hal ini terjadi karena perbedaan varie-tas kedelai yang digunakan.
Hasil ekstraksi xilan dari kulit kedelaidiperoleh sekitar 3,2%, yaitu untuk 1 kg
A = susu kedelai, difermentasi F3; B = susu kacang hijau, difermentasiNb3; C = susu kacang merah, difermentasi KKB5; D = susu kacangtanah, difermentasi NNB PR5; E = susu kacang tunggak, difermentasiSLB22; F - susu kacang tunggak, difermentasi L bulgaricus; G =yoghurt komersial
232326222225242124252420262222
42333223422333324
c.dG
44333332323432433
c,dF
i hasil fermentasi BAL
1,66 1,200,41 0,43
kacang tunggak kacang tunggakSLB22/ L. but/
0,861,53
47
334324434342332
b,cE
55444444334344343
aD
38223224223422422
dC
47334443333332333
b,cB
Tabel uji organoleptik susu asarr
1,030,16
kacang merah kacangtanahNNBPR5/KBB5/
Bakteri asam laktat/susu kacangI
1,640,56
52334334443333444
a,bA
.107.
Jumlah
15.14.13.12.11.10.9.8.7.6.5.4.3.2.1.
INO.Kin
Tabel III
Asam laktat 0,99Protein 4,53
F3/kedeb, kacang hijauNb3/
Ui
Tabel 111.106. Peningkatan kadar protein dan asam laktat setelah fermentasi
Laporan Tahunan Balitbiogen 2002

122
Ekstrak khamir berpengaruh terhadappertumbuhan isolat bakteri dan hasilmetabolisme skundernya. Semakin banyak
ekstrak khamir yang ditambahkan akanmeningkatkan aktivitas spesifik enzim.Penambahan ekstrak khamir terbaik ialah0,3%.
Xilan sebagai sumber karbon berpengaruh terhadap pertumbuhan isolat bakteri. Peningkatan xilan diikuti oleh pening-katan aktivitas spesifik enzim. Komposisimedia terbaik ialah polipepton 0,1%;ekstrak khamir 0,3%; dan xilan 1%. Hasil inidigunakan untuk penelitian berikutnya.
Hasil optimasi proses, yaitu pengaruhsuhu, pH, dan kecepatan shaker terhadappertumbuhan biomassa, aktivitas enzim,
maupun protein terlarut disajikan padaTabel III. 109, III. 110, dan III. 111. Pada kecepatan shaker 120 rpm temyata kenaikansuhu diiringi oleh kenaikan biomassa. Pengaruh pH terhadap biomassa, menunjuk-kan pada suhu 35 dan 40C, pH 7 merupa-kan pH optimum untuk menghasilkan biomassa. Sedangkan pada suhu 40 dan 45CpH 8 merupakan pH optimum untuk menghasilkan biomassa.
Pada suhu 35C, pH optimum untukmenghasilkan biomassa adalah pH 7,sedangkan pada suhu 40, 45, dan 50C pHoptimumnya adalah pH 8.
Pengamatan protein terlarut dari bebe-rapa kondisi pH, suhu, dan kecepatanshaker temyata tidak berbeda nyata. De-ngan demikian, berdasarkan pengamatanaktivitas enzim dan protein terlarut makahasil perhitungan aktivitas spesifik hampirsama dengan dengan aktivitas enzimnya.
Hasil terbaik berdasarkan aktivitas spesifikenzim, yaitu pada kondisi kecepatanshaker 120 rpm, suhu 35C, dan pH 9 yaitu612,14 U/mg protein.• .
Ekstrak khamir mengandung asamamino, peptida, vitamin, dan karbohidrat.
Komposisi penggunaan ekstrak khamiryang tepat sangat diperlukan dalam proseskultivasi. Penggunaan ekstrak khamir yangcukup tinggi dan adanya pengadukan/penggoyangan pada saat kultivasi menye-
babkan timbulnya buih. Hal tersebut tidakdiinginkan, karena akan menurunkan akti-
vitas enzim yang dihasilkan.
Aktivitas enzim sangat dipengaruhioleh formulasi dan komponen media, de-
ngan aktivitas terkecil 26,71 Umol/menitdan terbesar 186,37 Umol/menit. Hasil ana-lisis statistik temyata penambahan polipep-ton maupun ekstrak khamir akan menurunkan aktivitas enzim. Penambahan xilanakan meningkatkan aktivitas enzim. For
mulasi media terbaik berdasarkan aktivitasenzim ialah polipepton 0,1%; ekstrakkhamir 0,2%; dan xilan 1%.
Protein terlarut dari perlakuan formulasi media berkisar antara 187-596 mg/1.Hasil optimum pada komposisi polipepton0,5%, ekstrak khamir 0,1% dan xilan 1%.
Protein terlarut dapat ditingkatkan de-ngan penambahan sumber N dalam hal iniialah polipepton. Semakin tinggi polipeptonsemakin tinggi protein terlarut yang dihasilkan. Protein terlarut dalam penelitian ini di-asumsikan enzim (xilanase) yang dihasilkan. Temyata hasil tersebut tidak diikutioleh hasil aktivitas xilanasenya. Hal tersebut dapat dilihat dari hasil kondisi optimumberdasarkan aktivitas enzim tertinggi(144,44 Umol/menit), yaitu pada polipepton0,1% dan ekstrak khamir 0,2% (aktivitasenzim), sedangkan komposisi optimumberdasarkan hasil protein tertinggi (aktivitas enzim 113,23 Umol/menit) (Tabel111.108).
Hasil Penelitian 2002

123
86,64158,14436,45
85,79313,13219,66336,29303,13170,98180,43190,65350,63216,91193,26165,72170,62209,08207,70228,72206,99212,19156,92331,51278,43233,86204,42201,64203,67180,13178,67193,41180,64342,36307,71334,93397,96
0,3110,3400,4270,3150,3850,3490,4080,3700,4100,3170,3630,4180,4350,5480,4580,4170,5960,5010,4520,3990,2550,1870,2580,1870,4910,4610,4690,5140,6240,5400,4860,5900,2690,1950,2500,24296,13
84,0266,6692,07
102,5192,0396,62
112,42104,39
93,5192,28
113,2351,9285,8129,2353,0478,0392,63
101,47121,4271,5574,67
105,1191,27
144,4467,8255,6768,55
113,02127,44
76,34119,9241,63
186,3753,3626,71
Aktivitas enzim (
36.35.34.33.32.31.30.29.28.27.26.25.24.23.22.21.20.19.18.17.16.15.14.13.12.11.10.9.8.7.6.5.4.3.2.1.
Kode perlakuan
Tabel 111.108. Hasil pengamatan aktivitas enzim, protein terlarut, dan aktivitas spesifik
^^nenit) Protein terlarut (g/l) Aktivitas spesifik (U/g protein)
Laporan Tahunan Balitbiogen 2002
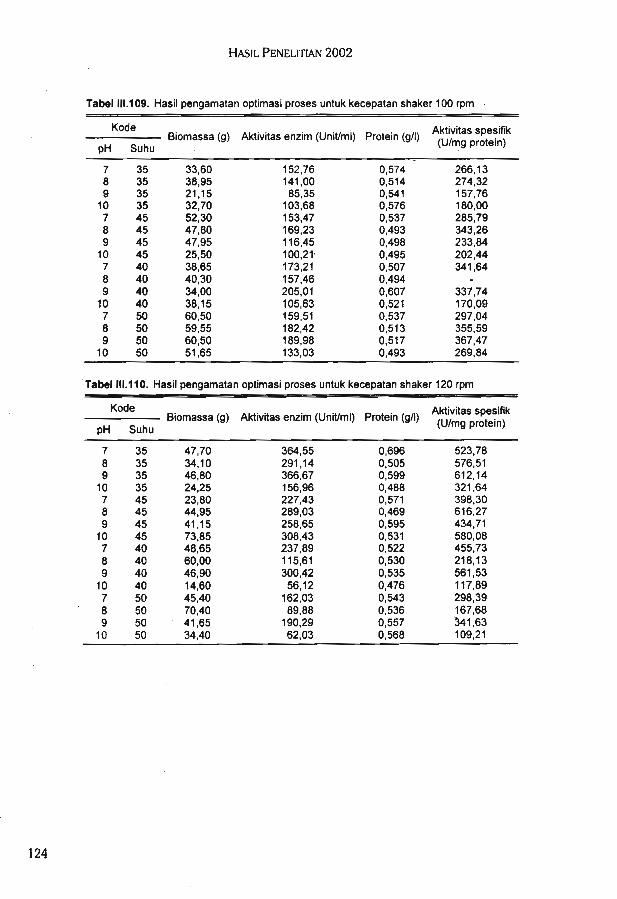
124
109,21341,63167,68298,39117,89561,53218,13455,73580,08434,71616,27398,30321,64612,14576,51523,78
(U/mg protein)Aktivitas spesifik
0,5680,5570,5360,5430,4760,5350,5300,5220,5310,5950,4690,5710,4880,5990,5050,696
Protein (g/l)
62,03190,2989,88
162,0356,12
300,42115,61237,89308,43258,65289,03227,43156,96366,67291,14364,55
Aktivitas enzim (Unit/ml)
optimasi proses untuk kecepatan shaker 120 rpm
269,84367,47355,59297,04170,09337,74
.
341,64202,44233,84343,26285,79180,00157,76274,32266,13
(U/mg protein)Aktivitas spesifik
0,4930,5170,5130,5370,5210,6070,4940,5070,4950,4980,4930,5370,5760,5410,5140,574
Protein (g/l)
133,03189,98182,42159,51105,63205,01157,46173,21100,21116,45169,23153,47103,68
85,35141,00152,76
Aktivitas enzim (Unit/ml)
34,4041,6570,4045,4014,6046,9060,0048,6573,8541,1544,9523,8024,2546,8034,1047,70
- Biomassa (g)
50505050404040404545454535353535
Suhu
10987
10987
10987
10987
PH
Kode
11.110. Hasil pengamatan
51,6560,5059,5560,5038,1534,0040,3038,6525,5047,9547,8052,3032,7021,1538,9533,60
- Biomassa (g)
50505050404040404545454535353535
Suhu
Tabel 1
10987
10987
10987
10987
pH
Kode
Tabel 111.109. Hasil pengamatan optimasi proses untuk kecepatan shaker 100 rpm
Hasil Penelitian 2002
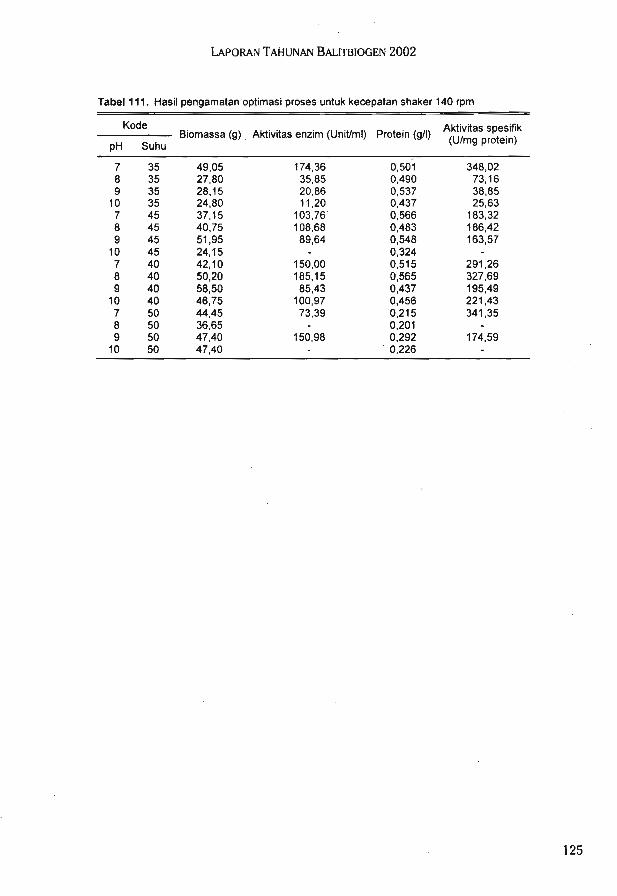
125
-
174,59-
341,35221,43195,49327,69291,26
.
163,57186,42183,3225,6338,8573,16
348,02
(U/mg protein)Aktivitas spesifik
0,2260,2920,2010,2150,4560,4370,5650,5150,3240,5480,4830,5660,4370,5370,4900,501
Protein (g/l)
-
150,98-
73,39100,9785,43
185,15150,00
.
89,64108,68103,76
11,2020,8635,85
174,36
Aktivitas enzim (Unit/ml)
47,4047,4036,6544,4546,7558,5050,2042,1024,1551,9540,7537,1524,8028,1527,8049,05
Biomassa (g)
50505050404040404545454535353535
Suhu
10987
10987
10987
10987
PH
Kode
Tabel 111. Hasil pengamatan optimasi proses untuk kecepatan shaker 140 rpm
Laporan Tahunan Balitbiogen 2002

126
8.Keragaman Genetik Strain Ralstoniasolanacearum berdasarkan Karak-
terisasi Menggunakan Teknik Berba-sis Asam Nukleat (Yadi Suryadi danM. Machmud)
9.Penyakit Hawar Daun (Phytophthorainfestans (Mont.) de Bary) pada Ken-tang dan Tomat: Identifikasi Per-masalahan di Indonesia (HaeniPurwanti)
?Warta Balitbio nomor 18, 19, dan 20,memuat artikel atau berita yang berkait-
an dengan penelitian bioteknologi danrumusan maupun kegiatan-kegiatan
yang ada di Balitbiogen?Laporan tahunan, memuat hasil pene
litian serta kegiatan pendukungnya
?Presiding Seminar Hasil PenelitianRintisan dan Bioteknologi Tanaman,memuat kumpulan makalah hasil penelitian tahun anggaran 2001 yang telahdipresentasikan pada Seminar HasilPenelitian Rintisan dan BioteknologiTanaman.
?Leaflet Bio-Lestari, Bioaktivator Perom-
bak Limbah Pertanian/Orlitan, Tekno-logi Produksi Bibit Abaka melalui KulturJaringan, Kit ELISA untuk Deteksi DiniPeanut Stripe Virus, dan NPS Biopesti-sida Unggulan
APRESIASI PROGRAM PENELITIANBIOTEKNOLOGI DAN SUMBER DAYA
GENETIK PERTANIANDalam rangka sinkronisasi program
penelitian bioteknologi pertanian BalaiPenelitian Bioteknologi dan SumberdayaGenetik Pertanian menyelenggarakan
Komunikasi, memegang peranan pen-
ting dalam upaya penyebarluasan hasilpenelitian. Selama TA 2002, Balitbiogenmenyelenggarakan berbagai kegiatandalam upaya penyebarluasan hasil penelitian. Kegiatan tersebut meliputi penerbitan,seminar, apresiasi, pameran/peragaan, dan
pelayanan informasi
PUBLIKASI DAN LAPORAN
Selama TA 2002, publikasi dan laporanyang ditetbitkan oleh Balitbiogen terdiridari:? Buletin Agrobio Vol. 5, No. 1 dan No. 2,
memuat artikel tinjauan ilmiah hasilpenelitian bidang biologi dan biotekno-logi tanaman. Dalam buletin inidisajikan 9 makalah, yaitu
1.Perakitan Tanaman Tahan SeranggaHama melalui Teknik RekayasaGenetik (Muhammad Herman)
2.Wild Rice (Oryza spp.): TheirExistence and Research in Indonesia(Ida Hanarida Somantri)
3.Penggunaan Bacillus thuringiensissebagai Bioinsektisida (Bahagiawati)
4.Produksi dan Prospek Enzim Xilan-ase dalam Pengembangan Bioindus-tri di Indonesia (Nur Richana)
5.Perkembangan Penelitian Regenera-si dan Transformasi pada TanamanKedelai (Saptowo J. Pardal)
6.Perkembangan Penelitian Kultur InVitro pada Tanaman Industri, Pa-ngan, dan Hortikultura (Ika Mariska)
7.Regenerasi Tanaman melalui Em-
briogenesis Somatik dan BeberapaGen yang Mengendalikannya (Raga-padmi Purnamaningsih)
IV. Penyebarluasan Hasil Penelitian
Laporan Tahunan Balitbiogen 2002© Copyright 2003, Balitbiogen

127
nologi IPB, Puslitbang Bioteknologi BPPT,serta Pusat Penelitian dan PengembanganTeknoiogi Isotop dan Radiasi BATAN.
PAMERAN DAN DEBAT PENDAPATBIOTEKNOLOGI
Teknoiogi rekayasa genetik mempu-nyai potensi yang sangat besar sebagai teknoiogi pelengkap dan pendukung pemuiia-an konvesional seandainya sumber geninteres yang ingin disilangkan tidak adaatau belum dijumpai di dalam koleksi plasma nutfah. Rekayasa genetik merupakanwahana baru bagi para pemulia untukmemperoleh gen bam, yang tidak lagi di-batasi oleh adanya hubungan kekerabatan.
Upaya untuk memperoleh tanamantransgenik tahan cekaman abiotik sepertitoleran kekeringan, suhu dingin, salinitastinggi, dan keracunan aluminium ataukeracunan besi telah dirintis oleh berbagailembaga penelitian di dalam dan luarnegeri. Rekayasa genetik membuka ke-mungkinan introduksi sifat bam ke varietasyang sudah ada seperti halnya padi yangmengandung vitamin A yang tinggi. Melaluirekayasa genetika telah dihasilkan berbagai tanaman transgenik yang memiliki sifatbam seperti ketahanan terhadap hama danpenyakit, toleran herbisida, serta pening-katan kualitas hasil.
Luas area! tanaman transgenik terns
meningkat pesat sejak tanaman transgenikdiintroduksi ke lapang. Selama periodeenam tahun dari 1996 sampai 2001, telahterjadi peningkatan luas areal pertanamanyang tajam, yaitu luas yang hanya 1,7 jutaha pada tahun 1996 menjadi 52,6 juta hapada tahun 2001 atau terjadi peningkatanlebih dari 30 kali dibandingkan periodeawai peiepasan tanaman transgenik. Adadelapan jenis tanaman transgenik yang
Apresiasi Program Penelitian Bioteknologidan Sumber Daya Genetik Pertaniandengan mengundang balai-balai komoditasdi Iingkup Badan Litbang Pertanian sertainstansi di luar Badan Litbang Pertanian.
Apresiasi diselenggarakan selama duahari, pada tanggal 17-18 September 2002dan dihadiri sekitar 100 peserta dari BalaiPenelitian Bioteknologi dan SumberdayaGenetik Pertanian, Unit Penelitian Bioteknologi Perkebunan, Balai Penelitian Ta-naman Padi, Balai Penelitian TanamanKacang-kacangan dan Umbi-umbian, BalaiPenelitian Serealia, Balai KesuburanTanah, Balai Penelitian Lahan Rawa, LokaPenelitian Pencemaran Lingkungan Pertanian, Balai Penelitian Tanaman Hias, BalaiPenelitian Tanaman Sayuran, Balai Penelitian Tanaman Buah, Balai Penelitian Pasca-panen, Balai Penelitian Tanaman Rempahdan Obat, Balai Penelitian Tanaman Tem-bakau dan Serat, Balai Penelitian Veteriner,Balai Penelitian Ternak, Pusat Penelitiandan Pengembangan Bioteknologi LIPI,Pusat Penelitian Bioteknologi IPB, PusatPenelitian dan Pengembangan Bioteknologi BPPT, dan Pusat Penelitian dan Pengembangan Teknoiogi Isotop dan RadiasiBATAN
Apresiasi ini dibuka oleh SekretarisBadan Litbang Pertanian, Dr. Djoko S.Damardjati dan diianjutkan dengan presen-tasi tentang (1) tugas dan fungsi Balitbio-gen yang berkaitan dengan balai komoditas, (2) program Balai Penelitian Bioteknologi dan Sumberdaya Genetik Pertaniandan Unit Penelitian Bioteknoiogi Perkebunan, dan (3) program balai komoditas yangterkait dengan program penelitian bioteknologi. Pada hari kedua, dipresentasikanprogram penelitian dari instansi di luarBadan Litbang Pertanian, yaitu PuslitbangBioteknologi LIPI, Pusat Penelitian Biotek
Laporan Tahunan Balttbiogen 2002

128
perusahaan swasta di Bogor yang bergerakdi bidang pengujian, yaitu PT SaraswantiIndo Genetech juga berpartisipasi dalampameran ini.
Seminar dua hari hanya membahasmakalah undangan yang dilaksanakan dalam bentuk diskusi panel. Pembicara dalam diskusi panel adalah pakar dalam bi-dangnya masing-masing yang berasal dariPusat Penelitian dan Pengembangan Bioteknologi LIPI Cibinong, Pusat Perlindung-an Varietas Tanaman, Universitas GadjahMada, Kementerian Lingkungan Hidup,Pusat Penelitian dan Pengembangan SosialEkonomi Pertanian, Institut PertanianBogor, dan Lembaga Swadaya Masyarakat.Mated yang disampaikan oleh masing-masing pembicara adalah sebagai berikut:1.Teknologi rekayasa genetik (Dr. Inez
H.S. Loedin, Puslitbang BioteknologiLIPI)
2.Ketentuan/peraturan-peraturan perun-dangan tentang KHKP PRG(Dr. Sugiono Moeljopawiro, PusatPerlindungan Varietas Tanaman)
3.Risiko tanaman transgenik ditinjau dariaspek lingkungan (Dr. Andi Trisyono,Universitas Gadjah Mada, Yogyakarta)
4.Manfaat tanaman transgenik ditinjaudari aspek lingkungan (Ir. B. SatyawanWardhana, Kementerian LingkunganHidup)
5.Manfaat tanaman transgenik ditinjaudari aspek sosial, ekonomi, dan budaya(Dr. Erna Maria Lokollo, PuslitbangSosial Ekonomi Pertanian)
6.Risiko tanaman transgenik ditinjau dariaspek sosial, ekonomi, dan budaya(Tejo Wahyu Jatmiko, Konphalindo)
7.Manfaat tanaman transgenik ditinjaudari aspek kesehatan (Prof. Dr. DeddyMuchtadi, MS, Institut Pertanian Bogor)
telah dikomersialkan, yaitu jagung, kanola,kapas, kedelai, kentang, papaya, squash,
dantomat.
Di samping perbaikan ketahanan ter-hadap cekaman biotik, abiotik, dan nutrisi,teknologi rekayasa genetik dapat diaplika-sikan untuk memproduksi vaksin tahan pe-nyakit kolera ke dalam pisang dan kentang,sehingga orang hanya dengan memakanpisang atau kentang transgenik yang me-ngandung vaksin penyakit kolera dapat ter-hindar dari serangan penyakit tersebut.Teknologi rekayasa genetik ini diharapkandapat meningkatkan produksi dan kualitashasil pertanian, yang pada gilirannya meningkatkan daya saing di pasar global.
Selain manfaat dan keuntungan yangdapat diharapkan dari tanaman transgenik,akhir-akhir ini telah beredar isu kekhawa-tiran bahwa tanaman tersebut akan berisi-ko mengganggu, merugikan, dan memba-
hayakan lingkungan dan kesehatan ma-nusia. Dengan memperhatikan kekhawatir-an tersebut, maka penelitian, pengem-bangan, dan pemanfaatan tanaman transgenik di Indonesia perlu dianalisis risiko-nya, diatur dengan cermat, dan harus
memperhatikan pendekatan kehati-hatian(precautionary approach) sebelum diko-mersialisasikan.
Pameran dan Seminar Bioteknologidiselenggarakan di Auditorium Dr. Ir. M.Ismunadji Bogor pada tangggal 30-31Oktober 2002, dibuka oleh SekretarisBadan Litbang Pertanian (Dr. Djoko S.Damardjati) dan ditutup oleh Kepala BadanLitbang Pertanian (Dr. Joko Budianto) sertadihadiri sekitar 200 peserta yang berasaldari lembaga penelitian, perguruan tinggi,swasta, dan lembaga swadaya masyarakat.
Sebanyak 22 poster yang berkaitan denganbioteknologi ditampilkan dalam pamerantersebut. Selain pameran poster, salah satu
Penyebarluasan Hasil Penelitian

129
8.On-lineProteinPurification(Ir. Elisabeth Maria, MSi)
9.DUS Testing in the Netherlands(Ir. N.P.A. Marrewijk)
UNIT KOMERSIALJSASI TEKNOLOGIPada tahun anggaran 2002, Unit
Komersialisasi Teknologi telah melaksana-kan kegiatan sebagai berikut:
1.Merintis kerja sama dengan Pemda DKIJakarta dalam rangka mencari teknologi produksi bibit buah-buahan tropikmelalui kultur in vitro,
2.Mengadakan pertemuan dalam rangkapromosi produk unggulan Balitbiogen.Pertemuan dihadiri oleh 135 orangpeserta yang berasal dari 25 instansibaik pemerintah maupun swasta.Dalam pertemuan tersebut diperkenal-kan beberapa produk unggulan hasilpenelitian Balitbiogen yang mempu-nyai nilai komersial. Selain itu, juga di-perkenalkan aset-aset Balitbiogen yangberpotensi untuk ditawarkan sebagaiObyek Wisata Ilmiah kepada wisata-wan minat khusus seperti para siswa,mahasiswa, peneliti, dan praktisi dibidang pertanian,
3.Mengikuti pelatihan Pengelolaan Wisa-ta Agro di Malang, Jawa Timur. Pelatihan ini diselenggarakan oleh Komisi Wi-sata Agro Departemen Pertanian beker-jasama dengan Asosiasi Wisata AgroIndonesia dan diikuti oleh peserta yangberasal dari 22 unit kerja baik yangberada di lingkup Badan Litbang Pertanian maupun swasta dan sekolah keju-
ruan,
4.Mengikuti pameran Agro Tourism Expo2002 yang diselenggarakan olehasi Wisata Agro Indonesia.
8. Risiko tanaman transgenik ditinjau danaspek kesehatan (Prof. Dr. Maggy T.Suhartono, Institut Pertanian Bogor)
PRESENTASI RUTINPresentasi rutin merupakan seminar
yang dilaksanakan secara mtin dan matedyang disampaikan tidak hanya hasil pene-litian, tetapi dapat berupa proposal ataurencana penelitian, hasil perjalanan dinasatau monitoring, dan hasil yang diperolehdari suatu pertemuan (seminar, lokakarya,pelatihan, temu ilmiah, temu lapang, dll.)di dalam maupun di luar negeri. Selain itu,masalah-masalah yang perlu segera di-informasikan ataupun masalah-masalah
yang memerlukan masukan lebih luas da-pat pula diseminarkan di luar jadwal yangtelah ditetapkan.
Beberapa materi yang telah diseminarkan, yaitu1.Emisi Gas Metana (CH4) pada Lahan
Sawah (Dr. A. Hidayat)2.Bioinformatika (Dr. M. Herman)
3.Interaction of Anti-insect Dietary Proteins in cowpea Bruchid Callosobru-chus maculatus (Dr. Bahagiawati Amir-husin)
4.Challenges and Options for AgricultureBiotechnology in Indonesia (Dr. EijaPehu)
5.Hak Pilih Petani dalam PemanfaatanTeknologi Pertanian: Studi Kasus PetaniSulawesi Selatan (Ir. Erizal)
6.Aplikasi Flow Cytometer-cell Counterdalam Biologi (Dr. Carsten Herrmann)
7.Pemetaan Sifat Ketahanan terhadapKekeringan pada Tanaman Padi:Laporan Kemajuan (Kurniawan RudiTrijatmiko, SP, MSi)
Laporan Tahunan Balitbiogen 2002
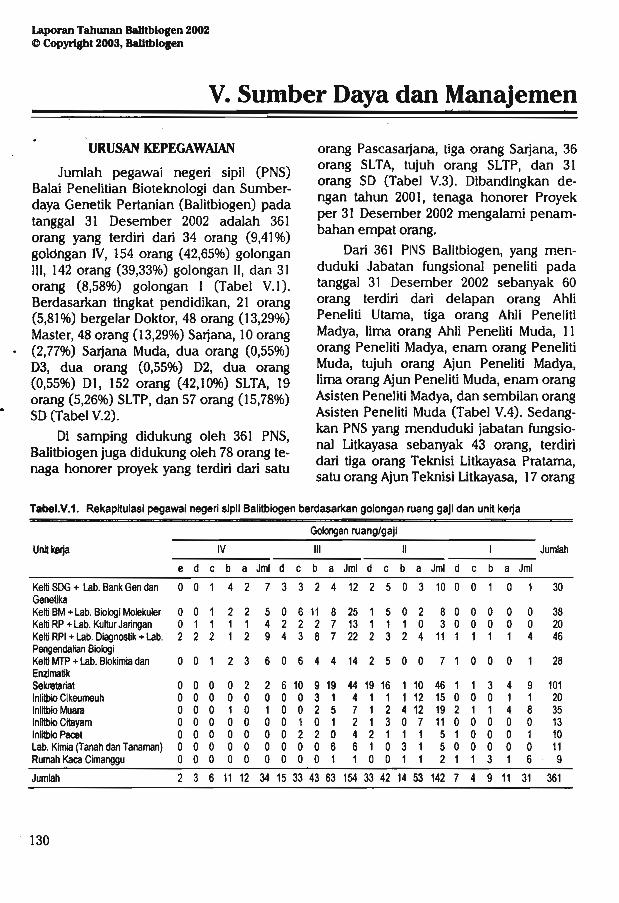
130
3619
1110133520
101
28
462038
30
Jumlah
316010819
1
400
1
Jml
111000414
0
100
0a
93000103
0
100
1bI
41000101
0
100
0c
71010201
1
100
0d
142255
11191546
7
1138
10
Jml
531117
121210
0
402
3a
141310411
0
210
0bII
42001321
16
5
315
5c
33012111
19
2
211
2d
154164274
44
14
221325
12Jml
63160151
19
4
778
4a
430020239
4
82
11
2bIII
Golongan ruang/gaji
33002100
10
6
326
3c
150000006
0
420
3d
340000102
6
945
7Jml
120000002
3
212
2a
110000100
2
112
4b
IV
60000000
1
211
1c
30000000
0
210
0d
20000000
0
200
0e
Jumlah
Rumah Kaca CimangguLab. Kimia (Tanah dan Tanaman)Inlitbio PacetInlitbio CitayamInlitbio MuaraInlitbio CikeumeuhSekretariatEnzimatikKelti MTP + Lab. Biokimia danPengendalian BiologiKelti RPI + Lab. Diagnostik + Lab.Kelti RP +Lab. Kultur JaringanKelti BM + Lab. Biologi MolekulerGenetikaKelti SDG+Lab. Bank Gen dan
Unit kerja
Tabel.V.1. Rekapitulasi pegawai negeri sipil Balitbiogen berdasarkan golongan ruang gaji dan unit kerja
orang Pascasarjana, tiga orang Sarjana, 36orang SLTA, tujuh orang SLTP, dan 31orang SD (Tabel V.3). Dibandingkan de-ngan tahun 2001, tenaga honorer Proyekper 31 Desember 2002 mengalami penam-bahan empat orang.
Dari 361 PNS Balitbiogen, yang men-duduki Jabatan fungsional peneliti padatanggal 31 Desember 2002 sebanyak 60orang terdiri dari delapan orang AhliPeneliti Utama, tiga orang Ahli PenelitiMadya, lima orang Ahli Peneliti Muda, 11orang Peneliti Madya, enam orang PenelitiMuda, tujuh orang Ajun Peneliti Madya,lima orang Ajun Peneliti Muda, enam orangAsisten Peneliti Madya, dan sembilan orangAsisten Peneliti Muda (Tabel V.4). Sedang-kan PNS yang menduduki jabatan fungsional Litkayasa sebanyak 43 orang, terdiridari tiga orang Teknisi Litkayasa Pratama,satu orang Ajun Teknisi Litkayasa, 17 orang
URUSAN KEPEGAWAIANJumlah pegawai negeri sipil (PNS)
Balai Penelitian Bioteknologi dan Sumber-daya Genetik Pertanian (Balitbiogen) padatanggal 31 Desember 2002 adalah 361orang yang terdiri dari 34 orang (9,41%)goldngan IV, 154 orang (42,65%) golonganIII, 142 orang (39,33%) golongan II, dan 31orang (8,58%) golongan I (Tabel V.I).Berdasarkan tingkat pendidikan, 21 orang(5,81%) bergelar Doktor, 48 orang (13,29%)Master, 48 orang (13,29%) Sarjana, 10 orang(2,77%) Sarjana Muda, dua orang (0,55%)D3, dua orang (0,55%) D2, dua orang(0,55%) Dl, 152 orang (42,10%) SLTA, 19orang (5,26%) SLTP, dan 57 orang (15,78%)SD (Tabel V.2).
Di samping didukung oleh 361 PNS,Balitbiogen juga didukung oleh 78 orang te-naga honorer proyek yang terdiri dari satu
V. Sumber Daya dan Manajemen
Laporan Tahunan Balltbiogen 2002© Copyright 2003, Balitblogen
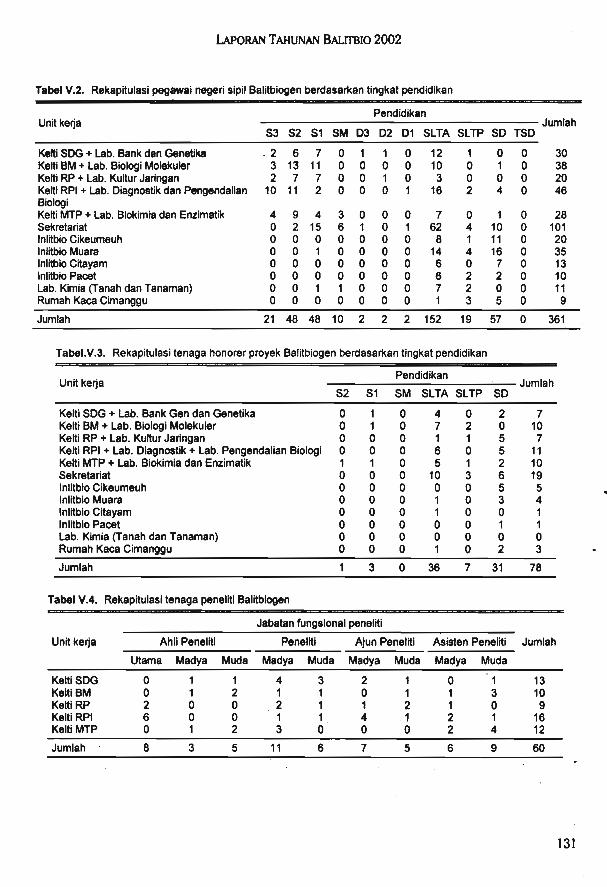
131
6011Jumiah12169
1013
41031
22110
01211
04102
01113
31214
20021
10011
06200
Kelti MTPKelti RPIKeKi RPKelti BMKelti SDG
Jumiah
78
Muda
Peneliti
31
Madya
Asisten
36 7
Madya Muda
Ajun Peneliti
Utama Madya Muda Madya Muda
Ahli Peneliti Peneliti
Jabatan fungsional peneliti
1 3 0
Rekapitulasi tenaga peneliti Balitbiogen
Unit kerja
Tabel V.4.
Jumiah
7107
111019541103
205526530102
021013000000
47165
10011001
000000000000
110010000000
000010000000
Kelti SDG + Lab. Bank Gen dan GenetikaKelti BM + Lab. Biologi MolekulerKelti RP + Lab. Kultur JaringanKelti RPI + Lab. Diagnostik + Lab. Pengendalian BiologiKeKi MTP + Lab. Biokimia dan EnzimatikSekretariatInlitbio CikeumeuhInlitbio MuaraInlitbio CitayamInlitbio PacetLab. Kimia (Tanah dan Tanaman)Rumah Kaca Cimanggu
- Jumiah
0 361
SD
57
S2 S1 SM SLTA SLTPPendidikan
Balitbiogen berdasarkan tingkat pendidikan
48 48 10 2 2 2 152 19
honorer proyek
21
Rekapitulasi tenaga
Unit kerja
Tabel.V.3.
Jumiah9
1110133520
10128
46203830
jumii
00000000
0000
TSD
5027
161110
1
4010
SD
32204140
2001
SLTP
1766
148
627
163
1012
SLTA
00000010
1000
D1
00000000
0101
D2
00000010
0001
D3
01000063
0000
SM
010010
154
27
117
S1
00000029
117
136
S2
00000004
10232
S3
Rumah Kaca CimangguLab. Kimia (Tanah dan Tanaman)Inlitbio PacetInlitbio CrtayamInlitbio MuaraInlitbio CikeumeuhSekretariatKeKi MTP + Lab. Biokimia dan EnzimatikBiologiKelti RPI + Lab. Diagnostik dan PengendalianKelti RP + Lab. Kultur JaringanKelti BM + Lab. Biologi MolekulerKeKi SOG + Lab. Bank dan Genetika
unit Keija
Tabel V.2. Rekapitulasi pegawai negeri sipil Balitbiogen berdasarkan tingkat pendidikan
Pendidikan
Laporan Tahunan Balitbio 2002
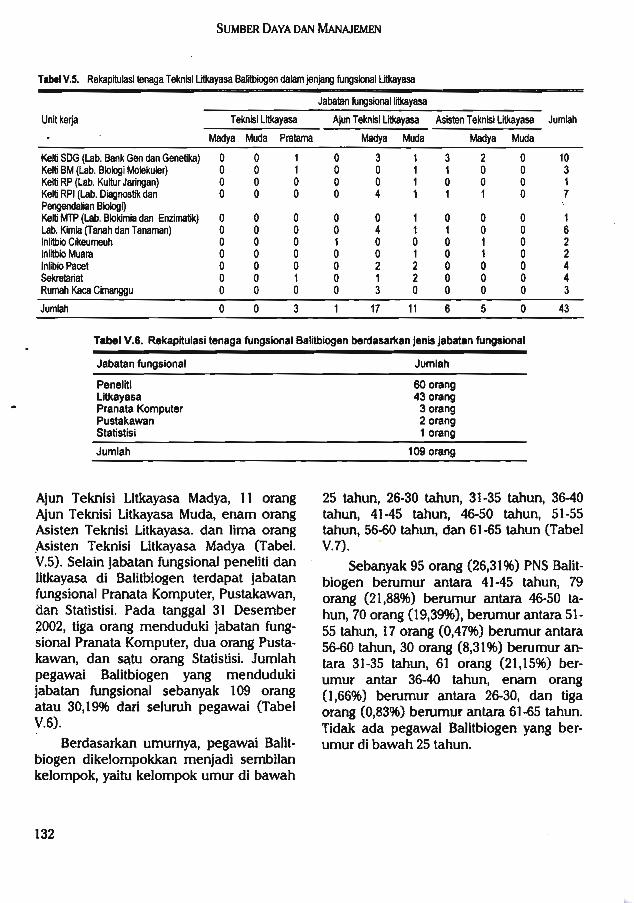
132
25 tahun, 26-30 tahun, 31-35 tahun, 36-40tahun, 41-45 tahun, 46-50 tahun, 51-55tahun, 56-60 tahun, dan 61-65 tahun (TabelV.7).
Sebanyak 95 orang (26,31%) PNS Balitbiogen berumur antara 41-45 tahun, 79orang (21,88%) berumur antara 46-50 tahun, 70 orang (19,39%), berumur antara 51-55 tahun, 17 orang (0,47%) berumur antara56-60 tahun, 30 orang (8,31%) berumur antara 31-35 tahun, 61 orang (21,15%) berumur antar 36-40 tahun, enam orang(1,66%) berumur antara 26-30, dan tigaorang (0,83%) berumur antara 61-65 tahun.Tidak ada pegawai Balitbiogen yang berumur di bawah 25 tahun.
Ajun Teknisi Litkayasa Madya, 11 orangAjun Teknisi Litkayasa Muda, enam orangAsisten Teknisi Litkayasa. dan lima orangAsisten Teknisi Litkayasa Madya (Tabel.V.5). Selain jabatan fungsional peneliti danlitkayasa di Balitbiogen terdapat jabatanfungsional Pranata Komputer, Pustakawan,dan Statistisi. Pada tanggal 31 Desember2002, tiga orang menduduki jabatan fungsional Pranata Komputer, dua orang Pustakawan, dan satu orang Statistisi. Jumlahpegawai Balitbiogen yang mendudukijabatan fungsional sebanyak 109 orangatau 30,19% dari seluruh pegawai (TabelV.6).
Berdasarkan umurnya, pegawai Balitbiogen dikelompokkan menjadi sembilankelompok, yaitu kelompok umur di bawah
433442261
713
10
Jumlah
109 orang
1 orang2 orang3 orang
43orang60 orang
Jumlah
Jumlah
StatistisiPustakawanPranata KomputerLitkayasaPeneliti
Jabatan fungsional
Tabel V.6. Rekapitulasi tenaga fungsional Balitbiogen berdasatkan jenis jabatan fungsional
00000000
0000
Muda
50001100
1002
Madya
11 60 02 02 01 00 01 11 0
1 11 01 11 3
Muda
173120040
4003
Madya
10000100
0000
Ajun Teknisi Litkayasa Asisten Teknisi Litkayasa
30100000
0011
Pratama
00000000
0000
Muda
00000000
0000
Madya
Teknisi Litkayasa
Jabatan fungsional litkayasa
Jumlah
Rumah Kaca CimangguSekretariatInlibio PacetInlitbio MuaraInlltbio CikeumeuhLab. Kimia (Tanah dan Tanaman)Kelt MTP (Lab. Biokimia dan Enzlmatk)Pengendalian Btologi)Kelt RPI (Lab. Diagnostk danKelti RP (Lab. Kultur Jaringan)Kelt BM (Lab. Btologi Molekuler)Kelti SDG (Lab. Bank Gen dan Genetika)
V
Unit kerja
Tibel V.5. Rekapitulasi tenaga Teknisi Litkayasa Balitbiogen dalam jenjang fungsional Litkayasa
SUMBER DAYA DAN MANAJEMEN
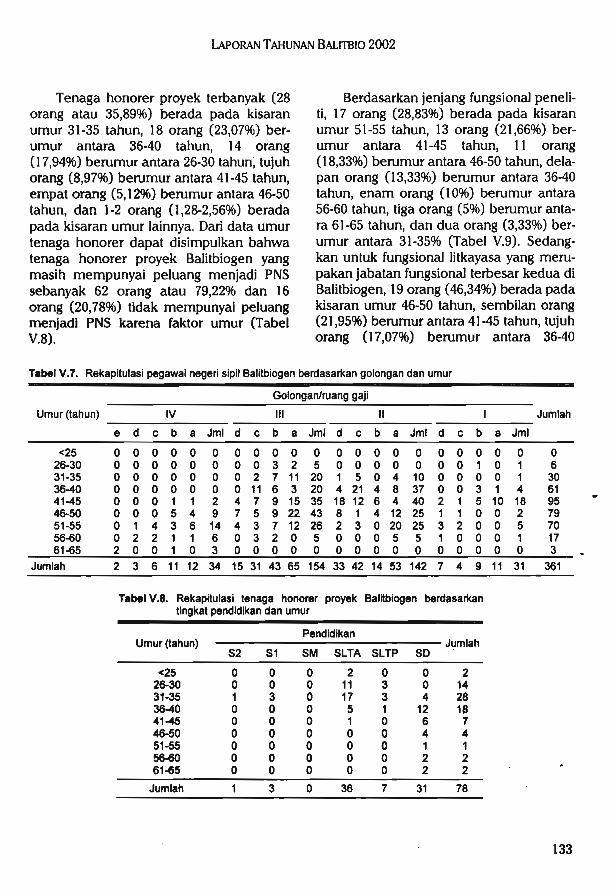
133
3613
17707995613060
Jumlah
310152
184110
Jml
110000
101000a
ih
7822147
1828142
3122146
12400
SD
7000001330
SLTP
Balitbiogen berdasarkan
9000053010
bI
4002110000c
7013120000
d
14205
252540371000
Jml
14 530 00 50 204 126 44 80 40 00 0
b a
II
36000015
17112
SLTA
0000000000
SM
Pendidikan
umurhonorer proyek
33 420 00 02 38 1
18 124 211 50 00 0
d c
15405
264335202050
Jml
65001222153
1120a
I
3000000300
S1
1000000100
S2
Jumlah
61-6556-6051-5546-5041-4536-4031-3526-30
<25
tingkat pendidikan dan
1 1,
Tabel V.8. Rekapitulasi tenaga I
43027996730
bIII
Golongan/ruang gaji
310335711200c
15004740000
d
3436
14920000
Jml
12016410000a
6 110 12 14 30 50 10 00 00 00 0
c b
IV
3021000000
d
2200000000e
Jumlah61-6556-6051-5546-5041-4536-4031-3526-30<25
Umur (tahun)
Tabel V.7. Rekapitulasi pegawai negeri sipil Balitbiogen berdasarkan golongan dan umur
Berdasarkan jenjang fungsional peneli-ti, 17 orang (28,83%) berada pada kisaranumur 51-55 tahun, 13 orang (21,66%) berumur antara 41-45 tahun, 11 orang
(18,33%) berumur antara 46-50 tahun, dela-pan orang (13,33%) berumur antara 36-40tahun, enam orang (10%) berumur antara56-60 tahun, tiga orang (5%) berumur antara 61-65 tahun, dan dua orang (3,33%) berumur antara 31-35% (Tabel V.9). Sedang-kan untuk fungsional litkayasa yang meru-pakan jabatan fungsional terbesar kedua diBalitbiogen, 19 orang (46,34%) berada padakisaran umur 46-50 tahun, sembilan orang(21,95%) berumur antara 41-45 tahun, tujuhorang (17,07%) berumur antara 36-40
Tenaga honorer proyek terbanyak (28orang atau 35,89%) berada pada kisaranumur 31-35 tahun, 18 orang (23,07%) ber-umur antara 36-40 tahun, 14 orang
(17,94%) berumur antara 26-30 tahun, tujuhorang (8,97%) berumur antara 41-45 tahun,empat orang (5,12%) berumur antara 46-50tahun, dan 1-2 orang (1,28-2,56%) beradapada kisaran umur lainnya. Dari data umurtenaga honorer dapat disimpulkan bahwatenaga honorer proyek Balitbiogen yangmasih mempunyai peluang menjadi PNSsebanyak 62 orang atau 79,22% dan 16orang (20,78%) tidak mempunyai peluangmenjadi PNS karena faktor umur (TabelV.8).
Laporan Tahunan Balitbio 2002
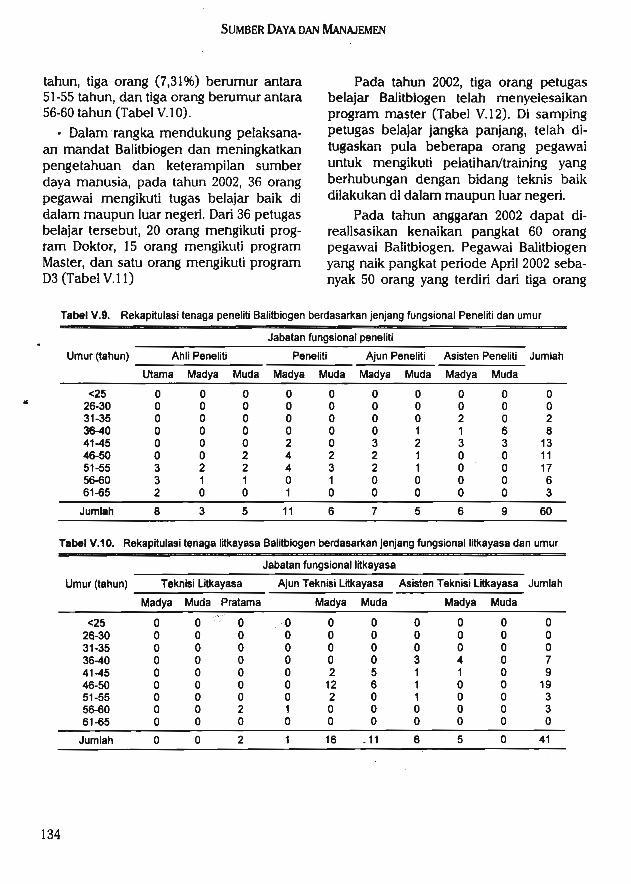
134
41
033
1997000
Jumlah
0000000000
Muda
5000014000
Madya
6001113000
Asisten Teknisi Litkayasa
-11
000650000
Muda
16002
1220000
Madya
1010000000
Ajun Teknisi Litkayasa .
Jabatan fungsional litkayasa
Balitbiogen berdasarkan jenjang fungsional litkayasa dan umur
6036
1711138200
Jumlah
9000036000
Muda
Peneliti
6000031200
Madya
Asisten
5001121000
Muda
7002230000
Madya
Ajun Peneliti
6013200000
Muda
11104420000
Madya
Peneliti
Jabatan fungsional peneliti
2020000000
Pratama
000000000o "-'"•
Muda
0000000000
Madya
Teknisi Litkayasa
Jumlah
61-6556-6051-5546-5041-4536-4031-3526-30
<25
Umur (tahun)
Tabel V.10. Rekapitulasi tenaga litkayasa
5012200000
Muda
3012000000
Madya
8233000000
Utama
Ahli Peneliti
Jumlah
61-6556-6051-5546-5041-4536-4031-3526-30<25
Umur (tahun)
Tabel V.9. Rekapitulasi tenaga peneliti Balitbiogen berdasarkan jenjang fungsional Peneliti dan umur
Pada tahun 2002, tiga orang petugasbelajar Balitbiogen telah menyelesaikanprogram master (Tabel V.I2). Di sampingpetugas belajar jangka panjang, telah di-tugaskan pula beberapa orang pegawaiuntuk mengikuti pelatihan/training yangberhubungan dengan bidang teknis baikdilakukan di dalam maupun luar negeri.
Pada tahun anggaran 2002 dapat di-reaiisasikan kenaikan pangkat 60 orangpegawai Balitbiogen. Pegawai Balitbiogenyang naik pangkat periode April 2002 seba-nyak 50 orang yang terdiri dari tiga orang
tahun, tiga orang (7,31%) berumur antara51-55 tahun, dan tiga orang berumur antara
56-60 tahun (TabelV. 10).
• Dalam rangka mendukung pelaksana-an mandat Balitbiogen dan meningkatkanpengetahuan dan keterampilan sumberdaya manusia, pada tahun 2002, 36 orangpegawai mengikuti tugas belajar baik didalam maupun luar negeri. Dari 36 petugasbelajar tersebut, 20 orang mengikuti program Doktor, 15 orang mengikuti programMaster, dan satu orang mengikuti programD3(TabelV.ll)
SUMBER DAYA DAN MANAJEMEN
135
IPB
JepangKoba Univarsity
IPBsaj^__ —j^^^ ri^i^^j,.•vagamngan DManoaUnivereity ofUnivereity KoreaSeoul NationalUSACornel UnivereitySouth Wales AustraliaUnivereity of New
IPB
Ul
IPBUnivereity JapanOsaka Perpecture
UNSWAustraka
UNSW AustraliaWageningen BotandaUtwereilyor
IPB
UGMUSACome* University
IPBPbWgJnaUniversity of The
IPBUSACornel University
UniversityPutra Malaysia
IPB
IPB
IPB
UNPAD
UNPAD
UNPAD
IPB
IPB
IPB
IPB
UNPADPNIiplnaUniversity of die
IPB
IPB
Unlversdas
ARMPII
MonBusho
PAATP
BIORIN
ARMPII
ADS
PAATP
PAATP
PAATP
Monbusho
PAATP
PAATP
BIORIN
PAATP
PAATP
PAATP
ARMPII
IRRI
ARMPIIFoundationRockefeller
PAATP
PAATP
PAATP
PAATP
ARMPII
ARMPII
ARMPII
ARMPII
ARMPII
Baktbiogen
BWDbJogen
ARMPII
ARMPII
ARMPII
ARMPII
Sponsor
Komputer
Biotechnology
Agronomi
Biotechnology
Biotechnology
Plant Breodfog
Biotechnology
Bioiogi
KbNaPangan
Agronomi
Biotaknologj
BMaknotogi
Bioteknologi
Bteknolo
Pamuiaan
PamWaai
BiotocbnologyMotekulerBMogi
Plant Virology
BWteknologiGenet*Plant Breedmg
Tissue CuWire
Bloteknologl
Bloteknologi
Bfotoknologi
Bloteknologi
Blotoknologi
Pemuliaan
Pemukaan
Pemukaan
Bloteknologi
BloteknologiTanamanPemuhaan
Biotechnology
fNotokook>of
Blotoknologi
Spesialisasi
D3(DN)
S3(LN)
S3(DN)
S3 (IN)
S3(LN)
S3(LN)
S3 (EN)
S3 (DM)
S3(DN)
S3(DN)
S3(LN)
S3 (IN)
S3(LN)
S3(LN)
S3(DN)
S3(DN)
S3(LN)
S3(DN)
S3 (IN)
S3 (ON)
S3 (IN)
S2(LN)
S2(DN)
S2(DN)
S2(DN)
S2(DN)
S2(DN)i
S2DN)
S2(DN)
S2(DN)
S2(DN)
S2(DN)
S2(DN)
S2(LN)
S2(DN)
S2(DN)
Program tugas belajar
1 September 2000
Oktober2002
1 September 2002
Februari2002
Febnwi2002
Januari2002
16Jaouarl2002
Agustus2001
September 2001
Agustus2001
Oktober1996
Februarl2001
Febnwl2001
15Pebr.2OO1
September 2000
1 Februari2000
31Ju81999
1 September 1999
Desember1998
1Aprit1998
19Agustus1996
Ju82001-Mei2003
Agustus2001
Agustus2001
1 September 2000
1 September 2000
1 September 2000
1 September 2000
1 September 2000
1 September 2000
1 September 1999
1 September 1999
1 September 1999
11Junl1999
1 September 1999
1 September 1999
TMTtugasbelajar
LitkayasaMudaAjunTekNsi
KeWBMPenelW Non Klas
Asisten PenelW MadyaKeWRP
KeWBMPenelW Non KlasKeWMTPAslsten PenelW MudaKeWRPIPenelW Non KlasKeWBMPenelW Non KlasKeWRPPenelW MudaKeWRPInew dmAjun PeneH Muda
PenelW Non KlasKale nu
KeWBMnvni M1 rPenelW Non Klas
PenelW Non KlasKoffiMTP
KeWMTPPenelW Non KlasKaWRPIPenelW Non KlasKeWSDGPenelW MadyaKeWSDGAjun PenelW MadyaKeWBMPenelW Non KlasKeWMTPAsisten Penelti MadyaKeWRPIAjun PenelW MadyaKeWBMAsisten PenelW MadyaKeWBMPenelW Non Klas
KeWRPPenelW Non KlasKaWRPPenelW Non Klas
Aslsten PenelW MadyaKeW BM
KeWBMPenelW Non KlasKeWBMPenelW Non KlasKeWMTPPenelW Non KlasKeWBMPenelW Non KlasKeWBMAsistenPsneWIMudaKeWSDGPenaHdNonKlaaKeWBMPenelW Non KlasKeltRPPenelrti Non KlasKeWBMAsisten Panaw ModflKettMTPAsistanPanalidMuda
Ajun Penetd Mudaa/_U| nn
KeWMTPAsisten PenelW Madya
Jabatan
ll/d
III*
III*
III*
III*
III*
Ill/a
Ma
llVa
III*
III*
III*
III*
III*
IV*
IV/a
in*
III*
III*
III*
Ill/b
Ill/a
Ill/a
Ill/c
Ill/a
Ill/a
Ill/a
IIVc
Ill/b
Ill/a
Ill/a
lll/a
in/b
Hl/b
Ill/b
Ill/to
Pangkatsemula
NIP. 0800703931. SyarifulHidayal
NIP. 080.115.831rRr • U0v< JUI-CU 120. TotoHadlarto.MSc
19. lr.Rag^>adn8P.,MSiNIP fWl 101 A31
NIP. 080.124.48818. K.RuWTryaMko.SP.MSI
NIP. 080.124.74017. Pup Lestari, MSi
NIP. 080.115.82816. DaniSatyawan.MSc
NIP. 080.115.83015. RanaVusvanaMSc
NIP. 080.079.76514. Dra.Endang6a8L.MSi
NIP. 080.085.308Nir. uou.i lr.kao
13. AsepNugrahaA.MSi
12. Dra. DwimtaW.Utani.MSimid nun 117 OTA
NIP. 080.110.824
10. Ir.PujoyuwonoM.MSiNIP. 080.115.869
11. SusSprijatno,SSi
NIP. 080.110.3939. Ir Eny Ida Riyanii. MSi
NIP. 080.113.9918. Ir. Chaerani, MSc
NIP. 080.054.4937. k.Sutoro.MS
NIP. 080.066.9186. k. Asa*, MS
NIP. 080.108.4685. Ir.M.Yunus.MSi
NIP. 080.107.9134. lr.EttyPra6wi,MSi
NIP. 080.052.6193. Ors. M. Muhsin, MSi
NIP. 080.098.2232. Drs.SaplowoJP.MSi
NIP. 080.118.3591. Ir.EndangM. Septiningsih
Program S3NIP. 080.124.355
16. Slid Rahayu, SSINIP. 080.126.046nir. uou.uaa.'Di14. MiaKosmiabn, SSi
13. Dra. Diani DamayanbSUP (Wl OQQ 7R1
NIP. 080.126.11512. Joko Prasetiyono, SP
NIP. 080.126.04711. TtiJokoSantoso,SP
NIP. 080.126.04810. NurAzizah, SP
NIP. 080.111.3259. Drs. Ahmad Warsun
NIP. 080.111.3268. DreEdyListanto
NIP. 080.124.4897. Hakim Kurniawan.SP
NIP. 080.130.4266. Atmitri Sishanrtni. SSI
NIP. 080.128.6375. IkaRoosdkaT.SP
NIP. 080.102.9394. Ir. Budi Santosa
NIP. 080.108.601Nir. UBU.lUy.Ol^
3. Ir. Rath D. Hastut
2. Dre.A8HusniMID t\fU\ 1 HO ATA
NIP. 080.114.680
Program S21. Ir. R. Cinta B. Gintin^
No. Nama
TaMV.11. Nama-namapetugasbelajarBalitbioven
Laporan Tahunan Balitbio 2002
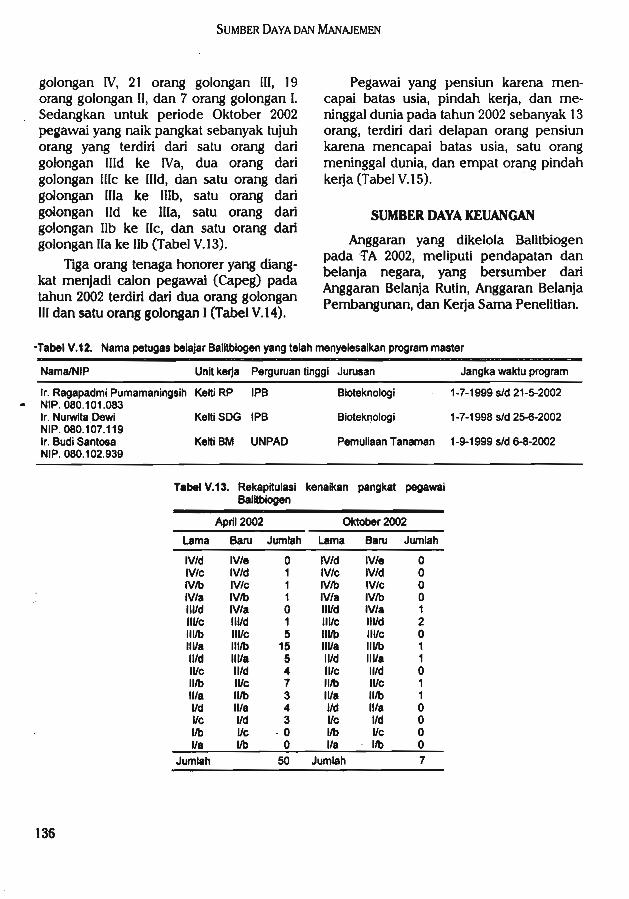
136
70000110110210000
Jumlah
l/bl/cl/d
ll/all/bll/cll/d
Ill/aIll/bIll/cIll/dIV/aIV/bIV/cIV/dIV/e
Baru
JumlahI/al/bl/cl/d
ll/all/bll/cll/d
Ill/aIll/bIll/cIll/dIV/aIV/bIV/cIV/d
Lama
Oktober 2002
pegawaipangkatkenaikan
500
- 0343745
155101110
Jumlah
l/bl/cl/d
ll/all/bll/cll/d
Ill/aIll/bIll/cIll/dIV/aIV/bIV/cIV/dIV/e
Baru
April 2002
Balitbiogen
JumlahI/al/bl/cl/d
ll/all/bll/cll/d
Ill/aIll/bIll/cIll/dIV/aIV/bIV/cIV/d
Lama
Tabel V.13. Rekapitulasi
Bioteknologi1-7-1999 s/d 21-5-2002
Bioteknologi1 -7-1998 s/d 25-6-2002
Pemuliaan Tanaman 1 -9-1999 s/d 6-8-2002
Ir. Ragapadmi Purnamaningsih Kelti RPNIP. 080.101.083Ir. Nurwita DewiKelti SDGNIP. 080.107.119Ir. Budi SantosaKelti BM UNPADNIP. 080.102.939
IPB
IPB
Jangka waktu programUnit kerja Perguruan tinggi JurusanNama/NIP
-Tabel V.12. Nama petugas belajar Balitbiogen yang telah menyelesaikan program master
Pegawai yang pensiun karena men-capai batas usia, pindah kerja, dan me-
ninggal dunia pada tahun 2002 sebanyak 13orang, terdiri dari delapan orang pensiunkarena mencapai batas usia, satu orang
meninggal dunia, dan empat orang pindahkerja (Tabel V.I5).
SUMBER DAYA KEUANGANAnggaran yang dikelola Balitbiogen
pada TA 2002, meliputi pendapatan danbelanja negara, yang bersumber dariAnggaran Belanja Rutin, Anggaran BelanjaPembangunan, dan Kerja Sama Penelitian.
golongan IV, 21 orang golongan HI, 19orang golongan II, dan 7 orang golongan I.Sedangkan untuk periode Oktober 2002pegawai yang naik pangkat sebanyak tujuhorang yang terdiri dari satu orang darigolongan Hid ke IVa, dua orang darigolongan Hie ke Hid, dan satu orang darigolongan Ilia ke IHb, satu orang darigolongan Hd ke Ilia, satu orang darigolongan lib ke lie, dan satu orang darigolongan Ha ke lib (Tabel V.13).
Tiga orang tenaga honorer yang diang-kat menjadi calon pegawai (Capeg) padatahun 2002 terdiri dari dua orang golonganIII dan satu orang golongan I (Tabel V.I4).
SUMBER DAYA DAN MANAJEMEN
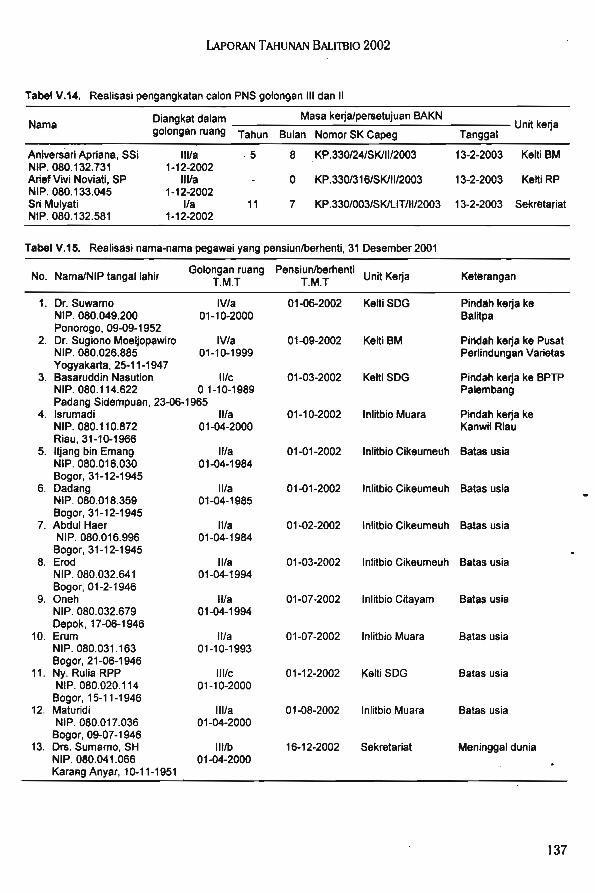
137
01-01-2002Inlitbio CikeumeuhBatasusia
01-01-2002Inlitbio CikeumeuhBatasusia
01-02-2002Inlitbio CikeumeuhBatasusia
01-03-2002Inlitbio CikeumeuhBatas usia
01-07-2002Inlitbio CitayamBatasusia
01-07-2002Inlitbio MuaraBatasusia
01-12-2002Kelti SDGBatasusia
01-08-2002Inlitbio MuaraBatasusia
16-12-2002SekretariatMeninggal dunia
Pindah kerja keBalitpa
Pindah kerja ke PusatPerlindungan Varietas
Pindah kerja ke BPTPPalembang
Pindah kerja keKanwH Riau
01-10-2002 Inlitbio Muara
01-03-2002 Kelti SDG
01-09-2002 Kelti BM
01-06-2002 Kelti SDG
01-04-2000Ill/b
01-04-2000Ill/a
01-10-2000Ill/c
01-10-1993ll/a
01-04-1994ll/a
01-04-1994ll/a
01-04-1984ll/a
01-04-1985ll/a
01-04-1984ll/a
01-04-2000I I/a
KarangAnyar, 10-11-1951NIP. 080.041.066
13. Drs. Sumarno, SHBogor, 09-07-1946NIP. 080.017.036
12. MaturidiBogor, 15-11-1946NIP. 080.020.114
11. Ny. RuliaRPPBogor, 21-06-1946NIP. 080.031.163
10. ErumDepok, 17-06-1946NIP. 080.032.679
9. OnehBogor, 01-2-1946NIP. 080.032.641
8. ErodBogor, 31-12-1945NIP. 080.016.996
7. Abdul HaerBogor, 31-12-1945NIP. 080.018.359
6. DadangBogor, 31-12-1945NIP. 080.016.030
5. Itjang bin EmangRiau, 31-10-1966NIP. 080.110.872
4. IsrumadiPadang Sidempuan, 23-06-1965
01-10-1989ll/c
01-10-1999IV/a
01-10-2000IV/a
NIP. 080.114.6223. Basaruddin Nasution
Yogyakarta, 25-11-1947NIP. 080.026.885
2. Dr. Sugiono MoeljopawiroPonorogo, 09-09-1952NIP. 080.049.200
1. Dr. Suwarno
KeteranganOolongan ruang Pensiun/berhenti .. .. ^ .TM.TT.M.TUmtKenaNo. Nama/NIP tangal lahir
Tabel V.15. Realisasi nama-nama pegawai yang pensiun/berhenti, 31 Desember 2001
58 KP.330/24/SK/II/200313-2-2003 Kelti BM
0 KP.330/316/SK/II/200313-2-2003 Kelti RP
117 KP.330/003/SK/LIT/H/2003 13-2-2003 Sekretariat1-12-2002
I/a1-12-2002
Ill/a1-12-2002
Ill/a
NIP. 080.132.581Sri MulyatiNIP. 080.133.045AriefViviNoviati, SPNIP. 080.132.731Aniversari Apriana, SSi
Unit kerjaTanggal
Diangkat dalamgolongan ruang Tahun Bulan NomorSKCapeg
Masa kerja/persetujuan BAKNNama
Tabel V.14. Realisasi pengangkatan calon PNS golongan III dan II
Laporan Tahunan Balitbio 2002
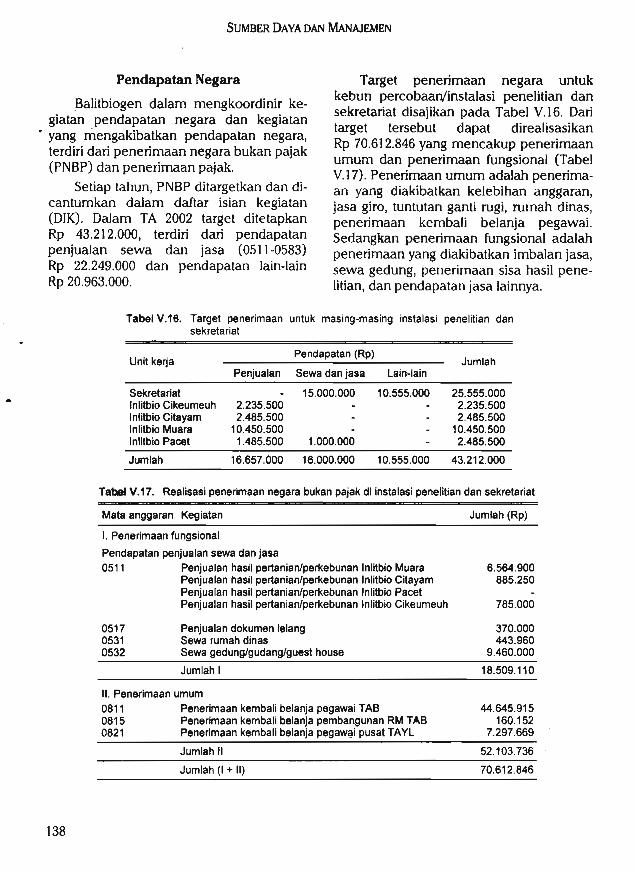
138
70.612.846Jumlah (I + II)
52.103.736
II. Penerimaan umum0811 Penerimaan kembali belanja pegawai TAB44.645.9150815 Penerimaan kembali belanja pembangunan RM TAB160.1520821 Penerimaan kembali belanja pegawai pusat TAYL7.297.669
18.509.110
Jumlah II
Jumlah I
6.564.900885.250
785.000
370.000443.960
9 460.000
I. Penerimaan fungsional
Pendapatan penjualan sewa dan jasa0511Penjualan hasil pertanian/perkebunan Inlitbio Muara
Penjualan hasil pertanian/perkebunan Inlitbio CitayamPenjualan hasil pertanian/perkebunan Inlitbio PacetPenjualan hasil pertanian/perkebunan Inlitbio Cikeumeuh
Jumlah (Rp)
0517Penjualan dokumen lelang0531Sewa rumah dinas0532Sewa gedung/gudang/guest house
Mata anggaran Kegiatan
Tabel V.17. Realisasi penerimaan negara bukan pajak di instalasi penelitian dan sekretariat
16.657.000 16.000.000 10.555.000 43.212.000Jumlah
-15.000.00010.555.00025.555.0002.235.500--2.235.5002.485.500--2.485.500
10.450.500--10.450.5001.485.5001.000.000-2.485.500
SekretariatInlitbio CikeumeuhInlitbio CitayamInlitbio MuaraInlitbio Pacet
JumlahPenjualan Sewa dan jasa Lain-lain
Pendapatan (Rp)Unit kerja
Tabel V.16. Target penerimaan untuk masing-masing instalasi penelitian dansekretariat
Target penerimaan negara untukkebun percobaan/instalasi penelitian dansekretariat disajikan pada Tabel V.I6. Daritarget tersebut dapat direalisasikanRp 70.612.846 yang mencakup penerimaanumum dan penerimaan fungsional (TabelV.I 7). Penerimaan umum adalah penerima
an yang diakibatkan kelebihan anggaran,jasa giro, tuntutan ganti rugi, rumah dinas,
penerimaan kembali belanja pegawai.Sedangkan penerimaan fungsional adalahpenerimaan yang diakibatkan imbalan jasa,sewa gedung, penerimaan sisa hasil pene
litian, dan pendapatan jasa lainnya.
Pendapatan Negara
Balitbiogen dalam mengkoordinir ke-giatan pendapatan negara dan kegiatanyang mengakibatkan pendapatan negara,terdiri dari penerimaan negara bukan pajak(PNBP) dan penerimaan pajak.
Setiap tahun, PNBP ditargetkan dan di-cantumkan dalam daftar isian kegiatan(DIK). Dalam TA 2002 target ditetapkanRp 43.212.000, terdiri dari pendapatanpenjualan sewa dan jasa (0511-0583)Rp 22.249.000 dan pendapatan Iain-lainRp 20.963.000.
SUMBER DAYA DAN MANAJEMEN

139
1.241.624
769.824471.800
281.996.052
12.310.604186.255.34157.006.81626.423.291
Jumlah (Rp)
6.785.868
5.822.489523.713439.666
4.849.542
3.607.918
1.241.624
PPh. 23
6.753.300
676.8005.563.500
513.000
9.395.443
1.925.689
6.785.868683.886
PPh. 22
-
-
212.418.391
27.000186.255.341
6.753.30019.382.750
PPh. 21
Jumlah pajak (Rp)
42.226.024
36.752.1792.542.7282.931.117
55.332.676
6.749.997
42.226.0246.356.655
PPN
Jumlah
BioteknologiPlasma nutfahManajemen varietas
Jumlah
Belanja rutinBelanja pegawaiBelanja proyekBelanja kerja sama penelitian
Sumber pajak
Tabel V.18. Realisasi penerimaan pajak Balitbiogen TA 2002
Rp 6.020.540.000 sedangkan untuk belanjanonpegawai mengalami kenaikanRp 177.597.000, yaitu dari Rp 1.100.002.000(TA 2001) menjadi Rp 1.277.599.000 (TA2002). Perkembangan anggaran rutin ling-kup Balitbiogen disajikan pada Tabel V.I9.
Dari rincian tersebut, anggaran belanjanonpegawai lebih banyak digunakan untukmata anggaran keluaran (MAK) 5230, yaituuntuk memenuhi langganan daya dan jasa(listrik, air, telepon, dan gas) yang dianggaprawan setiap tahun anggaran berjalansebesar Rp 865.656.000 atau 89% darianggaran belanja nonpegawai.
Realisasi anggaran rutin sampaidengan akhir Desember 2002 untuk belanjapegawai mencapai Rp 5.379.658.501 atau89% dari pagu DIK atau sisa Rp 640.881.499.Sedangkan realisasi belanja nonpegawaimencapai Rp 1.275.315.749 atau 99% dansisa dana sebesar Rp 2.283.251 disetorkanmelalui Kantor Perbendaharaan dan KasNegara (KPKN) Bogor (Tabel V.20).
Anggaran Belanja Pembangunan/Proyek -
Proyek Penelitian Genetika dan Biotek-nologi Bogor
Anggaran belanja pembangunan TA2002, Proyek Penelitian Genetika dan
Penerimaan negara pada TA 2002 khu-susnya pajak, yang telah dilaksanakan olehwajib pungut pajak (WAPU) meningkat di-bandingkan dengan TA 2001. Pada tahun2002 terdapat kenaikan pajak yang berasaldari pajak pribadi dan pajak penghasilan.
Pajak pribadi dikelola oleh Bendahara-wan Gaji di mana pemotongan dan penye-torannya dilaksanakan iangsung olehKantor Perbendaharaan dan Kas Negara(KPKN) Bogor, sedangkan pajak penghasilan bersumber dari pembayaran tunjanganpeiaksana proyek (TPP), pembelian barangatau jasa lainnya, dipungut Iangsung olehBendaharawan Proyek/Bagian Proyek,Bendaharawan Rutin maupun Bendaharawan Kerja Sama Penelitian. Secara kese-
luruhan pendapatan pajak sebesarRp 281.996.052. Realisasi pendapatan pajakselama TA 2002 disajikan pada Tabel V.I8.
Anggaran Belanja Rutin
Anggaran Belanja Rutin Balitbiogen un-tuk TA 2002 secara keseluruhan menunjuk-kan kenaikan. DIK TA 2002 sebesarRp 7.298.139.000 terdiri dari belanja pega-wai Rp 6.020.540.000 dan belanja nonpe-gawai Rp 1.277.599.000. Belanja pegawaimengalami kenaikan 10% dari TA 2001,yaitu dari Rp 5.478.698.000 menjadi
Laporan Tahunan Balitbio 2002
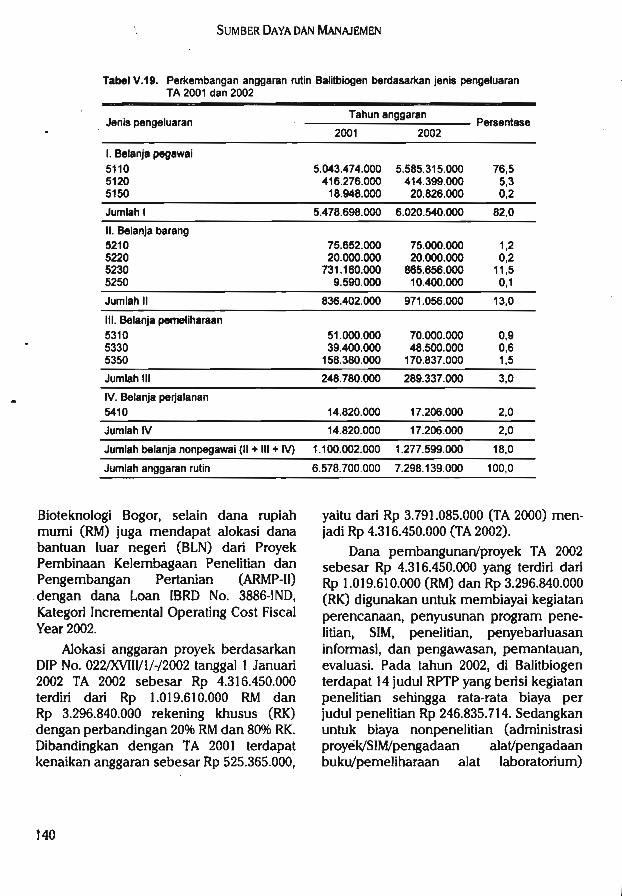
140
yaitu dari Rp 3.791.085.000 (TA 2000) men-jadi Rp 4.316.450.000 (TA 2002).
Dana pembangunan/proyek TA 2002sebesar Rp 4.316.450.000 yang terdiri dariRp 1.019.610.000 (RM) dan Rp 3.296.840.000(RK) digunakan untuk membiayai kegiatanperencanaan, penyusunan program penelitian, SIM, penelitian, penyebariuasaninformasi, dan pengawasan, pemantauan,
evaluasi. Pada tahun 2002, di Balitbiogenterdapat 14 judul RPTP yang berisi kegiatanpenelitian sehingga rata-rata biaya perjudul penelitian Rp 246.835.714. Sedangkanuntuk biaya nonpenelitian (administrasiproyek/SIM/pengadaan alat/pengadaanbuku/pemeliharaan alat laboratorium)
Bioteknologi Bogor, selain dana rupiahmurni (RM) juga mendapat alokasi danabantuan luar negeri (BLN) dari ProyekPembinaan Kelembagaan Penelitian danPengembangan Pertanian (ARMP-II)dengan dana Loan IBRD No. 3886-IND,Kategori Incremental Operating Cost FiscalYear 2002.
Alokasi anggaran proyek berdasarkanDIP No. 022/XVIII/1/-/2002 tanggal 1 Januari2002 TA 2002 sebesar Rp 4.316.450.000terdiri dari Rp 1.019.610.000 RM danRp 3.296.840.000 rekening khusus (RK)dengan perbandingan 20% RM dan 80% RK.Dibandingkan dengan TA 2001 terdapatkenaikan anggaran sebesar Rp 525.365.000,
100,0
18,0
2,0
2,0
3,0
0,90,61,5
13,0
7.298.139.000
1.277.599.000
17.206.000
17.206.000
289.337.000
70.000.00048.500.000
170.837.000
971.056.000
6.578.700.000
1.100.002.000
14.820.000
14.820.000
248.780.000
51.000.00039.400.000
158.380.000
836.402.000
Jumlah anggaran rutin
Jumlah belanja nonpegawai (II + III + IV)
Jumlah IV
IV. Belanja perjalanan
5410
Jumlah III
III. Belanja pemeliharaan
531053305350
Jumlah II
75.652.00075.000.0001,220.000.00020.000.0000,2
731.160.000865.656.00011,59.590.00010.400.0000,1
II. Belanja barang5210522052305250
82,05.478.698.000 6.020.540.000Jumlah I
5.043.474.000 5.585.315.000 76,5416.276.000 414.399.000 5,3
18.948.000 20.826.000 0,2
I. Belanja pegawai
511051205150
Persentase20022001
Tahun anggaranJenis pengeluaran
Tabel V.19. Perkembangan anggaran rutin Balitbiogen berdasarkan jenis pengeluaranTA 2001 dan 2002
SUMBER DAYA DAN MANAJEMEN
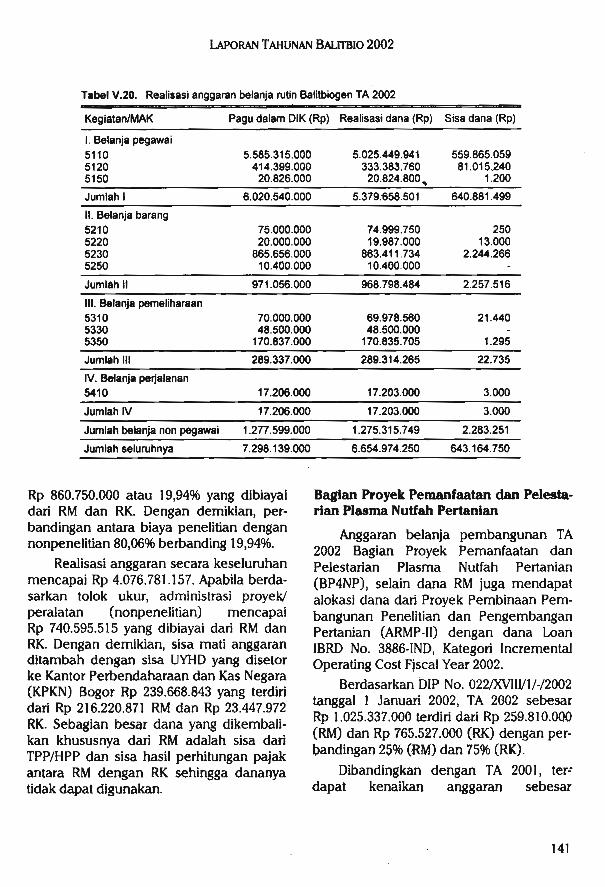
141
Bagian Proyek Pemanfaatan dan Pelesta-rian Plasma Nutfah Pertanian
Anggaran belanja pembangunan TA2002 Bagian Proyek Pemanfaatan danPelestarian Plasma Nutfah Pertanian(BP4NP), selain dana RM juga mendapatalokasi dana dari Proyek Pembinaan Pembangunan Penelitian dan PengembanganPertanian (ARMP-II) dengan dana LoanIBRD No. 3886-IND, Kategori IncrementalOperating Cost Fjscal Year 2002.
Berdasarkan DIP No. 022/XVIII/1/-/2002tanggal 1 Januari 2002, TA 2002 sebesarRp 1.025.337.000 terdiri dari Rp 259.810.000(RM) dan Rp 765.527.000 (RK) dengan per-bandingan 25% (RM) dan 75% (RK).
Dibandingkan dengan TA 2001, ter-dapat kenaikan anggaran sebesar
Rp 860.750.000 atau 19,94% yang dibiayaidari RM dan RK. Dengan demikian, per-bandingan antara biaya penelitian dengannonpenelitian 80,06% berbanding 19,94%.
Realisasi anggaran secara keseluruhanmencapai Rp 4.076.781.157. Apabila berda-sarkan tolok ukur, administrasi proyek/peraiatan (nonpenelitian) mencapaiRp 740.595.515 yang dibiayai dari RM danRK. Dengan demikian, sisa mati anggaranditambah dengan sisa UYHD yang disetorke Kantor Perbendaharaan dan Kas Negara(KPKN) Bogor Rp 239.668.843 yang terdiridari Rp 216.220.871 RM dan Rp 23.447.972RK. Sebagian besar dana yang dikembali-kan khususnya dari RM adalah sisa dariTPP/HPP dan sisa hasil perhitungan pajakantara RM dengan RK sehingga dananyatidak dapat digunakan.
643.164.750
2.283.251
3.000
3.000
22.735
21.440
1.295
2.257.516
6.654.974.250
1.275.315.749
17.203.000
17.203.000
289.314.265
69.978.56048.500.000
170.835.705
968.798.484
7.298.139.000
1.277.599.000
17.206.000
17.206.000
289.337.000
70.000.00048.500.000
170.837.000
971.056.000
Jumlah seluruhnya
Jumlah belanja non pegawai
Jumlah IV
IV. Belanja perjalanan5410
Jumlah III
III. Belanja pemeliharaan
531053305350
Jumlah II
2.244.26613.000
250
10.400.000863.411.734
19.987.00074.999.750
10.400.000865.656.000
20.000.00075.000.000
5250523052205210II. Belanja barang
640.881.499
559.865.05981.015.240
1.200
Sisa dana (Rp)
5.379.658.501
5.025.449.941333.383.760
20.824.800,
Realisasi dana (Rp)
6.020.540.000
5.585.315.000414.399.000
20.826.000
Pagu dalam DIK (Rp)
Jumlah I
1. Belanja pegawai
511051205150
Kegiatan/MAK
Tabel V.20. Realisasi anggaran belanja rutin Balitbiogen TA 2002
Laporan Tahunan Balitbio 2002

142
Loan IBRD No. 3886-IND, KategoriIncremental Operating Cost Fiscal Year2002.
Berdasarkan DIP No. 022/XVII1/1/-/2002 tanggal 1 Januari 2002, TA 2002sebesar Rp 485.233.000 terdiri dariRp 136.086.000 (RM) dan Rp 349.147.000(RK) dengan perbandingan 28% (RM) dan72% (RK).
Dana pembangunan/proyek TA 2002sebesar Rp 485.233.000 digunakan untukmembiayai kegiatan administrasi proyek,pengadaan perlengkapan sarana gedung,meubelair, pengembangan sistem informa
si, dan penyuluhan/penyebaran informasi.
Realisasi anggaran sampai akhirDesember 2002 mencapai Rp 451.429.211.Apabila berdasarkan tolok ukur administrasi proyek/non penelitian mencapaiRp 60.509.072 dibiayai dari RM dan RK.Dengan demikian, sisa mati anggaranditambah UYHD yang disetor ke KPKNBogor Rp 33.803.789 yang terdiri dariRp 19.646.237 (RM) dan Rp 14.157.552(RK).
Kerja Sama Penelitian
Biaya Penelitian Balitbiogen TA 2002selain diperoleh dari APBN, juga dari kerjasama penelitian. Jumlah dana kerja samapenelitian Rp 264.830.000 terdiri dari RisetUnggulan terpadu (RUT). Secara keselu-ruhan dana penelitian dan dana kerja samapenelitian mencapai Rp 4.847.402.000(TabelV.21).
Realisasi penerimaan anggaran kerjasama penelitian mencapai Rp 264.533.450sedangkan sisa anggaran yang tidak ter-serap Rp 296.550 (Tabel V.22).
Rp 656.000, yaitu dari Rp 1.024.681.000 (TA2001) menjadi Rp 1.025.337.000 (TA 2002).
Dana pembangunan/proyek TA 2002sebesar Rp 1.025.337.000 yang terdiri dariRp 259.810.000 (RM) dan Rp 765.527.000(RK) digunakan untuk membiayai kegiatanadministrasi proyek, pengelolaan keaneka-
ragaman hayati, pengadaan meubelair/alatpengolah data, penyuluhan dan penyebar-an informasi, serta kegiatan pemantauan
dan evaluasi.
Pada TA 2002, di BP4NP terdapat kegiatan Pengelolaan KeanekaragamanHayati (satu judul) yang terdiri dari dela-pan kegiatan dengan dana Rp 791.039.000sehingga rata-rata biaya per kegiatanRp 98.879.875. Sedangkan biaya nonpeneli-tian (administrasi proyek, pengadaan meu-
belair, alat pengolah data, penyuluhan, pe-nyebaran informasi dan pemantauan/eva-
luasi) Rp 234.298.000 (22%) yang dibiayaidari RM dan RK. Dengan demikian perban-dingan antara biaya kegiatan keanekaragaman hayati dengan kegiatan non pene-litian 78% berbanding 22%.
Realisasi anggaran mencapaiRp 928.721.055. Apabila berdasarkan tolokukur administrasi proyek/non penelitianmencapai Rp 200.417.091 dibiayai dari RMdan RK. Dengan demikian, sisa mati anggaran ditambah dengan sisa UYHD yangdisetor ke KPKN Bogor Rp 96.615.945 yangterdiri dari Rp 44.791.249 (RM) danRp 51.824.696 (RK).
Bagian Proyek Manajemen Varietas
Anggaran Belanja Pembangunan TA2002 Bagian Proyek Manajemen VarietasBogor, selain dana RM juga mendapatalokasi dana BLN dari Proyek PembinaanPembangunan Penelitian dan Pengem-bangan Pertanian (ARMP-II) dengan dana
SUMBER DAYA DAN MANAJEMEN

143
ministrasi kantor diperlukan alat tulis kantor (ATK). Realisasi pembelian ATK danalat kebersihan untuk tahun 2002'Rp 74.999.750. Tata cara pengadaan baranguntuk anggaran rutin mengacu kepadaKepres No. 16 tahun 2000.
Pada TA 2002, telah dilakukan pemeliharaan gedung kantor Kelompok Peneliti(Kelti) RP, Kelti SDG, dan sekretariat dengan biaya Rp 33.040.260, pengecatan KeltiBM Rp 28.438.300, dan pemeliharaan kantor Inlitbio Pacet dan Inlitbio Citayam sebe-sar Rp 8.500.000 yang diambil dari mataanggaran 5310.
Mata anggaran 5350 (Iain-lain pemeliharaan) digunakan untuk perbaikan garasi,bengkel, dan Laboratorium Bank Gen dan
PERLENGKAPAN DAN RUMAH TANGGADukungan Anggaran Rutin terhadap
Kegiatan Balitbiogen
Kegiatan Perlengkapan dan RumahTangga Balitbiogen nrteliputi pengadaanbarang, pemeliharaan gedung kantor danfasilitas peneiitian, surat menyurat peng-gajian, serta kendaraan dinas.
Pelaksanaan kegiatan penelitian Balitbiogen perlu ditunjang oleh bahan dan per-alatan yang memadai. Pengadaan baranginventaris pada TA 2002 mencapaiRp 19.987.000. Dibandingkan dengan TA2001, pengadaan barang inventaris meng-alami kenaikan Rp 897.000 (4,5%). Untukmenunjang kelancaran pelaksanaan ad-
264.830.000 264.533.450 296.550Jumlah
097.59/SK/RUT/2002, RUT/LIPI 79.000.000 78.992.800 7.20029 Januari 2002
097.59/SK/RUT/2002, RUT/LIPI 90.360.000 90.098.859 261.14129 Januari 2002
097.59/SK/RUT/2002, RUT/LIPI 95.470.000 95.441.791 28.20929 Januari 2002
Pembentukan buah partenokarpipada tanaman salak melaluirekayasa genetikaPeningkatan ketahanan terhadapaluminium pada tanaman kedelaimelalui kultur in vitroReduksi pencemaran lahan pertanian oleh senyawa logam beratdengan teknologi bioremediasi
3.
Sumber Jumlah dana Realisasi Sisadana(Rp)(Rp)(Rp)
Nomor kontrakNo. Judul penelitian
Tabel V.22. Realisasi anggaran kerja sama penelitian Balitbiogen TA 2002
anggaran pendapatan dan belanja negara, RUT = risetRPTP = rencana penelitian tingkat peneliti, APBNunggulan terpadu
4.847.402.0007920Jumlah
3.455.700.000791.039.000335.833.000
264.830.000
6682
1412
APBNProyek Penelitian Genetika dan Bioteknologi BogorBagian Proyek Pemanfaatan dan Peiestarian Plasma Nutfah PertanianBagian Proyek Manajemen Varietas
KERJA SAMA PENELITIANRUT
Biaya (Rp)RPTP Kegiatan
JumlahSumber dana
Tabel V.21. Jumlah RPTP, kegiatan dan biaya penelitian berdasarkan sumber dana TA 2002
Laporan Tahunan Balitbio 2002

144
mengalamai kenaikan Rp 14.250.000dibandingkan dengan TA 2001 sebesarRp 57.600.000. Sedangkan biaya operasional 10 unit kendaraan roda empat yangdialokasikan dari anggaran rutin sebesarRp 58.000.000 atau mengalami kenaikanRp 9.500.000 apabila dibandingkan denganTA 2001 sebesar Rp 48.500.000.
Biaya eksploitasi kendaraan tersebutselain digunakan untuk biaya pemelihara-an, juga untuk pembelian bahan bakar danpengurusan STNK.
Genetika, jalan kantor, LaboratoriumMikroskop Elektron, gedung mekanikal danelektrik serta Mess Inlitbio Pacet.
Kendaran dinas yang dimiliki Balitbio-gen pada TA 2002 sebanyak 23 unit kendaran roda empat dan 11 unit kendaraanroda dua. Sebanyak 13 unit kendaraan rodaempat dan 11 unit kendaraan roda dua di-biayai dari anggaran proyek sedangkan 10unit kendaraan dinas roda empat dibiayaidari anggaran rutin. Biaya operasional kendaraan dinas yang dialokasikan dari anggaran proyek sebesar Rp 71.850.000 atau
SUMBER DAYA DAN MANAJEMEN

146
Kepala Instalasi PerpustakaanKepala Instalasi Rumah KacaKepala Instalasi Bengkel Peralatan
Jabatan
Jetty SuhaetiMulyotoYusuf Sujatman
Nama
Personalia (lanjutan)
Hasil Penelitian 2001