prefrontal regulation of punished ethanol self-administration
TRANSCRIPT

iologicalsychiatry
Archival Report BP
Prefrontal Regulation of Punished EthanolSelf-administration
Lindsay R. Halladay, Adrina Kocharian, Patrick T. Piantadosi, Michael E. Authement,Abby G. Lieberman, Nathen A. Spitz, Kendall Coden, Lucas R. Glover, Vincent D. Costa,Veronica A. Alvarez, and Andrew HolmesISS
ABSTRACTBACKGROUND: A clinical hallmark of alcohol use disorder is persistent drinking despite potential adverse conse-quences. The ventromedial prefrontal cortex (vmPFC) and dorsomedial prefrontal cortex (dmPFC) are positioned toexert top-down control over subcortical regions, such as the nucleus accumbens shell (NAcS) and basolateralamygdala, which encode positive and negative valence of ethanol (EtOH)-related stimuli. Prior rodent studies haveimplicated these regions in regulation of punished EtOH self-administration (EtOH-SA).METHODS: We conducted in vivo electrophysiological recordings in mouse vmPFC and dmPFC to obtain neuronalcorrelates of footshock-punished EtOH-SA. Ex vivo recordings were performed in NAcS D1 receptor–expressingmedium spiny neurons receiving vmPFC input to examine punishment-related plasticity in this pathway.Optogenetic photosilencing was employed to assess the functional contribution of the vmPFC, dmPFC, vmPFCprojections to NAcS, or vmPFC projections to basolateral amygdala, to punished EtOH-SA.RESULTS: Punishment reduced EtOH lever pressing and elicited aborted presses (lever approach followed by rapidretraction). Neurons in the vmPFC and dmPFC exhibited phasic firing to EtOH lever presses and aborts, but only inthe vmPFC was there a population-level shift in coding from lever presses to aborts with punishment. Closed-loopvmPFC, but not dmPFC, photosilencing on a postpunishment probe test negated the reduction in EtOH leverpresses but not in aborts. Punishment was associated with altered plasticity at vmPFC inputs to D1 receptor–expressing medium spiny neurons in the NAcS. Photosilencing vmPFC projections to the NAcS, but not to thebasolateral amygdala, partially reversed suppression of EtOH lever presses on probe testing.CONCLUSIONS: These findings demonstrate a key role for the vmPFC in regulating EtOH-SA after punishment, withimplications for understanding the neural basis of compulsive drinking in alcohol use disorder.
Keywords: Alcohol, Amygdala, Compulsive, Corticostriatal, Infralimbic, Mouse, Nucleus accumbens
https://doi.org/10.1016/j.biopsych.2019.10.030
Excessive alcohol drinking is the third leading cause of pre-ventable death in the United States (1). By recent estimates,over 15 million adults are affected by alcohol use disorder,while a quarter of the adult population reports recentlyengaging in either heavy alcohol use or binge drinking (1). Ahallmark of alcohol use disorder is that drinking persistsdespite an awareness of the potential adverse consequences(2). Despite a large body of work describing brain mechanismsunderlying alcohol seeking, the neural circuits that exert con-trol over the drive to use alcohol and other drugs in the face ofpotential negative outcomes remain poorly understood (3–10).
There is growing evidence from human and rodent studiesthat the medial prefrontal cortex (mPFC) modulates ethanolself-administration (EtOH-SA), and the mPFC has been shownto either reduce or enable various measures of EtOH seekingand drinking (4,5). For instance, cue-induced reinstatement ofEtOH consumption is abolished by ablating neuronal ensem-bles in the ventromedial PFC (vmPFC) but not in the
SEE COMMENTARY ON PAGE 940; SE
ª 2019 Society of Biological Psychiatry. PubliCC BY-NC-ND
N: 0006-3223 B
dorsomedial PFC (dmPFC) (11,12). The mPFC is also wellpositioned, anatomically, to arbitrate between EtOH seekingand avoidance via its connections with amygdaloid and striatalregions (13–17).
Prior studies have found that the development of resistanceto EtOH-SA is associated with plasticity in mPFC projectionsto the nucleus accumbens (NAc) core (18), and that dis-connecting the dmPFC from the NAc core biases rats towardmaking disadvantageous decisions (19,20). Moreover, recentdata has shown that the shell region of the NAc (NAcS) and thebasolateral amygdala (BLA), as well as mPFC connections withthese regions, play a key role in regulating responding tonondrug rewards in the presence of reward-associated cuesand potentially aversive outcomes (21–27). These circuits arealso affected by EtOH exposure. For example, EtOH depen-dence was accompanied by insertion of GluA2-lacking AMPAreceptors at glutamatergic synapses in NAc medium spinyneurons (MSNs) (28) and increased excitability and amplitude
E ALSO VIDEO CONTENT ONLINE
shed by Elsevier Inc. This is an open access article under thelicense (http://creativecommons.org/licenses/by-nc-nd/4.0/).
967
iological Psychiatry June 1, 2020; 87:967–978 www.sobp.org/journal

Punishment and the Prefrontal CortexBiologicalPsychiatry
of NMDA receptor–mediated synaptic responses in NAc D1
receptor–expressing MSNs (D1-MSNs) (29). Furthermore,NMDA receptors in the NAc and mPFC are implicated inpunishment-resistant EtOH drinking in rats (18,30), and arecent elegant study found that activity of NAc-projectingvmPFC neurons predicted less cocaine seeking (27).
Taken together, these prior studies suggest that thevmPFC, BLA, and NAc are part of an integrated circuit regu-lating the seeking, consumption, and, in the face of negativeoutcomes, avoidance of EtOH and other drugs of abuse(31–36). Nonetheless, understanding of the contributions of thevmPFC and its downstream connections to the BLA and NAcSto the regulation of punished EtOH-SA remains incomplete.The aim of the current study was to clarify this role using acombination of in vivo and ex vivo neuronal recordings andoptogenetic manipulations in a mouse assay for punishedEtOH-SA.
METHODS AND MATERIALS
Subjects
Subjects were male (7–8 weeks old) C57BL/6J and B6.Cg-Tg(Drd1a-tdTomato)6Calak/J mice obtained from The Jack-son Laboratory (Bar Harbor, ME), gradually reduced to 85% oftheir free-feeding body weight before testing.
Ethical Considerations
All experimental procedures were approved by the NationalInstitute on Alcohol Abuse and Alcoholism Animal Care andUse Committee and followed the National Institutes of Healthguidelines outlined in Using Animals in Intramural Research.
Behavioral Testing
Mice were trained in a Med Associates operant chamber (MedAssociates, Inc., Fairfax, VT) to respond on the left of two le-vers on a continuous schedule of reinforcement to earn su-crose pellets during 40-minute sessions (the right, “inactive”lever had no programed consequences) until the criterion wasmet ($35 rewards), as previously described (37–39). The pelletwas then substituted with a progression of 10-mL liquid re-wards (“sucrose fading”; 10% sucrose, 10% sucrose 1 10%EtOH, 5% sucrose 1 10% EtOH, 10% EtOH). Training at eachEtOH concentration stage continued until performance wasstable (,20% coefficient between reward lever responses over3 sessions).
There was then a single punishment session consisting of abaseline period (10 rewarded/unpunished EtOH lever presses)immediately followed (after the 10th press) by a 40-minutepunishment period (EtOH lever presses alternatingly pro-duced 10% EtOH or no EtOH 1 footshock). A probe test(identical to rewarded training sessions) was conducted 24hours later. For in vivo recordings, a second probe was con-ducted 24 hours following the first, with data collapsed acrossprobes for analyses. EtOH lever presses, inactive presses, andhead entries into the reward receptacle were recorded by Med-PC IV software (Med Associates, Inc.). Approaches to theEtOH lever with an elongated body posture, followed by rapidbody retraction without execution of press, were scored
968 Biological Psychiatry June 1, 2020; 87:967–978 www.sobp.org/jo
manually (Supplemental Video 1) and referred to as aborts,keeping with the definition of a similar behavior in rats (40).
In Vivo Neuronal Recordings
Following training, fixed microelectrode arrays were surgicallyimplanted in the bilateral vmPFC and dmPFC of the samemouse (41,42). Recordings were conducted during punishmentand probe testing using OmniPlex Neural Data AcquisitionSystem (Plexon Inc., Dallas, TX) combined with simultaneousCinePlex Behavioral Research Systems (Plexon Inc.) (41). Themean firing rate of recorded cells was 6.33 6 0.90 for thevmPFC and 4.12 6 0.46 Hz for the dmPFC.
Spike timestamp information was integrated andanalyzed using NeuroExplorer, version 5 (Nex Technologies,Colorado Springs, CO). For each unit, spikes were time-locked to the onset of behavioral events. Spiking activity1 second prior to event onset was used as baseline. Usinga sliding window (300 ms) stepped in 10-ms bins from 22.5seconds to 12.5 seconds around each event, we computedt tests to determine when the activity surrounding eachevent deviated from baseline; units with significant de-viations were considered responsive to an event for thattime bin.
Fluorescence In Situ Hybridization
Immediately after probe testing, brains were collected toperform fluorescence in situ hybridization using the RNAscopeMultiplex Fluorescent Assay Kit (Advanced Cell Diagnostics,Inc., Newark, CA) (42) to label vmPFC fos (c-Fos), Slc17a7(VGLUT1), Gad1 (GAD67), Drd1a (DR1A), and Drd2 (DR2). Weused ImageJ, version 1.50 (National Institutes of Health,Bethesda, MD) to calculate percentage of DAPI-labeled neu-rons positive for fos, and of those, the percentage that wereSlc17a7-, Gad1-, Drd1a-, and Drd2-positive.
Ex Vivo Whole-Cell Recordings
AAV5/CaMKIIa-hChR2(H134R)-eYFP was bilaterally injectedinto the vmPFC of D1-tdTomato mice. Mice underwentbehavioral testing and were sacrificed on probe day. Cells inNAcS brain sections were voltage clamped at 140 mV (ingabazine). tdTomato-expressing D1-MSNs and nonfluorescentputative D2-MSNs were identified using blue light pulses tooptically evoke excitatory postsynaptic currents (EPSCs).AMPA-mediated EPSCs were pharmacologically isolated us-ing (R)-CPP, NMDA antagonist (ab120159) (Abcam, Cam-bridge, MA) and subtracted from combined EPSCs to obtainthe NMDA-mediated EPSC component (28). AMPA:NMDAratios were calculated by dividing the peak receptor-mediatedEPSC by the peak NMDA-mediated EPSC. For the rectificationindex, AMPA-mediated EPSCs were measured at both nega-tive and positive (255/140 mV) holding potentials, and sper-mine was included in the internal solution. The rectificationindex was calculated by dividing the peak current at 255 mVby the peak current at 140 mV.
In Vivo Photosilencing
rAAV8/CAG-ArchT-GFP or rAAV8/CAG-GFP (local PFCsilencing) or AAV5/CaMKIIa-eArchT3.0-eYFP or AAV5/CaMKII-eYFP (vmPFC projection silencing) was bilaterally injected into
urnal
B
CEtOH-lever
presses
Inactive-lever presses
Aborts
Number per session
EtOH-lever presses
Inactive-lever presses
Aborts
0 8 16 24 32 40 48 0 8 16 24 32 40 48 0 8 16 24 32 40 48
** *
pre-shockbaseline
shock
Star
t
End
Star
t
Star
t
End
*
EtOH-lever
Inactive-lever
EtOH-lever EtOH
Shock/no EtOHEtOH-lever
EtOH EtOH-lever EtOH
A Training Punishment Probe
EtOH-lever
Inactive-lever
EtOH-lever
Inactive-leverEtOH EtOH EtOH
1 day
1-2 days
0 8 16 24 32 40 48
*St
artrr
*
EtOH-lever EtOH
Probe
EtOH-lever
Inactive-leverEtOH
0 8 16 24 32 40 48
**
pre-shockbaseline
shock
Star
trr
End
Shock/no EtOHEtOH-lever
EtOH
Punishment
EtOH-lever
Inactive-leverEtOH
1-2days
0 8 16 24 32 40 48
Star
trr
End
EtOH-lever
Inactive-lever
EtOH-lever EtOH
Training
EtOH
1day
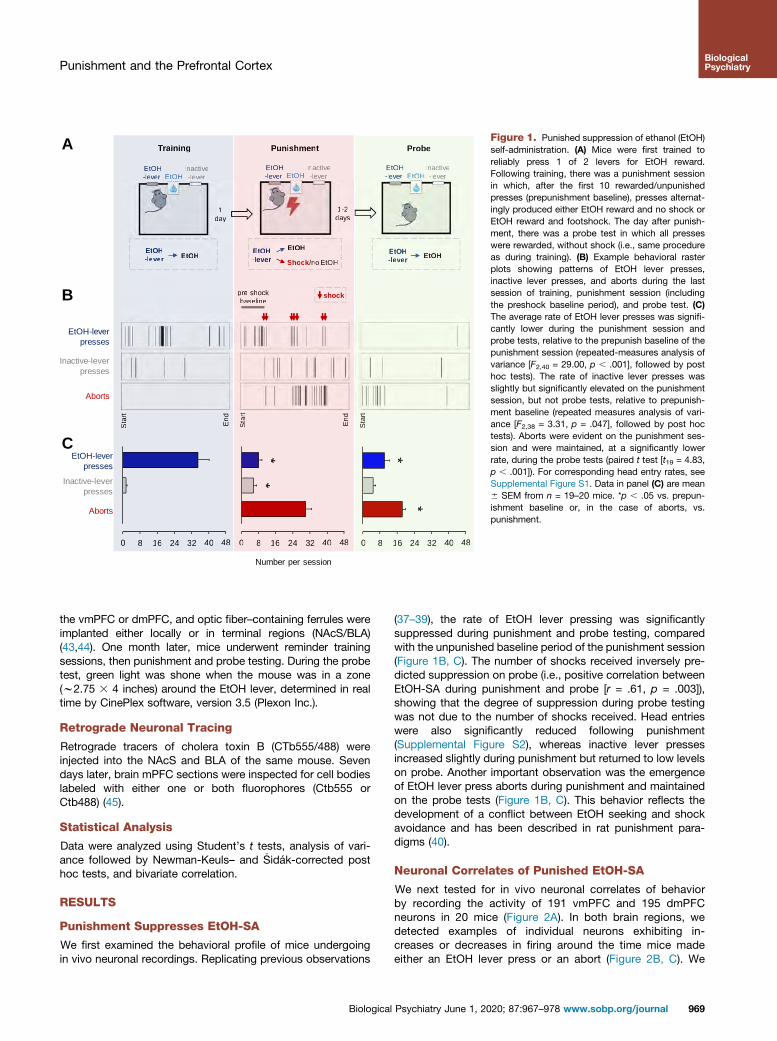
Figure 1. Punished suppression of ethanol (EtOH)self-administration. (A) Mice were first trained toreliably press 1 of 2 levers for EtOH reward.Following training, there was a punishment sessionin which, after the first 10 rewarded/unpunishedpresses (prepunishment baseline), presses alternat-ingly produced either EtOH reward and no shock orEtOH reward and footshock. The day after punish-ment, there was a probe test in which all presseswere rewarded, without shock (i.e., same procedureas during training). (B) Example behavioral rasterplots showing patterns of EtOH lever presses,inactive lever presses, and aborts during the lastsession of training, punishment session (includingthe preshock baseline period), and probe test. (C)The average rate of EtOH lever presses was signifi-cantly lower during the punishment session andprobe tests, relative to the prepunish baseline of thepunishment session (repeated-measures analysis ofvariance [F2,40 = 29.00, p , .001], followed by posthoc tests). The rate of inactive lever presses wasslightly but significantly elevated on the punishmentsession, but not probe tests, relative to prepunish-ment baseline (repeated measures analysis of vari-ance [F2,38 = 3.31, p = .047], followed by post hoctests). Aborts were evident on the punishment ses-sion and were maintained, at a significantly lowerrate, during the probe tests (paired t test [t19 = 4.83,p , .001]). For corresponding head entry rates, seeSupplemental Figure S1. Data in panel (C) are mean6 SEM from n = 19–20 mice. *p , .05 vs. prepun-ishment baseline or, in the case of aborts, vs.punishment.
Punishment and the Prefrontal CortexBiologicalPsychiatry
the vmPFC or dmPFC, and optic fiber–containing ferrules wereimplanted either locally or in terminal regions (NAcS/BLA)(43,44). One month later, mice underwent reminder trainingsessions, then punishment and probe testing. During the probetest, green light was shone when the mouse was in a zone(w2.75 3 4 inches) around the EtOH lever, determined in realtime by CinePlex software, version 3.5 (Plexon Inc.).
Retrograde Neuronal Tracing
Retrograde tracers of cholera toxin B (CTb555/488) wereinjected into the NAcS and BLA of the same mouse. Sevendays later, brain mPFC sections were inspected for cell bodieslabeled with either one or both fluorophores (Ctb555 orCtb488) (45).
Statistical Analysis
Data were analyzed using Student’s t tests, analysis of vari-ance followed by Newman-Keuls– and �Sidák-corrected posthoc tests, and bivariate correlation.
RESULTS
Punishment Suppresses EtOH-SA
We first examined the behavioral profile of mice undergoingin vivo neuronal recordings. Replicating previous observations
Biological
(37–39), the rate of EtOH lever pressing was significantlysuppressed during punishment and probe testing, comparedwith the unpunished baseline period of the punishment session(Figure 1B, C). The number of shocks received inversely pre-dicted suppression on probe (i.e., positive correlation betweenEtOH-SA during punishment and probe [r = .61, p = .003]),showing that the degree of suppression during probe testingwas not due to the number of shocks received. Head entrieswere also significantly reduced following punishment(Supplemental Figure S2), whereas inactive lever pressesincreased slightly during punishment but returned to low levelson probe. Another important observation was the emergenceof EtOH lever press aborts during punishment and maintainedon the probe tests (Figure 1B, C). This behavior reflects thedevelopment of a conflict between EtOH seeking and shockavoidance and has been described in rat punishment para-digms (40).
Neuronal Correlates of Punished EtOH-SA
We next tested for in vivo neuronal correlates of behaviorby recording the activity of 191 vmPFC and 195 dmPFCneurons in 20 mice (Figure 2A). In both brain regions, wedetected examples of individual neurons exhibiting in-creases or decreases in firing around the time mice madeeither an EtOH lever press or an abort (Figure 2B, C). We
Psychiatry June 1, 2020; 87:967–978 www.sobp.org/journal 969
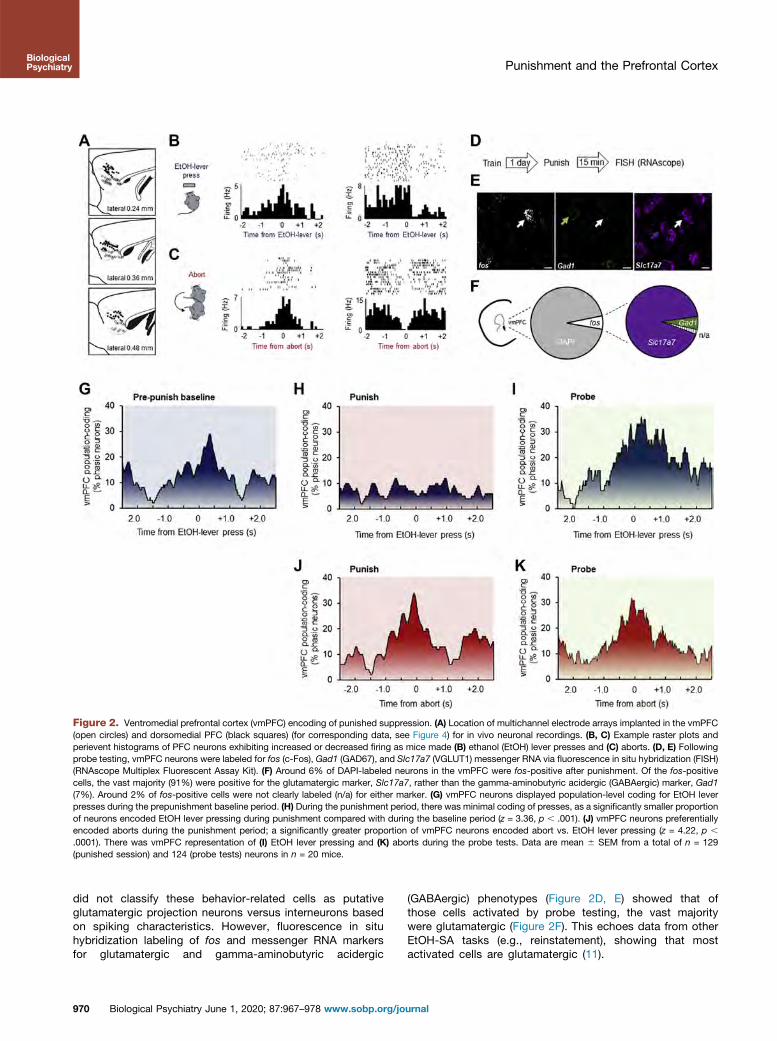
Figure 2. Ventromedial prefrontal cortex (vmPFC) encoding of punished suppression. (A) Location of multichannel electrode arrays implanted in the vmPFC(open circles) and dorsomedial PFC (black squares) (for corresponding data, see Figure 4) for in vivo neuronal recordings. (B, C) Example raster plots andperievent histograms of PFC neurons exhibiting increased or decreased firing as mice made (B) ethanol (EtOH) lever presses and (C) aborts. (D, E) Followingprobe testing, vmPFC neurons were labeled for fos (c-Fos), Gad1 (GAD67), and Slc17a7 (VGLUT1) messenger RNA via fluorescence in situ hybridization (FISH)(RNAscope Multiplex Fluorescent Assay Kit). (F) Around 6% of DAPI-labeled neurons in the vmPFC were fos-positive after punishment. Of the fos-positivecells, the vast majority (91%) were positive for the glutamatergic marker, Slc17a7, rather than the gamma-aminobutyric acidergic (GABAergic) marker, Gad1(7%). Around 2% of fos-positive cells were not clearly labeled (n/a) for either marker. (G) vmPFC neurons displayed population-level coding for EtOH leverpresses during the prepunishment baseline period. (H) During the punishment period, there was minimal coding of presses, as a significantly smaller proportionof neurons encoded EtOH lever pressing during punishment compared with during the baseline period (z = 3.36, p , .001). (J) vmPFC neurons preferentiallyencoded aborts during the punishment period; a significantly greater proportion of vmPFC neurons encoded abort vs. EtOH lever pressing (z = 4.22, p ,
.0001). There was vmPFC representation of (I) EtOH lever pressing and (K) aborts during the probe tests. Data are mean 6 SEM from a total of n = 129(punished session) and 124 (probe tests) neurons in n = 20 mice.
Punishment and the Prefrontal CortexBiologicalPsychiatry
did not classify these behavior-related cells as putativeglutamatergic projection neurons versus interneurons basedon spiking characteristics. However, fluorescence in situhybridization labeling of fos and messenger RNA markersfor glutamatergic and gamma-aminobutyric acidergic
970 Biological Psychiatry June 1, 2020; 87:967–978 www.sobp.org/jo
(GABAergic) phenotypes (Figure 2D, E) showed that ofthose cells activated by probe testing, the vast majoritywere glutamatergic (Figure 2F). This echoes data from otherEtOH-SA tasks (e.g., reinstatement), showing that mostactivated cells are glutamatergic (11).
urnal

Punishment and the Prefrontal CortexBiologicalPsychiatry
vmPFC Coding Punished of EtOH-SA
We next examined population-level mPFC coding (46) byaligning neuronal firing in a 5-second window around behav-ioral events during the prepunishment baseline, punishment,and probe tests. During baseline, we found clear population-level representation of EtOH lever presses in the vmPFC(w30% of units, n = 65); neuronal engagement increased overthe 1.5 seconds prior to execution of a lever press and peakedslightly after a press was made (Figure 2G). During the pun-ished phase, press-related activity of vmPFC neurons wasminimal (Figure 2H), but there was strong coding of aborts(w38% of vmPFC units) beginning around 0.5 second prior tothe mouse retracting away from the EtOH lever (Figure 2I).Notably, this vmPFC neuronal coding of aborts was main-tained during probe testing, while coding of EtOH leverpresses returned to levels evident during punishment, despitethe fact that the number of presses was still much lower(Figure 2J, K). To determine whether EtOH lever and abortevents were encoded by distinct neural populations in vmPFC,we conducted a correlational analysis on units that wereresponsive to either event. Interestingly, only 2 of all unitsrecorded exhibited firing rates during the 2 events that corre-lated significantly, suggesting that these behaviors are repre-sented by separate ensembles in vmPFC.
These in vivo neuronal data demonstrate that vmPFC neu-rons exhibit representations of the execution of a response forEtOH and avoidance of that same response in a manner thatshifts, depending on the prevailing task demands: prepunish-ment = press, ongoing punishment = abort. This agrees withprevious studies showing that the vmPFC can contribute to thepromotion or inhibition of reward seeking across differentexperimental situations (31,34,47–53). The current data alsoshow that during probe testing, when there is prior knowledgeof both the reward and punishment outcomes for responding,the vmPFC strongly codes for both lever pressing and aborts,consistent with a role in arbitrating between opposing actionsunder conflict but not with a simple response-suppressionfunction.
Silencing the vmPFC Abolishes PunishedSuppression of EtOH-SA
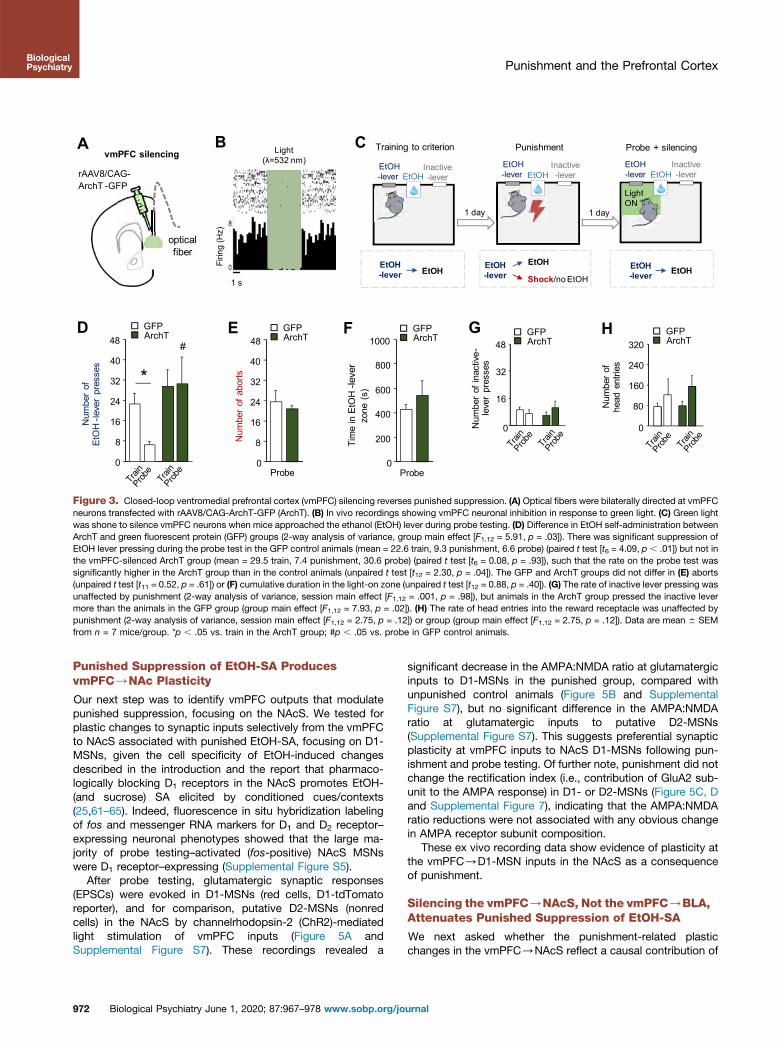
Our recording data demonstrate vmPFC neuronal correlates ofpunished suppression of EtOH-SA but do not address causalcontribution of vmPFC to performance. Therefore, we deviseda closed-loop optogenetic approach to test the consequencesof photosilencing archaerhodopsin T (ArchT)-transfectedvmPFC neurons when mice were near the EtOH lever duringthe probe test (Figure 3A–C).
We found that silencing vmPFC neurons in this mannerabolished punishment-induced suppression of EtOH-SA.Specifically, green fluorescent protein (GFP)-expressing con-trol mice exhibited a significant decrease in EtOH leverpressing during probe testing, relative to the last training dayprior to punishment, whereas there was no significant changein ArchT animals, such that their probe test lever pressing wassignificantly higher than in GFP mice (Figure 3D). Notably,despite this silencing-induced normalization of lever pressing,aborts exhibited by the groups did not differ (Figure 3E), nor didthe cumulative or average time spent in the light-on zone
Biological
proximal to the EtOH lever (Figure 3F), or inactive lever presses(Figure 3G). There was a small but significant increase in headentries in ArchT mice but not in GFP control animals(Figure 3H), which likely reflects more frequent reward collec-tion/checking in the higher-pressing ArchT group, rather thangreater motivation for the EtOH reward following vmPFCsilencing. Indeed, prior studies in rats reported negative (54,55)or demotivating (56–59) effects on reward seeking after vmPFClesioning and pharmacological inactivation.
In a control experiment, mice underwent the samebehavioral testing and surgical procedures, and then, onreaching the training criterion, were given a pseudo-probetest (i.e., without punishment), during which the vmPFC wassilenced (exactly as above). All behavioral parameters weresimilar between the ArchT and GFP groups (SupplementalFigure S13), demonstrating that vmPFC silencing was insuf-ficient to alter behavior in the absence of punishment,excluding the possibility that increased EtOH-SA in punishedmice following vmPFC silencing was due to a general in-crease in motivation for EtOH-SA.
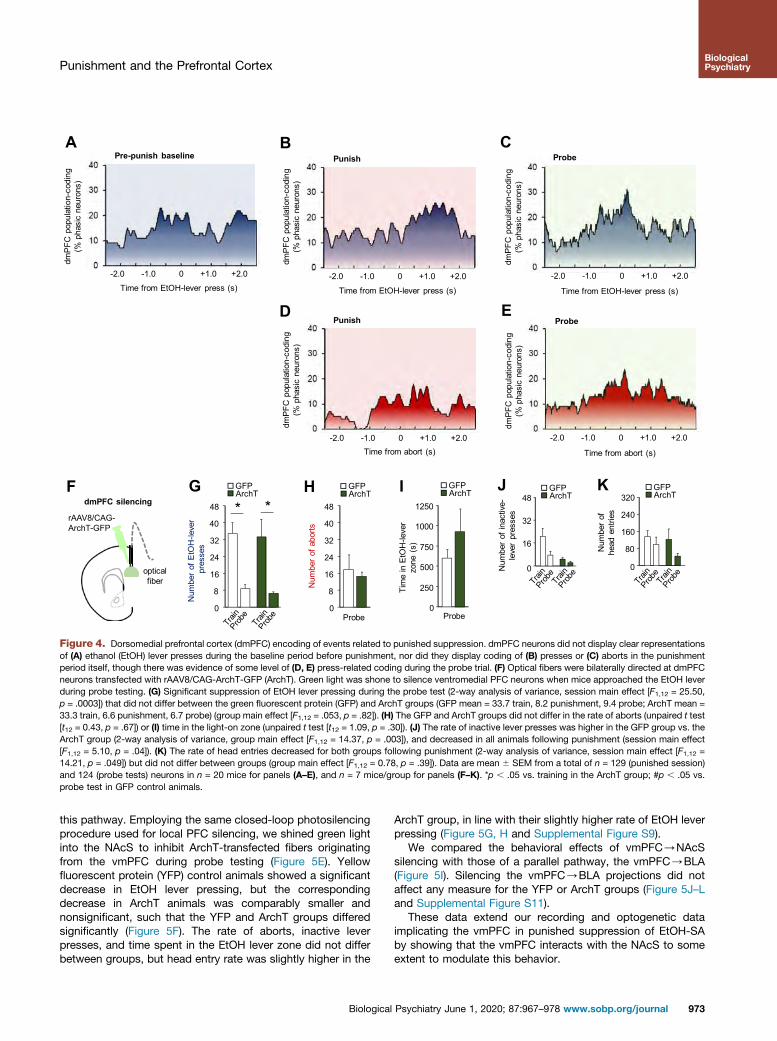
dmPFC Coding of Punished EtOH-SA
We next determined whether the contribution of the vmPFC topunished EtOH-SA was shared by the neighboring dmPFC.Recorded units in the dmPFC did not show the same shifts intask phase–related population coding seen in the vmPFC,though there was moderate dmPFC activity associated withEtOH lever and abort events during probe testing (Figure 4A–E).Prior work has suggested that the dmPFC promotes reward SA(31), a notion supported by recent reports of in vivo calciumtransients evident during sucrose cue presentation in thedmPFC and dmPFC/NAc core cells (60). Indeed, data hereindicate an absence of clear neuronal correlates in the dmPFCduring punishment, but it remains possible that representationsare extant but restricted to discrete dmPFC ensembles that aredifficult to resolve with neuronal recordings. Relatedly, a recentreport found that a small subset of shock-activated NAcS-projecting dmPFC cells exhibited calcium correlates duringavoidance of punished reward (22).
dmPFC Photosilencing Does Not Affect PunishedSuppression of EtOH-SA
We next assessed the consequences of photosilencing thedmPFC, employing the same closed-loop design describedabove (Figure 4F), anticipating that the lack of EtOH-SA cor-relates in the dmPFC would correspond with minimal func-tional relevance. In contrast to vmPFC effects, followingdmPFC silencing, EtOH lever press rates in the ArchT and GFPgroups were similarly decreased following punishment(Figure 4G). Likewise, aborts, time spent in the light-on EtOHlever zone, inactive lever presses, and head entries were allunaffected by dmPFC silencing (Figure 4H–K).
These optogenetic data show that in contrast to the vmPFC,the dmPFC is not necessary for the expression of punishedsuppression.
Psychiatry June 1, 2020; 87:967–978 www.sobp.org/journal 971
optical fiber
rAAV8/CAG-ArchT -GFP
B
0
8
16
24
32
40
48D
Num
ber
of
EtO
H-le
ver
pres
ses
#G
0
16
32
48
Num
ber
of in
activ
e-le
ver
pres
ses
GFPArchT
0
80
160
240
320
Num
ber
of
head
ent
ries
H
Firin
g (H
z)
1 s
Light (λ=532 nm)
GFPArchT GFP
ArchT
vmPFC silencingA
E
Num
ber
of a
borts
FTi
me
in E
tOH
-lev
er
zone
(s)
0
8
16
24
32
40
48
0
200
400
600
800
1000
Probe Probe
GFPArchT
GFPArchT
C Training to criterion Punishment Probe + silencing
EtOH-lever
EtOH-lever
Inactive-lever
EtOH-lever
Inactive-lever
Light ONLightON
1 day 1 day
Inactive-lever
EtOH-lever EtOH
Shock/no EtOHEtOH-lever
EtOH EtOH-lever EtOH
EtOH EtOH EtOH
*
Figure 3. Closed-loop ventromedial prefrontal cortex (vmPFC) silencing reverses punished suppression. (A) Optical fibers were bilaterally directed at vmPFCneurons transfected with rAAV8/CAG-ArchT-GFP (ArchT). (B) In vivo recordings showing vmPFC neuronal inhibition in response to green light. (C) Green lightwas shone to silence vmPFC neurons when mice approached the ethanol (EtOH) lever during probe testing. (D) Difference in EtOH self-administration betweenArchT and green fluorescent protein (GFP) groups (2-way analysis of variance, group main effect [F1,12 = 5.91, p = .03]). There was significant suppression ofEtOH lever pressing during the probe test in the GFP control animals (mean = 22.6 train, 9.3 punishment, 6.6 probe) (paired t test [t6 = 4.09, p , .01]) but not inthe vmPFC-silenced ArchT group (mean = 29.5 train, 7.4 punishment, 30.6 probe) (paired t test [t6 = 0.08, p = .93]), such that the rate on the probe test wassignificantly higher in the ArchT group than in the control animals (unpaired t test [t12 = 2.30, p = .04]). The GFP and ArchT groups did not differ in (E) aborts(unpaired t test [t11 = 0.52, p = .61]) or (F) cumulative duration in the light-on zone (unpaired t test [t12 = 0.88, p = .40]). (G) The rate of inactive lever pressing wasunaffected by punishment (2-way analysis of variance, session main effect [F1,12 = .001, p = .98]), but animals in the ArchT group pressed the inactive levermore than the animals in the GFP group (group main effect [F1,12 = 7.93, p = .02]). (H) The rate of head entries into the reward receptacle was unaffected bypunishment (2-way analysis of variance, session main effect [F1,12 = 2.75, p = .12]) or group (group main effect [F1,12 = 2.75, p = .12]). Data are mean 6 SEMfrom n = 7 mice/group. *p , .05 vs. train in the ArchT group; #p , .05 vs. probe in GFP control animals.
Punishment and the Prefrontal CortexBiologicalPsychiatry
Punished Suppression of EtOH-SA ProducesvmPFC/NAc Plasticity
Our next step was to identify vmPFC outputs that modulatepunished suppression, focusing on the NAcS. We tested forplastic changes to synaptic inputs selectively from the vmPFCto NAcS associated with punished EtOH-SA, focusing on D1-MSNs, given the cell specificity of EtOH-induced changesdescribed in the introduction and the report that pharmaco-logically blocking D1 receptors in the NAcS promotes EtOH-(and sucrose) SA elicited by conditioned cues/contexts(25,61–65). Indeed, fluorescence in situ hybridization labelingof fos and messenger RNA markers for D1 and D2 receptor–expressing neuronal phenotypes showed that the large ma-jority of probe testing–activated (fos-positive) NAcS MSNswere D1 receptor–expressing (Supplemental Figure S5).
After probe testing, glutamatergic synaptic responses(EPSCs) were evoked in D1-MSNs (red cells, D1-tdTomatoreporter), and for comparison, putative D2-MSNs (nonredcells) in the NAcS by channelrhodopsin-2 (ChR2)-mediatedlight stimulation of vmPFC inputs (Figure 5A andSupplemental Figure S7). These recordings revealed a
972 Biological Psychiatry June 1, 2020; 87:967–978 www.sobp.org/jo
significant decrease in the AMPA:NMDA ratio at glutamatergicinputs to D1-MSNs in the punished group, compared withunpunished control animals (Figure 5B and SupplementalFigure S7), but no significant difference in the AMPA:NMDAratio at glutamatergic inputs to putative D2-MSNs(Supplemental Figure S7). This suggests preferential synapticplasticity at vmPFC inputs to NAcS D1-MSNs following pun-ishment and probe testing. Of further note, punishment did notchange the rectification index (i.e., contribution of GluA2 sub-unit to the AMPA response) in D1- or D2-MSNs (Figure 5C, Dand Supplemental Figure 7), indicating that the AMPA:NMDAratio reductions were not associated with any obvious changein AMPA receptor subunit composition.
These ex vivo recording data show evidence of plasticity atthe vmPFC/D1-MSN inputs in the NAcS as a consequenceof punishment.
Silencing the vmPFC/NAcS, Not the vmPFC/BLA,Attenuates Punished Suppression of EtOH-SA
We next asked whether the punishment-related plasticchanges in the vmPFC/NAcS reflect a causal contribution of
urnal
A
F
optical fiber
rAAV8/CAG-ArchT-GFP
dmPFC silencingG JH K
0
8
16
24
32
40
48 * *
Num
ber
of E
tOH
-leve
r pr
esse
s
GFPArchT
Num
ber
of
head
ent
ries
GFPArchT
Num
ber
of in
activ
e-le
ver
pres
ses
GFPArchT
I
Num
ber
of a
borts
Tim
e in
EtO
H-le
ver
zone
(s)
Probe
GFPArchT
0
80
160
240
320
0
250
500
750
1000
1250
0
8
16
24
32
40
48
0
16
32
48
Probe
GFPArchT
B
D
C
EPunish
dmPF
C p
opul
atio
n-co
ding
(%
pha
sic
neur
ons)
Probe
dmPF
C p
opul
atio
n-co
ding
(%
pha
sic
neur
ons)
Time from abort (s)-2.0 -1.0 0 +1.0 +2.0
Time from abort (s)
-2.0 -1.0 0 +1.0 +2.0
Pre-punish baseline
noitalupop CFP
md-c
odin
g )snoruen cisahp
%(
Time from EtOH-lever press (s)-2.0 -1.0 0 +1.0 +2.0
Punish
dmPF
C p
opul
atio
n-co
ding
(%
pha
sic
neur
ons)
Time from EtOH-lever press (s)
-2.0 -1.0 0 +1.0 +2.0
Probe
dmPF
C p
opul
atio
n-co
ding
(%
pha
sic
neur
ons)
Time from EtOH-lever press (s)
-2.0 -1.0 0 +1.0 +2.0
Figure 4. Dorsomedial prefrontal cortex (dmPFC) encoding of events related to punished suppression. dmPFC neurons did not display clear representationsof (A) ethanol (EtOH) lever presses during the baseline period before punishment, nor did they display coding of (B) presses or (C) aborts in the punishmentperiod itself, though there was evidence of some level of (D, E) press-related coding during the probe trial. (F) Optical fibers were bilaterally directed at dmPFCneurons transfected with rAAV8/CAG-ArchT-GFP (ArchT). Green light was shone to silence ventromedial PFC neurons when mice approached the EtOH leverduring probe testing. (G) Significant suppression of EtOH lever pressing during the probe test (2-way analysis of variance, session main effect [F1,12 = 25.50,p = .0003]) that did not differ between the green fluorescent protein (GFP) and ArchT groups (GFP mean = 33.7 train, 8.2 punishment, 9.4 probe; ArchT mean =33.3 train, 6.6 punishment, 6.7 probe) (group main effect [F1,12 = .053, p = .82]). (H) The GFP and ArchT groups did not differ in the rate of aborts (unpaired t test[t12 = 0.43, p = .67]) or (I) time in the light-on zone (unpaired t test [t12 = 1.09, p = .30]). (J) The rate of inactive lever presses was higher in the GFP group vs. theArchT group (2-way analysis of variance, group main effect [F1,12 = 14.37, p = .003]), and decreased in all animals following punishment (session main effect[F1,12 = 5.10, p = .04]). (K) The rate of head entries decreased for both groups following punishment (2-way analysis of variance, session main effect [F1,12 =14.21, p = .049]) but did not differ between groups (group main effect [F1,12 = 0.78, p = .39]). Data are mean 6 SEM from a total of n = 129 (punished session)and 124 (probe tests) neurons in n = 20 mice for panels (A–E), and n = 7 mice/group for panels (F–K). *p , .05 vs. training in the ArchT group; #p , .05 vs.probe test in GFP control animals.
Punishment and the Prefrontal CortexBiologicalPsychiatry
this pathway. Employing the same closed-loop photosilencingprocedure used for local PFC silencing, we shined green lightinto the NAcS to inhibit ArchT-transfected fibers originatingfrom the vmPFC during probe testing (Figure 5E). Yellowfluorescent protein (YFP) control animals showed a significantdecrease in EtOH lever pressing, but the correspondingdecrease in ArchT animals was comparably smaller andnonsignificant, such that the YFP and ArchT groups differedsignificantly (Figure 5F). The rate of aborts, inactive leverpresses, and time spent in the EtOH lever zone did not differbetween groups, but head entry rate was slightly higher in the
Biological
ArchT group, in line with their slightly higher rate of EtOH leverpressing (Figure 5G, H and Supplemental Figure S9).
We compared the behavioral effects of vmPFC/NAcSsilencing with those of a parallel pathway, the vmPFC/BLA(Figure 5I). Silencing the vmPFC/BLA projections did notaffect any measure for the YFP or ArchT groups (Figure 5J–Land Supplemental Figure S11).
These data extend our recording and optogenetic dataimplicating the vmPFC in punished suppression of EtOH-SAby showing that the vmPFC interacts with the NAcS to someextent to modulate this behavior.
Psychiatry June 1, 2020; 87:967–978 www.sobp.org/journal 973
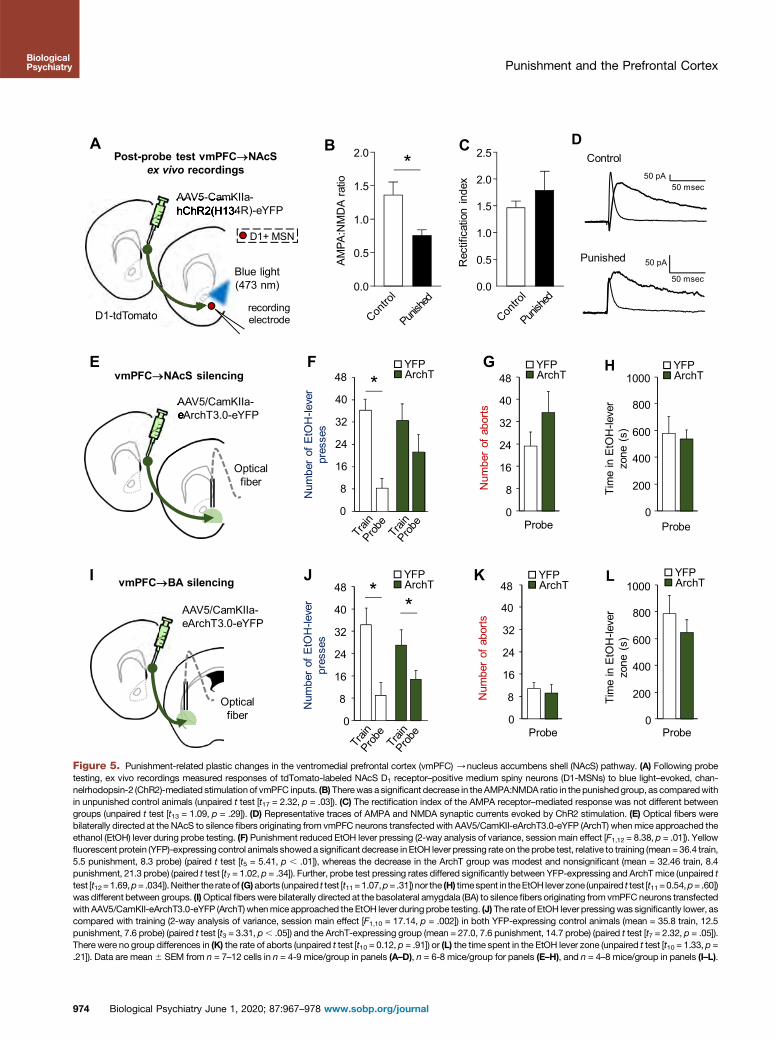
A
E
I
B C
F G
J K
Figure 5. Punishment-related plastic changes in the ventromedial prefrontal cortex (vmPFC) /nucleus accumbens shell (NAcS) pathway. (A) Following probetesting, ex vivo recordings measured responses of tdTomato-labeled NAcS D1 receptor–positive medium spiny neurons (D1-MSNs) to blue light–evoked, chan-nelrhodopsin-2 (ChR2)-mediated stimulation of vmPFC inputs. (B)Therewas a significant decrease in theAMPA:NMDA ratio in the punishedgroup, as comparedwithin unpunished control animals (unpaired t test [t17 = 2.32, p = .03]). (C) The rectification index of the AMPA receptor–mediated response was not different betweengroups (unpaired t test [t13 = 1.09, p = .29]). (D) Representative traces of AMPA and NMDA synaptic currents evoked by ChR2 stimulation. (E) Optical fibers werebilaterally directed at the NAcS to silence fibers originating from vmPFC neurons transfectedwith AAV5/CamKII-eArchT3.0-eYFP (ArchT) whenmice approached theethanol (EtOH) lever during probe testing. (F) Punishment reduced EtOH lever pressing (2-way analysis of variance, session main effect [F1,12 = 8.38, p = .01]). Yellowfluorescent protein (YFP)-expressing control animals showeda significant decrease inEtOH lever pressing rate on the probe test, relative to training (mean= 36.4 train,5.5 punishment, 8.3 probe) (paired t test [t5 = 5.41, p , .01]), whereas the decrease in the ArchT group was modest and nonsignificant (mean = 32.46 train, 8.4punishment, 21.3 probe) (paired t test [t7 = 1.02, p = .34]). Further, probe test pressing rates differed significantly between YFP-expressing and ArchTmice (unpaired ttest [t12=1.69,p= .034]). Neither the rateof (G)aborts (unpaired t test [t11=1.07,p= .31]) nor the (H) timespent in theEtOH lever zone (unpaired t test [t11=0.54,p= .60])was different between groups. (I)Optical fiberswere bilaterally directed at the basolateral amygdala (BA) to silence fibers originating from vmPFCneurons transfectedwith AAV5/CamKII-eArchT3.0-eYFP (ArchT) whenmice approached the EtOH lever duringprobe testing. (J) The rate of EtOH lever pressingwas significantly lower, ascompared with training (2-way analysis of variance, session main effect [F1,10 = 17.14, p = .002]) in both YFP-expressing control animals (mean = 35.8 train, 12.5punishment, 7.6 probe) (paired t test [t3 = 3.31, p, .05]) and the ArchT-expressing group (mean = 27.0, 7.6 punishment, 14.7 probe) (paired t test [t7 = 2.32, p = .05]).There were no group differences in (K) the rate of aborts (unpaired t test [t10 = 0.12, p = .91]) or (L) the time spent in the EtOH lever zone (unpaired t test [t10 = 1.33, p =.21]). Data are mean6 SEM from n = 7–12 cells in n = 4-9 mice/group in panels (A–D), n = 6-8 mice/group for panels (E–H), and n = 4–8 mice/group in panels (I–L).
Punishment and the Prefrontal Cortex
974 Biological Psychiatry June 1, 2020; 87:967–978 www.sobp.org/journal
BiologicalPsychiatry

Punishment and the Prefrontal CortexBiologicalPsychiatry
Minimal Collateralization of vmPFC Neurons to theNAcS and BLA
While optogenetic experiments were designed to selectivelytarget the vmPFC/NAcS/BLA, it is possible that nonselectiveeffects occurred through inhibition of vmPFC neurons sendingcollaterals to both regions. To address this, Alexa-conjugatedCTb (CTb555/488) was injected into the NAcS and BLA toretrogradely label neurons in the mPFC. Consistent with recentmethodologically similar work quantifying vmPFC collaterali-zation to NAcS and BLA (45), we saw very few double-labeledcells in the vmPFC or dmPFC (Supplemental Figure S12),suggesting that the attenuated punished suppression effect ofvmPFC/NAcS silencing is unlikely to stem from effects onvmPFC/NAcS/BLA collaterals, though we cannot excludethe contribution of vmPFC collaterals to other brain regions.
DISCUSSION
Collectively, these data provide correlative and causal evi-dence supporting an important contribution of the vmPFC inregulating EtOH-SA in the face of potential punishment.
Two key findings in the current study were that 1) followingpunishment, vmPFC neurons coded EtOH lever pressing andwere behaviorally required for the suppression of lever press-ing; and 2) while vmPFC neurons also strongly coded foraborting EtOH lever pressing after punishment, the vmPFCwas not necessary for expression of this avoidance beahvior.These data fit with the long-standing view of the importance ofthis region for inhibitory control over various processes at playin the current task, most pertinently reward seeking and threatavoidance (61,66).
As noted, a striking observation was that despite clear ev-idence for neural representation of abort behavior within thevmPFC, optogenetically silencing the vmPFC did not reducethe number of aborts made during probe testing. This di-chotomy suggests that despite the disinhibited EtOH seekinginduced by vmPFC silencing, the association between theEtOH lever response and the punished outcome was retainedand expressed. A related conclusion is that brain regions otherthan the vmPFC (e.g., periaqueductal gray or certain amyg-daloid nuclei) either normally mediate abort expression or canmaintain abort behavior when the vmPFC is nonfunctional.These data support a model in which, rather than being asimple substrate for inhibiting EtOH-SA, the vmPFC has amore complex role in signaling changes in response-outcomerelationships after punishment, which, in turn, enables appro-priate modification of some (lever pressing) but not all (aborts)behaviors.
It is becoming increasingly clear that the vmPFC anddmPFC can contribute to reward-related behaviors in com-plementary or contrasting ways, depending on factors notlimited to the testing context and history with or type of drug/reward (53). Further speaking to this complexity, we found thatin contrast to vmPFC silencing, silencing the dmPFC using thesame experimental design was without effect. On the onehand, these negative findings concur with recent work showingthat pharmacologically inactivating the rat dmPFC did notaffect punished EtOH lever pressing (55) or conditionedavoidance behavior (62). On the other hand, a lack of dmPFCsilencing effects is perhaps surprising in view of the neural
Biological
correlates of probe performance we detected in the dmPFC, aswell as earlier data from rats showing that dmPFC inactivationor photosilencing increased punished licking for water (63) andpromoted punished cocaine seeking (64,65).
There may be technical reasons for these discrepancies,such as species idiosyncrasies or differences in drug orbehavioral assays used. A more interesting possibility, how-ever, is that dmPFC does indeed contribute to punished EtOH-SA, but this effect is difficult to uncover owing to functionalheterogeneity across dmPFC neurons: our neuronal re-cordings indicated the presence of both inhibitory and excit-atory coding of presses and aborts in the dmPFC during probetesting. In this regard, a recent study found that photoexcita-tion of the dmPFC/NAcS reduced punished responding for amilkshake reward (albeit partially) only when shock-activateddmPFC neurons were selectively manipulated (22). Thus,similarly targeted manipulations of task-related neuronal en-sembles may be required to unmask a role for the dmPFC inpunished EtOH-SA.
Providing initial insight into downstream targets of thevmPFC that are recruited to regulate punished EtOH-SA, wefound evidence of plasticity of vmPFC inputs to NAcS D1-MSNs suggestive of a punishment-induced change in synap-tic strength of these inputs. This plasticity appeared restrictedto input onto D1-MSNs but not onto putative D2-MSNs. In-cubation of cocaine seeking has been associated withstrengthening of vmPFC inputs onto NAcS D1-MSNs, as evi-denced by the formation of silent synapses, which, whenontogenetically reversed, produces either increased ordecreased cocaine responding (67,68).
If the vmPFC reduced EtOH lever pressing after punishmentvia downstream connections to NAcS, the most parsimoniousprediction would be behavioral suppression associated withstrengthening of vmPFC inputs to NAcS D1-MSNs. Our findingof reduced D1-MSN AMPA:NMDA ratios, however, appearsmore in line with a depotentiation in this pathway. One possibleexplanation is that this change is indicative of a postsynaptichomeostatic change in NAcS MSNs that could relate to therecruitment of the vmPFC and increased drive onto these cellswith punishment. Equally plausible is the interpretation that thechanges result from altered inputs from other regions, or inmicrocircuits within the NAcS itself. Further work will beneeded to parse these possibilities.
Prior work has shown that ablating mPFC/NAcS neuronsreduces EtOH reinstatement (17), while activating thevmPFC/NAc reduces cocaine seeking (27), and dis-connecting the vmPFC/NAcS pathway produces heroinrelapse (69). We found that silencing the vmPFC/NAcSpathway attenuated a punishment-induced decrease in EtOHlever pressing. Of note, this effect was characterized by apartial reversal of suppression, rather than the full reversalseen with local vmPFC silencing. This difference could beexplained in various ways. One reason could be the use of aCaMKII promoter to express ArchT in the vmPFC. This pro-moter prefers, but is not exclusive to, glutamatergic neurons,leaving open the possible contribution of silencing long-rangeGABAergic interneurons, which could have different oropposite effects to those produced by glutamatergic projec-tion silencing (70). Another explanation is that vmPFC/NAcSsilencing does not fully recapitulate the effects of local vmPFC
Psychiatry June 1, 2020; 87:967–978 www.sobp.org/journal 975

Punishment and the Prefrontal CortexBiologicalPsychiatry
silencing because of the contribution of other efferent pro-jections of the vmPFC. Although the identity of these is un-clear, our negative optogenetic data indicate that thevmPFC/BLA does not critically mediate punished suppres-sion in the current task, despite evidence supporting a role forthe BLA in other assays of punished reward seeking (23,71).
A third, particularly intriguing possibility again raises theissue of functional heterogeneity at the level of the NAcSitself. If distinct subpopulations of NAcS cells promote EtOH-SA, while others mediate suppression, it would not be sur-prising that the net effect of indiscriminately silencing vmPFCinputs to the NAcS would be mixed. In this context, vmPFCinactivation has been found to disinhibit a subpopulation ofNAcS neurons encoding reward-seeking suppression, whilesimultaneously inhibiting another population of reward-encoding NAcS cells (72). Recent work also shows thatwith ventral tegmental area dopamine inputs to NAcS, ef-fects on reward seeking and aversion can be opposite,depending on the subcompartment and cell type targeted(26,73). Finally, in terms of their outputs, NAcS cells targetingthe ventral tegmental area versus the lateral hypothalamushave opposite effects on context-induced EtOH relapse (74).Thus, the vmPFC/NAcS circuit is positioned to exertinhibitory and excitatory influences on consumption andresponding for rewards, including EtOH (75–78). An impor-tant future question will be defining the contribution of inputsfrom the vmPFC (as well as the BLA) (17,79,80) to specificfunctional classes of NAcS neurons involved in punishedsuppression of EtOH-SA.
Functional perturbation of the PFC has been consistentlyreported in alcohol use disorder and in animal models of EtOHexposure, which can lead to a shift toward less flexible be-haviors mediated by subcortical structures (81–83). Theseshifts are thought to contribute to the progression from mod-erate drinking to uncontrolled, compulsive alcohol abuse.Consistent with this, the current study shows that the vmPFCcodes for behaviors associated with the avoidance of a pun-ished response for EtOH and is required for the suppression ofEtOH-SA after punishment. Further, the current results showthat the vmPFC interacts with the NAcS to modulate punishedEtOH-SA behavior. Collectively, these findings advance ourunderstanding of the neural circuits that may underliecompulsive drinking.
ACKNOWLEDGMENTS AND DISCLOSURESThis work was supported by National Institute on Alcohol Abuse andAlcoholism Intramural Research Program Grant No. ZIA-AA000401 (to AH),Intramural Programs of National Institute on Alcohol Abuse and Alcoholismand National Institute of Neurological Disorders and Stroke Grant No. ZIA-AA000421 (to VAA), a Postdoctoral Research Associate Training fellowshipfrom the Center on Compulsive Behaviors (to PTP and MEA), and a NationalInstitutes of Health Director’s Challenge Award and DDIR Innovation Award(to VAA).
The authors report no biomedical financial interests or potential conflictsof interest.
ARTICLE INFORMATIONFrom the Laboratory of Behavioral and Genomic Neuroscience (LRH, AK,PTP, AGL, NAS, KC, LRG, AH) and Laboratory on Neurobiology ofCompulsive Behaviors (MEA, VAA), National Institute on Alcohol Abuse andAlcoholism, National Institutes of Health; and Center on Compulsive
976 Biological Psychiatry June 1, 2020; 87:967–978 www.sobp.org/jo
Behaviors (PTP, MEA, VAA), Intramural Research Program, National In-stitutes of Health, Bethesda, Maryland; Department of Psychology (LRH),Santa Clara University, Santa Clara, California; and the Department ofBehavioral Neuroscience (VDC), Oregon Health Sciences University, Port-land, Oregon.
Address correspondence to Lindsay R. Halladay, Ph.D., Alumni ScienceHall, Room 204, Santa Clara University, 500 El Camino Real, Santa Clara,CA 95053; E-mail: [email protected].
Received Apr 10, 2019; revised Oct 8, 2019; accepted Oct 25, 2019.Supplementary material cited in this article is available online at https://
doi.org/10.1016/j.biopsych.2019.10.030.
REFERENCES1. Substance Abuse and Mental Health Services Administration (2015):
National Survey on Drug Use and Health. Available at: https://www.ncbi.nlm.nih.gov/books/NBK524967/. Accessed December 16, 2019.
2. American Psychiatric Association (2013): Diagnostic and StatisticalManual of Mental Disorders, 5th ed. Washington, DC: American Psy-chiatric Publishing.
3. Tuesta LM, Chen Z, Duncan A, Fowler CD, Ishikawa M, Lee BR, et al.(2017): GLP-1 acts on habenular avoidance circuits to control nicotineintake. Nat Neurosci 20:708–716.
4. Everitt BJ, Robbins TW (2005): Neural systems of reinforcement fordrug addiction: From actions to habits to compulsion. Nat Neurosci8:1481–1489.
5. Koob GF, Volkow ND (2010): Neurocircuitry of addiction. Neuro-psychopharmacology 35:217–238.
6. Hopf FW, Lesscher HM (2014): Rodent models for compulsive alcoholintake. Alcohol 48:253–264.
7. Jean-Richard-Dit-Bressel P, Killcross S, McNally GP (2018): Behav-ioral and neurobiological mechanisms of punishment: Implications forpsychiatric disorders. Neuropsychopharmacology 43:1639–1650.
8. Orsini CA, Moorman DE, Young JW, Setlow B, Floresco SB (2015):Neural mechanisms regulating different forms of risk-related decision-making: Insights from animal models. Neurosci Biobehav Rev 58:147–167.
9. Marchant NJ, Campbell EJ, Pelloux Y, Bossert JM, Shaham Y(2018): Context-induced relapse after extinction versus punishment:Similarities and differences. Psychopharmacology (Berl) 236:439–448.
10. Giuliano C, Pena-Oliver Y, Goodlett CR, Cardinal RN, Robbins TW,Bullmore ET, et al. (2018): Evidence for a long-lasting compulsivealcohol seeking phenotype in rats. Neuropsychopharmacology43:728–738.
11. Pfarr S, Meinhardt MW, Klee ML, Hansson AC, Vengeliene V,Schonig K, et al. (2015): Losing control: Excessive alcohol seekingafter selective inactivation of cue-responsive neurons in the infralimbiccortex. J Neurosci 35:10750–10761.
12. Meinhardt MW, Hansson AC, Perreau-Lenz S, Bauder-Wenz C,Stahlin O, Heilig M, et al. (2013): Rescue of infralimbic mGluR2 deficitrestores control over drug-seeking behavior in alcohol dependence.J Neurosci 33:2794–2806.
13. Groenewegen HJ, Wright CI, Beijer AV, Voorn P (1999): Convergenceand segregation of ventral striatal inputs and outputs. Ann N Y AcadSci 877:49–63.
14. Sesack SR, Deutch AY, Roth RH, Bunney BS (1989): Topographicalorganization of the efferent projections of the medial prefrontal cortexin the rat: An anterograde tract-tracing study with Phaseolus vulgarisleucoagglutinin. J Comp Neurol 290:213–242.
15. Krettek JE, Price JL (1977): Projections from the amygdaloid complexto the cerebral cortex and thalamus in the rat and cat. J Comp Neurol172:687–722.
16. Brog JS, Salyapongse A, Deutch AY, Zahm DS (1993): The patterns ofafferent innervation of the core and shell in the "accumbens" part ofthe rat ventral striatum: Immunohistochemical detection of retro-gradely transported fluoro-gold. J Comp Neurol 338:255–278.
17. Keistler CR, Hammarlund E, Barker JM, Bond CW, DiLeone RJ,Pittenger C, et al. (2017): Regulation of alcohol extinction and cue-induced reinstatement by specific projections among medial
urnal

Punishment and the Prefrontal CortexBiologicalPsychiatry
prefrontal cortex, nucleus accumbens, and basolateral amygdala.J Neurosci 37:4462–4471.
18. Seif T, Chang SJ, Simms JA, Gibb SL, Dadgar J, Chen BT, et al. (2013):Cortical activation of accumbens hyperpolarization-active NMDARsmediates aversion-resistant alcohol intake. Nat Neurosci 16:1094–1100.
19. St Onge JR, Stopper CM, Zahm DS, Floresco SB (2012): Separateprefrontal-subcortical circuits mediate different components of risk-based decision making. J Neurosci 32:2886–2899.
20. St Onge JR, Floresco SB (2010): Prefrontal cortical contribution to risk-based decision making. Cereb Cortex 20:1816–1828.
21. Burgos-Robles A, Kimchi EY, Izadmehr EM, Porzenheim MJ, Ramos-Guasp WA, Nieh EH, et al. (2017): Amygdala inputs to prefrontal cortexguide behavior amid conflicting cues of reward and punishment. NatNeurosci 20:824–835.
22. Kim CK, Ye L, Jennings JH, Pichamoorthy N, Tang DD, Yoo AW, et al.(2017): Molecular and circuit-dynamical identification of top-downneural mechanisms for restraint of reward seeking. Cell 170:1013–1027.e14.
23. Piantadosi PT, Yeates DCM, Wilkins M, Floresco SB (2017): Contri-butions of basolateral amygdala and nucleus accumbens subregionsto mediating motivational conflict during punished reward-seeking.Neurobiol Learn Mem 140:92–105.
24. Piantadosi PT, Yeates DCM, Floresco SB (2018): Cooperative anddissociable involvement of the nucleus accumbens core and shell inthe promotion and inhibition of actions during active and inhibitoryavoidance. Neuropharmacology 138:57–71.
25. Keistler C, Barker JM, Taylor JR (2015): Infralimbic prefrontal cortexinteracts with nucleus accumbens shell to unmask expression ofoutcome-selective Pavlovian-to-instrumental transfer. Learn Mem22:509–513.
26. de Jong JW, Afjei SA, Pollak Dorocic I, Peck JR, Liu C, Kim CK, et al.(2019): A neural circuit mechanism for encoding aversive stimuli in themesolimbic dopamine system. Neuron 101:133–151.e7.
27. Cameron CM, Murugan M, Choi JY, Engel EA, Witten IB (2019):Increased cocaine motivation is associated with degraded spatial andtemporal representations in IL-NAc neurons. Neuron 103:80–91.e7.
28. Laguesse S, Morisot N, Shin JH, Liu F, Adrover MF, Sakhai SA, et al.(2017): Prosapip1-dependent synaptic adaptations in the nucleusaccumbens drive alcohol intake, seeking, and reward. Neuron 96:145–159.e8.
29. Renteria R, Maier EY, Buske TR, Morrisett RA (2017): Selective alter-ations of NMDAR function and plasticity in D1 and D2 medium spinyneurons in the nucleus accumbens shell following chronic intermittentethanol exposure. Neuropharmacology 112:164–171.
30. LaLumiere RT, Smith KC, Kalivas PW (2012): Neural circuit competi-tion in cocaine-seeking: Roles of the infralimbic cortex and nucleusaccumbens shell. Eur J Neurosci 35:614–622.
31. Peters J, Kalivas PW, Quirk GJ (2009): Extinction circuits for fear andaddiction overlap in prefrontal cortex. Learn Mem 16:279–288.
32. Floresco SB (2015): The nucleus accumbens: An interface betweencognition, emotion, and action. Annu Rev Psychol 66:25–52.
33. Gourley SL, Taylor JR (2016): Going and stopping: Dichotomies inbehavioral control by the prefrontal cortex. Nat Neurosci 19:656–664.
34. Peters J, LaLumiere RT, Kalivas PW (2008): Infralimbic prefrontalcortex is responsible for inhibiting cocaine seeking in extinguishedrats. J Neurosci 28:6046–6053.
35. Orsini CA, Heshmati SC, Garman TS, Wall SC, Bizon JL, Setlow B(2018): Contributions of medial prefrontal cortex to decision makinginvolving risk of punishment. Neuropharmacology 139:205–216.
36. Hopf FW, Bowers MS, Chang SJ, Chen BT, Martin M, Seif T, et al.(2011): Reduced nucleus accumbens SK channel activity enhancesalcohol seeking during abstinence. Neuron 65:682–694.
37. Radke AK, Jury NJ, Kocharian A, Marcinkiewcz CA, Lowery-Gionta EG, Pleil KE, et al. (2017): Chronic EtOH effects on putativemeasures of compulsive behavior in mice. Addict Biol 22:423–434.
38. Halladay LR, Kocharian A, Holmes A (2017): Mouse strain differencesin punished ethanol self-administration. Alcohol 58:83–92.
Biological
39. Jury NJ, Pollack GA, Ward MJ, Bezek JL, Ng AJ, Pinard CR, et al.(2017): Chronic Ethanol during adolescence impacts corticolimbicdendritic spines and behavior. Alcohol Clin Exp Res 41:1298–1308.
40. Hunt HF, Brady JV (1955): Some effects of punishment and intercur-rent anxiety on a simple operant. J Comp Physiol Psychol 48:305–310.
41. Gamble-George JC, Baldi R, Halladay L, Kocharian A, Hartley N,Silva CG, et al. (2016): Cyclooxygenase-2 inhibition reduces stress-induced affective pathology. eLife 5:e14137.
42. Gunduz-Cinar O, Brockway E, Lederle L, Wilcox T, Halladay LR,Ding Y, et al. (2019): Identification of a novel gene regulatingamygdala-mediated fear extinction. Mol Psychiatry 24:601–612.
43. Bukalo O, Pinard C, Silverstein S, Brehm C, Hartley N, Whittle N, et al.(2015): Prefrontal inputs to the amygdala instruct fear extinctionmemory formation. Sci Adv 1:e1500251.
44. Bergstrom HC, Lipkin AM, Lieberman AG, Pinard CR, Gunduz-Cinar O,Brockway ET, et al. (2018): Dorsolateral striatum engagement in-terferes with early discrimination learning. Cell Rep 23:2264–2272.
45. Bloodgood DW, Sugam JA, Holmes A, Kash TL (2018): Fear extinctionrequires infralimbic cortex projections to the basolateral amygdala.Transl Psychiatry 8:60.
46. Seo M, Lee E, Averbeck BB (2012): Action selection and action value infrontal-striatal circuits. Neuron 74:947–960.
47. Marchant NJ, Furlong TM, McNally GP (2010): Medial dorsal hypo-thalamus mediates the inhibition of reward seeking after extinction.J Neurosci 30:14102–14115.
48. Rhodes SE, Killcross S (2004): Lesions of rat infralimbic cortexenhance recovery and reinstatement of an appetitive Pavlovianresponse. Learn Mem 11:611–616.
49. Peters J, Vallone J, Laurendi K, Kalivas PW (2008): Opposing roles forthe ventral prefrontal cortex and the basolateral amygdala on thespontaneous recovery of cocaine-seeking in rats. Psychopharmacol-ogy 197:319–326.
50. Barker JM, Torregrossa MM, Taylor JR (2012): Low prefrontal PSA-NCAM confers risk for alcoholism-related behavior. Nat Neurosci15:1356–1358.
51. Ferenczi EA, Zalocusky KA, Liston C, Grosenick L, Warden MR,Amatya D, et al. (2016): Prefrontal cortical regulation of brainwidecircuit dynamics and reward-related behavior. Science351:aac9698.
52. Barker JM, Glen WB, Linsenbardt DN, Lapish CC, Chandler LJ (2017):Habitual behavior is mediated by a shift in response-outcomeencoding by infralimbic cortex. eNeuro 4:ENEURO.0337-17.2017.
53. Moorman DE, James MH, McGlinchey EM, Aston-Jones G (2015):Differential roles of medial prefrontal subregions in the regulation ofdrug seeking. Brain Res 1628:130–146.
54. Pelloux Y, Murray JE, Everitt BJ (2013): Differential roles of the pre-frontal cortical subregions and basolateral amygdala in compulsivecocaine seeking and relapse after voluntary abstinence in rats. Eur JNeurosci 38:3018–3026.
55. Jean-Richard-Dit-Bressel P, McNally GP (2016): Lateral, not medial,prefrontal cortex contributes to punishment and aversive instrumentallearning. Learn Mem 23:607–617.
56. Bossert JM, Stern AL, Theberge FR, Cifani C, Koya E, Hope BT, et al.(2011): Ventral medial prefrontal cortex neuronal ensembles mediatecontext-induced relapse to heroin. Nat Neurosci 14:420–422.
57. Koya E, Uejima JL, Wihbey KA, Bossert JM, Hope BT, Shaham Y(2009): Role of ventral medial prefrontal cortex in incubation of cocainecraving. Neuropharmacology 56(suppl 1):177–185.
58. Warren BL, Mendoza MP, Cruz FC, Leao RM, Caprioli D, Rubio FJ,et al. (2016): Distinct Fos-expressing neuronal ensembles in theventromedial prefrontal cortex mediate food reward and extinctionmemories. J Neurosci 36:6691–6703.
59. Rogers JL, Ghee S, See RE (2008): The neural circuitry underlyingreinstatement of heroin-seeking behavior in an animal model ofrelapse. Neuroscience 151:579–588.
60. Otis JM, Namboodiri VM, Matan AM, Voets ES, Mohorn EP, Kosyk O,et al. (2017): Prefrontal cortex output circuits guide reward seekingthrough divergent cue encoding. Nature 543:103–107.
Psychiatry June 1, 2020; 87:967–978 www.sobp.org/journal 977

Punishment and the Prefrontal CortexBiologicalPsychiatry
61. Halladay LR, Blair HT (2015): Distinct ensembles of medial prefrontalcortex neurons are activated by threatening stimuli that elicit excitationvs. inhibition of movement. J Neurophysiol 114:793–807.
62. Diehl MM, Bravo-Rivera C, Rodriguez-Romaguera J, Pagan-Rivera PA, Burgos-Robles A, Roman-Ortiz C, et al. (2018): Activeavoidance requires inhibitory signaling in the rodent prelimbic pre-frontal cortex. eLife 7:e34657.
63. Resstel LB, Souza RF, Guimaraes FS (2008): Anxiolytic-like effectsinduced by medial prefrontal cortex inhibition in rats submitted to theVogel conflict test. Physiol Behav 93:200–205.
64. Chen BT, Yau HJ, Hatch C, Kusumoto-Yoshida I, Cho SL, Hopf FW,et al. (2013): Rescuing cocaine-induced prefrontal cortex hypoactivityprevents compulsive cocaine seeking. Nature 496:359–362.
65. Limpens JH, Damsteegt R, Broekhoven MH, Voorn P,Vanderschuren LJ (2015): Pharmacological inactivation of the pre-limbic cortex emulates compulsive reward seeking in rats. Brain Res1628:210–218.
66. Halladay LR, Blair HT (2017): Prefrontal infralimbic cortex mediatescompetition between excitation and inhibition of body movementsduring pavlovian fear conditioning. J Neurosci Res 95:853–862.
67. Ma YY, Lee BR, Wang X, Guo C, Liu L, Cui R, et al. (2014): Bidirectionalmodulation of incubation of cocaine craving by silent synapse-basedremodeling of prefrontal cortex to accumbens projections. Neuron83:1453–1467.
68. Pascoli V, Terrier J, Espallergues J, Valjent E, O’Connor EC, Luscher C(2014): Contrasting forms of cocaine-evoked plasticity control com-ponents of relapse. Nature 509:459–464.
69. Bossert JM, Stern AL, Theberge FR, Marchant NJ, Wang HL,Morales M, et al. (2012): Role of projections from ventral medial pre-frontal cortex to nucleus accumbens shell in context-induced rein-statement of heroin seeking. J Neurosci 32:4982–4991.
70. Lee AT, Vogt D, Rubenstein JL, Sohal VS (2014): A class of GABAergicneurons in the prefrontal cortex sends long-range projections to thenucleus accumbens and elicits acute avoidance behavior. J Neurosci34:11519–11525.
71. Jean-Richard-Dit-Bressel P, McNally GP (2015): The role of thebasolateral amygdala in punishment. Learn Mem 22:128–137.
72. Ghazizadeh A, Ambroggi F, Odean N, Fields HL (2012): Prefrontalcortex mediates extinction of responding by two distinct neuralmechanisms in accumbens shell. J Neurosci 32:726–737.
978 Biological Psychiatry June 1, 2020; 87:967–978 www.sobp.org/jo
73. Al-Hasani R, McCall JG, Shin G, Gomez AM, Schmitz GP,Bernardi JM, et al. (2015): Distinct subpopulations of nucleusaccumbens dynorphin neurons drive aversion and reward. Neuron87:1063–1077.
74. Gibson GD, Prasad AA, Jean-Richard-Dit-Bressel P, Yau JOY,Millan EZ, Liu Y, et al. (2018): Distinct accumbens shell output path-ways promote versus prevent relapse to alcohol seeking. Neuron98:512–520.e16.
75. Christakou A, Robbins TW, Everitt BJ (2004): Prefrontal cortical-ventralstriatal interactions involved in affective modulation of attentionalperformance: Implications for corticostriatal circuit function.J Neurosci 24:773–780.
76. Ambroggi F, Ghazizadeh A, Nicola SM, Fields HL (2011): Roles ofnucleus accumbens core and shell in incentive-cue responding andbehavioral inhibition. J Neurosci 31:6820–6830.
77. Taha SA, Fields HL (2006): Inhibitions of nucleus accumbens neuronsencode a gating signal for reward-directed behavior. J Neurosci26:217–222.
78. Krause M, German PW, Taha SA, Fields HL (2010): A pause in nucleusaccumbens neuron firing is required to initiate and maintain feeding.J Neurosci 30:4746–4756.
79. Millan EZ, Kim HA, Janak PH (2017): Optogenetic activation ofamygdala projections to nucleus accumbens can arrest conditionedand unconditioned alcohol consummatory behavior. Neuroscience360:106–117.
80. Lee BR, Ma YY, Huang YH, Wang X, Otaka M, Ishikawa M, et al.(2013): Maturation of silent synapses in amygdala-accumbens pro-jection contributes to incubation of cocaine craving. Nat Neurosci16:1644–1651.
81. Holmes A, Fitzgerald PJ, Macpherson KP, Debrouse L, Colacicco G,Flynn SM, et al. (2012): Chronic alcohol remodels prefrontal neuronsand disrupts NMDAR-mediated fear extinction encoding. Nat Neurosci15:1359–1361.
82. DePoy L, Daut R, Brigman JL, Macpherson K, Crowley N, Gunduz-Cinar O, et al. (2013): Chronic alcohol produces neuroadaptations toprime dorsal striatal learning. Proc Natl Acad Sci U S A 110:14783–14788.
83. Zahr NM, Pfefferbaum A, Sullivan EV (2017): Perspectives on fronto-fugal circuitry from human imaging of alcohol use disorders. Neuro-pharmacology 122:189–200.
urnal