elevated levels of circulating interleukin18 in human immunodeficiency virus-infected individuals:...
TRANSCRIPT

10.1128/JVI.76.24.12448-12456.2002.
2002, 76(24):12448. DOI:J. Virol. Morisset and Ali AhmadRasheed Ahmad, Sardar T. A. Sindhu, Emil Toma, Richard Implications for AIDS PathogenesisPeripheral Blood Mononuclear Cells and Virus-Infected Individuals: Role ofInterleukin-18 in Human Immunodeficiency Elevated Levels of Circulating
http://jvi.asm.org/content/76/24/12448Updated information and services can be found at:
These include:
REFERENCEShttp://jvi.asm.org/content/76/24/12448#ref-list-1at:
This article cites 48 articles, 18 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

JOURNAL OF VIROLOGY, Dec. 2002, p. 12448–12456 Vol. 76, No. 240022-538X/02/$04.00�0 DOI: 10.1128/JVI.76.24.12448–12456.2002Copyright © 2002, American Society for Microbiology. All Rights Reserved.
Elevated Levels of Circulating Interleukin-18 in HumanImmunodeficiency Virus-Infected Individuals: Role
of Peripheral Blood Mononuclear Cells andImplications for AIDS Pathogenesis
Rasheed Ahmad,1 Sardar T. A. Sindhu,1 Emil Toma,2 Richard Morisset,2and Ali Ahmad1*
Laboratory of Immunovirology, Pediatric Research Center, and Department of Microbiology andImmunology, University of Montreal and Sainte-Justine Hospital, Montreal, Quebec, Canada
H3T 1C5,1 and Department of Microbiology and Immunology, University of Montreal andHotel-Dieu Hospital, Montreal, Quebec, Canada H2W 1T82
Received 17 June 2002/Accepted 13 September 2002
Originally identified as the gamma interferon-inducing factor, interleukin-18 (IL-18) was rediscovered as aproinflammatory cytokine related to the IL-1 family of cytokines that plays an important role in both innateand adaptive immune responses against viruses and intracellular pathogens. Despite its importance ininducing and regulating immune responses, relatively little is known about its production in HIV infection. Wereport here significantly (P < 0.05) elevated levels of this cytokine in the sera of human immunodeficiency virus(HIV)-infected/AIDS patients compared to those of HIV-seronegative healthy persons. Surprisingly, the pe-ripheral blood mononuclear cells (PBMC) from HIV-infected/AIDS patients were compromised in the abilityto upregulate IL-18 gene expression and produce this cytokine with and without lipopolysaccharide (LPS)stimulation. A significant positive correlation (P < 0.05) existed between the concentration of IL-18 in serumand its production from PBMC of HIV-seronegative healthy individuals but not those of HIV-infected/AIDSpatients. Furthermore, the patients’ PBMC expressed relatively reduced levels of activated caspase-1 consti-tutively as well as in response to LPS stimulation. Our data suggest the involvement of transforming growthfactor beta (TGF-�) in suppressing IL-18 production from the patients’ PBMC for the following reasons. (i)In in vitro studies it suppressed the production of IL-18 from PBMC. (ii) Its levels were significantly higherin the plasma of patients compared to that of control subjects. (iii) A significant negative correlation existedbetween the concentrations of TGF-� in plasma and of IL-18 in serum of the patients. The elevated levels ofIL-18 in the serum of HIV-infected individuals may contribute to AIDS pathogenesis, whereas its compromisedproduction from their PBMC in response to stimuli may reduce their innate defense to opportunistic intra-cellular pathogens.
AIDS in humans is the ultimate outcome of an infection withhuman immunodeficiency virus type 1 (HIV-1). This viral in-fection causes several immune abnormalities in the infectedhost, rendering him or her unable to control the infection. Italso makes the host unusually susceptible to a variety of op-portunistic infections. These immune abnormalities occur, atleast in part, as a result of the deregulated expression of severalimmunologically important cytokines (6, 9, 36, 37). For exam-ple, aberrant production of tumor necrosis factor alpha (TNF-�), transforming growth factor beta (TGF-�), interleukin-12(IL-12), and IL-15, etc., has been well documented in HIV-infected/AIDS patients (1, 7, 8, 21). Optimal production ofthese cytokines is essential not only for innate host resistanceto pathogens but also for the induction, amplification, andmaintenance of pathogen-specific immunity. The qualitativenature of the immune response is also determined by the typeof cytokines induced in the host, i.e., whether they are of
T-helper 1 (TH1) or of TH2 type. A predominance of TH2-type cytokines has been reported in HIV-infected/AIDS pa-tients and is believed to contribute to the pathogenesis ofAIDS (36). Finally, cytokines induce and/or activate transcrip-tion factors directly or indirectly, which may regulate HIV-1replication in human cells. Despite the ability of IL-18 (seebelow) to enhance innate immunity, regulate TH1- and TH2-type immune responses, and enhance HIV-1 replication, littleis known about its expression in HIV-infected/AIDS patients.
Previously known as the gamma interferon (IFN-�)-inducingfactor, IL-18 was rediscovered as a novel cytokine that plays animportant role in promoting TH1 responses by its ability toinduce IFN-� from T and natural killer (NK) cells (24, 25, 28;reviewed in references 11 and 26). This pleiotropic cytokine isproduced by activated macrophages, dendritic cells, Kupffercells, keratinocytes, and enterocytes as well as by the adrenalcortex and neurohypophysis. These cells usually express theIL-18 gene and protein constitutively and increase their ex-pression in response to stress, infection, cold, and lipopolysac-charide (LPS). IL-18 and IL-1� have similar tertiary structures(all �-pleated folded forms). Furthermore, both these cyto-kines are produced as inactive precursors without a signal
* Corresponding author. Mailing address: Laboratory of Immuno-virology, Sainte-Justine Hospital, 3175 Cote Ste-Catherine, Montreal,Quebec, Canada H3T 1C5, Tel: (514) 345-4729, Fax: (514) 345-4801,E-mail: [email protected].
12448
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

peptide and require proteolytic cleavage by the IL-1�-convert-ing enzyme (ICE or caspase-1) to become mature biologicallyactive molecules, which are then readily released from the cells(14). ICE cleaves the 193-amino-acid precursor form of IL-18on the carboxyl side of aspartate 36 (14, 17). IL-18 uses aunique heterodimeric receptor, which comprises the ligand-binding � chain and the signal-transducing � chain, and be-longs to the IL-1 receptor superfamily. IL-18 potentiates in-nate immunity by increasing the cytolytic potential of NK andT cells and regulates adaptive immune responses to pathogens(4, 29). Its protective role for the host against intracellularpathogens, including viruses, has been well documented (11,31). Being a proinflammatory cytokine, its deregulated produc-tion has also been implicated in several chronic autoimmunedisorders (23, 44). Induction of IL-18 production seems to bean integral part of the host’s innate response to viral pathogens(26, 32, 41). However, little is known about the regulation ofthis cytokine in HIV-1 infection. This issue is addressed in thisreport.
MATERIALS AND METHODS
Patients. Peripheral blood was obtained for serum collection from 70 HIV-infected/AIDS patients of both genders (50 males and 20 females) from localAIDS clinics after their written informed consent. The clinical characteristics ofthe patients were: age, 28 to 65 years (median� 44); CD4� T cells, 16 to 774 per�l (median � 242); viral load, 101.70 to 105.66 (median � 103.24); infectionduration, 2 to 10 years (median� 6 years).
The viral load in plasma (i.e., number of HIV-1 RNA copies per ml) wasdetermined using a colorimetric reverse transcription-PCR-based commercial kit(HIV-1 monitor test; Roche Diagnostic Systems, Branchburg, N.J.). The pa-tients’ CD4� T-cell counts in the peripheral blood (number of cells per micro-liter) were determined by flow cytometry by use of the whole blood lysis method(BD Bioscience, Mississauga, Ontario, Canada). All the patients except six werereceiving highly active antiretroviral therapy (HAART) that comprised at leastone protease inhibitor (saquinavir, ritonavir, or indinavir) and one or two reversetranscriptase inhibitors (stavudine [D4T], lamivudine [3TC], zidovudine [AZT],or didanosine). Blood samples from 28 age-matched HIV-seronegative healthypersons were used as controls.
Separation of serum and PBMC. For the collection of serum samples, periph-eral blood was obtained in vacuum tubes without any additive. The blood wasallowed to clot a room temperature for 4 to 6 h. Thereafter, clear straw-coloredserum was obtained by centrifugation, divided into aliquots, and stored at �80°Cuntil used. Peripheral blood mononuclear cells (PBMC) were obtained from theperipheral blood from some of these patients (detailed in individual experi-ments). For this purpose, blood was collected in heparinized vacuum tubes bycentrifugation over Ficol-Hypaque (Pharmacia, Montreal, Ontario, Canada) asdescribed earlier (2). The buffy coat at the interface of Ficol-Hypaque and bloodwas collected, washed, and resuspended in RPM1-1640 medium containing 10%heat-inactivated fetal bovine serum and antibiotics (culture medium) as de-scribed previously (2).
PBMC cultures. PBMC (2 � 105) were incubated in triplicate in the wells ofa round-bottomed 96-well microculture plate in the culture medium with orwithout the presence of LPS (from Escherichia coli O55:B5; Sigma, St. Louis,Mo.; catalog no. L6529; 10 �g/ml). Supernatants from these microcultures werecollected 12, 24, 48, and 72 h later, filtered through low protein binding 0.45-nm-pore-size filters (Millex-HV; Millipore, Bedford, Mass.), divided into ali-quots, and stored at �80°C.
ELISA for IL-18. The concentrations of IL-18 were determined in serumsamples and culture supernatants using a commercial enzyme-linked immu-nosorbent assay (ELISA) kit (MBL, Naka-ku Nagoya, Japan), which has adetection limit of 12.5 pg/ml. The kit is based upon a sandwich ELISA thatutilizes two monoclonal antibodies directed against two different epitopes ofhuman IL-18. One antibody is coated onto the wells of the microculture plate tocapture IL-18 from biological fluids, and the second antibody is conjugated withhorseradish peroxidase. After being washed, the chromogenic substrate tetram-ethylbenzidine was added and the optical density was measured in an ELISAreader (Easy Reader EAR 400AT; STL Lab-instruments, Salzburg, Austria) at450 nm as directed by the manufacturer.
Biological activity of IL-18. The biological activity of IL-18 in the culturesupernatants was tested by its ability to induce IFN-� in mouse splenocytes aftertheir suboptimal stimulation with concanavalin A (ConA) (0.2 �g/ml; Sigma) asdescribed previously (39). Briefly, the culture supernatants were concentrated10-fold by a microconcentrator (Amicon, Beverly, Mass.; catalog no. 42406) andadded in a 50-�l volume to the ConA-prestimulated 5 � 106 mouse splenocytes.After 24 h, the quantity of IFN-� secreted into the culture supernatants wasmeasured with a commercial ELISA kit (R & D Systems, Minneapolis, Minn.).The specificity of the IFN-� production to IL-18 in this assay was confirmed bypreincubating the supernatants with an IL-18-neutralizing monoclonal antibody(clone 52713.11; 1 �g/ml; R & D Systems) and an isotype-matched (IgG1)control antibody (1 �g/ml; Serotec, Toronto, Ontario, Canada).
Determination of TGF-�. The contents of transforming growth factor beta(TGF-�) in the sera and plasma samples were determined with a commercialELISA kit (TGF-�1 Emax immunoassay system; Promega, Madison, Wis.) byfollowing the manufacturer’s recommendations. The kit determines the biolog-ically active form of the cytokine in an antibody sandwich format. The first(capture) antibody, used to coat the ELISA plate, is a mouse monoclonal anti-body that is specific for the activated form of TGF-�1. For determining totalTGF-�1, the samples were first treated with acid and neutralized as detailed inour previous publication (47).
Western blotting. The expression of different proteins within PBMC wasanalyzed by Western blotting as described in our earlier publications (3, 46).Briefly, 2 � 106 PBMC were incubated in the culture medium with and withouttreatment as detailed in individual experiments. Twenty-four hours after incu-bation, cells were washed with phosphate-buffered saline and lyzed in a lysisbuffer containing Tris HCl (pH 6.8; 50 mM), sodium dodecyl sulfate (SDS) (2%),leupeptin (1 mg/ml), phenylmethylsulfonyl fluoride (1 mM), and pepstatin (1mg/ml). The lysates were clarified by centrifugation at 14,000 � g for 15 min.Protein concentrations were determined in the lysates by using a commercial kit(Bio-Rad, Richmond, Calif.). Forty micrograms of the lysate proteins was mixedwith 2� SDS-polyacrylamide gel electrophoresis (PAGE) sample loading buffercontaining 1 mM dithiothreitol, boiled, run on SDS–12% PAGE gels, and elec-troblotted onto polyvinylidene difluoride membranes (Immobilon; Millipore).After blockage of the membranes in 1% casein for 30 min at room temperature,they were incubated on a shaker with protein-specific antibodies, i.e., antihumanIL-18 (Clone 52713.11; R & D Systems), anti-caspase-1 (clone B24-2; Pharmin-gen BD, Mississauga, Ontario, Canada), or anti-�-actin (Sigma), at room tem-perature for 1 h. The protein bands were revealed by autoradiography by usingbiotinylated goat antimouse antibodies and a commercial chemiluminiscent kit(Vectastain ABC-AmP; Vector Labs, Burlington, Calif.). Individual bands on theX-ray films were quantified by densitometry.
Fluorescence-activated cell sorter analysis. The percentage of PBMC express-ing CD14 on their surface was determined by membrane immunofluorescence.For this purpose, 0.5 � 106 PBMC were incubated with phycoerythrin-conju-gated antihuman CD14 (Pharmingen BD) on ice for 45 min. After being washedwith phosphate-buffered saline, the cells were resuspended in 2% paraformal-dehyde and analyzed by flow cytometry.
RPA. IL-18 transcripts were measured by using RNase protection assay(RPA). For this purpose, total cellular RNA was extracted from 106 PBMC,using Trizol reagent (Life Technologies, Burlington, Ontario, Canada), which isessentially a monophasic solution of phenol and guanidine isiothiocyanate. RNAwas stored as an ethanol precipitate at �80°C until used. High-specific-activityantisense probe for detecting IL-18 mRNA was prepared by in vitro transcriptionwith a commercial kit (Promega) and DNA-dependent RNA polymerase fromSP6 bacteriophage. The template vector pGEM-3zf(�) containing IL-18 cDNAhas been previously described and was linearized with SacI (32). One millioncounts per minute of the 32P-labeled antisense probe was hybridized overnight at56°C with the total cellular RNA and treated with RNase I using a kit fromPharmingen BD (RiboQuant). The reaction mixture was run on a 5% denaturingpolyacrylamide gel. The protected probe was revealed and quantitated by phos-phorimaging. For normalization, the transcripts of the housekeeping gene glyc-eraldehyde-3-phosphate dehydrogenase (GAPDH) were also determined alongwith the IL-18 transcripts. The antisense probe for GAPDH was prepared froma commercially available template (Pharmingen BD) as described above forIL-18; however, the DNA-dependent RNA polymerase from the T3 bacterio-phage was used. The ratio between IL-18 and GAPDH transcripts was deter-mined and compared between different PBMC samples by the ImageQuantsoftware of Molecular Dynamics (Sunnyvale, Calif.).
Statistical analysis. The viral loads (number of copies of HIV-1 RNA per mlof plasma) were log10 transformed. The differences between means of differentparameters were compared using Student’s unpaired t test. The forward corre-lation between two variables was determined using Pearson’s method. The for-
VOL. 76, 2002 PRODUCTION OF IL-18 IN HIV INFECTION 12449
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
ward stepwise multiple linear regression between IL-18 (independent variable)and dependent variables (CD4� T-cell count, viral load, CD8� T-cell counts, andCD4�/CD8� T-cell ratios) was determined by standard methods. For all analy-ses, statistical software SPSS (Statistical Package for Social Sciences; Graph PadInc., Chicago, Ill.) was used. P values �0.05 were considered significant.
RESULTS
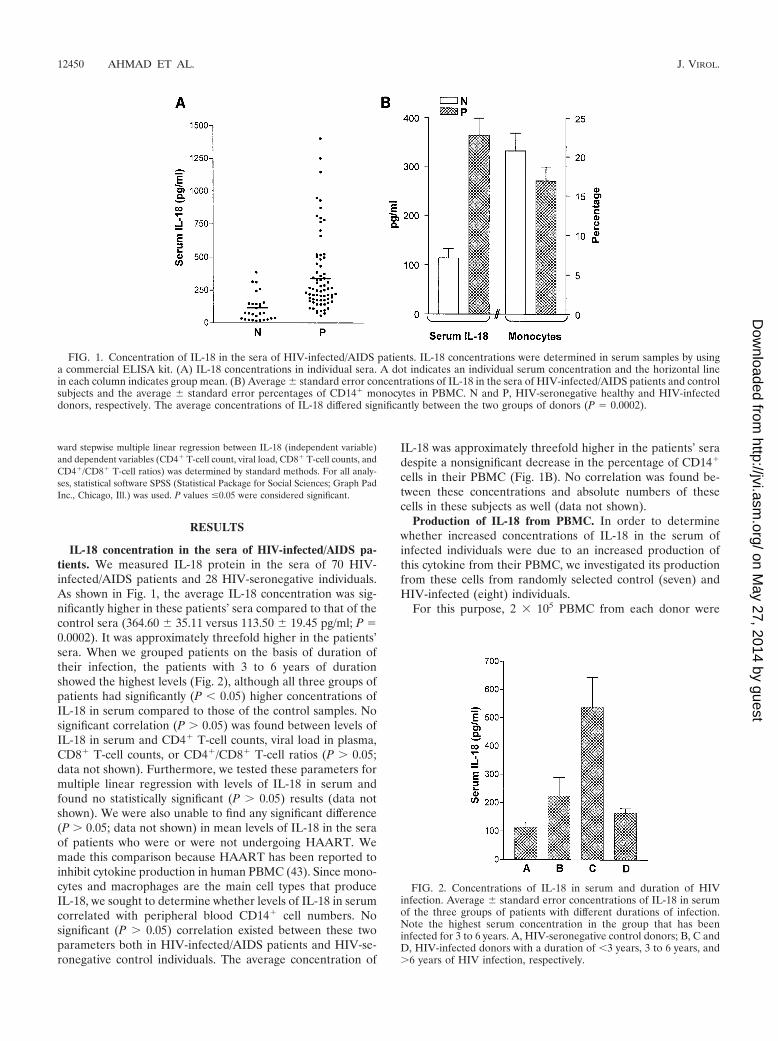
IL-18 concentration in the sera of HIV-infected/AIDS pa-tients. We measured IL-18 protein in the sera of 70 HIV-infected/AIDS patients and 28 HIV-seronegative individuals.As shown in Fig. 1, the average IL-18 concentration was sig-nificantly higher in these patients’ sera compared to that of thecontrol sera (364.60 35.11 versus 113.50 19.45 pg/ml; P �0.0002). It was approximately threefold higher in the patients’sera. When we grouped patients on the basis of duration oftheir infection, the patients with 3 to 6 years of durationshowed the highest levels (Fig. 2), although all three groups ofpatients had significantly (P � 0.05) higher concentrations ofIL-18 in serum compared to those of the control samples. Nosignificant correlation (P � 0.05) was found between levels ofIL-18 in serum and CD4� T-cell counts, viral load in plasma,CD8� T-cell counts, or CD4�/CD8� T-cell ratios (P � 0.05;data not shown). Furthermore, we tested these parameters formultiple linear regression with levels of IL-18 in serum andfound no statistically significant (P � 0.05) results (data notshown). We were also unable to find any significant difference(P � 0.05; data not shown) in mean levels of IL-18 in the seraof patients who were or were not undergoing HAART. Wemade this comparison because HAART has been reported toinhibit cytokine production in human PBMC (43). Since mono-cytes and macrophages are the main cell types that produceIL-18, we sought to determine whether levels of IL-18 in serumcorrelated with peripheral blood CD14� cell numbers. Nosignificant (P � 0.05) correlation existed between these twoparameters both in HIV-infected/AIDS patients and HIV-se-ronegative control individuals. The average concentration of
IL-18 was approximately threefold higher in the patients’ seradespite a nonsignificant decrease in the percentage of CD14�
cells in their PBMC (Fig. 1B). No correlation was found be-tween these concentrations and absolute numbers of thesecells in these subjects as well (data not shown).
Production of IL-18 from PBMC. In order to determinewhether increased concentrations of IL-18 in the serum ofinfected individuals were due to an increased production ofthis cytokine from their PBMC, we investigated its productionfrom these cells from randomly selected control (seven) andHIV-infected (eight) individuals.
For this purpose, 2 � 105 PBMC from each donor were
FIG. 1. Concentration of IL-18 in the sera of HIV-infected/AIDS patients. IL-18 concentrations were determined in serum samples by usinga commercial ELISA kit. (A) IL-18 concentrations in individual sera. A dot indicates an individual serum concentration and the horizontal linein each column indicates group mean. (B) Average standard error concentrations of IL-18 in the sera of HIV-infected/AIDS patients and controlsubjects and the average standard error percentages of CD14� monocytes in PBMC. N and P, HIV-seronegative healthy and HIV-infecteddonors, respectively. The average concentrations of IL-18 differed significantly between the two groups of donors (P � 0.0002).
FIG. 2. Concentrations of IL-18 in serum and duration of HIVinfection. Average standard error concentrations of IL-18 in serumof the three groups of patients with different durations of infection.Note the highest serum concentration in the group that has beeninfected for 3 to 6 years. A, HIV-seronegative control donors; B, C andD, HIV-infected donors with a duration of �3 years, 3 to 6 years, and�6 years of HIV infection, respectively.
12450 AHMAD ET AL. J. VIROL.
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
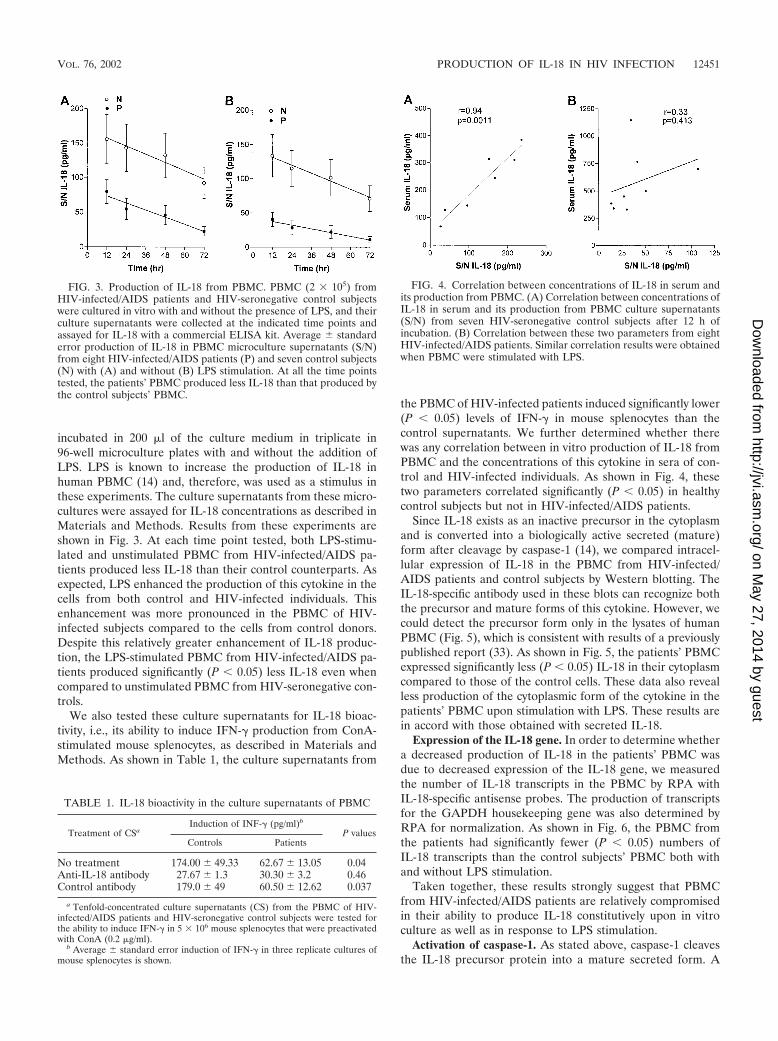
incubated in 200 �l of the culture medium in triplicate in96-well microculture plates with and without the addition ofLPS. LPS is known to increase the production of IL-18 inhuman PBMC (14) and, therefore, was used as a stimulus inthese experiments. The culture supernatants from these micro-cultures were assayed for IL-18 concentrations as described inMaterials and Methods. Results from these experiments areshown in Fig. 3. At each time point tested, both LPS-stimu-lated and unstimulated PBMC from HIV-infected/AIDS pa-tients produced less IL-18 than their control counterparts. Asexpected, LPS enhanced the production of this cytokine in thecells from both control and HIV-infected individuals. Thisenhancement was more pronounced in the PBMC of HIV-infected subjects compared to the cells from control donors.Despite this relatively greater enhancement of IL-18 produc-tion, the LPS-stimulated PBMC from HIV-infected/AIDS pa-tients produced significantly (P � 0.05) less IL-18 even whencompared to unstimulated PBMC from HIV-seronegative con-trols.
We also tested these culture supernatants for IL-18 bioac-tivity, i.e., its ability to induce IFN-� production from ConA-stimulated mouse splenocytes, as described in Materials andMethods. As shown in Table 1, the culture supernatants from
the PBMC of HIV-infected patients induced significantly lower(P � 0.05) levels of IFN-� in mouse splenocytes than thecontrol supernatants. We further determined whether therewas any correlation between in vitro production of IL-18 fromPBMC and the concentrations of this cytokine in sera of con-trol and HIV-infected individuals. As shown in Fig. 4, thesetwo parameters correlated significantly (P � 0.05) in healthycontrol subjects but not in HIV-infected/AIDS patients.
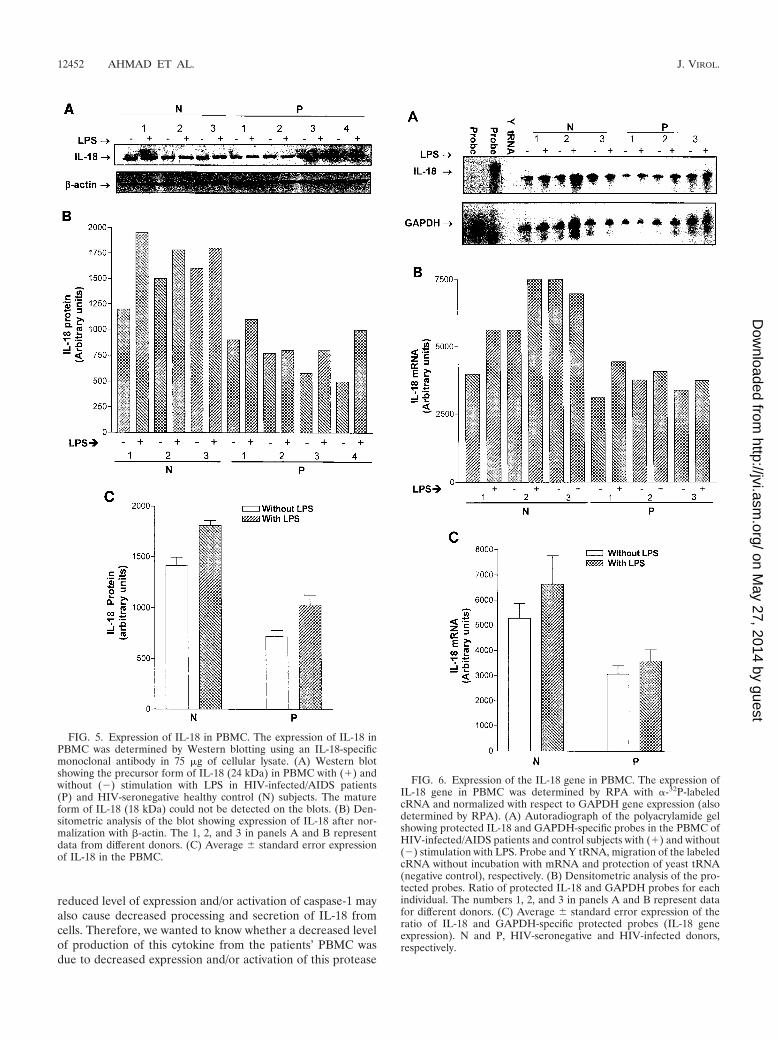
Since IL-18 exists as an inactive precursor in the cytoplasmand is converted into a biologically active secreted (mature)form after cleavage by caspase-1 (14), we compared intracel-lular expression of IL-18 in the PBMC from HIV-infected/AIDS patients and control subjects by Western blotting. TheIL-18-specific antibody used in these blots can recognize boththe precursor and mature forms of this cytokine. However, wecould detect the precursor form only in the lysates of humanPBMC (Fig. 5), which is consistent with results of a previouslypublished report (33). As shown in Fig. 5, the patients’ PBMCexpressed significantly less (P � 0.05) IL-18 in their cytoplasmcompared to those of the control cells. These data also revealless production of the cytoplasmic form of the cytokine in thepatients’ PBMC upon stimulation with LPS. These results arein accord with those obtained with secreted IL-18.
Expression of the IL-18 gene. In order to determine whethera decreased production of IL-18 in the patients’ PBMC wasdue to decreased expression of the IL-18 gene, we measuredthe number of IL-18 transcripts in the PBMC by RPA withIL-18-specific antisense probes. The production of transcriptsfor the GAPDH housekeeping gene was also determined byRPA for normalization. As shown in Fig. 6, the PBMC fromthe patients had significantly fewer (P � 0.05) numbers ofIL-18 transcripts than the control subjects’ PBMC both withand without LPS stimulation.
Taken together, these results strongly suggest that PBMCfrom HIV-infected/AIDS patients are relatively compromisedin their ability to produce IL-18 constitutively upon in vitroculture as well as in response to LPS stimulation.
Activation of caspase-1. As stated above, caspase-1 cleavesthe IL-18 precursor protein into a mature secreted form. A
FIG. 3. Production of IL-18 from PBMC. PBMC (2 � 105) fromHIV-infected/AIDS patients and HIV-seronegative control subjectswere cultured in vitro with and without the presence of LPS, and theirculture supernatants were collected at the indicated time points andassayed for IL-18 with a commercial ELISA kit. Average standarderror production of IL-18 in PBMC microculture supernatants (S/N)from eight HIV-infected/AIDS patients (P) and seven control subjects(N) with (A) and without (B) LPS stimulation. At all the time pointstested, the patients’ PBMC produced less IL-18 than that produced bythe control subjects’ PBMC.
FIG. 4. Correlation between concentrations of IL-18 in serum andits production from PBMC. (A) Correlation between concentrations ofIL-18 in serum and its production from PBMC culture supernatants(S/N) from seven HIV-seronegative control subjects after 12 h ofincubation. (B) Correlation between these two parameters from eightHIV-infected/AIDS patients. Similar correlation results were obtainedwhen PBMC were stimulated with LPS.
TABLE 1. IL-18 bioactivity in the culture supernatants of PBMC
Treatment of CSaInduction of INF-� (pg/ml)b
P valuesControls Patients
No treatment 174.00 49.33 62.67 13.05 0.04Anti-IL-18 antibody 27.67 1.3 30.30 3.2 0.46Control antibody 179.0 49 60.50 12.62 0.037
a Tenfold-concentrated culture supernatants (CS) from the PBMC of HIV-infected/AIDS patients and HIV-seronegative control subjects were tested forthe ability to induce IFN-� in 5 � 106 mouse splenocytes that were preactivatedwith ConA (0.2 �g/ml).
b Average standard error induction of IFN-� in three replicate cultures ofmouse splenocytes is shown.
VOL. 76, 2002 PRODUCTION OF IL-18 IN HIV INFECTION 12451
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
reduced level of expression and/or activation of caspase-1 mayalso cause decreased processing and secretion of IL-18 fromcells. Therefore, we wanted to know whether a decreased levelof production of this cytokine from the patients’ PBMC wasdue to decreased expression and/or activation of this protease
FIG. 5. Expression of IL-18 in PBMC. The expression of IL-18 inPBMC was determined by Western blotting using an IL-18-specificmonoclonal antibody in 75 �g of cellular lysate. (A) Western blotshowing the precursor form of IL-18 (24 kDa) in PBMC with (�) andwithout (�) stimulation with LPS in HIV-infected/AIDS patients(P) and HIV-seronegative healthy control (N) subjects. The matureform of IL-18 (18 kDa) could not be detected on the blots. (B) Den-sitometric analysis of the blot showing expression of IL-18 after nor-malization with �-actin. The 1, 2, and 3 in panels A and B representdata from different donors. (C) Average standard error expressionof IL-18 in the PBMC.
FIG. 6. Expression of the IL-18 gene in PBMC. The expression ofIL-18 gene in PBMC was determined by RPA with �-32P-labeledcRNA and normalized with respect to GAPDH gene expression (alsodetermined by RPA). (A) Autoradiograph of the polyacrylamide gelshowing protected IL-18 and GAPDH-specific probes in the PBMC ofHIV-infected/AIDS patients and control subjects with (�) and without(�) stimulation with LPS. Probe and Y tRNA, migration of the labeledcRNA without incubation with mRNA and protection of yeast tRNA(negative control), respectively. (B) Densitometric analysis of the pro-tected probes. Ratio of protected IL-18 and GAPDH probes for eachindividual. The numbers 1, 2, and 3 in panels A and B represent datafor different donors. (C) Average standard error expression of theratio of IL-18 and GAPDH-specific protected probes (IL-18 geneexpression). N and P, HIV-seronegative and HIV-infected donors,respectively.
12452 AHMAD ET AL. J. VIROL.
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
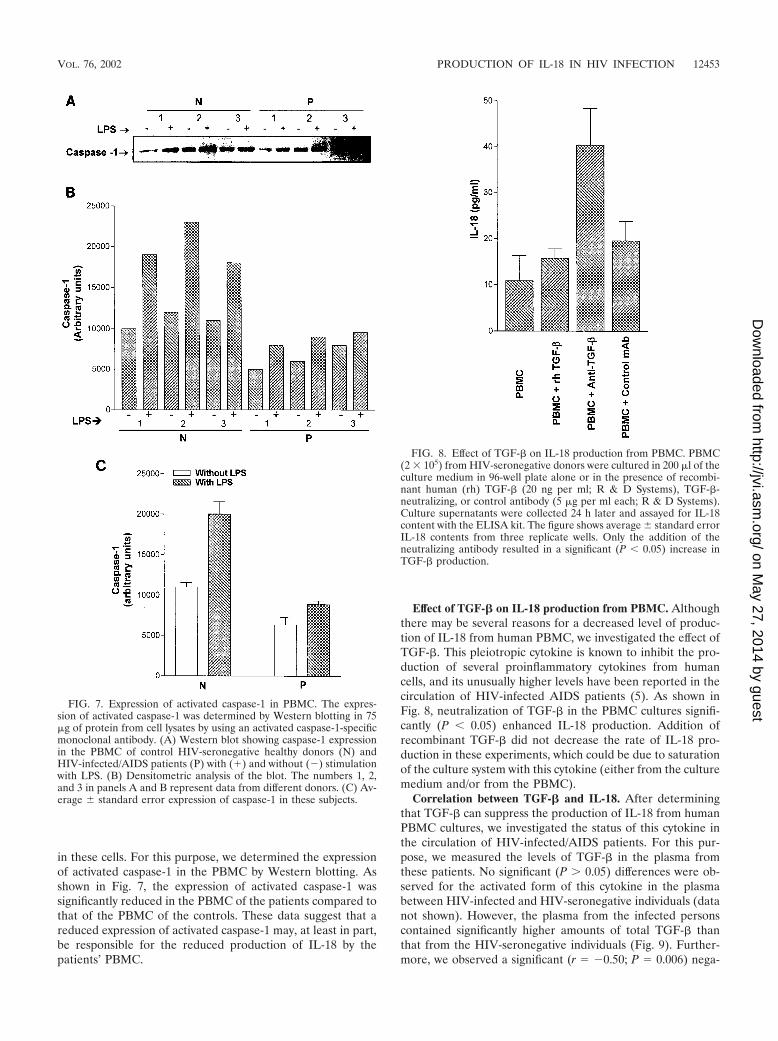
in these cells. For this purpose, we determined the expressionof activated caspase-1 in the PBMC by Western blotting. Asshown in Fig. 7, the expression of activated caspase-1 wassignificantly reduced in the PBMC of the patients compared tothat of the PBMC of the controls. These data suggest that areduced expression of activated caspase-1 may, at least in part,be responsible for the reduced production of IL-18 by thepatients’ PBMC.
Effect of TGF-� on IL-18 production from PBMC. Althoughthere may be several reasons for a decreased level of produc-tion of IL-18 from human PBMC, we investigated the effect ofTGF-�. This pleiotropic cytokine is known to inhibit the pro-duction of several proinflammatory cytokines from humancells, and its unusually higher levels have been reported in thecirculation of HIV-infected AIDS patients (5). As shown inFig. 8, neutralization of TGF-� in the PBMC cultures signifi-cantly (P � 0.05) enhanced IL-18 production. Addition ofrecombinant TGF-� did not decrease the rate of IL-18 pro-duction in these experiments, which could be due to saturationof the culture system with this cytokine (either from the culturemedium and/or from the PBMC).
Correlation between TGF-� and IL-18. After determiningthat TGF-� can suppress the production of IL-18 from humanPBMC cultures, we investigated the status of this cytokine inthe circulation of HIV-infected/AIDS patients. For this pur-pose, we measured the levels of TGF-� in the plasma fromthese patients. No significant (P � 0.05) differences were ob-served for the activated form of this cytokine in the plasmabetween HIV-infected and HIV-seronegative individuals (datanot shown). However, the plasma from the infected personscontained significantly higher amounts of total TGF-� thanthat from the HIV-seronegative individuals (Fig. 9). Further-more, we observed a significant (r � �0.50; P � 0.006) nega-
FIG. 7. Expression of activated caspase-1 in PBMC. The expres-sion of activated caspase-1 was determined by Western blotting in 75�g of protein from cell lysates by using an activated caspase-1-specificmonoclonal antibody. (A) Western blot showing caspase-1 expressionin the PBMC of control HIV-seronegative healthy donors (N) andHIV-infected/AIDS patients (P) with (�) and without (�) stimulationwith LPS. (B) Densitometric analysis of the blot. The numbers 1, 2,and 3 in panels A and B represent data from different donors. (C) Av-erage standard error expression of caspase-1 in these subjects.
FIG. 8. Effect of TGF-� on IL-18 production from PBMC. PBMC(2 � 105) from HIV-seronegative donors were cultured in 200 �l of theculture medium in 96-well plate alone or in the presence of recombi-nant human (rh) TGF-� (20 ng per ml; R & D Systems), TGF-�-neutralizing, or control antibody (5 �g per ml each; R & D Systems).Culture supernatants were collected 24 h later and assayed for IL-18content with the ELISA kit. The figure shows average standard errorIL-18 contents from three replicate wells. Only the addition of theneutralizing antibody resulted in a significant (P � 0.05) increase inTGF-� production.
VOL. 76, 2002 PRODUCTION OF IL-18 IN HIV INFECTION 12453
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
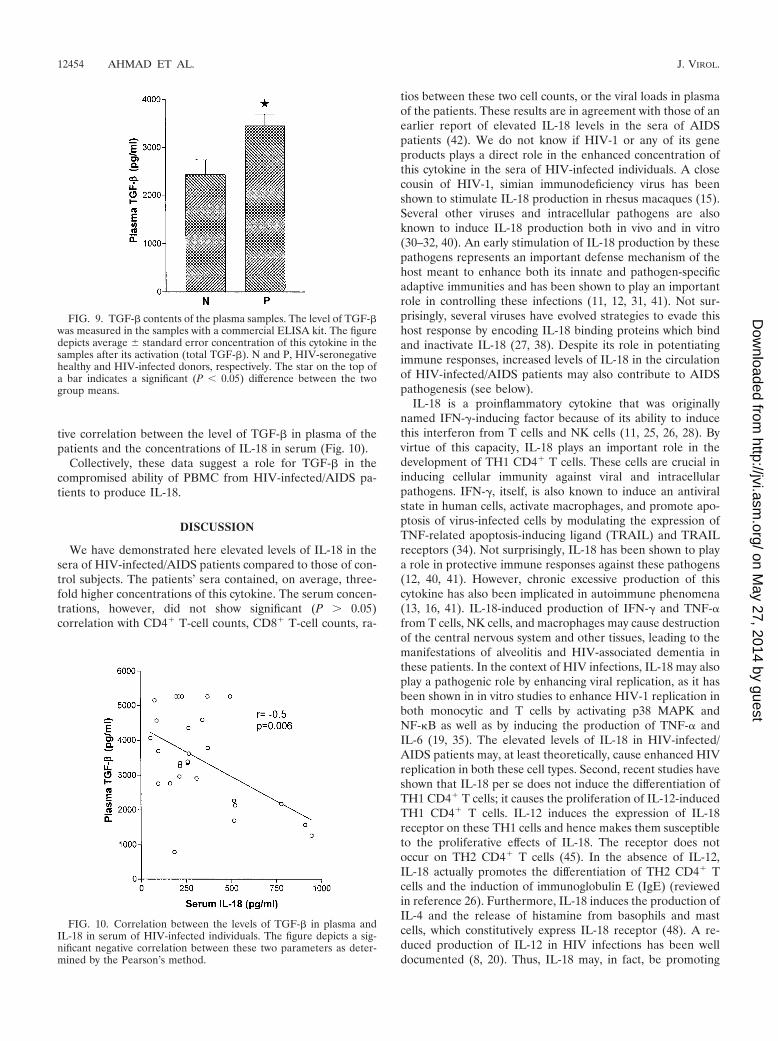
tive correlation between the level of TGF-� in plasma of thepatients and the concentrations of IL-18 in serum (Fig. 10).
Collectively, these data suggest a role for TGF-� in thecompromised ability of PBMC from HIV-infected/AIDS pa-tients to produce IL-18.
DISCUSSION
We have demonstrated here elevated levels of IL-18 in thesera of HIV-infected/AIDS patients compared to those of con-trol subjects. The patients’ sera contained, on average, three-fold higher concentrations of this cytokine. The serum concen-trations, however, did not show significant (P � 0.05)correlation with CD4� T-cell counts, CD8� T-cell counts, ra-
tios between these two cell counts, or the viral loads in plasmaof the patients. These results are in agreement with those of anearlier report of elevated IL-18 levels in the sera of AIDSpatients (42). We do not know if HIV-1 or any of its geneproducts plays a direct role in the enhanced concentration ofthis cytokine in the sera of HIV-infected individuals. A closecousin of HIV-1, simian immunodeficiency virus has beenshown to stimulate IL-18 production in rhesus macaques (15).Several other viruses and intracellular pathogens are alsoknown to induce IL-18 production both in vivo and in vitro(30–32, 40). An early stimulation of IL-18 production by thesepathogens represents an important defense mechanism of thehost meant to enhance both its innate and pathogen-specificadaptive immunities and has been shown to play an importantrole in controlling these infections (11, 12, 31, 41). Not sur-prisingly, several viruses have evolved strategies to evade thishost response by encoding IL-18 binding proteins which bindand inactivate IL-18 (27, 38). Despite its role in potentiatingimmune responses, increased levels of IL-18 in the circulationof HIV-infected/AIDS patients may also contribute to AIDSpathogenesis (see below).
IL-18 is a proinflammatory cytokine that was originallynamed IFN-�-inducing factor because of its ability to inducethis interferon from T cells and NK cells (11, 25, 26, 28). Byvirtue of this capacity, IL-18 plays an important role in thedevelopment of TH1 CD4� T cells. These cells are crucial ininducing cellular immunity against viral and intracellularpathogens. IFN-�, itself, is also known to induce an antiviralstate in human cells, activate macrophages, and promote apo-ptosis of virus-infected cells by modulating the expression ofTNF-related apoptosis-inducing ligand (TRAIL) and TRAILreceptors (34). Not surprisingly, IL-18 has been shown to playa role in protective immune responses against these pathogens(12, 40, 41). However, chronic excessive production of thiscytokine has also been implicated in autoimmune phenomena(13, 16, 41). IL-18-induced production of IFN-� and TNF-�from T cells, NK cells, and macrophages may cause destructionof the central nervous system and other tissues, leading to themanifestations of alveolitis and HIV-associated dementia inthese patients. In the context of HIV infections, IL-18 may alsoplay a pathogenic role by enhancing viral replication, as it hasbeen shown in in vitro studies to enhance HIV-1 replication inboth monocytic and T cells by activating p38 MAPK andNF-B as well as by inducing the production of TNF-� andIL-6 (19, 35). The elevated levels of IL-18 in HIV-infected/AIDS patients may, at least theoretically, cause enhanced HIVreplication in both these cell types. Second, recent studies haveshown that IL-18 per se does not induce the differentiation ofTH1 CD4� T cells; it causes the proliferation of IL-12-inducedTH1 CD4� T cells. IL-12 induces the expression of IL-18receptor on these TH1 cells and hence makes them susceptibleto the proliferative effects of IL-18. The receptor does notoccur on TH2 CD4� T cells (45). In the absence of IL-12,IL-18 actually promotes the differentiation of TH2 CD4� Tcells and the induction of immunoglobulin E (IgE) (reviewedin reference 26). Furthermore, IL-18 induces the production ofIL-4 and the release of histamine from basophils and mastcells, which constitutively express IL-18 receptor (48). A re-duced production of IL-12 in HIV infections has been welldocumented (8, 20). Thus, IL-18 may, in fact, be promoting
FIG. 9. TGF-� contents of the plasma samples. The level of TGF-�was measured in the samples with a commercial ELISA kit. The figuredepicts average standard error concentration of this cytokine in thesamples after its activation (total TGF-�). N and P, HIV-seronegativehealthy and HIV-infected donors, respectively. The star on the top ofa bar indicates a significant (P � 0.05) difference between the twogroup means.
FIG. 10. Correlation between the levels of TGF-� in plasma andIL-18 in serum of HIV-infected individuals. The figure depicts a sig-nificant negative correlation between these two parameters as deter-mined by the Pearson’s method.
12454 AHMAD ET AL. J. VIROL.
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

TH2-type responses and the production of IgE in HIV infec-tion especially when IL-12 levels are decreased in the laterstages of the infection. This is supported by the facts thatTH2-type responses predominate and there is enhanced pro-duction of IgE in HIV-infected/AIDS patients (22, 36, 37). Thepredominance of TH2-like responses in these infections hasbeen implicated in the pathogenesis of AIDS (9, 36). Theelevated levels of IL-18 in the sera of HIV-infected/AIDSpatients, shown here, may play a role in promoting TH2-likeresponses in these patients.
IL-18 is produced mainly by monocytes/macrophages in thehuman body. Apart from these cell types, Kupffer cells, kera-tinocytes, dendritic cells, and cells of the adrenal cortex havealso been reported to produce this cytokine (reviewed in ref-erence 11). Surprisingly, our results show that despite elevatedserum levels of IL-18 in HIV-infected/AIDS patients, theirPBMC are compromised in the ability to produce this cytokine.They not only produced less IL-18 spontaneously upon in vitroculture, but also did so after stimulation with LPS, which is wellknown to upregulate IL-18 production in PBMC. The reducedproduction of IL-18 from these PBMC was not due to reducednumbers of monocytes, because we did not find any significantdifference in the percentages and absolute numbers of mono-cytes in PBMC from infected versus control subjects. Theseresults are in accord with those of a previous report (10) thatdescribed a decreased production of IL-18 and IL-18 mRNAin the PBMC of HIV-infected/AIDS patients compared tothose of control subjects. However, He et al. (18) reported adecreased production of this cytokine from the PBMC of HIV-seropositive hemopholiacs but not from PBMC of HIV-sero-positive individuals in response to stimulation with Staphylo-coccus aureus Cowan strain 1. These differences may resultfrom the use of a different stimulating agent.
We also found a decreased expression of activated caspase-1in the PBMC of HIV-infected individuals. Because of the in-volvement of this protease in the processing of precursor IL-18into a biologically active secreted form, its decreased expres-sion may result in a decreased secretion of this cytokine fromthe PBMC of HIV-infected individuals. These findings suggestthat PBMC may not be the source of elevated IL-18 in thesepatients. It is possible, however, that IL-18-producing mono-cytes/macrophages may have migrated out of the circulationinto body tissues. Alternatively, cell types other than PBMC,e.g., Kupffer cells, keratinocytes, dendritic cells, etc., may beproducing more IL-18 in these patients. This is supported by alack of correlation between serum concentration and theamount of production of this cytokine from the PBMC ofHIV-infected donors. It is noteworthy that these two parame-ters correlated significantly (P � 0.05) with each other in HIV-seronegative healthy donors. A likely reason for the reducedproduction of IL-18 from the patients’ PBMC could be anincreased production of immunosuppressive cytokines, e.g.,TGF-� and IL-10 in HIV-infected individuals.
We investigated the role of TGF-� in this study. As reportedin the literature (5; reviewed in reference 21), we found ele-vated levels of this cytokine in the circulation of the patients. Inour in vitro experiments, TGF-� neutralization increased IL-18production from cultured human PBMC. Furthermore, theconcentrations of IL-18 in serum of the patients correlatednegatively with the concentration of TGF-� in plasma. Collec-
tively, these results strongly implicate TGF-� in the compro-mised production of IL-18 from the patients’ PBMC. In fact,this immunosuppressive cytokine is known to inhibit the pro-duction of several other proinflammatory cytokines fromPBMC and has been implicated in AIDS pathogenesis (re-viewed in reference 21). In addition to TGF-�, IL-10 may alsobe involved in the reduced production of IL-18 from the pa-tients’ PBMC. Clearly, further studies are required to learnmore about the role of these cytokines in the regulation ofIL-18 production in HIV-infected individuals.
In summary, we have demonstrated elevated levels of IL-18in the sera of HIV-infected/AIDS patients, which do not cor-relate with their viral loads in plasma or with their CD8� orCD4� T-cell counts. We further demonstrated that despiteelevated levels of this cytokine in their sera, the PBMC fromHIV-infected/AIDS patients are compromised in the ability toproduce IL-18 in in vitro cultures both constitutively and inresponse to LPS stimulation. Elevated levels of TGF-� in thecirculation of these patients may be, at least in part, responsi-ble for this compromise. Our results discount the role ofPBMC in these patients as a source of this enhanced IL-18.Further studies are needed to identify the cells and tissues thatproduced the elevated levels of IL-18 in these patients.
ACKNOWLEDGMENTS
We thank Ilkka Julkunen of the National Public Health InstituteHelsinki (Finland) for providing the pGEM-3zf(�) vector containinghuman IL-18 cDNA, Jose Menezes for useful discussions, DevendraAmre for help in statistical analysis of the data, Mary Blagdon for flowcytometry, and Sylvie Julien for excellent secretarial work.
The work was supported by a grant and a scholarship award from theCanadian Institutes of Health Research (CIHR) to A.A.
REFERENCES
1. Ahmad, A., R. Ahmad, E. Toma, R. Morisset, and J. Menezes. 2000. Im-paired induction of IL-15 in response to herpes simplex virus type 1 infectionin peripheral blood mononuclear cells of HIV-infected patients. AIDS 14:744–746.
2. Ahmad, A., E. Sharif-Askari, L. Fawaz, and J. Menezes. 2000. Innate im-mune response of the human host to exposure with herpes simplex virus type1: in vitro control of the virus infection by enhanced natural killer activity viainterleukin-15 induction. J. Virol. 74:7196–7203.
3. Ahmad, R., L. Knafo, J. Xu, S. T. Sindhu, J. Menezes, and A. Ahmad. 2000.Thrombin induces apoptosis in human tumor cells. Int. J. Cancer 87:707–715.
4. Akira, S. 2000. The role of IL-18 in innate immunity. Curr. Opin. Immunol.12:59–63.
5. Allen, J. B., H. L. Wong, P. M. Guyre, G. L. Simon, and S. M. Wahl. 1991.Association of circulating receptor Fc gamma RIII-positive monocytes inAIDS patients with elevated levels of transforming growth factor-beta.J. Clin. Investig. 87:1773–1779.
6. Cease, K. B., and J. A. Berzofsky. 1994. Toward a vaccine for AIDS: theemergence of immunobiology-based vaccine development. Annu. Rev. Im-munol. 12:923–989.
7. Chehimi, J., S. E. Starr, I. Frank, A. D’Andrea, X. Ma, R. R. MacGregor, J.Sennelier, and G. Trinchieri. 1994. Impaired interleukin 12 production inhuman immunodeficiency virus-infected patients. J. Exp. Med. 179:1361–1366.
8. Chougnet, C., T. A. Wynn, M. Clerici, A. L. Landay, H. A. Kessler, J. Rusnak,G. P. Melcher, A. Sher, and G. M. Shearer. 1996. Molecular analysis ofdecreased interleukin-12 production in persons infected with human immu-nodeficiency virus. J. Infect. Dis. 174:46–53.
9. Clerici, M., and G. M. Shearer. 1994. The Th1-Th2 hypothesis of HIVinfection: new insights. Immunol. Today 15:575–581.
10. David, D., D. Chevrier, M. P. Treilhou, M. Joussemet, B. Dupont, J. Theze,and J. L. Guesdon. 2000. IL-18 underexpression reduces IL-2 levels duringHIV infection: a critical step towards the faulty cell-mediated immunity?AIDS 14:2212–2214.
11. Dinarello, C. A. 1999. IL-18: a TH1-inducing, proinflammatory cytokine andnew member of the IL-1 family. J. Allergy Clin. Immunol. 103:11–24.
12. Fujioka, N., R. Akazawa, K. Ohashi, M. Fujii, M. Ikeda, and M. Kurimoto.
VOL. 76, 2002 PRODUCTION OF IL-18 IN HIV INFECTION 12455
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

1999. Interleukin-18 protects mice against acute herpes simplex virus type 1infection. J. Virol. 73:2401–2409.
13. Furlan, R., G. Martino, F. Galbiati, P. L. Poliani, S. Smiroldo, A. Bergami,G. Desina, G. Comi, R. Flavell, M. S. Su, and L. Adorini. 1999. Caspase-1regulates the inflammatory process leading to autoimmune demyelination.J. Immunol. 163:2403–2409.
14. Ghayur, T., S. Banerjee, M. Hugunin, D. Butler, L. Herzog, A. Carter, L.Quintal, L. Sekut, R. Talanian, M. Paskind, W. Wong, R. Kamen, D. Tracey,and H. Allen. 1997. Caspase-1 processes IFN-gamma-inducing factor andregulates LPS-induced IFN-gamma production. Nature 386:619–623.
15. Giavedoni, L. D., M. C. Velasquillo, L. M. Parodi, G. B. Hubbard, and V. L.Hodara. 2000. Cytokine expression, natural killer cell activation, and phe-notypic changes in lymphoid cells from rhesus macaques during acute infec-tion with pathogenic simian immunodeficiency virus. J. Virol. 74:1648–1657.
16. Gracie, J. A., R. J. Forsey, W. L. Chan, A. Gilmour, B. P. Leung, M. R. Greer,K. Kennedy, R. Carter, X. Q. Wei, D. Xu, M. Field, A. Foulis, F. Y. Liew, andI. B. McInnes. 1999. A proinflammatory role for IL-18 in rheumatoid arthri-tis. J. Clin. Investig. 104:1393–1401.
17. Gu, Y., K. Kuida, H. Tsutsui, G. Ku, K. Hsiao, M. A. Fleming, N. Hayashi,K. Higashino, H. Okamura, K. Nakanishi, M. Kurimoto, T. Tanimoto, R. A.Flavell, V. Sato, M. W. Harding, D. J. Livingston, and M. S. Su. 1997.Activation of interferon-gamma inducing factor mediated by interleukin-1�converting enzyme. Science 275:206–209.
18. He, L., H. Terunuma, H. Hanabusa, A. Iwamoto, S. Oka, F. Tanabe, N.Chiba, M. Kurimoto, M. Ikeda, H. Okamura, J. Dai, Y. Iwatani, T. Ishida,and M. Ito. 2000. Interleukin 18 and interleukin 1� production is decreasedin HIV type 1-seropositive hemophiliacs but not in HIV type 1-seropositivenonhemophiliacs. AIDS Res. Hum. Retrovir. 16:345–353.
19. Klein, S. A., C. Klebba, D. Kauschat, M. Pape, L. Ozmen, D. Hoelzer, O. G.Ottmann, and U. Kalina. 2000. Interleukin-18 stimulates HIV-1 replicationin a T-cell line. Eur. Cytokine Netw. 11:47–52.
20. Liebowitz, D. 1994. Nasopharyngeal carcinoma: the Epstein-Barr virus asso-ciation. Semin. Oncol. 21:376–381.
21. Lotz, M., and P. Seth. 1993. TGF beta and HIV infection. Ann. N. Y. Acad.Sci. 685:501–511.
22. Mazza, D. S., M. H. Grieco, M. M. Reddy, and D. Meriney. 1995. Serum IgEin patients with human immunodeficiency virus infection. Ann. AllergyAsthma Immunol. 74:411–414.
23. McInnes, I. B., J. A. Gracie, B. P. Leung, X. Q. Wei, and F. Y. Liew. 2000.Interleukin 18: a pleiotropic participant in chronic inflammation. Immunol.Today 21:312–315.
24. Nakamura, K., H. Okamura, K. Nagata, T. Komatsu, and T. Tamura. 1993.Purification of a factor which provides a costimulatory signal for gammainterferon production. Infect. Immun. 61:64–70.
25. Nakamura, K., H. Okamura, M. Wada, K. Nagata, and T. Tamura. 1989.Endotoxin-induced serum factor that stimulates gamma interferon produc-tion. Infect. Immun. 57:590–595.
26. Nakanishi, K., T. Yoshimoto, H. Tsutsui, and H. Okamura. 2001. Interleu-kin-18 regulates both th1 and th2 responses. Annu. Rev. Immunol. 19:423–474.
27. Novick, D., S. H. Kim, G. Fantuzzi, L. L. Reznikov, C. A. Dinarello, and M.Rubinstein. 1999. Interleukin-18 binding protein: a novel modulator of theTh1 cytokine response. Immunity 10:127–136.
28. Okamura, H., H. Tsutsi, T. Komatsu, M. Yutsudo, A. Hakura, T. Tanimoto,K. Torigoe, T. Okura, Y. Nukada, and K. Hattori. 1995. Cloning of a newcytokine that induces IFN-gamma production by T cells. Nature 378:88–91.
29. Okamura, H., H. Tsutsui, S. Kashiwamura, T. Yoshimoto, and K. Nakan-ishi. 1998. Interleukin-18: a novel cytokine that augments both innate andacquired immunity. Adv. Immunol. 70:281–312.
30. Pirhonen, J. 2001. Regulation of il-18 expression in virus infection. Scand.J. Immunol. 53:533–539.
31. Pirhonen, J., T. Sareneva, I. Julkunen, and S. Matikainen. 2001. Virusinfection induces proteolytic processing of IL-18 in human macrophages viacaspase-1 and caspase-3 activation. Eur. J. Immunol. 31:726–733.
32. Pirhonen, J., T. Sareneva, M. Kurimoto, I. Julkunen, and S. Matikainen.1999. Virus infection activates IL-1 beta and IL-18 production in humanmacrophages by a caspase-1-dependent pathway. J. Immunol. 162:7322–7329.
33. Puren, A. J., G. Fantuzzi, and C. A. Dinarello. 1999. Gene expression,synthesis, and secretion of interleukin 18 and interleukin 1� are differentiallyregulated in human blood mononuclear cells and mouse spleen cells. Proc.Natl. Acad. Sci. USA 96:2256–2261.
34. Sedger, L. M., D. M. Shows, R. A. Blanton, J. J. Peschon, R. G. Goodwin, D.Cosman, and S. R. Wiley. 1999. IFN-gamma mediates a novel antiviralactivity through dynamic modulation of TRAIL and TRAIL receptor ex-pression. J. Immunol. 163:920–926.
35. Shapiro, L., A. J. Puren, H. A. Barton, D. Novick, R. L. Peskind, R. Shenkar,Y. Gu, M. S. Su, and C. A. Dinarello. 1998. Interleukin 18 stimulates HIVtype 1 in monocytic cells. Proc. Natl. Acad. Sci. USA 95:12550–12555.
36. Shearer, G. M. 1998. HIV-induced immunopathogenesis. Immunity 9:587–593.
37. Shearer, G. M., and M. Clerici. 1998. Cytokine profiles in HIV type 1 diseaseand protection. AIDS Res. Hum. Retroviruses 14(Suppl. 2):S149-S152.
38. Smith, V. P., N. A. Bryant, and A. Alcami. 2000. Ectromelia, vaccinia andcowpox viruses encode secreted interleukin-18-binding proteins. J. Gen.Virol. 81:1223–1230.
39. Stoll, S., G. Muller, M. Kurimoto, J. Saloga, T. Tanimoto, H. Yamauchi, H.Okamura, J. Knop, and A. H. Enk. 1997. Production of IL-18 (IFN-gamma-inducing factor) messenger RNA and functional protein by murine keratin-ocytes. J. Immunol. 159:298–302.
40. Sugawara, I. 2000. Interleukin-18 (IL-18) and infectious diseases, with spe-cial emphasis on diseases induced by intracellular pathogens. Microbes In-fect. 2:1257–1263.
41. Tanaka-Kataoka, M., T. Kunikata, S. Takayama, K. Iwaki, K. Ohashi, M.Ikeda, and M. Kurimoto. 1999. In vivo antiviral effect of interleukin 18 in amouse model of vaccinia virus infection. Cytokine 11:593–599.
42. Torre, D., F. Speranza, R. Martegani, A. Pugliese, F. Castelli, C. Basilico,and G. Biondi. 2000. Circulating levels of IL-18 in adult and paediatricpatients with HIV-1 infection. AIDS 14:2211–2212.
43. Tovo, P. A. 2000. Highly active antiretroviral therapy inhibits cytokine pro-duction in HIV-uninfected subjects. AIDS 14:743–744.
44. Tsutsui, H., K. Matsui, H. Okamura, and K. Nakanishi. 2000. Pathophysi-ological roles of interleukin-18 in inflammatory liver diseases. Immunol. Rev.174:192–209.
45. Xu, D., W. L. Chan, B. P. Leung, D. Hunter, K. Schulz, R. W. Carter, I. B.McInnes, J. H. Robinson, and F. Y. Liew. 1998. Selective expression andfunctions of interleukin 18 receptor on T helper (Th) type 1 but not Th2cells. J. Exp. Med. 188:1485–1492.
46. Xu, J. W., A. Ahmad, M. D’Addario, L. Knafo, J. F. Jones, U. Prasad, R.Dolcetti, E. Vaccher, and J. Menezes. 2000. Analysis and significance ofanti-latent membrane protein-1 antibodies in the sera of patients with EBV-associated diseases. J. Immunol. 164:2815–2822. (Erratum, 164:5532.)
47. Xu, J. W., J. Menezes, U. Prasad, and A. Ahmad. 1999. Elevated serum levelsof transforming growth factor beta1 in Epstein-Barr virus-associated naso-pharyngeal carcinoma patients. Int. J. Cancer 84:396–399.
48. Yoshimoto, T., H. Tsutsui, K. Tominaga, K. Hoshino, H. Okamura, S. Akira,W. E. Paul, and K. Nakanishi. 1999. IL-18, although antiallergic whenadministered with IL-12, stimulates IL-4 and histamine release by basophils.Proc. Natl. Acad. Sci. USA 96:13962–13966.
12456 AHMAD ET AL. J. VIROL.
on May 27, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from