effects of ganglioside administration on experimental autoimmune neuritis induced by peripheral...
TRANSCRIPT

Journal of Neuroimmunology, 43 (1993) 103-112 103 © 1993 Elsevier Science Publishers B.V. All rights reserved 0165-5728/93/$06.00
JNI 02315
Effects of ganglioside administration on experimental autoimmune neuritis induced by peripheral nerve myelin or P2-specific T cell lines
Jiirgen Zielasek a, Stefan Jung a Beate Schmidt a, Gerd Ritter b, Hans-Peter Har tung a
and Klaus Toyka a
a Department of Neurology, University of Wiirzburg, Wiirzburg, Germany and b Department of Immunology, FIDIA Research Laboratories, Abano Terme, Italy
(Received 21 July 1992) (Revision received 15 October 1992)
(Accepted 15 October 1992)
Key words: Demyelination; Nerve inflammation; Drug therapy; Ganglioside antibodies
Summary
We studied the effects of ganglioside administration in two animal models of inflammatory demyelinating polyneuropathy. We administered a mixture of bovine brain gangliosides intraperitoneally to Lewis rats with myelin-induced or T cell line-mediated experimental autoimmune neuritis (EAN). Under the experimental condi- tions we had chosen, we only detected marginal but not statistically significant effects on disease course and severity, as evidenced by motor function, electrophysiological findings, and morphological signs of inflammation and demyelination. There was no significant induction of antibody production against gangliosides, and we did not detect signs of increased cellular reactivity towards gangliosides. We conclude that the administration of ganglio- sides modulates EAN at best marginally, and does not induce a cellular or humoral immune reaction.
Introduction
Gangliosides have been used as a treatment for various degenerative, toxic, and metabolic neuro- pathies in Europe although their efficacy remains con- troversial (Bradley, 1990). More recently, a deleterious effect of ganglioside preparations has been claimed, i.e., induction or even causation of the Guillain-Barr6 syndrome (GBS), an inflammatory demyelinating polyneuropathy, which appears to involve both hu- moral and cellular immune reactions against compo- nents of peripheral nerve myelin (Hartung et al., 1993; Hughes, 1990; Toyka and Hartung, 1992). Although the relevant (auto)antigens are still unknown in this dis- ease, a significant fraction of GBS patients contain in their sera antibodies against gangliosides (Ilyas et al., 1988, 1991a,b, 1992a,b; Fredman et al., 1991; Nobile- Orazio et al., 1992; Quarles et al., 1990; van der Berg,
Correspondence to: J. Zielasek, Neurologische Universit~itsklinik, Josef-Schneider-Str. 11, D-8700 Wiirzburg, Germany.
1992), and these patients appear to have a poorer prognosis (Walsh et al., 1991). Furthermore, a newly defined subgroup of patients with multifocal motor neuropathy has high-titer serum antibodies against the ganglioside GM1 (reviewed by Pestronk, 1991). The pathogenetic significance of these antibodies, mostly of the immunoglobulin M (IgM) class, is unclear. Since gangliosides, especially GM1, are present at a high concentration at nodes of Ranvier (Ganser et al., 1983), an immune reaction to gangliosides at these structures may conceivably impair conduction and lead to struc- tural damage (Santoro et al., 1992).
A recent experimental study found no immunogenic activity of bovine brain gangliosides when co-adminis- tered with the neuritogenic peptide P2 and complete Freund's adjuvant (CFA) in experimental autoimmune neuritis (EAN), an animal model of GBS (Ponzin et al., 1991). In the light of the undetermined role of ganglio- sides as immunogens or immunomodulators in inflam- matory demyelinating polyneuropathies, we studied the effects of ganglioside administration in EAN. Here we investigated the possible immunomodulatory role of
104
gangliosides by injecting gangliosides during or before clinical signs of EAN.
M a t e r i a l s a n d M e t h o d s
One hundred and ten female Lewis rats (8-12 weeks of age) were obtained from the Charles River Wiga Institute (Schweinfurt, Germany), from the Zentralin- stitut ftir Versuchstierkunde (Hannover, Germany), or from our local breeding stock. EAN was induced as described (Jung et al., 1992) by either subcutaneous injection of bovine peripheral nerve and root myelin in CFA, or by intravenous injection of P2 protein-specific T lymphocytes (AT-EAN). A mixture of bovine brain gangliosides (GA; batches 103 and 105; approximate composition 21% GM1, 2% GD3, 40% GDla , 16% GDlb, 19% GTlb and 2% GQlb; F IDIA Research Laboratories, Abano Terme, Italy) was injected in- traperitoneally once daily at a dose of 5 or 50 mg/kg. The latter dose was chosen because it had been shown to partially suppress autoimmune diabetes in NOD mice (Wilberz et al., 1988). Details of the experimental design are given in Table 1. Animals were monitored daily for body weight and disease severity by a masked investigator. Clinical scores were given according to a modified scale (Jung et al., 1992; modified after King et al., 1983): 0 = normal, 1 = reduced tone of the tail, hanging tail tip, 2 = limp tail, 3 = impaired righting, 4 = gait ataxia, abnormal positioning, 5 = mild para- paresis of the hind limbs, 6 = moderate paraparesis of
the hind limbs, 7 = severe paraparesis or paraplegia of the hind limbs, 8 = tetraparesis, 9 = moribund, 10 = death.
Electrophysiological tests Details of electrophysiological testing have been de-
scribed (Heininger et al., 1986). In brief, animals were anesthetized with 60-100 /xl Hypnorm ® (Janssen, Beerse, Belgium) and the left sciatic nerve was investi- gated using an MS92a electromyograph (Medelec, Sur- rey, UK). The protocol included measurement of the amplitudes and latencies of proximally and distally elicited compound muscle action potentials (CMAPs), distally evoked F-waves (latencies and frequency of elicitation after repeated stimulation with a repetition rate of approx. 0.5/s, i.e., F-wave persistence rate), and mixed afferent nerve potential latencies. Both mo- tor and mixed afferent nerve conduction velocities (NCVs) were calculated from these determinations.
Histological studies Animals were killed at the end of the experiment in
pentobarbital anesthesia (Narcoren®, Iffa Merieux, Laupheim, Germany) and perfused through the left cardiac ventricle with heparinized Ringer solution (Fresenius, Bad Homburg, Germany) followed by 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) (pH 7.4). The right sciatic nerve and the lumbar and spinal roots were dissected, fixed overnight in 4% P F A / P B S and used for paraffin (Histowax®, Reichert-Jung, Nussloch, Germany) or epoxy resin em-
TABLE 1
Experimental design
Exp. Type of EAN Treatment Number Duration of Time of no. of rats treatment follow up
1 myelin EAN GA 50 mg/kg 3 d 7-d 21 d 21 PBS 3
2 myelin EAN GA 50 mg/kg 6 d - 2-d 27 d 27 GA 5 mg/kg 6 PBS 6
3 myelin EAN GA 50 mg/kg 10 d - 2-d 20 d 30 PBS 10
4 myelin EAN GA 50 mg/kg 10 d 9-d 38 d 52 PBS 10
5 myelin EAN GA 50 mg/kg 5 d 1-d 9 d 9 PBS 5
6 AT-EAN GA 50 mg/kg 6 d - 2-d 20 d 20 GA 5 mg/kg 6 PBS 6
7 AT-EAN GA 50 mg/kg 6 d -2 d 21 d 21 GA 5 mg/kg 6 PBS 6

bedding. The severity of the inflammatory infiltrate and demyelination were rated in epoxy-embedded 1 /xm thick sections of nerve and roots stained with toluidine blue as described (Heininger et al., 1988). In brief, intraneural blood vessels from the sciatic nerve and lumbar and sacral nerve roots were identified and the surrounding areas examined. Coded sections were evaluated and graded semiquantitatively using the fol- lowing scale: 0 = normal perivascular area, 1 = mild cellular infiltrate adjacent to a vessel, 2 = cellular infil- trate plus demyelinated fibers adjacent to a vessel and in the immediate surroundings, 3 = cellular infiltrates plus demyelinated fibers around a vessel, confluence with infiltrates from more distant vessels. Immunohis- tological studies were performed on 5/~m thick paraf- fin sections using mouse monoclonal antibodies ED1 (rat macrophages, monocytes and dendritic ceils; Serotec, Kidlington, UK), OX6 (polymorphic rat class II antigen, Serotec), W3/13 (rat T lymphocytes, plasma cells, polymorphonuclear cells, stem cells and some brain ceils, Serotec) and 5B5 (negative isotype control, reactive with human prolyl 4-hydroxylase, DAKO, Glostrup, Denmark) using the DAKO alkaline phos- phatase anti-alkaline phosphatase system. All sections were evaluated by an investigator who did not know the treatment group of the specimen.
Antibody studies Blood was obtained from the tail vein of rats during
anesthesia for electrophysiological studies. A masked investigator analyzed coded sera from one experiment with myelin-induced EAN (experiment no. 2; d - 2 and d 26) and one experiment with AT-EAN (experiment no. 7; d 0, 8 and 20) for the presence of antibodies to gangliosides and other antigens using a previously pub- lished dot blot method (Ritter et al., 1990a). We looked for the presence of IgM or IgG antibodies to the individual gangliosides GM1, GM2, GM3, GDla, GDlb, GD3, GTlb, and GQlb; to the ganglioside mixture used for treating the experimental animals; and to the neutral glycolipids asialo-GM1 and galacto- syl ceramide.
Cellular reactivity against gangliosides All reagents were obtained from Gibco, Eggenstein,
Germany, unless otherwise indicated. Mixed spleen cells were obtained from experimental animals at the end of the experiments. In addition, we obtained lymph node tissue for cellular proliferation assays from rats with myelin-induced EAN before symptoms started (day 9; cf. Table 1, exp. no. 5).
We assessed the proliferative response of spleen or lymph node cells in vitro (1 x 105/well in RPMI 1640 containing 10 -5 M 2-mercaptoethanol, 2 mM L-gluta- mine, 0.1 mM non-essential amino acids, 1 mM sodium pyruvate, and 10 mM HEPES) from selected animals
105
by measuring 3H-thymidine incorporation (Amersham Buchler, Braunschweig, Germany) after adding 0.5-2 /xg/ml concanavalin A (Sigma, Deisenhofen, Ger- many) in combination with 0-100/xg/ml mixed bovine brain gangliosides (GA, composition as above) or highly purified single bovine brain gangliosides GM1, GDla, GDlb or GTlb (10 /zg/ml, provided by FIDIA Re- search Laboratories, Abano Terme, Italy). The stimu- lation index was defined as the counts per minute (cpm) obtained from mitogen-stimulated cells divided by the cpm obtained from unstimulated cells. Incorpo- rated radioactivity was measured using the Betaplate® cell harvester and liquid scintillation counting system (Wallac, Turku, Finland).
Statistical analysis Following the initial hypothesis that gangliosides do
influence disease severity, the null hypothesis had to be tested statistically. The two-sided chi-square test was used to compare the presence of clinical scores of 5 or greater between groups, and to compare the persis- tence rates of distally elicited F-waves between groups. The two-sided non-parametric U-test (corrected for ties where necessary) was used to compare the differ- ences of the histological scores, nerve conduction ve- locities, compound muscle action potentials, and F- wave latencies between groups. A P-value of less than 0.05 was considered significant to reject the null hy- pothesis.
Results
Myelin-induced EAN Five experiments were performed. In the pilot ex-
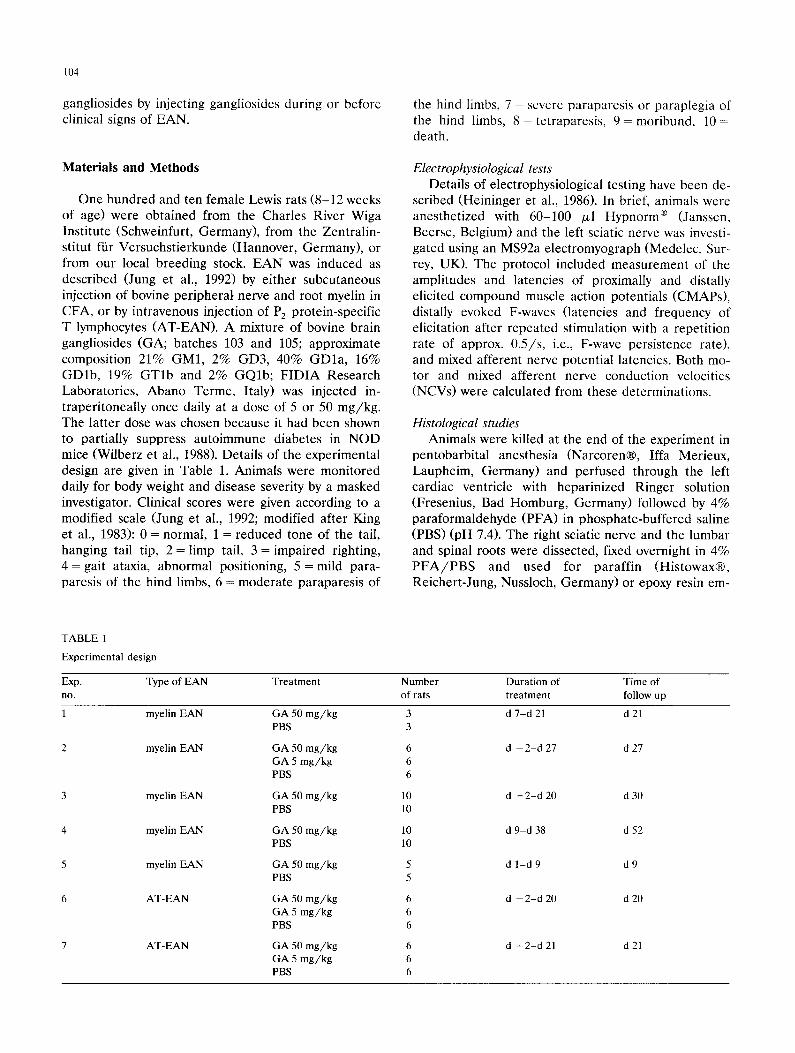
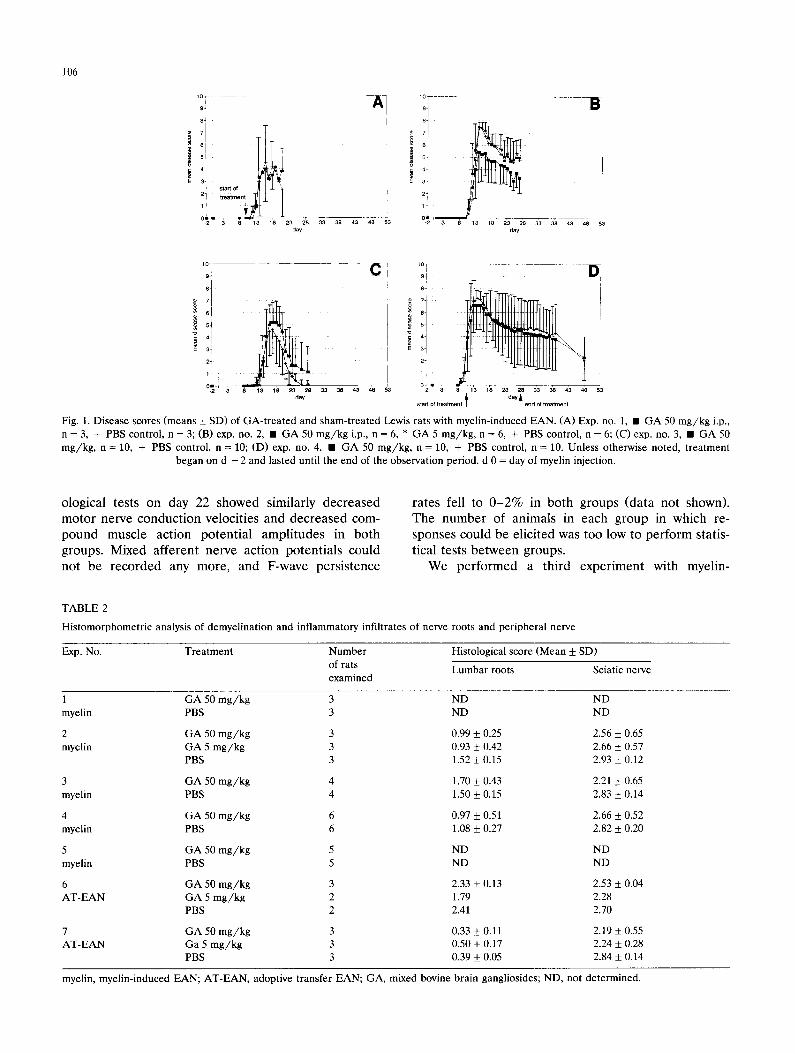
periment (exp. no. 1, cf. Table 1 and Fig. 1A) with a mild disease course (induced by s.c. injection of ap- prox. 15 mg myelin/kg body weight in CFA), ganglio- side administration was begun on day 10 shortly before onset of symptoms but had no effects. However, this experiment indicated that ganglioside administration at the desired concentration was safe and well tolerated. In a more severe form of EAN (exp. no. 2; approx. 19 mg myelin/kg), ganglioside administration started 2 days before myelin injection and appeared to have a mild but statistically non-significant effect on the mean disease score (d 15, Fig. 1B). Since there was a slight reduction in disease severity in rats treated with the high dose (50 mg/kg) of gangliosides, we investigated whether electrophysiological tests or histological analy- ses of peripheral nerves and roots corroborated any evidence for ganglioside effects. A histomorphometric analysis of the extent of demyelination and inflamma- tory infiltration of the roots and of the sciatic nerve showed only a mild or marginal reduction in ganglio- side-treated animals (Fig. 2 and Table 2). Electrophysi-
106
84 i
°1 ! 54 I
3" start of
lJ
day
I0~
E 3
2~ 1-
om ; -2
B]
,'3 t ~ ~ 3 ~ 3'8 4 3 . 'o -~3 day
Io] . . . . . .
2~
o I 5
2 3 8 13 18 23 28 33 38 43 48 53 day
~o N 9~
8-
i3
o = 3
stad of treatment
r !
,3 t . 2%ay~3. 33 38 .3 . . .3
end of treatment
Fig. 1. Disease scores (means _+ SD) of GA-treated and sham-treated Lewis rats with myelin-induced EAN. (A) Exp. no. 1, • GA 50 mg/kg i.p., n = 3, + PBS control, n = 3; (B) exp. no. 2, • GA 50 m g / k g i.p., n = 6, * GA 5 mg/kg, n = 6, + PBS control, n = 6; (C) exp. no. 3, • GA 50 mg/kg, n = 10, + PBS control, n = 10; (D) exp. no. 4, • GA 50 mg/kg, n = 10, + PBS control, n = 10. Unless otherwise noted, treatment
began on d - 2 and lasted until the end of the observation period, d 0 = day of myelin injection.
ological tests on day 22 showed similarly decreased motor nerve conduction velocities and decreased com- pound muscle action potential amplitudes in both groups. Mixed afferent nerve action potentials could not be recorded any more, and F-wave persistence
rates fell to 0 -2% in both groups (data not shown). The number of animals in each group in which re- sponses could be elicited was too low to perform statis- tical tests between groups.
We performed a third experiment with myelin-
TABLE 2
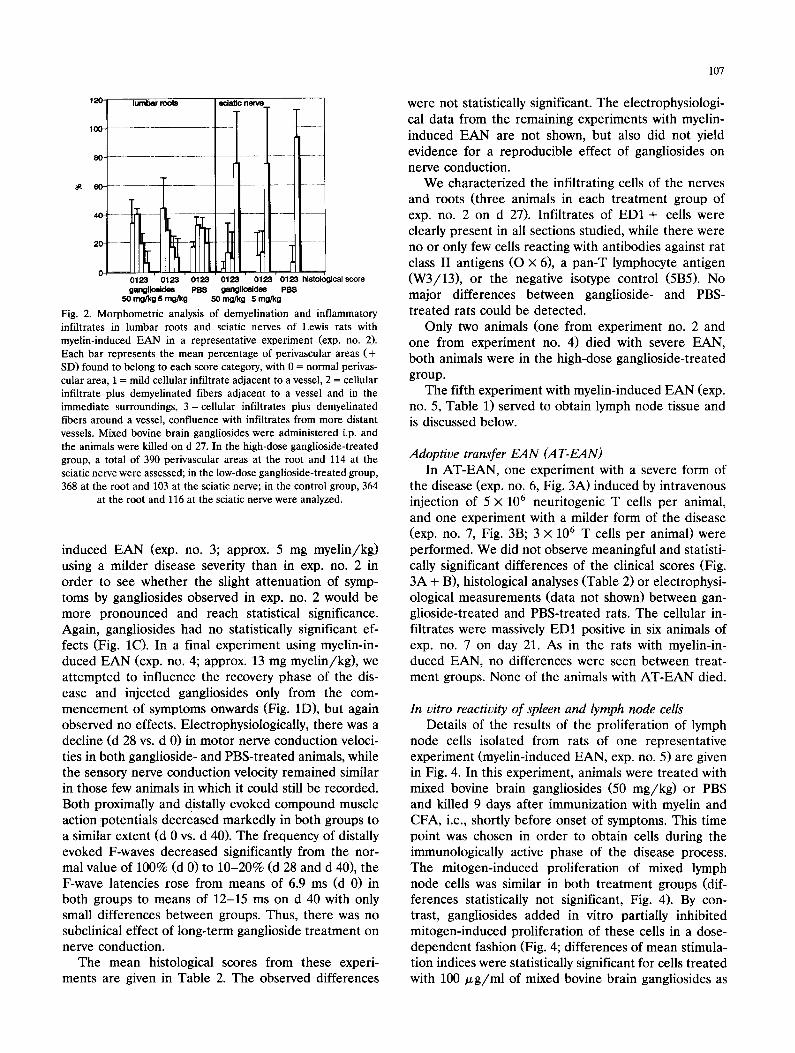
Histomorphometric analysis of demyelination and inflammatory infiltrates of nerve roots and peripheral nerve
Exp. No. Treatment Number of rats examined
Histological score (Mean _+ SD)
Lumbar roots Sciatic nerve
1 GA 50 mg /kg 3 ND ND myelin PBS 3 ND ND
2 GA 50 mg /kg 3 0.99 + 0.25 2.56 _+ 0.65 myelin GA 5 mg /kg 3 0.93 _+ 0.42 2.66 _+ 0.57
PBS 3 1.52 + 0.15 2.93 + 0.12
3 GA 50 mg /kg 4 1.70 + 0.43 2.21 _+ 0.65 myelin PBS 4 1.50 + 0.15 2.83 + 0.14
4 GA 50 mg /kg 6 0.97 _+ 0.51 2.66 + 0.52 myelin PBS 6 1.08 + 0.27 2.82 _+ 0.20
5 GA 50 mg /kg 5 ND ND myelin PBS 5 ND ND
6 GA 50 mg /kg 3 2.33 + 0.13 2.53 + 0.04 AT-EAN GA 5 m g / k g 2 1.79 2.28
PBS 2 2.41 2.70
7 GA 50 m g / k g 3 0.33 + 0.11 2.19 + 0.55 AT-EAN Ga 5 mg /kg 3 0.50 + 0.17 2.24 + 0.28
PBS 3 0.39 + 0.05 2.84 + 0.14
myelin, myelin-induced EAN; AT-EAN, adoptive transfer EAN; GA, mixed bovine brain gangliosides; ND, not determined.
107
120-
100-
80-
40-
20-
0
lumbar roots
0123 0123 0123 0123 0123 0123 histolo icaE score gangllaeides PB8 gangUoelde6 PBS
50 mg/kg 5 mg/kg 50 mg/kg 5 mg/kg
Fig. 2. Morphometric analysis of demyelination and inflammatory infiltrates in lumbar roots and sciatic nerves of Lewis rats with myelin-induced EAN in a representative experiment (exp. no. 2). Each bar represents the mean percentage of perivascular areas ( + SD) found to belong to each score category, with 0 = normal perivas- cular area, 1 = mild cellular infiltrate adjacent to a vessel, 2 = cellular infiltrate plus demyelinated fibers adjacent to a vessel and in the immediate surroundings, 3 = cellular infiltrates plus demyelinated fibers around a vessel, confluence with infiltrates from more distant vessels. Mixed bovine brain gangliosides were administered i.p. and the animals were killed on d 27. In the high-dose ganglioside-treated group, a total of 390 perivascular areas at the root and 114 at the sciatic nerve were assessed; in the low-dose ganglioside-treated group, 368 at the root and 103 at the sciatic nerve; in the control group, 364
at the root and 116 at the sciatic nerve were analyzed.
induced EAN (exp. no. 3; approx. 5 mg myelin/kg) using a milder disease severity than in exp. no. 2 in order to see whether the slight attenuation of symp- toms by gangliosides observed in exp. no. 2 would be more pronounced and reach statistical significance. Again, gangliosides had no statistically significant ef- fects (Fig. 1C). In a final experiment using myelin-in- duced EAN (exp. no. 4; approx. 13 mg myelin/kg), we attempted to influence the recovery phase of the dis- ease and injected gangliosides only from the com- mencement of symptoms onwards (Fig. 1D), but again observed no effects. Electrophysiologically, there was a decline (d 28 vs. d 0) in motor nerve conduction veloci- ties in both ganglioside- and PBS-treated animals, while the sensory nerve conduction velocity remained similar in those few animals in which it could still be recorded. Both proximally and distally evoked compound muscle action potentials decreased markedly in both groups to a similar extent (d 0 vs. d 40). The frequency of distally evoked F-waves decreased significantly from the nor- mal value of 100% (d 0) to 10-20% (d 28 and d 40), the F-wave latencies rose from means of 6.9 ms (d 0) in both groups to means of 12-15 ms on d 40 with only small differences between groups. Thus, there was no subclinical effect of long-term ganglioside treatment on nerve conduction.
The mean histological scores from these experi- ments are given in Table 2. The observed differences
were not statistically significant. The electrophysiologi- cal data from the remaining experiments with myelin- induced EAN are not shown, but also did not yield evidence for a reproducible effect of gangliosides on nerve conduction.
We characterized the infiltrating cells of the nerves and roots (three animals in each treatment group of exp. no. 2 on d 27). Infiltrates of ED1 + cells were clearly present in all sections studied, while there were no or only few cells reacting with antibodies against rat class II antigens (O x 6), a pan-T lymphocyte antigen (W3/13), or the negative isotype control (5B5). No major differences between ganglioside- and PBS- treated rats could be detected.
Only two animals (one from experiment no. 2 and one from experiment no. 4) died with severe EAN, both animals were in the high-dose ganglioside-treated group.
The fifth experiment with myelin-induced EAN (exp. no. 5, Table 1) served to obtain lymph node tissue and is discussed below.
Adoptive transfer EAN (A T-EAN) In AT-EAN, one experiment with a severe form of
the disease (exp. no. 6, Fig. 3A) induced by intravenous injection of 5 x 106 neuritogenic T cells per animal, and one experiment with a milder form of the disease (exp. no. 7, Fig. 3B; 3 x 106 T cells per animal) were performed. We did not observe meaningful and statisti- cally significant differences of the clinical scores (Fig. 3A + B), histological analyses (Table 2) or electrophysi- ological measurements (data not shown) between gan- glioside-treated and PBS-treated rats. The cellular in- filtrates were massively ED1 positive in six animals of exp. no. 7 on day 21. As in the rats with myelin-in- duced EAN, no differences were seen between treat- ment groups. None of the animals with AT-EAN died.
In vitro reactivity of spleen and lymph node cells Details of the results of the proliferation of lymph
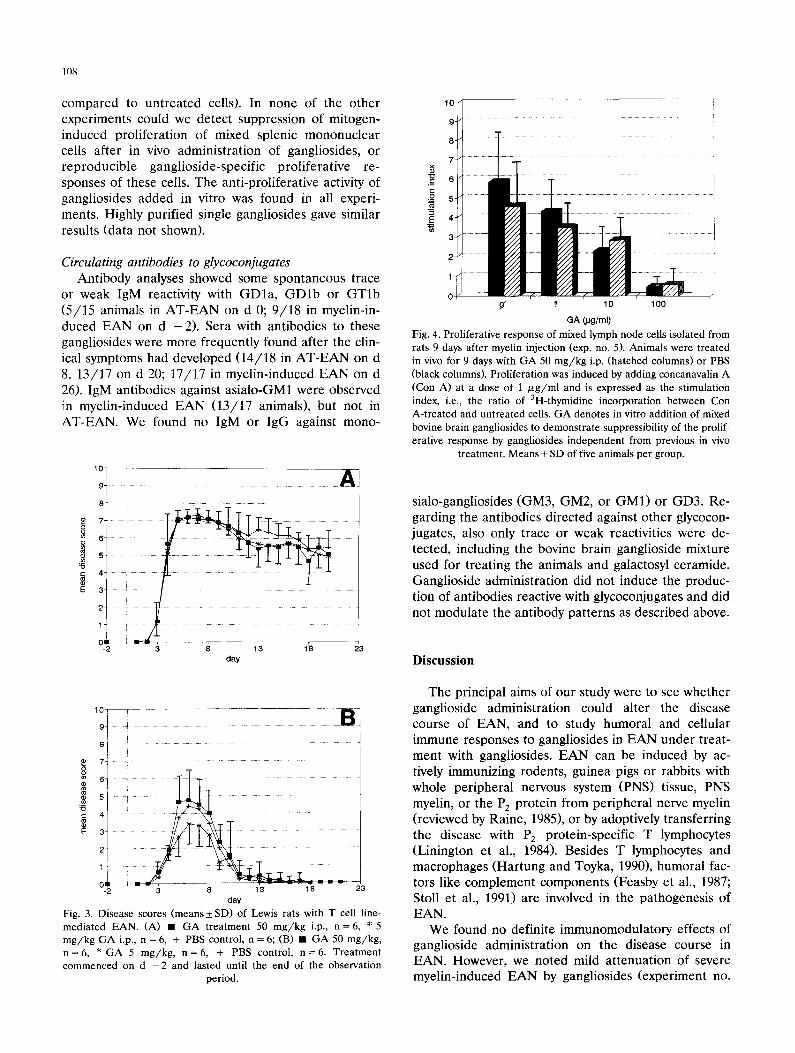
node cells isolated from rats of one representative experiment (myelin-induced EAN, exp. no. 5) are given in Fig. 4. In this experiment, animals were treated with mixed bovine brain gangliosides (50 mg/kg) or PBS and killed 9 days after immunization with myelin and CFA, i.e., shortly before onset of symptoms. This time point was chosen in order to obtain cells during the immunologically active phase of the disease process. The mitogen-induced proliferation of mixed lymph node cells was similar in both treatment groups (dif- ferences statistically not significant, Fig. 4). By con- trast, gangliosides added in vitro partially inhibited mitogen-induced proliferation of these cells in a dose- dependent fashion (Fig. 4; differences of mean stimula- tion indices were statistically significant for cells treated with 100 /zg /ml of mixed bovine brain gangliosides as
108
compared to untreated cells). In none of the other experiments could we detect suppression of mitogen- induced proliferation of mixed splenic mononuclear cells after in vivo administration of gangliosides, or reproducible ganglioside-specific proliferative re- sponses of these cells. The anti-proliferative activity of gangliosides added in vitro was found in all experi- ments. Highly purified single gangliosides gave similar results (data not shown).
Circulating antibodies to glycoconjugates Antibody analyses showed some spontaneous trace
or weak IgM reactivity with GDla, GDlb or GTlb (5/15 animals in AT-EAN on d 0; 9/18 in myelin-in- duced EAN on d -2). Sere with antibodies to these gangliosides were more frequently found after the clin- ical symptoms had developed (14/18 in AT-EAN on d 8, 13/17 on d 20; 17/17 in myelin-induced EAN on d 26). IgM antibodies against asialo-GM1 were observed in myelin-induced EAN (13/17 animals), but not in AT-EAN. We found no IgM or IgG against mono-
=o
E
1 0 ¸
9 . . . . . . . . A
8 . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5°iiiii I i I i 4
3 . . . . . . . . . . . . . . . .
-2 3 8 13 18 23 day
7 8
E 3-
2 . . . . . . .
o2 . . . . . -2 3 8 13 18 23
day
Fig. 3. Disease scores (means 4-_ SD) of Lewis rats with T cell line- mediated EAN. (A) • GA treatment 50 m g / k g i.p., n = 6 , * 5 m g / k g GA i.p., n = 6, + PBS control, n = 6; (B) • G A 50 m g / k g , n = 6 , * GA 5 m g / k g , n = 6 , + PBS control, n = 6 . Trea tment commenced on d - 2 and lasted until the end of the observation
period.
10-
9-
8-
7-
5-
-,~ 3
2
1
0 ~" I 1o 1oo
GA QJglml)
Fig. 4. Proliferative response of mixed lymph node cells isolated from rats 9 days after myelin injection (exp. no. 5). Animals were treated in vivo for 9 days with GA 50 m g / k g i.p. (hatched columns) or PBS (black columns). Proliferation was induced by adding concanavalin A (Con A) at a dose of 1 /~g /ml and is expressed as the stimulation index, i.e., the ratio of 3H-thymidine incorporation between Con A-treated and untreated cells. GA denotes in vitro addition of mixed bovine brain gangliosides to demonstra te suppressibility of the prolif- erative response by gangliosides independent from previous in vivo
treatment. Means + SD of five animals per group.
sialo-gangliosides (GM3, GM2, or GM1) or GD3. Re- garding the antibodies directed against other glycocon- jugates, also only trace or weak reactivities were de- tected, including the bovine brain ganglioside mixture used for treating the animals and galactosyl ceramide. Ganglioside administration did not induce the produc- tion of antibodies reactive with glycoconjugates and did not modulate the antibody patterns as described above.
Discussion
The principal aims of our study were to see whether ganglioside administration could alter the disease course of EAN, and to study humoral and cellular immune responses to gangliosides in EAN under treat- ment with gangliosides. EAN can be induced by ac- tively immunizing rodents, guinea pigs or rabbits with whole peripheral nervous system (PNS) tissue, PNS myelin, or the P2 protein from peripheral nerve myelin (reviewed by Raine, 1985), or by adoptively transferring the disease with P2 protein-specific T lymphocytes (Linington et el., 1984). Besides T lymphocytes and macrophages (Hartung and Toyka, 1990), humoral fac- tors like complement components (Feasby et el., 1987; Stoll et al., 1991) are involved in the pathogenesis of EAN.
We found no definite immunomodulatory effects of ganglioside administration on the disease course in EAN. However, we noted mild attenuation of severe myelin-induced EAN by gangliosides (experiment no.

2), and we cannot rule out the possibility that testing a larger number of animals per group would have led to statistically significant results. In our experience, such small differences in clinical scores are difficult to con- firm given the high inter-individual variation of disease scores in myelin-induced EAN. Thus, even a statisti- cally significant result would have left us in doubt about the biological significance of such a finding. Therefore, we hypothesized that mild immunomodula- tory effects of ganglioside administration may be more pronounced in a milder form of EAN, and tested the same dose of gangliosides with a less severe form of myelin-induced EAN in a larger group of animals (experiment no. 3). Although the clinical effect was again statistically non-significant, administration of gangliosides rather increased disease severity and led to our conclusion that gangliosides have at best marginal effects in myelin-induced EAN. Our results are in contrast to another study reporting a reduced mortality rate of Lewis rats with EAN under treatment with gangliosides (Ledeen et al., 1990). A detailed quantitative analysis was not given by these authors and it is therefore not possible to assess peripheral nerve disease in their animals. Moreover, mortality is generally very low in our experiments (two animals in this study).
More recently, two groups reported reduced severity of EAN in Lewis rats, when mixed bovine brain gan- gliosides were added to the PE protein emulsion at the time of immunization (Ponzin et al., 1991; Wieth61ter et al., 1992). The gangliosides added to the antigen emulsion may have interfered with processing and pre- sentation of P2 protein at the injection site or at the regional lymph nodes. Indeed, a marginal to mild beneficial effect of gangliosides was seen only in the experimental group in which ganglioside administra- tion was commenced before myelin injection (exp. no. 2), which suggests that a similar mechanism may have interfered with antigen processing and presentation in this experiment. However, this mild beneficial effect was not reproduced in a milder form of myelin-induced EAN (exp. no. 3).
Our histological analyses do not reveal a major change in the quantity or the composition of the in- flammatory infiltrate in the ganglioside-treated rats as compared to the PBS-treated rats. This adds to our interpretation of absent immunomodulatory effects.
In addition to experiments with actively induced EAN, we examined the adoptive transfer model, AT- EAN. Here the individual animal has never immuno- logically 'seen' myelin and CFA, and comes down with an inflammatory but less demyelinating disorder of the peripheral nervous system (Heininger et al., 1988). Theoretically, the early recruitment of the host's (re- cipient's) immune cells by the transferred CD4 ÷ T cells might be particularly sensitive to immunomodulat-
109
ing agents (Hartung et al., 1988; Hartung and Toyka, 1990). Our findings with high and low doses of ganglio- sides would not support such an action of gangliosides.
We cannot detect a major effect of ganglioside ad- ministration on cellular or humoral immune reactions in rats with EAN. As regards cellular immune reac- tions, there is no established in vitro test system to assess T cell reactivity against gangliosides. The only reproducible biological effect of gangliosides on cellu- lar immune reactions in vitro is the suppression of mitogen-stimulated proliferation. Mononuclear spleen or lymph node cells from ganglioside-treated animals with EAN reacted towards mitogens or gangliosides in vitro just as did cells from PBS-treated controls. Fur- thermore, gangliosides added in vitro still reduced mi- togen-induced proliferation, which rules out a ganglio- side-refractory state of the mononuclear cells in our experiments and is in keeping with work from other laboratories (reviewed by Marcus, 1984). Regarding humoral immune reactions, we find only trace or weak IgM reactivity against asialo-GM1 or highly sialylated gangliosides in both ganglioside- and PBS-treated rats, but we cannot find antibodies against monosialo-gang- liosides like GM1, which have been implicated in the pathogenesis of lower motor neuron syndromes (see Pestronk, 1991, for a review). Our observations suggest that the disease process by itself may non-specifically trigger a low level of unspecific humoral immune re- sponse against a variety of glycoconjugates including gangliosides, which is not enhanced by parenteral gan- glioside administration, even with multiple doses. Our findings are in keeping with previous reports on low immunogenicity of natural gangliosides in mice and man (Livingston et al., 1989; Ritter et al., 1990b; Gallo et al., 1992). The lack of immunosuppression after in vivo administration of gangliosides is in agreement with a report on ganglioside-treated mice (Presti et al., 1989). This observation also supports our conclusion of a lack of major immunomodulation in vivo induced by gangliosides.
The second question which was addressed by our experiments was whether gangliosides could speed up nerve repair in EAN. In the 20 rats treated and ob- served for a prolonged period of time we did not observe better regeneration or remyelination by admin- istering a high dose of gangliosides in vivo (exp. no. 4).
Our observations do not support claims for a dis- ease-modulating role of gangliosides in inflammatory neuropathies, or major effects on humoral and cellular immune reactions in vivo. While the concept of a disease-modulating effect of systemically administered gangliosides may appear attractive, we cannot confirm such a role for gangliosides in EAN under the experi- mental conditions chosen. A recent epidemiological study on Italian neurological patients receiving ganglio- sides also suggests that exposure to gangliosides does

110
not increase the risk of GBS (Granieri et al., 1991). Further studies are needed to clarify the significance of ganglioside antibodies in sera from patients with in- flammatory neuropathies or lower motor neuron syn- dromes associated with high-titer ganglioside anti- bodies.
Acknowledgements
The authors wish to acknowledge the expert techni- cal assistance of S. Hilbert, V. Preisend6rfer, I. Tschertner and H. Weinand. This study was supported in part by a research grant to J.Z. from FIDIA Phar- maforschung, Munich, Germany.
References
Bradley, W.G. (1990) Critical review of gangliosides and thyrotropin-releasing hormone in peripheral neuromuscular dis- eases. Muscle Nerve 13, 833-842.
Feasby, T.E., Gilbert, J.J., Hahn, A.F. and Neilson, M. (1987) Complement depletion suppresses Lewis rat experimental allergic neuritis. Brain Res. 419, 97 103.
Fredman, P., Vedeler, C.A., Nyland, H., Aarli, J.A. and Svenner- holm, L. (1991) Antibodies in sera from patients with inflamma- tory demyelinating polyradiculoneuropathy react with ganglioside LM1 and sulphatide of peripheral nerve myelin. J. Neurol. 238, 75-79.
Gallo, P., Piccinno, M.G., Tavolato, B., Innocenti, M., Callegaro, L., Kirschner, G., Bruno, R., Chizzolini, C. and Fiori, M.G. (1992) Effect of parenteral administration of GM1 on cytokines and anti-ganglioside antibody patterns. Preliminary report in normal human individuals. J. Neuroimmunol. 36, 81 86.
Ganser, A.L., Kirschner, D.A. and Willinger, M. (1983) Ganglioside localization on myelinated nerve fibers by cholera toxin binding. J. Neurocytol. 12, 921-938.
Granieri, E., Casetta, I., Govoni, V., Tola, M.R., Paolino, E. and Rocca, W.A. (1991) Ganglioside therapy and Guillain-Barr~ syn- drome. Neuroepidemiology 10, 161-169.
Hartung, H.P. and Toyka, K.V. (1990) T cell and macrophage activa- tion in experimental autoimmune neuritis and Guillain-Barr~ syndrome. Ann. Neurol. 27 (Suppl.), $57-$63.
Hartung, H.P., Heininger, K., Schiifer, B., Fierz, W. and Toyka, K.V. (1988) Immune mechanisms in inflammatory polyneuropathy. Ann. NY Acad. Sci. 540, 122-161.
Hartung, H.P., Toyka, K.V. and Pollard, J.D. (1993) Immunology of the Guillain-Barr~-Syndrome. Neurology (in press).
Heininger, K., Stoll, G., Linington, C., Toyka, K.V. and Wekerle, H. (1986) Conduction failure and nerve conduction slowing in exper- imental allergic neuritis induced by P2-specific T-cell lines. Ann. Neurol. 19, 44 49.
Heininger, K., Schfifer, B., Hartung, H.P., Fierz, W., Linington, C. and Toyka, K.V. (1988) The role of macrophages in experimental autoimmune neuritis induced by a P2-specific T-cell line. Ann. Neurol. 23, 326-331.
Hughes, R.A.C. (1990) Guillain-Barr6 Syndrome. Springer, Heidel- berg.
llyas, A.A., Willison, H.J., Quarles, R.H., Jungalwala, F.B., Corn- blath, D.R., Trapp, B.D., Griffin, D.E., Griffin, J.W. and Mc- Khann, G.M. (1988) Serum antibodies to gangliosides in Gudlain-Barr6 syndrome. Ann. Neurol. 23, 440-447.
llyas, A.A., Mithen, F.A., Dalakas, M.C., Wargo, M., Chen, Z.W.. Bielory, L. and Cook, S.D. (1991a) Antibodies to sulfated glyco- lipids in Guillain-Barr& syndrome. J. Neurol. Sci. 105, 108-117.
llyas, A.A., Mithen, F.A., Chen, Z.W. and Cook, S.D. (1991b) Search for antibodies to neutral glycolipids in sera of patients with Guillain-Barr6 syndrome. J. Neurol. Sci. 102, 67 75.
Ilyas, A.A., Mithen, F.A.. Chen, Z.W. and Cook, S.D. (1992a) Anti-GMl IgA antibodies in Guillain-Barr~ syndrome. J. Neuro- immunol. 36, 69-76.
Ilyas, A.A., Mithen, F.A., Dalakas, M.C., Chen, Z.W. and Cook, S.D. (1992b) Antibodies to acidic glycolipids in Guillain-Barr~ syn- drome and chronic inflammatory demyelinating polyneuropathy. J. Neurol. Sci. 107, 111-121.
Jung, S., Kr~imer, S., Schluesener, H.J., Hiinig, T., Toyka, K. and Hartung, H.P. 11992) Prevention and therapy of experimental autoimmune neuritis by an antibody against T cell receptors-a//3. J. lmmunol. 148, 3768-3775.
King, R.H.M., Craggs, R.I., Gross, M.L.P., Tompkins, C. and Thomas, P.K. (1983) Suppression of experimental allergic neuritis by cy- closporin-A. Acta Neuropathol. (Berl.) 59, 262-268.
Ledeen, R.W., Oderfeld-Nowak, B., Brosnan, C.F. and Cervone, A. (1990) Gangliosides offer partial protection in experimental aller- gic neuritis. Ann. Neurol. 27 (Suppl.), $69-$74.
Linington, C., lzumo, S., Suzuki, M., Uyemura, K., Meyermann, R. and Wekerle, H. (1984) A permanent rat T cell line that mediates experimental allergic neuritis in the Lewis rat in vivo. J. Immunol. 133, 1946-1950.
Livingston, P.O., Ritter, G., Oettgen, H.F. and Old, L.J. 11989) Immunization of melanoma patients with purified gangliosides. In: H.F. Oettgen (Ed.), Gangliosides and Cancer. VCH, Wein- heim, pp. 293-299.
Marcus, D.M. (1984) A review of the immunogenic and immunomod- ulatory properties of glycosphingolipids. Mol. Immunol. 21, 1083-1091.
Nobile-Orazio, E., Carpo, M., Meucci, N., Grassi, M.P., Capitani, E., Sciacco, M., Mangoni, A. and Scarlato, G. (1992) Guillain-Barr& syndrome associated with high titers of anti-GM1 antibodies. J. Neurol. Sci. 109, 200-206.
Pestronk, A. 11991) Motor neuropathies, motor neuron disorders, and antiglycolipid antibodies. Muscle Nerve 14, 927-936.
Ponzin, D., Menegus, A.M., Kirschner, G., Nunzi, M.G., Fiori, M.G. and Raine, C.S. (1991) Effects of gangliosides upon the expres- sion of autoimmune demyelination in the peripheral nervous system. Ann. Neurol. 30, 678-685.
Presti, D., Callegaro, L., Toffano, G. and Marcus, D.M. (1989) Lack of suppression by gangliosides of humoral or cellular immunity in vivo. J. Neuroimmunol. 22, 233-239.
Quarles, R.H., Ilyas, A.A. and Willison, H,J. (1990) Antibodies to gangliosides and myelin proteins in Guillain-Barr~ syndrome. Ann. Neurol. 27 (Suppl.), $48-$52.
Raine, C.S. (1985) Experimental allergic encephalomyelitis and ex- perimental allergic neuritis. In: J.C. Koetsie (Ed.), Handbook of Clinical Neurology, Vol. 47. Elsevier, Amsterdam, pp. 429-466.
Ritter, G., Boosfeld, E., Markstein, E., Yu, R.K., Ren, S., Stallcup, W.B., Oengen, H.F., Old, L.J. and Livingston, P.O. (1990a) Biochemical and serological characteristics of natural 9-O-acetyl GD3 from human melanoma and bovine buttermilk and chemi- cally modified O-acetylated GD3. Cancer Res. 50, 1403-1410.
Riner, G., Boosfeld, E., Calves, IVI.J., Oettgen, H.F., Old, L.J. and Livingston, P.O. (1990b) Antibody response to immunization with purified GD3 ganglioside and GD3 derivatives (lactones, amide and gangliosidol) in the mouse. Immunobiology 182, 32-43.
Santoro, M., Uncini, A., Corbo, M., Staugaitis, S.M., Thomas, F.P., Hays, A.P. and Latov, N. (1992) Experimental conduction block induced by serum from a patient with anti-GM1 antibodies. Ann. Neurol, 31, 385-39/I.
Stoll, G., Schmidt, B., Jander, S., Toyka, K.V. and Hartung, H.P.

(1991) Presence of the terminal complement complex (C5b-9) precedes myelin degradation in immune-mediated demyelination of the rat peripheral nervous system. Ann. Neurol. 30, 147-155.
Toyka, K.V. and Hartung, H.P. (1992) Circulating immune factors. In: A.K. Asbury, G.M. McKhann and W.I. McDonald (Eds.), Diseases of the Nervous System: Clinical Neurobiology, 2nd edn., Vol. II, W.B. Saunders, Philadelphia, PA, pp. 1396-1409.
Van den Berg, L.H., Marrink, J., de Jager, A.E.J., de Jong, H.J., van Imhoff, G.W., Latov, N. and Sadiq, S.A. (1992) Anti-GM1 anti- bodies in patients with Guillain-Barr6 syndrome. J. Neurol. Neu- rosurg. Psychiatry 55, 8-11.
111
Walsh, F.S., Cronin, M., Koblar, S., Doherty, P., Winer, J., Leon, A. and Hughes, R.A.C. (1991) Association between glycoconjugate antibodies and Campylobacter infection in patients with Guillain-Barr6 syndrome. J. Neuroimmunol. 34, 43-51.
Wieth61ter, H., Schabet, M., Stevens, A., Melms, A., Sommer, N. and Weller, M. (1992) Influence of gangliosides on experimental allergic neuritis. J. Neuroimmunol. 38, 221-228.
Wilberz, S., Herberg, L. and Renold, A.E. (1988) Gangliosides in vivo reduce diabetes incidence in non-obese diabetic mice. Dia- betologia 31, 855-857.