three novel clade b serine protease inhibitors from disk abalone, haliotis discus discus: molecular...
TRANSCRIPT

lable at ScienceDirect
Fish & Shellfish Immunology 45 (2015) 334e341
Contents lists avai
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate/ fs i
Full length article
Three novel clade B serine protease inhibitors from disk abalone,Haliotis discus discus: Molecular perspectives and responses toimmune challenges and tissue injury
S.D.N.K. Bathige a, b, Navaneethaiyer Umasuthan a, b, G.I. Godahewa a, b, Ilson Whang a, b,Chul Kim c, Hae-Chul Park d, **, Jehee Lee a, b, *
a Department of Marine Life Sciences, School of Marine Biomedical Sciences, Jeju National University, Jeju Self-Governing Province 690-756,Republic of Koreab Fish Vaccine Research Center, Jeju National University, Jeju Special Self-Governing Province 690-756, Republic of Koreac Informatics Development & Management Group, Korea Institute of Oriental Medicine, 1672 Yuseongdae-ro, Yuseong-gu, Daejeon 305-811,Republic of Koread Graduate School of Medicine, Korea University, Ansan, Gyeonggido 425-707, Republic of Korea
a r t i c l e i n f o
Article history:Received 20 February 2015Received in revised form7 April 2015Accepted 16 April 2015Available online 24 April 2015
Keywords:Serine protease inhibitor (SERPIN)Haliotis discus discusImmune responseTissue injury
* Corresponding author. Marine Molecular GeneticLife Sciences, College of Ocean Science, Jeju NationalAra-Dong, Jeju 690-756, Republic of Korea. Tel.: þ82fax: þ82 64 756 3493.** Corresponding author. Tel.: þ82 31 412 6712; fax
E-mail address: [email protected] (J. Lee).
http://dx.doi.org/10.1016/j.fsi.2015.04.0201050-4648/© 2015 Elsevier Ltd. All rights reserved.
a b s t r a c t
Serine protease inhibitors (SERPINs) control cellular protease activity in order to maintain cellular ho-meostasis. The immune and inflammatory responses of invertebrate clade B SERPINs have not beenwidely reported. In the present study, three proteins with high similarity to clade B SERPINs, referred toas AbSERPIN-1, AbSERPIN-2 and AbSERPIN-3, were identified from disk abalone (Haliotis discus discus).While AbSERPIN-1 (399 aa) was of a typical size for this protein class, AbSERPIN-2 (506 aa) andAbSERPIN-3 (532 aa) were relatively larger. Bioinformatic analysis revealed the characteristic SERPINdomain in each AbSERPIN. In addition, the N-terminal region of both AbSERPIN-2 and AbSERPIN-3contained a predicted low complexity region (LCR) and a signal peptide, suggesting that these pro-teins are secretory proteins and are, thus, novel peptides. Tertiary structural models of the AbSERPINshighlighted their structural and functional conservation. Ubiquitous expression of AbSERPIN transcriptswas evaluated by quantitative real time PCR (qPCR) analysis in seven tissue types. AbSERPIN-1, AbSERPIN-2, and AbSERPIN-3 transcript levels were highest in mantle, hemocytes, and muscles, respectively.Temporal expression analysis revealed that AbSERPINs were significantly (P < 0.05) elevated in hemo-cytes during the early/middle stages following the injection of a bacterial pathogen (Vibrio para-haemolyticus or Listeria monocytogenes) or an immuno-stimulant (lipopolysaccharide). Moreover, mantletissue injury led to significant changes in the temporal expression of AbSERPIN mRNA. Specifically,transcription of AbSERPIN-1 and AbSERPIN-3 was considerably up-regulated, while expression ofAbSERPIN-2 was suppressed. These results suggest a potential role of AbSERPINs in response to pathogeninvasion and tissue injury in disk abalone.
© 2015 Elsevier Ltd. All rights reserved.
1. Introduction
Protease inhibitors (PIs) are the main inhibitors of protease ac-tivity. Proteases are involved in various innate immune processes,
s Lab, Department of MarineUniversity, 66 Jejudaehakno,64 754 3472;
: þ82 31 412 6729.
including phagocytosis, synthesis of antimicrobial peptides, com-plement activation, blood coagulation, as well as in general meta-bolism [1,2]. PIs are found in all biological entities, from viruses tohumans [3], and play crucial roles, since unnecessary or excessprotease activity may damage cells, organs, or the entire organism.At present, 74 PI families are described in the MEROPS database(http://merops.sanger.ac.uk) [4]. According to the mechanism ofinhibition, there are two major classes of PIs: tight-binding in-hibitors and trapping inhibitors. Tight-binding inhibitors form astrong but reversible complex with their target proteases. Incontrast, trapping inhibitors form irreversible complexes. Trapping

S.D.N.K. Bathige et al. / Fish & Shellfish Immunology 45 (2015) 334e341 335
inhibitors include PI types I39, I50, and I4, the class known as serineprotease inhibitors or SERPINs [3].
SERPINs, which make up the largest and most broadly distrib-uted PI family, have been identified in animals, plants, bacteria, andviruses [4,5]. Thirteen SERPIN members belonging to clade B arewell-documented in human [6,7]; however, these remain to be fullycharacterized in other species. Particularly, the characterization ofSERPIN B proteins in mollusks is very limited. Typically, SERPINsconsist of ~400 amino acids and a characteristic three-dimensional(3D) folding structurewith three b-sheets (AeC) and nine a-helices.The region important for inhibitory activity, known as the reactive-center loop (RCL), is positioned at the C-terminus. The RCL containsa scissile bond between two residues (P1 and P10) that can becleaved by the cognate target protease. The specificity of inhibitoryactivity depends on P1, the key amino acid residue. Interactionbetween the SERPIN and the target protease via the RCL inactivatesthe protease [6,8]. Although the majority of SERPINs inhibit serineproteases, some SERPINs also inhibit caspases [8] and papain-likecysteine proteases [9]. A number of studies have also demon-strated that PIs, including SERPIN members, have a potential role inresponses to tissue injury [10e12].
In mollusks, several PIs have been identified and reported to beinvolved in host defense via inhibition of proteases secreted by theparasitic pathogen Perkinsus marinus [13e15]. In addition, severalstudies have analyzed the effects of PIs against bacterial invasion.For instance, Faisal et al. report that plasma from Crassostrea vir-ginica and Crassostrea gigas inhibit Vibrio vulnificus protease activity[15]. PIs identified from Argopecten irradians [12] and Chlamys far-reri [16] demonstrated immune response upon Vibrio anguillaruminfection.
Disk abalone, Haliotis discus discus, is a gastropod species ofgreat economic importance in Eastern Asia. However, in the pastfew decades, frequent disease outbreaks due to microbial patho-gens have affected the aquaculture production of abalone [17e20].The study of PIs, such as SERPINs, may contribute to our knowledgeof immune defense mechanisms and wound healing capacity,thereby enabling improved disease management strategies ingastropod farming. Accordingly, the main objectives of this studywere (1) to characterize the cDNA and translated sequences of threeSERPIN members from disk abalone, (2) to determine SERPINmRNAexpression levels in hemocytes upon bacterial infection, and (3) tostudy the role of SERPINs in the wound healing process.
2. Materials and methods
2.1. Identification of SERPIN genes from disk abalone
Three SERPIN homologs were isolated from a previously con-structed disk abalone transcriptome database [21]. NCBI BLASTanalysis revealed high similarity to SERPIN clade B. The threeidentified genes were designated AbSERPIN-1, AbSERPIN-2, andAbSERPIN-3.
2.2. Bioinformatic analysis
Analysis of the nucleotide and deduced amino acid sequences ofthe identified AbSERPINs, and the search for orthologous se-quences, were performed using BLAST (http://www.ncbi.nlm.nih.gov/blast). AbSERPIN sequences were further analyzed withBLAST in the MEROPS database under default parameters [4].SERPIN signature sequences and domain architectural featureswere identified by ScanProsite (http://prosite.expasy.org/scanprosite/) and SMART (Simple Modular Architecture ResearchTool, http://smart.embl-heidelberg.de/), respectively. SignalP 3.0(http://www.cbs.dtu.dk/services/SignalP/) was used to predict
signal peptides and ClustalW2 (http://www.Ebi.ac.uk/Tools/clustalw2) [22] was used to align multiple sequences. Based onthe alignments, a phylogenetic tree was constructed with MEGAversion 5.05 [23] using a maximum likelihood (ML) algorithm [23].The three-dimensional (3D) structure of each AbSERPIN wasmodeled using SWISS-MODEL [24] and visualized with PyMOLmolecular graphic software version 1.3 [25].
2.3. Challenge experiments
2.3.1. Animal rearingHealthy disk abalones (average weight of 50 g) were purchased
from Youngsoo commercial abalone farm in Jeju Island, Republic ofKorea, and acclimated to laboratory conditions (sand-filteredseawater at a salinity of 34 ± 0.6‰ and a temperature of 20 ± 1 �C)in theMarine and Environmental Research Institute of Jeju NationalUniversity. Animals were fed fresh marine seaweed (Undaria pin-natifida) prior to the experiment.
2.3.2. Tissue distribution analysisTo analyze the tissue-specific expression of AbSERPIN mRNA,
several tissues (including that of the gills, digestive tract, gonads,hepatopancreas, muscles, and mantle) were dissected from fourhealthy disk abalones. To obtain hemocytes, hemolymph wascollected from the pericardial cavities of healthy animals andimmediately centrifuged at 3000�g for 10 min at 4 �C. All tissueswere snap frozen in liquid nitrogen and stored at �80 �C untilfurther use.
2.3.3. Immune challenge experimentsTo study the expression patterns of AbSERPINs in response to
pathogen infection, acclimatized disk abalones were randomlyassigned to one of four groups: two groups were subjected tobacterial challenge, one group was subjected to immune stimulantchallenge, and the remaining group served as a control. For thebacterial challenge, 100 mL of 1 � 104 CFUmL�1 live bacteria (eitherListeria monocytogenes or Vibrio parahaemolyticus) in saline (0.9%NaCl) was administered intramuscularly. Animals in the third groupwere injected with 100 mL LPS (5 mg mL�1, Escherichia coli 055:B5;Sigma), dissolved in saline. Similarly, an equal volume (100 mL) ofsaline was injected in the control group. Four disk abalones fromeach group were sacrificed to collect hemocytes at different timepoints (3, 6, 12, 24, 48, 72, and 120 h) post-injection (p.i.). Inaddition, hemocytes collected from four un-injected abalones wereused to establish baseline expression levels (0 h). All collected tis-sues were snap frozen in liquid nitrogen and stored at �80 �C untilfurther use.
2.3.4. Tissue injuryTo understand the involvement of AbSERPINs in wound healing,
a group of disk abalones was subjected to tissue injury, which wasachieved by damaging the abalone mantle and shell as previouslydescribed [26]. Mantle tissue was collected from three individualsat different time points post-injury (3, 6, 9, 12, 24, 48, 96, and120 h). Mantle tissue collected from three uninjured abalonesserved as a control (0 h). All collected tissues were stored at �80 �Cuntil RNA extraction.
2.4. RNA extraction and quantitative real-time PCR (qPCR)
Pools of isolated tissues from four abalones were homogenizedusing a homogenizer (Polytron®, USA) and total RNA was extractedusing TRIzol® reagent (Sigma). Total RNA concentration wasdetermined by measuring absorbance at 260 nm and purity wasdetermined by comparing absorbance at 260 and 280 nm. Then,

S.D.N.K. Bathige et al. / Fish & Shellfish Immunology 45 (2015) 334e341336
first-strand cDNAwas synthesized using PrimeScript™ First StrandcDNA Synthesis Kit (TaKaRa, Japan). In brief, a mixture of 2.5 mg oftotal RNA, 50 mM oligo dT primer, 10 mM dNTP mix, and RNase-freewater was incubated at 65 �C for 5 min, then transferred to an icebath for at least 1 min. Subsequently, 5� PrimeScript™ buffer, 20 URNase inhibitor, and 200 U PrimeScript™ RTase were added to afinal volume of 20 mL. The reaction mixture was incubated at 42 �Cfor 1 h, followed by incubation at 70 �C for 15 min to stop the re-action. Finally, the synthesized cDNA was diluted 40-fold in prep-aration for SYBR® Green fluorescent qPCR assay.
Gene-specific primers for AbSERPIN-1, AbSERPIN-2, and AbSER-PIN-3 (Supplementary Table 1) were designed according to MIQEguidelines [27], with 50% GC content, 60 �C Tm, and amplicon sizesof 151, 142, and 131 bp, respectively. qPCR was performed usingReal Time System TP800 Thermal Cycler Dice™ (TaKaRa, Japan)with fluorescence active SYBR Green and using abalone ribosomalprotein L5 (GenBank accession: EF103443) as an internal control[28]. The qPCR amplificationwas executed in a total volume of 15 mLcomprising 7.5 mL of 2 � SYBR® Premix Ex Taq™ (Takara, Japan),0.6 mL of each gene-specific primer (10 pmol mL�1), 4 mL dilutedcDNA, and 2.3 mL PCR-grade water. qPCR was performed by meansof a three-step method, including incubation at 95 �C for 10 s, 45cycles at 95 �C for 5 s, 58 �C for 20 s and 72 �C for 20 s, and finally adissociation condition of 95 �C for 15 s, 60 �C for 30 s, and 95 �C for15 s to assess the amplification of single product. The qPCR wasperformed in triplicate and each AbSERPIN Ct value was normalizedwith respect to ribosomal protein L5 Ct values. The Livak method[29] was applied to calculate relative expression levels of AbSERPINsand fold changes at each time point, relative to the saline control.
2.5. Statistical analysis
For tissue-specific expression data, the mean ± standard devi-ation (SD) was calculated for each tissue type, and compared be-tween tissues by one-way ANOVA followed by Duncan's MultipleRange test, using SPSS 11.5 software (Chicago, IL, USA). All the otherexperimental results were subjected to t-test analysis usingGraphPad (GraphPad Software, Inc., USA). A P value of <0.05 wasconsidered statistically significant.
3. Results
3.1. In silico analysis of AbSERPINs
3.1.1. AbSERPIN sequence characterizationAbSERPIN-1 cDNA was consisted of a 1928 bp sequence (Gen-
Bank accession number: KP681239), which included an 1197 bpORF encoding 399 amino acids (aa). The predicted molecular massof AbSERPIN-1 was 44.1 kDa and its theoretical isoelectric point (pI)was 5.80. SMART server predicted a characteristic SERPIN domain(E-value¼ 3.41� 10�150) covering almost the entiremature protein(amino acid positions 26e385). The ScanProsite tool predicted aSERPIN signature (358FKADHPFLFLI368) at the C-terminus of theSERPIN domain. Four RNA instability motifs (ATTTA) were identi-fied at the 30-untranslated region (UTR) (Supplementary Fig. 1a).
The 2637 bp cDNA sequence of AbSERPIN-2 (GenBank accessionnumber: KP681240) contained a 1518 bp ORF encoding 506 aa,constituting a protein with an estimated molecular mass of55.8 kDa and theoretical pI of 9.49. SignalP predicted a putativesignal peptide of 20 aa at the N-terminus. SMART predicted both alow-complexity region (LCR) and a SERPIN domain (E-value ¼ 8.02 � 10�106) in the mature protein. Three putative sitesfor N-linked glycosylation (212NATL215, 244NISS247, and 491NVSK494)were recognized within the SERPIN domain. The SERPIN signature(478FRCDHPFLFFI488) was identified at the C-terminus of this
domain. In addition, two RNA instability motifs and a poly-adenylation signal (AATAAA) were detected at the 30-UTR(Supplementary Fig. 1b).
The AbSERPIN-3 cDNA sequence was 2089 bp in length (Gen-Bank accession number: KP681241) with a 1596 bp ORF encoding532 aa. SignalP revealed a 21 aa signal peptide at the N-terminus.The predicted molecular mass and theoretical pI of the maturepeptide were 55.7 kDa and 9.46, respectively. SMART programpredicted an LCR and characteristic SERPIN domain (E-value ¼ 2.45 � 10�110) in the mature AbSERPIN-3 peptide. Withinthe SERPIN domain, three potential N-glycosylation sites(240NATL243, 272NISL275, and 517NVSK520) were identified. The SER-PIN signature (504IRCDHPFLFII514) was predicted at the N-terminalregion of AbSERPIN-3 by ScanProsite. A single RNA instability motifand a polyadenylation signal (AATAAA) were also identified in theAbSERPIN-3 gene (Supplementary Fig. 1c).
3.1.2. Sequence comparison, tertiary structure, and phylogeneticanalysis
The SERPIN domain of each disk abalone SERPIN protein wascompared with those of several orthologs, from both invertebratesand vertebrates, particularly those in the SERPIN B family(Supplementary Table 2), which NCBI BLAST showed to have greathomology to AbSERPINs. The SERPIN domain of AbSERPIN-1exhibited 37e44% overall identity with vertebrate and inverte-brate orthologs. C. gigas SERPIN B8 shared the highest degree ofidentity with AbSERPIN-1 (43.5%), but comparatively lower identitywith AbSERPIN-2 (39.6%) and AbSERPIN-3 (37.1%). AbSERPIN-2 andAbSERPIN-3 shared 64.4% identity between them and less than 40%identity with other homologs. Multiple SERPIN sequence align-ments revealed a conserved hinge region and SERPIN signature inthe RCL region (Fig. 1b). The amino acid residues in the putativescissile bond (P1eP10) of AbSERPIN-1 (Arg350 and Ser351),AbSERPIN-2 (Arg466 and Ser467), and AbSERPIN-3 (Gln494 andThr495) were recognized in the central portion of each RCL. SWISS-MODEL, an automated homology modeling server, was used togenerate 3D structures of the AbSERPINs, using human SERPIN B3(PDB: 2zv6) as a template. Fig. 1c shows the native structures ofAbSERPINs; each was made up of three b sheets (A, B, and C) andnine prominent a helices. The residues in the RCL responsible forinteraction with target proteases (P1eP10) are located near the topof each SERPIN scaffold (Fig. 1c). According to the ML phylogeneticanalysis (Fig. 2), AbSERPIN-1 clustered with oyster (C. gigas) SERPINB8. However, AbSERPIN-2 and AbSERPIN-3 formed a clade with thecrustacean mitten crab SERPIN, even though the other crustaceanhomologs formed a separate clade. In addition, a distinct evolu-tionary branch consisted of mammalian and other vertebrateSERPINs emerged as a major branch.
3.2. Transcriptional profiles of AbSERPINs
3.2.1. Tissue-specific expression of AbSERPIN transcriptsExpression of AbSERPIN transcripts was assessed by the SYBR
Green qPCR method. Amplification efficiencies for the internalreference (ribosomal protein L5), AbSERPIN-1, AbSERPIN-2 andAbSERPIN-3 were 2.05, 2.01, 2.03, and 2.04, respectively. qPCRanalysis demonstrated that AbSERPIN transcripts were ubiquitouslyexpressed, at different levels, in all healthy tissues tested (Fig. 3).Expression of AbSERPIN-1, AbSERPIN-2, and AbSERPIN-3 mRNA wasmost abundant in mantle, hemocytes, and muscle, respectively(P < 0.05). Moreover, the relative expression of AbSERPIN-2 in he-mocytes and of AbSERPIN-3 in muscle and mantle tissues wasextremely high, compared to other tissues (P < 0.05).

Fig. 1. (a) Molecular domain organization of AbSERPINs showing signal peptide (SP), low-complexity region (LCR), SERPIN domain, hinge region (HR), and SERPIN signature (SS) inreactive center loop (RCL). (b) Multiple sequence alignment of deduced amino acid sequences of RCL region of AbSERPINs and their SERPIN clade B orthologs. Completely conserved,semi-conserved, and weakly conserved residues are indicated by asterisk (*), semicolon (:), and period (.), respectively. The conserved HR and SS regions are boxed with broken andcontinuous lines, respectively. Predicted P1eP10 amino acid positions are shaded gray. (c) Predicted tertiary structures of the SERPIN domains of (i) AbSERPIN-1, (ii) AbSERPIN-2, and(iii) AbSERPIN-3. The conserved hinge regions and SERPIN signatures are colored pink and orange, respectively. The characteristic b-sheets A, B, and C are colored green, red, andyellow, respectively. Scissile bond residues (P1eP10) in the reactive center loop (RCL) are located at the top of each molecule. (For interpretation of the references to color in thisfigure legend, the reader is referred to the web version of this article.)
S.D.N.K. Bathige et al. / Fish & Shellfish Immunology 45 (2015) 334e341 337
3.2.2. Effect of live bacteria and PAMP on the mRNA expression ofAbSERPINs
To investigate the mRNA expression profiles of AbSERPINsfollowing in vivo challenges with pathogenic bacteria or LPS, weisolated hemocytes at different time points p.i. for qPCR analysis.As shown in Fig. 4, more or less similar mRNA expression
patterns for each AbSERPINs were observed in hemocytes afterV. parahaemolyticus, L. monocytogenes, and LPS challenges. mRNAexpression levels of AbSERPIN-1 (3.02-fold) and AbSERPIN-2 (4.5-fold) peaked at 12 h p.i. of V. parahaemolyticus and were subse-quently down-regulated until 72 h p.i. AbSERPIN-3 (4.4-fold)expression peaked at 3 h p.i, and was then significantly (P < 0.05)
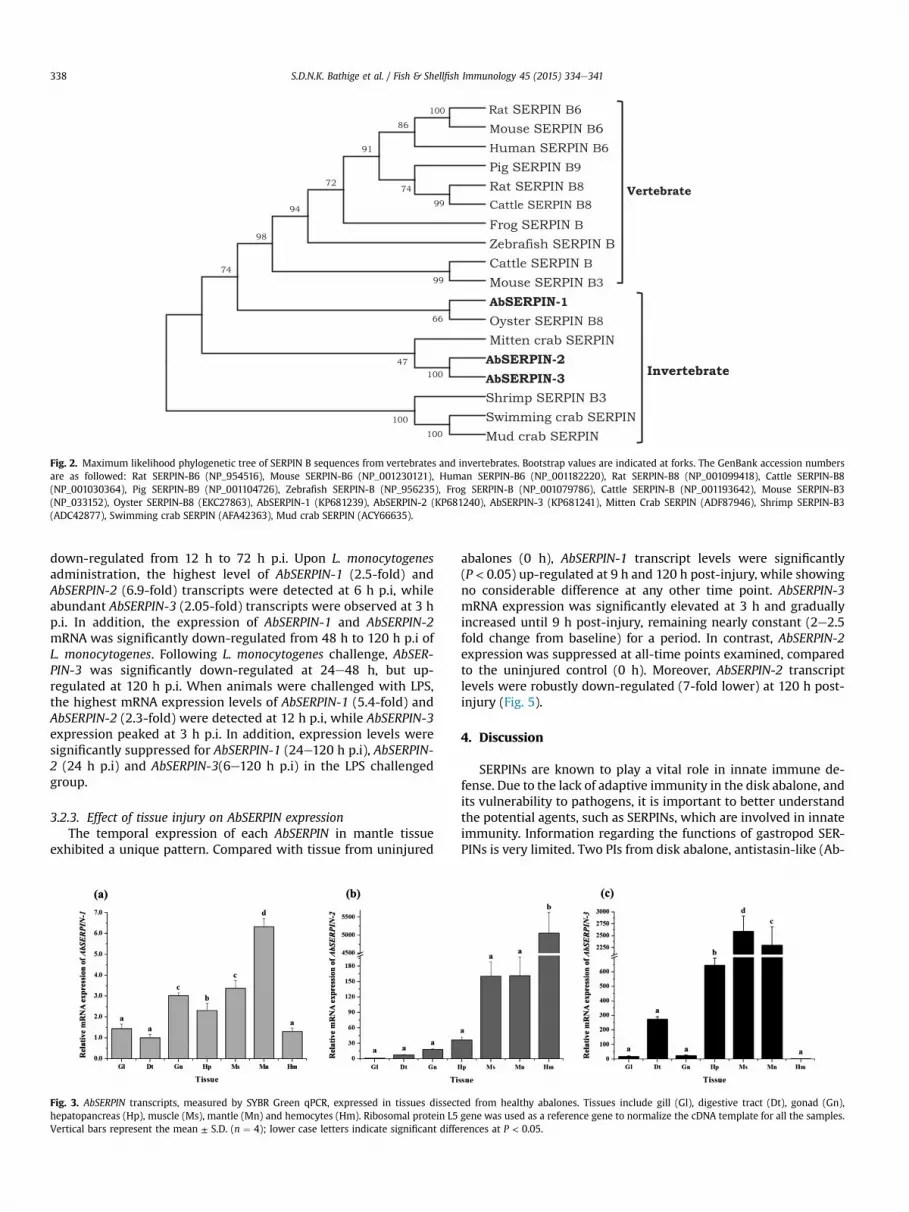
Fig. 2. Maximum likelihood phylogenetic tree of SERPIN B sequences from vertebrates and invertebrates. Bootstrap values are indicated at forks. The GenBank accession numbersare as followed: Rat SERPIN-B6 (NP_954516), Mouse SERPIN-B6 (NP_001230121), Human SERPIN-B6 (NP_001182220), Rat SERPIN-B8 (NP_001099418), Cattle SERPIN-B8(NP_001030364), Pig SERPIN-B9 (NP_001104726), Zebrafish SERPIN-B (NP_956235), Frog SERPIN-B (NP_001079786), Cattle SERPIN-B (NP_001193642), Mouse SERPIN-B3(NP_033152), Oyster SERPIN-B8 (EKC27863), AbSERPIN-1 (KP681239), AbSERPIN-2 (KP681240), AbSERPIN-3 (KP681241), Mitten Crab SERPIN (ADF87946), Shrimp SERPIN-B3(ADC42877), Swimming crab SERPIN (AFA42363), Mud crab SERPIN (ACY66635).
S.D.N.K. Bathige et al. / Fish & Shellfish Immunology 45 (2015) 334e341338
down-regulated from 12 h to 72 h p.i. Upon L. monocytogenesadministration, the highest level of AbSERPIN-1 (2.5-fold) andAbSERPIN-2 (6.9-fold) transcripts were detected at 6 h p.i, whileabundant AbSERPIN-3 (2.05-fold) transcripts were observed at 3 hp.i. In addition, the expression of AbSERPIN-1 and AbSERPIN-2mRNA was significantly down-regulated from 48 h to 120 h p.i ofL. monocytogenes. Following L. monocytogenes challenge, AbSER-PIN-3 was significantly down-regulated at 24e48 h, but up-regulated at 120 h p.i. When animals were challenged with LPS,the highest mRNA expression levels of AbSERPIN-1 (5.4-fold) andAbSERPIN-2 (2.3-fold) were detected at 12 h p.i, while AbSERPIN-3expression peaked at 3 h p.i. In addition, expression levels weresignificantly suppressed for AbSERPIN-1 (24e120 h p.i), AbSERPIN-2 (24 h p.i) and AbSERPIN-3(6e120 h p.i) in the LPS challengedgroup.
3.2.3. Effect of tissue injury on AbSERPIN expressionThe temporal expression of each AbSERPIN in mantle tissue
exhibited a unique pattern. Compared with tissue from uninjured
Fig. 3. AbSERPIN transcripts, measured by SYBR Green qPCR, expressed in tissues dissecthepatopancreas (Hp), muscle (Ms), mantle (Mn) and hemocytes (Hm). Ribosomal protein L5Vertical bars represent the mean ± S.D. (n ¼ 4); lower case letters indicate significant diffe
abalones (0 h), AbSERPIN-1 transcript levels were significantly(P < 0.05) up-regulated at 9 h and 120 h post-injury, while showingno considerable difference at any other time point. AbSERPIN-3mRNA expression was significantly elevated at 3 h and graduallyincreased until 9 h post-injury, remaining nearly constant (2e2.5fold change from baseline) for a period. In contrast, AbSERPIN-2expression was suppressed at all-time points examined, comparedto the uninjured control (0 h). Moreover, AbSERPIN-2 transcriptlevels were robustly down-regulated (7-fold lower) at 120 h post-injury (Fig. 5).
4. Discussion
SERPINs are known to play a vital role in innate immune de-fense. Due to the lack of adaptive immunity in the disk abalone, andits vulnerability to pathogens, it is important to better understandthe potential agents, such as SERPINs, which are involved in innateimmunity. Information regarding the functions of gastropod SER-PINs is very limited. Two PIs from disk abalone, antistasin-like (Ab-
ed from healthy abalones. Tissues include gill (Gl), digestive tract (Dt), gonad (Gn),gene was used as a reference gene to normalize the cDNA template for all the samples.rences at P < 0.05.

Fig. 4. Time course of AbSERPIN expression in hemocytes after bacterial- and LPS-injections for (a) AbSERPIN-1, (b) AbSERPIN-2, and (c) AbSERPIN-3 transcripts. Foldchanges in expression are relative to that of saline-injected control. Vertical barsrepresent the mean ± S.D. (n ¼ 4) and significant up-regulation (at P < 0.05) is indi-cated with asterisk.
Fig. 5. Time course of AbSERPIN expression in mantle in response to tissue injury.Vertical bars represent the mean ± S.D. (n ¼ 3) and significant difference (P < 0.05)from uninjured control is indicated with asterisk.
S.D.N.K. Bathige et al. / Fish & Shellfish Immunology 45 (2015) 334e341 339
Antistasin) [30] and Kazal-type inhibitor (Ab-KPI) [31], have beenpreviously identified in our laboratory. In the present study, weidentified and characterized three novel members of the SERPINfamily in disk abalone, revealing homology to known homologs of
SERPIN clade B. These findings were further confirmed by theMEROPS database. Typically, SERPINs are 350e400 amino acids inlength [5] and invertebrate SERPIN peptides are usually less than450 aa long [32e34]. AbSERPIN-1, discovered in this study, had atypical size (399 aa). However, AbSERPIN-2 and AbSERPIN-3 werecomparatively larger (>500 aa) and each contained an additionalpeptide region, an LCR, thought to facilitate proteineproteininteraction [35]. More importantly, AbSERPIN-2 and AbSERPIN-3each comprised a signal sequence at the N-terminal region, sug-gesting that these are secretory proteins. In contrast, previous re-ports have characterized most clade B SERPINs, including humanclade B SERPINs from B1 to B13, as intracellular proteins [6]. Thesefindings indicate that AbSERPIN-2 and AbSERPIN-3, as potentialsecretory proteins, may represent novel SERPIN proteins.
Alignment of RCL region sequences from various speciesrevealed highly conserved residues in the hinge region and theSERPIN signature region. The hinge region is functionally importantand provides the flexibility essential for changing the structuralconformation of the protein from S (stressed) to R (relaxed) [36]. Byalignment with other sequences, the P1eP10 site (scissile bond) wasrecognized within the RCL. The P1 residue that interacts with thetarget protease to cleave the scissile bond is crucial for specificity ofinhibition [8,37]. The predicted P1 residue in AbSERPIN-1 andAbSERPIN-2 was arginine, whilst glutamine was predicted forAbSERPIN-3. However, more extensive studies are required in orderto confirm the P1 site and characterize its inhibitory effect.
To study evolutionary relationships, a ML phylogenetic tree wasconstructed based on the available sequences of SERPIN orthologssharing high similarity with AbSERPINs. While AbSERPIN-1 clus-tered with a mollusk homolog, AbSERPIN-2 and AbSERPIN-3 clus-tered with a crustacean homolog. The NCBI BLAST analysis ofAbSERPINs revealed that most of the homologs those belong toSERPIN clade B were identified from crustaceans. Therefore, addi-tional studies are required to understand the proper evolutionarymechanism of AbSERPINs. In addition, we have modeled the ter-tiary structures of AbSERPINs by SWISS-MODEL server andrevealed the structural conservation to show the possible commoninhibitory activity.
Examination of tissue-specific AbSERPIN expression may revealtheir involvement in diverse biological functions. The current studyexamined the mRNA expression of each AbSERPIN. In previousstudies, disk abalone Ab-Antistasin [30] and Ab-KPI [31] PIs showed

S.D.N.K. Bathige et al. / Fish & Shellfish Immunology 45 (2015) 334e341340
the highest expression in the digestive tract and hepatopancreas,respectively. A recent study in octopus reported that the highestexpression level of a SERPIN occurred in hepatopancreas [38]. Inaddition, a number of studies characterizing crustacean SERPINsrevealed prominent expression in hemocytes [33,34,39] andhepatopancreas [32]. In contrast, Chinese mitten crab exhibited thehighest SERPIN expression in gonad and muscle tissues [40]. Thewidely distributed expression of SERPINs in disk abalone tissuesobserved in the current study provokes interest regarding theirroles in diverse biological functions.
SERPINs are known to play various essential roles in in-vertebrates. A number of studies have described possible roles inimmune responses in crustaceans, as well as in mollusks, againstbacterial infection. Recent studies in Eriocheir sinensis [40] andMarsupenaeus japonicus [32] revealed significantly up-regulatedSERPIN transcription at 24 h post-infection with V. anguillarum.Studies examining Penaeus monodon hemocytes found upregulatedmRNA expression of PmSERPIN8 and PmSERPINB3 at 24 and 48 hafter Vibrio harveyi challenge, respectively [34]. A molluskan SERPINfrom Octopus ocellatus demonstrated prominently elevated tran-script levels at 48 h after Listonella anguillarum infection and 24 hafter Micrcoccus luteus infection [38]. Previous studies of diskabalone PIs such as Ab-Antistasin [30] and Ab-KPI [31] alsodemonstrated elevated mRNA transcription at 6e12 h followingbacterial infection [28,29]. In the present study, we examinedtranscription levels of SERPINs in hemocytes, since they areresponsible for the recognition and removal of invading microbialpathogens in invertebrates [41]. Our results demonstrated thatbacterial infections (V. parahaemolyticus and L. monocytogenes) andLPS administration resulted in elevated mRNA expression levels ofAbSERPINs, at the early or middle time points following challenge.Gram-positive bacterial infection has been shown to activate theToll pathway via peptidoglycan recognition proteins (PGRPs),which induce the serine protease cascade in insects [42,43].Meanwhile, another study in Tenebrio molitor found that PGRP-SAcan recognize both Gram-positive and Gram-negative bacteriaand stimulates the serine protease cascade to cleave Spazle [44].We have previously found that disk abalone has functional PGRPsthat act as pattern recognition receptors [45], and that mighttrigger downstream immune signaling cascades. These signalingcascades are equipped with a diverse array of proteases, and inorder to control proteolysis, PIs should be in action. Accordingly,data from the current study suggest that AbSERPIN genes areregulated at the transcription level, and responded against Gram-positive and Gram-negative bacterial infections. Additionalstudies are warranted to determine the exact function of AbSER-PINs during bacterial infection.
To further understand the role of AbSERPINs in inflammatoryresponses, transcript levels were quantified over a time coursesubsequent to mantle tissue injury. Interestingly, different resultswere obtained for each AbSERPIN. Up-regulation of AbSERPIN-1 andAbSERPIN-3 was detected during the middle/late phase followingthe injury, whereas AbSERPIN-2 was down-regulated post-injury.Several studies have described the role of SERPINs in response totissue injury. The levels of proteases are primarily governed by theirrespective SERPINs, since uncontrolled protease activity may harmliving tissues. L. Zhu et al. (2006) found considerable up-regulationof A. irradians Kazal-type PI in the middle phase after adductormuscle injury [12]. Moreover, our previous report on Kazal-type PIfrom disk abalone also demonstrated significant elevation aftermantle tissue injury [31]. Previous studies in mammals have re-ported a possible role for SERPIN E2 and protease nexin-1 (PN-1) atthe site of tissue injury in the mouse vascular system [46]. A reportfound elevated levels of various SERPINs, including PN-1, plas-minogen activator inhibitor 1 (PAI-1), and kallikrein-binding
protein (KBP), in response tomuscle injury in rat [47]. However, fewreports described the down-regulation of SERPINs. One report findsthat migration of up-regulated proteases is influenced by down-regulation of the relevant SERPIN, such as hepatocyte nuclear fac-tor 4 alpha [48]. Another study in rock bream also reports thatexpression of PN-1 and heparin cofactor II are down-regulated inorder to maintain the proper balance between proteases and theircognate inhibitors [49]. These evidences imply that AbSERPINsmight be potentially involved in wound healing processes in diskabalone.
In conclusion, we identified and transcriptionally characterizedthree gastropod SERPIN clade B orthologs in disk abalone. Two ofthem (AbSERPIN-2 and AbSERPIN-3) appeared to be novel SERPINproteins due to the presence of N-terminal signal sequences char-acteristic of secretory proteins. Tertiary structures of the AbSERPINsindicated functional similarity with regard to interaction withpertinent substrates. Ubiquitous expression of AbSERPIN transcriptsunderlined their broad functional roles. Transcriptional up-regulation in the early phase following bacterial- and LPS-challenges suggested that AbSERPINs are potentially involved ininnate immune defense against invadingmicrobial pathogens. Overthe course of injury, the potent inflammatory responses of AbSER-PINs suggested their possible contribution to the wound healingprocess. Consequently, an increased understanding of the biologicalfunctions and host immune defense mechanisms of AbSERPINsmay provide new approaches for preventing bacterial infection infarmed disk abalone.
Acknowledgments
This research was supported by the Golden Seed Project, Min-istry of Agriculture, Food and Rural Affairs (MAFRA), the Ministry ofOceans and Fisheries (MOF), the Rural Development Administra-tion (RDA), and the Korea Forest Service (KFS).
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.fsi.2015.04.020.
References
[1] S. Iwanaga, B.L. Lee, Recent advances in the innate immunity of invertebrateanimals, J. Biochem. Mol. Biol. 38 (2005) 128e150.
[2] Y. He, H. Yu, Z. Bao, Q. Zhang, X. Guo, Mutation in promoter region of a serineprotease inhibitor confers Perkinsus marinus resistance in the eastern oyster(Crassostrea virginica), Fish Shellfish Immunol. 33 (2012) 411e417.
[3] N.D. Rawlings, D.P. Tolle, A.J. Barrett, Evolutionary families of peptidase in-hibitors, Biochem. J. 378 (2004) 705e716.
[4] N.D. Rawlings, A.J. Barrett, A. Bateman, MEROPS: the database of proteolyticenzymes, their substrates and inhibitors, Nucleic Acids Res. 40 (2012)D343eD350.
[5] J.A. Irving, R.N. Pike, A.M. Lesk, J.C. Whisstock, Phylogeny of the serpin su-perfamily: implications of patterns of amino acid conservation for structureand function, Genome Res. 10 (2000) 1845e1864.
[6] M.S. Mangan, D. Kaiserman, P.I. Bird, The role of serpins in vertebrate im-munity, Tissue Antigens. 72 (2008) 1e10.
[7] R.H. Law, Q. Zhang, S. McGowan, A.M. Buckle, G.A. Silverman, W. Wong, et al.,An overview of the serpin superfamily, Genome Biol. 7 (2006) 216.
[8] J.A. Huntington, Serpin structure, function and dysfunction, J. Thromb. Hae-most. 9 (Suppl. 1) (2011) 26e34.
[9] J.A. Irving, R.N. Pike, W. Dai, D. Bromme, D.M. Worrall, G.A. Silverman, et al.,Evidence that serpin architecture intrinsically supports papain-like cysteineprotease inhibition: engineering alpha(1)-antitrypsin to inhibit cathepsinproteases, Biochemistry 41 (2002) 4998e5004.
[10] R. Subramaniam, R.P. Dassanayake, J. Norimine, W.C. Brown, D.P. Knowles,S. Srikumaran, Molecular cloning, characterization and in vitro expression ofSERPIN B1 of bighorn sheep (Ovis canadensis) and domestic sheep (Ovisaries), and comparison with that of other species, Vet. Immunol. Immuno-pathol. 137 (2010) 327e331.
[11] G.A. Silverman, P.I. Bird, R.W. Carrell, F.C. Church, P.B. Coughlin, P.G. Gettins, etal., The serpins are an expanding superfamily of structurally similar but

S.D.N.K. Bathige et al. / Fish & Shellfish Immunology 45 (2015) 334e341 341
functionally diverse proteins. Evolution, mechanism of inhibition, novelfunctions, and a revised nomenclature, J. Biol. Chem. 276 (2001)33293e33296.
[12] L. Zhu, L. Song, Y. Chang, W. Xu, L. Wu, Molecular cloning, characterizationand expression of a novel serine proteinase inhibitor gene in bay scallops(Argopecten irradians, Lamarck 1819), Fish Shellfish Immunol. 20 (2006)320e331.
[13] Q. Xue, N. Itoh, K.L. Schey, R.K. Cooper, J.F. La Peyre, Evidence indicating theexistence of a novel family of serine protease inhibitors that may be involvedin marine invertebrate immunity, Fish Shellfish Immunol. 27 (2009) 250e259.
[14] J.F. La Peyre, Q.G. Xue, N. Itoh, Y. Li, R.K. Cooper, Serine protease inhibitor cvSI-1 potential role in the eastern oyster host defense against the protozoanparasite Perkinsus marinus, Dev. Comp. Immunol. 34 (2010) 84e92.
[15] M. Faisal, E.A. Maclntyre, K.G. Adham, B.D. Tail, M.H. Kothary, J.F. La Peyre,Evidence for the presence of protease inhibitors in eastern (Crassostrea vir-ginica) and pacific (Crassostrea gigas) oysters, Comp. Biochem. Physiol. Part BBiochem. Mol. Biol. 121 (1998) 161e168.
[16] B. Wang, J. Zhao, L. Song, H. Zhang, L. Wang, C. Li, et al., Molecular cloning andexpression of a novel Kazal-type serine proteinase inhibitor gene from Zhi-kong scallop Chlamys farreri, and the inhibitory activity of its recombinantdomain, Fish Shellfish Immunol. 24 (2008) 629e637.
[17] J. Cai, Z. Wang, C. Cai, Y. Zhou, Characterization and identification of virulentKlebsiella oxytoca isolated from abalone (Haliotis diversicolor supertexta)postlarvae with mass mortality in Fujian, China, J. Invertebr. Pathol. 97 (2008)70e75.
[18] P.C. Liu, Y.C. Chen, K.K. Lee, Pathogenicity of Vibrio alginolyticus isolated fromdiseased small abalone Haliotis diversicolor supertexta, Microbios 104 (2001)71e77.
[19] P.H. Chang, S.T. Kuo, S.H. Lai, H.S. Yang, Y.Y. Ting, C.L. Hsu, et al., Herpes-likevirus infection causing mortality of cultured abalone Haliotis diversicolorsupertexta in Taiwan, Dis. Aquat. Org. 65 (2005) 23e27.
[20] C.L. Goggin, R.J.G. Lester, Perkinsus, a protistan parasite of abalone inAustralia: a review, Mar. Freshwater Res. 46 (1995) 639e646.
[21] S.D. Bathige, N. Umasuthan, I. Whang, B.S. Lim, H.B. Jung, J. Lee, Evidences forthe involvement of an invertebrate goose-type lysozyme in disk abaloneimmunity: cloning, expression analysis and antimicrobial activity, FishShellfish Immunol. 35 (2013) 1369e1379.
[22] J.D. Thompson, D.G. Higgins, T.J. Gibson, CLUSTAL W: improving the sensi-tivity of progressive multiple sequence alignment through sequenceweighting, position-specific gap penalties and weight matrix choice, NucleicAcids Res. 22 (1994) 4673e4680.
[23] K. Tamura, D. Peterson, N. Peterson, G. Stecher, M. Nei, S. Kumar, MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolu-tionary distance, and maximum parsimony methods, Mol. Biol. Evol. 28(2011) 2731e2739.
[24] K. Arnold, L. Bordoli, J. Kopp, T. Schwede, The SWISS-MODEL workspace: aweb-based environment for protein structure homology modelling, Bioin-formatics 22 (2006) 195e201.
[25] W.L. DeLano, The PyMOL Molecular Graphics System, USA, San Carlos, CA,2002.
[26] M. De Zoysa, C. Nikapitiya, Y. Lee, S. Lee, C. Oh, I. Whang, et al., First molluscantranscription factor activator protein-1 (Ap-1) member from disk abalone andits expression profiling against immune challenge and tissue injury, FishShellfish Immunol. 29 (2010) 1028e1036.
[27] S.A. Bustin, V. Benes, J.A. Garson, J. Hellemans, J. Huggett, M. Kubista, et al., TheMIQE guidelines: minimum information for publication of quantitative real-time PCR experiments, Clin. Chem. 55 (2009) 611e622.
[28] Q. Wan, I. Whang, C.Y. Choi, J.S. Lee, J. Lee, Validation of housekeeping genes asinternal controls for studying biomarkers of endocrine-disrupting chemicalsin disk abalone by real-time PCR, Comp. Biochem. Physiol. Toxicol. Pharmacol.153 (2011) 259e268.
[29] K.J. Livak, T.D. Schmittgen, Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(-Delta Delta C(T)) Method, Methods 25(2001) 402e408.
[30] C. Nikapitiya, M. De Zoysa, C. Oh, Y. Lee, P.M. Ekanayake, I. Whang, et al., Diskabalone (Haliotis discus discus) expresses a novel antistasin-like serine pro-tease inhibitor: molecular cloning and immune response against bacterialinfection, Fish Shellfish Immunol. 28 (2010) 661e671.
[31] W.D. Wickramaarachchi, M. De Zoysa, I. Whang, Q. Wan, J. Lee, Kazal-typeproteinase inhibitor from disk abalone (Haliotis discus discus): molecularcharacterization and transcriptional response upon immune stimulation, FishShellfish Immunol. 35 (2013) 1039e1043.
[32] Y.R. Zhao, Y.H. Xu, H.S. Jiang, S. Xu, X.F. Zhao, J.X. Wang, Antibacterial activityof serine protease inhibitor 1 from kuruma shrimp Marsupenaeus japonicus,Dev. Comp. Immunol. 44 (2014) 261e269.
[33] S. Wang, Z. Cui, Y. Liu, Q. Li, C. Song, Identification and characterization of aserine protease inhibitor (PtSerpin) in the swimming crab Portunus tritu-berculatus, Fish Shellfish Immunol. 32 (2012) 544e550.
[34] S. Somnuk, A. Tassanakajon, V. Rimphanitchayakit, Gene expression andcharacterization of a serine proteinase inhibitor PmSERPIN8 from the blacktiger shrimp Penaeus monodon, Fish Shellfish Immunol. 33 (2012) 332e341.
[35] A. Coletta, J.W. Pinney, D.Y. Solis, J. Marsh, S.R. Pettifer, T.K. Attwood, Low-complexity regions within protein sequences have position-dependent roles,BMC Syst. Biol. 4 (2010) 43.
[36] J. Potempa, E. Korzus, J. Travis, The serpin superfamily of proteinase in-hibitors: structure, function, and regulation, J. Biol. Chem. 269 (1994)15957e15960.
[37] T. Homvises, A. Tassanakajon, K. Somboonwiwat, Penaeus monodon SERPIN,PmSERPIN6, is implicated in the shrimp innate immunity, Fish ShellfishImmunol. 29 (2010) 890e898.
[38] X. Wei, J. Xu, J. Yang, X. Liu, R. Zhang, W. Wang, et al., Involvement of a Serpinserine protease inhibitor (OoSerpin) from mollusc Octopus ocellatus in anti-bacterial response, Fish Shellfish Immunol. 42 (2015) 79e87.
[39] Y. Liu, F. Li, B. Wang, B. Dong, X. Zhang, J. Xiang, A serpin from Chinese shrimpFenneropenaeus chinensis is responsive to bacteria and WSSV challenge, FishShellfish Immunol. 26 (2009) 345e351.
[40] L. Wang, Z. Ma, J. Yang, Y. Gai, Z. Zhou, L. Wang, et al., Identification andcharacterization of a serine protease inhibitor Esserpin from the Chinesemitten crab Eriocheir sinensis, Fish Shellfish Immunol. 34 (2013) 1576e1586.
[41] E. Bachere, Y. Gueguen, M. Gonzalez, J. de Lorgeril, J. Garnier, B. Romestand,Insights into the anti-microbial defense of marine invertebrates: the penaeidshrimps and the oyster Crassostrea gigas, Immunol. Rev. 198 (2004) 149e168.
[42] N. Buchon, N.A. Broderick, B. Lemaitre, Gut homeostasis in a microbial world:insights from Drosophila melanogaster, Nat. Rev. Microbiol. 11 (2013)615e626.
[43] C.M. Cirimotich, Y. Dong, L.S. Garver, S. Sim, G. Dimopoulos, Mosquito immunedefenses against Plasmodium infection, Dev. Comp. Immunol. 34 (2010)387e395.
[44] Y. Yu, J.W. Park, H.M. Kwon, H.O. Hwang, I.H. Jang, A. Masuda, et al., Diversityof innate immune recognition mechanism for bacterial polymeric meso-diaminopimelic acid-type peptidoglycan in insects, J. Biol. Chem. 285 (2010)32937e32945.
[45] H.K.A. Premachandra, D.A.S. Elvitigala, I. Whang, J. Lee, Identification of anovel molluscan short-type peptidoglycan recognition protein in disk abalone(Haliotis discus discus) involved in host antibacterial defense, Fish ShellfishImmunol. 39 (2014) 99e107.
[46] S. Selbonne, F. Azibani, S. Iatmanen, Y. Boulaftali, B. Richard, M. Jandrot-Per-rus, et al., In vitro and in vivo antiangiogenic properties of the serpin proteasenexin-1, Mol. Cell Biol. 32 (2012) 1496e1505.
[47] B.W. Festoff, R.B. Reddy, M. VanBecelaere, I. Smirnova, J. Chao, Activation ofserpins and their cognate proteases in muscle after crush injury, J. Cell.Physiol. 159 (1994) 11e18.
[48] J.J. Castronuovo Jr., R.M. Price, The sequence of gene expression in culturedhuman saphenous vein after injury, J. Vasc. Surg. 35 (2002) 146e151.
[49] N. Umasuthan, I. Whang, J.O. Kim, M.J. Oh, S.J. Jung, C.Y. Choi, et al., Rockbream (Oplegnathus fasciatus) serpin, protease nexin-1: transcriptional anal-ysis and characterization of its antiprotease and anticoagulant activities, Dev.Comp. Immunol. 35 (2011) 785e798.