effects of reduced ph on the growth and survival of postlarvae of the donkey's ear abalone,...
TRANSCRIPT

Effects of reduced pH on the growth and survivalof postlarvae of the donkey’s ear abalone,Haliotis asinina (L.)
Abduraji S. Tahil • Danilo T. Dy
Received: 2 February 2014 / Accepted: 4 June 2014� Springer International Publishing Switzerland 2014
Abstract High atmospheric CO2 levels primarily from burning fossil fuels have
increased CO2 concentration in the surface water of the ocean, this in turn has resulted in a
drop in seawater pH known as ‘‘ocean acidification’’ (OA) and presents a potential threat to
calcifying marine organisms. Up to the present, however, the impacts of OA on the early
developmental stages of tropical abalone were not known. For the first time in the Phil-
ippines, we investigated the direct effects of reduced pH on the growth and survival of the
postlarvae of the donkey’s ear abalone, Haliotis asinina L. H. asinina is the only com-
mercial species of haliotid in the country. We reduced the ambient pH (7.99) of experi-
mental seawater by bubbling food-grade CO2 to obtain the desired pH levels as expected
by the year 2100. Survival of H. asinina postlarvae was negatively affected by reduced pH.
Mean survival of postlarvae significantly decreased from 86.3 % at ambient conditions to
47.2 and 18.3 % at pH 7.62 and 7.42, respectively, after 20 days (P \ 0.001 for both).
Survival at pH 7.81 (69.9 %) was comparable with ambient conditions (P [ 0.05). Mean
shell lengths of postlarvae also differed significantly among treatments and across exposure
period (P = 0.000 for both). Postlarvae reared at pH 7.62 and pH 7.42 exhibited smaller
shell size (138 and 124 lm, respectively) compared to those at ambient conditions
(175 lm). Postlarvae reared at pH 7.81 showed larger shell length (162 lm) compared to
the postlarvae reared at pH 7.42 (124 lm). Across time, significant increase in shell size
from day 5 to day 20 was observed only in postlarvae reared at ambient conditions
(P = 0.000) and in the pH 7.81 treatment (P = 0.006). Daily growth rate decreased sig-
nificantly from 2.83 lm day-1 (ambient) to 0.95 and 0.30 lm day-1 at pH 7.62 and 7.42,
respectively. Our results suggest that growth and survival of postlarval H. asinina was
found to be sensitive to reduced pH of seawater. This implies that future decrease in pH
unit of the world’s oceans will most likely threaten the natural population of this eco-
nomically important species.
A. S. Tahil � D. T. DyDepartment of Biology, University of San Carlos Talamban, Nasipit, Cebu City, Philippines
A. S. Tahil (&)USC Marine Research Station, 6015 Maribago, Lapu-Lapu City, Cebu, Philippinese-mail: [email protected]
123
Aquacult IntDOI 10.1007/s10499-014-9804-4

Keywords Abalone seed production � Daily growth rates �Mortality � Ocean acidification
Introduction
Concern over the negative effects of ocean acidification (OA) on marine ecosystem par-
ticularly on calcifying organisms is gaining more attention all over the world, especially in
the temperate region. Increasing concentration of CO2 in the world’s ocean due to high
anthropogenic CO2 emissions into the atmosphere could result in a decrease in the
availability of carbonate ions and a drop in seawater pH (Crim et al. 2011). Depending on
geographic region and season, the pH of the open sea (upper 100 m) varies between 7.9
and 8.3 (Andersen et al. 2013). This variation may be more pronounced in coastal waters
where relatively rapid changes in pH or pCO2 can occur (Kerrison et al. 2011). In the last
200 years, the oceans have already absorbed approximately 50 % of the atmospheric CO2,
which decreased surface ocean pH by an average of 0.1 units (Royal Society 2005; Orr
et al. 2005). This current drop in seawater pH units will continue to decline at a slower rate
to 0.3–0.5 units by 2100 (Caldeira and Wickett 2003). Recognizing the potential impact of
OA on the marine ecosystem, ecologically relevant CO2 perturbation experiments have
been conducted worldwide, revealing substantial adverse effects of OA on a wide range of
marine organisms (reviewed in Fabry et al. 2008).
Several studies have reported negative effects of OA on many early developmental
stages of calcifying invertebrates including fertilization, larval growth and survival,
shell calcification and dissolution, and larval duration and settlement (e.g., Albright
et al. 2012; Amaral et al. 2012; Andersen et al. 2013; Jansson et al. 2013; Kim et al.
2013; Kroeker et al. 2013; O’Donnell et al. 2013). It was shown that even minor
changes in pH can significantly impair larval development, survival and growth in
marine calcifying species (Dupont et al. 2008; Crim et al. 2011). This negative effect
may have severe consequences for commercially important species (i.e., abalone)
whose population hinges on larval recruitment, growth and survival of postlarvae in
their respective habitats. With all these observations, OA has now been recognized as
affecting key life history stages and transitions (including larval development, growth
and survival) of marine invertebrates. To date, however, the deleterious effect of OA
has not been investigated for tropical abalones or any high-value species of molluscs in
tropical areas.
Haliotis asinina is the only commercial species of abalone in the Philippines. In
1996, the country’s harvest of wild H. asinina was recorded at 448 metric tons with
China as the largest market for dried abalone meat, and the shells are also sold as
buttons, jewelry and inlay work (Fermin 2001). Juveniles of this species are produced
in the hatchery for grow-out culture and restocking in natural habitat. As an aquaculture
species, studying the postlarvae from the potential effects of reduced pH may help
enhance the sustainable seed production for the aquaculture industry. Here, we
investigated the effects of reduced seawater pH brought about by elevated CO2 con-
centration on the growth and survival of H. asinina postlarvae. The results presented
here provide a better understanding of the negative impact of low pH and the predicted
effects of OA on an ecologically and economically important gastropod species in a
tropical region, such as the Philippines.
Aquacult Int
123

Materials and methods
Experimental design
The experiment was conducted at the Tawi–Tawi Multi-Species Hatchery in Latu–Latu,
Bongao, Tawi–Tawi, Philippines, from August–November 2013 using a randomized
complete block design (RCBD). An epoxy-coated wooden tank with eight compartments
(4 9 2) and aeration system was used for the experiment. Each compartment has a
dimension of 60 9 60 9 60 cm. Each tank compartment was provided with UV-treated
and filtered (sand-filter and 5 lm mesh filter bag) seawater pumped from the subtidal area
of Tawi–Tawi Bay. The experiment was conducted in four successive trials with two
replicates for each treatment per trial, and a new group of one-month-old postlarvae was
used during each trial. Part of the water quality management was a daily siphoning of fecal
material and dirt from the bottom of each tank compartment. The volume of water taken
out was approximately 5 L and replaced with an equal volume of UV-treated and filtered
seawater via the flow-through system. A flow-through, non-recirculating system was done
daily for 8 h (2 min L-1) to allow exchange with new filtered (sand-filter and 5-lm mesh
filter bag) seawater in the experimental units.
Manipulation and maintenance of pH
Immediately after stocking of postlarvae, the pH levels were adjusted daily and maintained
through manual addition of a food-grade CO2 delivered via thin-walled silicon tubings.
Based on the preliminary simulation to determine the amount of CO2 needed to maintain
the pH within the range, CO2 was added every 2.5–3 h for an average duration of 7–50 s
depending on the desired pH level for each treatment. Based on the predicted decrease of
0.3–0.5 units in the ocean pH by 2100 (Caldeira and Wickett 2003, 2005), four pH levels
each ranging from 7.95 to 8.02 (mean ± SD; 7.99 ± 0.01—Ambient), 7.78–7.83
(7.81 ± 0.02—Treatment I), 7.59–7.65 (7.62 ± 0.02—Treatment II) and 7.39–7.45
(7.42 ± 0.02—Treatment III) were used in this experiment. The pH and temperature of the
water were measured with a calibrated pH meter.
Total alkalinity (TA) was measured every 5 days throughout the experiment using an
alkalinity titration kit (Precision: ± 0.05 lmol kg-1). Concentrations of CO2, carbonate
(CO3-2) and bicarbonate (HCO3
-); partial pressure of CO2 (pCO2); saturation state of
calcite (Xcalc) and aragonite (Xarag) were then calculated from measured TA and pH using
the software CO2SYS (Pierrot et al. 2006) and by using the dissociation constants of
carbonic acid from Mehrbach et al. (1973) refitted by Dickson and Millero (1987) (Moulin
et al. 2011). Salinity and dissolved oxygen was measured with a refractometer and DO
meter, respectively.
Preparation of diatom slurry and plate substrates
Before the main experiment, several hatching protocols were done to ensure adequate
supply of one-month-old abalone postlarvae. A pure stock culture of benthic diatoms
(Amphora sp. and Navicula ramosissima) was acquired and cultured at the larval food
laboratory of the Tawi–Tawi Multi-Species Hatchery. The diatoms were cultured from the
(starter) stock to a larger culture vessel of 500 L using the multi-step batch culture method
(De la Pena et al. 2010). Diatom species were harvested during their exponential growth
Aquacult Int
123

phase before transferring them to a larger culture vessel. Initially, 150 mL of diatom starter
was scaled up into 3 L of seawater in the laboratory using F/2 medium (Guillard and
Ryther 1962). After 3 days, this was scaled up into a 30-L culture using a white plastic
basin with TMRL (The Media Research Laboratory) medium (prepared from technical-
grade reagents composed of silicate, sodium phosphate, ferric chloride and sodium nitrate).
After another 3 days, the 30-L culture was inoculated into a 500-L concrete tank filled
with 300 L of seawater pumped from the subtidal area that passed through a sand filter and
a 5-lm mesh filter bag. The culture was vigorously aerated from the bottom of the tank and
enriched using the modified Commercial II medium (Renaud et al. 1991) composed of
technical- and agricultural-grade reagents (ammonium sulfate 21-0-0, 150.0 mg L-1; urea
46-0-0, 7.5 mg L-1; super phosphate 16-20-0, 25 mg L-1; FeCl3�6H2O, 5.0 mg L-1;
Na2EDTA, 5.0 mg L-1; and sodium silicate 15.0 mg L-1) (de la Pena et al. 2010). After
3 days of culture, additional seawater and a proportional amount of fertilizer were added to
fill-up the tank to full capacity (500 L).
A separate concrete tank was fitted with 100 pieces of 45 cm 9 40 cm corrugated
plastic sheet with crustose coralline algae (CCA) and arranged vertically on PVC pipes to
induce the growth of benthic diatoms on the plates. The plates with CCA were old plates
which have been placed in the sea for at least 3 months to allow the settlement of CCA
identified as Mesophyllum sp. on the top and Hydrolliton samoense at the bottom of the
pink crust (de la Pena et al. 2010). After 3–4 days in the tank, the corrugated plastic sheets
were examined as to the presence of diatom films growing on their surfaces. This hatchery
procedure was done to induce the abalone planktonic trochophores to settle onto the plates
with CCA and diatoms.
Stocking and rearing of abalone postlarvae
One-month-old postlarvae were obtained from the larval rearing facilities of the abalone
hatchery. Three replicate plates with CCA and diatom films were taken from the concrete
tank containing 100 plates and placed in each experimental unit during each trial as
substrate for the postlarvae. Each plate contained 20 postlarvae that have already settled
and developed for a month on the same plate. Plates with\20 postlarvae were augmented
by carefully detaching postlarvae from other plates using a soft paint brush or small plastic
spatula and placed on those plates containing \20 postlarvae. The postlarvae were reared
for 20 days in each of the four treatments. To sustain the food supply of the postlarvae
during the culture period, diatom slurry was added to each tank compartment every 3 days
for a period of 20 days.
To minimize counting error by avoiding repetitive counting of the same individuals, a
transparent plastic/acetate tape with grids of 2 cm2 was laid on the plate during counting of
surviving postlarvae.
Monitoring of growth and survival rates
The initial shell length (in lm) of the postlarvae was recorded at the time of stocking to the
experimental units using a micrometer scale inserted in the eyepiece of a monocular 409
microscope. Shell length was measured from the posterior end to the anterior end of the shell.
Subsequent measurement of shell length and monitoring of surviving postlarvae was done
every 5 days over a period of 20 days. For each sampling period, a total of 15 postlarvae were
randomly selected and dislodged from the three plates using a soft paint brush or small
spatula. All plates were numbered from which postlarval samples were collected using
Aquacult Int
123

random numbers. Increase in shell size of postlarvae in each treatment was calculated during
each sampling period, and the daily growth rate (in lm day-1) was expressed as the differ-
ence between the final and initial shell lengths divided by the number of days during which
measurements were taken. Postlarval survival for each treatment was determined by census of
individual postlarva during each sampling period and at the end of the 20-day rearing period
per trial. This was expressed in % and calculated from the accumulated mortality divided by
the initial number of postlarvae 9 100.
Statistical analysis
Results of the two-way ANOVA (factors pH and trials) on survival and shell size data did
not show significant differences among trials (survival: P = 0.254; shell size: P = 0.104 at
5 % level of significance). With these observations and since the postlarvae used for each
trial were obtained from the same group of broodstock and the conditions during the
experiment were similar, the data from the four trials were pooled for the statistical
analyses. Data on shell lengths were analyzed using repeated measures ANOVA to
determine the levels of differences in mean values among treatments and the effect of time
(day) on the growth and survival of postlarvae. Homoscedasticity of the shell length data
was done using Levene’s test. Differences between means were tested for significance
(P \ 0.05) using the post hoc Tukey HSD test. Differences in the daily growth rate and
survival of postlarvae at the end of the 20-day rearing period were determined using a
nonparametric Kruskal–Wallis ANOVA and multiple comparisons of mean ranks for all
treatment groups.
Results
Diffusion of CO2 into the experimental seawater resulted in reduced pH from ambient
conditions by 0.18, 0.37 and 0.56 units for treatment 1 (T-I), treatment 2 (T-II) and treatment
3 (T-III), respectively. The mean seawater pH was consistent within a desired level for each
treatment. At ambient conditions, the mean pH for the whole duration of the experiment was
7.99 ± 0.01. In CO2-treated waters, the mean pH levels were 7.81 ± 0.02 (T-I),
7.62 ± 0.02 (T-II) and 7.42 ± 0.02 (T-III) (Table 1). For all cases, the mean temperature
and salinity showed minimal variations across treatments which ranged from 26.8 ± 0.6 to
27.3 ± 0.5 �C and 34.1 ± 0.3 to 34.2 ± 0.4 psu, respectively (Table 1). The same variation
was recorded for the TA which ranged from 2,092.2 to 2,194.2 lmol kg-1 across treatments.
Minimal variations in O2 concentration were also observed among treatments (7.9 ± 0.6 to
8.2 ± 0.3 mg L-1), suggesting that the amount of CO2 addition did not significantly affect
the oxygen concentration of the seawater.
The carbonate chemistry of the experimental seawater is also shown in Table 1. Car-
bonate ion concentration was lowest in waters with high CO2 or lowest pH
(63.7 ± 2.4 lmol kg-1) and highest at ambient conditions (210.9 lmol kg-1) (Table 1).
The partial pressure of CO2 (pCO2) in the experimental seawater increased with increasing
CO2 concentration and was highest (pCO2 = 1,886.4 ± 73.6 latm) in T-III
(CO2 = 51.3 ± 1.8 lmol kg-1) compared to ambient conditions (CO2 = 11.8 ± 0.4
lmol kg-1) with a pCO2 of 440.6 ± 19.1 latm. Saturation states of aragonite (Xarag) and
calcite (Xcalc) decreased with increasing CO2 concentration or reduced pH level. In T-III
(pH = 7.42; CO2 = 51.3 lmol kg-1), Xarag and Xcalc of the experimental seawater were
only 1.0 ± 0.0 and 1.5 ± 0.1, respectively, compared to the ambient conditions
Aquacult Int
123
(Xarag = 3.4 ± 0.1 and Xcalc = 5.1 ± 0.1) (Table 1). Calculations of these parameters
were based on the measured pH and TA as with other similar studies (i.e., Cunningham
2013; Jansson et al. 2013; Timmins-Schiffman et al. 2012).
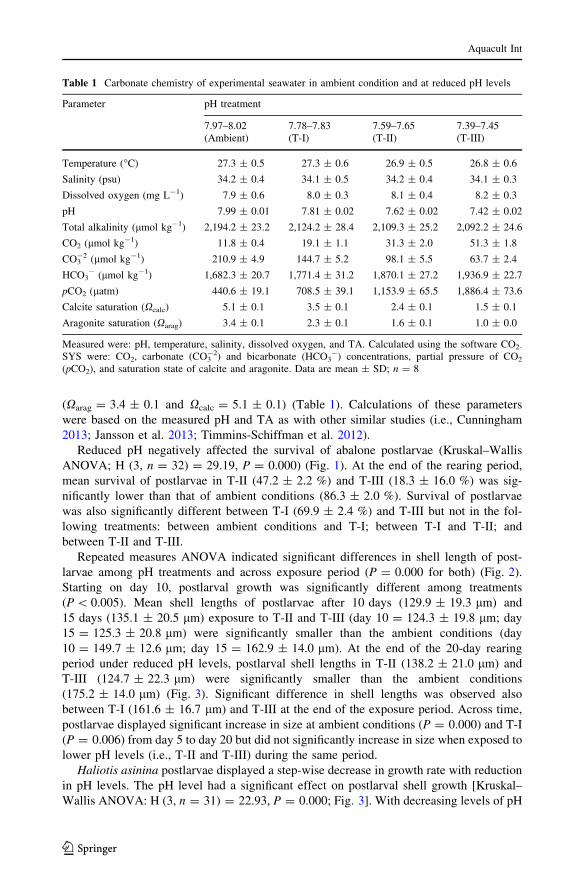
Reduced pH negatively affected the survival of abalone postlarvae (Kruskal–Wallis
ANOVA; H (3, n = 32) = 29.19, P = 0.000) (Fig. 1). At the end of the rearing period,
mean survival of postlarvae in T-II (47.2 ± 2.2 %) and T-III (18.3 ± 16.0 %) was sig-
nificantly lower than that of ambient conditions (86.3 ± 2.0 %). Survival of postlarvae
was also significantly different between T-I (69.9 ± 2.4 %) and T-III but not in the fol-
lowing treatments: between ambient conditions and T-I; between T-I and T-II; and
between T-II and T-III.
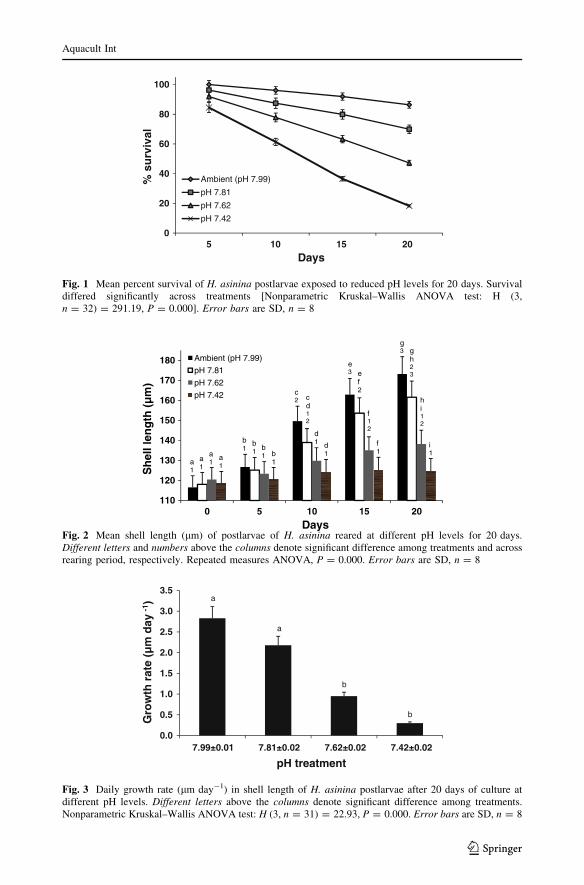
Repeated measures ANOVA indicated significant differences in shell length of post-
larvae among pH treatments and across exposure period (P = 0.000 for both) (Fig. 2).
Starting on day 10, postlarval growth was significantly different among treatments
(P \ 0.005). Mean shell lengths of postlarvae after 10 days (129.9 ± 19.3 lm) and
15 days (135.1 ± 20.5 lm) exposure to T-II and T-III (day 10 = 124.3 ± 19.8 lm; day
15 = 125.3 ± 20.8 lm) were significantly smaller than the ambient conditions (day
10 = 149.7 ± 12.6 lm; day 15 = 162.9 ± 14.0 lm). At the end of the 20-day rearing
period under reduced pH levels, postlarval shell lengths in T-II (138.2 ± 21.0 lm) and
T-III (124.7 ± 22.3 lm) were significantly smaller than the ambient conditions
(175.2 ± 14.0 lm) (Fig. 3). Significant difference in shell lengths was observed also
between T-I (161.6 ± 16.7 lm) and T-III at the end of the exposure period. Across time,
postlarvae displayed significant increase in size at ambient conditions (P = 0.000) and T-I
(P = 0.006) from day 5 to day 20 but did not significantly increase in size when exposed to
lower pH levels (i.e., T-II and T-III) during the same period.
Haliotis asinina postlarvae displayed a step-wise decrease in growth rate with reduction
in pH levels. The pH level had a significant effect on postlarval shell growth [Kruskal–
Wallis ANOVA: H (3, n = 31) = 22.93, P = 0.000; Fig. 3]. With decreasing levels of pH
Table 1 Carbonate chemistry of experimental seawater in ambient condition and at reduced pH levels
Parameter pH treatment
7.97–8.02(Ambient)
7.78–7.83(T-I)
7.59–7.65(T-II)
7.39–7.45(T-III)
Temperature (�C) 27.3 ± 0.5 27.3 ± 0.6 26.9 ± 0.5 26.8 ± 0.6
Salinity (psu) 34.2 ± 0.4 34.1 ± 0.5 34.2 ± 0.4 34.1 ± 0.3
Dissolved oxygen (mg L-1) 7.9 ± 0.6 8.0 ± 0.3 8.1 ± 0.4 8.2 ± 0.3
pH 7.99 ± 0.01 7.81 ± 0.02 7.62 ± 0.02 7.42 ± 0.02
Total alkalinity (lmol kg-1) 2,194.2 ± 23.2 2,124.2 ± 28.4 2,109.3 ± 25.2 2,092.2 ± 24.6
CO2 (lmol kg-1) 11.8 ± 0.4 19.1 ± 1.1 31.3 ± 2.0 51.3 ± 1.8
CO3-2 (lmol kg-1) 210.9 ± 4.9 144.7 ± 5.2 98.1 ± 5.5 63.7 ± 2.4
HCO3- (lmol kg-1) 1,682.3 ± 20.7 1,771.4 ± 31.2 1,870.1 ± 27.2 1,936.9 ± 22.7
pCO2 (latm) 440.6 ± 19.1 708.5 ± 39.1 1,153.9 ± 65.5 1,886.4 ± 73.6
Calcite saturation (Xcalc) 5.1 ± 0.1 3.5 ± 0.1 2.4 ± 0.1 1.5 ± 0.1
Aragonite saturation (Xarag) 3.4 ± 0.1 2.3 ± 0.1 1.6 ± 0.1 1.0 ± 0.0
Measured were: pH, temperature, salinity, dissolved oxygen, and TA. Calculated using the software CO2-
SYS were: CO2, carbonate (CO3-2) and bicarbonate (HCO3
-) concentrations, partial pressure of CO2
(pCO2), and saturation state of calcite and aragonite. Data are mean ± SD; n = 8
Aquacult Int
123
0
20
40
60
80
100
5 10 15 20
Ambient (pH 7.99)
pH 7.81
pH 7.62
pH 7.42
Days
% s
urv
ival
Fig. 1 Mean percent survival of H. asinina postlarvae exposed to reduced pH levels for 20 days. Survivaldiffered significantly across treatments [Nonparametric Kruskal–Wallis ANOVA test: H (3,n = 32) = 291.19, P = 0.000]. Error bars are SD, n = 8
110
120
130
140
150
160
170
180
0 5 10 15 20
Ambient (pH 7.99)
pH 7.81
pH 7.62
pH 7.42
Sh
ell l
eng
th (
µm
)
Days
a1
a1
b1 b
1
d1
f 12
d1
f1
a1
a1
b1
b1
c2
e3
i1
ef2
cd12
gh 2 3
hi 1 2
g3
Fig. 2 Mean shell length (lm) of postlarvae of H. asinina reared at different pH levels for 20 days.Different letters and numbers above the columns denote significant difference among treatments and acrossrearing period, respectively. Repeated measures ANOVA, P = 0.000. Error bars are SD, n = 8
a
a
b
b
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
7.99±0.01 7.81±0.02 7.62±0.02 7.42±0.02
Gro
wth
rat
e (µ
m d
ay -1
)
pH treatment
Fig. 3 Daily growth rate (lm day-1) in shell length of H. asinina postlarvae after 20 days of culture atdifferent pH levels. Different letters above the columns denote significant difference among treatments.Nonparametric Kruskal–Wallis ANOVA test: H (3, n = 31) = 22.93, P = 0.000. Error bars are SD, n = 8
Aquacult Int
123

from 7.99 (ambient), 7.81, 7.62 to 7.42, daily growth rates of the postlarvae decreased from
2.83 ± 0.26, 2.18 ± 0.47, 0.95 ± 0.81, to 0.30 ± 0.42 lm day-1 at the end of the 20-day
exposure period (Fig. 3).
Discussion
The reduction in pH units (0.18–0.56) as a result of CO2 infusion into seawater was within
the ocean surface water pH reduction (0.3–0.5 units) expected by 2100 (Caldeira and
Wickett 2003). However, reducing the ambient pH by 0.56 units is relatively lower than
that resulting from the expected worst-case scenario (-0.5 units). We used this lowest pH
to forecast how future worst scenario may affect the postlarvae of this abalone species
should the emission of anthropogenic CO2 into the atmosphere continue with its current
trend of ‘‘business-as-usual’’ in the coming century. Here, we provide the first direct
experimental evidence that the expected pH reduction of the ocean’s surface waters will
negatively impact the tropical abalone species (H. asinina). We recorded an ambient pH
ranging from 7.97 to 8.02 (mean = 7.99 ± 0.01) for seawater pumped from the subtidal
area (2.5–4.5 m depth) of Tawi–Tawi Bay, which is partly surrounded by mangrove forest
and where pH levels are expected to be relatively low and variable compared to the open
ocean. The calculated pCO2 (441 latm) for ambient conditions was closely similar to the
reports of other workers studying molluscs from the Pacific area [e.g., 468 latm (Timmins-
Schiffman et al. 2012); 469 latm (Andersen et al. 2013)]. H. asinina is most common in
the shallow reef flat (ca. 0.5–4.5 m depth) around the bay where they may be exposed to
such variations in pH levels in nature. Thus, the use of reduced pH levels here revealed
how these variations in nature may affect the post-settlement stage and population integrity
of this abalone species. We calculated also low saturation states of calcium carbonate (1.5)
and aragonite (1.0) at the lowest pH (7.42) treatment, suggesting that shell dissolution of
postlarvae may be rapid at reduced pH levels. For shell-forming molluscs (i.e., abalone)
living in such conditions, high organism energy expenditure is expected to maintain a
normal shell structure and obtain better survival and growth (Jansson et al. 2013).
Survival
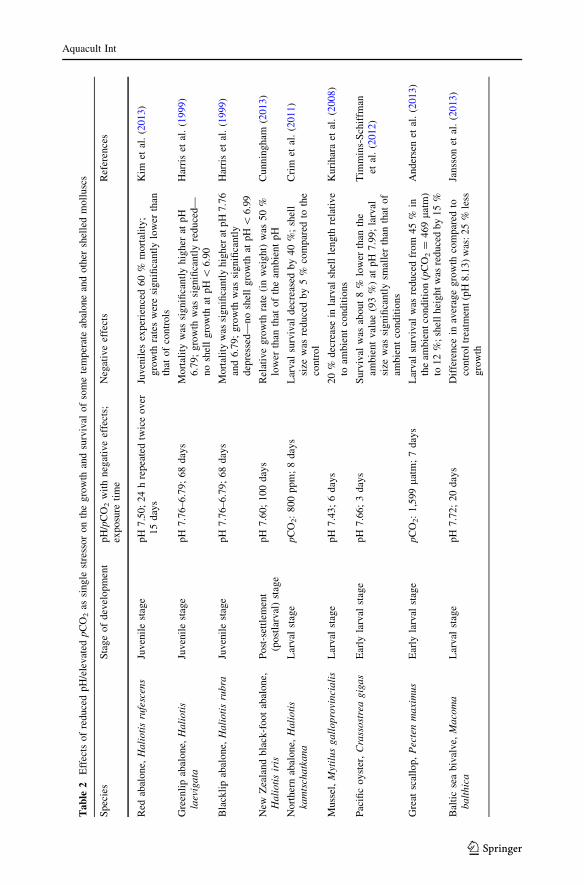
In the context of tropical conditions, our results on larval survival of H. asinina exposed to
reduced pH for 20 days indicated that OA has highly negative impact on the postlarval
stage of this tropical abalone. The observed reduction in % survival of postlarvae from
86.3 % at ambient conditions to 47.2 % at pH 7.62 and 18.3 % at pH 7.42 is a clear
evidence of such negative effects. These observations corroborated other studies for some
temperate abalone and shelled molluscs (Table 2). In the southern California red abalone
H. rufescens, only 40 % of the juveniles survived after 15 days of exposure to pH 7.50
(Kim et al. 2013). Significantly, higher mortalities were also reported for two species of
abalone from Australia after 68 days at pH 6.79 (for H. laevigata) and pH 7.76 and 6.79
(for H. rubra) (Harris et al. 1999). Considering that the pH of the open sea (upper 100 m)
varies between 7.9 and 8.3 (Andersen et al. 2013), the use of pH 6.79 (ca. -1.3 units
relative to the ambient value) for these two species is very much lower than that resulting
from the worst-case scenario (pH decrease of 0.5 units) expected by the end of the present
century. Such pH reduction (-1.3 units) can be considered highly hypothetical and most
likely far from reality even after the next century. In a short-term experiment conducted by
Crim et al. (2011), the survival of the northern abalone (H. kamtschatkana) larvae was
Aquacult Int
123
Tab
le2
Eff
ects
of
redu
ced
pH
/ele
vat
edpC
O2
assi
ng
lest
ress
or
on
the
gro
wth
and
surv
ival
of
som
ete
mp
erat
eab
alo
ne
and
oth
ersh
elle
dm
oll
usc
s
Sp
ecie
sS
tag
eo
fd
evel
op
men
tp
H/p
CO
2w
ith
neg
ativ
eef
fect
s;ex
po
sure
tim
eN
egat
ive
effe
cts
Ref
eren
ces
Red
abal
on
e,H
ali
oti
sru
fesc
ens
Juven
ile
stag
epH
7.5
0;
24
hre
pea
ted
twic
eover
15
day
sJu
ven
iles
exp
erie
nce
d6
0%
mort
alit
y;
gro
wth
rate
sw
ere
sign
ifica
ntl
ylo
wer
than
that
of
con
trols
Kim
etal
.(2
01
3)
Gre
enli
pab
alo
ne,
Ha
lio
tis
laev
iga
taJu
ven
ile
stag
ep
H7
.76
–6
.79
;6
8d
ays
Mo
rtal
ity
was
sig
nifi
can
tly
hig
her
atp
H6
.79;
gro
wth
was
sig
nifi
can
tly
red
uce
d—
no
shel
lg
row
that
pH
\6
.90
Har
ris
etal
.(1
99
9)
Bla
ckli
pab
alone,
Ha
lio
tis
rub
raJu
ven
ile
stag
ep
H7
.76
–6
.79
;6
8d
ays
Mo
rtal
ity
was
sig
nifi
can
tly
hig
her
atp
H7
.76
and
6.7
9;
gro
wth
was
sign
ifica
ntl
yd
epre
ssed
—n
osh
ell
gro
wth
atp
H\
6.9
9
Har
ris
etal
.(1
99
9)
New
Zea
lan
db
lack
-foo
tab
alo
ne,
Ha
lio
tis
iris
Post
-set
tlem
ent
(po
stla
rval
)st
age
pH
7.6
0;
10
0d
ays
Rel
ativ
eg
row
thra
te(i
nw
eig
ht)
was
50
%lo
wer
than
that
of
the
amb
ien
tp
HC
un
nin
gh
am(2
01
3)
Nort
her
nab
alone,
Ha
lio
tis
kam
tsch
atk
ana
Lar
val
stag
epC
O2:
80
0p
pm
;8
day
sL
arv
alsu
rviv
ald
ecre
ased
by
40
%;
shel
lsi
zew
asre
du
ced
by
5%
com
par
edto
the
con
tro
l
Cri
met
al.
(20
11)
Mu
ssel
,M
ytil
us
ga
llo
pro
vin
cia
lis
Lar
val
stag
ep
H7
.43
;6
day
s2
0%
dec
reas
ein
larv
alsh
ell
len
gth
rela
tiv
eto
amb
ien
tco
nd
itio
ns
Ku
rihar
aet
al.
(20
08)
Pac
ific
oy
ster
,C
rass
ost
rea
gig
as
Ear
lyla
rval
stag
ep
H7
.66
;3
day
sS
urv
ival
was
abo
ut
8%
low
erth
anth
eam
bie
nt
val
ue
(93
%)
atp
H7
.99
;la
rval
size
was
sig
nifi
can
tly
smal
ler
than
that
of
ambie
nt
con
dit
ion
s
Tim
min
s-S
chif
fman
etal
.(2
01
2)
Gre
atsc
allo
p,
Pec
ten
ma
xim
us
Ear
lyla
rval
stag
epC
O2:
1,5
99
latm
;7
day
sL
arv
alsu
rviv
alw
asre
du
ced
fro
m4
5%
inth
eam
bie
nt
con
dit
ion
(pC
O2
=4
69
latm
)to
12
%;
shel
lh
eig
ht
was
redu
ced
by
15
%
An
der
sen
etal
.(2
01
3)
Bal
tic
sea
biv
alv
e,M
aco
ma
ba
lth
ica
Lar
val
stag
epH
7.7
2;
20
day
sD
iffe
rence
inav
erag
egro
wth
com
par
edto
con
tro
ltr
eatm
ent
(pH
8.1
3)
was
:2
5%
less
gro
wth
Jan
sson
etal
.(2
01
3)
Aquacult Int
123

significantly reduced by 40 % after 8 days of exposure to elevated pCO2 of 800 ppm
(Table 2).
A correlation between larval survival and pH levels has also been reported for other
shell-forming molluscs. In the Pacific oyster Crassostrea gigas, a small reduction in larval
survival by 7.3 % was observed after 3 days of exposure to pH 7.66 (Timmins-Schiffman
et al. 2012). In our experiment, survival of H. asinina was reduced by 39 % when the
postlarvae were exposed to a similar pH level (7.62) relative to the ambient conditions (pH
7.99). Andersen et al. (2013) reported a significant reduction in larval survival of the Great
scallop (Pecten maximus) from 45 % to ca. 20 and 12 % on day 7 post fertilization when
the ambient pH (7.98) was reduced to pH 7.78 and pH 7.51, respectively (Table 2). For
H. asinina, we obtained 47.2 % postlarval survival at pH 7.62 and 18.3 % at pH 7.42 after
20 days of exposure. These variations in survival rates among shelled molluscs indicate
inter-species differences at certain life stages and exposure period, and some species may
be robust to the perturbations related to seawater chemistry changes.
Growth
Our results show that decreases in pH due to increased CO2 concentration negatively
affected the growth of H. asinina postlarvae. The shell growth of the postlarvae was
reduced by 28 % at pH 7.62 and 20 % at pH 7.42 relative to ambient conditions. In the
New Zealand black-foot abalone (H. iris), the relative growth of the postlarvae was 50 %
lower than that of ambient conditions after 100 days of exposure to pH 7.60 (Cunningham
2013) (Table 2). In the Green-lip abalone (H. laevigata) and Black-lip abalone (H. rubra)
from Australia, juvenile growth rates of both species were significantly depressed at lower
pH levels, and no shell growth was observed after 68 days of exposure to a very low pH of
\6.99 (Harris et al. 1999). In the red abalone (H. rufescens), juvenile shell growth was
significantly reduced when exposed to low pH (Kim et al. 2013). With these negative
effects on growth and in the context of natural scenario, a reduction in seawater pH can be
considered as an important factor influencing the maintenance of abalone populations. This
is so because reproductive success and susceptibility to predation of a species is influenced
by its growth (Rossetto et al. 2012).
Reduced shell growth under reduced pH conditions was observed also in some species
of shell-forming molluscs. The average shell growth of the Baltic Sea bivalve, Macoma
balthica, larvae was 25 % lower than that of the control treatment after 20 days at reduced
pH of 7.72 (Jansson et al. 2013). A mussel species, Mytilus galloprovincialis, displayed
20 % decrease in shell length of larvae when the ambient pH (8.21) was reduced to pH
7.43 over a period of 6 days (Kurihara et al. 2008). In the Pacific oyster C. gigas, the shell
length of larvae became smaller by 20 % after the 2-day exposure to pH 7.4 (Kurihara et al.
2007). These observations strongly corroborate our results under closely similar pH levels
and CO2 concentrations (Table 1; Fig. 2).
Slow growth of H. asinina postlarvae at reduced pH may be attributed to the effect of
pH stressor on the development of larval shell structure. Shell deformity and dissolution
was observed in postlarvae exposed to pH 7.42 during the experiment (Fig. 4). This
condition was observed also in H. rufescens larvae that showed decreased shell length after
exposure to the low pH treatment, suggesting that low pH may have caused the dissolution
of larval shells (Kim et al. 2013). The decrease in growth rates as manifested in the
abnormal shell development of larvae due to reduced pH had been observed also in the
great scallop, P. maximus, larvae (Andersen et al. 2013). Normally shaped shells decreased
from 64 % at ambient pH (7.98) to 28 % when the larvae were exposed to pH 7.51 for
Aquacult Int
123
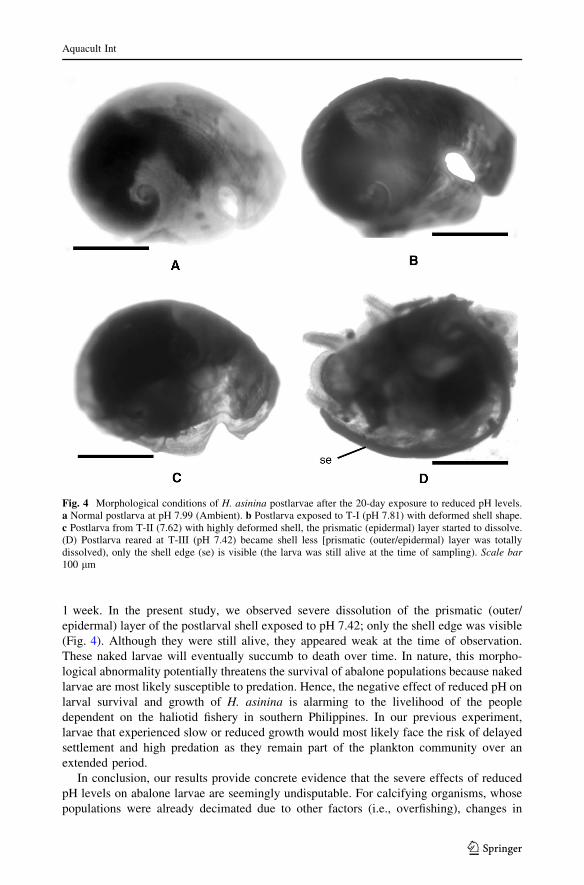
1 week. In the present study, we observed severe dissolution of the prismatic (outer/
epidermal) layer of the postlarval shell exposed to pH 7.42; only the shell edge was visible
(Fig. 4). Although they were still alive, they appeared weak at the time of observation.
These naked larvae will eventually succumb to death over time. In nature, this morpho-
logical abnormality potentially threatens the survival of abalone populations because naked
larvae are most likely susceptible to predation. Hence, the negative effect of reduced pH on
larval survival and growth of H. asinina is alarming to the livelihood of the people
dependent on the haliotid fishery in southern Philippines. In our previous experiment,
larvae that experienced slow or reduced growth would most likely face the risk of delayed
settlement and high predation as they remain part of the plankton community over an
extended period.
In conclusion, our results provide concrete evidence that the severe effects of reduced
pH levels on abalone larvae are seemingly undisputable. For calcifying organisms, whose
populations were already decimated due to other factors (i.e., overfishing), changes in
Fig. 4 Morphological conditions of H. asinina postlarvae after the 20-day exposure to reduced pH levels.a Normal postlarva at pH 7.99 (Ambient). b Postlarva exposed to T-I (pH 7.81) with deformed shell shape.c Postlarva from T-II (7.62) with highly deformed shell, the prismatic (epidermal) layer started to dissolve.(D) Postlarva reared at T-III (pH 7.42) became shell less [prismatic (outer/epidermal) layer was totallydissolved), only the shell edge (se) is visible (the larva was still alive at the time of sampling). Scale bar100 lm
Aquacult Int
123

ocean carbonate chemistry and in particular reduction in surface water pH may pose severe
impact on their existence. The reduced pH-mediated decline in larval survival and growth
that we observed in our study suggested that highly commercial and hence heavily
exploited species, such as abalone, may succumb to higher rates of natural mortality. More
investigations between larval growth and survival of abalones and seawater pH or CO2
concentration should be done across the region as there are possibly other haliotid species
which could possibly have higher tolerance to environmental stressors and could serve as
alternate candidate for aquaculture. Together with fishing mortality, it may further threaten
the existence of the natural populations. Since OA is seemingly irreversible and may
continue to pose detrimental conditions, long-term studies across biogeographical regions
are needed to elucidate the negative effects of OA on the early life history of both tem-
perate and tropical species of abalone. Such studies should involve investigation on the
additive effects of pH together with other environmental parameters (i.e., temperature,
salinity, dissolved oxygen) associated with OA.
Acknowledgments The authors thank the two anonymous reviewers for their critical review and helpfulsuggestions. We also thank the management of the Tawi–Tawi Multi-Species Hatchery, Latu–Latu, Bongao,Tawi–Tawi, Philippines, for allowing the authors to conduct the experiment using their facilities andmanpower. Due recognition and gratitude are afforded to Ms. Arlyn C. Carroz, Hatchery Manager; Ms.Juliet F. Francisco, Senior Hatchery Technician; and our Research Assistants, Mr. Japrin L. Hadji-Amin andMr. Abdurasul S. Sarail for their support and assistance during the study. This paper is a joint marine sciencecontribution of the University of San Carlos and the Mindanao State University Tawi–Tawi, Philippines.
References
Albright R, Bland C, Gillette P, Serafy JE, Langdon C, Capo TR (2012) Juvenile growth of the tropical seaurchin Lytechinus variegatus exposed to near-future ocean acidification scenarios. J Exp Mar Biol Ecol426–427:12–17
Amaral V, Cabral HN, Bishop MJ (2012) Moderate acidification affects growth but not survival of 6-month-old oysters. Aquat Ecol 46:119–127
Andersen S, Grefsrud ES, Harboe T (2013) Effect of increased pCO2 level on early shell development ingreat scallop (Pecten maximus Lamarck) larvae. Biogeoscience 10:6161–6184
Caldeira K, Wickett ME (2003) Anthropogenic carbon and ocean pH. Nature 425:365Caldeira K, Wickett ME (2005) Ocean model predictions of chemistry changes from carbon dioxide
emissions to the atmosphere and ocean. J Geophys Res 110:C09S04Crim RN, Sunday JM, Harley CDG (2011) Elevated seawater CO2 concentrations impair larval development
and reduce larval survival in endangered northern abalone (Haliotis kamtschatkana). J Exp Mar BiolEcol 400:272–277
Cunningham SC (2013) The effects of ocean acidification on juvenile Haliotis iris. Thesis, Master ofScience. University of Otago, New Zealand
De La Pena MR, Bautista JI, Buen-Ursua SM, Bayona N, Titular VST (2010) Settlement, growth andsurvival of the donkey’s ear abalone Haliotis asinina (Linne) in response to diatom diets andattachment substrate. Philos J Sci 139:27–34
Dickson AG, Millero FJ (1987) A comparison of the equilibrium constants for the dissociation of carbonicacid in seawater media. Deep Sea Res A 34:1733–1743
Dupont S, Havenhand J, Thorndyke W, Peck L, Thorndyke M (2008) Near-future level of CO2-driven oceanacidification radically affects larval survival and development in the brittlestar Ophiothrix fragilis. MarEcol Prog Ser 373:285–294
Fabry VJ, Seibel BA, Feely RA, Orr JC (2008) Impacts of ocean acidification on marine fauna andecosystem processes. ICES J Mar Sci 65:414–432
Fermin AC (2001) Tropical abalone culture in the Philippines. The Advocate 3:68–69Guillard RRL, Ryther JH (1962) Studies on the marine planktonic diatoms. I. Cyclotella nana Hustedt, and
Detonela confervacea (Cleve) gran. Can J Microbiol 8:229–239Harris JO, Maguire GB, Edwards SJ, Hindrum SM (1999) Effect of pH on growth rate, oxygen consumption
rate, and histopathology of gill and kidney tissue for juvenile greenlip abalone, Haliotis laevigataDonovan and blacklip abalone, Haliotis rubra Leach. J Shellfish Res 18:611–619
Aquacult Int
123

Jansson A, Norkko J, Norkko A (2013) Effects on reduced pH on Macoma balthica larvae from a systemwith naturally fluctuating pH-dynamics. PLoS One 8:e68198. doi:10.1371/journal.pone.0068198
Kerrison P, Hall-Spencer JM, Suggett DJ, Hepburn LJ, Steinke M (2011) Assessment of pH variability at acoastal CO2 vent for ocean acidification studies. Estuar Coast Shelf Sci 94:129–137. doi:10.1016/j.ecss.2011.05.025
Kim TW, Barry JP, Micheli F (2013) The effects of intermittent exposure to low pH and oxygen conditionson survival and growth of juvenile red abalone. Biogeosci Discuss 10:3559–3576
Kroeker KJ, Kordas RL, Crim R, Hendricks IE, Ramajo L, Singh GS, Duarte CM, Gattuso JP (2013)Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction withwarming. Glob Change Biol. doi:10.1111/gcb.12179
Kurihara H, Kato S, Ishimatsu A (2007) Effects of increased seawater pCO2 on early development of theoyster Crassostrea gigas. Aquat Biol 1:91–98
Kurihara H, Asai T, Kato S, Ishimatsu A (2008) Effects of elevated pCO2 on early development in themussel Mytilus galloprovincialis. Aquat Biol 4:225–233. doi:10.3354/ab00109
Mehrbach C, Culberso CH, Hawley JE, Pytkowic RM (1973) Measurement of apparent dissociation-con-stants of carbonic-acid in seawater at atmospheric pressure. Limnol Oceanogr 18:897–907
Moulin L, Catarino AI, Claessens T, Dubois P (2011) Effects of seawater acidification on early developmentof the intertidal sea urchin Paracentrotus lividus (Lamarck 1816). Mar Pollut Bull 62:48–54
O’Donnell MJ, George MN, Carrington E (2013) Mussel byssus attachment weakened by ocean acidifi-cation. Nat Clim Change 3:587–590
Orr JC, Fabry VJ, Aumont O, Bopp L, Doney SC, Feely RA, Gnanadesikan A, Gruber N, Ishida A, Joos F,Key RM, Lindsay K, Maier-Reimer E, Matear E, Monfray P, Mouchet A, Najjar RG, Plattner GK,Rodgers KB, Sabine CL, Sarmiento JL, Schlitzer R, Slater RD, Totterdell IJ, Weirig M, Yamanaka Y,Yool A (2005) Anthropogenic ocean acidification over the twenty-first century and its impact oncalcifying organisms. Nature 437:681–686
Pierrot D, Lewis E, Wallace DWR (2006) MS Excel Program developed for CO2 system calculations.ORNL/CDIAC-105a. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory.US Department of Energy, Oak Ridge, Tennessee
Renaud SM, Parry DL, Luong-Van T, Kuo C, Padovan A, Sammy N (1991) Effect of light intensity in theproximate biochemical and fatty acid composition of Isochrysis sp. and Nannochloropsis oculata foruse in tropical aquaculture. J Appl Phycol 3:43–53
Rossetto M, De Leo GA, Bevacqua D, Micheli F (2012) Allometric scaling of mortality rates with bodymass in abalones. Oecologia 168:989–996
Royal Society (2005) Ocean acidification due to atmospheric carbon dioxide. Working Group members:Raven J, Caldeira K, Elderfield H, Hoegh-Guldberg O, Liss P, Riebesell U, Shepherd J, Turley C,Watson A. The Royal Society London. http://www.royalsoc.ac.uk
Timmins-Schiffman E, O’Donnell MJ, Friedman CS, Roberts SB (2012) Elevated pCO2 causes developmentdelay in early larval Pacific oysters, Crassostrea gigas. Mar Biol. doi:10.1007/s00227-012-2055-x
Aquacult Int
123