factors influencing food selection in the abalone haliotis rubra (mollusca: gastropoda
TRANSCRIPT

J. Exp. Mar. Biol. Ecol. 176 (1994) 27-37
JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY
Factors influencing food selection in the abalone Haliotis rubra (Mollusca: Gastropoda)
Paul E. McShane”,*, H. K. Gorfine”, I. A. KnuckeyaTb
“Marine Science Laboratories, Department qf Conservatim and Environment. Queensclt$ Vietovio, Australirr;
“Research Branch. Fisheries Division, Department qf Primary Industry and Fisheries, GPO Box 990, Darwin.
NT 0801, Australiu
(Received 12 November 1992; revision received 21 September 1993; accepted 11 October 1993)
Abstract The feeding rates of the abalone Huliotis rubru were measured for a range of macroalgal diets. The macroalgae Gigartina radula, Plocamium mertensii, Ecklonia mdiata, Phyllospora con~osc~, Jenerettia lobata, Ulva lactuca and Macrocystis augustofolia showed interspecific variation in ash, nitrogen and energy content; toughness (resistance to penetration); and rates of assimilation and food conversion efficiency by H. rubru. Tests with an artificial diet (agar) showed that feeding rates by H. rubra were influenced by toughness. Toughness also accounted for much of the variation (60%) in the feeding rate on the macroalgal diets. The results of our study show little evidence for chemical deterrence of herbivory for H. rubra, but suggest that food toughness is
a primary factor in the feeding preferences of abalone.
Key words: Abalone; Haliotis rubra; Macroalgal diet
1. Introduction
Studies of the feeding biology of abalone (Hal&is spp.) reveal that Australasian species have a preference for red macroalgae (division Rhodophyta) over brown macro- algae (division Phaeophyta) (Poore, 1972; Shepherd, 1973; Wells & Keesing, 1989);
whereas the reverse, in general, is true for abalone from North America (Cox, 1962; Leighton & Boolotian, 1963; Breen, 1980; Tutschulte & Connell, 1988), South Africa (Barkai & Griffiths, 1986) and Japan (Uki et al., 1986). One hypothesis explaining these
food preferences in abalone is the presence of unpalatable chemicals which act as chemical deterrents against herbivory (Steinberg 1988, 1989; Shepherd & Steinberg,
* Corresponding author. Present address: Ministry of Agriculture, Fisheries, Fisheries Research Division,
P. 0. Box 297, Wellington, New Zealand
0022-0981/94/$7.00 0 1994 Elsevier Science B.V. All rights reserved
SSDI 0022-098 1(93)E0149-S

28 P. E. McShane et cd. : J E.Yp. Mu. Bid. Ed I76 (I 994) 2 7-37
1992, Steinberg & van Altema, 1992). The main chemicals are polyphenolic (poly-
phloroglucinol) compounds, although other secondary metabolites such as terpenoids
and prenylated or alkylated phenolics are present in brown macroalgae and may have
anti-herbivore properties (Steinberg, 1989; Steinberg & van Altema, 1992). Abalone are generalist herbivores feeding primarily on macroalgal drift in proportion
to the general abundance of acceptable species (Cox, 1962; Leighton & Boolootian, 1963; Shepherd 1973, Breen, 1980; Barkai & Griffiths, 1986). The concentration of polyphenols can be maintained even in drift macroalgae and deter herbivores (Ragan & Glombitza, 1986; Steinberg, 1989). Acceptance or rejection of food, according to the chemical defence hypothesis, depends on the relative concentration of polyphenols, which are natural compounds present in marine brown algae (Steinberg, 1984). Much of the evidence in support of this hypothesis is correlative (e.g., Shepherd & Cannon,
1988; Steinberg, 1985, 1988; Shepherd & Steinberg, 1992; Steinberg & van Altema, 1992), yet recently Winter & Estes (1992) demonstrated a dose-dependent effect of phenolics on feeding by abalone (H. rufescens).
Evidence contrary to chemical defense of herbivory includes lack of avoidance by abalone of brown algae with high levels of polyphenolics (Barkai & Griffiths, 1986;
Shepherd & Steinberg, 1992; Steinberg, 1992) avoidance by herbivores of brown algae (kelps) with low levels of phenolics (Steinberg, 1985, 1989; Shepherd & Steinberg, 1992; Steinberg & van Altema, 1992) the fact that phenolics in living brown algae arc gen- erally not exuded and must be ingested to be sensed (Ragan, 1976; Winter & Estes, 1992; but see Ragan & Glombitza, 1986) the high intraspecific variation in phenolic concentration (Ragan & Glombitza, 1986; Steinberg & van Altema, 1992) and adap- tations by herbivores to counter the digestive-reducing effects of polyphenols (Martin & Martin, 1983; Martin et al., 1985; but see Steinberg & van Altema, 1992, and Tugwell
& Branch, 1992). The morphology or texture of macroalgae is also a factor in the diet preference of
marine herbivores (Littler & Littler, 1980; Steneck & Watling, 1982; Steinberg, 1985; Padilla, 1985; 1989). Abalone, which lack hardened teeth on the radula, are morpho- logically constrained to consume macroalgae with soft rather than leathery thalli
(Steneck & Watling, 1982). Hdiotis vubm is a commercially important abalone widely distributed off southern
Australia (Shepherd, 1973; McShane & Smith, 1991). It has previously been observed
that H. rubra consumes more fresh macroalgae than artificial food, probably because the alga (Jenneretia lobata) was not as tough as the artificial food pellets and was therefore easier to breakdown mechanically (Gorfine, unpublished data). Here we reinforce the importance of toughness in the feeding preference of abalone with evidence from studies of the feeding rate of H. rubra on a variety of diets.
2. Materials and methods
2. I Test systems
Juvenile Haliotis rubra (mean shell length 43 mm) were collected near Beaumaris in Port Philip Bay (McShane et al., 1986) and maintained without food for 4 weeks prior to tests in flow-through tanks. Juveniles were used because incremental growth (shell

P.E. McShane et al. /J. E-up. Mar. Bid. Ed. I76 (1994) 27-37 29
length or total weight) is higher and easier to detect than for adult H. rubra (McShane & Smith, 1992). Some animals were starved and then dissected after increasing periods
revealing that guts were empty after 5 days.
Acclimation of H. rubra to aquaria was considered important. Koike et al. (1979) showed that H. tuberculata required an acclimation period of approximately 2 months and Day & Fleming (1992) demonstrated reduced growth rates in aquaria for H. rubra during the first month after collection, but they also showed that individuals fed on single species diets had reduced growth after 50 days. Because we were inter- ested more in relative rather than absolute performance of various diets, we considered
an acclimation period of 4 weeks to be adequate. The weight of food provided was about 5 % of the total wet weight of each H. rubra.
We used the laminae of fresh Macrocystis augustofolia, Phyllospora comosa, Ulva lactuca,
Plocamium mertensii, Gigartina radula, Jennerettia lobata and Ecklonia radiata as test diets. We also used dried Jennerettia lobata (oven dried at 80 “C for 48 h). Each treatment (species of macroalgae) involved six H. rubra. For testing of diets, individual H. rubra were maintained in plastic “bait buckets” (N 11 capacity) within separate tanks (-41 capacity) supplied with sand filtered seawater (1 l.min-I). Temperature
was controlled (17 & 0.5 “C) and test animals were given approximately 10% shaded sunlight on a natural cycle.
Unconsumed food was removed and weighed each morning and fresh diets were provided daily in the late afternoon. During tests, daily measurements of the relative proportion of each diet ingested, the amount of each diet uneaten (g dry wt.), and the amount of faecal production (g dry wt.) were recorded. Tests were of 5 days duration.
Time-lapse video photography was used to record and compare movements of H. rubra feeding on J. lobata and an artificial diet. Illumination was provide for 15 min periods each hour using a timer-coupled switch. Individual animals were en-
closed in two shallow (40 mm) perspex trays located on a grid of 50 mm squares. In one tray, H. rubra were provided with an artificial diet (pasta), distributed randomly, which mimicked the appearance of J. lobata. In another tray, pieces of J. lobata were provided in the same distribution pattern. This experiment was repeated four times.
2.2. Analyses of nitrogen, ash and energy content
The nitrogen, ash and energy content of six macroalgal species: Macrocystis augusto-
,folia, Phyllospora comosa, Plocamium mertensii, Gigartina radula and Ecklonia radiata was assessed. The analyses were performed on each of three replicates for each species
of alga. The stipe and pneumatocysts of M. augustofolia and P. comosa were removed prior
to analysis, as these were not eaten by H. rubra. The laminae of each alga were ground with a mortar and pestle and dried to constant weight at 60 “C before analysis. Ash
of algae was weighed to the nearest 10 mg after heating to 500 “C for at least 12 h. A Kjeltec Auto 1030 analyser was used to analyse the total nitrogen content of the algae.
Pellets of the laminae of the algae were made using an air-driven pellet press. The energy content of the pelleted algae was determined with a microbomb calorimeter, calibrated using Benzoic acid.

30 P.E. McShane et al. /J. Exp. Mar. Bid. Ed. 176 (1994) 27-37
2.3. Feeding studies
For feeding studies, each replicate comprised two containers, each with two ran-
domly chosen H. rubru. A known weight of each species of macroalgae, including Ulva luctuca, was added to each container at weekly intervals and the macroalgae remain-
ing after each week was weighed, dried and weighed again (to the nearest mg). A similar procedure was followed in the absence of H. rubra. The faecal material was siphoned from each container daily, rinsed in fresh water and sieved to remove any small pieces of macroalgae. The macroalgae retained in the sieve were replaced in the container and the faecal material collected for each replicate was placed in an oven at 70 o C and dried. Analyses of the macroalgal and faecal material, together with the measurements of total ingestion and faecal excretion enabled accurate measurement of the nutritional value
of each alga to the abalone. Huliotis rubra were fed Jennerettia lobatu and Phyllospora comosa at a rate of 5% of
the live weight of the abalone per day. The food rations were indexed to growth and were adjusted proportionally to the wet weight of H. rubra on a weekly basis. Experi- ments for ten individuals per treatment were run over 9 weeks. During the experiment the length of each H. rubra was recorded weekly to the nearest 0.1 mm and weights
were measured at the beginning and end of the trial to the nearest mg. Increases in weight were used to determine the food conversion efficiency (FCE) of the diet (gain in the live weight of the individual divided by the dry weight of macroalgae supplied).
2.4. Food texture
To test for feeding preference of H. rubra related to texture, agar was used because it is the main structural polysaccharide found in red macroalgae (Stewart, 1974) is
known to be palatable and to support the growth of abalone (McShane & Gorfine, unpublished data) and because we could vary the texture (toughness) in experimental diets. Laboratory grade agar (Ajax No. 863) was used to manufacture experimental diets because of its excellent gel-forming properties and its palatability to H. rubru. The compound forms gels of a range of toughness according to concentration. The agar was combined with demineralised water and 5% (w/v) sodium chloride to render the pellets negatively buoyant in seawater. The pellets were disc shaped with a diameter of 15 mm and 2-4 mm in thickness to mimic pieces of drift algae that H. rubru consume naturally.
Pellets of 2, 4, 6 and 8% agar were pre-weighed and supplied in the late afternoon to nine animals per treatment. We were constrained in this range of agar concentra- tions because agar failed to gel at concentrations greater than S”/, and less than 1%. Nonetheless, the pellets showed a range of toughness values consistent with the inter- specific variation measured for natural diets.
The following morning uneaten and partly eaten pellets were collected, “damped” dry, reweighed and observations recorded. The final pellet weight was subtracted from the initial weight to obtain the quantity of food ingested for each treatment replicate. This was repeated on 12 consecutive days to provide a total of 108 observations per treatment.
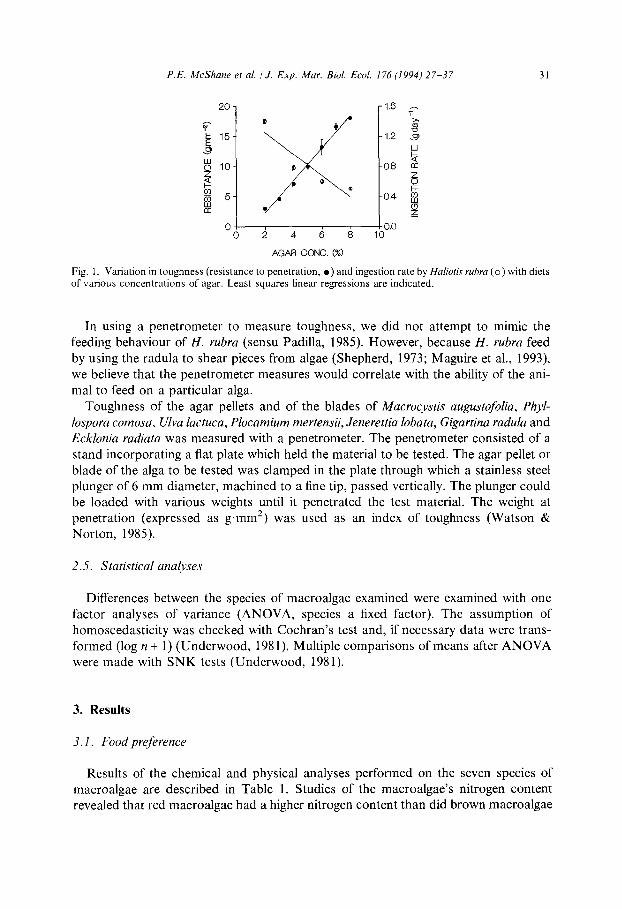
P.E. McShane et al. /J. Exp. Mar. Bid. Ed. I76 (1994) 27-37 31
1.6 -7
B 1.2 5
p
0.8 2
g
0.4 fi
9
- 0.0 10
AGAR CGNC. (%)
Fig. 1. Variation in toughness (resistance to penetration, l ) and ingestion rate by Haliotis rubru (0) with diets of various concentrations of agar. Least squares linear regressions are indicated.
In using a penetrometer to measure toughness, we did not attempt to mimic the feeding behaviour of H. rubra (sensu Padilla, 1985). However, because H. rubra feed by using the radula to shear pieces from algae (Shepherd, 1973; Maguire et al., 1993) we believe that the penetrometer measures would correlate with the ability of the ani- mal to feed on a particular alga.
Toughness of the agar pellets and of the blades of Macrocystis augustofolia, Phyl-
lospora comosa, Ulva lactuca, Plocamium mertensii, Jenerettia lobata, Gigartina radula and Ecklonia radiata was measured with a penetrometer. The penetrometer consisted of a
stand incorporating a flat plate which held the material to be tested. The agar pellet or blade of the alga to be tested was clamped in the plate through which a stainless steel plunger of 6 mm diameter, machined to a fine tip, passed vertically. The plunger could be loaded with various weights until it penetrated the test material. The weight at penetration (expressed as g.mm2) was used as an index of toughness (Watson & Norton, 1985).
2.5. Statistical analyses
Differences between the species of macroalgae examined were examined with one factor analyses of variance (ANOVA, species a fixed factor). The assumption of homoscedasticity was checked with Cochran’s test and, if necessary data were trans- formed (log n + 1) (Underwood, 198 1). Multiple comparisons of means after ANOVA were made with SNK tests (Underwood, 1981).
3. Results
3.1. Food preference
Results of the chemical and physical analyses performed on the seven species of macroalgae are described in Table 1. Studies of the macroalgae’s nitrogen content revealed that red macroalgae had a higher nitrogen content than did brown macroalgae
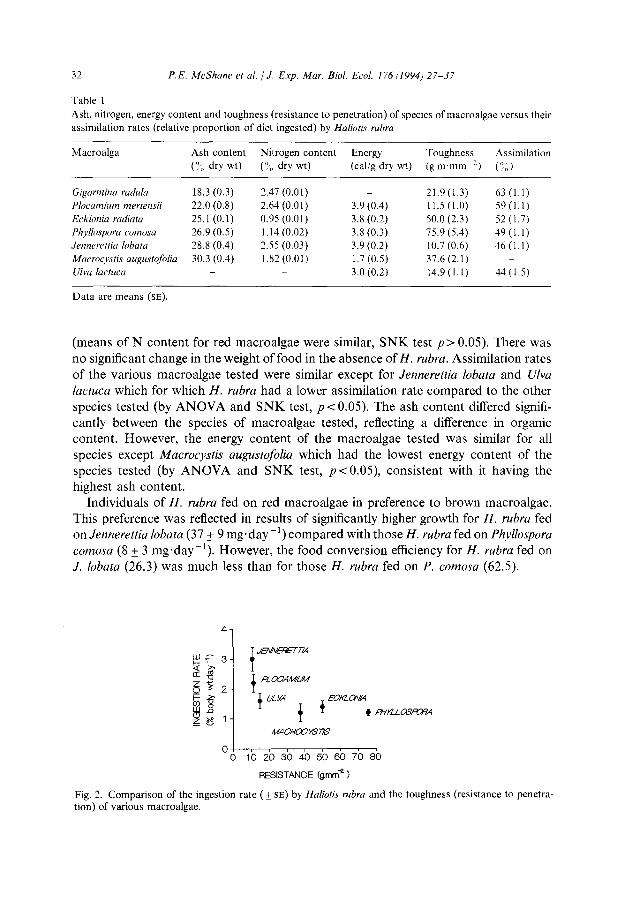
32 P.E. McShane et al. 1 J. Exp. Mar. Biol. Ecol. 176 (1994) 27-37
Table 1
Ash, nitrogen, energy content and toughness (resistance to penetration) of species of macroalgae versus their
assimilation rates (relative proportion of diet ingested) by Haliotis rubra
Macroalga Ash content Nitrogen content Energy Toughness Assimilation (u; dry wt) (4: dry wt) (Cal/g dry wt) (g m.mm-‘) (“,)
Gigarntina radula 18.3 (0.3) 2.47 (0.01) 21.9(1.3) 63 (1.1)
Plocamium inertensii 22.0 (0.8) 2.64 (0.01) 3.9 (0.4) 11.5 (1.0) 59(1.1) Ecklonia radiata 25.1 (0.1) 0.95 (0.01) 3.8 (0.2) 50.0 (2.3) 52 (1.7)
Phyllospora comosa 26.9 (0.5) 1.14(0.02) 3.8 (0.3) 75.9 (5.4) 49(1.1) Jennerettia lobata 28.8 (0.4) 2.55 (0.03) 3.9 (0.2) 10.7 (0.6) 46 (1.1) Macrocystis augustofolia 30.3 (0.4) 1.82 (0.01) 1.7 (0.5) 37.6 (2.1) _
Ulna lactuca 3.0 (0.2) 14.9(1.1) 44 (1.5)
Data are means (SE).
(means of N content for red macroalgae were similar, SNK test p>O.O5). There was
no significant change in the weight of food in the absence of H. rubra. Assimilation rates of the various macroalgae tested were similar except for Jennerettia lobata and Ulva
lactuca which for which H. rubra had a lower assimilation rate compared to the other
species tested (by ANOVA and SNK test, p< 0.05). The ash content differed signifi- cantly between the species of macroalgae tested, reflecting a difference in organic content. However, the energy content of the macroalgae tested was similar for all
species except Macrocystis augustofolia which had the lowest energy content of the species tested (by ANOVA and SNK test, p<O.O5), consistent with it having the highest ash content.
Individuals of H. rubra fed on red macroalgae in preference to brown macroalgae. This preference was reflected in results of significantly higher growth for H. rubra fed on Jennerettia lobata (37 + 9 mg*day -‘) compared with those H. rubra fed on Phyllospora
comosa (8 + 3 mg.day -‘). However, the food conversion efficiency for H. rubra fed on J. lobata (26.3) was much less than for those H. rubra fed on P. comosa (62.5).
01 0 1020304050607080
RESISTANCE (g.mm* )
Fig. 2. Comparison of the ingestion rate ( k SE) by Huliotis rubra and the toughness (resistance to penetra- tion) of various macroalgae.

P.E. McShane et al. / .I. Exp. Mu. Biol. Ed. 176 11994) 27-37 33
Consumption rates of dried versus fresh Jennerettia lobata were similar at 40 k 2.4
and 38? 2.3 mg*day-’ respectively (by ANOVA: df 1,ll; F= 0.38, p= 0.87). Time- lapse video photography confirmed that H. rubra fed actively after dusk (sensu
Shepherd, 1973) and that they were observed to be attracted to pieces ofJ. lobata rather than the artificial diet. The results suggest foraging behaviour by H. rubra and a chemo- tactic response to J. lobata.
There was significantly greater faecal production by H. rubra fed on dried J. lobata (69 k 4 pellets/individual) compared with fresh J. lobata (38 + 3 pellets/individual) ANOVA p < 0.00 1).
3.2. Food texture
The ingestion rate of agar pellets was related to the concentration of agar. The re- lationship of ingestion rate (g.day -‘) (Y) with agar concentration (%) (X) was highly significant (v2 = 0.9, p = 0.0001) where Y= - 2.6X+ 22.0. The toughness of the agar pellets (measured as resistance to penetration) increased proportionally with the con-
centration of agar. Thus the ingestion rate of agar by H. rubra (Y) was inversely pro- portional to the toughness (resistance, X) of the pellet (Y= - 0.05X+ 1.38; r2 = 0.7). Examination of intact pellets on the day after feeding revealed that each of the remaining pellets in any given container had been grazed across the flat surface of the discs as well as around the perimeter. Examination of the feeding behaviour of H. rubra on a preferred diet of J. lobata revealed that individuals fed selectively on the membranous regions and avoided the tougher mid-ribs and fruiting bodies of the laminae.
Toughness of the laminae varied between the seven species tested; Plocamium
mertensii and J. lobata had similar toughness (by ANOVA and SNK test, p< 0.05; Table 1). The consumption rates of H. rubra were related to the toughness of the various species of macroalga (comparison of ingestion rate with toughness $ = 0.6). The most preferred algae, J. lobata and P. mertensii were the least tough of the algae tested.
4. Discussion
We tested a range of diets commonly encountered in the natural habitat of Haliotis
rubra (see Shepherd, 1973; Foale & Day 1992). Feeding and growth rates of H. rubra
on all diets were relatively low (cf. Uki et al., 1986; Day & Fleming, 1992; Foale & Day, 1992) possibly because the acclimation period of the experimental animals was not long enough (Day & Fleming, 1992). However, growth rates of abalone show high intra- specific variation and absolute feeding rates on monospecific diets vary according to several factors (reviewed by Day & Fleming, 1992). We argue below that the morphol- ogy of macroalgae, specifically their toughness, is preeminent in determining the rela- tive feeding rates of H. rubra on various diets.
Feeding rates were positively correlated with nitrogen content but not with the calorific value or the ash content of the diets that we tested. However, in another study Fleming (1991) showed that feeding rate of H. rubra varied independently of the nitrogen content for several species including J. lobata. Other studies of marine herbi-

34 P.E. McShane et al. /J. Exp. Mar. Bid. Ed I76 (1994) 27-37
vores reveal that feeding preference is not correlated with the energy content of macro-
algae perhaps because much of the energy content derives from non-palatable struc-
tural polysaccharides (Paine & Vadas, 1969; Nicotri, 1980). Steinberg (1985) showed
that feeding preference of the intertidal gastropod Tegzda funebralis was positively correlated with nitrogen content, but this result was attributed to a co-association with low levels of phenolics. Ecklonia radiata, with a low organic nitrogen content did not support the long term growth of H. rubra even though it was a preferred diet (Fleming, 1991). Haliotis rubra are evidently not choosing diets with a high nutritive value. There was some evidence from our study that Haliotis rubra were attracted to Jenerettia lobata.
Jenerettia lobata is preferred food for H. rubra (Fleming, 1991; Foale & Day, 1992; our study) but the attraction to J. lobata dried at 80 “C casts doubt on the influence of volatile organic phagostimulants (cf. Sakata & Ina, 1983). Non-volatile nutrients such as sugars or amino acids may be important in stimulating feeding (Carefoot, 1982).
We showed that of the diets that were eaten, over 50% was lost in the form of faeces with the exception of Gigartina radula, Plocamium mertensii and Ecklonia radiata in which assimilation efficiencies ranged from 5243%. Our findings parallel those of Barkai & Griffiths (1988) who postulated that low assimilation of natural diets could
be attributed to the toughness or texture of the algae preferred by abalone (Haliotis
midae). However, efficiencies of food absorption for marine gastropods are generally low (Bayne & Newell, 1983).
The results of our study support suggestions by other workers (e.g., Littler & Littler, 1980; Steneck & Watling, 1982; Watson & Norton, 1985; Padilla, 1985, 1989; Winter
& Estes, 1992) that algal morphology is important in influencing feeding preference by marine invertebrate herbivores. Abalone have a rhipodoglossan radula which, because
of a lack of hardened teeth, mitigates against grazing on tough macrophytes such as fucoids or on the stipes of other seaweeds (see Steneck & Watling, 1982). Indeed, rhipodoglossan grazers rarely graze leathery macroalgae although some large abalone graze the detached thalli of some kelps (Steneck & Watling, 1982). This was clearly demonstrated in our experiments with agar where feeding rates by H. rubra were in- versely related to toughness. Conversely, gastropods with harder radulae show feed-
ing preferences less influenced by toughness (e.g., Steinberg, 1985 for Tegulajiinebralis).
Such gastropods are negatively responsive to polyphenols in brown macroalgae (see also Gielselman & McConnell, 1981). The use of juveniles in our study could have exaggerated the effects of toughness as applied to adult H. rubra. However, Shepherd & Steinberg (1992) found that feeding rates of adult H. rubra were negatively correlated with toughness as we found with juveniles.
The results of our study do not support the hypothesis that chemicals present in brown algae are the main deterrents to herbivory (Steinberg, 1985, 1988; Tugwell & Branch, 1992). One explanation is that Australasian species have evolved a tolerance
to high levels of phenolics in some macroalgae (Estes & Steinberg, 1988; Steinberg, 1989; Shepherd & Steinberg, 1992; Steinberg & van Altema, 1992). Studies of the phenolic content of Australian species of brown algae reveal that Ecklonia radiata has the highest concentration of phenolics (5.5% of dry weight) followed by Phyllospora
comosa (2.7% of dry weight) and Macrocystis augustifolia (0.7pb) (Steinberg, 1989).

P.E. McShane et al. / .I. Exp. Mar. Bid. Ed. 176 (1994) 27-37 35
However, these concentrations relate to whole tissue measurements and concentrations of polyphenolics could be substantially higher in the meristoderm (Tugwell & Branch,
1989). Furthermore, there is intraspecific variation in the phenolic content of some
macroalgae (e.g. E. radiata) depending on age or location (Ragan & Glombitza, 1986; Steinberg & van Altema, 1992). Even so, Steinberg (1989) concluded that brown macroalgae from Australian waters had typically high levels of polyphenols compared with species from North American waters.
In feeding principally on drift, and in being morphologically constrained to consum- ing the softer laminae of macroalgae, abalone would be expected to target the meris- toderm tissue in herbivory. The kelp Ecklonia maxima has extraordinarily high levels of polyphenolics in the meristoderm tissue (- 39y10 dry weight, Tugwell & Branch, 1989) but it is the preferred natural diet of the abalone H. midae (Barkai & Griffiths, 1986). We showed that the laminae of Macrocystis augustofolia had lower toughness than Ecklonia radiata. With both low toughness and low phenolics, M. augustofolia had little defence against herbivory. Indeed, M. augustofolia ranks as a preferred diet for H. rubra (Foale & Day, 1992). Laminarians such as M. augustofolia are characteristi- cally fast growing and there may be a trade off between defence and growth rate as
suggested for terrestrial plants (Coley, 1983). Foale & Day (1992) showed that the digestibility of the three species of brown algae
listed above was inversely correlated with phenolic concentration, reinforcing the sug- gestion that polyphenolics retard digestion (reviewed by Tugwell & Branch, 1992). However, in contrast, we showed that food conversion efficiency for Phyllospora comosa
was much higher than the preferred alga Jenerettia lobata. Moreover, there was no difference in the assimilation rates of these two algae by H. rubra. Haliotis rubra, can digest 74% of the dry matter in P. comosa (Maguire et al., 1993) casting further doubt on the digestion-reducing role of polyphenolic compounds in macroalgae (see also Steinberg & van Altema, 1992).
Plants are unlikely to evolve defensive strategies against herbivores such as abalone which feed primarily on detached (drift) macroalgae. Nonetheless, abalone as drift
feeders would be expected to respond to defensive strategies developed against herbi- vores, such as echinoids, which directly graze macroalgae. Such defensive strategies include the presence of unpalatable chemicals and morphological adaptations such as toughness. Our study demonstrates that toughness of the diet is a significant factor
influencing consumption rates by H. rubra. The general difference in toughness can explain the preference by Australian species of abalone for red over brown macroalgae. However, this preference pattern is not shown by species of abalone elsewhere. The apparent preference of abalone from California, Canada, South Africa and Japan for kelps can be explained by their relative abundance and by their relatively low toughness compared with fucoid macroalgae (Steneck & Watling, 1982; Steinberg
1985). Our study shows that while the presence of unpalatable or digestion inhibiting chemi-
cals can be effective defenses against some marine herbivores, abalone are morpho- logically constrained to consuming relatively soft tissue. Toughness of macroalgae may be the primary factor governing the natural diet preference of abalone.

36 P.E. McShane et al. ! J. Exp. Mar. Biol. Eeol. I76 (1994) 27-37
5. Acknowledgements
We thank Dr. Ann Fleming for technical assistance and advice on abalone hus-
bandry. Two anonymous reviewers provided constructive comments on a previous draft.
6. References
Barkai, R. & CL. Griffiths, 1986. Diet of the South African abalone Huliofis midae. S. A,fr. J. Mar. Sci., Vol. 4, pp. 37-44.
Barkai, R. & C.L. Griffiths, 1988. An energy budget for the South African abalone Ha1iori.y midoe Linnaeus.
J. MON. Stud., Vol. 54, pp. 43-51.
Bayne, B.L. & R.C. Newell, 1983. Physiological energetics of marine molluscs. In, The mollusca edited by
(K.M. Wilbur & A.S.M. Saleuddin), Vol. 4, Academic Press, NY, pp. 407-515.
Breen, P.A., 1980. Measuring fishing intensity and annual production in the abalone fishery of British
Columbia. Can. Tech. Rep. Fi.sh. Aquat. Sri., Vol. 941, 49 pp.
Carefoot, T., 1982. Phagostimulatory properties of various chemicals to sea hares (Ap!v.k kurodai and Aply~io dactylomelu). Mar. Biol., Vol. 68, pp. 207-215.
Coley, P.D., 1983. Herbivory and defensive characteristics of tree species in a lowland tropical forest. Ewl. Monogr., Vol. 53, pp. 209-233.
Cox, K.W., 1962. California abalones, family Haliotidae. Fi.rh. Bull. Cal$, Vol. 118, pp. 1-129.
Day. R.W. & A.E. Fleming, 1992. The determinants and measurement of abalone growth. In, Ahukme of the world: biology. fiheries and culture. edited by S.A. Shepherd, M.J. Tegner & S. Guzman del Proo,
Blackwell Scientific, Oxford, pp. 141-168.
Estes, J.A. & P.D. Steinberg, 1988. Predation. herbivory and kelp evolution. Paleohiolog~~. Vol. 14, 19-36.
Fleming, A.E., 1991. The nutritional biology of the black lip abalone, Holiotis ruhra. Dissertation, Dept of
Zoology, University of Melbourne.
Foale, S. & R.W. Day, 1992. The recognizability of algae ingested by abalone. Aust. J. Mar. Frexhwater Rex., Vol. 43, pp. 1331-1338.
Gieselman, J.A. & O.J. McConnell, 1981. Polyphenols in the brown algae mucus w.riculo.ws and Ascoph~Yhrm nodo.vum: chemical defenses against the herbivorous snail Littorinu lirtorea. J. Chem. Ecol., Vol. 7. pp. 11 15-
1133.
Koikc, Y., J. Flassch & J. Mazurier, 1979. Biological and ecological studies on the propagation of the ormer.
Huliotis tuherculuta (L.). II. Influence of food and density on the growth of juveniles. LN Mer, Vol. 17.
pp. 43-52.
Leighton, D. & R.A. Boolootian, 1963. Diet and growth in the black abalone, Haliotis cmcherodii. Ecolog>~. Vol. 44, pp. 227-238.
Littler. M.M. & D.S. Littler, 1980. The evolution of thallus form and survival strategies in benthic marine
macroalgae: field and laboratory tests of a functional form model. Am. IVat.. Vol. 116. pp. 25-44.
Maguirc, G.B., K.W. Wee & S.M. Hindrum, 1993. Digestibility studies - the “ins” and “outs” of abalone
guts. Aust. Asia Aquaculture, Vol. 7, pp. 42-49.
Martin, J.S. & M.M. Martin, 1983. tannin assays in ecological studies: precipitation of ribulose-l.5-
biphosphate carboxylaseioxygenasc by tannic acid, quebracho and oak foliage extracts. J. Chem. Ewl.. Vol. 9, pp. 285-294.
Martin, M.M., D.C. Rockholm & J.S. Martin, 1985. Effects of surfactants, pH, and certain cations on
precipitation of proteins by tannins. J. Chem. EC&., Vol. 11, pp. 485-494.
McShane. P.E. & M.G. Smith, 1991, Recruitment variation in sympatric populations of Halioti.~ ruhrrr (Mollusca:Gastropoda) in southeast Australian waters. Mar. Ecol. Prog. Ser., Vol. 73, pp. 203-210.
McShane, P.E. & M.G. Smith, 1992. Shell growth checks are unreliable indicators of age of the abalone
Haliotis ruhra (Mollusca: Gastropoda). Aust. J. Mar. Freshwater Res., Vol. 43, pp. 1215-12 19.
McShane. P.E., K.H.H. Beinssen & S. Foley, 1986. Abalone reefs in Victoria - a resource atlas. Mar. Sci. Lab. Tech. Rep. No. 47 (Dept. of Conservation Forests and Lands, Melbourne).
Nicotri, M.E., 1980. Factors involved in herbivore food preference. J. E.Y~. Mu. BiolEcol., Vol. 42, pp. 13-26.

P.E. McShme et al. / J. Ex,v. Mar. Biol. Ecol. 176 (1994) 27-37 37
Padilla, D.K., 1985. Structural resistance of algae to herbivores a biochemical approach. Mm. Biol.,
Vol. 90, pp. 103-109.
Padilla, D.K., 1989. Algal structural defenses: form and calcification in resistance to tropical limpets. Eco/og,r,
Vol. 70, pp. 835-842.
Paine, R.T. & R.L. Vadas, 1969. Calorific values of marine benthic algae and their postulated relation to invertebrate food preference. Mar. Biol., Vol. 4, pp. 79-86.
Poore, G.C.B., 1972. Ecology of New Zealand abalones, H&h species (Mollusca:Gastropoda). 1. Fccd-
ing. N.Z. J. Mar. Freshwuter Res., Vol. 6, pp. 11-22.
Ragan. M.A., 1976. Physodes and the phenolic compounds of brown algae. Composition and significance
of physodcs it2 t’iiao. Bar. Muv.. Vol. 19, pp. 145-154.
Ragan. M.A. & K.W. Glombitza 1986. Phlorotannins, brown algal polyphenols. Prong. Phj,col. Rev..
Vol. 4, pp. 130-241.
Sakata, K. & K. Ina, 1983. Digalactosyldiacylglycerols isolated from a brown alga as effective phagostnnu-
lants for a young abalone. A~ic. Biol. Chem., Vol. 47, pp. 2957-2960.
Sakata, K., Y. Iwase, K. Kato, K. Ina & Y. Machiguchi, 1991. A simple feeding inhibitor assay for marine
herbivorous gastropods and the sea urchin S/rongdocentrotus intennedius and its application to unpalat-
able algal extracts. Nippon Suisar~ Gakkaishi, Vol. 57, pp. 261-265.
Shcphcrd. S.A., 1973. Studies on southern Australian abalone (genus H&fix) I. Ecology of five sympat-
ric spccics. Aust. J. Mar. Freshwater Rex., Vol. 24, pp. 217-257.
Shepherd. S.A. & .I. Cannon, 1988. Studies on southern Australian abalone (genus Haliori.~) X. Food and
feeding of juveniles. J. M&c. Sot. Atrst., Vol. 9, pp. 21-26.
Shepherd, S.A. & P.D. Stcinbcrg, 1992. Food preferences of three Australian abalone species with a rev&
of the algal food of abalone. In, Abalone of the world: biology, ,jkheries and culture, (edited by S.A. Shepherd,
M.J. Tegner & S. Guzman de1 Proo, Blackwell Scientific, Oxford, pp. 169-181.
Steinberg, P.D., 1984. Algal chemical dcfcnsc against herbivores: allocation of phenolic compounds in the
kelp Alaria nwrginata. Science, Vol. 223, pp. 405-407.
Steinberg, P.D., 1985. Feeding preferences of Tegulafitnebraalis (Gastropoda) on marine brown algae: cffccts
of phenolic compounds, nitrogen content, and algal toughness. Ecol. Momgr., Vol. 55, pp. 333-349.
Steinberg. P.D., 1988. Effects of quantitative and qualitative variation in phcnolic compounds on feeding in
three species of marine herbivores. J. E.wp. Mar. Biol. Ecol., Vol. 120, pp. 221-237.
Steinberg, P.D., 1989. Biogeographical variation in brown algal polyphenolics and other secondary metabo-
lites: comparison between temperate Australasia and North America. Oecnl@r, Vol. 78, pp. 373-382.
Steinberg, P.D. & I. van Altema, 1992. Tolerance of marine invertebrate herbivores to brown algal phloro-
tannins in temperate Australasia. Ecol. Mom~gr., Vol. 62, pp. 189-222.
Steneck, R.S. & L. Watling, 1982. Feeding capabilities and limitation of herbivorous molluscs: A functional
group approach. Mrrr. Biol., Vol. 68, pp. 299-319.
Stewart, W.D.P. Algal phpriolog?) a,ld biochemiqv. Blackwell, Oxford, 989 pp.
Tugwell, S. & G.M. Branch, 1989. Differential polyphenolic distribution among tissues in the kelps Ecklmicr
ma\-itttcl, Lamir~aria pallida and Macroqxtis augusf~fijlia in relation to plant-dcfcncc theory. J. E.Y~. Mtrr.
Biol. Ecol., Vol. 129, pp. 219-230.
Tugwell. S. & G.M. Branch, 1992. Effects of herbivore gut surfactants on kelp polyphenol defenses. E&
OW’, Vol. 73, pp. 205-215.
Tutschulte, T.C. & J.H. Connell, 1988. Feeding behaviour and algal food of three species of abalones
(Ha&k) in Southern California. Mar. Ecol. Pmg. Ser., Vol. 49. pp. 57-64.
Uki. N.M. Sugiura & T. Watanabe, 1986. Dietary value of seaweeds occurring on the Pacific coast ofTohoku
for growth of the abalone Haliotis discus hamui. Bull. Jp. Sot. Fish.. Vol. 52, pp. 257-266.
Underwood. A.J., 1981. Techniques of analysis of variance in experimental marine biology and ecology.
Oceanogr. Mar. Biol A17n. Rev., Vol. 19, pp. 513-605.
Watson, D.C. & T.A. Norton, 1985. The physical characteristics of scawccd thalli as dctcrrcnts to litorinc
grazers. Bat. Mu., Vol. 28, pp. 383-387. Wells. F. & J. Kecsing, 1989. Reproduction and feeding in Halioti.~ roei (Gray). Amt. J. Mar. Freshwater Rex..
Vol. 40, pp. 187-197.
Winter, F.C. & J.A. Estes, 1992. Experimental evidence for the effects of polyphcnolic compounds from
Dict~meurum cal~firrzicum Ruprecht (Phaeophyta:Laminariales) on feeding rate and growth on the red
abalone Halioti.7 ryfkscetzs Swainson. J. Esp. Mar. Biol. Ecol.. Vol. 155, pp. 263-277.