pacific dulse ( palmaria mollis) as a food and biofilter in recirculated, land-based abalone culture...
TRANSCRIPT

Pacific dulse (Palmaria mollis) as a food
and biofilter in recirculated, land-based
abalone culture systems
Carl Demetropoulosa,*, Chris Langdonb
aDepartment of Biology, California Lutheran University, 185 Erten St.,
Thousand Oaks, CA 91360, USAbCoastal Oregon Marine Experiment Station, Hatfield Marine Science Center,
Oregon State University, Newport, OR 93367, USA
Received 14 July 2004; accepted 13 August 2004
Abstract
Palmaria mollis (Pacific dulse) is increasingly being used as an in situ biofilter and feed in land-
based abalone culture. This work reviews physical and nutrient requirements for P. mollis culture and
its nutritional value for three abalone species, the Japanese abalone Haliotis discus hannai, the red
abalone Haliotis rufescens, and the U.S. Federally endangered white abalone Haliotis sorenseni.
P. mollis growth was positively correlated with light over specific light densities (SLD) ranging
from 0.0048 to 0.036 mol photons g�1 fresh weight d�1, under tumble culture conditions. Under low
light conditions, growth was greatest at a temperature of 12 8C (SGR = 8.69 � 0.29% d�1), while
under high light conditions growth was greatest at a temperature of 14–18 8C (SGR = 10.5 �0.12% d�1). Addition of Guillard’s f nutrient medium [Guillard, R.R.L., Ryther, J.H., 1962. Studies
on marine plankton diatoms. I. Cyclothella nana Huntedt and Detonula confercacae (Cleve). Gran.
Can. J. Microbiol. 8, 229–239] significantly increased Pacific dulse growth in cultures receiving one
seawater exchange per day; furthermore, growth was significantly improved by addition of trace
metals. The use of NaNO3 as an N source in the f medium resulted in better growth compared with
NH4NO3. The use of phosphorus additions above 83.3 mM P d�1 depressed P. mollis growth rates. P.
www.elsevier.com/locate/aqua-online
Aquacultural Engineering 32 (2004) 57–75
* Corresponding author. Tel.: +1 805 4977 517; fax: +1 805 4977 517.
E-mail address: [email protected] (C. Demetropoulos), [email protected]
(C. Langdon).
0144-8609/$ – see front matter # 2004 Published by Elsevier B.V.
doi:10.1016/j.aquaeng.2004.08.005

mollis growth was positively correlated with concentrations of dissolved inorganic carbon, including
HCO3�. Growth rates were not significantly different over a pH range of 8.3–8.9.
There were no significant toxic effects on Japanese abalone, H. discus hannai cultured in seawater
supplemented with (0.75f + 1.37 mM Zn) trace metals d�1. H. discus hannai showed the highest
growth rates when fed on P. mollis fertilized with f medium and nitrate (NO3�-N) loads ranging from
1176 to 2353 mM d�1 while red abalone, H. rufescens, grew best when fed on P. mollis fertilized with
f medium and nitrate loads of 2353–2942 mM d�1. Both abalone species grew better on P. mollis
fertilized with (0.75f + 1.37 mM Zn) d�1 trace metal solution compared with P. mollis without trace
metal additions. H. sorenseni, grew well when fed on P. mollis, showing higher linear shell growth
rates and higher survival rates compared with those of abalone fed on commercially harvested and
abundant giant kelp, Macrocystis pyrifera.
# 2004 Published by Elsevier B.V.
Keywords: Palmaria mollis; Pacific dulse; Cultivation; Growth factors; Haliotis; Recirculating systems
1. Nomenclature and abbreviations
1. Diffusive boundary layer (DBL): A thin layer of static or unstirred water adjacent to
the algal surface through which transport of gases and nutrients is diffusive rather than
convective.
2. Dissolved inorganic carbon (DIC): Total amount of in organic carbon, generally
available as a combination of three species; CO2, HCO3� and CO3.
3. Dry weight (dw): Algae that was dried at 60 8C for 24 h.
4. Epiphytes: Algae that tends to be found growing on other algae.
5. Linear growth rate (LGR): A measure of abalone shell growth on a per day basis.
LGR = (Lf � Li)/t where Lf and Li were the final and initial abalone lengths and t was
time in days.
6. Photosynthetically active radiation (PAR): The broad region of irradiance absorbed by
chlorophyll and other light harvesting pigments typically encompassing a range of
wavelengths from 300 to 700 nm.
7. Photon flux density (PDF): The number of photons of photosynthetically active
radiation (PAR) received by a surface per unit time. PDF = mol photons m�2 d�1.
8. Plasmalemma: Membrane separating the cytoplasm of a cell from the algal cell wall.
9. Rosette: Cluster or ball of thalli.
10. Specific growth rate (SGR): A measure of whole body abalodne growth on a per day
basis. SGR = 100 � (ln Wf � ln Wi)/t, where Wf and Wi were the final and initial
abalone dry meat weights (g) and t was time in days.
11. Specific light density (SLD): A model predicting growth of P. mollis as a function of
incident light supplied to the algae per day on a wet weight basis. SLD = PFD � SA/
FW where PFD was the photon flux density in mol photons, SA was the area of
illuminated surface in m2, FW was the fresh weight of P. mollis in grams.
12. Specific-pathogen-free (SPF): Used to denote cultured organisms that are free of a
specific pathogen known to cause disease and/or mortality in cultured and wild
organisms.
13. Thalli: The plant body of algae.
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7558

2. Seaweed-abalone co-culture systems
The major limiting factor to expansion of the abalone industry in the U.S. continues to
be availability of suitable and inexpensive foods. Traditional U.S. commercial abalone
culture systems depend on supplying abalone with harvested kelp. Unfortunately,
nutritional value and availability of kelp varies with season; furthermore, kelp generally
degrades in abalone culture systems, leading to deterioration in water quality. Co-culture
systems depend on the culture of abalone and seaweed in the same culture water. Abalone
feed on seaweed and excrete NH4+ and CO2 that are, in turn, taken up by the seaweed and
converted into algal biomass (cf., Neori et al., 1996; Evans et al., this volume). In this way
nutrients are recycled within the system resulting in both lower seawater exchange rates
and lower pumping costs that can represent as much as 30% of operational budgets for
land-based systems (Huguenin, 1976; Shpigel and Neori, 1996; Neori et al., 2000). Further,
a significant reduction of effluent nutrient loads into the environment acts to mitigate the
need for discharge regulation and monitoring.
Seaweed biofilters have several significant advantages over bacterial biofilters.
Generally, macroalgae can remove all forms of inorganic nitrogenous waste products from
culture systems, while nitrifying bacteria filters do not remove NO3�. Macroalgae are also
more reliably maintained than bacterial biofilters and are not typically subject to
unpredictable crashes due to changes in microbial communities. Further, macroalgae are
very efficient at removing CO2 and can increase dissolved oxygen concentrations by as
much as 233% as a result of photosynthesis (Demetropoulos, unpublished data). Finally,
seaweed from biofilters can be used to provide food for cultured organisms, such as
abalone, bivalves, and fish (Shpigel and Neori, 1996).
There are also important limitations associated with establishing successful land-based
abalone farms that revolve around prevention of disease epizootics; for example, the
California abalone industry has recently been severely impacted by ‘‘withering foot
syndrome’’, a disease caused by a Rickettsia-like intracellular bacterial pathogen (RLP),
Candidatus xenohaliotis californiensis (Gardner et al., 1995; Friedman et al., 2000, 2002;
Moore et al., 2000). These bacteria are present in coastal waters of Southern California and
abalone cultured in open systems can become infected as transmission requires no
intermediate host and direct contact between infected and uninfected abalone is not
required for disease transmission to occur. Use of co-culture recirculation systems, with P.
mollis or another suitable macrophyte acting as both biofilter and food, can avoid potential
infections of cultured abalone from wild harvested seaweed, such as currently experienced
with the use of Macrocystis pyrifera as food for Haliotis sorenseni (Demetropoulos,
unpublished data). Thus, co-culture recirculation systems can play a significant role in
producing ‘‘High health’’ or specific-pathogen-free (SPF) abalone seed that are valuable
for both abalone restoration efforts and commercial production.
3. Physical parameters of Pacific dulse culture
3.1. Tank design and aeration
Tank design and water motion are important in supporting the best growth of P. mollis.
Air-generated tumble culture cells provided turbulent water motion for efficient mass
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–75 59

transfer of gas and nutrients across the diffusive boundary layer (DBL) adjacent to thalli of
P. mollis. This was generally accomplished by vigorous aeration of tanks. Proper
placement of air lines was necessary for (1) efficiently cycling rosettes through a zone of
high light at the surface, (2) decreasing DBL thickness (Gonen et al., 1993) so as to increase
mass transfer of CO2 and O2, (3) dislodging epiphytes and (4) constantly breaking up large
rosettes to further improve growth (Craigie and Shacklock, 1989).
Tumble culture exposed rosettes to the same amount of light, allowing vegetative
growth of specific P. mollis strains to be sustained for long periods of time (years) under
daily photon flux densities (PFD’s) up to 72 mol photons m�2 d�1. These are considerably
higher than PFD’s typical of temperate regions where P. mollis is found (up to
1800 mmol photons m�2 s�1 from central California to Alaska). Cycling plants between
light and dark in air-generated cells probably allows P. mollis to grow well under higher
PFD’s (cf., Bidwell et al., 1985; Terry, 1986; Kubler and Raven, 1996a, 1996b). Aeration is
not typically required at night (Demetropoulos, unpublished data) and additional cost
savings may be realized by using interrupted aeration during the day (Friedlander and Ben-
Amotz, 1991; Demetropoulos, unpublished data).
Wheeler (1980) showed increased uptake rates of inorganic carbon for M. pyrifera were
a function of decreasing the thickness of the DBL and Gonen et al. (1993) found the relative
water velocity to algal thalli for Gracilaria conferta in culture had a significant affect on
photosynthetic rates with optimal relative velocities of 8 cm s�1. The highest growth rates
for P. mollis under adequate nutrient and pH conditions were observed with a relative water
velocity >10 cm s�1, requiring an aeration rate of between 0.47 and 0.68 l air l water�1
min�1 in 3.8 l vessels (Demetropoulos and Langdon, 2004a,b) and 0.16–
0.23 l air l water�1 min�1 in 15,000 l tanks.
Tube tanks (65 cm deep; 125 l) with transparent side walls and airlines placed
across one side of the bottom, opposite the outlet at the top, were found to accomplish the
highest growth rates and yields as a function of incident light (Demetropoulos and
Langdon, 2004b,c), allowing yields up to 173–197 g [dw] m�3 d�1 (Demetropoulos and
Langdon, 2004b) under a natural PFD of 55 mol photons m�2 d�1. However, practical
considerations such as labor costs suggested culture scale-up in rectangular trough tanks
(70 cm deep; 15,000 l) with curved, opaque sidewalls and a single central airline.
These tanks produced P. mollis yields up to 62 g [dw] m�2 d�1 (�89 g [dw] m�3 d�1)
under a natural PFD from 39 to 52 mol photons m�2 d�1 (Demetropoulos and Langdon,
2004b).
While aerated tanks were reported to be the most energy-efficient methods for moving
seaweed under cultivation (Schramm, 1991), paddle wheel raceways have been shown to
produce growth rates comparable to aerated systems (Neish et al., 1977; Hansen, 1984),
provided the relative velocity of the water medium to the algae was high (>10 cm s�1) (cf.,
Bird and Sanchez, 1990; Gonen et al., 1993; Demetropoulos and Langdon, 2004b). Our
experience with cultivating P. mollis in paddlewheel raceways (34 cm deep; 105,000 l)
suggests that a slow relative velocity (<7 cm s�1) was responsible for producing lower than
expected P. mollis yields on a m�2 basis (39 g [dw] m�2 d�1 or �111 g [dw] m�3 d�1)
under a natural PFD from 36 to 53 mol photons m�2 d�1 and a rosette morphology with
many fine bladelets and a course and somewhat tough texture (Demetropoulos, unpublished
data; cf., Rivero and Viana, 1996).
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7560

3.2. Salinity
When salinity was lowered from 35% to 30 � 1% with municipal water or brackish
water, P. mollis specific growth rates improved by approximately 24% (SGR =
10.92 � 0.64% d�1; Demetropoulos and Langdon, 2004b). The lower salinity treatments
were relatively free of epiphytes as was reported by Friedlander (1992) for G. conferta.
Lobban and Harrison (1997) suggest salinities above 32% can increase energy required for
osmotic adjustment and negatively affect the ability of seaweeds to utilize inorganic carbon
for growth.
3.3. Stocking density, light, and temperature
Understanding light requirements under variable culture conditions as well as designing
culture systems that supply adequate light are among the most important challenges of
land-based seaweed culture systems. Provided other factors are not limiting, P. mollis
cultures become light-limited by self-shading, absorption of light by water and reflection of
light off of air bubbles and culture tank surfaces.
Aerial densities of P. mollis cultures (kg wet weight m�2 d�1) can be adjusted to
produce the best yields as a function of light density (mol photons m�2 d�1) measured at
the tank surface. This typically results in an optimal stocking density for P. mollis between
2 and 4 kg m�2 (Demetropoulos and Langdon, 2004b). This optimal stocking density for
Pacific dulse generally agrees with work reported for Gracilaria sp. and other cultured
macrophytes (Lapointe and Ryther, 1978; McLachlan, 1991; Ugarte and Santelices, 1992;
Friedlander and Levy, 1995). However, because aerial densities can vary with tank depth or
culture design (translucent versus opaque side walls), we employed a biomass-specific
light measurement to create a growth model (Demetropoulos and Langdon, 2004b). The
model allowed prediction of growth rates as a function of specific light density
(SLD = mol photons g�1 wet weight d�1). Ambient light (PFD; mol photons m�2 d�1)
was quantified using a combination of LI-192S (2p) and LI-193S (4p) sensors connected to
a LICOR LI-1000 data logger. Specific light density (SLD) was calculated as:
SLD ¼ PFD � SA
FW(1)
where PFD was photon-flux density in mol photons m�2 d�1, SA was the area of
illuminated surface in m2, FW was the weight of P. mollis in grams, expressed in fresh
weight to make it more useful for commercial applications.
Measurements of specific growth rates of P. mollis as a log function of SLD (in
mol photons g�1 wet weight d�1) generated the following equation:
SGR ¼ 40:896 þ 16:107 log SLD (2)
Using this equation, culture densities can be adjusted according to light densities to maximize
SGR and yields. For example, under the highest PDF tested (51.64 mol photons m�2 d�1;
average culture temperature = 16 � 1 8C), a condition typical of a sunny July day on the
Oregon coast, P. mollis showed SGR’s up to 18% d�1 [dw] (Demetropoulos and Langdon,
2004b), with no light-saturation being observed. Therefore, light saturation must occur at
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–75 61

SLD values above 0.036 mol photons g�1 damp weight d�1 (the highest light density tested
in these experiments); however, light-saturation is likely to occur at lower SLD values for P.
mollis at lower temperatures (Demetropoulos and Langdon, 2004b).
3.4. Temperature and light interaction
While growth as a function of temperature has not been previously reported for P.
mollis, our results agree with those of Robbins (1978) and Morgan et al. (1980) who
showed that P. palmata grew best at 14–15 8C under similar, low light conditions (20–
35 mol photons m�2 d�1). However, under high natural light (55 mol photons m�2 d�1) P.
mollis SGR’s and yields at 18 � 1 8C were not significantly different from those cultured at
14 � 1 to 16 � 1 8C (SGR = 9.2 � 0.5 to 11.1 � 0.5% d�1 and yields = 173–
197 g [dw] m�3 d�1; Demetropoulos and Langdon, 2004b). High SGR’s and yields were
due to the use of translucent tanks in combination with moderate P. mollis stocking
densities (SLD = 0.015 mol photons g�1 wet weight d�1), high seawater exchange rates
(60 vol d�1; Demetropoulos and Langdon, 2004c) and relative velocities >12 cm s�1.
Furthermore, high light conditions supported greater growth rates at the higher temperature
of 18 � 1 8C (Demetropoulos and Langdon, 2004b). Mathieson and Norall (1975) reported
a similar finding for Chondrus crispus where photosynthesis under summer light was
greater at a higher temperature than was optimal during lower light intensities of winter.
4. Nutrient and dissolved inorganic carbon requirements of Palmaria mollis
Nutrient experiments with P. mollis were conducted at either the Hatfield Marine
Science Center (HMSC), Oregon State University, or the Natural Energy Laboratory of
Hawaii (NELH) using a fast-growing strain of P. mollis (C-3 strain) developed by
Demetropoulos and Langdon (2001). Culture were not maintained using sterile technique
so as to approach the commercial reality of air/water-born contamination. Culture vessels
were stocked at P. mollis densities between 2.6 and 11 g wet weight l�1. The nutrient loads
[(concentration of added nutrients in culture) � culture volume/frequency of addition] for
the experimental treatments were held constant by adjusting the concentration of nutrient
additions according to the application rates tested. For example, treatment 1 � 7 d�1
received seven times the concentration on day 7 versus treatment 1 � d�1, which received
nutrients every day. Cultures were ‘‘batch fertilized’’ at night when water could be shut off
such that nutrients were not lost due to flushing yet culture temperatures remained
relatively constant.
4.1. Timing of nutrient addition
Demetropoulos and Langdon (2004a) found P. mollis stored enough nutrients for up to 7
days of growth while maintaining SGR’s of 8.77 and 11.17% d�1 under light levels of 22.5
and 42.3 mol photons m�2 d�1 (respectively) that were not significantly different
(P < 0.001) from those attained by daily nutrient additions. This is in agreement with
results of Morgan and Simpson (1981) for P. palmata and other rhodophytes (Lapointe and
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7562

Duke, 1984; Hanisak, 1990; Pickering et al., 1993). Due to apparent free space located
exterior to the plasmalemma in the cells, macrophytes can often store excess nutrients for
later use (Thomas and Harrison, 1985). It is only when these internal nutrient supplies are
depleted that growth rates decline. An important advantage of pulsing nutrients every seven
days is a dramatic reduction of epiphytes that would otherwise develop in Pacific dulse
cultures (cf., Friedlander et al., 1991; Demetropoulos, personal observation). A further
reduction in epiphyte growth was obtained by pulse fertilizing at night (Hanisak, 1987;
Bidwell et al., 1985; Demetropoulos and Langdon, 2004a).
4.2. Ammonium
Generally, NH4+-N has been considered superior to NO3
�-N for growth of most
macrophytes, because NH4+ can be directly incorporated into amino acids while NO3
� must
be reduced prior use (Lobban and Harrison, 1997). While growth of P. mollis fertilized with
NH4+ and NO3
� as a combined source of nitrogen was not significantly different compared
with that with NO3� alone in a 21-day experiment, the use of NH4
+ as a nitrogen source
depressed growth over a 56-day period (Demetropoulos and Langdon, 2004a). The
advantage of using NH4+-N versus NO3
�-N for cultured rhodophytes appears to vary with
species and under high light, NO3�-N is typically preferred by many rhodophyte species,
including P. palmata (Iwasaki, 1967; Lapointe and Ryther, 1979; Morgan and Simpson,
1981). Further, Maestrini et al. (1982) suggested that in the presence of even low
concentrations (<2 mM) of NH4+-N, NO3
�-N uptake can be inhibited in some microalgae.
Thus, the use of NH4+-N as a nitrogen source for P. mollis culture should be considered with
caution, especially under high light and pulse fertilizing conditions described above.
4.3. Nitrate
Greatest growth of P. mollis occurred with NO3�-N additions of 2942 mM N d�1 with
NO3�-N above this level causing P. mollis growth to decline slightly. The critical tissue N
concentration, correlated with such declines in growth, was 2.97–4.35% [dw] under light
conditions of 23.68–51.64 mol photons m�2 d�1, respectively (Demetropoulos and
Langdon, 2004a). Tissue N and protein concentrations of P. mollis correlated with
highest growth rates were approximately 4.9% and 30.6% [dw] under low light
(23.68 mol photons m�2 d�1) and 4.6% and 28.8% [dw] at high light
(51.64 mol photons m�2 d�1), respectively (Demetropoulos and Langdon, 2003d).
Davison (1991) reported a greater N requirement to support increased photosynthetic
pigment content of cultures under high temperature and light conditions (cf.,
Demetropoulos and Langdon, 2003b); therefore, we recommend providing high nitrogen
(NO3�-N) fertilization rates of 2353–2942 mM d�1 in order to maintain high growth rates.
4.4. Phosphate
Surprisingly, moderate to high daily PO4� loads (125–500 mM d�1 supplied as
NaH2PO4) depressed SGR’s of P. mollis (Demetropoulos and Langdon, 2004a). Tissue P
concentrations were inversely correlated with P. mollis growth at PO4� fertilization loads
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–75 63
above 83 mM d�1. Tissue P concentrations >0.70% [dw] were positively correlated with
daily P loads and inversely correlated growth rates; thus, providing the correct level of P is
critical in producing high yields of P. mollis.
4.5. N:P and C:N
The optimal molar N:P ratios of nutrient additions for maximum P. mollis growth were
approximately 70 (2942 mM N d�1 and 42 mM P d�1) and 35 (2942 mM N d�1 and
83 mM P d�1) at low (23.68 mol photons m�2 d�1) and high (51.64 mol photons m�2 d�1)
light conditions, respectively (Demetropoulos and Langdon, 2004a). These nutrient
additions resulted in tissue N:P ratios of approximately 6.7 (4.6% N, 0.69% P [dw]) under
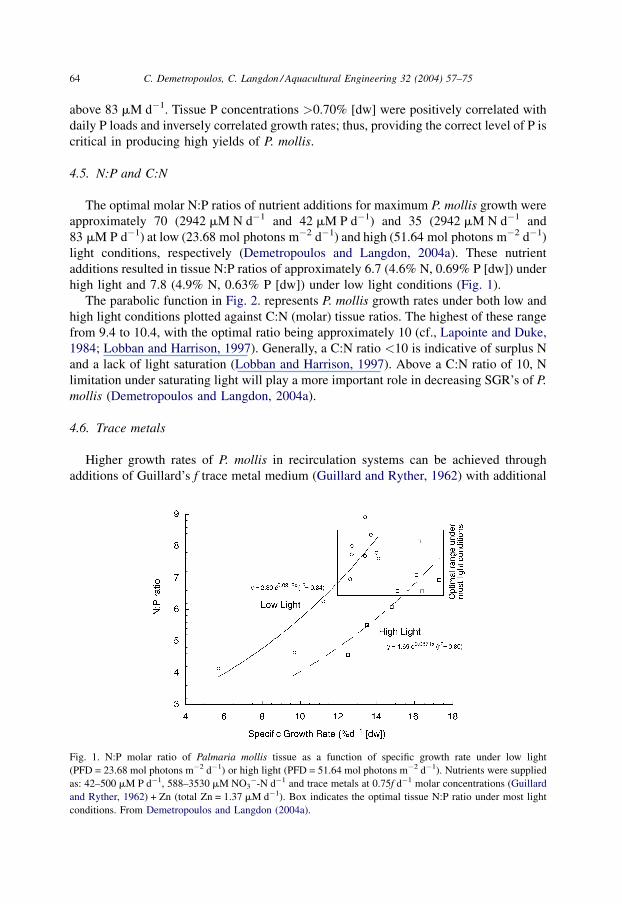
high light and 7.8 (4.9% N, 0.63% P [dw]) under low light conditions (Fig. 1).
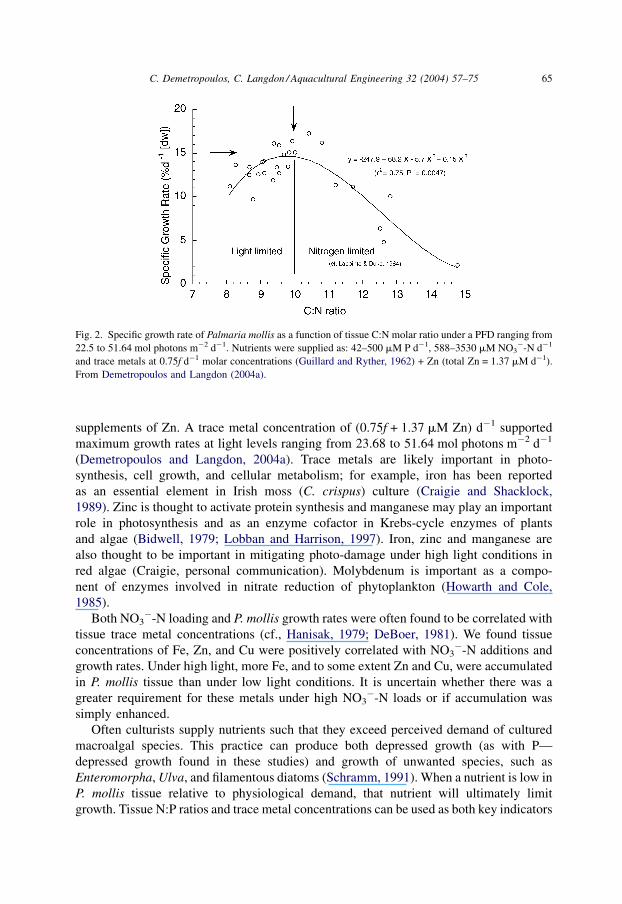
The parabolic function in Fig. 2. represents P. mollis growth rates under both low and
high light conditions plotted against C:N (molar) tissue ratios. The highest of these range
from 9.4 to 10.4, with the optimal ratio being approximately 10 (cf., Lapointe and Duke,
1984; Lobban and Harrison, 1997). Generally, a C:N ratio <10 is indicative of surplus N
and a lack of light saturation (Lobban and Harrison, 1997). Above a C:N ratio of 10, N
limitation under saturating light will play a more important role in decreasing SGR’s of P.
mollis (Demetropoulos and Langdon, 2004a).
4.6. Trace metals
Higher growth rates of P. mollis in recirculation systems can be achieved through
additions of Guillard’s f trace metal medium (Guillard and Ryther, 1962) with additional
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7564
Fig. 1. N:P molar ratio of Palmaria mollis tissue as a function of specific growth rate under low light
(PFD = 23.68 mol photons m�2 d�1) or high light (PFD = 51.64 mol photons m�2 d�1). Nutrients were supplied
as: 42–500 mM P d�1, 588–3530 mM NO3�-N d�1 and trace metals at 0.75f d�1 molar concentrations (Guillard
and Ryther, 1962) + Zn (total Zn = 1.37 mM d�1). Box indicates the optimal tissue N:P ratio under most light
conditions. From Demetropoulos and Langdon (2004a).
supplements of Zn. A trace metal concentration of (0.75f + 1.37 mM Zn) d�1 supported
maximum growth rates at light levels ranging from 23.68 to 51.64 mol photons m�2 d�1
(Demetropoulos and Langdon, 2004a). Trace metals are likely important in photo-
synthesis, cell growth, and cellular metabolism; for example, iron has been reported
as an essential element in Irish moss (C. crispus) culture (Craigie and Shacklock,
1989). Zinc is thought to activate protein synthesis and manganese may play an important
role in photosynthesis and as an enzyme cofactor in Krebs-cycle enzymes of plants
and algae (Bidwell, 1979; Lobban and Harrison, 1997). Iron, zinc and manganese are
also thought to be important in mitigating photo-damage under high light conditions in
red algae (Craigie, personal communication). Molybdenum is important as a compo-
nent of enzymes involved in nitrate reduction of phytoplankton (Howarth and Cole,
1985).
Both NO3�-N loading and P. mollis growth rates were often found to be correlated with
tissue trace metal concentrations (cf., Hanisak, 1979; DeBoer, 1981). We found tissue
concentrations of Fe, Zn, and Cu were positively correlated with NO3�-N additions and
growth rates. Under high light, more Fe, and to some extent Zn and Cu, were accumulated
in P. mollis tissue than under low light conditions. It is uncertain whether there was a
greater requirement for these metals under high NO3�-N loads or if accumulation was
simply enhanced.
Often culturists supply nutrients such that they exceed perceived demand of cultured
macroalgal species. This practice can produce both depressed growth (as with P—
depressed growth found in these studies) and growth of unwanted species, such as
Enteromorpha, Ulva, and filamentous diatoms (Schramm, 1991). When a nutrient is low in
P. mollis tissue relative to physiological demand, that nutrient will ultimately limit
growth. Tissue N:P ratios and trace metal concentrations can be used as both key indicators
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–75 65
Fig. 2. Specific growth rate of Palmaria mollis as a function of tissue C:N molar ratio under a PFD ranging from
22.5 to 51.64 mol photons m�2 d�1. Nutrients were supplied as: 42–500 mM P d�1, 588–3530 mM NO3�-N d�1
and trace metals at 0.75f d�1 molar concentrations (Guillard and Ryther, 1962) + Zn (total Zn = 1.37 mM d�1).
From Demetropoulos and Langdon (2004a).

of P. mollis health and by extension, yields as a function of ambient light (Hanisak, 1979;
DeBoer, 1981). Based on our results (Demetropoulos and Langdon, 2004a) tissue nutrient
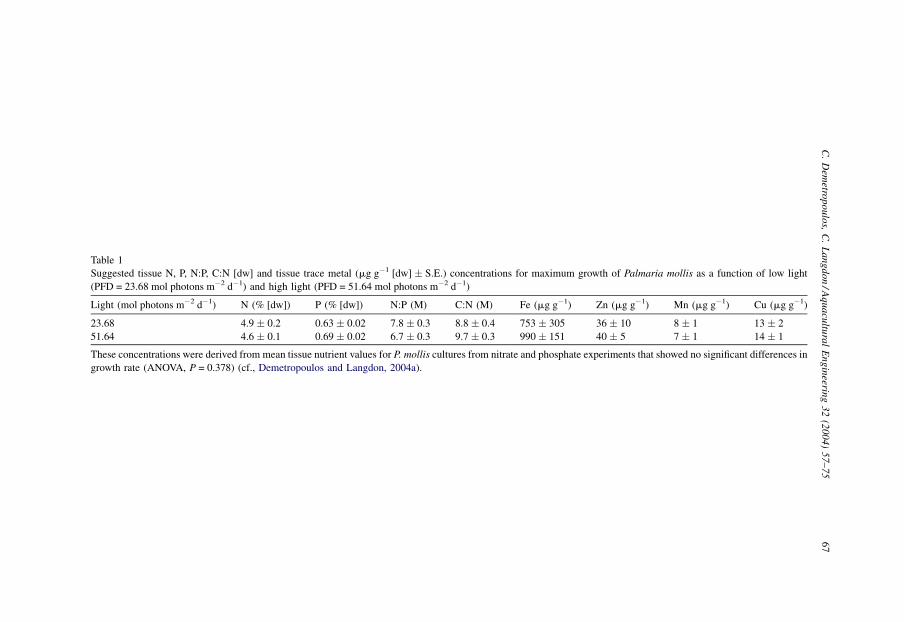
concentrations should be kept within the ranges set forth in Table 1.
4.7. Dissolved inorganic carbon sources and concentration
P. mollis cultures are often limited by lack of dissolved inorganic carbon (DIC).
Addition of DIC in the form of both CO2 and up to 6 mM HCO3� (the highest
concentration tested) increased P. mollis yields by approximately 53% compared to
cultures receiving no supplemental DIC at exchange rates of 3 vol d�1 (Demetropoulos and
Langdon, 2004c).
Like P. palmata, P. mollis probably utilizes CO2 directly as well as HCO3� indirectly
after dehydration to CO2 by extracellular carbonic anhydrase (CA) (cf., Robbins, 1978;
Johnston et al., 1992; Kubler and Raven, 1994). In studies conducted with P. palmata,
relative utilization of each DIC species varied with light intensity. Under low light P.
palmata was less likely to use HCO3� compared with CO2 (Kubler and Raven, 1994).
Under high light both HCO3� and CO2 were utilized, with HCO3
� being used
preferentially over CO2 (Bidwell and McLachlan, 1985; Johnston et al., 1992; Maberly
et al., 1992; Kubler and Raven, 1995).
4.8. Dissolved inorganic carbon and pH
Seawater pH can be used to predict the relative concentrations of CO2 and HCO3�
(Craigie and Shacklock, 1989). At an average pH of 8.2, nearly all of the DIC present in
seawater (>95%) is in the form of HCO3� (Falkowski and Raven, 1997). Further, Lyman
(1956) showed that as natural seawater pH increased from 8.0 to 8.9, the relative abundance
of CO2 was reduced by over 73% while the relative abundance of HCO3� decreased by
only 13%, with both species contributing to the relative abundance of CO32� and in the
extreme to the precipitation of CaCO3. At a pH of 9 or greater, free CO2 is generally
unavailable (Ryther and Debusk, 1982; Hanisak and Ryther, 1984), and most of the DIC
(>60%) is in the form of CO32�, which is unusable as a carbon source for macroalgae
(Reiskind et al., 1989). Under conditions of low CO2, high rates of carbon fixation in many
algae are maintained by increasing the activity of carbonic anhydrase (CA) such that
HCO3� is dehydrated to release CO2 and OH� ions, causing a rise in pH (Bidwell and
McLachlan, 1985; Haugland and Pedersen, 1992).
Under commercial stocking densities and light conditions in our experiments (2–
4 kg m�2, 23.68–51.64 mol photons m�2 d�1) we found a pH of 8.1 or less resulted in
highest growth rates of P. mollis (Demetropoulos and Langdon, 2004c). While growth rates
of P. mollis decreased by about 14% over pH values ranging from 8.1 to 8.9, there were no
significant differences in P. mollis growth rates between 8.3 and 8.9, indicating the range of
pH values (8.3–8.9) over which P. mollis growth can be economically cultured (Bruad and
Amat, 1996; Demetropoulos and Langdon, 2004c). P. mollis cultures experiencing a daily
pH of 9 or greater are likely dependent on both HCO3� and limited CO2 from exchange
water, culture aeration, and respiration at night. Thus, frequent pH values of 9.2 or greater
should be avoided to achieve the best growth for this species.
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7566
C.
Dem
etrop
ou
los,
C.
La
ng
do
n/A
qu
acu
ltura
lE
ng
ineerin
g3
2(2
00
4)
57
–7
56
7
Table 1
Suggested tissue N, P, N:P, C:N [dw] and tissue trace metal (mg g�1 [dw] � S.E.) concentrations for maximum growth of Palmaria mollis as a function of low light
(PFD = 23.68 mol photons m�2 d�1) and high light (PFD = 51.64 mol photons m�2 d�1)
Light (mol photons m�2 d�1) N (% [dw]) P (% [dw]) N:P (M) C:N (M) Fe (mg g�1) Zn (mg g�1) Mn (mg g�1) Cu (mg g�1)
23.68 4.9 � 0.2 0.63 � 0.02 7.8 � 0.3 8.8 � 0.4 753 � 305 36 � 10 8 � 1 13 � 2
51.64 4.6 � 0.1 0.69 � 0.02 6.7 � 0.3 9.7 � 0.3 990 � 151 40 � 5 7 � 1 14 � 1
These concentrations were derived from mean tissue nutrient values for P. mollis cultures from nitrate and phosphate experiments that showed no significant differences in
growth rate (ANOVA, P = 0.378) (cf., Demetropoulos and Langdon, 2004a).

5. Abalone growth experiments
Laboratory studies with H. discus hannai and Haliotis rufescens were initially
conducted at the Hatfield Marine Science Center (HMSC), Oregon State University, and
scale-up was extended to Big Island Abalone Corporation (BIAC), at the Natural Energy
Laboratory of Hawaii (NELH). Co-culture techniques were then used in a project for the
restoration of federally endangered H. sorenseni, held at Channel Islands Marine Resource
Institute, Port Heuneme, California, in conjunction with Pacific Aquaculture Research &
Technology Institute and California Lutheran University, Thousand Oaks, California.
5.1. Nutrient bioassay experiment
Abalone bioassays with H. discus hannai were used to determine the direct effects of
nutrients used to fertilize P. mollis cultures, on abalone survival and growth
(Demetropoulos and Langdon, in press). Nutrient additions of N (0–2942 mM d�1), P
(0–83 mM d�1), and trace metals (0–0.75f + 1.37 mM Zn d�1) delivered to cultures had no
observable negative effects (P > 0.117) on H. discus hannai survival, linear shell growth
rates (LGR as mm d�1) or specific growth rates (SGR as % d�1) (cf., Flemming, 1995a,b).
LGR ¼ Lf � Li
t(3)
SGR ¼ 100 � ln Wf � ln Wi
t
� �(4)
On the contrary, abalone growth rates tended to increase in treatments where nutrients were
added. This may have been due to abalone grazing on enhanced diatom growth due to the
presence of added nutrients (Austin et al., 1990; Demetropoulos, unpublished data).
5.2. Growth experiments with H. discus hannai, H. rufescens, and H. sorenseni
P. mollis diets differing in biochemical composition were produced by supplying
adequate PO4� and varying NO3
� loads in the presence or absence of trace metals. PO4�
was fixed at a load of 83.3 mM d�1 (as NaH2PO4). Nitrate was supplied at loads of 1176,
1765, 2353 or 2942 mM d�1 (as NaNO3) with or without addition of trace metals supplied
as Guillard and Ryther’s (1962) f medium in the form of 0.75f + 1.37 mM Zn d�1
(Demetropoulos and Langdon, 2004a). P. mollis diets were first evaluated by feeding them
to H. discus hannai and H. rufescens and measuring the effect on LGR and SGR. The best
P. mollis diet was subsequently evaluated as a rearing diet for juvenile H. sorenseni.
Growth rates of these three species are summarized in Fig. 3.
Highest shell growth rates for H. discus hannai were obtained with P. mollis diets cultured
under NO3� loads of 1176–1765 mM NO3
� d�1 [linear growth rate (LGR) = 140.5 mm d�1
for abalone fed on 1176 NO3� load + trace metals] and for H. rufescens highest growth rates
were obtained with NO3� loads of 2353–2942 mM NO3
� d�1 (LGR = 198 � 2.7 mm d�1
for abalone fed on 2942 [NO3� load] + trace metals; Fig. 3; Demetropoulos and Langdon,
in press). Abalone shell lengths under these treatments ranged from 25.1 to 34.5 mm for
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7568
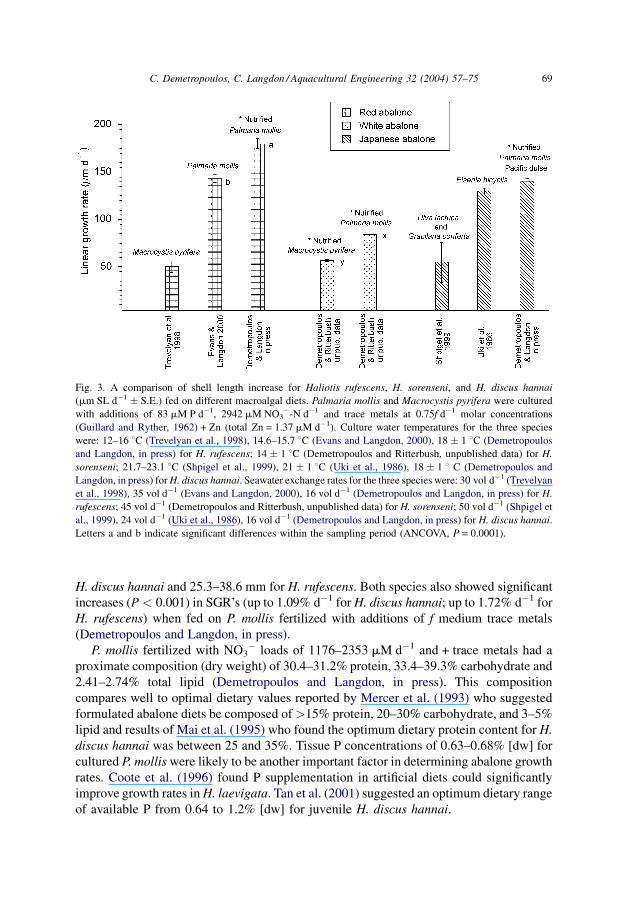
H. discus hannai and 25.3–38.6 mm for H. rufescens. Both species also showed significant
increases (P < 0.001) in SGR’s (up to 1.09% d�1 for H. discus hannai; up to 1.72% d�1 for
H. rufescens) when fed on P. mollis fertilized with additions of f medium trace metals
(Demetropoulos and Langdon, in press).
P. mollis fertilized with NO3� loads of 1176–2353 mM d�1 and + trace metals had a
proximate composition (dry weight) of 30.4–31.2% protein, 33.4–39.3% carbohydrate and
2.41–2.74% total lipid (Demetropoulos and Langdon, in press). This composition
compares well to optimal dietary values reported by Mercer et al. (1993) who suggested
formulated abalone diets be composed of >15% protein, 20–30% carbohydrate, and 3–5%
lipid and results of Mai et al. (1995) who found the optimum dietary protein content for H.
discus hannai was between 25 and 35%. Tissue P concentrations of 0.63–0.68% [dw] for
cultured P. mollis were likely to be another important factor in determining abalone growth
rates. Coote et al. (1996) found P supplementation in artificial diets could significantly
improve growth rates in H. laevigata. Tan et al. (2001) suggested an optimum dietary range
of available P from 0.64 to 1.2% [dw] for juvenile H. discus hannai.
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–75 69
Fig. 3. A comparison of shell length increase for Haliotis rufescens, H. sorenseni, and H. discus hannai
(mm SL d�1 � S.E.) fed on different macroalgal diets. Palmaria mollis and Macrocystis pyrifera were cultured
with additions of 83 mM P d�1, 2942 mM NO3�-N d�1 and trace metals at 0.75f d�1 molar concentrations
(Guillard and Ryther, 1962) + Zn (total Zn = 1.37 mM d�1). Culture water temperatures for the three species
were: 12–16 8C (Trevelyan et al., 1998), 14.6–15.7 8C (Evans and Langdon, 2000), 18 � 1 8C (Demetropoulos
and Langdon, in press) for H. rufescens; 14 � 1 8C (Demetropoulos and Ritterbush, unpublished data) for H.
sorenseni; 21.7–23.1 8C (Shpigel et al., 1999), 21 � 1 8C (Uki et al., 1986), 18 � 1 8 C (Demetropoulos and
Langdon, in press) for H. discus hannai. Seawater exchange rates for the three species were: 30 vol d�1 (Trevelyan
et al., 1998), 35 vol d�1 (Evans and Langdon, 2000), 16 vol d�1 (Demetropoulos and Langdon, in press) for H.
rufescens; 45 vol d�1 (Demetropoulos and Ritterbush, unpublished data) for H. sorenseni; 50 vol d�1 (Shpigel et
al., 1999), 24 vol d�1 (Uki et al., 1986), 16 vol d�1 (Demetropoulos and Langdon, in press) for H. discus hannai.
Letters a and b indicate significant differences within the sampling period (ANCOVA, P = 0.0001).

While trace metal additions to P. mollis cultures significantly increased its protein
content (from 26.1 � 2.8% to 30.3 � 4.1% [dw]; P = 0.0063), it is unlikely that this in
itself produced significantly higher growth rates for H. discus hannai and H. rufescens.
Increased abalone growth was likely due to the abalone’s direct use of trace metals
accumulated in P. mollis tissue or more optimal concentrations of other, unmeasured
dietary biochemical constituents, such as vitamins or specific fatty acids. Mai and Tan
(2000) and Tan and Mai (2001) showed growth rates and LGR’s for H. discus hannai were
improved with artificial diets supplemented with Fe and Zn and they suggested a minimum
dietary requirement of 65–70 mg g�1 Fe and 16–35 mg g�1 Zn when culturing juvenile H.
discus hannai (cf. Mai et al., 2003). P. mollis fertilized with 2353 to 2942 mM NO3-N d�1
and (0.75f + 1.37 mM Zn) trace metals contained 296–1220 mg g�1 Fe and 18–
49 mg g�1 Zn (Demetropoulos and Langdon, 2004a, in press) that should have fully
met the abalone’s requirements for these trace metals according to Mai et al. (2003).
Shell growth rates for H. discus hannai maintained at 18 � 1 8C and supplied with the
most nutritive P. mollis diets were high compared to those reviewed by Flemming et al.
(1996) for various abalone species fed on different artificial diets at temperatures ranging
from 18 to 22 8C. They reported LGR’s of 50–138 mm d�1 for a variety of species
including H. discus hannai compared to LGR’s of 94.8–140.5 mm d�1 for the same sized
animals in this study (Demetropoulos and Langdon, in press). Further, at a cultivation
temperature of 18 � 1 8C a mean LGR of 139 mm d�1 for H. discus hannai fed on the most
nutritive P. mollis diet was comparable to the highest values reported by Uki et al. (1986)
for abalone fed Eisenia bicyclis at a more optimal temperature of 21 � 1 8C (Fig. 3).
Shell growth rates of H. rufescens typically range from 33 to 50 mm d�1 when abalone
are fed on the giant kelp, M. pyrifera at temperatures between 12 and 16 8C (Trevelyan
et al., 1998) (Fig. 3). Evans and Langdon (2000) showed H. rufescens LGR’s could be as
high as 123 mm d�1 on diets of P. mollis fertilized with nitrate and phosphate (Fig. 3). In
our recent study, LGR’s were as high as 198 mm d�1 during the first 67 days and averaged
171 mm d�1 for the entire 100-day experiment (Demetropoulos and Langdon, in press).
Higher LGRs were likely due to higher seawater temperatures (18 � 1 8C) than those of
Evans and Langdon (2000) (15 � 1 8C) coupled with the addition of 0.75f + 1.37 mM Zn
trace metals to the P. mollis cultures (Fig. 3).
H. rufescens fed on dulse in our experiments reached sexual maturity as small as 27 mm
in shell length. Ault (1985) reported a minimum size of 50 mm for sexual maturity of H.
rufescens males and Giorgi and DeMartini (1977) reported a minimum size of 39.5 mm for
females. Further, Demetropoulos and Langdon (in preparation) found P. mollis enhanced
the onset of gonad development in H. rufescens compared to other macroalgal and artificial
feeds fed to the species under the same culture conditions. Aspects of the culture system
itself, including use of P. mollis as a diet, probably led to H. rufescens to becoming sexually
mature at a smaller size (cf., Ault, 1982; Leighton, 2000). Future investigations should look
at testing mixed diets (e.g., Palmaria sp., Egregia sp., Laminaria sp., Nereocystis
luetkeana, Macrocystis sp., and artificial feeds) and temperature as ways to manipulate the
strength and timing of gametogenesis.
In subsequent studies, P. mollis and commercially harvested M. pyrifera were fertilized
with 83 mM P, 2353 mM NO3�-N and 0.75f + 1.37 mM Zn trace metals and fed to juvenile
white abalone, H. sorenseni maintained at 14 � 1 8C to compare growth rates. Growth
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7570

rates produced by these diets were significantly different from one another (P = <0.0001).
LGR’s of H. sorenseni (SL = 35.7–46.5 mm) averaged 84.14 m d�1 and SGR’s averaged
0.89% d�1 when fed on P. mollis, while LGR’s averaged only 56.67 m d�1 and SGR’s
averaged only 0.62% d�1 with nutrient supplemented M. pyrifera (Fig. 3) (Demetropoulos
and Ritterbush, unpublished data). Over a 147-day experiment, H. sorenseni survival rates
were higher when fed on a diet of P. mollis (survival = 100%) compared to a diet of M.
pyrifera (survival = 93%) and, as was the case with H. rufescens, P. mollis accelerated the
onset of gonad development in H. sorenseni compared to M. pyrifera. Furthermore, as with
previous results for H. discus hannai and H. rufescens, P. mollis produced a mottled brick
red shell color characteristic of cryptic color patterns found in wild H. sorenseni, whereas
M. pyrifera produced a green coloration (Demetropoulos and Langdon, in press). In
conjunction with the advantages of co-culture outline above, these combined results make
P. mollis highly valuable to H. sorenseni restoration efforts now under way.
6. Conclusions
Partial or complete recirculation systems for co-culture of Pacific dulse and abalone
offer many advantages compared with flow-through systems. By providing optimum
conditions for Pacific dulse growth, culture operations can ensure a reliable source of
nutritious food for abalone while simultaneously maximizing biofilter functions and
decreasing discharge of nutrients into the environment. Reduced seawater exchange rates
coupled with nutrient additions are likely to be a more favorable economic strategy than
pumping large volumes of seawater through a facility to provide sufficient nutrients for
high Pacific dulse productivity. The complementary strategy of adding nutrients during the
dark cycle every 5–7 days has several advantages: (1) at night, water can be shut off so that
nutrients are not lost due to flushing yet culture temperatures remain relatively constant, (2)
costs associated with nutrient additions are reduced because nutrient uptake is more
efficient, (3) epiphyte growth is better controlled, and (4) chlorophyte species are not
provided with light necessary for their nutrient uptake (cf., Hanisak, 1987; Lobban and
Harrison, 1997). Further, decreasing the volume of seawater pumped through a facility will
avoid sustained exposure to abalone pathogens present in coastal waters.
Often there is a tendency for the seaweed culturist to supply nutrients such that they
exceed demand. Unfortunately, this can easily produce both depressed growth (e.g.,
phosphate depressed growth as shown here) and blooms of weed species such as Ulva,
Enteromorpha, and filamentous diatoms (Schramm, 1991). It is far better to supply
nutrients on a schedule that takes advantage of the demand for nutrients based on growth.
Tissue nutrient assays, similar to those used by terrestrial farmers, provide a reliable
indicator of macroalgal health and potential yields as a function of incident light (Hanisak,
1979, 1982; DeBoer, 1981). Tissue analysis should be adopted as a standard method for
determining the nutritional status of P. mollis. Moreover, tissue nutrient status should be
tracked closely since nutrient demand will vary as a function of culture conditions such as
temperature and especially that of incident light.
Results of these studies suggest daily nutrient concentrations that should be applied to P.
mollis cultures to produce abalone feed of high nutritional value are approximately 83 mM
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–75 71

P, 1765–2353 mM NO3�-N, and 0.75f + 1.37 mM Zn trace metals for H. discus hannai and
83 mM P, 2353–2942 mM NO3�-N, and 0.75f + 1.37 mM Zn trace metals for H. rufescens
and H. sorenseni, under natural light conditions ranging from 20 to
55 mol photons m�2 d�1.
Acknowledgements
Thanks to Amgen Inc. for valuable donations of equipment that were central to the white
abalone work and to California Lutheran University for supplying a number of eager
undergraduate students to help in the data collection. The white abalone research was
conducted at the Channel Islands Marine Resource Institute and funded by the Pacific
Aquaculture Research and Technology Institute, which promote hands on training of
undergraduate students in science.
References
Ault, J., 1982. Aspects of laboratory reproduction and growth of the red abalone, Haliotis rufescens Swainson.
M.S. Thesis. Humboldt State University, Arcata, CA, p. 77.
Ault, J., 1985. Some quantitative aspects of reproduction and growth of the red abalone, Haliotis rufescens
Swainson. J. World Maricult. Soc. 16, 398–425.
Austin, A.P., Ridley-Thomas, C.I., Lucey, W.P., Austin, J.D., 1990. Effects of nutrient enrichment on marine
periphyton: implications for abalone culture. Bot. Mar. 33, 235–239.
Bidwell, R.G.S., 1979. Plant Physiology, second ed. Macmillan, New York, 311 pp.
Bidwell, R.G.S., McLachlan, J., 1985. Carbon nutrition of seaweeds: photosynthesis, respiration and photo-
respiration. J. Exp. Mar. Biol. Ecol. 86, 15–46.
Bidwell, R.G.S., McLachlan, J., Lloyd, N.D.H., 1985. Tank cultivation of Irish Moss Chondrus crispus Stackh.
Bot. Mar. 28, 87–97.
Bird, K.T., Sanchez, F.X., 1990. Paddle wheel raceway cultivation of Gracilaria verrucosa. J. Phycol. Suppl. 26, 13.
Bruad, J.-P., Amat, M.A., 1996. Chondrus crispus (Gigartinaceae, Rhodophyta) tank cultivation: optimizing
carbon input by fixed pH and use of a salt water well. Hydrobiology 326–327, 335–340.
Coote, T.A., Hone, P.W., Kenyon, R., Maguire, G.B., 1996. The effect of different combinations of dietary calcium
and phosphorus on the growth of juvenile Haliotis laevigata. Aquaculture 145, 267–279.
Craigie, J.S., Shacklock, P.F., 1989. Culture of Irish moss. In: Boghen, A. (Ed.), Cold Water Aquaculture in
Atlantic Canada. Canadian Institute for Research on Regional Development, Moncton, Canada, pp. 243–270.
Davison, I.R., 1991. Environmental effects on algal photosynthesis: temperature. J. Phycol. 27, 2–8.
Day, R.W., Fleming, A.E., 1992. The determinants and measurement of abalone growth. In: Shepard, S., Tegner,
M., Guzman del Proo, S. (Eds.),Abalone of the World, Biology, Fisheries, and Culture. Fishing News Books,
Oxford, London, Edinburgh, Cambridge, Victoria, pp. 141–169.
DeBoer, J.A., 1981. Nutrients. In: Lobban, C., Wynne, M. (Eds.), The Biology of Seaweeds. Blackwell Scientific,
Oxford, pp. 356–391.
Demetropoulos, C.L., Langdon, C.J., Co-inventors 2001. Palmaria algal strains and methods for their use. U.S.
Patent 6258588.
Demetropoulos, C.L., Langdon, C.J., 2004a. Enhanced production of Pacific dulse (Palmaria mollis) for co-
culture with abalone in a land-based system: nitrogen, phosphorus, and trace metal nutrition. Aquaculture 235,
433–455.
Demetropoulos, C.L., Langdon, C.J., 2004b. Enhanced production of Pacific dulse (Palmaria mollis) for co-
culture with abalone in a land-based system: effects of stocking density, light, salinity, and temperature.
Aquaculture 235, 471–488.
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7572

Demetropoulos, C.L., Langdon, C.J., 2004c. Enhanced production of Pacific dulse (Palmaria mollis) for co-
culture with abalone in a land-based system: effects of seawater exchange, pH, and inorganic carbon
concentration on growth. Aquaculture 235, 457–470.
Demetropoulos, C.L., Langdon, C.J., in press. Enhanced production of Pacific dulse (Palmaria mollis) for co-
culture with abalone in a land-based system: effects of algal nutrient enrichment and biochemical composition
on abalone growth and condition. J. Exp. Mar. Biol. Ecol.
Evans, F., Langdon, C.J., 2000. Co-culture of dulse Palmaria mollis and red abalone Haliotis rufescens under
limited flow conditions. Aquaculture 185, 137–158.
Evans, F., Langdon, C.J., Demetropoulos, C.L. 2000. An environmentally-sustainable, integrated, co-culture
system for dulse and abalone production. Aquacultural Eng. (this volume).
Falkowski, P.G., Raven, J.A., 1997. Aquatic Photosynthesis. Blackwell Science Inc., Malden, MA, 375 pp.
Flemming, A.E., 1995a. Growth, intake, feed conversion efficiency and chemosensory preference of the
Australian abalone, Haliotis rubra. Aquaculture 132, 297–311.
Flemming, A.E., 1995b. Digestive efficiency of the Australian abalone Haliotis rubra in relation to growth and
feed preference. Aquaculture 134, 279–293.
Flemming, A.E., Van Barnard, R.J., Hone, P.W., 1996. The development of artificial diets for abalone: a review and
future directions. Aquaculture 140, 5–53.
Friedlander, M., 1992. Gracilaria conferta and its epiphytes. The effect of outdoor culture conditions on growth.
Bot. Mar. 35, 423–428.
Friedlander, M., Ben-Amotz, A., 1991. The effect of outdoor culture conditions on growth and epiphytes of
Gracilaria conferta. Aquat. Bot. 39, 315–333.
Friedlander, M., Krom, M.D., Ben-Amotz, A., 1991. The effect of light and ammonium on growth, epiphytes and
chemical constituents of Gracilaria conferta in outdoor cultures. Bot. Mar. 34, 161–166.
Friedlander, M., Levy, I., 1995. Cultivation of Gracilaria in outdoor tanks and ponds. J. Appl. Phycol. 7, 315–
324.
Friedman, C.S., Andree, K.B., Beauchamp, K.A., Moore, J.D., Robbins, T.T., Shields, J.D., Hedrick, R.P., 2000.
’Candidatus Xenohaliotis californiensis’, a newly described pathogen of abalone, Haliotis spp., along the west
coast of North America. Int. J. Syst. Evol. Microbiol. 50 (2), 847–855.
Gardner, G.R., Harshbarger, J.C., Lake, J.L., Sawyer, T.K., Price, K.I., Stephenson, M.D., Haaker, P.L., Togstad,
H.A., 1995. Association of Prokaryotes with symptomatic appearance of withering syndrome in Black
Abalone Haliotis cracherodii. J. Inv. Pathol. 66, 111–120.
Giorgi, A.E., DeMartini, J.D., 1977. A study of the reproductive biology of the red abalone near Mendocino, CA.
Calif. Fish Game 63 (2), 80–94.
Gonen, Y., Kimmel, E., Friedlander, M., 1993. Effect of relative water motion on photosynthetic rate of the red
alga Gracilaria conferta. Hydrobiologia 260–261, 493–498.
Guillard, R.R.L., Ryther, J.H., 1962. Studies on marine plankton diatoms I. Cyclothella nana Huntedt and
Detonula confercacae (Cleve). Gran. Can. J. Microbiol. 8, 229–239.
Hanisak, M.D., 1979. Nitrogen limitation of Codium fragile ssp. tomentosoides as determined by tissue analysis.
Mar. Biol. 5, 333–337.
Hanisak, M.D., 1982. The nitrogen status of Gracilaria tikvahiae in natural and mariculture systems as measured
by tissue analysis. Int. Phycol. Congress Abstracts 1, 20.
Hanisak, M.D., 1987. Cultivation of Gracilaria and other macroalgae in Florida for energy production. In: Bird,
K., Benson, P. (Eds.), Seaweed Cultivation for Renewable Resources. Elsevier, Amsterdam, pp. 191–218.
Hanisak, M.D., 1990. The use of Gracilaria tikvahiae (Gracilariales Rhodophyta) as a model system to understand
the nitrogen limitation of cultured seaweeds. Hydrobiologia 204–205, 79–87.
Hanisak, M.D., Ryther, J.H., 1984. Cultivation biology of Gracilaria tikvahiae in the United States. Proc. Int.
Seaweed Symp. 11, 295–298.
Hansen, J., 1984. Strain selection and physiology in the development of Gracilaria mariculture. In: Bird, C.J.,
Ragan, M.A. (Eds.), Eleventh International Seaweed Symposium. Developments in Hydrobiology, vol. 22. Dr
W. Junk Publishers, Dordrecht, pp. 89–94 (Reprinted from Hydrobiologia 116/117).
Haugland, K., Pedersen, M., 1992. Growth of the red alga Gracilaria tenuistipitata at high pH. Influence of
some environmental factors and correlation to an increased carbonic-anhydrase activity. Bot. Mar. 35,
579–587.
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–75 73

Howarth, R.W., Cole, J.J., 1985. Molybdenum availability, nitrogen limitation and phytoplankton growth in
natural waters. Science 229, 653–655.
Huguenin, J.E., 1976. An examination of problems and potentials for future large-scale intensive seaweed culture
systems. Aquaculture 9, 313–342.
Iwasaki, H., 1967. Nutritional studies of the edible seaweed Porphyra tenera. II. Nutrition of conchocelis. J.
Phycol. 3, 30–34.
Johnston, A.M., Maberly, S.C., Raven, J.A., 1992. The acquisition of inorganic carbon by four red macroalgae.
Oecologia 92, 317–326.
Kubler, J.E., Raven, J.A., 1994. Consequences of light limitation for carbon acquisition in three Rhodophytes.
Mar. Prog. Ser. 110 (2–3), 203–209.
Kubler, J.E., Raven, J.A., 1995. The interaction between inorganic carbon acquisition and light supply in Palmaria
palmata (Rhodophyta). J. Phycol. 31, 369–375.
Kubler, J.E., Raven, J.A., 1996a. Nonequilibrium rates of photosynthesis and respiration under dynamic light
supply. J. Phycol. 32, 963–969.
Kubler, J.E., Raven, J.A., 1996b. Inorganic carbon acquisition by red seaweeds grown under dynamic light
regimes. Hydrobiolia 326–327, 401–406.
Lapointe, B.E., Duke, C.S., 1984. Biochemical strategies for growth for Gracilaria tikvahiae (Rhodophyceae) in
relation to light intensity and nitrogen availability. J. Phycol. 20, 465–488.
Lapointe, B.E., Ryther, J.H., 1978. Some aspects of the growth and yield of Gracilaria tikvahiae in culture.
Aquaculture 15, 185–193.
Lapointe, B.E., Ryther, J.H., 1979. The effects of nitrogen and seawater flow rate on the growth and biochemical
composition of Gracilaria foliifera v. angustissima in mass outdoor cultures. Bot. Mar. 22, 529–537.
Leighton, D.L., 2000. The Biology and Culture of the California Abalones. Dorrance Publishing Co Inc.,
Pittsburgh, pp. 64–75.
Lobban, C.S., Harrison, P.J., 1997. Seaweed Ecology and Physiology. Cambridge University Press, Cambridge,
NJ, 366 pp.
Lyman, J., 1956. Buffer mechanism of seawater. Ph.D. thesis. University of California, Los Angeles, CA, 317 pp.
Maberly, S., Johnston, A.M., Raven, J.A., 1992. Discrimination between 13C and 12C by marine plants. Oecologia
91, 481–492.
Maestrini, S.Y., Robert, J.M., Truquet, I., 1982. Simultaneous uptake of ammonium and nitrate by oyster-pond
algae. Mar. Biol. Lett. 3 (3), 143–153.
Mai, K., Mercer, J., Donlon, P.J., 1995. Comparative studies on the nutrition of two species of abalone, Haliotis
tuberculata L. and Haliotis discus hannai Ino. III. Response of abalone to various levels of dietary lipid.
Aquaculture 134, 65–80.
Mai, K., Tan, B., 2000. Iron methionine (FeMet) and iron sulfate (FeSO4) as sources of dietary iron for juvenile
abalone, Haliotis discus hannai Ino. J. Shellfish Res. 19 (2), 861–868.
Mai, K., Zhang, W., Tan, B., He, G., 2003. Effects of dietary zinc on the shell biomineralization in abalone Haliotis
discus hannai Ino. J. Exp. Mar. Biol. Ecol. 283 (1–2), 51–62.
Mathieson, A.C., Norall, T.L., 1975. Photosynthetic studies of Chondrus crispus. Mar. Biol. 33, 207–213.
McLachlan, J., 1991. General principles of onshore cultivation of seaweeds: effects of light on production.
Hydrobiologia 221, 125–135.
Mercer, J.P., Mai, K.S., Donlon, J., 1993. Comparative studies on the nutrition of two species of abalone, Haliotis
tuberculata and Haliotis discus hannai Ino: I. Effects of algal diets on growth and biochemical composition.
Invertebr. Reprod. Dev. 23, 75–88.
Moore, J.D., Robbins, T.T., Friedman, C.S., 2000. Withering syndrome in farmed red abalone, Haliotis rufescens:
Thermal induction and association with a gastrointestinal Rickettsiales-like prokaryote. J. Aquat. Anim.
Health 12, 26–34.
Morgan, K.C., Shacklock, P.F., Simpson, F.J., 1980. Some aspects of the culture of Palmaria palmata in
greenhouse tanks. Bot. Mar. 23, 765–770.
Morgan, K.C., Simpson, F.J., 1981. The cultivation of Palmaria palmate. Effect of high concentrations of nitrate
and ammonium on growth and nitrogen uptake. Aquat. Bot. 11, 167–171.
Neish, A.C., Shacklock, P.F., Fox, C.H., Simpson, F.J., 1977. The cultivation of Chondrus crispus: factors
affecting growth under greenhouse conditions. Can. J. Bot. 55, 2263–2271.
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–7574

Neori, A., Krom, M.D., Ellner, S.P., Boyd, C.E., Popper, D., Rabinovitch, R., Davidson, P., Dvir, O., Zuber, D.,
Ucko, M., Angel, D., Gordin, H., 1996. Seaweed biofilters as regulators of water quality in integrated fish-
seaweed culture units. Aquaculture 141, 183–199.
Neori, A., Shpigel, M., Ben-Ezra, D., 2000. A sustainable integrated system for culture of fish, seaweed, and
abalone. Aquaculture 186, 279–291.
Pickering, T.D., Gordon, M.E., Tong, L.J., 1993. Effect of nutrient pulse concentration and frequency on growth of
Gracilaria chilensis plants and levels of epiphytic algae. J. Appl. Phycol. 5, 525–533.
Reiskind, J.B., Meer, S., Bowes, G., 1989. Photosynthesis, photorespiration, and ecophysiological interactions in
marine macroalgae. Aquat. Bot. 34, 131–152.
Rivero, L.E., Viana, M.T., 1996. Effects of pH, water stability and toughness of artificial diets on the palatability
for juvenile abalone Haliotis fulgens. Aquaculture 144, 353–362.
Robbins, J.V., 1978. Effects of physical and chemical factors on photosynthetic and respirator rates of Palmaria
palmata (Florideaophyceae). In: Jensen, A., Stein, J. (Eds.), Proceedings of the 9th International Seaweed
Symposium, Santa Barbara, California, Science Press, Cambridge, NJ, pp. 273–283.
Ryther, J.H., Debusk, T.A., 25–29 January 1982. Significance of carbon-dioxide and bicarbonate-carbon uptake in
marine biomass production. In: Proceedings of the IGT Symposium on Energy from Biomass and Wastes, vol.
IV, Lake Buena Vista, Florida.
Schramm, W., 1991. Cultivation of unattached seaweeds. In: Guiry, M., Blunden, H. (Eds.), Seaweed Resources
in Europe, Uses and Potential. John Wiley and Sons, Chichester, pp. 379–408.
Shpigel, M., Neori, A., 1996. The integrated culture of seaweed, abalone, fish, and clams in modular intensive
land-based systems: I. Proportions of size and projected revenues. Aquaculture Eng. 15, 313–326.
Shpigel, M., Ragg, N.L., Lupatsch, I., Neori, A., 1999. Protein content determines the nutritional value of the
seaweed Ulva lactuca L. for the abalone Haliotis tuberculata L. and H. discus hannai Ino. J. Shellfish Res. 18
(1), 227–233.
Tan, B., Mai, K., Liufu, Z., 2001. Response of juvenile abalone, Haliotis discus hannai, to dietary calcium,
phosphorus, and calcium/phosphorus ratio. Aquaculture 198 (1–2), 141–158.
Tan, B., Mai, K., 2001. Zinc methionine and zinc sulfate as sources of dietary zinc for juvenile abalone, Haliotis
discus hannai Ino. Aquaculture 192 (1), 141–158.
Terry, K.L., 1986. Photosynthesis in modulated light: quantitative dependence on photosynthetic enhancement on
flashing rate. Biotech. Bioeng. 28, 988–995.
Thomas, T.E., Harrison, P.J., 1985. Effects of nitrogen supply on nitrogen uptake, accumulation and assimilation
in Porphyra perforata (Rhodophyta). Mar. Biol. 85, 269–278.
Trevelyan, G.A., Mendoza, J.L., Buckley, B., 1998. Increasing the yield of red abalone with the alga, Microcladia
coulteri. J. Shellfish Res. 17, 631–633.
Ugarte, R., Santelices, B., 1992. Experimental tank cultivation of Gracilaria chilensis in central Chile.
Aquaculture 101, 7–16.
Uki, N., Sugiura, M., Watanabe, T., 1986. Dietary value of seaweeds occurring on the Pacific coast of Tohoku for
growth of the abalone Haliotis discus hannai. Bull. Jpn. Soc. Sci. Fish 52 (2), 257–266.
Wheeler, W., 1980. The effect of boundary layer transport on the fixation of carbon by the giant kelp, Macrocystis
pyrifera (L.) C. Agric. Mar. Biol. 56, 103–110.
C. Demetropoulos, C. Langdon / Aquacultural Engineering 32 (2004) 57–75 75