development of the radula and digestive system of juvenile blacklip abalone ( haliotis rubra):...
TRANSCRIPT

www.elsevier.com/locate/aqua-online
Aquaculture 250 (
Development of the radula and digestive system of juvenile
blacklip abalone (Haliotis rubra): Potential factors responsible for
variable weaning success on artificial diets
Danielle JohnstonT, Natalie Moltschaniwskyj, Jarrod Wells
School of Aquaculture, Tasmanian Aquaculture and Fisheries Institute, University of Tasmania, Locked Bag 1370, Launceston,
Tasmania 7250, Australia
Received 7 December 2004; received in revised form 9 March 2005; accepted 9 March 2005
Abstract
We investigated the structural and physiological changes in the radula and digestive system in juvenile blacklip abalone
Haliotis rubra, between 80 and 158 days post settlement (PS) to determine if variable growth on artificial diets (and prevalence
of runts) are due to an inability to efficiently ingest and digest the diet. Between 80 and 102 days PS, L5 teeth appeared on the
radula and there were fewer lateral serrations, consistent with the adult form of the animal, suggesting that this development is
in preparation for feeding on macroalgae. Digestive gland complexity (tubule number and density) increased between 80 and
102 days PS and is consistent with greater enzyme production and increased digestive efficiency. Of the enzymes studied,
laminarinase and lipase exhibited the highest activities in animals fed a diatom diet, both significantly increasing with age of the
abalone. High laminarinase activities reflect higher utilisation of the algal polysaccharide chrysolaminarin in the diatom diet.
Ingestion of artificial diet had no adverse effects on the morphological development of the digestive system, but trypsin activity
in abalone fed the artificial diet was significantly higher than diatom-fed abalone of similar age, indicative of higher levels of
protein in the artificial diet. Similarly, lipase activity was significantly lower in abalone fed the artificial diet and may reflect an
inability to digest the fish oil component, which is not found in their natural diet. Future development of artificial diets,
especially for juvenile abalone, should focus on the levels and type of lipid provided. Runt abalone (i.e. under-developed
compared to their siblings) had radulae similar to much younger 80 days PS abalone and digestive tissue degradation at 137
days PS is evidence that runts have limited ability to ingest food and are nutritionally compromised.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Abalone; Weaning; Artificial diet; Digestive enzymes; Radula; Digestive system; Aquaculture
1. Introduction
0044-8486/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.aquaculture.2005.03.012
T Corresponding author. WA Marine Research Laboratories, P.O.
Box 20, North Beach, Western Australia 6020, Australia. Tel.: +61
8 92468460; fax: +61 8 94473062.
E-mail address: [email protected] (D. Johnston).
Australian abalone aquaculture is an expanding
industry focused on the commercially important
blacklip (Haliotis rubra Leach) and greenlip (Haliotis
2005) 341–355

D. Johnston et al. / Aquaculture 250 (2005) 341–355342
laevigata Donovan) species (Shepherd and Hearn,
1983; Hahn, 1989b; Dunstan et al., 1996). Abalone
growers culture naturally occurring diatoms on settle-
ment plates as diets for newly settled juveniles,
however, maintaining sufficient diatom numbers as
the juveniles grow is a major obstacle in production
(Ebert and Houk, 1984; Hahn, 1989a; Knauer et al.,
1996). As juveniles increase in size and are ready to
switch from diatoms to macroalgae farmers have two
choices; provide natural macroalgae or use an
artificial diet (Hahn, 1989a; Dunstan et al., 1996).
Providing sufficient macroalgae is time consuming
and expensive with supply from wild harvesting of
local macroalgal beds unreliable and environmentally
unsustainable (Hahn, 1989a; Dunstan et al., 1996).
Consequently, growers wean abalone onto artificial
diets that are cheaper and may produce faster growth
if they meet the nutritional requirements of abalone
better than an individual algal species (Britz, 1996a,b;
Viana et al., 1996).
When using artificial diets growers need to develop
a successful weaning protocol, one element of which
is identifying the age or size that abalone are able to
consume and digest artificial diet successfully. Vari-
able weaning success is a major problem for the
industry and results in starvation and slow growth,
with some individuals displaying only 20–30% of the
average growth of their siblings (Hahn, 1989a).
Differential growth results in considerable size varia-
tion within a cohort, which requires costly and labour
intensive grading of the animals. The reasons for
variable growth are unclear, and may in part be due to
a poor understanding of changes in structure and
function of the abalone digestive system during
development and how these changes may be asso-
ciated with changes in diet.
The nocturnal and cryptic habit of juvenile
abalone (Shepherd, 1973; Saito, 1981) make it
difficult to observe changes in behaviour that are
likely to be important in determining their patterns of
mortality and growth (McShane, 1992). This has
resulted in little being known about ontogenetic
development associated with diet changes from
microalgae to macroalgae. Immediately after settle-
ment, abalone post-larvae feed on biofilm and mucus
trails (Shepherd, 1973; Saito, 1981) and once the
radula is developed, at around 0.8 mm shell length
(SL) (Kawamura et al., 2001), juveniles start feeding
on diatoms, turf, and crustose coralline algae
(Dunstan et al., 1996). Juveniles maintain this diet
until they are large enough to undergo the final diet
transition from diatoms to macroalgae (Jarayabhand
and Paphavasit, 1996; Kawamura et al., 2001). The
size of individuals at this final transition varies
among species, ranging from 5 to 10 mm for
Haliotis discus hannai (Kawamura et al., 2001), 7
to 8 mm for Haliotis rufescens (Hahn, 1989a) and 10
to 20 mm for Haliotis asinina and Haliotis ovina
(Jarayabhand and Paphavasit, 1996).
It is thought that the diet change occurs because
increasing diatom consumption of growing abalone
results in longer foraging trips (Shepherd and Turner,
1985). This in turn increases energy expenditure
associated with locomotion making it necessary to
exploit higher energy macroalgae as a food source
(McShane, 1992). However, developmental changes
in structure and function of the digestive system may
also possibly influence the timing of dietary tran-
sitions and therefore weaning success. Changes in
radula structure and organisation in Haliotis iris and
H. discus hannai are thought to be in preparation for
the feeding transition from micro- to macroalgae
(Roberts et al., 1999; Kawamura et al., 2001). There is
currently no information on the development of the
radula and gut of blacklip or greenlip abalone cultured
in Australia.
The basic structure of the adult abalone digestive
system has been described by Harris (1994). It
consists of an oesophagus that extends posteriorly of
the buccal region to a large crop organ, where food is
stored before entering the stomach. A crop extends
posteriorly into the stomach, which forms a 1808 loopat the posterior end of the abalone to extend anteriorly
adjacent to the crop. A voluminous digestive gland,
overlying the crop and stomach, occupies most of the
visceral mass and is connected with the other
digestive organs by a network of tubules and ducts.
The stomach continues anteriorly extending into the
style sac with the protostyle. The style sac is
connected to a long and complex intestine with five
regions (I–V). Food from the style sac enters region I
and exits region V (also known as the rectum) which
terminates at the anus.
Ontogenetic changes in the types and concentra-
tions of digestive enzymes are indicative of shifts in
the ability to hydrolyse dietary components and have

D. Johnston et al. / Aquaculture 250 (2005) 341–355 343
been used to identify dietary shifts in a range of
invertebrates (Hammer et al., 2000; Johnston, 2003).
For example, changes in protease and a-amylase
activities indicate changes in feeding preferences of
the black tiger prawn Penaeus monodon from
herbivory in nauplius, to carnivory in mysis, and
omnivory in juveniles and adults (Fang and Lee,
1992). Similarly, in Macrobranchium rosenbergii
enzyme activity changes and hepatopancreas develop-
ment occurs when individuals shift from exclusive
carnivory to omnivory during the larval period
(Kamarudin et al., 1994). Although this approach is
common in invertebrates there is no quantitative data
on ontogenetic changes in digestive enzymes in
abalone which may assist in identifying the critical
ages and sizes in relation to dietary changes.
In this study we describe the structure of the radula
and digestive system and document the digestive
physiology (digestive enzyme activities) of juvenile
blacklip abalone H. rubra between 80 and 158 days
post settlement fed on diatoms. We also compare these
structures and digestive physiology with abalone of
the same age weaned onto artificial diet and with
runts. This information will (1) identify changes in
structure and/or physiology associated with dietary
shift from diatoms to macroalgae, (2) determine when
the digestive physiology of juvenile blacklip abalone
is best suited to the transition from a natural diatom to
artificial diet and (3) identify whether the digestive
physiology or structural development in these runts
can explain reduced growth.
2. Materials and methods
2.1. Sampling
Juvenile H. rubra were collected from stock
produced and reared at ABTAS Seafoods, Garden
Island, Tasmania. Settled juveniles were fed encrust-
ing diatoms and biofilm for 158 days. To document
developmental changes in the structure and function
of the digestive system abalone from the same cohort
were sampled at 80 days (meanFS.E. maximum
shell length, 3.5F0.04 mm), 102 days (8.5F0.06
mm), 137 days (10.5F0.11 mm), and 158 days
(11.3F0.13 mm) post settlement (PS). Juveniles
were selected from individuals at the top end of
the size range to ensure that growing juveniles were
sampled. At 137 days PS small juveniles (5.5F0.37
mm) in the cohort identified as bruntsQ (approx-
imately 50% smaller than larger individuals) were
sampled on the assumption that these individuals
were not growing normally.
To determine the effects of weaning on the
digestive system, juvenile abalone weaned onto an
artificial diet were compared to juveniles of the same
age maintained on diatoms. At 105 days PS juveniles
from the same cohort were transferred to a clean tank
with no diatom biofilm and given an artificial diet
manufactured by Adam and Amos, Mt Barker, South
Australia. Abalones were fed the diet for 57 days until
162 days PS and 58 juveniles were sampled for
histology and enzyme analysis. Only successfully
weaned individuals were sampled, these were identi-
fied by a coloured band of new shell growth
associated with the artificial diet. To fully assess the
success of the artificial diet we compared the animals
fed the artificial diet with starved animals. To starve
juveniles, 20 individuals at 142 days PS were moved
into a clean tank and maintained for 20 days without
food. These animals were kept in the dark to ensure no
algal film grew to provide a food source for these
juveniles. The structure of the digestive system of
starved juveniles was examined using histological and
histochemical techniques.
All animals were sampled early in the morning to
ensure enzyme activity was at its greatest after they
had been feeding during the night. Following collec-
tion abalone were transported in seawater or on ice for
dissection in the laboratory. Prior to dissection or
freezing maximum shell length (SL) of all abalone
was measured to the nearest 0.1 mm using callipers.
Wild juvenile abalone (20–30 mm) were collected
from southern Tasmania and transferred to liquid
nitrogen for storage within 8 h of collection. Digestive
enzyme profiles of wild abalone were compared to the
cultured abalone.
2.2. Radula structure
Whole abalones were fixed in 10% seawater
formalin for 24 h and the radula removed with
forceps. The radula was placed in 1.5% sodium
hypochlorite (NaOCl) for 1 h to dissolve attached
connective tissue and mucous, then washed with

D. Johnston et al. / Aquaculture 250 (2005) 341–355344
distilled water and stored separately. Radulae were
dehydrated in an ethanol series to 100%, critical point
dried, gold sputter coated, and examined on an Electro
Scan 2020 environmental scanning electron micro-
scope at 15 kV. Morphology and the presence or
absence of different tooth types (rachidian, laterals 1
and 2, laterals 3–5, and marginals) was described.
2.3. Histology and histochemistry
Abalone were fixed in FAACC (10% formalin, 5%
glacial acetic acid and 1.3% calcium chloride) for 24
h, after which the abalone tissue was removed from
the shell and returned to fresh FAACC for storage at
room temperature until processing. Whole abalones
were processed for wax histology using standard
methods and transverse serial sections (5 Am) cut and
mounted on poly-l-lysine coated microscope slides.
Sections were stained with Mallory-Heidenhain Tri-
chrome and examined using an Olympus BH-2
microscope.
Individuals used for histochemical analysis were
fixed in 10% seawater formalin for 24 h and processed
and sectioned as above. Sections of digestive gland,
stomach, crop and intestine were stained with
Mercuric Bromophenol Blue for 15 min at room
temperature (Chapman, 1975). This stain binds free
protein within tissue to identify where digestive
enzymes were being produced and secreted.
2.4. Enzyme analyses
The digestive gland from each abalone was
dissected out on ice and pooled into one of four
replicate tubes for each age group, the numbers
pooled per age group depending on size of the
abalone (80 and 102 days PS, n =25; 137 days PS,
n =16; 158 days PS, n =13; artificial diet n =12). The
digestive glands from wild abalone (12 in total) were
dissected out on ice and four were pooled into each of
three replicate tubes. Tissue was frozen in liquid
nitrogen and stored at �80 8C until extraction.
Thawed abalone were homogenised in chilled 0.1 M
Tris 0.02 M NaCl buffer pH 7.5 using an Ultra Turrax
homogeniser fitted with a S8N-8G dispersing tool
(IKA-Works Germany). The homogenate was centri-
fuged at 10,000�g for 10 min at 4 8C and aliquots of
supernatant stored at �20 8C.
One enzyme unit is defined as the amount of
enzyme that catalysed the release of 1 nmol of
product/min and was calculated using the appropriate
molar extinction coefficient (e) for the assay con-
ditions or standard curve. Specific activity was
defined as enzyme activity per mg of abalone protein
(units mg protein�1) and total activity was defined as
enzyme activity per abalone (units abalone�1). Protein
concentration was determined by the method of
Bradford (1976) using bovine serum albumin as the
standard. Spectrophotometric enzyme assays (200 Almicro-assays) were performed in duplicate at 37 8C(a-glucosidase, h-glucosidase and laminarinase) or 32
8C (lipase and trypsin) in IWAKI flatbottom micro-
plates and absorbances read in a Tecan Spectro
Rainbow Thermo microplate reader. Appropriate
controls were included with each analysis.
Trypsin was assayed using N-a-benzoylarginine-U-nitroanalide (BAPNA) dissolved in dimethylforma-
mide (DMF) as substrate. Each assay contained a final
concentration of 1.25 mM BAPNA in 0.1 M citrate
0.2 M phosphate buffer pH 5.5. Assays were initiated
by the addition of enzyme extract and the release of U-nitroanalide measured at A400–410. Under these assay
conditions the molar extinction coefficient was 9300
M�1 cm�1 for U-nitroanaline (Stone et al., 1991). A
positive control of 3 mg ml�1 porcine pancreas
trypsin in 1 mM HCl was used.
a-Glucosidase and h-glucosidase activities were
determined using U-nitrophenyl a-d-glucopyranoside(Sigma N1377) and U-nitrophenyl h-d-glucopyrano-side (Sigma N7006) as substrates, respectively. Each
assay contained a final concentration of 4 mM
substrate in 0.1 M citrate 0.2 M phosphate buffer
pH 5.5. Assays were initiated with the addition of
enzyme extract. Aliquots of assay mixture were
removed at time intervals and added to 1 M Na2CO3
(pH 11), to terminate the reaction. Liberation of U-nitrophenol was measured at A400. The molar
extinction coefficient is 18 300 M�1 cm�1 for U-nitrophenol at pH N9 (Erlanger et al., 1961).
Laminarinase activity was measured using lami-
narin as substrate. Each assay mixture contained 10
mg ml�1 of laminarin dissolved in 0.1 M phosphate
buffer pH 5.5 and the reaction was initiated by
addition of enzyme extract. Assay mixture was
removed at 0 min and after 60 min incubation at 37
8C and the concentration of glucose liberated was

D. Johnston et al. / Aquaculture 250 (2005) 341–355 345
measured by adding this to glucose (HK) assay
reagent (Sigma G2020). Glucose (HK) reagent con-
verts glucose in the assay mixture to NADH via a
coupled enzyme reaction catalysed by hexokinase and
glucose-6-dehydrogenase. After 15 min incubation at
room temperature the absorbance of NADH, which is
proportional to the concentration of glucose, was read
at A340. The amount of glucose liberated min�1 mg�1
was calculated using a standard curve that was
generated by incubating known amounts of glucose
with glucose (HK) reagent.
Lipase activity was determined using a method
modified from Gjellesvik et al. (1992) using 4-
nitrophenyl caproate (4-NPC) dissolved in ethanol
as substrate. Each assay contained a final concen-
tration of 2.5 mM 4-NPC in 6 mM sodium taurocho-
late, 500 mM Tris, 100 mM NaCl buffer pH 7.4.
Assays were initiated by the addition of enzyme
extract and the release of nitrophenol was measured at
A405. Under these assay conditions the molar extinc-
tion coefficient was 19,800 M�1 cm�1 for nitrophenol
(Gjellesvik et al., 1992).
2.5. Statistical analyses
Specific enzyme activity was compared among the
age groups using a one-way ANOVA. A second one-
way ANOVA was used to compare the enzyme
activity among juveniles fed diatoms (157 days PS),
juveniles fed artificial diet (162 days PS) and wild
juveniles (unknown age). Data were tested for
heterogeneity of variance using residual plots. Sig-
nificant differences among means were determined
using Tukeys HSD post-hoc test (P b0.05).
3. Results
3.1. Ontogenetic changes
3.1.1. Radula structure
At 80 days post settlement abalone had well
differentiated teeth, typical of a Rhipidoglossan radula
(Fig. 1A–C). The radula was asymmetrical with four
types of teeth present in pairs within a tooth row with
one either side of a central rachidian tooth; one
rachidian, a pair of 1st laterals, a pair of 2nd laterals, a
pair of 3rd laterals, a pair of 4th laterals. On the
outside of laterals, many marginals extended in pairs
to the outermost edge of the radula tooth row (Fig.
1A,B). The rachidian tooth was slightly cuspid (curled
tip), with the cusp towards the posterior end of the
radula and its edge flattened (Fig. 1B). The 1st laterals
interlocked with the rachidian and were different in
morphology having a larger cusp (same direction) and
a rounded spade shaped tip (Fig. 1B). The 2nd laterals
were narrower than the 1st but also had a posterior
facing pronounced cusp (Fig. 1B). There was a
distinct change in tooth morphology between the
2nd lateral and the 3rd lateral. The 3rd lateral was the
largest tooth in the radula and the tip (facing
posteriorly) was extended and pointed, almost
canine-like (Fig. 1C). The 4th lateral shared the same
general morphology as the 3rd, but smaller in size
with serrations on both sides of the tooth, whereas the
3rd lateral only had serrations on the outer side away
from the rachidian (Fig. 1C). The marginals were
distinguishable from the 4th lateral by increased
serrations on both sides of the tooth and a rounded
end as opposed to the point of the 3rd and 4th laterals
(Fig. 1C). Marginals also orientate differently to the
3rd and 4th laterals being closely folded on top of
adjacent marginals with their cusps pointing more
outer laterally as compared to posteriorly. As the
marginals moved to the outer edge of the radula the
number of serrations on both sides increase.
The radula of 102 days PS abalone had an
additional 5th lateral that was the same shape as the
3rd and 4th laterals, but smaller in size (Fig. 1D). The
rachidian was less cuspid (having a reduced curl in the
tip) and the 1st and 2nd laterals appeared to increase
in width. In addition to these changes the rachidian
and 1st lateral of 137 days PS abalone had lost the
cuspid shape and the 3rd, 4th and 5th laterals were
larger and had sharper points. There was no change in
the radula structure between 137 days and 158 days
post settlement.
3.1.2. Structure of digestive system
In 80 days PS abalone the oesophagus was a
longitudinally folded tubular organ (Fig. 2A), with an
epithelium of two cell types; tall columnar ciliated,
and mucus cells. Ciliation was sparse but extended
around the entire internal perimeter of the organ
(Fig. 2A). Longitudinal folding of the oesophagus
increased between 80 days PS and 102 days PS and
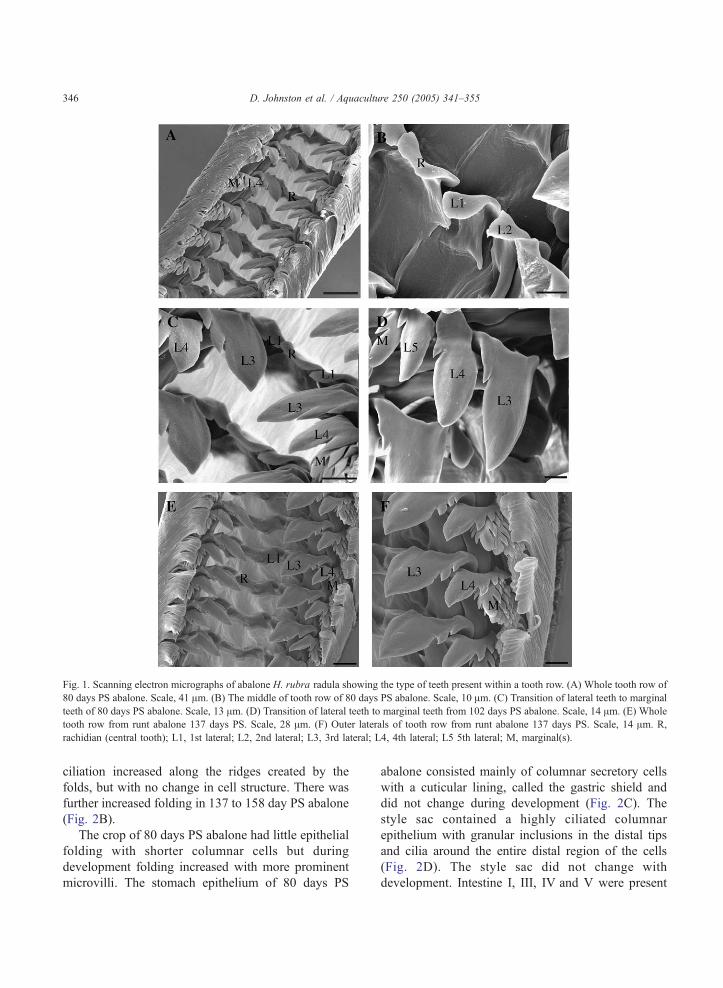
Fig. 1. Scanning electron micrographs of abalone H. rubra radula showing the type of teeth present within a tooth row. (A) Whole tooth row of
80 days PS abalone. Scale, 41 Am. (B) The middle of tooth row of 80 days PS abalone. Scale, 10 Am. (C) Transition of lateral teeth to marginal
teeth of 80 days PS abalone. Scale, 13 Am. (D) Transition of lateral teeth to marginal teeth from 102 days PS abalone. Scale, 14 Am. (E) Whole
tooth row from runt abalone 137 days PS. Scale, 28 Am. (F) Outer laterals of tooth row from runt abalone 137 days PS. Scale, 14 Am. R,
rachidian (central tooth); L1, 1st lateral; L2, 2nd lateral; L3, 3rd lateral; L4, 4th lateral; L5 5th lateral; M, marginal(s).
D. Johnston et al. / Aquaculture 250 (2005) 341–355346
ciliation increased along the ridges created by the
folds, but with no change in cell structure. There was
further increased folding in 137 to 158 day PS abalone
(Fig. 2B).
The crop of 80 days PS abalone had little epithelial
folding with shorter columnar cells but during
development folding increased with more prominent
microvilli. The stomach epithelium of 80 days PS
abalone consisted mainly of columnar secretory cells
with a cuticular lining, called the gastric shield and
did not change during development (Fig. 2C). The
style sac contained a highly ciliated columnar
epithelium with granular inclusions in the distal tips
and cilia around the entire distal region of the cells
(Fig. 2D). The style sac did not change with
development. Intestine I, III, IV and V were present
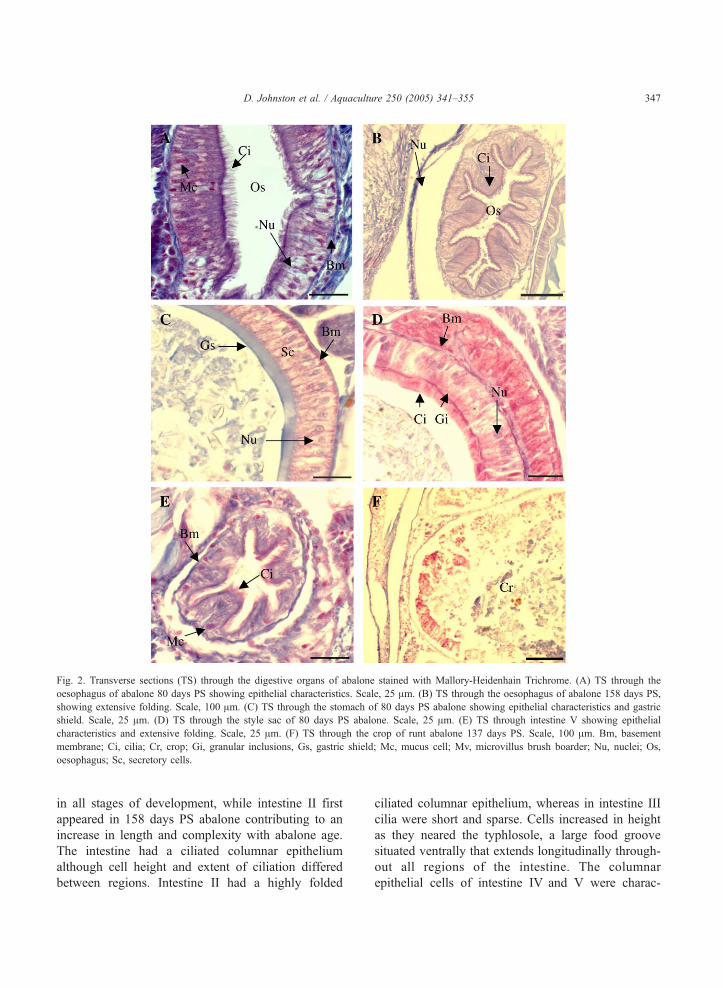
Fig. 2. Transverse sections (TS) through the digestive organs of abalone stained with Mallory-Heidenhain Trichrome. (A) TS through the
oesophagus of abalone 80 days PS showing epithelial characteristics. Scale, 25 Am. (B) TS through the oesophagus of abalone 158 days PS,
showing extensive folding. Scale, 100 Am. (C) TS through the stomach of 80 days PS abalone showing epithelial characteristics and gastric
shield. Scale, 25 Am. (D) TS through the style sac of 80 days PS abalone. Scale, 25 Am. (E) TS through intestine V showing epithelial
characteristics and extensive folding. Scale, 25 Am. (F) TS through the crop of runt abalone 137 days PS. Scale, 100 Am. Bm, basement
membrane; Ci, cilia; Cr, crop; Gi, granular inclusions, Gs, gastric shield; Mc, mucus cell; Mv, microvillus brush boarder; Nu, nuclei; Os,
oesophagus; Sc, secretory cells.
D. Johnston et al. / Aquaculture 250 (2005) 341–355 347
in all stages of development, while intestine II first
appeared in 158 days PS abalone contributing to an
increase in length and complexity with abalone age.
The intestine had a ciliated columnar epithelium
although cell height and extent of ciliation differed
between regions. Intestine II had a highly folded
ciliated columnar epithelium, whereas in intestine III
cilia were short and sparse. Cells increased in height
as they neared the typhlosole, a large food groove
situated ventrally that extends longitudinally through-
out all regions of the intestine. The columnar
epithelial cells of intestine IV and V were charac-
D. Johnston et al. / Aquaculture 250 (2005) 341–355348
terised by the increased presence of mucus cells (Fig.
2E). Intestine V was highly folded and possessed the
longest cilia in the intestine (Fig. 2E).
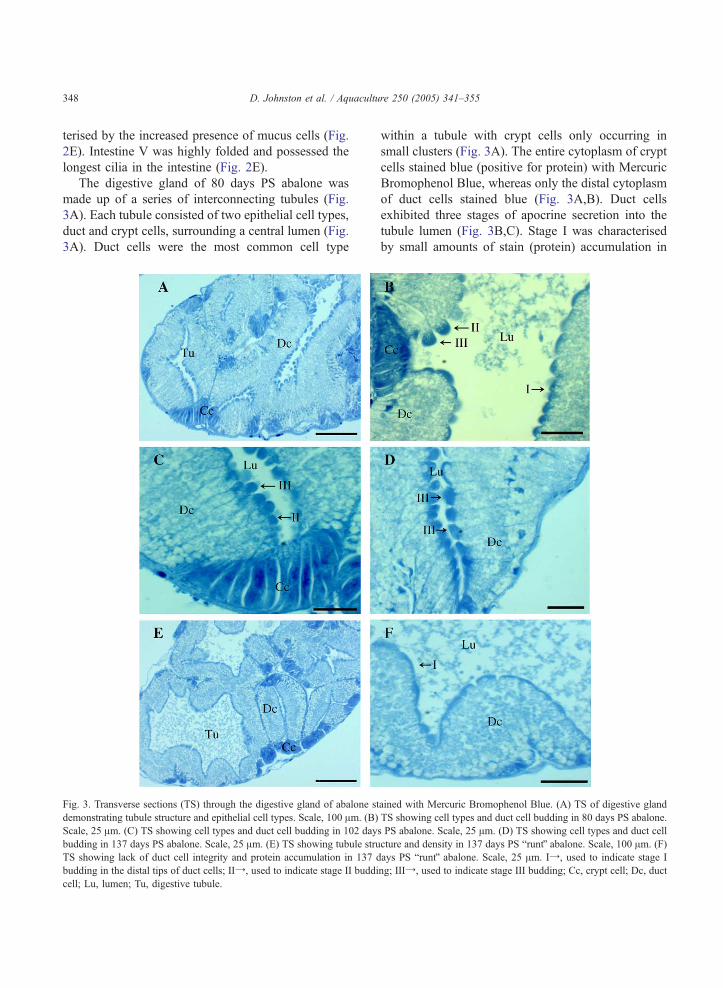
The digestive gland of 80 days PS abalone was
made up of a series of interconnecting tubules (Fig.
3A). Each tubule consisted of two epithelial cell types,
duct and crypt cells, surrounding a central lumen (Fig.
3A). Duct cells were the most common cell type
Fig. 3. Transverse sections (TS) through the digestive gland of abalone st
demonstrating tubule structure and epithelial cell types. Scale, 100 Am. (B)
Scale, 25 Am. (C) TS showing cell types and duct cell budding in 102 day
budding in 137 days PS abalone. Scale, 25 Am. (E) TS showing tubule stru
TS showing lack of duct cell integrity and protein accumulation in 137 d
budding in the distal tips of duct cells; IIY, used to indicate stage II buddi
cell; Lu, lumen; Tu, digestive tubule.
within a tubule with crypt cells only occurring in
small clusters (Fig. 3A). The entire cytoplasm of crypt
cells stained blue (positive for protein) with Mercuric
Bromophenol Blue, whereas only the distal cytoplasm
of duct cells stained blue (Fig. 3A,B). Duct cells
exhibited three stages of apocrine secretion into the
tubule lumen (Fig. 3B,C). Stage I was characterised
by small amounts of stain (protein) accumulation in
ained with Mercuric Bromophenol Blue. (A) TS of digestive gland
TS showing cell types and duct cell budding in 80 days PS abalone.
s PS abalone. Scale, 25 Am. (D) TS showing cell types and duct cell
cture and density in 137 days PS bruntQ abalone. Scale, 100 Am. (F)
ays PS bruntQ abalone. Scale, 25 Am. IY, used to indicate stage I
ng; IIIY, used to indicate stage III budding; Cc, crypt cell; Dc, duct
D. Johnston et al. / Aquaculture 250 (2005) 341–355 349
the distal tip; stage II by increased accumulation
starting to form a distended bud; and stage III by a
distinct bud at the distal tip or separation of the bud
from the duct cell (Fig. 3B,C).
The number of digestive tubules increased in 102
days PS abalone and duct cells showed more
concentrated blue staining in the distal tips and more
cells were budding into the lumen (Fig. 3C). There
was no change in staining, location or prevalence of
crypt cells within a tubule. There did not appear to be
any change in tubule density within the digestive
gland between 102 days PS and 137 days PS,
although duct cells showed increased stain accumu-
lation and budding in the distal tips as indicated by the
increase in stage III duct cells (Fig. 3D). There was no
change in digestive gland structure between 137 days
PS and 158 days PS.
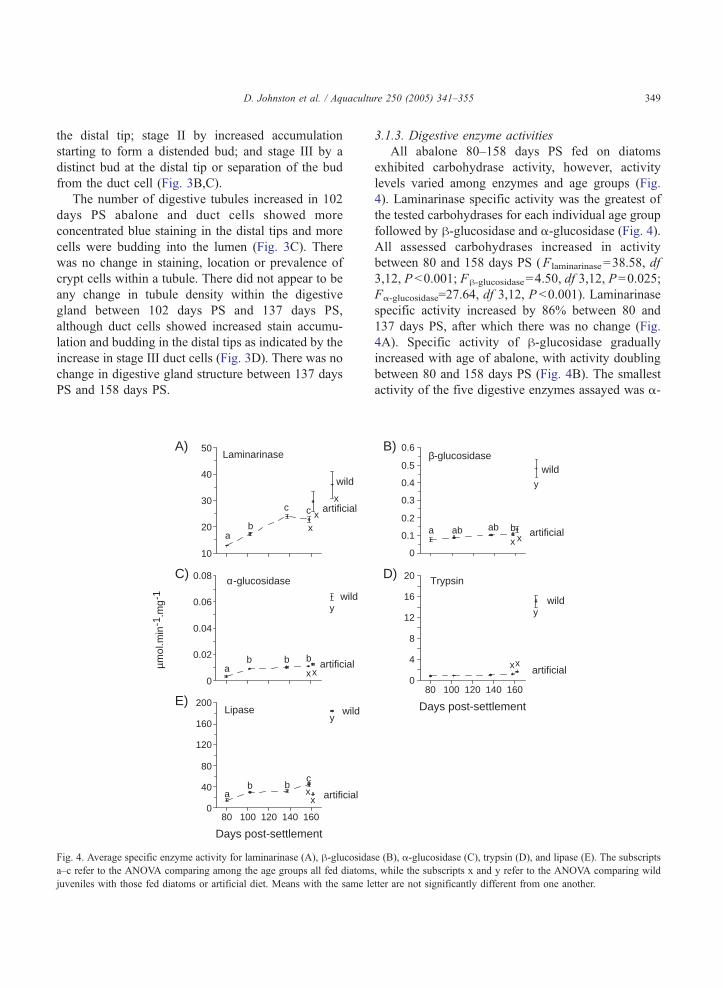
A)
0
0.02
0.04
0.06
0.08 α-glucosidase
80 100 120 140 1600
40
80
120
160
200
C)
µmo
l.min
-1.m
g-1
Lipase
10
20
30
40
50
E)
Laminarinase
wild
artificial
Days post-settlement
wild
artificial
ab
c c
ab b b
xx
y
artificial
wild
ab b
cxx
y
xx
x
Fig. 4. Average specific enzyme activity for laminarinase (A), h-glucosidaa–c refer to the ANOVA comparing among the age groups all fed diatoms
juveniles with those fed diatoms or artificial diet. Means with the same le
3.1.3. Digestive enzyme activities
All abalone 80–158 days PS fed on diatoms
exhibited carbohydrase activity, however, activity
levels varied among enzymes and age groups (Fig.
4). Laminarinase specific activity was the greatest of
the tested carbohydrases for each individual age group
followed by h-glucosidase and a-glucosidase (Fig. 4).
All assessed carbohydrases increased in activity
between 80 and 158 days PS (Flaminarinase=38.58, df
3,12, P b0.001; Fh-glucosidase=4.50, df 3,12, P=0.025;
Fa-glucosidase=27.64, df 3,12, P b0.001). Laminarinase
specific activity increased by 86% between 80 and
137 days PS, after which there was no change (Fig.
4A). Specific activity of h-glucosidase gradually
increased with age of abalone, with activity doubling
between 80 and 158 days PS (Fig. 4B). The smallest
activity of the five digestive enzymes assayed was a-
0
0.1
0.2
0.3
0.4
0.5
0.6β-glucosidase
B)
0
4
8
12
16
20D)Trypsin
wild
80 100 120 140 160
Days post-settlement
artificial
wild
artificial
a ab ab bx x
y
xx
y
se (B), a-glucosidase (C), trypsin (D), and lipase (E). The subscripts
, while the subscripts x and y refer to the ANOVA comparing wild
tter are not significantly different from one another.

D. Johnston et al. / Aquaculture 250 (2005) 341–355350
glucosidase (Fig. 4C), however, there was a signifi-
cant increase in activity between 80 and 102 days PS
abalone, after which time activity was constant.
Trypsin specific activity did not change signifi-
cantly from 80 days PS to 158 days PS abalone
(F =2.52, df 3,12, P=0.11). The average specific
activity of trypsin for the four age groups was
1.02F0.017 units mg protein�1 (Fig. 4D). Lipase
specific activity increased significantly from 80 days
PS to 157 days PS (F =24.86, df 3,12, P b0.001),
with activity more than doubling (Fig. 4E).
3.2. Changes associated with weaning onto an
artificial diet
3.2.1. Structure of digestive system
In starved animals the number of tubules remained
similar but the lumen area was enlarged and empty.
Duct cells were smaller and showed minimal distal
cytoplasm staining with no evidence of budding. Duct
cells were often indistinguishable due to cell wall
rupture and cell nuclei were not visible. In contrast,
abalone that had been weaned onto artificial diet
showed no visible differences in digestive system
structure to those abalone fed diatoms of the same
age.
3.2.2. Digestive enzyme activities
In a comparison between juvenile abalone fed
diatoms, artificial diet, and wild juveniles there was a
significant difference in specific activity of h-gluco-sidase (F =61.06, df 2,8, P b0.001), a-glucosidase
(F=430.04, df 2,8, P b0.001), lipase (F =79.21, df
2,8, P b0.001), and trypsin (F =212.42, df 2,8,
P b0.001), but not in laminarinase (F =3.45, df 2,8,
P=0.083). Abalone fed the artificial diet did not have
significantly different specific enzyme activities in the
five enzymes when compared to siblings fed on
diatoms (Fig. 4). Wild juveniles had specific activities
3.6–8.7 times greater than those juveniles fed the
artificial diet in all the digestive enzymes except for
laminarinase (Fig. 4).
3.3. Runts
3.3.1. Radula structure
The radula in runt abalone 137 days PS had only
four lateral teeth (1st, 2nd, 3rd 4th) plus marginal
teeth, compared with the full complement of laterals
in their larger siblings of the same age (Fig. 1E,F).
The 137 days PS runts had a radula structure similar
to that of full size 80 day PS abalone, with a narrow
cuspid rachidian, 1st and 2nd laterals, and highly
serrated 4th laterals (Fig. 1F).
3.3.2. Structure of digestive system
There were fewer digestive gland tubules in runt
abalone than in siblings of the same age, but crypt
cells were similar in structure (Fig. 3E). Duct cells had
lost structural integrity, with individual duct cells
indistinguishable due to cell wall rupture and cell
nuclei were no longer visible (Fig. 3F). There was also
loss of epithelium from the basal lamina and very little
accumulation of secretory products (blue staining
protein) in the distal tips of duct cells (Fig. 3F).
The epithelium of digestive organs in runt abalone
was also very different to that of their 137 days PS
siblings. Runts showed variable levels of degradation
and tissue histology indicated severe epithelial vacuo-
lation, loss of cell integrity, epithelial sloughing, and
lack of visible nuclei (Fig. 2F), as well as decom-
position of the foot muscle. Where the epithelium was
identifiable, it had the same cell structure as 80 days
PS abalone. Intestine IV was contorted into an
irregular morphology.
4. Discussion
4.1. Ontogenetic development
The radula structure of H. rubra juveniles was of
typical Rhipidoglossan form: consisting of a central
column of heavier central (rachidian) and lateral teeth,
a fan-like arrangement of marginals on the outer
perimeter, and made from superficially hardened
chitin (Sollas, 1907; Crofts, 1929; Hickman, 1980).
Major structural changes occurred in the radula of
juvenile blacklip abalone H. rubra between 80 and
102 days PS. The rachidian of H. rubra was initially
curled, but as the abalone aged the rachidian
straightened resulting in a greater cutting ability by
changing the angle that the tip makes contact with the
substrate (Padilla, 1985). The development of L5 teeth
and an increase in the size of L3–L5 suggests these
teeth are more suited for gouging and collecting large

D. Johnston et al. / Aquaculture 250 (2005) 341–355 351
food particles (Roberts et al., 1999; Kawamura et al.,
2001). Fewer serrations on the outer laterals L3–L5
suggest that the radula has become less specialised for
handling small food particles as the animal developed
(Roberts et al., 1999; Kawamura et al., 2001).
Increased grazing efficiency and capability of the
radula with development in H. rubra is presumably
needed for the transition to feeding on macroalgae
allowing them to exploit larger and tougher food
material and increase mastication efficiency. The
radula of the older 102 days PS animals was similar
to the adult form (Crofts, 1929; Herbert, 1990;
Chitramvong et al., 1998; Roberts et al., 1999;
Kawamura et al., 2001) and there were only minor
morphological changes to the teeth after 102 days PS,
suggesting that H. rubra are already fully equipped to
consume a macroalgal diet by this age. The develop-
ment of the radula in H. rubra was slower than other
temperate species, such as H. iris and H. discus
hannai, which have a full adult complement of 5
lateral teeth and associated tooth morphology by 60–
63 days PS (Leighton, 1974; Shepherd and Hearn,
1983; Roberts et al., 1999; Kawamura et al., 2001).
The histology of individual digestive organs in H.
rubra was similar to the adults of other abalone
species, H. tuberculata (Crofts, 1929), Haliotis
cracherodii (Campbell, 1965), H. rufescens (McLean,
1970; Bevelander, 1988) and H. laevigata (Harris,
1994; Harris et al., 1998). Elaboration of oesophageal
folding between 80 and 158 days PS increases the
epithelial surface area creating greater interaction
between food particles and the cilia and the mucus,
thereby increasing the rate of food passage into the
crop (Roberts et al., 1999; Kawamura et al., 2001).
Increased folding and expansion of the crop between
80 and 158 days PS would maximize the uptake of
nutrients with more of the distal microvilli coming
into contact with digesting food (McLean, 1970). The
appearance of intestine II at 158 days PS greatly
increases the area for nutrient absorption in older
juveniles (Crofts, 1929; Campbell, 1965; McLean,
1970; Bevelander, 1988). This looped morphology
slows the passage of food, increasing digestion and
absorption (Oozeki and Bailey, 1995).
Positive stain for protein in distal cytoplasmic
granules in duct cells of the digestive gland indicates
that these cells are producing and accumulating
digestive enzymes. In contrast, crypt cells did not
appear have an enzyme secretory function, given no
distal accumulation of protein in the cells, no evidence
of budding, and no difference in staining between fed
and starved abalone. This is consistent with other
studies that suggest duct cells synthesise enzymes
capable of digesting proteins, lipids, and carbohy-
drates (Bevelander, 1988) and secrete enzymes to
other organs such as the caecum (Purchon, 1968;
Morton, 1979), crop (McLean, 1970) and stomach
(Purchon, 1968; McLean, 1970; Morton, 1979).
Hence the increase in the number of digestive gland
tubules and digestive gland complexity as well as
distal cytoplasm staining in duct cells in 80–102 days
PS abalone indicates that digestive capacity increases
with age. The enzyme secretory role of duct cells is
evident in H. rubra, as enzymes are accumulated
within the distal region of duct cells forming a bud
before being released into the lumen. This is
consistent with apocrine secretion described in other
invertebrate species, e.g. the surf barnacle Tetraclita
squamosa (Johnston et al., 1993).
Juvenile H. rubra fed a diatom diet exhibited
extremely high laminarinase activity reflecting the
high concentration of the algal storage polysaccharide,
chrysolaminarin, in the ingested diatoms. Wild
abalone of similar age also had high levels of
laminarinase suggesting that carbohydrases such as
laminarinase play a major role in digestion for all
abalone; a consequence of the high carbohydrate
composition of their natural diet (Mori, 1953; Takami
et al., 1998). Laminarin and chrysolaminarin are h-glucans that are also present in brown macroalgae
(Phaeophyta), and the same enzymes are necessary for
the utilisation of macroalgae as a diet. Laminarinase
activity increased from 80 to 158 days PS coinciding
with changes in the radula structure, digestive gland,
and digestive organs; supporting the hypothesis that
these individuals are preparing for the shift in diet
from diatoms to macroalgae. A similar pattern of
development occurs in H. discus hannai, where
laminarinase specific activity increased in conjunction
with development of L5 teeth and differentiation of
laterals in 63 days PS abalone (Takami et al., 1998;
Kawamura et al., 2001). This would allow the dietary
transition from microalgae to the macroalgae in this
species (Takami et al., 1998; Kawamura et al., 2001).
The specific activity of h-1,4-glucosidase, a struc-
tural polysaccharide degrading enzyme (McCandless,

D. Johnston et al. / Aquaculture 250 (2005) 341–355352
1981) increased between 80 and 158 days PS also
reflecting the increasing rate of diatom consumption in
older abalone and subsequently macroalgae. In con-
trast, a-1,4-glucosidase activity, more typically seen in
terrestrial herbivores (Bennett et al., 1971), was low,
approximately a 10th of h-1,4-glucosidase activity,
and similar to the levels of a-1,4-glucosidase reported
in adult H. rufescens (Bennett et al., 1971).
There has been extensive research on the digestive
proteases of Haliotis, compared to other enzymes, as
industry pushes to incorporate higher protein levels
into the diet (Fleming et al., 1996; Edwards and
Condon, 2001). Trypsin activity in H. rubra did not
change from 80 to 158 days PS, which does not
exclude dietary transitions, rather reflects the low
protein levels in both micro- and macroalgae (Viana et
al., 1996; Edwards and Condon, 2001). Abalone
cannot synthesise 10 of the 20 l-amino acids required
to assemble proteins, and their growth is dependent on
utilising what protein is available in the diet (Serviere-
Zaragoza et al., 1997). Trypsin activities increase in
adult H. fulgens, whereas juvenile H. fulgens lack
carboxypeptidases A and B (Serviere-Zaragoza et al.,
1997) and the carboxypeptidase-like enzyme of the
adults (Hernandez-Santoyo et al., 1998). Future
research on proteases of juvenile abalone approaching
the age when diet transition takes place should focus
on a suite of digestive proteases rather than an
individual enzyme.
Lipase activity in juvenile H. rubra is high
compared with other enzymes and reflects the high
content of lipid in their diatom diet, with a need to
rapidly digest lipid rather than store it, as abalone
contain only low lipid reserves (Dunstan et al., 1996).
Lipase activity increased with age of H. rubra
suggesting an increased ability to digest lipids as they
develop and reflects the increase in lipid consumption
with the higher diatom grazing rates of older abalone.
Lipid digesting enzymes may prove useful in identi-
fying a diet transition between wild juvenile and adult
abalone, with a potential decline in activity indicating
the utilisation of low lipid composition macroalgae
(Ragan, 1981) as compared to higher lipid composi-
tion diatoms.
In most cases, the digestive enzyme activities of
wild H. rubra were significantly higher than in
diatom or artificial diet fed cultured abalone of
similar size. It is not clear what factors are responsible
for these differences. As the level of enzyme activity
is a function of the diet it is likely that the
composition of the diet differed rather that differences
in quantity consumed. It may be that the natural diet
ingested was considerably higher in protein, carbohy-
drate, and lipid than diatoms ingested by cultured
animals. The exception to this trend was laminarinase
activity which was not significantly different from
cultured abalone. This suggests that the content of
laminarin consumed in the diet of cultured and wild
abalone was similar. It is clear that digestive enzyme
analysis of wild abalone needs further investigation to
fully understand the digestive physiology of these
animals.
4.2. Artificial diet
There were no differences in the morphology of the
radula and the digestive system of H. rubra, feed
natural and artificial diets; a similar situation for H.
laevigata (Harris, 1994; Harris et al., 1998). There
were however, significant differences in their diges-
tive physiology. Artificial abalone diets are generally
remarkably similar in their proximate composition,
usually containing a high level of protein to increase
growth rates (Viana et al., 1993, 1996; Britz, 1996a,b;
Knauer et al., 1996; Bautista-Teruel and Millamena,
1999), a high level of carbohydrate and low levels of
lipid (Fleming et al., 1996).
Trypsin activity in the abalone fed the artificial diet
was significantly greater than those of similar age fed
diatoms, indicative of higher levels of protein in the
artificial diet. Marine algae generally have a protein
composition approximately 10–20% of dry weight
(Piscos-Garcia et al., 2000; Edwards and Condon,
2001), whereas artificial diets have a protein content
of 30–50% of dry weight (Fleming et al., 1996). This
study showed juvenile H. rubra increased their
trypsin activity to use the higher amounts of protein
in the artificial diet. In contrast, adult H. rubra do not
change trypsin activity in response to a high protein
diet (Edwards and Condon, 2001) and it is possible
that adult H. rubra do not share the flexible
enzymology of the juveniles. However, adult trial
was only for 3 weeks which may have been too short
for a response. Juvenile H. midae also show increased
protease activity after being fed an artificial diet
(Knauer et al., 1996) and similar trends have been

D. Johnston et al. / Aquaculture 250 (2005) 341–355 353
shown using more crude enzymatic methods for a
number of other species on high protein diets, H.
midae (Britz, 1996a); H. fulgens (Viana et al., 1996);
H. discus hannai (Bautista-Teruel and Millamena,
1999).
Feeding on artificial diet had no effect on the
carbohydrase activities of juvenile H. rubra presum-
ably because both the natural (micro- and macroalgae)
and artificial diet of abalone contain mostly carbohy-
drate (Fleming et al., 1996). In contrast, lipase activity
was significantly lower in abalone fed the artificial
diet reflecting the decreased amount of lipid compared
to diatoms. The natural diet of adult abalone contains
only small quantities of fatty acids and lipids (Ragan,
1981; Dunstan et al., 1996). Artificial diets usually
contain lipid at approximately 5% of dry matter
(Fleming et al., 1996), as higher levels have detri-
mental impacts on growth (Dunstan et al., 1996).
Unfortunately the investigation into lipid tolerance is
limited to adults; interestingly juveniles may be better
equipped to digest greater quantities of lipid. Certainly
this study has revealed that juvenile H. rubra have a
greater ability to digest lipid than previously thought
and the level of lipid inclusion in artificial diets for
blacklip abalone, H. rubra, may need to be revised to
best reflect their lipid digestion capability. The type of
lipid used also needs careful consideration, as the
typical lipid sources for abalone artificial diets are fish
oil and vegetable oil (Fleming et al., 1996), which are
not utilised by abalone naturally.
4.3. Runt abalone
Tissue degradation in under-developed (runt)
abalone 137 days PS suggests that they were nutri-
tionally compromised. Runts had radula similar to
much younger (80 days PS) abalone and this lack of
radula development and the resultant inability to
ingest the adult diet may limit their survivorship.
The under-developed radula structure of runt abalone
may have been inefficient at removing diatoms, which
had little or no adhesion to the substrate. The presence
of food in the stomach is one of the main stimuli for
enzyme secretion (Takami et al., 1998). Therefore,
given that the duct cells had little or no enzyme
budding present, and that minimal digestive enzyme
activity was detected in these individuals it appears
that runt abalone ingested little or no food.
H. rubra runts will start growing again when
removed from their siblings and fed an artificial diet
(Hindrum, personal communication). Similarly H.
fulgens exhibits compensatory growth when changed
from a low nutrition value diet to one higher in
nutritional value (Viana et al., 1996). Haliotis
kamschatkana will also survive and make a full
recovery after 27 days of starvation when feeding
resumes (Carefoot et al., 1993). The actual cause of
runting in these species is unclear, although avail-
ability or provision of the correct food appears to be a
factor. Carefully controlled experiments that account
for the effects of genetics, density, and feeding rates
will provide clearer insights to causes of variability in
growth rates. Grading individuals in each cohort and
providing additional food and/or different food may
be a way to manage stock to reduce variability in size
caused by ineffective weaning of abalone juveniles
from diatoms to artificial diet.
Acknowledgements
We thank ABTAS Seafood, Garden Island, Tasma-
nia, for supporting this project and supplying abalone
for the duration of the experimental period. In
particular, we thank Steve Hindrum and Andrew
McArther for their co-operation and assistance. Thank
you to Craig Mundy for collection of the wild abalone
and Martin Lourey for critical review of a final draft
of the manuscript.
References
Bautista-Teruel, M.N., Millamena, O.M., 1999. Diet development
and evaluation for juvenile abalone Haliotis asinina: protein/
energy levels. Aquaculture 178, 117–126.
Bennett Jr., R., Thannassi, N., Nakada, H.I., 1971. Hepatopancraes
glycosidases of the abalone (Haliotis rufescens). Comp. Bio-
chem. Physiol. 40B, 807–811.
Bevelander, G., 1988. Abalone: Gross and Fine Structure. Boxwood
Press, California, pp. 24–39.
Bradford, M.M., 1976. A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing
the principle of protein–dye binding. Anal. Biochem. 72,
248–254.
Britz, P.J., 1996a. Effect of dietary protein level on growth
performance of South African abalone Haliotis midae, fed
fishmeal-based semi-purified diets. Aquaculture 140, 55–61.

D. Johnston et al. / Aquaculture 250 (2005) 341–355354
Britz, P.J., 1996b. The suitability of selected protein sources for
inclusion in formulated diets for the South African abalone
Haliotis midae. Aquaculture 140, 63–73.
Campbell, J.L., 1965. The structure and function of the alimentary
canal of the black abalone Haliotis cracherodii leach. Amer.
Microsc. Soc. 84, 376–395.
Carefoot, T.H., Qian, P.-Y., Taylor, B.E., West, T., Osborne, J.,
1993. Effect of starvation on energy reserves and metabolism of
the Northern Abalone Haliotis kamschatkana. Aquaculture 118,
315–325.
Chapman, D.M., 1975. Dichromatism of bromophenol blue, with an
improvement in the mercuric bromophenol blue technique for
protein. Stain Technol. 50, 25–30.
Chitramvong, Y.P., Upatham, E.S., Kruatrachue, M., Sobhon, P.,
Limthong, V., 1998. Scanning electron microscope study of
radulae in Haliotis asinina Linnaeus, 1758 and Haliotis ovina
Gmelin, 1791 (Gastropoda: Haliotidae). J. Shellfish Res. 17,
755–759.
Crofts, D.R., 1929. Haliotis. Liverpool Marine Biological Commit-
tee Memoirs no. XXIX. University Press, Liverpool, pp. 50–76.
Dunstan, G.A., Baillie, H.J., Barrett, S.M., Volkman, J.K., 1996.
Effect of diet on the lipid composition of wild and cultured
abalone. Aquaculture 140, 115–127.
Ebert, E.E., Houk, J.L., 1984. Elements and innovations in the
culture of red abalone Haliotis rufescens. Aquaculture 39,
375–392.
Edwards, S., Condon, C., 2001. Digestive protease characterization,
localization and adaptation in blacklip abalone (Haliotis rubra
leach). Aquac. Res. 32, 95–102.
Erlanger, B.F., Kokowsky, N., Cohen, W., 1961. The preparation
and properties of two chromogenic substrates of trypsin. Arch.
Biochem. Biophys. 95, 271–278.
Fang, L.S., Lee, B.N., 1992. Ontogenetic change of the digestive
enzymes in Panaeus monodon. Comp. Biochem. Physiol. 103B,
1033–1037.
Fleming, A.E., Van Barneveld, R.J., Hone, P.W., 1996. The
development of artificial diets for abalone: a review and future
directions. Aquaculture 140, 5–53.
Gjellesvik, D.R., Lombardo, D., Walther, B.T., 1992. Pancreatic bile
salt dependent lipase from cod (Gadus morhua): purification
and properties. Biochim. Biophys. Acta 1124, 123–134.
Hahn, K.O., 1989a. Nutrition and growth of abalone. In: Hahn,
K.O. (Ed.), Handbook of Culture of Abalone and other Marine
Gastropods. CRC Press, Florida, pp. 135–156.
Hahn, K.O., 1989b. Larval development of abalone. In: Hahn, K.O.
(Ed.), Handbook of Culture of Abalone and other Marine
Gastropods. CRC Press, Florida, pp. 71–100.
Hammer, H.S., Bishop, C.D., Watts, S.A., 2000. Activities of three
digestive enzymes during development in the crayfish Procam-
barus clarkii (Decapoda). J. Crustac. Biol. 20, 614–620.
Harris, J.O., 1994. Digestive physiology of the greenlip abalone
Haliotis laevigata Donovan: a preliminary investigation of
digestive anatomy, microbial environment, occurrence and
algal digestion. Honours thesis: University of Tasmania,
Launceston.
Harris, J.O., Burke, C.M., Maguire, G.B., 1998. Characterization of
the digestive tract of greenlip abalone, Haliotis laevigata
Donovan: I. Morphology and histology. J. Shellfish Res. 17,
979–988.
Herbert, D.G., 1990. Designation of lectotype and type locality for
Haliotis rugosa Lamarack, 1822 (Mollusca: Gastropoda:
Haliotidae). Ann. Natal Mus. 31, 201–213.
Hernandez-Santoyo, A., Hernandez-Arana, A., Arrequin-Eespinosa,
R., Rodriquez-Romero, A., 1998. Purification and character-
ization of several digestive proteases from the blue abalone,
Haliotis fulgens. Aquaculture 159, 203–216.
Hickman, C.S., 1980. Gastropod radulae and the assessment of form
in evolutionary paleontology. Paleobiology 6, 276–294.
Jarayabhand, P., Paphavasit, N., 1996. A review of the culture of
tropical abalone with special reference to Thailand. Aquaculture
140, 159–168.
Johnston, D.J., 2003. Ontogenetic changes in digestive enzymology
of the spiny lobster, Jasus edwardsii hutton (Decapoda,
Palinuridae). Mar. Biol. 143, 1071–1082.
Johnston, D.J., Alexander, C.G., Yellowlees, D., 1993. Histology,
histochemistry and enzyme biochemistry of the digestive glands
in the tropical surf barnacle Tetraclita squamosa. J. Mar. Biol.
Assoc. U.K. 73, 1–14.
Kamarudin, M.S., Jones, D.A., le Vay, L., Abidin, A.Z., 1994.
Ontogenetic change in digestive enzyme activity during larval
development ofMacrobranchium rosenbergii. Aquaculture 123,
323–333.
Kawamura, T., Takami, H., Roberts, R.D., Yamashita, Y., 2001.
Radula development in abalone Haliotis discus hannai from
larval to adult in relation to feeding transitions. Fish. Sci. 67,
596–605.
Knauer, J., Britz, P., Hecht, T., 1996. Comparative growth
performance and digestive enzyme activity of juvenile South
African abalone, Haliotis midae, fed on diatoms and a practical
diet. Aquaculture 140, 75–85.
Leighton, D.L., 1974. The influence of temperature on larval and
juvenile growth in three species of Southern California
abalones. Fish. Bull. 72, 1137–1145.
McCandless, E.L., 1981. Polysaccharides of seaweeds. In: Lobban,
C.S., Wynne, M.J. (Eds.), The Biology of Seaweeds. University
of California Press, Berkeley, pp. 559–588.
McLean, N., 1970. Digestion in Haliotis rufescens Swainson
(Gastropoda: Prosobranchia). J. Exp. Zool. 173, 303–318.
McShane, P.E., 1992. Early life history of abalone: a review. In:
Shepherd, S.A., Tegner, M.J., Gunzman del Proo, S. (Eds.),
Abalone of the World: Biology, Fisheries and Culture. Fishing
News books, Cambridge, pp. 120–140.
Mori, T., 1953. Seaweed polysaccharides. Adv. Carbanion Chem. 8,
315–350.
Morton, J.E., 1979. Feeding and digestion. In: Morton, J.E. (Ed.),
Molluscs, 5th edition. The Anchor Press Ltd, Essex, pp. 97–123.
Oozeki, Y., Bailey, K.M., 1995. Ontogenetic development of
digestive enzymes activities in larval walleye pollock, Theragra
chalcogramma. Mar. Biol. 122, 177–186.
Padilla, D.K., 1985. The structural resistance of algae to herbivores,
a biomechanical approach. Mar. Biol. 90, 103–109.
Piscos-Garcia, C., Garcia-Carreno, F.L., Serviere-Zaragoza, E.,
2000. Digestive proteases in juvenile Mexican green abalone,
Haliotis fulgens. Aquaculture 181, 157–170.

D. Johnston et al. / Aquaculture 250 (2005) 341–355 355
Purchon, R.D., 1968. Digestion. In: Purchon, R.D. (Ed.), The
Biology of theMollusca. Pergamon Press, London, pp. 207–264.
Ragan, M.A., 1981. Chemical constituents of seaweeds. In: Lobban,
C.S., Wynne, M.J. (Eds.), The Biology of Seaweeds. University
of California Press, Berkeley, pp. 589–626.
Roberts, R.D., Kawamura, T., Takami, H., 1999. Morphological
changes in the radula of abalone (Haliotis iris) during post-
larval development. J. Shellfish Res. 18, 637–644.
Saito, K., 1981. The appearance and growth of 0-year-old Ezo
abalone. Bull. Jpn. Soc. Sci. Fish. 47, 1393–1400.
Serviere-Zaragoza, E., Nararrete del Toro, M.A., Garcia-Carreno,
F.L., 1997. Protein-hydrolysing enzymes in the digestive
systems of the adult Mexican blue abalone, Haliotis fulgens
(Gastropoda). Aquaculture 157, 325–336.
Shepherd, S.A., 1973. Studies on southern Australian abalone
(Genus Haliotis): I. Ecology of five sympatric species. Aust. J.
Mar. Freshw. Res. 24, 217–257.
Shepherd, S.A., Hearn, W.S., 1983. Studies on Southern Australian
abalone (Genus Haliotis): IV. Growth of H. laevigata and H.
ruber. Aust. J. Mar. Freshw. Res. 34, 461–475.
Shepherd, S.A., Turner, J.A., 1985. Studies on Southern
Australian abalone (Genus Haliotis): VI. Habitat preference,
abundance and predators of juveniles. J. Exp. Mar. Biol. Ecol.
93, 285–298.
Sollas, I.B.J., 1907. The molluscan radula: its chemical composition
and some points in its development. Queens J. Microsc. Sci. 51,
115–136.
Stone, S.T., Betz, A., Hofsteenge, J., 1991. Mechanistic studies on
thrombin catalysis. Biochemistry 30, 9841–9848.
Takami, H., Kawamura, T., 1998. Development of polysaccharide
degradation activity in postlarval abalone Haliotis discus
hannai. J. Shellfish Res. 17, 723–727.
Viana, M.T., Lopez, L.M., Salas, A., 1993. Diet development for
juvenile abalone Haliotis fulgens, evaluation of two artificial
diets and macroalgae. Aquaculture 117, 149–156.
Viana, M.T., Lopez, L.M., Garcia-Esquivel, Z., Mendez, E., 1996.
The use of silage made from fish and abalone viscera as an
ingredient in abalone feed. Aquaculture 140, 87–98.