the endovanilloid/endocannabinoid system: a new potential target for osteoporosis therapy
TRANSCRIPT

Bone 48 (2011) 997–1007
Contents lists available at ScienceDirect
Bone
j ourna l homepage: www.e lsev ie r.com/ locate /bone
The endovanilloid/endocannabinoid system: A new potential target forosteoporosis therapy
Francesca Rossi a, Giulia Bellini b, Livio Luongo b, Marco Torella c, Silvia Mancusi a,b, Luciano De Petrocellis d,Stefania Petrosino d, Dario Siniscalco b, Pierangelo Orlando d, Mariantonietta Scafuro b, Nicola Colacurci c,Silverio Perrotta a, Bruno Nobili a, Vincenzo Di Marzo d, Sabatino Maione b,⁎and Endocannabinoid Research Group (ERG), Italy b,d
a Department of Pediatrics, Second University of Naples, via De Crecchio 4, 80138 Naples, Italyb Department of Experimental Medicine, Second University of Naples, via Costantinopoli 16, 80138 Naples, Italyc Department of Gynecology, Obstetrics and Reproductive Sciences, Second University of Naples, Largo Madonna delle Grazie, 80138 Naples, Italyd Institute of Biomolecular Chemistry, Institute of Cybernetics and Institute of Protein Biochemistry National Research Council (CNR), via Campi Flegrei 34,80078 Pozzuoli and via P.castellino M, Naples, Italy
⁎ Corresponding author. Fax: +390815667503.E-mail addresses: [email protected] (F. Rossi), giu
(G. Bellini), [email protected] (L. Luongo), [email protected] (S. Mancusi), l.depetrocellis@[email protected] (S. Petrosino), [email protected]@ibp.cnr.it (P. Orlando), [email protected] (N. Colacurci), [email protected] (B. Nobili), [email protected]@unina2.it (S. Maione).
8756-3282/$ – see front matter © 2011 Published by Edoi:10.1016/j.bone.2011.01.001
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 July 2010Revised 20 December 2010Accepted 2 January 2011Available online 13 January 2011
Edited by: J. Aubin
Keywords:MenopauseHuman boneOsteoclastsVanilloid and cannabinoid receptorsBone-turnover markers
Human osteoclasts express functional TRPV1 channels, CB1/CB2 cannabinoid receptors and endocannabinoid/endovanilloid synthetic/catabolic enzymes. Pharmacologic manipulation of this system can modulateosteoclast activity. Here, through multidisciplinary approaches, we demonstrate that enzymes and receptorsof the endocannabinoid/endovanilloid system are differently expressed in osteoclasts from menopausalwomen without or with osteoporosis. We report that in osteoclasts from osteoporotic patients, TRPV1channels are upregulated and, if persistently stimulated with resiniferatoxin, become clustered to the plasmamembrane while inducing a massive over-expression of CB2 receptors. By providing new evidence for acritical functional cross-talk between CB2 and TRPV1 receptors in osteoporosis, we speculate that TRPV1desensitization, or its enhanced trafficking, together with TRPV1 agonist-induced CB2 receptor over-expression, might be critical to minimize calcium entry in osteoclasts, which could be in turn responsible ofcell over-activation and higher bone resorption. Our data pave the way to the use of TRPV1 agonist togetherwith CB2 agonists or CB1 antagonists in osteoporosis.
[email protected]@unina2.it (M. Torella),.na.cnr.it (L. De Petrocellis),edu (D. Siniscalco),[email protected] (M. Scafuro),[email protected] (S. Perrotta),nr.it (V. Di Marzo),
lsevier Inc.
© 2011 Published by Elsevier Inc.
Introduction
Osteoporosis (OP) is a skeletal disorder characterized by low bonemass and micro-architectural bone deterioration. The associatedfractures and the subsequent morbidity and mortality make of OP asan enormous public health concern [1–3]. The etiology of OP isheterogeneous, the main predictors are age, gender and a positivefamily history. Indeed, while OP is thought to be a polygenic disorderand affects both sexes, postmenopausal women are at the highest risk[4,5]. Postmenopausal OP represents a classical type of primary OP,which is characterized by a progressive bone mass loss, with an onsetoverlapping with the beginning of menopause, and fractures that
occur within 15–20 years from gonad function interruption [6].Moreover, it represents the bone disease with the highest epidemi-ological impact, which makes it one of the most important public andeconomic health problems. Although a low bone mass index, life-style, dietary habits and age-related bone mass loss contribute to theestablishment of OP, the main etiopathological mechanisms seem tobe hormone-related [7,8]. Treatment options include general mea-sures on lifestyle, calcium and vitamin D supplements, hormonetherapy, raloxifene, and bisphosphonates [9–11].
Recently, we provided evidence that human osteoclasts (OCs),derived byperipheral bloodmonocyte (PBMC) fromhealthy volunteers,express functional transient receptor potential vanilloid subtype 1(TRPV1) channels together with types 1 and 2 cannabinoid receptors(CB1/CB2) and anandamide (AEA) synthetic and catabolic enzymes,NAPE-PLD (N-acyl-phosphatidylethanolamine phospholipase) andFAAH (fatty acid amide hydrolase), respectively [12]. It was shownthat cannabinoid/vanilloid agonists, alone or in combination withselective antagonists, are able to modulate osteoclast formation andactivity. AEA has been recognized to have endovanilloid activity [13,14]and pharmacological blockade of FAAH, by enhancingAEA levels, causesTRPV1-mediated stimulatory effects on osteoclast formation and bone

998 F. Rossi et al. / Bone 48 (2011) 997–1007
turnover [12]. Consistent with the data from rodents pointing to theendovanilloid/endocannabinoid (EV/EC) system as a novel therapeutictarget for treating some bone diseases, i.e. OP, [15–19], in this study wehave considered it interesting to investigate the co-expression of CB1/CB2 receptors and TRPV1 channels, as well as the specific synthetic orcatabolic EV/EC enzymatic machinery, in OCs derived by PBMC frommenopausal women screened by Dual Energy X-ray Absorptiometry(DEXA) to measure Bone Mineral Density (BMD), and then categorizedusing WHO criteria. Accordingly, subjects from three different popula-tion were selected and three corresponding groups of OCs wereobtained: i) from control women (BMDb1.0 SD below the mean),ii) from women with osteopenia (1.0NBMDb2.5 SD), and iii) fromwomen with OP (BMDN2.5 SD). In these three different human OCscultures we have performed: 1) biomolecular and immunohistochem-ical analysis to reveal a possible alteration in the coexpression of CB1,CB2 and TRPV1; or in the expression of the enzymes NAPE-PLD andFAAH, but also of DAGL-α (diacylglycerol lipase alpha), and MAGL(monoacylglycerol lipase), involved in the biosynthesis and catabolism,respectively, of the endocannabinoid 2-arachydonoylglycerol (2-AG),which has little activity at TRPV1; 2) TRPV1 functional activity assays asmeasured by capsaicin-induced intracellular calcium entry in vitro; 3)measurements of the levels of the EV/EC AEA, the EV N-oleoylethano-lamine (OEA), the EC 2-AG, and the AEA congener, N-palmitoylethano-lamine (PEA), which, among others, can enhance AEA effects atcannabinoid and TRPV1 receptors; these measures were made withand without application of URB597, a FAAH blocker; 4) enzymatic andbiomolecular assay for evaluating the activity of Tartrate-Resistant AcidPhosphatase (TRAP), a specific biomarker of osteoclast activation, beforeand after pharmacologicalmanipulation of TRPV1, CB1 or CB2 receptors,or of the enzyme FAAH. Finally, immunohistochemical analyses toreveal co-expression of native CB1, CB2 and TRPV1 receptors in OCsfrom osteoporotic human bone were also performed.
Materials and methods
Patients
Post-menopausal women (n=142) admitted for the first time toMenopause Unit of Department of Gynaecology of the SecondUniversity of Naples, were enrolled after formal approval of theEthics Committee. A signed study informed consent was obtainedfrom all enrolled women. Patients were screened by DEXA to evaluatebone loss and to measure BMD. DEXA results were categorized on thebasis of T-scores and Z-scores using WHO criteria. According to thesecriteria, all enrolledmenopausal womenwere divided in 3 subgroups:OP (n=53) (BMDN2.5 SD below the mean for adult women);osteopenia (n=45) (1.0NBMDb2.5 SD); normal (n=44) (BMDb1.0SD). Past history of fractures and bone disease treatments wereexcluded. For each experimental procedure the following number ofpatients were used: cell-signalling, n=6 for each group; molecularbiology, n=15 for each group; endocannabinoid level measurements,n=6–8 for each group; immunocytochemistry, n=4 for each group;TRAP assays, n=6 for each group). Seventy-four patients weregenotyped for the rs35761398 variant of the CNR2 gene encoding forCB2 receptor. Moreover, pre-menopausal women (n=50) were alsoenrolled, in order to confirm previously reported data (n=18) [12]and to check genotype distributions of the investigated SNP. Allexperiments were conducted according to the Declaration of Helsinki.
Human cell cultures
OCs were obtained from PBMC of menopausal women anddifferentiated for 21 days as described [12]. The mature fullydifferentiated OCs, strongly adherent to plastic, were characterizedby reverse transcriptase-polymerase chain reaction (RT-PCR) for theexpression of the specific biomarker TRAP. Seeding density was:
4×107 cells for biomolecular experiments; 3×106 cells for immuno-cytochemical and calcium assay experiments; 5×105 cells for TRAPassay experiments.
RNA isolation
Before RNA extraction, cell cultures were trypsinized to removeearly or late OCs precursors. mRNA extractionwas performed by usingan RNA Tri-Reagent (Molecular Research Center Inc., Cincinnati, OH),according to the manufacturer's instructions. RNA concentrationswere determined by UV spectrophotometer (NanoDrop ND 1000,NanoDrop Technologies, LLC, Wilmington, USA).
DNA isolation
According to manufacturer's protocol, genomic DNA wasextracted from whole blood by using the Wizard Genomic kit(Promega, Milan, Italy) or from the residual phase of RNA extractionby using the Tri-Reagent (Molecular Research Center Inc., Cincin-nati, OH).
RT–PCR, semiquantitative amplification, real-time quantitative PCR
Reverse transcriptase from Avian Myeloblastosis Virus (Promega,Madison, WI, USA) was used [12]. Specific amplification reactions forTRAP (alias ACPS, GeneID54), TRPV1 (transcript variants 1 and 3)(GeneID7442), CB1 (isoforms a and b) (alias CNR1, GeneID1268),CB2 (alias CNR2 GeneID1269), FAAH (GeneID2166), NAPE-PLD(GeneID222236), DAGL-α (GeneID221955), MAGL (alias MGLL,GeneID11343), CRIP1A (alias CNRIP1 GeneID25927) and NF-κB(GeneID4720) and for the housekeeping β-actin cDNAs were carriedout. In addition, mRNA levels were normalized also with respect toosteoclast number.
Amplimers were resolved into 2.0% agarose gel, detected by the“Gel Doc 2000 UV System” (Bio-Rad, Hercules, CA, USA) and verifiedby sequencing using the Big-Dye Terminators reaction kit and an ABIPRISM 310 (Applied Biosystem, Forster City, USA).
Three serial 2× cDNA dilutions obtained from 300 ng and 750 ngtotal mRNA were amplified by real-time PCR, using Sybr green asfluorophore, in order to quantify the expression levels of CB1, CB2and TRPV1 respect to β-actin. Assays were performed in triplicate. Atwenty-five-microliter reaction contained: 2 μl cDNA, 12.5 μl Sybrgreen Master Mix (Biorad), 10 μl primers mix (10 mM). The thermalcyclingprogramwas:95 °C–10 min, followedby40cyclesof95 °C–15 sand 60 °C–1 min. Gene expression profiling was achieved using thecomparative cycle threshold method of relative quantization to thehousekeeping gene. Real-time PCR products were analyzed by Icyclersoftware (Biorad).
TRAP assay
TRAPwas valuated as specific OCs biochemical activity marker andquantified using the ACP method (Takara Bio, Japan) as previouslydescribed [12]. Experiments included positive and negative controls.
Cell counting
We counted TRAP(+) multinucleated (n≥3) OCs in threedifferent wells per each group of treatment using an opticalmicroscope (Nikon Eclipse TS100, Nikon Instruments, Badhoevedorp,The Netherlands).
Western blot
TRPV1 channels and CB2 receptors in total lysates from osteoclastcultures were analyzed by Western blot experiment. Membrane strips

999F. Rossi et al. / Bone 48 (2011) 997–1007
were incubated overnight at 4 °C with goat polyclonal anti-TRPV1antibody (1:200 dilution; sc-12502, Santa Cruz, CA, USA) or rabbitpolyclonal anti-CB2 (1:1000; Abcam, Cambridge, UK); reactive bandswere detected by chemiluminescence (SuperSignal,West Femto, Pierce,USA) on a ChemiDoc station (BioRad). An anti-β-actin (1:5000; Sigma,Milan, Italy) was used to check for identical protein loading and ashousekeeping protein. In addition, protein levels were normalized alsowith respect to osteoclast number. Images were captured, stored, andanalyzed with the Quantity One software (BioRad).
Immunofluorescence
OCs cultures were prepared as described previously [12]. OCs wereincubated 3 hours with goat polyclonal anti-TRPV1 (1:100; Santa CruzBiotechnology, CA, USA); rabbit polyclonal TRAP (1:100; Santa CruzBiotechnology, CA, USA) and mouse vimentine (1:1000; Abcam,Cambridge,UK). Secondarydonkeyantibodyanti-rabbit IgG-conjugatedAlexa FluorTM488, or antigoat IgG-conjugated Alexa FluorTM568, oranti-mouse IgG-conjugated Alexa FluorTM350 (1:1000;MolecularProbes, USA) was used.
In vitro calcium assay
The effect of test substances on intracellular [Ca2+] was deter-mined by using Fluo-4, a selective intracellular fluorescent probe forCa2+ [12]. Experiments were carried out by measuring cell fluores-cence at 25 °C (λEX=488 nm, λEM=516 nm) before and after theaddition of the test compounds at various concentrations. Agonistactivity was determined in comparison to the maximum increase ofintracellular Ca2+ due to the application of 4 μM ionomycin (AlexisBiochemicals, Lausen Switzerland). EC50 values were determined asthe concentration of test substances required to produce half-maximal increase in [Ca2+]i. All determinations were performed atleast in triplicate. Curve fitting (sigmoidal dose–response variableslope) and parameter estimation was performed with GraphPadPrism® (GraphPad Software Inc., San Diego, CA). Statistical analysis ofthe data was performed at each point using ANOVA followed byBonferroni's test.
Endocannabinoid extraction and quantification
Cell cultures were treated with URB597 [1nM] or vehicle (DMSO)for 10 min and endocannabinoid extracted as previously described[12]. Results were expressed as pmol/mg of extracted lipids.
CNR2 rs35761398 genotyping
Seventy-four patients and 50 healthy female volunteers werescreened for the CNR2 rs35761398 variant. Detection of thepolymorphism was performed by PCR followed by direct sequencing.PCR, consisting of an initial denaturation at 94 °C for 4 min followedby 31 cycles consisting of 94 °C for 30 s, 60 °C for 30 s and 72 °C for30 s, generated an amplimer of 280 bp. Primers were picked by usingPrimer3 software (Forward 5′-GAGTGGTCCCCAGAAGACAG-3′ andReverse 5’′-CACAGAGGCTGTGAAGGTCA-3′). PCR products were ana-lyzed by direct sequencing using an ABI PRISM 310 automatedsequencer (Applied Biosystem, Foster City, CA).
TRPV1 silencing
TRPV1mRNAwas silenced by using human TRPV1 Accell SMARTpoolsiRNA reagent (Dharmacon, Thermo Scientific, Chicago, IL). Celltransfectionwasperformedaccording tomanufacturer'sprotocol. Briefly,OCs were differentiated as described above and plated on 96-well plateswith α-MEM and 10% fetal bovine serum (FBS) at density 50,000 cells/well. Cells were then transfected by using 1 μM control or experimental
Accell siRNA combined in Accell delivery 1%-supplemented FBSmedium.Cells were incubated at 37 °C in a CO2 incubator for 4 days, after whichthey were collected for biomolecular examination. Transfection minuscontrolswere carried out using non-targeting siRNA inorder to check theefficiency of transfection. I-RTX 2.5 μM was added after 4 days for48 hours in order to evaluate TRAP levels reduction.
Human bone immunohistochemistry
Human bone samples were kindly supplied by the Department ofOrthopaedics of the Second University of Naples after the formalapproval of the Ethics Committee.
Bone samples were obtained from osteoporotic women whounderwent elective bone surgeries for accidental fractures andprocessed as described [12]. A signed informed consent was obtainedfrom each patient.
Statistics
Molecular data are shown as means±SD. One-way ANOVAfollowed by Student–Newman–Keuls post hoc test was used. Biomo-lecular and biochemical data have been normalized with respect tohousekeeping gene, protein and osteoclast number. Cell counting dataare shown as means±SEM. One-way ANOVA, followed by Tukey's posthoc test was used. pb0.05 was considered statistically significant.Regression analysis was performed by using Stat-Graphic software.
Drugs and treatments
Resiniferatoxin (RTX), 5′-iodo-resiniferatoxin (I-RTX), capsaicin,AM251, AM630, JWH-133, ACEA (Tocris, Avonmouth, UK) andURB597(Alexis Biochemicals, Lausen Switzerland) were dissolved in PBScontaining DMSO. DMSO final concentration on cultures was 0.01%.OCs were treated with RTX, I-RTX, AM251, AM630, JWH-133, ACEA orURB597, alone or in combination, for 48 hours. Unless otherwisestated, the concentrations used for each drug was determinedfollowing pilot dose–response experiments and were the onesproducing the strongest effect. For single treatment cultured OCswere treated with RTX [0.1–1.25–2.5–5–7–10 μM], I-RTX [1 or 3 μM],AM251 [10 μM], AM630 [10 μM], JWH-133 [5–10–20 μM], ACEA [10–20–40 nM] or URB597 [1nM]. For co-treatment, RTX [2.5 or 5 μM]wasadded 5 min after application of I-RTX [1 or 3 μM], alone or incombination with AM630 [10 μM] and/or AM251 [10 μM]. For the RTX[5 μM]/AM630 or AM251 [10 μM] co-treatment, the cannabinoidantagonists were co-applied with RTX or added after 16 hours. Theapplication of URB597 [1nM] was blocked after 10 min by changingthe culture medium. Duration of challenge was the same for alltreatments. Cells were cultured for at least 21 days. This time intervalis enough to observe a complete osteoclast differentiation. Treatmentswere applied for 48 hours only when OCs were fully differentiated.Non-treated cultured OCs were maintained in incubation mediaduring the relative treatment time with or without vehicle (DMSO0.01%). Differences in OCs number rather than in differentiationdegreewere observed amongpatients relatively to T-score and Z-scorevalues. Ligandswere added on 22th day. RNA extraction or TRAP assayor immunostaining were performed 48 hours after each treatment.
Results
Expression of TRPV1 channels, CB1/CB2 receptors, FAAH, NAPE-PLD,DAGL-α, MAGL and TRAP mRNAs in post-menopausal human OCs inculture
RT-PCR analysis followed by the cDNA amplification for theosteoclastic marker TRAP demonstrated that cells in culture wereOCs. A regression analysis between TRAP expression and T-score and

1000 F. Rossi et al. / Bone 48 (2011) 997–1007
Z-score values suggested that TRAP levels were significantly related toBMD (supplementary data, S1).
RT-PCR showed also the presence of mature mRNA for TRPV1, CB1/CB2 receptors, as well as for the EV/EC metabolic enzymes NAPE-PLD,FAAH, DAGL-α and MAGL (supplementary data, S2). In particular, CB1expression could be detected starting from 100 ng mRNA for the RTreaction and both the long and short CB1 isoforms, namely CB1a andCB1b [20] were expressed, whereas we had previously shown that onlythe CB1a band could be detected in healthy OCs and only by using aminimal amount of 300 ngmRNA [12]. Semiquantitative PCR revealed alower expression of CB2, which was previously found to be fourfoldexpressed with respect to CB1 in healthy OCs [12]. Therefore, aquantitative PCR analysis was performed in order to better investigatethe relative expression of CB1, CB2 and TRPV1 in the three differentgroups (menopause, osteopenic, osteoporotic). We observed that CB1expression was enhanced in osteoporotic OCs, as compared toosteopenic or non-osteopenic OCs (Fig. 1A). Moreover, specificamplification for CB1a and CB1b evidenced that the CB1b isoform wasover-expressed (Fig. 1B). It is noteworthy that the CB2 expression wasfound unmodified in all of the three osteoclast groups (Fig. 1C).Accordingly, a regression analysis between CB2 levels and T-score andZ-score values showed lack of association between CB2 expressionwithrespect to BMD (supplementary data, S3). TRPV1 expression wassignificantly up-regulated in OCs from both osteopenic and osteopo-rotic women (Fig. 1D). In particular, this increase in TRPV1mRNA levelsresulted from a higher expression of the TRPV1 variant 1 (Fig. 1E),whereas we have previously shown that healthy OCs preferentiallyexpress TRPV1 variant 3 [12]. The difference in the expression of FAAH,NAPE-PLD, DAGL-α and MAGL was also analyzed (Fig. 1F, G). ThemRNA levels of DAGLα and MAGL, involved in the biosynthesis and
Fig. 1. Expression of receptors and enzymes of the endocannabinoid/endovanilloid system inisoform b, Cannabinoid Receptor type 2 (CB2), transient receptor potential vanilloid type-1(MAGL), N-acyl phosphatidylethanolamine phospholipase D (NAPE-PLD) and fatty acid amidfrom 300 ng of total mRNA for the RT reaction.
catabolism of the endocannabinoid 2-AG, respectively, resulted inenhanced OCs from both osteopenic and osteoporotic women,indicating an increased turnover of 2-AG in the pathological group(Fig. 1F). The expression of the NAPE-PLD resulted unmodified in all ofthe osteoclast groups, but the signal for FAAH resulted dramaticallydecreased in osteoporotic OCs (Fig. 1G). Normalization with respect tohousekeeping gene and osteoclast number showed same results.
TRPV1 pharmacological modulation affects the expression of TRAP inosteoporotic OCs
The osteoclast biomarker TRAP was significantly enhanced in OCsfrom post-menopausal women respect to pre-menopausal. In particu-lar, osteoporotic OCs, as compared to both osteopenic and nonosteoporotic, showed the highest values (pre-menopausal 0.37±0.12;menopause 1±0.11; osteopenia 1.29±0.23; OP 1.48±0.08) (Fig. 2A).Pharmacological treatment with the TRPV1 agonist RTX at differentconcentrations [0.1–2.5–5–7 μM] caused a dose-related reduction inTRAPmRNA levels (Fig. 2B), in contrast with our previous data showingthat TRPV1 agonism was able to enhance TRAP expression in healthyOCs [12]. Also the selective TRPV1 antagonist I-RTX [2.5 μM] inhibitedTRAP expression in both menopausal and osteoporotic OCs (Fig. 2B),similar to what was previously found in OCs from healthy subjects [12].I-RTX [2.5 μM] partly reversed the inhibitory effect of RTX inosteoporotic OCs.
TRPV1 receptors in osteoporotic activated OCs
TRPV1 channel was up-regulated in OCs derived from osteoporoticwomen as compared to the menopausal-derived cells. The increased
human osteoclast cultures: Expression of total Cannabinoid Receptor type 1 (CB1), CB1(TRPV1), TRPV1 variant 1, diacylglycerol lipase alpha (DAGLA), monoglyceride lipasee hydrolase (FAAH) mRNAs in human OCs in culture revealed by real-time PCR starting
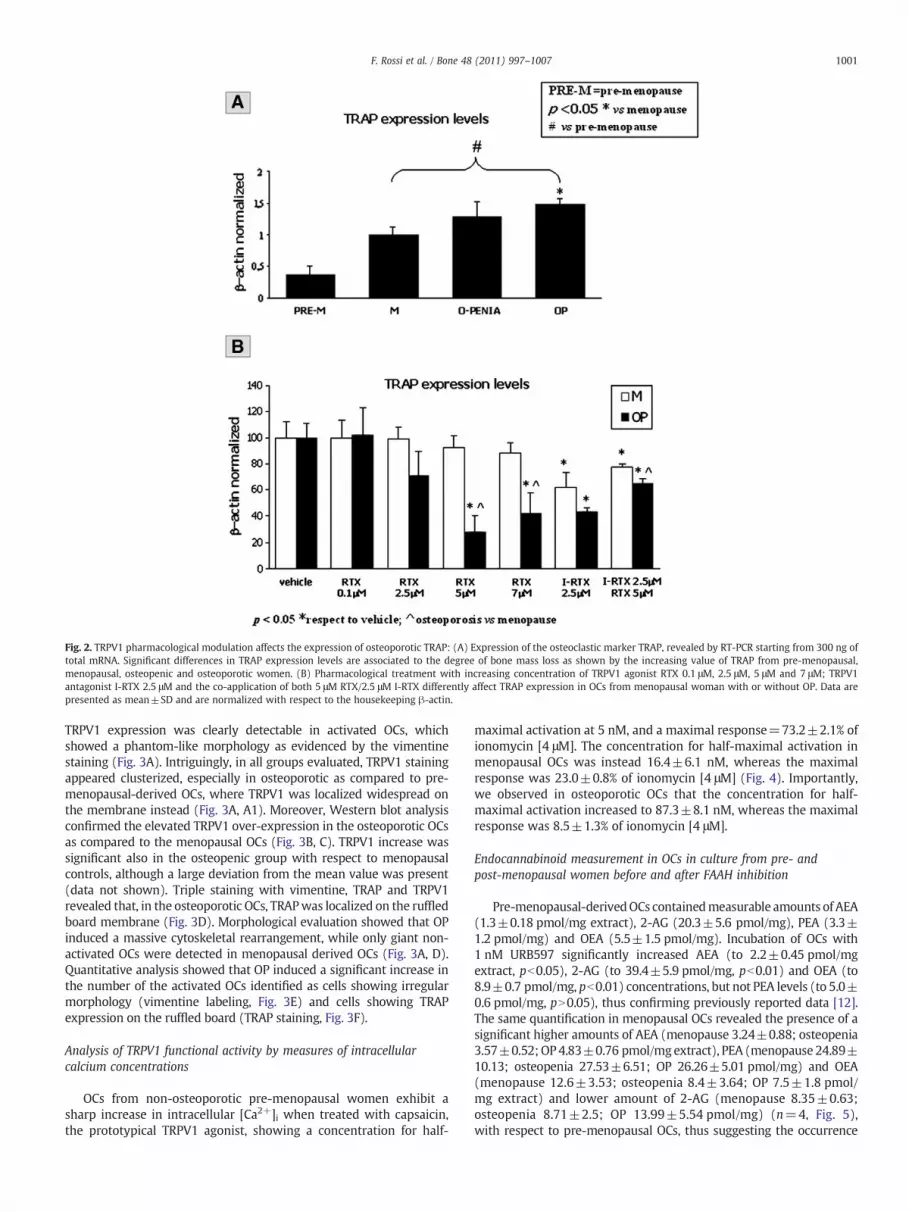
Fig. 2. TRPV1 pharmacological modulation affects the expression of osteoporotic TRAP: (A) Expression of the osteoclastic marker TRAP, revealed by RT-PCR starting from 300 ng oftotal mRNA. Significant differences in TRAP expression levels are associated to the degree of bone mass loss as shown by the increasing value of TRAP from pre-menopausal,menopausal, osteopenic and osteoporotic women. (B) Pharmacological treatment with increasing concentration of TRPV1 agonist RTX 0.1 μM, 2.5 μM, 5 μM and 7 μM; TRPV1antagonist I-RTX 2.5 μM and the co-application of both 5 μM RTX/2.5 μM I-RTX differently affect TRAP expression in OCs from menopausal woman with or without OP. Data arepresented as mean±SD and are normalized with respect to the housekeeping β-actin.
1001F. Rossi et al. / Bone 48 (2011) 997–1007
TRPV1 expression was clearly detectable in activated OCs, whichshowed a phantom-like morphology as evidenced by the vimentinestaining (Fig. 3A). Intriguingly, in all groups evaluated, TRPV1 stainingappeared clusterized, especially in osteoporotic as compared to pre-menopausal-derived OCs, where TRPV1 was localized widespread onthe membrane instead (Fig. 3A, A1). Moreover, Western blot analysisconfirmed the elevated TRPV1 over-expression in the osteoporotic OCsas compared to the menopausal OCs (Fig. 3B, C). TRPV1 increase wassignificant also in the osteopenic group with respect to menopausalcontrols, although a large deviation from the mean value was present(data not shown). Triple staining with vimentine, TRAP and TRPV1revealed that, in the osteoporotic OCs, TRAPwas localized on the ruffledboard membrane (Fig. 3D). Morphological evaluation showed that OPinduced a massive cytoskeletal rearrangement, while only giant non-activated OCs were detected in menopausal derived OCs (Fig. 3A, D).Quantitative analysis showed that OP induced a significant increase inthe number of the activated OCs identified as cells showing irregularmorphology (vimentine labeling, Fig. 3E) and cells showing TRAPexpression on the ruffled board (TRAP staining, Fig. 3F).
Analysis of TRPV1 functional activity by measures of intracellularcalcium concentrations
OCs from non-osteoporotic pre-menopausal women exhibit asharp increase in intracellular [Ca2+]i when treated with capsaicin,the prototypical TRPV1 agonist, showing a concentration for half-
maximal activation at 5 nM, and a maximal response=73.2±2.1% ofionomycin [4 μM]. The concentration for half-maximal activation inmenopausal OCs was instead 16.4±6.1 nM, whereas the maximalresponse was 23.0±0.8% of ionomycin [4 μM] (Fig. 4). Importantly,we observed in osteoporotic OCs that the concentration for half-maximal activation increased to 87.3±8.1 nM, whereas the maximalresponse was 8.5±1.3% of ionomycin [4 μM].
Endocannabinoid measurement in OCs in culture from pre- andpost-menopausal women before and after FAAH inhibition
Pre-menopausal-derivedOCs containedmeasurable amounts of AEA(1.3±0.18 pmol/mg extract), 2-AG (20.3±5.6 pmol/mg), PEA (3.3±1.2 pmol/mg) and OEA (5.5±1.5 pmol/mg). Incubation of OCs with1 nM URB597 significantly increased AEA (to 2.2±0.45 pmol/mgextract, pb0.05), 2-AG (to 39.4±5.9 pmol/mg, pb0.01) and OEA (to8.9±0.7 pmol/mg, pb0.01) concentrations, but not PEA levels (to 5.0±0.6 pmol/mg, pN0.05), thus confirming previously reported data [12].The same quantification in menopausal OCs revealed the presence of asignificant higher amounts of AEA (menopause 3.24±0.88; osteopenia3.57±0.52; OP4.83±0.76 pmol/mg extract), PEA (menopause 24.89±10.13; osteopenia 27.53±6.51; OP 26.26±5.01 pmol/mg) and OEA(menopause 12.6±3.53; osteopenia 8.4±3.64; OP 7.5±1.8 pmol/mg extract) and lower amount of 2-AG (menopause 8.35±0.63;osteopenia 8.71±2.5; OP 13.99±5.54 pmol/mg) (n=4, Fig. 5),with respect to pre-menopausal OCs, thus suggesting the occurrence
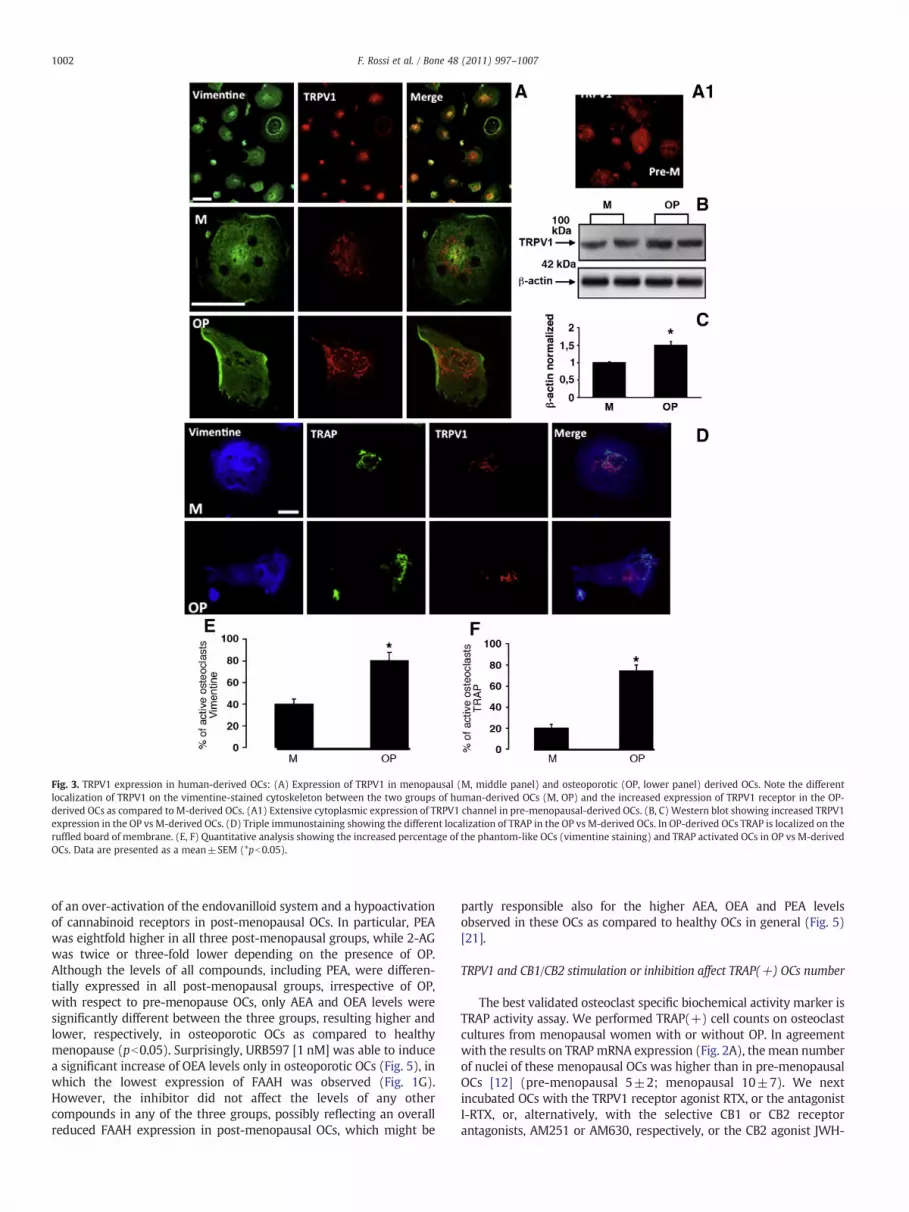
Fig. 3. TRPV1 expression in human-derived OCs: (A) Expression of TRPV1 in menopausal (M, middle panel) and osteoporotic (OP, lower panel) derived OCs. Note the differentlocalization of TRPV1 on the vimentine-stained cytoskeleton between the two groups of human-derived OCs (M, OP) and the increased expression of TRPV1 receptor in the OP-derived OCs as compared to M-derived OCs. (A1) Extensive cytoplasmic expression of TRPV1 channel in pre-menopausal-derived OCs. (B, C) Western blot showing increased TRPV1expression in the OP vs M-derived OCs. (D) Triple immunostaining showing the different localization of TRAP in the OP vs M-derived OCs. In OP-derived OCs TRAP is localized on theruffled board of membrane. (E, F) Quantitative analysis showing the increased percentage of the phantom-like OCs (vimentine staining) and TRAP activated OCs in OP vs M-derivedOCs. Data are presented as a mean±SEM (*pb0.05).
1002 F. Rossi et al. / Bone 48 (2011) 997–1007
of an over-activation of the endovanilloid system and a hypoactivationof cannabinoid receptors in post-menopausal OCs. In particular, PEAwas eightfold higher in all three post-menopausal groups, while 2-AGwas twice or three-fold lower depending on the presence of OP.Although the levels of all compounds, including PEA, were differen-tially expressed in all post-menopausal groups, irrespective of OP,with respect to pre-menopause OCs, only AEA and OEA levels weresignificantly different between the three groups, resulting higher andlower, respectively, in osteoporotic OCs as compared to healthymenopause (pb0.05). Surprisingly, URB597 [1 nM] was able to inducea significant increase of OEA levels only in osteoporotic OCs (Fig. 5), inwhich the lowest expression of FAAH was observed (Fig. 1G).However, the inhibitor did not affect the levels of any othercompounds in any of the three groups, possibly reflecting an overallreduced FAAH expression in post-menopausal OCs, which might be
partly responsible also for the higher AEA, OEA and PEA levelsobserved in these OCs as compared to healthy OCs in general (Fig. 5)[21].
TRPV1 and CB1/CB2 stimulation or inhibition affect TRAP(+) OCs number
The best validated osteoclast specific biochemical activity marker isTRAP activity assay. We performed TRAP(+) cell counts on osteoclastcultures from menopausal women with or without OP. In agreementwith the results on TRAP mRNA expression (Fig. 2A), the mean numberof nuclei of these menopausal OCs was higher than in pre-menopausalOCs [12] (pre-menopausal 5±2; menopausal 10±7). We nextincubated OCs with the TRPV1 receptor agonist RTX, or the antagonistI-RTX, or, alternatively, with the selective CB1 or CB2 receptorantagonists, AM251 or AM630, respectively, or the CB2 agonist JWH-
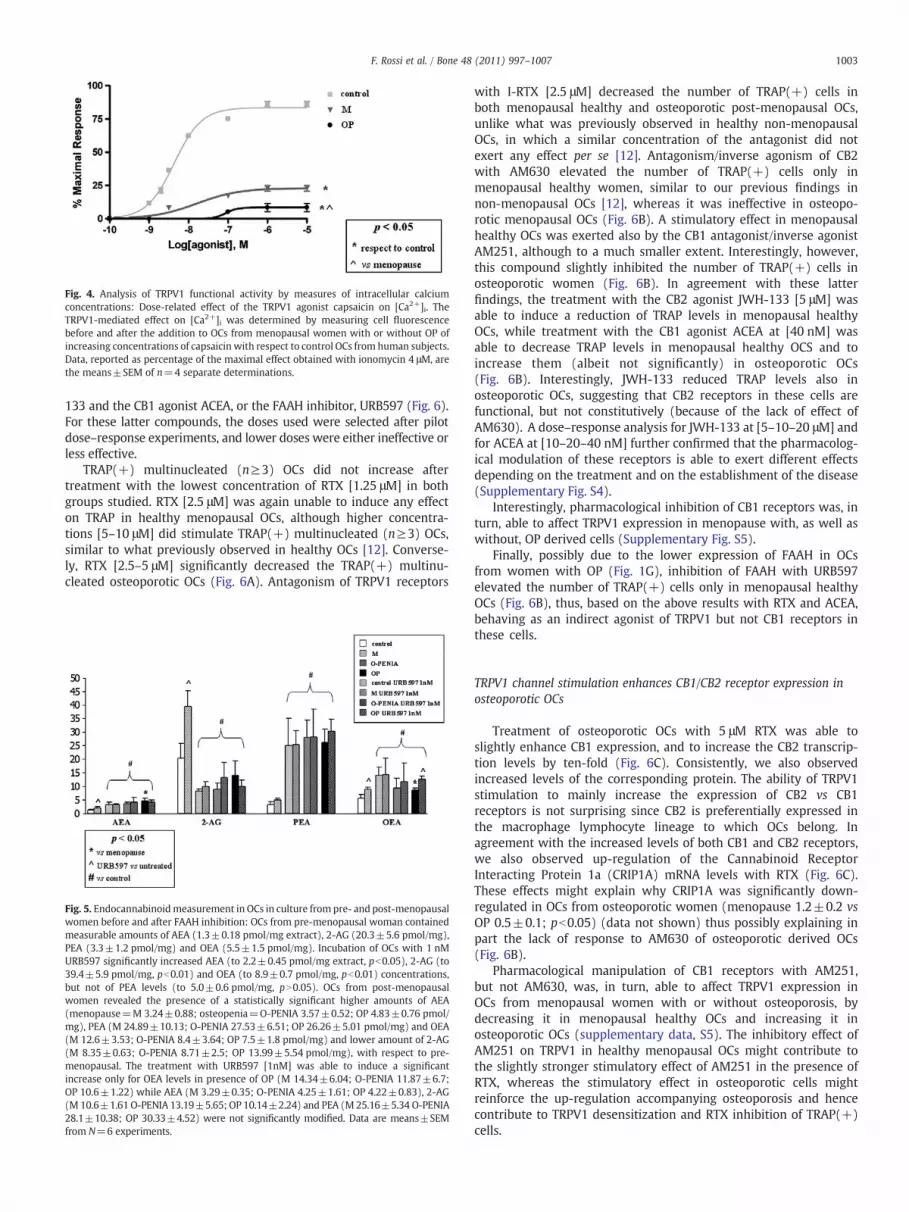
Fig. 4. Analysis of TRPV1 functional activity by measures of intracellular calciumconcentrations: Dose-related effect of the TRPV1 agonist capsaicin on [Ca2+]i. TheTRPV1-mediated effect on [Ca2+]i was determined by measuring cell fluorescencebefore and after the addition to OCs from menopausal women with or without OP ofincreasing concentrations of capsaicin with respect to control OCs from human subjects.Data, reported as percentage of the maximal effect obtained with ionomycin 4 μM, arethe means±SEM of n=4 separate determinations.
1003F. Rossi et al. / Bone 48 (2011) 997–1007
133 and the CB1 agonist ACEA, or the FAAH inhibitor, URB597 (Fig. 6).For these latter compounds, the doses used were selected after pilotdose–response experiments, and lower doses were either ineffective orless effective.
TRAP(+) multinucleated (n≥3) OCs did not increase aftertreatment with the lowest concentration of RTX [1.25 μM] in bothgroups studied. RTX [2.5 μM] was again unable to induce any effecton TRAP in healthy menopausal OCs, although higher concentra-tions [5–10 μM] did stimulate TRAP(+) multinucleated (n≥3) OCs,similar to what previously observed in healthy OCs [12]. Converse-ly, RTX [2.5–5 μM] significantly decreased the TRAP(+) multinu-cleated osteoporotic OCs (Fig. 6A). Antagonism of TRPV1 receptors
Fig. 5. Endocannabinoidmeasurement in OCs in culture from pre- and post-menopausalwomen before and after FAAH inhibition: OCs from pre-menopausal woman containedmeasurable amounts of AEA (1.3±0.18 pmol/mg extract), 2-AG (20.3±5.6 pmol/mg),PEA (3.3±1.2 pmol/mg) and OEA (5.5±1.5 pmol/mg). Incubation of OCs with 1 nMURB597 significantly increased AEA (to 2.2±0.45 pmol/mg extract, pb0.05), 2-AG (to39.4±5.9 pmol/mg, pb0.01) and OEA (to 8.9±0.7 pmol/mg, pb0.01) concentrations,but not of PEA levels (to 5.0±0.6 pmol/mg, pN0.05). OCs from post-menopausalwomen revealed the presence of a statistically significant higher amounts of AEA(menopause=M 3.24±0.88; osteopenia=O-PENIA 3.57±0.52; OP 4.83±0.76 pmol/mg), PEA (M 24.89±10.13; O-PENIA 27.53±6.51; OP 26.26±5.01 pmol/mg) and OEA(M 12.6±3.53; O-PENIA 8.4±3.64; OP 7.5±1.8 pmol/mg) and lower amount of 2-AG(M 8.35±0.63; O-PENIA 8.71±2.5; OP 13.99±5.54 pmol/mg), with respect to pre-menopausal. The treatment with URB597 [1nM] was able to induce a significantincrease only for OEA levels in presence of OP (M 14.34±6.04; O-PENIA 11.87±6.7;OP 10.6±1.22) while AEA (M 3.29±0.35; O-PENIA 4.25±1.61; OP 4.22±0.83), 2-AG(M 10.6±1.61 O-PENIA 13.19±5.65; OP 10.14±2.24) and PEA (M 25.16±5.34 O-PENIA28.1±10.38; OP 30.33±4.52) were not significantly modified. Data are means±SEMfrom N=6 experiments.
with I-RTX [2.5 μM] decreased the number of TRAP(+) cells inboth menopausal healthy and osteoporotic post-menopausal OCs,unlike what was previously observed in healthy non-menopausalOCs, in which a similar concentration of the antagonist did notexert any effect per se [12]. Antagonism/inverse agonism of CB2with AM630 elevated the number of TRAP(+) cells only inmenopausal healthy women, similar to our previous findings innon-menopausal OCs [12], whereas it was ineffective in osteopo-rotic menopausal OCs (Fig. 6B). A stimulatory effect in menopausalhealthy OCs was exerted also by the CB1 antagonist/inverse agonistAM251, although to a much smaller extent. Interestingly, however,this compound slightly inhibited the number of TRAP(+) cells inosteoporotic women (Fig. 6B). In agreement with these latterfindings, the treatment with the CB2 agonist JWH-133 [5 μM] wasable to induce a reduction of TRAP levels in menopausal healthyOCs, while treatment with the CB1 agonist ACEA at [40 nM] wasable to decrease TRAP levels in menopausal healthy OCS and toincrease them (albeit not significantly) in osteoporotic OCs(Fig. 6B). Interestingly, JWH-133 reduced TRAP levels also inosteoporotic OCs, suggesting that CB2 receptors in these cells arefunctional, but not constitutively (because of the lack of effect ofAM630). A dose–response analysis for JWH-133 at [5–10–20 μM] andfor ACEA at [10–20–40 nM] further confirmed that the pharmacolog-ical modulation of these receptors is able to exert different effectsdepending on the treatment and on the establishment of the disease(Supplementary Fig. S4).
Interestingly, pharmacological inhibition of CB1 receptors was, inturn, able to affect TRPV1 expression in menopause with, as well aswithout, OP derived cells (Supplementary Fig. S5).
Finally, possibly due to the lower expression of FAAH in OCsfrom women with OP (Fig. 1G), inhibition of FAAH with URB597elevated the number of TRAP(+) cells only in menopausal healthyOCs (Fig. 6B), thus, based on the above results with RTX and ACEA,behaving as an indirect agonist of TRPV1 but not CB1 receptors inthese cells.
TRPV1 channel stimulation enhances CB1/CB2 receptor expression inosteoporotic OCs
Treatment of osteoporotic OCs with 5 μM RTX was able toslightly enhance CB1 expression, and to increase the CB2 transcrip-tion levels by ten-fold (Fig. 6C). Consistently, we also observedincreased levels of the corresponding protein. The ability of TRPV1stimulation to mainly increase the expression of CB2 vs CB1receptors is not surprising since CB2 is preferentially expressed inthe macrophage lymphocyte lineage to which OCs belong. Inagreement with the increased levels of both CB1 and CB2 receptors,we also observed up-regulation of the Cannabinoid ReceptorInteracting Protein 1a (CRIP1A) mRNA levels with RTX (Fig. 6C).These effects might explain why CRIP1A was significantly down-regulated in OCs from osteoporotic women (menopause 1.2±0.2 vsOP 0.5±0.1; pb0.05) (data not shown) thus possibly explaining inpart the lack of response to AM630 of osteoporotic derived OCs(Fig. 6B).
Pharmacological manipulation of CB1 receptors with AM251,but not AM630, was, in turn, able to affect TRPV1 expression inOCs from menopausal women with or without osteoporosis, bydecreasing it in menopausal healthy OCs and increasing it inosteoporotic OCs (supplementary data, S5). The inhibitory effect ofAM251 on TRPV1 in healthy menopausal OCs might contribute tothe slightly stronger stimulatory effect of AM251 in the presence ofRTX, whereas the stimulatory effect in osteoporotic cells mightreinforce the up-regulation accompanying osteoporosis and hencecontribute to TRPV1 desensitization and RTX inhibition of TRAP(+)cells.
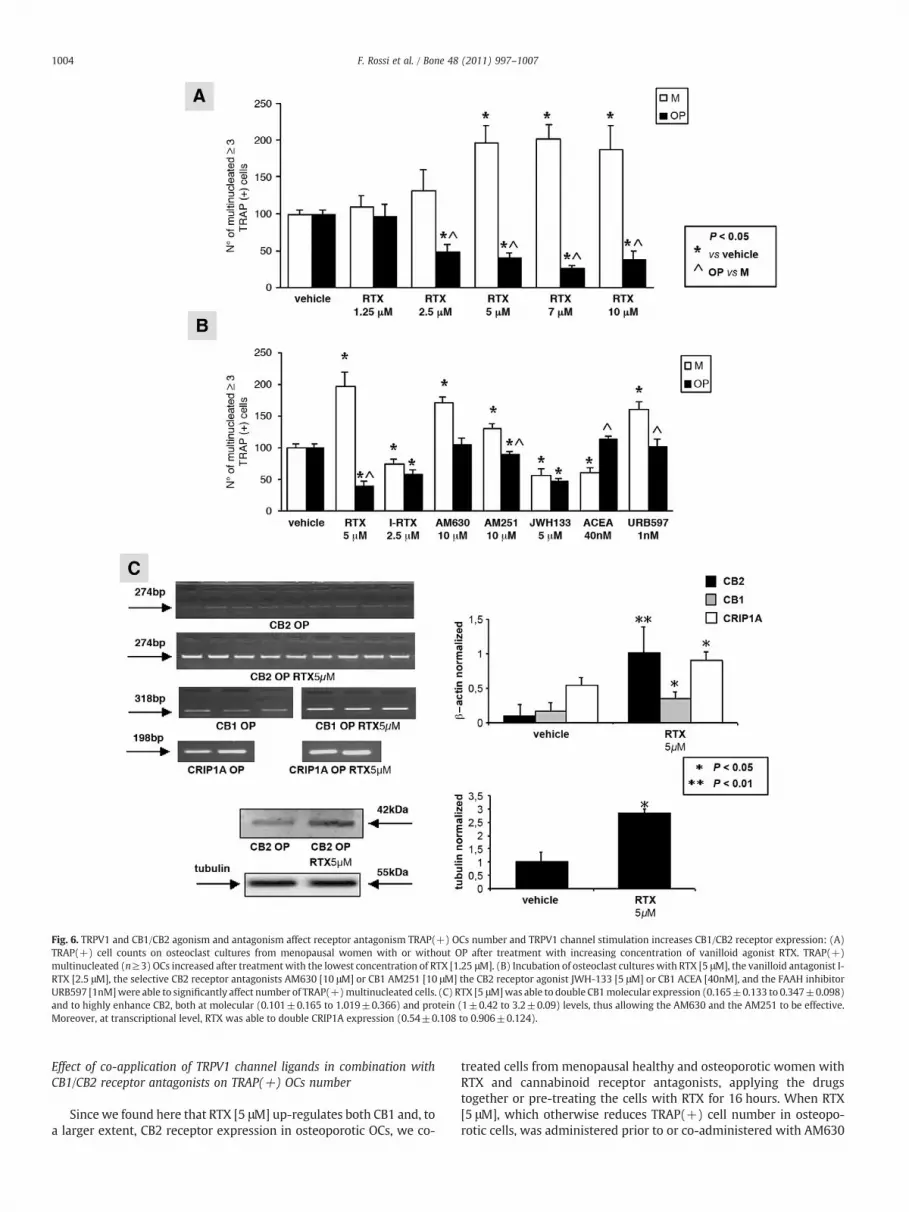
Fig. 6. TRPV1 and CB1/CB2 agonism and antagonism affect receptor antagonism TRAP(+) OCs number and TRPV1 channel stimulation increases CB1/CB2 receptor expression: (A)TRAP(+) cell counts on osteoclast cultures from menopausal women with or without OP after treatment with increasing concentration of vanilloid agonist RTX. TRAP(+)multinucleated (n≥3) OCs increased after treatment with the lowest concentration of RTX [1.25 μM]. (B) Incubation of osteoclast cultures with RTX [5 μM], the vanilloid antagonist I-RTX [2.5 μM], the selective CB2 receptor antagonists AM630 [10 μM] or CB1 AM251 [10 μM] the CB2 receptor agonist JWH-133 [5 μM] or CB1 ACEA [40nM], and the FAAH inhibitorURB597 [1nM]were able to significantly affect number of TRAP(+)multinucleated cells. (C) RTX [5 μM]was able to double CB1molecular expression (0.165±0.133 to 0.347±0.098)and to highly enhance CB2, both at molecular (0.101±0.165 to 1.019±0.366) and protein (1±0.42 to 3.2±0.09) levels, thus allowing the AM630 and the AM251 to be effective.Moreover, at transcriptional level, RTX was able to double CRIP1A expression (0.54±0.108 to 0.906±0.124).
1004 F. Rossi et al. / Bone 48 (2011) 997–1007
Effect of co-application of TRPV1 channel ligands in combination withCB1/CB2 receptor antagonists on TRAP(+) OCs number
Since we found here that RTX [5 μM] up-regulates both CB1 and, toa larger extent, CB2 receptor expression in osteoporotic OCs, we co-
treated cells from menopausal healthy and osteoporotic women withRTX and cannabinoid receptor antagonists, applying the drugstogether or pre-treating the cells with RTX for 16 hours. When RTX[5 μM], which otherwise reduces TRAP(+) cell number in osteopo-rotic cells, was administered prior to or co-administered with AM630

1005F. Rossi et al. / Bone 48 (2011) 997–1007
[10 μM], which causes no effect on TRAP(+) cell number in thesecells, a strong increase of TRAP(+) cells was observed (Figs. 7, S6),whereas the effect in menopausal healthy OCs, in which both drugselevate TRAP(+) cell number, was not different from the effect ofAM630 alone. The effect of RTX+AM630 was abolished by co-treatment with I-RTX [2.5 μM], in both osteoporotic and menopausalOCs. However, I-RTX also antagonized the effects of AM630 alone. Co-incubation or pre-treatment with RTX [5 μM] also significantlyenhanced AM251 inhibitory effect on osteoporotic cells, and slightly,although not significantly, increased the effect of the antagonist inhealthy OCs. Antagonism of TRPV1 with I-RTX [2.5 μM] antagonizedalso these effects of RTX on AM251, and, in the absence of RTX, insteadattenuated the effect of AM251 in menopausal healthy OCs butstrengthened it in healthy OCs (Figs. 7, S6). These data further confirmthe tight functional cross-talk between TRPV1 and CB1/CB2 receptorsin osteoclast metabolism, which we have previously investigated [12].The number of multinucleated TRAP(+) OCs was normalized vs thevehicle (DMSO 0.01%).
Molecular screening of CNR2 rs35761398 variant
The rs35761398 variant represents a commonmissensemutation ofthe CNR2 gene resulting in the nucleotide transition AA to GG at secondand third bases of codon63.We didnotfindany significant difference inallelic frequencies and genotype distributions of the polymorphism in74 patients with respect to 50 controls (supplementary data, S7).
TRPV1 silencing affects TRAP expression
In order to further confirm TRPV1-mediated OCs activation wesilenced TRPV1 mRNA in absence or presence of pharmacologicalblockade (I-RTX 2.5 μM). TRPV1 silencing significantly reduced TRAPexpression in osteoporotic OCs, whereas the non-targeting siRNAcontrol was ineffective. Treatment with I-RTX 2.5 μM confirmed thereduced TRAP expression only in cells treated with the non-targetingsiRNA control (supplementary data, S8), thus indicating that theantagonist acts selectively via TRPV1.
Fig. 7. TRPV1 agonism and CB1/ CB2 antagonism affect TRAP(+) OCs number: changes in nwere observed also when drugs where co-applied. AM630 [10 μM] alone, i.e., was unable tosignificant enhancement, that was reverted by the pre-incubation with I-RTX. Also the CB1toward CB2 but only in the osteoporotic cultures.
TRPV1 stimulation increases NF-κB expression
The application of RTX (2.5–5–7 μM) increased in concentration-dependent manner NF-κB mRNA levels in osteoporotic OCs (supple-mentary data, S9).
Immunohistochemistry of TRPV1 in human native bone
We revealed the presence of TRPV1 in OCs that were identified ascells with more than 3 nuclei co-expressing TRAP (supplementarydata, S10). TRPV1/TRAP double staining showed more than 50% of co-localization, while we have previously shown that only 35% of the twoproteins co-localize in healthy OCs [12].
Discussion
We have recently shown a role for the EV/EC system in human OCsderived from PBMC of healthy volunteers and its possible implicationin bone mass and biomechanical function [12]. In the current study,we have instead investigated the alterations of EV/EC function, andrelated enzymatic machinery, in OCs obtained from menopausalwomen with or without OP, a skeletal disorder characterized by lowbone mass and micro-architectural bone deterioration.
The first finding of this study is that OCs from women with post-menopausal OP express more CB1 and TRPV1 receptors than humanhealthy volunteers [12] and healthy menopausal women. In consid-eration of the fact that CB1 or TRPV1 activation stimulates osteoclastactivity [16,17,19] whereas CB2 activation inhibits osteoclast activity[22–24], and CB2 polymorphisms are associated with OP [25], it istempting to speculate that increased TRPV1 and CB1 levels are amongthe underlying causes of OP, especially if accompanied by theelevation, as reported here, of the levels of an endogenous mediatorcapable of activating both receptors (i.e. AEA), and by the down-regulation of another mediator that is instead capable of activatingCB2 (as well as CB1) receptors (i.e. 2-AG).
In further agreement with a role of up-regulated TRPV1 in osteoclastover-activation, we found that, in both the trabecular and the compactbone matrix from osteoporotic human bone biopsies, at least 50% of the
umber of multinucleated TRAP(+) OCs from menopausal women with or without OPaffect osteoporotic OCs number, while the co-application with RTX [5 μM] resulted in aantagonist AM251 was able to inhibit the OCs number increase induced by antagonism

1006 F. Rossi et al. / Bone 48 (2011) 997–1007
TRPV1 signal was on mature and active OCs, as identified by doubleTRAP/TRPV1 labelling. However, it is worth noting that TRPV1 was alsoexpressed by several other cell types, whichwere not stained with TRAPand exhibited different functional or maturation states. Interestingly, wefound a different localization of TRPV1 receptor in menopausal andosteoporotic OCs as compared to premenopausal-derived. While in theOCs from healthy pre-menopausal subjects TRPV1 staining was spreadon the cell surface, in themenopausal, and evenmore in the osteoporotic,subjects, although more expressed, TRPV1 staining appeared to be“clusterized”onaspecificportionof the cytoskeleton, as evidencedby thedouble staining with vimentine. In addition, vimentine staininghighlighted that 80% of osteoporotic OCs shows a phantom-likemorphology, representing the activated/macrophagic state, whereas inthe menopausal-derived healthy OCs, although there were many giantscells, only 40% of these exhibited an activated morphology.
In the OCs from both osteopenic and osteoporotic women, the CB1bisoforms were strongly over-expressed, whereas in OCs from healthyvolunteers only the CB1a subtype was present and hardly detectable[12]. Even though the two isoforms seem to exhibit a similarpharmacology, future studies will have to address the possibility thatthe CB1b, strongly expressed in osteoporotic OCs, plays a different rolefrom CB1a [16,17,19]. One possible reason for increased CB1 over-expression might be related to the decreased levels, observed here, ofthe only full endogenous agonist of this receptor, 2-AG, which, in turn,may be the consequence of increased basal levels of the endovanilloidsAEA and OEA, as well as of PEA, and of TRPV1 receptors. The enhancedAEAandOEA levels, possibly due to the strongdown-regulationof FAAHcoupled to unaltered NAPE-PLD expression levels, also observed here inosteoporotic OCs, might have inhibited 2-AG biosynthesis in theosteoporotic condition, since it was shown in other systems thatTRPV1over-stimulation is capable to inhibitDAGL-α activity and, hence,decrease 2-AG levels [26]. Finally, decreased 2-AG levels could lead toCB1 over-expression as a compensatory or counterbalancing mecha-nism. Although this matter is still quite controversial, it is noteworthythat the orphan G protein-coupled receptor GPR55 has also beensuggested tobeactivatedbymultiple different cannabinoid, endocanna-binoid andendocannabinoid-like ligands [27], includingAEA, PEA, 2-AG,and THC. Interestingly, GPR55 activation is able to stimulate cellpolarization and resorption in human osteoclasts [28]. Our resultsshow that OP is able to increase PEA levels (Fig. 5), which in turn couldactivate theGPR55receptor. Theassociationof bothGPR55andPEAwithinflammatory signalling [29] reinforces speculation that increased PEAcould enhance inflammatory response in bone tissue and osteoclastactivation throughGPR55 and possibly counteract the endocannabinoidprotective anti-osteoclastic effect. For these reasons, further studies areneeded to elucidate GPR55 involvement in EV/EC signalling and the roleof PEA in regulating bonemetabolism.
Our present experiments suggest that CB1 and, particularly, CB2receptors tonically inhibit the activity of OCs from menopausal, butnot from osteoporotic women. In fact, osteoclast activity, as measuredby morphological, functional and molecular experiments, wasincreased in the presence of AM251 and, particularly, AM630, twoselective CB1 and CB2 antagonists/inverse agonists, respectively, butthese effects were not observed in OP-derived cells. This observationcan be explained in part by our present finding of strongly reduced2-AG levels in post-menopausal OCs, which, together with thepresence of the elevated expression of CB1 receptors in osteoporoticOCs, might render CB1 antagonists ineffective at stimulating activityin these cells. However, the expression of CB2 receptors was notfound to be altered in OCs from menopausal osteoporotic women.Nevertheless, the present finding that TRPV1 channel expression wasstrongly up-regulated in OCs from these subjects, together with theobservations that: 1) TRPV1 activation enhances CB1 and, particu-larly, CB2 receptor expression in osteoporotic OCs; 2) the levels ofendogenous TRPV1 modulators, AEA, OEA and PEA, are elevated inpost-menopausal OCs, and those of AEA also in osteoporotic OCs, 3)
the levels of the endogenous agonist of CB2 receptors, 2-AG, areinstead generally reduced, and 4) also the expression levels of CRIP1aare reduced in osteoporotic OCs, might explain why the CB2antagonist AM630 was ineffective at elevating activity in osteopo-rotic OCs. Also consistent with our molecular data is the finding that,while the FAAH blocker URB597 increased the number of poly-nucleate OCs, as well as their basal activity, as suggested by TRAPover-expression and increased enzymatic activity, in menopausalOCs, the same treatment resulted ineffective in osteoporotic OCs,where FAAH was strongly down-regulated.
We found here that agonism of TRPV1 receptors causes activation inmenopausal healthy OCs and inhibition in osteoporotic OCs, whereasTRPV1 antagonism reduces osteoclast activity in both groups, whilebeing ineffective inOCs fromhealthyhumanvolunteers [12]. The formerresult can be explained by increased TRPV1 activity and susceptibility todesensitization in osteoporotic OCs. This, in turn, is likely caused byTRPV1 up-regulation coupled to elevation of AEA levels, and is such thatRTX causes immediate desensitization in these cells, thus leading to thesame overall effect of TRPV1 antagonism, as previously observed inother pathological conditions in which TRPV1 and its ligands are up-regulated, such as neuropathic pain [30,31], during which both agonistsand antagonists inhibit pain [32]. By contrast, in non-osteoporotic OCs,in which TRPV1 expression is lower, TRPV1 agonism and antagonismcause increased and decreased osteoclast activity, respectively. Indeed,the fact that the TRPV1 increased expression is accompanied by itsincreased capability of being immediately desensitized by agonists isalso strongly suggested by the fact that the TRPV1 agonist capsaicinwasfound here to be increasingly less efficacious and potent at elevatingintracellular calciumwhenpassing fromOCs fromhealthy volunteers, tohealthymenopausal women and, eventually, to osteoporotic women. Itis important to underline, however, that increased constitutivedesensitization of TRPV1 in osteoporotic OCs does not imply thatthese receptors are completely refractory to stimulation, sincewe foundhere that RTX treatment of these cells did cause biological effects. Forexample, we found that TRPV1 activation causes stimulation of NF-κBexpression, an effect that, however, should stimulate osteoclast activityrather than decreasing it. Nevertheless, such an increased NF-κBexpressionwas only 50%with respect to basal levels, possibly necessaryto guarantee the functioning of key signalling pathways vital forosteoclast generation but not enough for osteoclast activation. Thus,osteoclast activation was reduced (i.e. TRAP positive cell number)whereas the total cell number remained unchanged as compared to thevehicle-treated cultures.
Desensitization of TRPV1 to capsaicin or RTX action is a processwith both fast and slow components: the former depends on calciuminflux itself, whereas the slow component may depend on calcium-dependent and/or protein kinase A-dependent phosphorylation/dephosphorylation cycle [33]. As one of the major factors for theclinical manifestation of OP is the decreased level of estrogens, whichhave a key role at inhibiting the cAMP-dependent PKA pathway, onemight hypothesize that estrogen withdrawal during menopausemight raise cAMP levels thus inducing TRPV1 phosphorylation andclusterization, and resulting in its more rapidly responding toactivation and subsequent desensitization [34].
The above observations indicate that when OP is clinicallyestablished, osteoclast activation might be unaffected by the phar-macological manipulation of CB1, CB2 or FAAH, but still responding toTRPV1 agonists and antagonists. However, the co-localization ofTRPV1 channels with CB1 and CB2 receptors, and the up-regulation ofcannabinoid receptors following 48-hour incubation with a TRPV1agonist, raise the possibility that cross-talk between these receptormight modify this scenario and be exploited to interfere withexcessive osteoclast activity. Indeed, we found here that whencoincubated with RTX, OCs frommenopausal women with OP becameagain sensitive to antagonism of CB2 receptors, by being activated,and also more sensitive to CB1 antagonism, by being inactivated.

1007F. Rossi et al. / Bone 48 (2011) 997–1007
These effects are very probably explained to a large extent by ourpresent finding that, in osteoporotic OCs, prolonged stimulation ofTRPV1 increases the expression of CB1 and, particularly CB2 receptors,whereas antagonism of CB1, but not CB2, receptors can in turnstimulate the expression of TRPV1, thus rendering even moredramatic the effects of the up-regulation of this channel observed inuntreated osteoporotic cells. Nevertheless, the mutual modificationsobserved here might be explained to some extent also by previousfindings showing that CB1 and TRPV1 receptors, when co-expressedin cells, can produce functional cross-talk. For example, the endog-enous CB1 and TRPV1 agonist, AEA, was more efficacious at inducingTRPV1-mediated stimulation of calcium influx in CB1-TRPV1 co-expressing cells than in cells only expressing TRPV1, and such aneffect was inhibited by selective CB1 antagonists [35]. On the otherhand, inverse agonism at CB1 or CB2 receptors may enhanceintracellular calcium concentrations and, hence, contribute to PLCactivation, which is critical for TRPV1 sensitization [36]. Also thepresent finding that TRPV1 antagonism erases the pro-osteoclasticeffect of CB2 antagonism might be explained by opposing effects onintracellular calcium by these two receptors. At any rate, RTX-inducedTRPV1 desensitization via channel clusterization or increased cellmembrane trafficking, and CB2 receptor over-expression mightrepresent adaptive mechanisms against massive calcium entry andexcitotoxicity. This cross-talk might pave the way to the use of TRPV1modulators, alone or together with CB2 agonists or CB1 antagonists,for the treatment of OP.
Conclusions
In conclusion,we have provided here unprecedented evidence thatthe expression of TRPV1 channels and of CB1 vs CB2 cannabinoidreceptors, as well as of some other important proteins involved in EV/EC metabolism, is deeply modified in OCs from osteoporotic women.We found that, even though TRPV1 might be involved in osteoclastactivation and, hence, bone resorption in healthy conditions, it is alsocapable to induce a strong over-expression of the co-expressed CB2cannabinoid receptor, the counterpart receptor system for bonemineralization and remodelling via osteoclast inhibition. Thus, it ispossible that persistently maintained higher levels of AEA (or of anyother endovanilloid), as it happens in several chronic degenerativediseases [37], might generate an unbalance between TRPV1 and CB1/CB2 receptor stimulation, which might be responsible per se forpathological bone remodelling. In this direction, drug cocktails orhybridmolecules, designed to simultaneously stimulate CB2 receptorsand activate/desensitize or antagonize TRPV1 channels [31], mayprove useful in pathological conditions where an unbalancedosteoblast/osteoclast activity is observed. Future studies will have tobe aimed at further investigating how a proper osteoclast/osteoblastbalance in bonemineralization and resorption can be restored in OP byexploiting TRPV1 and CB2 receptor co-expression also in osteoblasts.
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.bone.2011.01.001.
References
[1] Datta HK, Ng WF, Walker JA, Tuck SP, Varanasi SS. The cell biology of bonemetabolism. J Clin Pathol 2008;61:577–87.
[2] Teitelbaum SL. Bone resorption by osteoclasts. Science 2000;289:504–1508.[3] Zethraeus N, Borgström F, Ström O, Kanis JA, Jönsson B. Cost-effectiveness of the
treatment and prevention of osteoporosis—a review of the literature and areference model. Osteopor Int 2007;18:9–23.
[4] Guggenbuhl P. Osteoporosis in males and females: is there really a difference?Joint Bone Spine 2009;76:595–601.
[5] Curtis JR, McClure LA, Delzell E, Howard VJ, Orwoll E, Saag KG, Safford M, HowardG. Population-based fracture risk assessment and osteoporosis treatmentdisparities by race and gender. J Gen Intern Med 2009;24:956–62.
[6] Stevenson JC, Whitehead MI. Postmenopausal osteoporosis. Br Med J 1982;285:585–8.
[7] Christiansen C, Christensen MS, Transbøl I. Bone mass in postmenopausal womenafterwithdrawal of oestrogen/gestagen replacement therapy. Lancet1981;1:459–61.
[8] Wasnich RD, Bagger YZ, Hosking DJ, McClung MR, Wu M, Mantz AM, Yates JJ, RossPD, Alexandersen P, Ravn P, Christiansen C, Santora AC. Changes in bone densityand turnover after alendronate or estrogen withdrawal. Menopause 2004;11:622–30.
[9] Poole KES, Compston JE. Osteoporosis and its management. B M J 2006;333:1251–6.
[10] Pines A. Lifestyle and diet in postmenopausal women. Climacteric 2009;12:62–5.[11] Compston J. Clinical and therapeutic aspects of osteoporosis. Eur J Radiol 2009;71:
388–91.[12] Rossi F, Siniscalco D, Luongo L, De Petrocellis L, Bellini G, Petrosino S, Torella M,
Santoro C, Nobili B, Perrotta S, Di Marzo V, Maione S. The endovanilloid/endocannabinoid system in human osteoclasts: possible involvement in boneformation and resorption. Bone 2009;44:476–84.
[13] Smart D, Gunthorpe MJ, Jerman JC, Nasir S, Gray J, Muir AI, Chambers JK, RandallAD, Davis JB. The endogenous lipid anandamide is a full agonist at the humanvanilloid receptor (hVR1). Br J Pharmacol 2000;129:227–30.
[14] Ahluwalia J, Urban L, Be Test S. Anandamide regulates neuropeptide release fromcapsaicin-sensitive primary sensory neurons by activating both the cannabinoid 1receptor and the vanilloid receptor 1 in vitro. Eur J Neurosci 2003;7:2611–8.
[15] Abed E, Labelle D, Martineau C, Loghin A, Moreau R. Expression of transientreceptor potential (TRP) channels in human and murine osteoblast-like cells. MolMembr Biol 2009;26:146–58.
[16] Idris AI, van't Hof RJ, Greig IR, Ridge SA, Baker D, Ross RA, Ralston SH. Regulation ofbone mass, bone loss and osteoclast activity by cannabinoid receptors. Nat Med2005;11:774–9.
[17] Idris AI, Sophocleous A, Landao-Bassonga E, van't Hof RJ, Ralston SH. Regulation ofbone mass, osteoclast function, and ovariectomy-induced bone loss by the type 2cannabinoid receptor. Endocrinology 2008;149:5619–26.
[18] Idris AI, Sophocleous A, Landao-Bassonga E, Canals M, Milligan G, Baker D, van'tHof RJ, Ralston SH. Cannabinoid receptor type 1 protects against age-relatedosteoporosis by regulating osteoblast and adipocyte differentiation in marrowstromal cells. Cell Metab 2009;10:139–47.
[19] Idris AI, Landao-Bassonga E, Ralston SH. The TRPV1 ion channel antagonistcapsazepine inhibits osteoclast and osteoblast differentiation in vitro andovariectomy induced bone loss in vivo. Bone 2010;46:1089–99.
[20] Ryberg E, Vu HK, Larsson N, Groblewski T, Hjorth S, Elebring T, Sjögren S, GreasleyPJ. Identification and characterisation of a novel splice variant of the human CB1receptor. FEBS Lett 2005;579:259–64.
[21] Engeli S, Böhnke J, Feldpausch M, Gorzelniak K, Janke J, Bátkai S, Pacher P, Harvey-White J, Luft FC, Sharma AM, Jordan J. Activation of the peripheral endocanna-binoid system in human obesity. Diabetes 2005;54:2838–43.
[22] Ofek O, Karsak M, Leclerc N, Fogel M, Frenkel B, Wright K, Tam J, Attar-Namdar M,Kram V, Shohami E, Mechoulam R, Zimmer A, Bab I. Peripheral cannabinoidreceptor, CB2, regulates bone mass. Proc Natl Acad Sci USA 2006;103:696–701.
[23] Bab I, Ofek O, Tam J, Rehnelt J, Zimmer A. Endocannabinoids and the regulation ofbone metabolism. J Neuroendocrinol 2008;1:69–74.
[24] Bab I, Zimmer A, Melamed E. Cannabinoids and the skeleton: from marijuana toreversal of bone loss. Ann Med 2009;41:560–7.
[25] Karsak M, Cohen-Solal M, Freudenberg J, Ostertag A, Morieux C, Kornak U, Essig J,Erxlebe E, Bab I, Kubisch C, de Vernejoul MC, Zimmer A. Cannabinoid receptor type2 gene is associated with human osteoporosis. HumMol Genet 2005;14:3389–96.
[26] Maccarrone M, Rossi S, Bari M, De Chiara V, Fezza F, Musella A, Gasperi V,Prosperetti C, Bernardi G, Finazzi-Agrò A, Cravatt BF, Centone D. Anandamideinhibits metabolism and physiological actions of 2-arachidonoylglycerol in thestriatum. Nat Neurosci 2008;11:152–9.
[27] Brown AJ. Novel cannabinoid receptors. Br J Pharmacol 2007;152:567–75.[28] Whyte LS, Ryberg E, Sims NA, Ridge SA, Mackie K, Greasley PJ, et al. The putative
cannabinoid receptor GPR55 affects osteoclast function in vitro and bone mass invivo. Proc Natl Acad Sci USA 2009;106:16511–6.
[29] Mackie K, Stella N. Cannabinoid receptors and endocannabinoids: evidence fornew players. AAPS J 2006;8:E298–306.
[30] Maione S, De Petrocellis L, de Novellis V, Moriello AS, Petrosino S, Palazzo E, RossiFS, Woodward DF, Di Marzo V. Analgesic actions of N-arachidonoyl-serotonin, afatty acid amide hydrolase inhibitor with antagonistic activity at vanilloid TRPV1receptors. Br J Pharmacol 2007;150:766–81.
[31] Petrosino S, Palazzo E, de Novellis V, Bisogno T, Rossi F, Maione S, Di Marzo V.Changes in spinal and supraspinal endocannabinoid levels in neuropathic rats.Neuropharmacol 2007;52:415–22.
[32] Premkumar LS, Targeting TRPV1 as an Alternative Approach to Narcotic Analgesicsto Treat Chronic Pain Conditions, AAPS J. (2010) Epub ahead of print.
[33] Tominaga M, Tominaga T. Structure and function of TRPV1. Pflugers Arch2005;451:143–50.
[34] Garcìa-Sanz N, Fernández-Carvajal A, Morenilla-Palao C, Planells-Cases R, Fajardo-Sánchez E, Fernández-Ballester G, Ferrer-Montiel A. Identification of a tetramer-ization domain in the C terminus of the vanilloid receptor. J Neuroscience2004;24:5307–14.
[35] Hermann H, De Petrocellis L, Bisogno T, Schiano Moriello A, Lutz B, Di Marzo V.Dual effect of cannabinoid CB1 receptor stimulation on a vanilloid VR1 receptor-mediated response. Cell Mol Life Sci 2003;60:607–16.
[36] De Petrocellis L, Di Marzo V. Lipids as regulators of the activity of transientreceptor potential type V1 (TRPV1) channels. Life Sci 2005;77:1651–66.
[37] Palazzo E, Staraowicz K, Maione S, Di Marzo V. TRPV1 in neuropathic pain andneurological and neuropsyhiatric disorders. In: Faltynek, Gomtsyen, editors.Vanilloid Receptor TRPV1 in drug discovery. John Wiley & Sons; 2010. Chapter 10.