proteomics of extracellular vesicles: exosomes and ectosomes
TRANSCRIPT

PROTEOMICS OF EXTRACELLULAR VESICLES: EXOSOMES ANDECTOSOMES
Dong-Sic Choi, Dae-KyumKim, Yoon-Keun Kim, and Yong SongGho*Department of Life Sciences, Pohang University of Science and Technology,Pohang, Republic of Korea
Received 15 July 2013; revised 9 December 2013; accepted 9 December 2013
Published online 14 January 2014 in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/mas.21420
Almost all bacteria, archaea, and eukaryotic cells shed extracel-lular vesicles either constitutively or in a regulated manner.These nanosized membrane vesicles are spherical, bilayeredproteolipids that harbor specific subsets of proteins, DNAs,RNAs, and lipids. Recent research has facilitated conceptualadvancements in this emerging field that indicate that extracel-lular vesicles act as intercellular communicasomes by transfer-ring signals to their target cell via surface ligands anddelivering receptors and functional molecules. Recent progressin mass spectrometry-based proteomic analyses of mammalianextracellular vesicles derived from diverse cell types and bodyfluids has resulted in the identification of several thousandvesicular proteins that provide us with essential clues to themolecular mechanisms involved in vesicle cargo sorting andbiogenesis. Furthermore, cell-type- or disease-specific vesicularproteins help us to understand the pathophysiological functionsof extracellular vesicles and contribute to the discovery ofdiagnostic and therapeutic target proteins. This review focuseson the high-throughput mass spectrometry-based proteomicanalyses of mammalian extracellular vesicles (i.e., exosomesand ectosomes), EVpedia (a free web-based integrated databaseof high-throughput data for systematic analyses of extracellularvesicles; http://evpedia.info), and the intravesicular protein–protein interaction network analyses of mammalian extracellu-lar vesicles. The goal of this article is to encourage furtherstudies to construct a comprehensive proteome database forextracellular vesicles that will help us to not only decode thebiogenesis and cargo-sorting mechanisms during vesicle forma-tion but also elucidate the pathophysiological roles of thesecomplex extracellular organelles. # 2014 Wiley Periodicals,Inc. Mass Spec Rev 34:474–490, 2015
Keywords: intercellular communicasomes; nanocosmos; intra-vesicular protein–protein interaction networks; proteomics;systems biology
I. INTRODUCTION
Throughout the evolutionary process, bacteria, archaea, andeukaryotic cells have secreted nanosized membrane vesicles intothe extracellular space (Fig. 1) (Lee et al., 2008, 2009; Deatherage&Cookson, 2012; Choi et al., 2013a). These extracellular vesiclesare spherical, bilayered proteolipids that are enriched with variousproteins, nucleic acids, and lipids. Although they have long beenconsidered cellular artifacts or dust, recent progress in this areaindicates that extracellular vesicles are intercellular communica-somes, that is, extracellular organelles that have multifacetedphysiological and pathological functions in intercellular commu-nication as well as inter-species and inter-kingdom communica-tion (Choi et al., 2007, 2011; Lee et al., 2008, 2010; Honget al., 2009). Furthermore, the secretion of extracellular vesiclescould have similar features in different organisms, and extracellu-lar vesicle-mediated intercellular communication is evolutionari-ly conserved (Lee et al., 2009; Deatherage & Cookson, 2012;Choi et al., 2013a). Publications regarding prokaryotic andeukaryotic extracellular vesicles have grown rapidly during thelast 10 years (Fig. 2); this rapid growth indicates that the field ofextracellular vesicles is expanding intensively (Kim et al., 2013).Therefore, the study of extracellular vesicles provides crucialclues for understanding the intercellular communication net-work in living organisms and the evolutionary connectionsamong bacteria, archaea, and eukaryotes.
A. Overview of Mammalian Extracellular Vesicles:Exosomes and Ectosomes
Extracellular vesicles are secreted by various mammalian cellsunder physiological conditions and various disease states,including cancer, in which the release of extracellular vesicles isaberrantly increased (Gyorgy et al., 2011). Two mechanisms ofmammalian extracellular vesicle biogenesis have been sug-gested (Fig. 3A): (1) exosomes that are 50–100 nm in diameterare secreted from the endosomal membrane compartment afterthe fusion of multivesicular bodies (MVBs) with a plasmamembrane or (2) ectosomes (also known as microparticles andmicrovesicles) with a diameter of 100 nm to 1mm that are sheddirectly from the plasma membrane. These two types ofmammalian extracellular vesicles harbor various bioactivematerials including proteins, genetic materials (mRNAs andmiRNAs), and lipids (Fig. 3B). The term exosomes is consistent-ly used to indicate the exocytic vesicles from MVBs, whereasectosomes are known by diverse names, including extracellularmembrane vesicles, membrane particles, exovesicles, nano-particles, microvesicles, microparticles, matrix vesicles, onco-somes, and shedding vesicles (Choi et al., 2013a).
Additional supporting information may be found in the online version
of this article at the publisher’s web-site.
Contract grant sponsor: National Research Foundation of Korea (NRF);
Contract grant numbers: 2013035248, 2012R1A1A2042534; Contract
grant sponsor: Ministry of Health and Welfare; Contract grant number:
A120273; Contract grant sponsor: Korea Basic Science Institute;
Contract grant number: D33400.�Correspondence to: Yong Song Gho, Department of Life Sciences,
Pohang University of Science and Technology, 77 Cheongam-Ro,
Nam-Gu, Pohang, Gyeongbuk 790-784, Republic of Korea.
E-mail: [email protected]
Mass Spectrometry Reviews, 2015, 34, 474–490# 2014 by Wiley Periodicals, Inc.

Despite apparent differences in the mechanism of biogene-sis between exosomes and ectosomes, it is experimentallydifficult to discriminate exosomes from ectosomes after they aresecreted from cells. Furthermore, there is no clearly descriptivephysical property or molecular marker that can unambiguouslydistinguish exosomes from ectosomes (Deneka et al., 2007;Simons & Raposo, 2009; Choi et al., 2012b). Therefore, we referto these mixed vesicle populations as extracellular vesicles(Fig. 3B) (Choi et al., 2012b).
Upon release, both types of extracellular vesicles circulatein the local extracellular space and travel long distances bydiffusion through body fluids, such as blood and lymph.Extracellular vesicles deliver complex signals via stimulating
target cells (Fig. 3A) using the following mechanisms (Ratajc-zak et al., 2006): (1) inducing receptor-mediated intracellularsignal transduction by surface-expressed or bound ligands, (2)transferring surface receptors to target cells, and (3) deliveringfunctional proteins, RNAs, and lipids into the target cells byfusion with plasma membrane or internalization into theendocytic compartment. These vesicle-mediated intercellularcommunications have several advantages. First, extracellularvesicles display the membrane proteins with a high localconcentration. For example, vesicular ICAM1 has a lowermobility than soluble ICAM1, which likely increases the localdensity and overall avidity of ICAM1 (Lee et al., 2010). Second,extracellular vesicles have an endogenous ability to target their
FIGURE 1. Secretion of extracellular vesicles and extracellular vesicle-mediated intercellular communication isevolutionarily conserved from prokaryotes (archaea, Gram-negative bacteria, and Gram-positive bacteria) toeukaryotes includingmammalian cells.
0
200
400
600
800
1967 1970 1973 1976 1979 1982 1985 1988 1991 1994 1997 2000 2003 2006 2009 2012
Num
ber o
f pub
licat
ions
Year
Eukaryote Prokaryote
FIGURE 2. Publication trends on prokaryotic and eukaryotic extracellular vesicle studies. Publications onprokaryotic and eukaryotic extracellular vesicles that were manually curated after searching candidate papers inPubMed (http://www.ncbi.nlm.nih.gov/pubmed) using the following keywords exosome�, exovesicle�, “mem-brane vesicle,” “membrane vesicles,” “extracellular vesicle,” “extracellular vesicles,” microvesicle�, argosome�,ectosome�, tolerosome�, “matrix vesicle,” “matrix vesicles,” oncosome�, dexosome�, “membrane particle,”“membrane particles,” prostasome�, nanovesicle�, “budding vesicle,” “budding vesicles,” “shedding vesicle,”“shedding vesicles,” “blebbing vesicle,” “blebbing vesicles,” “outer membrane bleb,” and “outer membraneblebs” (Kim et al., 2013).
Mass Spectrometry Reviews DOI 10.1002/mas 475
PROTEOMICS OF EXTRACELLULAR VESICLES &

cells of origin. Third, extracellular vesicles can transfer func-tional hydrophobic molecules, such as bioactive lipids andmembrane proteins. For example, vesicular sphingomyelinexhibits angiogenic activity by inducing the migration andproliferation of endothelial cells (Kim et al., 2002). In addition,extracellular vesicles promote the oncogenic transformation oftarget cells by transferring vesicular EGFRvIII (Al-Nedawiet al., 2008). Fourth, luminal components of extracellularvesicles are protected from degradation. For example, RNAs(mRNAs and miRNAs) are present in extracellular vesicles thatare separated from body fluids, which are rich in RNase (Valadiet al., 2007).
B. “Multi-Omics” Research on Extracellular Vesicles
To decode the molecular mechanisms that are involved invesicular cargo sorting and biogenesis as well as the diversephysiological and pathological functions of these complexextracellular organelles, several thousand protein and RNAcomponents of extracellular vesicles that are derived fromvarious cell types and body fluids have been catalogued usingproteomic and transcriptomic methods (Choi et al., 2013a).However, relatively few vesicular lipids have been identified(Subra et al., 2007; Choi et al., 2013a). Systematic analyses ofvesicular components have revealed that extracellular vesicles
A
B
: Phosphatidylserine : Sphingomyelin
: Ceramide : Cholesterol
Ligand-receptorinteraction
Fusion
Endocytosis
Nucleus
Ceramide
Sphingomyelin
ESCRTs
SyndecanSyntenin
[Ca2+]
CalpainGelsolin
Actin cytoskeleton
nSMase2
Exosomes
Ectosomes
MVB
TGN
Nucleus
Lipids
Vesicle trafficking-related proteins: Annexins, ARFs, ALIX, ESCRTs, TSG101, RABs
Tetraspanins:CD9, CD63, CD81
Cytosolic proteins:14-3-3 proteins, enolases, GAPDH, HSPs
Cytoskeleton proteins: Actin, cofilins, ezrin/radixin/moesin, myosin, tubulin
Receptors:EGFRs, FGFRs, PDGFRs
Adhesion proteins:CD44, EPCAM, ICAMs, Integrins
Secreted proteins:Complements, fibronectins, MIF
Transporters and channels: Solute carrier family, ATP-binding cassettes
Genetic materialsmRNAsmiRNAs
FIGURE 3. Two types of mammalian extracellular vesicles and their molecular components. A: Based on theirbiogenesis mechanisms, mammalian cells secrete two types of extracellular vesicles: exosomes and ectosomes.Exosomes, which are 50–100 nm in diameter, are secreted by the fusion of late endosomes or multivesicularbodies with the plasma membrane. Ectosomes (also known as microparticles and microvesicles), which are100 nm to 1mm in diameter, are shed directly from the plasma membrane. Released extracellular vesicles delivercomplex signals to the target cells via ligand-receptor interaction, fusion with the plasma membrane,internalization by endocytosis, or unidentified mechanisms. B: Mammalian extracellular vesicles are enrichedwith cytoskeletal proteins, cytosolic proteins, secreted proteins, plasma membrane proteins (e.g., adhesionproteins, receptors, tetraspanins, and transporters), and vesicle trafficking-related proteins. Their membranes areenriched with cholesterol, ceramide, phosphatidylserine, and sphingolipids. Moreover, mammalian extracellularvesicles contain vesicle-specific genetic materials, such as DNAs, mRNAs, and miRNAs.
476 Mass Spectrometry Reviews DOI 10.1002/mas
& CHOI ET AL.

harbor a specific subset of proteins, genetic materials (mRNAsand miRNAs), and lipids rather than random cellular compo-nents (Fig. 3B). Moreover, these vesicular proteins, mRNAs,miRNAs, and lipids can be grouped as either common vesicularor cell-type-specific vesicular subsets. The common vesicularsubset of proteins, mRNAs, miRNAs, and lipids provide ussome essential clues regarding the molecular mechanisms thatare involved in vesicle cargo sorting and biogenesis, whereas thecell-type-specific vesicular cargo help us to understand thepathophysiological functions of extracellular vesicles and couldprove to be a rich source of biomarkers (Kosaka, Iguchi, &Ochiya, 2010; Raimondo et al., 2011; Choi et al., 2013a,b).
This review focuses primarily on the high-throughput massspectrometry-based proteomic analyses of mammalian extracel-lular vesicles (i.e., exosomes and ectosomes). A recent review ofthe proteomic analysis of Gram-negative bacteria extracellularvesicles (also known as outer membrane vesicles) has beenpublished (Lee et al., 2008). In addition, we introduce EVpedia,which is a free web-based integrated database of high-through-put data for the systematic analyses of extracellular vesicles(http://evpedia.info), and we highlight the protein–proteininteraction network analyses of mammalian extracellularvesicles that represent the interrelationship between vesicularproteins and their functional activities. The goal of this review isto encourage further studies on the high-throughput massspectrometry-based proteomics and on the intravesicular pro-tein–protein interaction network analyses of extracellularvesicles that provide us with essential clues to the molecularmechanisms involved in vesicle cargo sorting and biogenesis.Furthermore, cell-type- or disease-specific vesicular proteinshelp us to understand the pathophysiological functions of thesecomplex extracellular organelles and contribute to the discoveryof diagnostic and therapeutic target proteins.
II. BIOGENESIS OF MAMMALIANEXTRACELLULAR VESICLES
A. Exosomes
Exosomes were first observed approximately 30 years ago byelectron microscopy as small intracellular vesicles that containselect plasma membrane proteins in maturing reticulocytes (Panet al., 1985). These small vesicles were segregated within amembrane-bound organelle forming a MVB and were releasedvia fusion of the MVB’s external membrane with the plasmamembrane of the cell (Fig. 3A). This process does not proceedby default or at random, but rather it is highly regulated byspecific modulators. The endosomal sorting complexes requiredfor transport (ESCRTs) have been shown to play a key role inthe generation of vesicles in MVBs via the recognition ofubiquitinylated membrane proteins (Simons & Raposo, 2009).ESCRT complexes are composed of ESCRT-0, ESCRT-1,ESCRT-2, ESCRT-3, and other accessory proteins (Hurley &Hanson, 2010). ESCRT-0, ESCRT-1, and ESCRT-2 haveubiquitin-interacting modules that are necessary for the sequen-tial sorting of cargo that is destined for the MVBs. ESCRT-0plays a role in the clustering of ubiquitinylated cargo. ESCRT-1and ESCRT-2 are responsible for membrane budding, andESCRT-3 cleaves the budded membrane by fission. Recently,syndecan heparan sulfate proteoglycans and their cytoplasmicadaptor syntenin have been shown to interact with ALIX, an
ESCRT-3-binding protein, thus controlling the formation ofexosomes (Baietti et al., 2012). However, an alternative pathwayfor exosome formation has been suggested in which themetabolic pathway for ceramide synthesis from sphingomyelin,which is catalyzed by sphingomyelinase, induces exosomalprotein sorting into MVBs (Trajkovic et al., 2008). However, theexact molecular mechanisms that are involved in exosomebiogenesis have not yet been elucidated.
B. Ectosomes
The first description of ectosomes, also frequently referred to byother terms, such as microparticles or microvesicles, wasreported in 1967 (Wolf, 1967). In human plasma, the fragmentsthat were derived from platelets were identified, and they wereshown to have shapes that resembled small membrane vesicles(Wolf, 1967). It has been suggested that ectosome shedding ispreceded by the budding of protrusions on the plasma membranefollowed by detachment by fission of the budding site (Fig. 3A)(Ratajczak et al., 2006; Cocucci, Racchetti, & Meldolesi, 2009).Importantly, the shedding rate of the ectosomes increasesdramatically upon stimulation rather than upon release fromresting cells. In particular, an increased level of intracellularCa2þ induces a strong ectosome-shedding response in variouscell types (Pilzer et al., 2005; Ratajczak et al., 2006; Moskovich& Fishelson, 2007). The intracellular Ca2þ concentration altersthe asymmetric phospholipid distribution of the plasma mem-brane by activating scramblase and floppase, resulting in theexposure of phosphatidylserine and phosphatidylethanolaminein the outer leaflet of the plasma membrane (Pap et al., 2009).Moreover, the level of Ca2þ contributes to the degradation of thecytoskeleton via the activation of Ca2þ-dependent proteases,such as calpain and gelsolin, followed by ectosome release (Papet al., 2009). Recently, it was reported that RhoA signaling leadsto the downstream activation of a Rho-associated coiled-coil-containing protein kinase that stimulates the generation ofectosomes to regulate the actin cytoskeleton via cofilin (Li et al.,2012). Moreover, the GTP-binding protein ARF6 modulates therelease of protease-loaded ectosomes (Muralidharan-Chariet al., 2009a). By acting through phospholipase D and ERK,ARF6 activates myosin light chain kinase, and the subsequentphosphorylation of myosin light chain regulates the releaseof ectosomes from invasive cells (Muralidharan-Chari et al.,2009a,b). However, the mechanisms of ectosome biogenesis andthe sorting of proteins into ectosomes are more obscure than themechanisms of exosome biogenesis and the sorting of proteinsinto exosomes.
III. MASS SPECTROMETRY-BASED PROTEOMICANALYSES OF MAMMALIAN EXTRACELLULARVESICLES
Early mass spectrometry-based proteomic analyses of mamma-lian extracellular vesicles mainly focused on the identificationof highly abundant vesicular proteins. In these studies, proteinsfrom purified extracellular vesicles were separated using gelelectrophoresis, and then, abundant proteins were analyzed bymass spectrometry (Thery et al., 1999, 2001; van Niel et al.,2001; Skokos et al., 2003). Using these approaches, some ofthe important vesicular marker proteins, such as tetraspanins,
Mass Spectrometry Reviews DOI 10.1002/mas 477
PROTEOMICS OF EXTRACELLULAR VESICLES &

14-3-3 proteins, actin, annexins, and heat shock proteins, wereidentified, but only a small number of vesicular proteins(approximately 10–30 proteins per study) were discovered.
During the last 20 years, as protein separation and massspectrometry technology have improved, several thousandproteins of extracellular vesicles from various cell typesand body fluids, including serum, urine, saliva, ascites, andbreast milk, have been catalogued by high-throughput massspectrometry-based proteomic studies (Choi et al., 2013a; Kimet al., 2013). The lists of all vesicular proteins that have beenidentified by these studies have been deposited in the free web-based EVpedia (http://evpedia.info) database (Fig. 4) (Choiet al., 2013a; Kim et al., 2013). An overview of the currentmethodological approaches in the high-throughput mass spec-trometry-based proteomic analyses of extracellular vesicles isprovided in Figure 5. Although all of the steps described in thisoverall scheme have influenced the quality of the results, thefollowing two methodologies are critical prerequisites for high-throughput mass spectrometry-based proteomic analyses: (1)efficient isolation of mammalian extracellular vesicles withoutany contamination by non-vesicular components and (2) frac-tionation of peptides or proteins prior to mass spectrometricanalysis to reduce the vesicular proteome complexity.
A variety of vesicle isolation methods, including ultracen-trifugation, immunoaffinity beads, flow field-flow fractionation,and gel filtration, are used for the high-throughput massspectrometry-based proteomic analyses of mammalian extracel-lular vesicles from cell culture medium or body fluids, includingserum, urine, saliva, ascites, and breast milk (Fig. 5). Thedetailed vesicle isolation methods for high-throughput massspectrometry-based proteomic analyses are listed in the EVpe-dia (http://evpedia.info) database (Choi et al., 2013a; Kim
et al., 2013). The most commonly used vesicle isolation methodis ultracentrifugation (Thery et al., 2006; Choi et al., 2013a).After the pre-clearing of cells, cell debris, and large apoptoticbodies by differential centrifugation with or without 0.1–0.2mmfiltration, mammalian extracellular vesicles are pelleted byultracentrifugation or sucrose cushion ultracentrifugation (e.g.,100,000–200,000g) (Hong et al., 2009). The vesicle isolationyield is adequate, but various different types of particles (e.g.,lipoproteins and viruses), small apoptotic bodies, aggregates,protein oligomers, or proteins that are non-specifically bound toextracellular vesicles should be co-sedimented by this isolationmethod.
Recent progress in high-throughput mass spectrometry-based proteomic analyses of extracellular vesicles indicates thatbuoyant density gradient ultracentrifugation is one of the bestvesicle isolation methods to remove the above-mentionedcontaminants that are caused by ultracentrifugation- or sucrosecushion ultracentrifugation-based vesicle isolation methods.Recently, several alternative vesicle isolation methods, includ-ing immunoaffinity beads, flow field-flow fractionation, andgel filtration, were introduced for efficient vesicle isolation(Kang et al., 2008; Dean et al., 2009; Looze et al., 2009;Mathivanan et al., 2010; Choi et al., 2013a). However, thetechnical standardization for challenging methods that are usedto isolate extracellular vesicles is urgently needed because thelists of vesicular proteins identified by mass spectrometry-basedproteomic analyses are largely dependent on the methods thatare used for the isolation of extracellular vesicles. Recentreviews of this subject are available (Choi et al., 2013a; Witweret al., 2013).
Prior to high-throughput mass spectrometry-based proteo-mic analysis, dynamic range reduction of vesicular proteins or
Database: Vesicular protein, mRNA, miRNA, and lipid
Search: Protein
Browse: Protein, mRNA, miRNA, and lipid
Gene Ontology Analysis Protein and mRNA
Network AnalysisProtein and mRNA
Set Analysis
Analysis
Search and Browse
Prokaryotes Eukaryotes
All Mammals Non-mammalian
eukaryotes
High-throughput proteomesa
Datasets 23 97 9 129
Proteins 3,526 47,759 1,491 52,786
High-throughput transcriptomes
mRNA
Datasets 0 19 0 19
mRNAs 0 164,519 0 164,519
miRNA
Datasets 0 22 0 22
miRNAs 0 13,287 0 13,287
Lipidomes
Datasets 1 17 2 20
Lipids 16 300 29 345
aHigh-throughput proteomes in which at least 50 vesicular proteins were identified (Choi et al., 2013a; Kim et al., 2013).
FIGURE 4. EVpedia: an integrated database of high-throughput data for the systematic analyses of prokaryoticand eukaryotic extracellular vesicles. A total of 230,937 vesicular proteins, mRNAs, miRNAs, and lipids ofprokaryotic, non-mammalian eukaryotic, and mammalian extracellular vesicles from 190 high-throughputdatasets are deposited in the free web-based database of EVpedia (http://evpedia.info) (Kim et al., 2013). For thesystematic analyses of prokaryotic and eukaryotic extracellular vesicles, EVpedia provides an array of analyticaltools: (1) search for and browse vesicular components, (2) Gene Ontology and network analyses of vesicularcomponents, and (3) set analysis: a comparison of vesicular datasets by ortholog identification.
478 Mass Spectrometry Reviews DOI 10.1002/mas
& CHOI ET AL.

peptides by fractionation is critical to identify the highlyconfident vesicular proteins and to catalogue a large number ofvesicular proteins (Choi et al., 2013a). As shown in Figure 5,three main strategies have been used to reduce the complexity ofthe vesicular proteome: (1) sodium dodecyl sulfate–polyacryl-amide gel electrophoresis-based vesicular protein fractionation(Utleg et al., 2003; Pisitkun, Shen, & Knepper, 2004), (2) two-dimensional liquid chromatography-based vesicular peptidefractionation, known as multidimensional protein identificationtechnology (Gonzalez-Begne et al., 2009), and (3) isoelectricfocusing-based fractionation of vesicular proteins or peptides(Choi et al., 2012a).
In the following subsections, we summarize previous massspectrometry-based proteomic studies of mammalian extracel-lular vesicles by introducing an integrated database of high-throughput data for the systematic analyses of extracellularvesicles (EVpedia) and discuss in detail the quantitation-basedcomparative proteomic analyses of mammalian extracellular
vesicles and the roles of proteomics in understanding mammali-an extracellular vesicles.
A. EVpedia: An Integrated Database of High-Throughput Data for the Systematic Analyses ofExtracellular Vesicles
Currently, three public online databases of vesicular proteins,mRNAs, miRNAs, and lipids (EVpedia, ExoCarta, andVesiclepedia) are available. EVpedia provides lists of thecomprehensive vesicular proteins, mRNAs, miRNAs, and lipidsof prokaryotic, non-mammalian eukaryotic, and mammalianextracellular vesicles (Choi et al., 2013a; Kim et al., 2013),while ExoCarta (Mathivanan & Simpson, 2009) and Vesiclepe-dia (Kalra et al., 2012) collects only the vesicular componentsof non-mammalian eukaryotic and mammalian extracellularvesicles. In the current EVpedia database (http://evpedia.info), a
FIGURE 5. Overview of methodological approaches in high-throughput mass spectrometry-based proteomicanalyses of extracellular vesicles.
Mass Spectrometry Reviews DOI 10.1002/mas 479
PROTEOMICS OF EXTRACELLULAR VESICLES &

total of 230,937 vesicular proteins, mRNAs, miRNAs, and lipidsfrom 190 high-throughput datasets were collected: 47,759vesicular proteins deposited from the 97 high-throughput massspectrometry-based proteomic datasets of mammalian extracel-lular vesicles (Fig. 4). Moreover, EVpedia provides an array ofanalytical tools: (1) search for and browse vesicular compo-nents, (2) Gene Ontology enrichment analysis of vesicularcomponents, (3) network analysis of vesicular components, and(4) set analysis: a comparison of vesicular datasets by orthologidentification. In addition, the detailed methods for the isolationof extracellular vesicles and publications on extracellular vesiclestudies are listed in the database. An overall comparison ofEVpedia with Exocarta and Vesiclepedia has been recentlypublished (Choi et al., 2013a; Kim et al., 2013). By providingthese vesicle-specific components that are derived from varioustypes of cells and body fluids as well as an array of analyticaltools for systematic analyses, these integrated online databasesfacilitate communication and resource sharing between re-searchers in this emerging field of biology. Researchers canupload their high throughput datasets to the EVpedia databasethrough the “Upload” menu (Kim et al., 2013).
B. Quantitation-Based Comparative ProteomicAnalyses of Mammalian Extracellular Vesicles
Although most mammalian cells secrete extracellular vesicles,their release rates and their molecular components, includingproteins, mRNAs, and miRNAs, are dependent on cell type.Furthermore, similar to the cellular proteome, vesicular proteincomponents and their abundance are influenced by cellularstatus, including stimulation by growth factor (Di Vizio et al.,2009), exposure to the aging process (Lehmann et al., 2008), andtransfection with infectious molecules (Coleman et al., 2012).Therefore, quantitation-based comparative proteomic analysesof mammalian extracellular vesicles that are derived fromdiverse cell types and cellular conditions are expected to provideinsights into the emerging biology of extracellular vesicles, toelucidate the biogenesis and pathophysiological functions ofextracellular vesicles and to contribute to the discovery ofdiagnostic and therapeutic target proteins.
Two technical approaches are applied for protein quantita-tion in proteomics: label-based and label-free quantitation(Fig. 5). Label-based quantitation methods, such as isobaric tagsfor relative and absolute quantification (iTRAQ) and stable-isotope labeling of amino acids (SILAC) are commonly used invarious comparative proteomic analyses, but these laboriousapproaches are expensive, multistep labeling processes and arenot always feasible due to the insufficient labeling of tagsaccording to the discrimination of samples (Bachi & Bonaldi,2008). Alternatively, label-free quantitation methods, based onthe extracted chromatograms of spectral intensity or spectralcounts, provide inexpensive, convenient, and reliable methodsfor quantitative proteomics studies (Bachi & Bonaldi, 2008).The spectral counting method is based on the assumption thatmore abundant proteins have more abundant peptides. Theabsolute protein expression (APEX) tool is based on spectralcounting of each protein, and it is corrected by the priorexpectation of observing each peptide (Lu et al., 2007). Thisapproach provides absolute protein concentrations across ap-proximately three to four orders of magnitude. Moreover, theprotein abundance that is quantified by APEX is consistent with
Western blotting and two-dimensional gel electrophoresis (Luet al., 2007).
The currently available high-throughput quantitation-basedcomparative proteomic analyses of mammalian extracellularvesicles are summarized in Table 1. For example, we recentlyconducted APEX-based label-free quantitative proteomic analy-ses of extracellular vesicles derived from human primarycolorectal cancer cells (SW480) and their metastatic derivatives(SW620) (Choi et al., 2012a). From the comparison of theestimated abundance of vesicular proteins, 368 and 359 enrichedproteins were identified in SW480- and SW620-derived extra-cellular vesicles, respectively. These differentially regulatedproteins are closely related to the cellular status. For example,SW620-enriched vesicular proteins are associated with cancerprogression and multidrug resistance as potential diagnosticmarkers of metastatic cancer, whereas SW480-enriched vesicu-lar proteins play a role in cell adhesion.
C. Role of Proteomics in Understanding MammalianExtracellular Vesicles
As more vesicular proteins are identified, it has become apparentthat extracellular vesicles contain a specific subpopulation ofproteins rather than randomly selected molecules from theircells of origin (de Gassart et al., 2004; Choi et al., 2013a).Vesicular proteins are mainly derived from the plasma mem-brane, cytosol, and internal vesicles and not from otherintracellular organelles, such as the endoplasmic reticulum,Golgi apparatus, mitochondria, and nucleus. Moreover, extra-cellular vesicles harbor a common set of vesicular proteins andcell-type-specific components (Choi et al., 2013a). For example,annexins, cytoskeletal proteins (actins, cofilin-1, ezrin/radixin/moesin, profilin-1, and tubulins), heat-shock proteins, integrins,metabolic enzymes (enolases, glyceraldehyde 3-phosphate de-hydrogenase, peroxiredoxins, and pyruvate kinase), ribosomalproteins, tetraspanins (CD9, CD63, and CD81), and vesicletrafficking-related proteins (TSG101, ALIX, and RAB proteins,syntenin-1) are frequently identified vesicular proteins(Fig. 3B). Table 2 presents the top 100 vesicular proteins,ordered from the highest to lowest identification count, that arefound most frequently in mammalian extracellular vesicles. Theidentification count of each vesicular protein represents thenumber of identifications of that protein in high-throughputproteomic datasets (Choi et al., 2013a). EVpedia (http://evpedia.info) provides a long list of vesicular proteins and theiroccurrences within prokaryotic and mammalian extracellularvesicles (Choi et al., 2013a; Kim et al., 2013). In addition,diverse functional proteins, including cell-surface antigens,immune-modulating proteins, proteases, and angiogenic mole-cules, have been identified (Choi et al., 2007). Therefore, thesevesicular proteomes provide diverse information on the natureof extracellular vesicles and help us to decode the molecularmechanisms that are involved in vesicular cargo sorting andbiogenesis as well as the diverse physiological and pathologicalfunctions of these complex extracellular organelles (Choi et al.,2007, 2013a). Finally, high-throughput mass spectrometry-based proteomic studies on extracellular vesicles facilitatebiomarker discovery based on the protein signature of theoriginating cells. Recent reviews of this subject are available inthe literature (Raimondo et al., 2011; Choi et al., 2013a,b).
480 Mass Spectrometry Reviews DOI 10.1002/mas
& CHOI ET AL.

TABLE
1.High-throughputquantitation-based
comparativeproteomicanalysesofmam
malianextracellularvesicles
ESI,electrosprayionization;iTRAQ,isobarictagsforrelativeandabsolutequantification;LC,liquidchromatography;MALDI,matrix-assistedlaserdesorption;MS/M
S,tandem
massspectrometry.
Sa
mpl
e ty
pe
Ultr
acen
trifu
gatio
n-ba
sed
vesi
cle
isol
atio
n
Pr
otei
n or
pep
tide
sepa
ratio
n st
rate
gies
M
ass
spec
trom
eter
Pr
otei
n Q
uant
itatio
n Ye
ar
Ref
eren
ce
Filtr
atio
n
(0.1
-0.2
µm
) U
ltrac
entri
fuga
tion
Sucr
ose
cush
ion
ultra
cent
rifug
atio
n D
ensi
ty g
radi
ent
ultra
cent
rifug
atio
n
Prot
ein
sepa
ratio
n an
d in
-gel
di
gest
ion
In-s
olut
ion
dige
stio
n an
d pe
ptid
e se
para
tion
Cel
l cul
ture
med
ium
H
omo
sapi
ens
C
olor
ecta
l ca
ncer
cel
l (D
Ks-
8, D
LD-
1, a
nd D
KO
-1)
DK
s-8
(Wild
-type
KR
AS)
, D
LD-1
(Par
enta
l; he
tero
zygo
us K
RA
S), a
nd
DK
O-1
(Mut
ant K
RA
S)
+ +
− −
−
+ ES
I-M
S/M
S Sp
ectra
l co
unts
20
13
Bec
kler
et
al.,
2013
C
olor
ecta
l ca
ncer
cel
l (S
W48
0 an
d SW
620)
SW48
0 (P
rimar
y ca
ncer
) an
d SW
620
(Met
asta
tic
canc
er)
− −
+ +
+
− ES
I-M
S/M
S Sp
ectra
l co
unts
20
12
Cho
i et a
l.,
2012
b
En
doth
elia
l ce
ll (H
UV
EC)
Uns
timul
ated
and
st
imul
ated
by
TNF-
alph
a an
d PA
I-1
− +
− −
+
− ES
I-M
S/M
S To
tal i
on
curr
ent
2008
Pe
ters
on e
t al
., 20
08
N
eona
tal
myo
blas
t cel
l Ex
osom
es a
nd
mic
ropa
rticl
es in
pat
ient
s of
dev
elop
men
tal h
eart
defe
ct
− +
− −
−
− ES
I-M
S/M
S Sp
ectra
l co
unts
20
12
Le B
ihan
et
al.,
2012
R
ed b
lood
cel
l St
orag
e da
y 3,
21,
and
42
− +
− −
+
− ES
I-M
S/M
S Sp
ectra
l co
unts
20
08
Bos
man
et
al.,
2008
R
attu
s nor
vegi
cus
R
etic
uloc
yte
Cul
ture
d fo
r 36
h, 8
4 h,
an
d 15
6 h
− +
− −
+
− ES
I-M
S/M
S Sp
ectra
l co
unts
20
11
Car
ayon
et
al.,
2011
Bo
dy fl
uid
H
omo
sapi
ens
B
reas
t milk
C
olos
trum
and
mat
ure
milk
+
+ −
+
− −
ESI-
MS/
MS
Spec
tral
coun
ts
2007
A
dmyr
e et
al
., 20
07
Pl
asm
a D
eep
veno
us th
rom
bosi
s −
+ −
−
− +
MA
LDI-
MS/
MS
iTR
AQ
20
10
Ram
acci
otti
et a
l., 2
010
U
rine
Bla
dder
can
cer
− +
− −
+
− ES
I-M
S/M
S Sp
ectra
l co
unts
20
08
Smal
ley
et
al.,
2008
Urin
e Ea
rly Ig
A n
ephr
opat
hy a
nd
base
men
t mem
bran
e ne
phro
path
y
− −
− +
+
− ES
I-M
S/M
S Sp
ectra
l in
tens
ity
2011
M
oon
et a
l.,
2011
U
rine
Bla
dder
can
cer a
nd h
erni
a −
+ −
−
− +
ESI-
MS/
MS
Spec
tral
inte
nsity
20
12
Che
n et
al.,
20
12
Mass Spectrometry Reviews DOI 10.1002/mas 481
PROTEOMICS OF EXTRACELLULAR VESICLES &

TABLE 2. Top 100 vesicular proteins that are most often found in mammalian extracellular vesiclesa
noitacifitnedI slobmys eneG eman nietorP count
19 2GTCA ,2LBTCA ,1ATCA ,1CTCA ,2ATCA ,BTCA snietorp detaler dna nitcA 1 ,JETOP ,EETOP ,PKETOP ,1GTCA ,IETOP ,FETOP niryknA 2
POTEB 91
APSH ,A1APSH 07 nietorp kcohs taeH 3 5, HSPA8, HSPA6, HSPA2, HSPA1L, HSPA7
90
28 NSM ,XDR ,RZE niseoM/nixidaR/nirzE 45 Glyceraldehyde-3-phosphate dehydrogenase/erythrose-
4-phosphate dehydrogenase 87 SHDPAG ,HDPAG
67 2P2AXNA ,2AXNA 2A nixennA 6 67 A6LAHDL ,BHDL ,AHDL esanegordyhed etatcal-L 7 67 3XDRP ,2XDRP ,1XDRP nixoderixoreP 8 57 3ONE ,2ONE ,1ONE esalonE 9 27 EAHWY nolispe nietorp 3-3-41 01
,1BBUT ,BBUT ateb nilubuT 11 TUBB2A, TUBB2B, TUBB3, TUBB4A, TUBB4B, TUBB6, TUBB8
72
17 1LFC 1-nilifoC 211AA09PSH 09 nietorp kcohs taeH 31 , HSP90AB1, HSP90B1, HSP90AB3P,
HSP90AB4P, TRAP1 70
14 Translation elongation factor EF-1alpha (G 07 L1SBH ,5P1A1FEE ,2A1FEE ,1A1FEE )esaPT 96 RLKP ,2MKP ,MKP esanik etavuryP 51
ABUT ,A4ABUT ahpla nilubuT 61 1C, TUBA8, TUBA3C, TUBA1A, TUBA1B, TUBA3E, TUBAL3
68
76 2KGP ,1KGP esanik etarecylgohpsohp-3 7118 Guanine nucleotide-binding protein G( 76 3IANG ,2IANG ,1IANG ahpla tinubus )i
56 4NTCA ,3NTCA ,2NTCA ,1NTCA ninitca-ahplA 9120 Ras-related protein 56 C5BAR ,B5BAR 5-baR
46 ZAHWY atled/atez nietorp 3-3-41 12 46 5AXNA 5A nixennA 22 46 CTLC ,1LCTLC niahc yvaeh nirhtalC 32 36 BAHWY ahpla/ateb nietorp 3-3-41 42
1PTA ,1A1PTA esaPTA tropsnart noitaC 52 A2, ATP1A3, ATP1A4, ATP12A, ATP4A 63 36 4BNG ,2BNG ,1BNG nietorp gnidnib-editoelcun eninauG 62 36 1NFP 1-niliforP 72 16 AIPP esaremosi snart-sic lylorp-lyditpeP 82 06 A1BAR A1-baR nietorp detaler-saR 92
30 Programmed cell death 6-inte 95 PI6DCDP nietorp gnitcar 85 QAHWY ateht nietorp 3-3-41 13 75 GAHWY ammag nietorp 3-3-41 23 75 BLA nimubla mureS 33 65 4DHE ,3DHE ,2DHE ,1DHE nietorp gniniatnoc-niamod HE 43 65 B1PAR b1-paR nietorp detaler-saR 53 55 1FRA 1 rotcaf noitalysobir-PDA 63 55 7BGTI ,6BGTI ,5BGTI ,3BGTI ,2BGTI ,1BGTI ateb nirgetnI 73 55 6XDRP 6-nixoderixoreP 83 45 IPG esaremosi etahpsohp-6-esoculG 93 45 PBCDS 1-ninetnyS 04 45 1IPT esaremosi etahpsohpesoirT 14
42 Ubiquitin-like modifier-activa 45 7ABU ,6ABU ,1ABU emyzne gnit 35 6AXNA 6A nixennA 34
44 Cell division control prot 35 24CDC golomoh 24 nie 25 AODLA ,CODLA esalodla etahpsohpsib-6,1-esotcurF 54 25 2DUTFE ,2FEE )sesaPTG( srotcaf noitagnole noitalsnarT 64 15 9DC negitna 9DC 74 15 B11BAR ,A11BAR 11-baR nietorp detaler-saR 84 05 11AXNA ,7AXNA 11/7A nixennA 94
482 Mass Spectrometry Reviews DOI 10.1002/mas
& CHOI ET AL.

TABLE 2. (Continued)
05 1CILC 1 nietorp lennahc ralullecartni edirolhC 05 05 NSAF esahtnys dica yttaF 15
,9HYM ,8HYM ,7HYM ,4HYM ,3HYM ,1HYM niahc yvaeh nisoyM 25MYH10, MYH11, MYH14, MYH13, MYH15
50
05 A7BAR a7-baR nietorp detaler-saR 3554 4F2 cell-surface antigen 94 2A3CLS niahc yvaeh55 Eukaryotic initiation fact 94 2A4FIE ,1A4FIE A4 ro56 Ras-related C3 botulinum t 94 2CAR ,1CAR etartsbus nixo
94 2TCC ateb tinubus 1 nietorp xelpmoc-T 75 84 YCHA esanietsycomohlysonedA 85 84 B5PTA lairdnohcotim ,ateb tinubus esahtnys PTA 95 84 5C ,B4C ,A4C ,3C tnemelpmoC 06
,A1OYM 1 niahc yvaeh nisoyM 16 MYO1B, MYO1C, MYO1D, MYO1E, MYO1F, MYO1G
48
84 2IDG ,1IDG rotibihni noitaicossid PDG baR 26 74 01TRK 01 lateleksotyc I epyt ,nitareK 36
64 Large extracellular alpha-helical 74 PZP ,901DC ,1LM2A ,M2A nietorp 65 Nucleoside diphosphate ki 74 3EMN ,2EMN ,1EMN esan66 Phosphatidylethanolamine-b 74 1PBEP 1 nietorp gnidni
74 01BAR ,B1BAR 01/B1 baR nietorp detaler-saR 76 64 1PTSG P esarefsnart-S enoihtatulG 86 64 1HDM esanegordyhed etalaM 96 64 ALAR A-laR nietorp detaler-saR 07 64 2NLGAT 2-nilegsnarT 17 64 PCV esaPTA muluciter cimsalpodne lanoitisnarT 27 54 YLCA esahtnys etartic-PTA 37
74 Ca2+-binding actin-bundling protein fimbrin/plastin (EF-Hand superfamily)
DST, FLNA, FLNB, FLNC, LCP1, PLEC, PLS1, PLS3 45
54 1FCAM ,PSD nietorp gnidnib noi muiclaC 57 54 1BPSH 1-ateb nietorp kcohs taeH 67 54 B2PAR ,41BAR 41/B2-baR nietorp detaler-saR 77
,RGF ,KLB esanik nietor enisoryT 87 FYN, HCK, LCK, LYN, SRC, YES1 45 79 6-phosphogluconate dehydroge 44 DGP gnitalyxobraced ,esan
44 18DC negitna 18DC 08 44 ULC niretsulC 18 44 1LIV ,NICS ,NSG nilosleG 28 44 2MAGP ,1MAGP esatum etarecylgohpsohP 38 44 1PCT ahpla tinubus 1 nietorp xelpmoc-T 48
85 Guanine nucleotide-binding pr 34 QANG ,11ANG )q(G nieto86 Guanine nucleotide-binding pr 34 31ANG 31-ahpla tinubus nieto
34 1TRK 1 lateleksotyc II epyt ,nitareK 78 34 3AIDP 3A esaremosi-ediflusid nietorP 88 34 A2BAR A2-baR nietorp detaler-saR 98 34 2NLT ,1NLT nilaT 09 24 MOTS nietorp enarbmem largetni 7 dnab etycorhtyrE 19 24 SANG ,SANG ,LANG sahpla G tinubus nietorp G 29
,A-ALH negitna ytilibitapmocotsiH 39 HLA-B, HLA-C, HLA-E, HLA-H 42 24 A8BAR A8-baR nietorp detaler-saR 49 24 A1V6PTA A tinubus citylatac esaPTA notorp epyt-V 59 14 8AMSP ,7AMSP stinubus ateb dna ahpla,emosaetorp S02 69
97 Ca2+-binding actin-bundling protein fimbrin/plastin (EF-Hand superfamily)
SPTB, SPTBN1, SPTBN2, SPTBN4, SPTBN5 41
98 Ca2+-binding protein (EF- 14 1NATPS )ylimafrepus dnaH 14 PB3SLAGL nietorp gnidnib-3-nitcelaG 99 14 VAGTI ,5AGTI ,B2AGTI 5/b2-ahpla nirgetnI 001
EVpedia (http://evpedia.info) provides a large list of vesicular proteins and their occurrences of prokaryotic and mammalian extracellular
vesicles (Choi et al., 2013a; Kim et al., 2013).aThe proteins are ordered from the highest to lowest identification count, which represents the number of identifications of that protein in high-throughput proteomic datasets (Choi et al., 2013a).
Mass Spectrometry Reviews DOI 10.1002/mas 483
PROTEOMICS OF EXTRACELLULAR VESICLES &
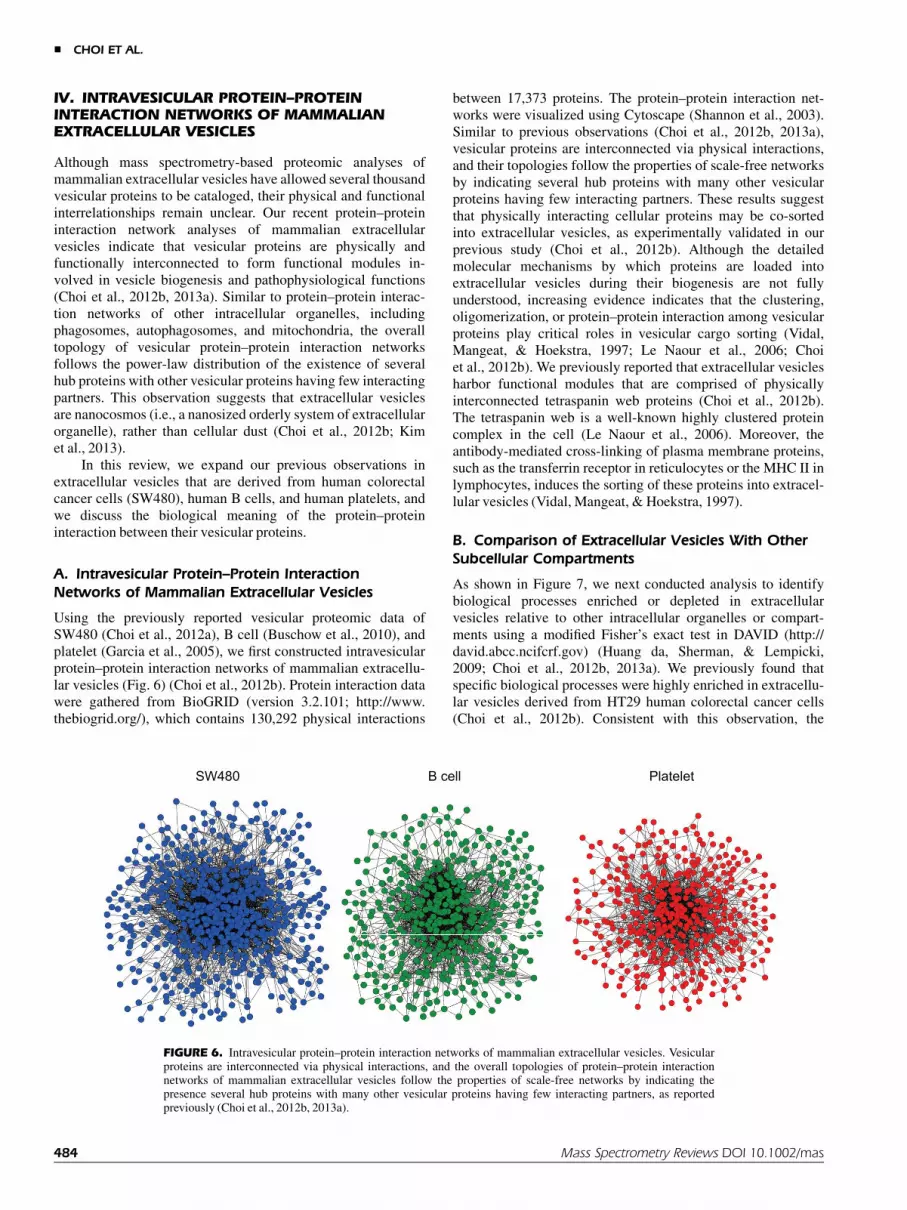
IV. INTRAVESICULAR PROTEIN–PROTEININTERACTION NETWORKS OF MAMMALIANEXTRACELLULAR VESICLES
Although mass spectrometry-based proteomic analyses ofmammalian extracellular vesicles have allowed several thousandvesicular proteins to be cataloged, their physical and functionalinterrelationships remain unclear. Our recent protein–proteininteraction network analyses of mammalian extracellularvesicles indicate that vesicular proteins are physically andfunctionally interconnected to form functional modules in-volved in vesicle biogenesis and pathophysiological functions(Choi et al., 2012b, 2013a). Similar to protein–protein interac-tion networks of other intracellular organelles, includingphagosomes, autophagosomes, and mitochondria, the overalltopology of vesicular protein–protein interaction networksfollows the power-law distribution of the existence of severalhub proteins with other vesicular proteins having few interactingpartners. This observation suggests that extracellular vesiclesare nanocosmos (i.e., a nanosized orderly system of extracellularorganelle), rather than cellular dust (Choi et al., 2012b; Kimet al., 2013).
In this review, we expand our previous observations inextracellular vesicles that are derived from human colorectalcancer cells (SW480), human B cells, and human platelets, andwe discuss the biological meaning of the protein–proteininteraction between their vesicular proteins.
A. Intravesicular Protein–Protein InteractionNetworks of Mammalian Extracellular Vesicles
Using the previously reported vesicular proteomic data ofSW480 (Choi et al., 2012a), B cell (Buschow et al., 2010), andplatelet (Garcia et al., 2005), we first constructed intravesicularprotein–protein interaction networks of mammalian extracellu-lar vesicles (Fig. 6) (Choi et al., 2012b). Protein interaction datawere gathered from BioGRID (version 3.2.101; http://www.thebiogrid.org/), which contains 130,292 physical interactions
between 17,373 proteins. The protein–protein interaction net-works were visualized using Cytoscape (Shannon et al., 2003).Similar to previous observations (Choi et al., 2012b, 2013a),vesicular proteins are interconnected via physical interactions,and their topologies follow the properties of scale-free networksby indicating several hub proteins with many other vesicularproteins having few interacting partners. These results suggestthat physically interacting cellular proteins may be co-sortedinto extracellular vesicles, as experimentally validated in ourprevious study (Choi et al., 2012b). Although the detailedmolecular mechanisms by which proteins are loaded intoextracellular vesicles during their biogenesis are not fullyunderstood, increasing evidence indicates that the clustering,oligomerization, or protein–protein interaction among vesicularproteins play critical roles in vesicular cargo sorting (Vidal,Mangeat, & Hoekstra, 1997; Le Naour et al., 2006; Choiet al., 2012b). We previously reported that extracellular vesiclesharbor functional modules that are comprised of physicallyinterconnected tetraspanin web proteins (Choi et al., 2012b).The tetraspanin web is a well-known highly clustered proteincomplex in the cell (Le Naour et al., 2006). Moreover, theantibody-mediated cross-linking of plasma membrane proteins,such as the transferrin receptor in reticulocytes or the MHC II inlymphocytes, induces the sorting of these proteins into extracel-lular vesicles (Vidal, Mangeat, & Hoekstra, 1997).
B. Comparison of Extracellular Vesicles With OtherSubcellular Compartments
As shown in Figure 7, we next conducted analysis to identifybiological processes enriched or depleted in extracellularvesicles relative to other intracellular organelles or compart-ments using a modified Fisher’s exact test in DAVID (http://david.abcc.ncifcrf.gov) (Huang da, Sherman, & Lempicki,2009; Choi et al., 2012b, 2013a). We previously found thatspecific biological processes were highly enriched in extracellu-lar vesicles derived from HT29 human colorectal cancer cells(Choi et al., 2012b). Consistent with this observation, the
B cell Platelet SW480
FIGURE 6. Intravesicular protein–protein interaction networks of mammalian extracellular vesicles. Vesicularproteins are interconnected via physical interactions, and the overall topologies of protein–protein interactionnetworks of mammalian extracellular vesicles follow the properties of scale-free networks by indicating thepresence several hub proteins with many other vesicular proteins having few interacting partners, as reportedpreviously (Choi et al., 2012b, 2013a).
484 Mass Spectrometry Reviews DOI 10.1002/mas
& CHOI ET AL.

functional annotations of vesicular proteins indicate that specificbiological processes are highly enriched or depleted in threemammalian extracellular vesicles when compared with otherintracellular compartments (Fig. 7). The vesicular protein list ofenriched or depleted biological processes is available fromEVpedia (http://evpedia.info) as Supplementary Table. All threeextracellular vesicles are highly enriched with the followingvesicle structure- and vesicle biogenesis-related biologicalprocesses: protein localization, protein transport, vesicle-medi-ated transport, protein targeting, regulation of protein ubiquiti-nation, membrane organization, protein complex biogenesis,
regulation of organelle organization, cytoskeleton organization,actin cytoskeleton organization, and small GTPase-mediatedsignal transduction (Raimondo et al., 2011; Choi et al., 2012b,2013a). These results suggest that extracellular vesicles areextracellular organelles that are distinct from other intracellularorganelles and compartments (Choi et al., 2012b, 2013a).
Interestingly, SW480-, B cell- and platelet-derived extra-cellular vesicles are enriched with cell-type-specific biologicalprocesses. For example, extracellular vesicles that are derivedfrom SW480 human colorectal cancer cells are specificallyenriched with cell-cell junction assembly, cell morphogenesis,
SW
480
B c
ell
Pla
tele
t P
lasm
a m
embr
ane
Cyt
osol
N
ucle
us
End
opla
smic
retic
ulum
G
olgi
app
arat
us
Lyso
som
e Ext
race
llula
r ve
sicl
es
Enriched biological process in B cell-derived
extracellular vesicles
Enriched biological process in Platelet-derived
extracellular vesicles
Enriched biological process in SW480-derived
extracellular vesicles
P-value
1 10-1 10-4 10-2 10-3
Protein localization Protein transport Vesicle-mediated transport Protein targeting Protein folding Regulation of protein modification process Regulation of protein ubiquitination Membrane organization Protein complex biogenesis Regulation of organelle organization Cytoskeleton organization Actin cytoskeleton organization Regulation of cell death Cell adhesion Cell motion Cell cycle Response to protein stimulus Small GTPase mediated signal transduction Generation of precursor metabolites and energy Glycolysis Cell-cell junction organization Cell-cell junction assembly Cell morphogenesis Cell morphogenesis involved in differentiation Cell projection morphogenesis Cell adhesion mediated by integrin Leukocyte activation Lymphocyte activation T cell activation Regulation of body fluid levels Hemostasis Wound healing Coagulation Platelet activation
FIGURE 7. Comparison of extracellular vesicles with other subcellular organelles or compartments. Thefunctional annotations of vesicular proteins with specific biological processes are highly enriched or depleted inthree mammalian extracellular vesicles when compared with other intracellular organelles or compartments.Colors indicate the statistical significance of enriched or depleted Gene Ontology biological process annotations.The vesicular protein list of enriched or depleted biological processes is available from EVpedia (http://evpedia.info) as Supplementary Table.
Mass Spectrometry Reviews DOI 10.1002/mas 485
PROTEOMICS OF EXTRACELLULAR VESICLES &

cell morphogenesis involved in differentiation, cell projectionmorphogenesis, and cell adhesion mediated by integrin process-es. These are well-known biological processes that are involvedin epithelial-mesenchymal transitions in human cancer (Yang &Weinberg, 2008; Choi et al., 2012a). B cell-derived extracellularvesicles are enriched with leukocyte, lymphocyte, and T cellactivation-associated proteins, whereas platelet-derived extra-cellular vesicles are specifically enriched with the regulation ofbody fluid levels, hemostasis, wound healing, coagulation, andplatelet activation. These observations suggest that extracellularvesicles contain not only the common sets of vesicular proteinsbut also cell-type-specific proteins that are associated with cell-type-specific pathophysiological functions (de Gassart et al.,2004; Choi et al., 2013a).
C. Functional Modules in the IntravesicularProtein–Protein Interaction Networks ofMammalian Extracellular Vesicles
Based on functional enrichment analyses (Fig. 7) and literaturemining (Choi et al., 2012b), we constructed subnetworks to
define common functional modules that are clustered withvesicle-enriched common biological processes: vesicle-mediat-ed transport (Fig. 8A) and cytoskeleton organization (Fig. 8B).The functional modules of the coatomer family, ESCRTs,exocytosis, RAB proteins, and 14-3-3 proteins are clustered inthe vesicle-mediated transport subnetwork (Fig. 8A), whereasthe cytoskeleton organization-related subnetwork is clusteredwith functional modules of the actin cytoskeleton, Arp2/3proteins, and ezrin/radixin/moesin (Fig. 8B). These commonfunctional modules in the vesicle-mediated transport andcytoskeleton organization-related subnetworks are known to beinvolved in the biogenesis and structure of extracellular vesicles(Laulagnier et al., 2004, 2005; Choi et al., 2012b, 2013a).
We also found that three different extracellular vesiclesharbor cell-type-specific functional modules that are related byphysical and functional interconnections (Fig. 8C). The func-tional modules of junction proteins, leukocyte activation, andcoagulation are only present in extracellular vesicles that arederived from SW480 human colorectal cancer cells, B cells, andplatelets, respectively. Junction proteins are important formaintaining the cellular integrity of and polarity in intestinalepithelial cells, and a loss of normal junctions could be a
FIGURE 8. Functional modules clustered with enriched biological processes in mammalian extracellularvesicles. Common functional modules in the vesicle-mediated transport subnetworks (A) and cytoskeletonorganization-related subnetworks (B) are present in all extracellular vesicles that are derived from SW480 humancolorectal cancer cells, B cells, and platelets. These common functional modules are known to be involved in thebiogenesis and structure of extracellular vesicles (Laulagnier et al., 2004, 2005; Choi et al., 2012b, 2013a). C:Cell-type-specific functional modules in mammalian extracellular vesicles. The functional modules of junctionproteins, leukocyte activation, and coagulation are only present in extracellular vesicles derived from SW480human colorectal cancer cells, B cells, and platelets, respectively.
486 Mass Spectrometry Reviews DOI 10.1002/mas
& CHOI ET AL.

hallmark of a human carcinoma (Conacci-Sorrell, Zhurinsky, &Ben-Ze’ev, 2002; Martin & Jiang, 2009; Dusek & Attardi,2011). Moreover, the abnormal regulation of the junctionproteins leads to a loss of epithelial polarity and an over-proliferation of cells (Lu & Bilder, 2005). It has been reportedthat B cell-derived extracellular vesicles play roles in immunemodulation, such as B cell and T cell activation (McLellan,2009). For example, MS4A1 and CD22 are involved in B cellactivation, whereas CD48 and SLAMF1 are involved in T cellactivation. B cell-derived extracellular vesicles play roles inimmune modulation, such as B cell and T cell activation(McLellan, 2009). For example, MS4A1 and CD22 are involvedin B cell activation, whereas CD48 and SLAMF1 are involved inT cell activation. Platelet-derived extracellular vesicles containthe coagulation-associated functional module. This result isconsistent with the previous observation that procoagulantenzymatic activity is one of the main pathophysiologicalfunctions of platelet-derived extracellular vesicles (Cocucci,Racchetti, & Meldolesi, 2009). Taken together, all theseobservations suggest that extracellular vesicles contain commonand cell-type-specific functional modules.
V. CONCLUSIONS AND PERSPECTIVES
Extracellular vesicle-mediated communication is an evolution-arily conserved process among bacteria, archaea, and eukaryotes(Lee et al., 2008, 2009; Deatherage & Cookson, 2012; Choiet al., 2013a). Growing evidence indicates that these extracellu-lar vesicles, including mammalian exosomes and ectosomes, areextracellular organelles (i.e., nanocosmoses) that play diverseroles in intercellular communication in multicellular communi-ties. In this review, we focused on the high-throughput massspectrometry-based proteomic analyses of mammalian extracel-lular vesicles (i.e., exosomes and ectosomes), introduced anintegrated database of high-throughput data for the systematicanalyses of extracellular vesicles (EVpedia), and highlightedintravesicular protein–protein interaction network analyses.
As purification technologies and proteomic analyticalmethods have been developed, more reliable vesicular pro-teomes have been identified. Moreover, a systems biologyapproach via protein–protein interaction network analysisprovides the functional relationship between these vesicularproteins (Choi et al., 2012b, 2013a). These approaches providecomprehensive information for understanding vesicle biogene-sis and cargo-sorting mechanisms as well as their pathophysio-logical roles. Furthermore, extracellular vesicles can be isolatedfrom various biological fluids with enriched surface proteins oftheir cells of origin, leading to the discovery of biomarkers.Together with conventional biological research, advancedproteomic analyses with the quantitation of vesicular proteinswill provide new knowledge and aid in the comprehension of thecomplex extracellular vesicle-mediated intercellular communi-cation system.
However, the identification of vesicular proteins with highconfidence will be critical for the global understanding of theemerging biology of extracellular vesicles. Previous high-throughput mass spectrometry-based proteomic studies haveidentified several hundred or, in some cases, more than 1,000,vesicular proteins. Moreover, the list and number of identifiedvesicular proteins varies between proteomic studies that wereconducted on extracellular vesicles derived from the same cells
or body fluids (Choi et al., 2012a; Ji et al., 2013). Theseobservations raise an issue regarding whether these proteins aregenuine components of extracellular vesicles because there is aconcern that falsely detected proteins could be included in thesingle proteomics dataset (Stuart et al., 2007; Choi et al.,2012b). First, the isolation of mammalian extracellular vesicleswithout any contamination by non-vesicular components, in-cluding serum abundant proteins, apoptotic bodies, and/orprotein aggregates, is a critical prerequisite for high-throughputmass spectrometry-based proteomic analyses (Choi et al.,2011). Therefore, after the isolation of extracellular vesicles, thecharacterization of the purified extracellular vesicles to indicatetheir presence without other non-vesicular contaminants shouldbe essential to identify the highly confident vesicular proteinsby subsequent mass spectrometry-based proteomic analysis(Fig. 5). The combination of different types of vesicle isolationtechnologies or repetitive density gradient ultracentrifugationsteps should be a promising isolation method to purify theextracellular vesicles without any of the potential contamina-tions that have been previously reported (Choi et al., 2011; Tauroet al., 2013).
In addition, to be more confident that the identified proteinsare components of extracellular vesicles, researchers will needto acquire more than two proteomic datasets including quantita-tive proteomic analyses. Recently, we provided a platform fordefining candidate vesicular proteins by integrating proteomicdata using genomics and systems approaches (Choi et al.,2012b). From the 1,364 proteins that were identified from fiveproteomic analyses of human colorectal cancer cell-derivedextracellular vesicles, we defined 957 vesicular proteins that areexpressed in parental cells based on microarray data and are alsointerconnected proteins that form an intravesicular protein–protein interaction network. We believe that this approach willprovide a rationale for cataloging the genuine proteins withinextracellular vesicles that are derived from various types of cellsand body fluids.
The other important issue that is raised by current proteo-mic studies is the uncertainty of how many different types ofextracellular vesicles are secreted by a single cell. Consideringthe size of mammalian extracellular vesicles, the presence ofseveral vesicular proteins and nucleic acids, including mRNAsand miRNAs, suggests the possibility that a single cell secretesmany different types of extracellular vesicles (Choi et al.,2012b). Further studies to either provide experimental evidenceof how many proteins an extracellular vesicle can harbor ordetermine how many vesicular proteins are present in a singletype of extracellular vesicles or in a single extracellular vesiclewill be of great value to understand the pathophysiological rolesof mammalian extracellular vesicles as well as their biogenesisand cargo-sorting mechanisms (Choi et al., 2012b).
VI. ABBREVIATIONS
APEX absolute protein expressionESCRT endosomal sorting complexes required for transportMVB multivesicular body
ACKNOWLEDGMENTS
We thank to Yae Jin Yoon, Ji Hyun Kim, Su Chul Jang, Kyong-Su Park, Oh Youn Kim, Jaewook Lee, Sae Rom Kim, and
Mass Spectrometry Reviews DOI 10.1002/mas 487
PROTEOMICS OF EXTRACELLULAR VESICLES &

Gyeongyun Go for manual curation of publications on extracel-lular vesicle studies in Figure 2. This work was supported by theNational Research Foundation of Korea (NRF) grant fundedby the Korea Government (MEST) (no. 2013035248 and no.2012R1A1A2042534), the Ministry of Health andWelfare grantfunded by the Korea Government (no. A120273), and a grantKorea Basic Science Institute (D33400).
REFERENCES
Admyre C, Johansson SM, Qazi KR, Filen JJ, Lahesmaa R, Norman M,
Neve EP, Scheynius A, Gabrielsson S. 2007. Exosomes with immune
modulatory features are present in human breast milk. J Immunol
179:1969–1978.
Al-Nedawi K, Meehan B, Micallef J, Lhotak V, May L, Guha A, Rak J.
2008. Intercellular transfer of the oncogenic receptor EGFRvIII by
microvesicles derived from tumour cells. Nat Cell Biol 10:619–624.
Bachi A, Bonaldi T. 2008. Quantitative proteomics as a new piece of the
systems biology puzzle. J Proteomics 71:357–367.
Baietti MF, Zhang Z, Mortier E, Melchior A, Degeest G, Geeraerts A,
Ivarsson Y, Depoortere F, Coomans C, Vermeiren E, Zimmermann P,
David G. 2012. Syndecan-syntenin-ALIX regulates the biogenesis of
exosomes. Nat Cell Biol 14:677–685.
Beckler MD, Higginbotham JN, Franklin JL, Ham AJ, Halvey PJ, Imasuen
IE,Whitwell C, Li M, Liebler DC, Coffey RJ. 2013. Proteomic analysis
of exosomes from mutant KRAS colon cancer cells identifies
intercellular transfer of mutant KRAS. Mol Cell Proteomics 12:343–
355.
Bosman GJ, Lasonder E, Luten M, Roerdinkholder-Stoelwinder B, Novotny
VM, Bos H, De Grip WJ. 2008. The proteome of red cell membranes
and vesicles during storage in blood bank conditions. Transfusion
48:827–835.
Buschow SI, van Balkom BW, Aalberts M, Heck AJ, Wauben M, Stoorvogel
W. 2010. MHC class II-associated proteins in B-cell exosomes and
potential functional implications for exosome biogenesis. Immunol
Cell Biol 88:851–856.
Carayon K, Chaoui K, Ronzier E, Lazar I, Bertrand-Michel J, Roques V,
Balor S, Terce F, Lopez A, Salome L, Joly E. 2011. Proteolipidic
composition of exosomes changes during reticulocyte maturation.
J Biol Chem 286:34426–34439.
Chen CL, Lai YF, Tang P, Chien KY, Yu JS, Tsai CH, Chen HW, Wu CC,
Chung T, Hsu CW, Chen CD, Chang YS, Chang PL, Chen YT. 2012.
Comparative and targeted proteomic analyses of urinary microparticles
from bladder cancer and hernia patients. J Proteome Res 11:5611–
5629.
Choi DS, Lee JM, Park GW, Lim HW, Bang JY, Kim YK, Kwon KH, Kwon
HJ, Kim KP, Gho YS. 2007. Proteomic analysis of microvesicles
derived from human colorectal cancer cells. J Proteome Res 6:4646–
4655.
Choi DS, Park JO, Jang SC, Yoon YJ, Jung JW, Choi DY, Kim JW, Kang
JS, Park J, Hwang D, Lee KH, Park SH, Kim YK, Desiderio DM,
Kim KP, Gho YS. 2011. Proteomic analysis of microvesicles
derived from human colorectal cancer ascites. Proteomics 11:2745–
2751.
Choi DS, Choi DY, Hong BS, Jang SC, Kim DK, Lee J, Kim YK, Kim KP,
Gho YS. 2012a. Quantitative proteomics of extracellular vesicles
derived from human primary and metastatic colorectal cancer cells. J
Extracell Ves 1:18704.
Choi DS, Yang JS, Choi EJ, Jang SC, Park S, Kim OY, Hwang D, Kim KP,
Kim YK, Kim S, Gho YS. 2012b. The protein interaction network of
extracellular vesicles derived from human colorectal cancer cells. J
Proteome Res 11:1144–1151.
Choi DS, Kim DK, Kim YK, Gho YS. 2013a. Proteomics, transcriptomics
and lipidomics of exosomes and ectosomes. Proteomics 13:1554–
1571.
Choi DS, Lee J, Go G, Kim YK, Gho YS. 2013b. Circulating extracellular
vesicles in cancer diagnosis and monitoring: An appraisal of clinical
potential. Mol Diagn Ther 17:265–271.
Cocucci E, Racchetti G, Meldolesi J. 2009. Shedding microvesicles:
Artefacts no more. Trends Cell Biol 19:43–51.
Coleman BM, Hanssen E, Lawson VA, Hill AF. 2012. Prion-infected cells
regulate the release of exosomes with distinct ultrastructural features.
FASEB J 26:4160–4173.
Conacci-Sorrell M, Zhurinsky J, Ben-Ze’ev A. 2002. The cadherin-catenin
adhesion system in signaling and cancer. J Clin Invest 109:987–991.
de Gassart A, Geminard C, Hoekstra D, Vidal M. 2004. Exosome secretion:
The art of reutilizing nonrecycled proteins? Traffic 5:896–903.
Dean WL, Lee MJ, Cummins TD, Schultz DJ, Powell DW. 2009. Proteomic
and functional characterisation of platelet microparticle size classes.
ThrombHaemost 102:711–718.
Deatherage BL, Cookson BT. 2012. Membrane vesicle release in bacteria,
eukaryotes, and archaea: A conserved yet underappreciated aspect of
microbial life. Infect Immun 80:1948–1957.
DenekaM, Pelchen-Matthews A, Byland R, Ruiz-Mateos E, MarshM. 2007.
In macrophages, HIV-1 assembles into an intracellular plasma
membrane domain containing the tetraspanins CD81, CD9, and CD53.
J Cell Biol 177:329–341.
Di Vizio D, Kim J, Hager MH, Morello M, Yang W, Lafargue CJ, True LD,
Rubin MA, Adam RM, Beroukhim R, Demichelis F, Freeman MR.
2009. Oncosome formation in prostate cancer: Association with a
region of frequent chromosomal deletion in metastatic disease. Cancer
Res 69:5601–5609.
Dusek RL, Attardi LD. 2011. Desmosomes: New perpetrators in tumour
suppression. Nat Rev Cancer 11:317–323.
Garcia BA, Smalley DM, Cho H, Shabanowitz J, Ley K, Hunt DF. 2005. The
platelet microparticle proteome. J Proteome Res 4:1516–1521.
Gonzalez-Begne M, Lu B, Han X, Hagen FK, Hand AR, Melvin JE, Yates
JR. 2009. Proteomic analysis of human parotid gland exosomes by
multidimensional protein identification technology (MudPIT). J Prote-
ome Res 8:1304–1314.
Gyorgy B, Szabo TG, Pasztoi M, Pal Z, Misjak P, Aradi B, Laszlo V,
Pallinger E, Pap E, Kittel A, Nagy G, Falus A, Buzas EI. 2011.
Membrane vesicles, current state-of-the-art: Emerging role of extracel-
lular vesicles. Cell Mol Life Sci 68:2667–2688.
Hong BS, Cho JH, Kim H, Choi EJ, Rho S, Kim J, Kim JH, Choi DS, Kim
YK, Hwang D, Gho YS. 2009. Colorectal cancer cell-derived micro-
vesicles are enriched in cell cycle-related mRNAs that promote
proliferation of endothelial cells. BMCGenomics 10:556.
Huang da W, Sherman BT, Lempicki RA. 2009. Systematic and integrative
analysis of large gene lists using DAVID bioinformatics resources. Nat
Protoc 4:44–457.
Hurley JH, Hanson PI. 2010. Membrane budding and scission by the ESCRT
machinery: It’s all in the neck. Nat RevMol Cell Biol 11:556–566.
Ji H, Greening DW, Barnes TW, Lim JW, Tauro BJ, Rai A, Xu R, Adda C,
Mathivanan S, Zhao W, Xue Y, Xu T, Zhu HJ, Simpson RJ. 2013.
Proteome profiling of exosomes derived from human primary and
metastatic colorectal cancer cells reveal differential expression of key
metastatic factors and signal transduction components. Proteomics
13:1672–1686.
Kalra H, Simpson RJ, Ji H, Aikawa E, Altevogt P, Askenase P, Bond VC,
Borras FE, Breakefield X, Budnik V, Buzas E, Camussi G, Clayton A,
Cocucci E, Falcon-Perez JM, Gabrielsson S, Gho YS, Gupta D, Harsha
HC, Hendrix A, Hill AF, Inal JM, Jenster G, Kramer-Albers EM, Lim
SK, Llorente A, Lotvall J, Marcilla A, Mincheva-Nilsson L, Nazarenko
I, Nieuwland R, Nolte-’t Hoen EN, Pandey A, Patel T, Piper MG,
Pluchino S, Prasad TS, Rajendran L, Raposo G, Record M, Reid GE,
Sanchez-Madrid F, Schiffelers RM, Siljander P, Stensballe A, Stoorvo-
gel W, Taylor D, Thery C, Valadi H, van Balkom BW, Vazquez J, Vidal
M, Wauben MH, Yanez-Mo M, Zoeller M, Mathivanan S. 2012.
Vesiclepedia: A compendium for extracellular vesicles with continuous
community annotation. PLoS Biol 10:e1001450.
488 Mass Spectrometry Reviews DOI 10.1002/mas
& CHOI ET AL.

Kang D, Oh S, Ahn SM, Lee BH, Moon MH. 2008. Proteomic analysis of
exosomes from human neural stem cells by flow field-flow fraction-
ation and nanoflow liquid chromatography-tandem mass spectrometry.
J Proteome Res 7:3475–3480.
Kim CW, Lee HM, Lee TH, Kang C, Kleinman HK, Gho YS. 2002.
Extracellular membrane vesicles from tumor cells promote angiogene-
sis via sphingomyelin. Cancer Res 62:6312–6317.
Kim DK, Kang B, Kim OY, Choi DS, Lee J, Kim SR, Go G, Yoon YJ, Kim
JH, Jang SC, Park KS, Choi EJ, Kim KP, Desiderio DM, Kim YK,
Lotvall J, Hwang D, Gho YS. 2013. EVpedia: An integrated database
of high-throughput data for systemic analyses of extracellular vesicles.
J Extracell Ves 2:20384.
Kosaka N, Iguchi H, Ochiya T. 2010. Circulating microRNA in body fluid: A
new potential biomarker for cancer diagnosis and prognosis. Cancer
Sci 101:2087–22092.
Laulagnier K, Grand D, Dujardin A, Hamdi S, Vincent-Schneider H, Lankar
D, Salles JP, Bonnerot C, Perret B, Record M. 2004. PLD2 is enriched
on exosomes and its activity is correlated to the release of exosomes.
FEBS Lett 572:11–14.
Laulagnier K, Vincent-Schneider H, Hamdi S, Subra C, Lankar D, Record
M. 2005. Characterization of exosome subpopulations from RBL-2H3
cells using fluorescent lipids. Blood Cells Mol Dis 35:116–121.
Le Bihan MC, Bigot A, Jensen SS, Dennis JL, Rogowska-Wrzesinska A,
Laine J, Gache V, Furling D, Jensen ON, Voit T, Mouly V, Coulton GR,
Butler-Browne G. 2012. In-depth analysis of the secretome identifies
three major independent secretory pathways in differentiating human
myoblasts. J Proteomics 77:344–356.
Le Naour F, Andre M, Boucheix C, Rubinstein E. 2006. Membrane
microdomains and proteomics: Lessons from tetraspanin microdo-
mains and comparison with lipid rafts. Proteomics 6:6447–6454.
Lee EY, Choi DS, Kim KP, Gho YS. 2008. Proteomics in gram-negative
bacterial outer membrane vesicles. Mass Spectrom Rev 27:535–555.
Lee EY, Choi DY, Kim DK, Kim JW, Park JO, Kim S, Kim SH, Desiderio
DM, Kim YK, Kim KP, Gho YS. 2009. Gram-positive bacteria
produce membrane vesicles: Proteomics-based characterization of
Staphylococcus aureus-derived membrane vesicles. Proteomics
9:5425–5436.
Lee HM, Choi EJ, Kim JH, Kim TD, Kim YK, Kang C, Gho YS. 2010. A
membranous form of ICAM-1 on exosomes efficiently blocks leuko-
cyte adhesion to activated endothelial cells. Biochem Biophys Res
Commun 397:251–256.
Lehmann BD, Paine MS, Brooks AM, McCubrey JA, Renegar RH, Wang R,
Terrian DM. 2008. Senescence-associated exosome release from
human prostate cancer cells. Cancer Res 68:7864–7871.
Li B, Antonyak MA, Zhang J, Cerione RA. 2012. RhoA triggers a specific
signaling pathway that generates transforming microvesicles in cancer
cells. Oncogene 31:4740–4749.
Looze C, Yui D, Leung L, Ingham M, Kaler M, Yao X, Wu WW, Shen RF,
Daniels MP, Levine SJ. 2009. Proteomic profiling of human plasma
exosomes identifies PPARgamma as an exosome-associated protein.
BiochemBiophys Res Commun 378:433–438.
Lu H, Bilder D. 2005. Endocytic control of epithelial polarity and
proliferation in Drosophila. Nat Cell Biol 7:1232–1239.
Lu P, Vogel C, Wang R, Yao X, Marcotte EM. 2007. Absolute protein
expression profiling estimates the relative contributions of transcrip-
tional and translational regulation. Nat Biotechnol 25:117–124.
Martin TA, Jiang WG. 2009. Loss of tight junction barrier function and its
role in cancer metastasis. Biochim Biophys Acta 1788:872–891.
Mathivanan S, Simpson RJ. 2009. ExoCarta: A compendium of exosomal
proteins and RNA. Proteomics 9:4997–5000.
Mathivanan S, Lim JW, Tauro BJ, Ji H, Moritz RL, Simpson RJ. 2010.
Proteomics analysis of A33 immunoaffinity-purified exosomes re-
leased from the human colon tumor cell line LIM1215 reveals a tissue-
specific protein signature. Mol Cell Proteomics 9:197–208.
McLellan AD. 2009. Exosome release by primary B cells. Crit Rev Immunol
29:203–217.
Moon PG, Lee JE, You S, Kim TK, Cho JH, Kim IS, Kwon TH, Kim CD,
Park SH, Hwang D, Kim YL, Baek MC. 2011. Proteomic analysis of
urinary exosomes from patients of early IgA nephropathy and thin
basement membrane nephropathy. Proteomics 11:2459–2475.
Moskovich O, Fishelson Z. 2007. Live cell imaging of outward and inward
vesiculation induced by the complement c5b-9 complex. J Biol Chem
282:29977–29986.
Muralidharan-Chari V, Clancy J, Plou C, Romao M, Chavrier P, Raposo
G, D’Souza-Schorey C. 2009a. ARF6-regulated shedding of tumor
cell-derived plasma membrane microvesicles. Curr Biol 19:1875–
1885.
Muralidharan-Chari V, Hoover H, Clancy J, Schweitzer J, Suckow MA,
Schroeder V, Castellino FJ, Schorey JS, D’Souza-Schorey C. 2009b.
ADP-ribosylation factor 6 regulates tumorigenic and invasive proper-
ties in vivo. Cancer Res 69:2201–2209.
Pan BT, Teng K,WuC, AdamM, Johnstone RM. 1985. Electron microscopic
evidence for externalization of the transferrin receptor in vesicular
form in sheep reticulocytes. J Cell Biol 101:942–948.
Pap E, Pallinger E, Pasztoi M, Falus A. 2009. Highlights of a new type of
intercellular communication: Microvesicle-based information transfer.
InflammRes 58:1–8.
Peterson DB, Sander T, Kaul S, Wakim BT, Halligan B, Twigger S, Pritchard
KA, Jr., Oldham KT, Ou JS. 2008. Comparative proteomic analysis of
PAI-1 and TNF-alpha-derived endothelial microparticles. Proteomics
8:2430–2446.
Pilzer D, Gasser O, Moskovich O, Schifferli JA, Fishelson Z. 2005. Emission
of membrane vesicles: Roles in complement resistance, immunity and
cancer. Springer Semin Immunopathol 27:375–387.
Pisitkun T, Shen RF, Knepper MA. 2004. Identification and proteomic
profiling of exosomes in human urine. Proc Natl Acad Sci USA
101:13368–13373.
Raimondo F, Morosi L, Chinello C, Magni F, Pitto M. 2011. Advances in
membranous vesicle and exosome proteomics improving biological
understanding and biomarker discovery. Proteomics 11:709–720.
Ramacciotti E, Hawley AE, Wrobleski SK, Myers DD, Jr., Strahler JR,
Andrews PC, Guire KE, Henke PK, Wakefield TW. 2010. Proteomics
of microparticles after deep venous thrombosis. Thromb Res 125:
e269–274.
Ratajczak J, Wysoczynski M, Hayek F, Janowska-Wieczorek A, Ratajczak
MZ. 2006. Membrane-derived microvesicles: Important and underap-
preciated mediators of cell-to-cell communication. Leukemia
20:1487–1495.
Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N,
Schwikowski B, Ideker T. 2003. Cytoscape: A software environment
for integrated models of biomolecular interaction networks. Genome
Res 13:2498–2504.
Simons M, Raposo G. 2009. Exosomes–vesicular carriers for intercellular
communication. Curr Opin Cell Biol 21:575–581.
Skokos D, Botros HG, Demeure C, Morin J, Peronet R, Birkenmeier G,
Boudaly S, Mecheri S. 2003. Mast cell-derived exosomes induce
phenotypic and functional maturation of dendritic cells and elicit
specific immune responses in vivo. J Immunol 170:3037–3045.
Smalley DM, Sheman NE, Nelson K, Theodorescu D. 2008. Isolation and
identification of potential urinary microparticle biomarkers of bladder
cancer. J Proteome Res 7:2088–2096.
Stuart LM, Boulais J, Charriere GM, Hennessy EJ, Brunet S, Jutras I,
Goyette G, Rondeau C, Letarte S, Huang H, Ye P, Morales F, Kocks C,
Bader JS, Desjardins M, Ezekowitz RA. 2007. A systems biology
analysis of the Drosophila phagosome. Nature 445:95–101.
Subra C, Laulagnier K, Perret B, Record M. 2007. Exosome lipidomics
unravels lipid sorting at the level of multivesicular bodies. Biochimie
89:205–212.
Tauro BJ, Greening DW, Mathias RA, Mathivanan S, Ji H, Simpson RJ.
2013. Two distinct populations of exosomes are released from
LIM1863 colon carcinoma cell-derived organoids. Mol Cell Proteo-
mics 12:587–598.
Mass Spectrometry Reviews DOI 10.1002/mas 489
PROTEOMICS OF EXTRACELLULAR VESICLES &

Thery C, Regnault A, Garin J, Wolfers J, Zitvogel L, Ricciardi-Castagnoli P,
Raposo G, Amigorena S. 1999. Molecular characterization of dendritic
cell-derived exosomes. Selective accumulation of the heat shock
protein hsc73. J Cell Biol 147:599–610.
Thery C, Boussac M, Veron P, Ricciardi-Castagnoli P, Raposo G, Garin J,
Amigorena S. 2001. Proteomic analysis of dendritic cell-derived
exosomes: A secreted subcellular compartment distinct from apoptotic
vesicles. J Immunol 166:7309–7318.
Thery C, Amigorena S, Raposo G, Clayton A. 2006. Isolation and
characterization of exosomes from cell culture supernatants and
biological fluids. Curr Protoc Cell Biol Chapter 3: Unit 3.22.
Trajkovic K, Hsu C, Chiantia S, Rajendran L,Wenzel D,Wieland F, Schwille
P, Brugger B, Simons M. 2008. Ceramide triggers budding of exosome
vesicles into multivesicular endosomes. Science 319:1244–1247.
Utleg AG, Yi EC, Xie T, Shannon P, White JT, Goodlett DR, Hood L, Lin B.
2003. Proteomic analysis of human prostasomes. Prostate 56:150–161.
Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall J. 2007.
Exosome-mediated transfer of mRNAs and microRNAs is a novel
mechanism of genetic exchange between cells. Nat Cell Biol 9:654–
659.
van Niel G, Raposo G, Candalh C, Boussac M, Hershberg R, Cerf-Bensussan
N, Heyman M. 2001. Intestinal epithelial cells secrete exosome-like
vesicles. Gastroenterology 121:337–349.
Vidal M, Mangeat P, Hoekstra D. 1997. Aggregation reroutes molecules
from a recycling to a vesicle-mediated secretion pathway during
reticulocyte maturation. J Cell Sci 110:1867–1877.
Witwer KW, Buzas EI, Bemis LT, Bora A, Lasser C, Lotvall J, Nolte-’t
Hoen EN, Piper MG, Sivaraman S, Skog J, Thery C, Wauben MH,
Hochberg F. 2013. Standardization of sample collection, isolation and
analysis methods in extracellular vesicle research. J Extracell Ves
2:20360.
Wolf P. 1967. The nature and significance of platelet products in human
plasma. Br J Haematol 13:269–2288.
Yang J, Weinberg RA. 2008. Epithelial-mesenchymal transition: At the
crossroads of development and tumor metastasis. Dev Cell 14:818–
829.
Dong-Sic Choi received B.Sc. and Ph.D. degrees in the Department of Life Sciences fromPohang University of Science and Technology, Republic of Korea, in 2006 and 2012,respectively. He is now a Research Assistant Professor at Pohang University of Science andTechnology, Republic of Korea. His current research areas focus on the proteomic analysis andsystems biology of extracellular vesicles derived from eukaryotes and prokaryotes and clinicalapplications based on extracellular vesicles, including biomarker discovery, diagnosis tooldevelopment, and drug delivery systems.
Dae-Kyum Kim received a B.Sc. degree in the Department of Life Sciences from PohangUniversity of Science and Technology, Republic of Korea, in 2009. He is now a Ph.D. Candidateat Pohang University of Science and Technology, Republic of Korea. His current research areasfocus on the high-throughput analysis and systems biology of extracellular vesicles derivedfrom eukaryotes and prokaryotes. More recently, he reported a study regarding the integrateddatabase of high-throughput data for the systematic analysis of extracellular vesicles: EVpedia(http://evpedia.info).
Yoon-Keun Kim, M.D., Ph.D. is a physician scientist. He received his M.D. from SeoulNational University in 1987 and received his Ph.D. in 1997. From 2002 to 2003, he was aVisiting Assistant Professor at Yale University, USA. He is currently a Professor in theDepartment of Life Sciences at the Pohang University of Science and Technology, Republic ofKorea. His research interest is the pathogenesis of allergic diseases and chronic inflammatorydiseases, such as asthma, atopic dermatitis, diabetes, obesity, gastritis, and inflammatory boweldiseases. His research focuses on the physiological relevance of the pathogenesis of allergicdiseases, and he published several papers on the immunopathogenesis of asthma and allergicdiseases. More recently, his research has focused on the relationship between extracellularvesicles from bacteria and inflammatory and metabolic diseases. He has reported several studiesregarding the medical importance of bacteria-derived extracellular vesicles on the pathogenesisof immune-based inflammatory diseases.
Yong Song Gho received his B.Sc. and M.Sc. degrees in Chemistry from Seoul NationalUniversity, Republic of Korea, in 1987 and 1989, respectively. He obtained his Ph.D. degree inBiochemistry and Biophysics from the University of North Carolina at Chapel Hill, USA, in1997. Currently, he is an Associate Professor in the Department of Life Sciences at the PohangUniversity of Science and Technology, Republic of Korea. His group is primarily studying thegenomic and proteomic components of extracellular vesicles using systems biology approachesand the pathophysiological functions of the extracellular vesicles derived from eukaryotes andprokaryotes. He is now serving as an Executive Board Member of the “International Society ofExtracellular Vesicles (ISEV)” and as the Editor-in-Chief of the “Journal of ExtracellularVesicles” launched by the ISEV.
490 Mass Spectrometry Reviews DOI 10.1002/mas
& CHOI ET AL.