paradoxical activation of endothelial nitric oxide synthase by nadph oxidase
TRANSCRIPT

FultonChantemele, Botond Banfi, Mario B. Marrero, R. Daniel Rudic, David W. Stepp and David J.R.
Qian Zhang, Pulkit Malik, Deepesh Pandey, Sonali Gupta, Davin Jagnandan, Eric Belin deParadoxical Activation of Endothelial Nitric Oxide Synthase by NADPH Oxidase
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2008 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/ATVBAHA.108.1682782008;28:1627-1633; originally published online June 12, 2008;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/28/9/1627World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2008/06/12/ATVBAHA.108.168278.DC1.htmlData Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from

Paradoxical Activation of Endothelial Nitric Oxide Synthaseby NADPH Oxidase
Qian Zhang, Pulkit Malik, Deepesh Pandey, Sonali Gupta, Davin Jagnandan, Eric Belin de Chantemele,Botond Banfi, Mario B. Marrero, R. Daniel Rudic, David W. Stepp, David J.R. Fulton
Objectives—Increased formation of reactive oxygen species (ROS) has been identified as a causative factor in endothelialdysfunction by reducing NO bioavailability and uncoupling endothelial nitric oxide synthase (eNOS). However, thespecific contribution of ROS to endothelial function is not well understood.
Methods and Results—A major source of intracellular ROS is the NADPH oxidase (Nox) family of enzymes. The goalof the current study was to directly assess the contribution of NADPH oxidase derived superoxide to eNOS function byexpressing Nox5, a single gene product that constitutively produces superoxide within cells. Paradoxically, we foundthat instead of inhibiting eNOS, coexpression of Nox5 increased NO release from both bovine and human endothelialcells. To establish the functional significance of this observation in intact blood vessels, the endothelium of mouse aortawas transduced with Nox5 or control adenoviruses. Nox5 potently inhibited Ach-induced relaxation and potentiatedcontractile responses to phenylephrine. In precontracted aortae, acute exposure to superoxide dismutase inducedsignificant vascular relaxation in vessels exposed to Nox5 versus control and unmasked the ability of Nox5 to activateeNOS in blood vessel endothelium.
Conclusions—These findings suggest that ROS inhibit eNOS function via consumption of NO rather than direct inhibitionof enzymatic activity. (Arterioscler Thromb Vasc Biol. 2008;28:1627-1633)
Key Words: nitric oxide � endothelial nitric oxide synthase � NADPH oxidase � endothelial cells� superoxide dismutase
Reactive oxygen species (ROS) are leading candidates inthe etiology of endothelial dysfunction and ensuing
cardiovascular disease.1 Increased superoxide formation inendothelial cells (EC) has been identified as a causative factorin this process by reducing NO bioavailability, uncouplingeNOS via BH4 depletion or homodimer disruption, and alsoby altering redox-sensitive signaling cascades. However,elevated superoxide production is frequently accompanied bychanges in blood pressure, cellular signaling, hormones, andthe composition of the extracellular milieu, making it difficultto ascertain the independent effects of intracellular ROS. Forexample, endothelial function is reduced in animal models oftype II diabetes, angiotensin-dependent hypertension, andatherosclerosis, and this deficit is accompanied by significantincreases in superoxide formation.2–4 However, whether in-creased superoxide is the causative factor, a participant, orrequires the cooperation of other factors present in theextracellular milieu is not yet known.
See accompanying article on page 1580Within vascular cells, there are several sources of ROS
including the mitochondrial electron transport chain, xanthineoxidase, arachidonic acid oxygenases (lipogenase, cyclooxy-genase, cytochrome P450), uncoupled eNOS, and NADPH
oxidases (Noxs).5 However, superoxide production is anunintended secondary product in all of these enzymes exceptfor the Nox isoforms, which have acquired the uniquecapacity to exclusively synthesize superoxide. Endothelialcells primarily express Nox2 and Nox4,6,7 and changes in theexpression or activity of these enzymes have been reported indiabetes,8,9 hypertension, and in response to angiotensinII.10–12 The functional consequences of superoxide produc-tion from NADPH oxidases versus other cellular sources suchas the mitochondria are not well defined. Increased superox-ide anion and other reactive oxygen species have beenproposed to contribute to endothelial dysfunction,3 elevatedblood pressure,13,14 and increased cellular proliferation andhypertrophy.15,16 The restricted intracellular distribution ofNoxs to specific locations such as the endoplasmic reticulum(ER) suggests that they may have important roles in modu-lating discrete aspects of intracellular signaling. However, theindividual role of NADPH oxidases in these processes re-mains uncertain because of the presence of multiple Noxisoforms and their binding partners and the lack of specificinhibitors.17
Thus, the goal of our current study is to identify thefunctional consequences of increased Nox activity in the
Original received April 4, 2008; final version accepted June 2, 2008.From the Departments of Pharmacology, Physiology, and the Vascular Biology Center, Medical College of Georgia, Augusta.Correspondence to David J.R. Fulton, PhD, Vascular Biology Center, Medical College of Georgia, Augusta, GA 30912-2500. E-mail [email protected]© 2008 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.108.168278
1627 by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from

vascular endothelium as a functional correlate to the elevatedNox activity seen in cardiovascular disease. To achieve thiswe have adopted a novel genetic approach of using the Nox5gene to produce superoxide at a time and place of ourchoosing. This strategy has several advantages. First, theability of Nox5 to produce superoxide is contained within asingle gene. This is important, as the protein-binding partnersrequired to activate the other Nox isoforms are not necessaryand thus their absence or inactivity cannot impede superoxideformation.14 For example, Nox2 requires the coexpression ofat least 4 different gene products just to have the capacity toproduce superoxide and then requires a stimulus, such asPMA, to induce activity. Second, Nox5 is a calcium-activatedenzyme and produces low levels of superoxide constitutivelyand therefore is active in the vast majority of vascular cells.18
Third, the intracellular distribution of Nox5 is consistent withthat described for Nox4 and Nox2 in endothelium,19–22 andconsequently the location of superoxide production fromNox5 should reliably replicate that derived from otherNox(s).
Materials and MethodsMaterialsAll chemicals and reagents were purchased from Sigma. Antibodies:GAPDH (Ambion), HA (Roche), hsp90, phosphorylated eNOS,(Cell Signaling, Millipore).
AnimalsC57bl6 mice (Jackson Laboratories Bar Harbor, Maine) were used inaccordance with the guidelines for animal use of the InstitutionalAnimal Care and Use Committee of the Medical College of Georgia.
Cell Culture and TransfectionCOS-7 cells, bovine aortic endothelial cells (BAECs), human aorticendothelial cells (HAEC) were grown as previously described.23
Replication-deficient adenoviruses encoding the control virusesGFP, �-gal or HA-Nox5 were generated as described previously.19,23
Adenoviral gene delivery into blood vessel endothelium has beendescribed in detail elsewhere.24
NO ReleaseThirty-six hours after transfection or adenoviral transduction, net NOrelease was calculated by NO specific chemiluminescence aftersubtracting background levels from cells treated with L-NMMA,cells without eNOS, and unstimulated basal release as describedpreviously.25
cGMP Reporter AssayHAECs were grown on glass coverslips and acutely placed over thetop of HAVSMCs, a source of soluble guanylyl cyclase (sGC). TheHAECs were stimulated with 1 �mol/L ionomycin in the presence of100U/mL SOD and 300 �mol/L IBMX for 10 minutes. cGMPcontent was measured in HVSMCs using a cGMP specific EIA(Cayman).
Isometric Measurements of Aortic ToneAortic rings (1 mm in length) were mounted on 2 wires in a 6-mLchamber vessel myograph (Danish Myo Technology) with 1 g basaltension. After equilibration, rings were tested for reproduciblecontraction using 32 mmol/L KCl. Concentration-response curveswere then constructed to phenylephrine (PE) and subsequently toACh in vessels precontracted with a submaximal concentrationof PE.26
Statistical AnalysisData are expressed as means�SEM. Comparisons were made using2-tailed Student t test or analysis of variance with a posthoc testwhere appropriate. Differences were considered significant atP�0.05.
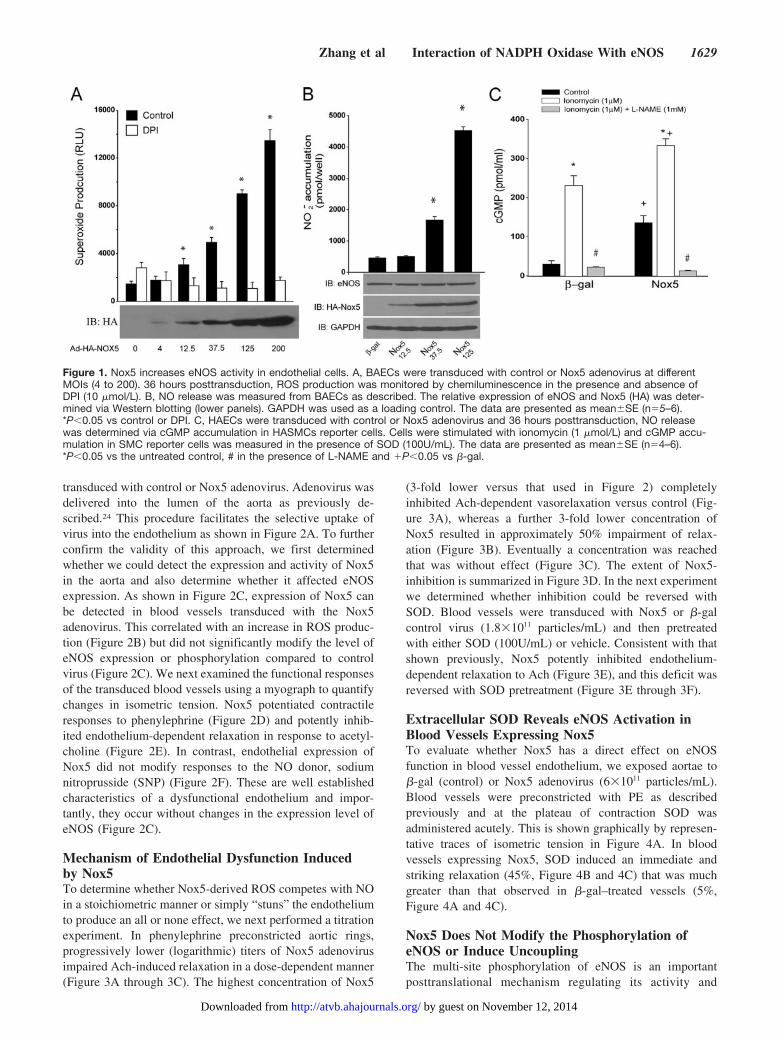
ResultsNox5 Increases eNOS Activity in Both TransfectedCOS-7 Cells and Endothelial CellsTo evaluate the direct interaction between eNOS andNADPH oxidase, we transfected COS-7 cells with cDNAsencoding eNOS, with or without the epitope tagged, HA-Nox5. In contrast to our expectations, we found that Nox5increased eNOS activity as determined by the detection ofnitrite (NO2
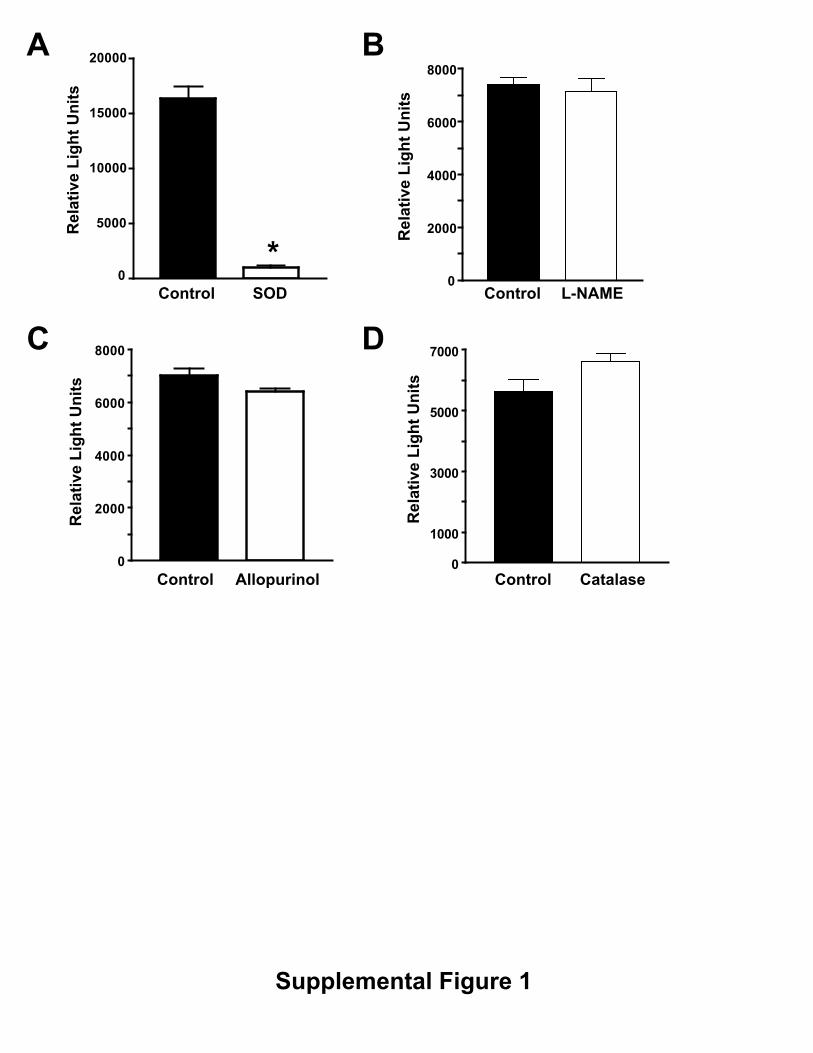
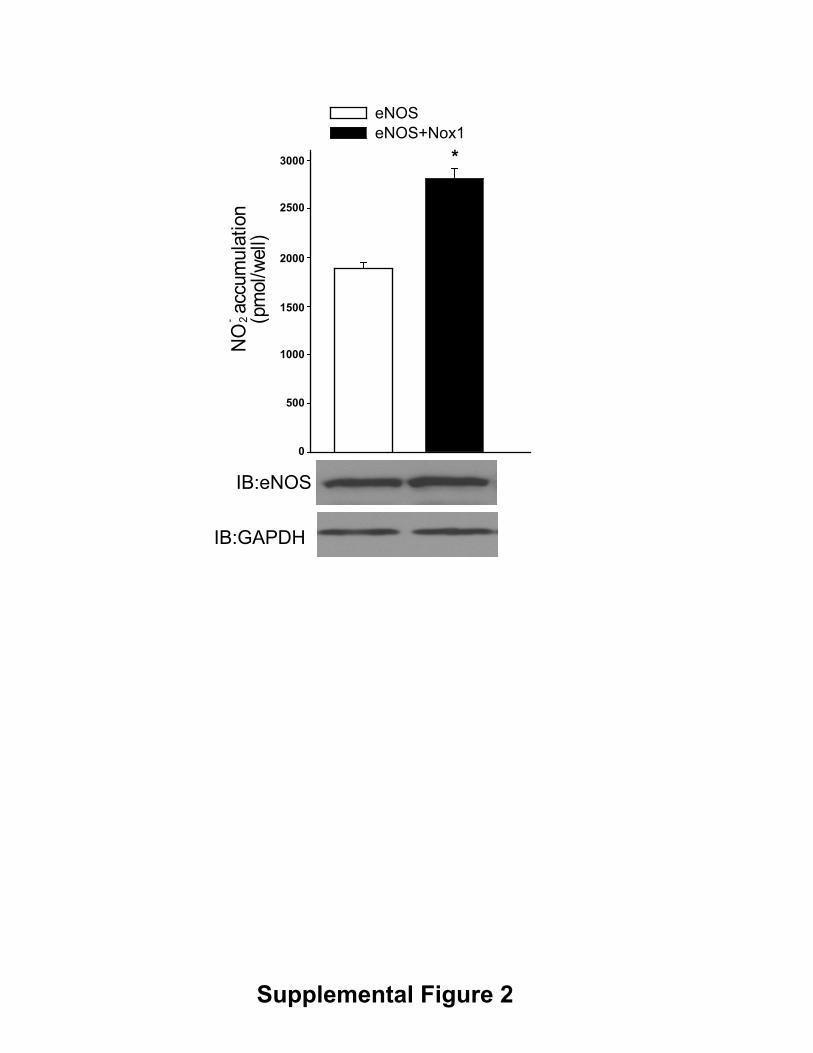
�) in the extracellular media. This was apparentunder both basal and 100 nmol/L thapsigargin stimulatedconditions (data not shown). We next extended these studiesto bovine aortic endothelial cells (BAECs), which weretransduced with control (�-gal) or Nox5 expressing adenovi-ruses. Increased expression of Nox5 (MOI ranging from 4 to200) resulted in the dose-dependent increases in superoxideproduction (Figure 1A) and corresponding dose-dependentincreases in NO release (Figure 1B). The Nox5-dependentincrease in superoxide production in BAECs was sensitive toDPI and SOD inhibition, but was not inhibited by L-NAME,catalase, or allopurinol (supplemental Figure 1, availableonline at http://atvb.ahajournals.org). The increase in eNOSactivity occurred without variation in total eNOS expression(Figure 1B lower panel) suggesting that the mechanismsresponsible for the increase in NO release are posttransla-tional. Furthermore, coexpression of active Nox1 was alsocapable of increasing eNOS activity (supplemental Figure II).Our next goal was to assess the effect of increased ROS onthe release of biologically active NO. To achieve this we useda coculture cGMP bioassay in which glass coverslips coatedwith human aortic endothelial cells (HAECs) are placed overthe top of human aortic vascular smooth muscle cells(HAVSMCs, a source of soluble guanylyl cyclase [sGC] thatfunctions as a bio-detector). cGMP content in HAVSMCswas then measured using a cGMP specific EIA. In thepresence of extracellular SOD, selective expression of Nox5in HAECs increased cGMP production under both basalconditions and following ionomycin stimulation (Figure 1C),data consistent with that obtained in BAECs and COS cells.eNOS expression in HAECs was unchanged (data notshown). These findings demonstrate that the increase incGMP levels in vascular smooth muscle cell (VSMCs) deriveexclusively from Nox5 modulation of endothelial eNOSactivity. In the absence of extracellular SOD, basal cGMPproduction dropped significantly to levels equivalent toL-NAME treatment (11.4746�1.1357 versus5.4924�0.8834pmol/mL). Collectively, these data show thatincreased superoxide via expression of Nox5 activates, ratherthan inhibits, eNOS activity and that the major inhibitoryaction of superoxide anion is to consume biologically activeNO.
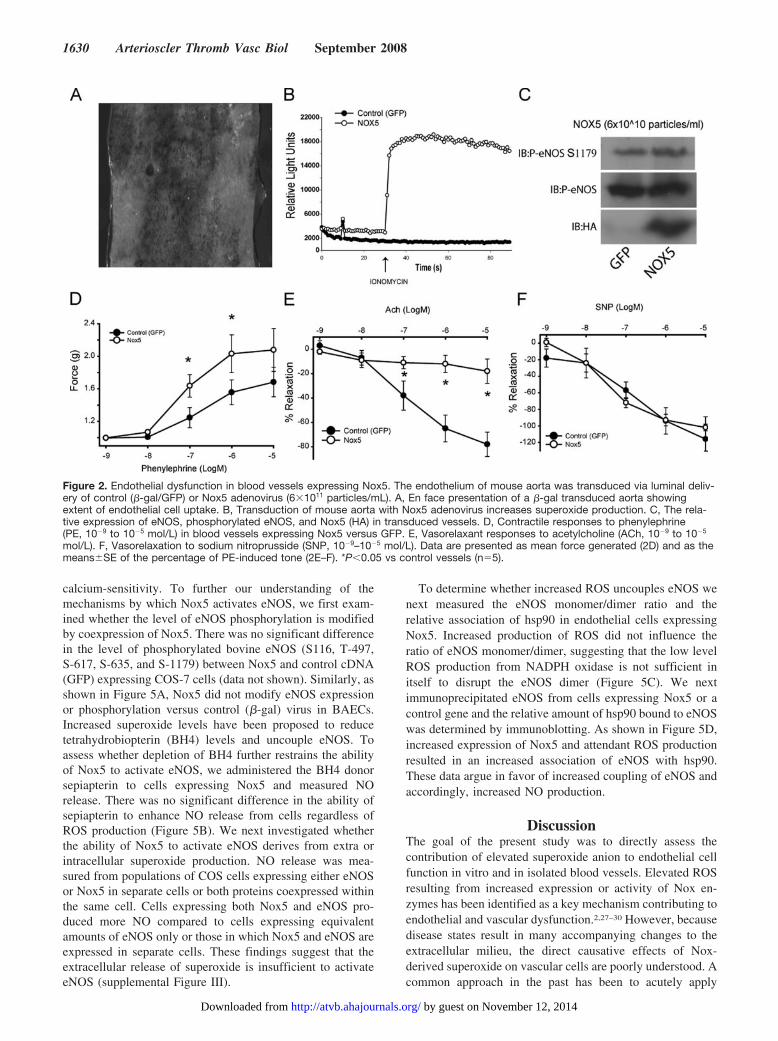
Nox5 Induces Endothelial Dysfunction in IsolatedBlood VesselsTo establish the vascular significance of this relationship inthe intact blood vessel, the endothelium of mouse aorta was
1628 Arterioscler Thromb Vasc Biol September 2008
by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from
transduced with control or Nox5 adenovirus. Adenovirus wasdelivered into the lumen of the aorta as previously de-scribed.24 This procedure facilitates the selective uptake ofvirus into the endothelium as shown in Figure 2A. To furtherconfirm the validity of this approach, we first determinedwhether we could detect the expression and activity of Nox5in the aorta and also determine whether it affected eNOSexpression. As shown in Figure 2C, expression of Nox5 canbe detected in blood vessels transduced with the Nox5adenovirus. This correlated with an increase in ROS produc-tion (Figure 2B) but did not significantly modify the level ofeNOS expression or phosphorylation compared to controlvirus (Figure 2C). We next examined the functional responsesof the transduced blood vessels using a myograph to quantifychanges in isometric tension. Nox5 potentiated contractileresponses to phenylephrine (Figure 2D) and potently inhib-ited endothelium-dependent relaxation in response to acetyl-choline (Figure 2E). In contrast, endothelial expression ofNox5 did not modify responses to the NO donor, sodiumnitroprusside (SNP) (Figure 2F). These are well establishedcharacteristics of a dysfunctional endothelium and impor-tantly, they occur without changes in the expression level ofeNOS (Figure 2C).
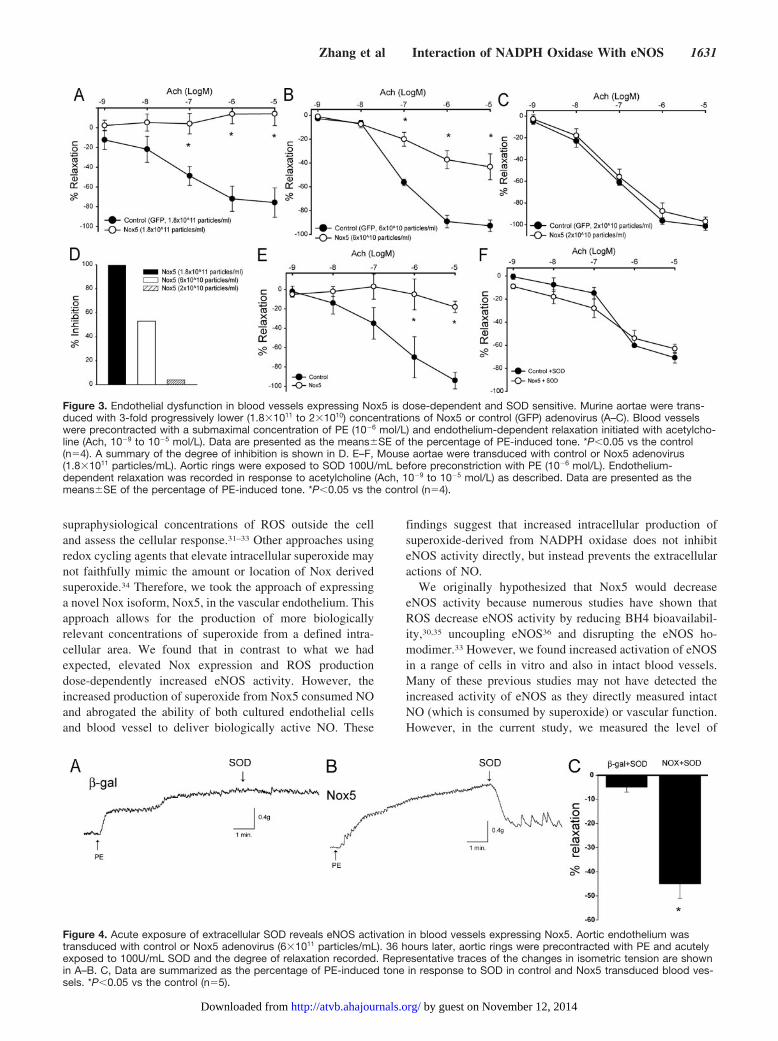
Mechanism of Endothelial Dysfunction Inducedby Nox5To determine whether Nox5-derived ROS competes with NOin a stoichiometric manner or simply “stuns” the endotheliumto produce an all or none effect, we next performed a titrationexperiment. In phenylephrine preconstricted aortic rings,progressively lower (logarithmic) titers of Nox5 adenovirusimpaired Ach-induced relaxation in a dose-dependent manner(Figure 3A through 3C). The highest concentration of Nox5
(3-fold lower versus that used in Figure 2) completelyinhibited Ach-dependent vasorelaxation versus control (Fig-ure 3A), whereas a further 3-fold lower concentration ofNox5 resulted in approximately 50% impairment of relax-ation (Figure 3B). Eventually a concentration was reachedthat was without effect (Figure 3C). The extent of Nox5-inhibition is summarized in Figure 3D. In the next experimentwe determined whether inhibition could be reversed withSOD. Blood vessels were transduced with Nox5 or �-galcontrol virus (1.8�1011 particles/mL) and then pretreatedwith either SOD (100U/mL) or vehicle. Consistent with thatshown previously, Nox5 potently inhibited endothelium-dependent relaxation to Ach (Figure 3E), and this deficit wasreversed with SOD pretreatment (Figure 3E through 3F).
Extracellular SOD Reveals eNOS Activation inBlood Vessels Expressing Nox5To evaluate whether Nox5 has a direct effect on eNOSfunction in blood vessel endothelium, we exposed aortae to�-gal (control) or Nox5 adenovirus (6�1011 particles/mL).Blood vessels were preconstricted with PE as describedpreviously and at the plateau of contraction SOD wasadministered acutely. This is shown graphically by represen-tative traces of isometric tension in Figure 4A. In bloodvessels expressing Nox5, SOD induced an immediate andstriking relaxation (45%, Figure 4B and 4C) that was muchgreater than that observed in �-gal–treated vessels (5%,Figure 4A and 4C).
Nox5 Does Not Modify the Phosphorylation ofeNOS or Induce UncouplingThe multi-site phosphorylation of eNOS is an importantposttranslational mechanism regulating its activity and
Figure 1. Nox5 increases eNOS activity in endothelial cells. A, BAECs were transduced with control or Nox5 adenovirus at differentMOIs (4 to 200). 36 hours posttransduction, ROS production was monitored by chemiluminescence in the presence and absence ofDPI (10 �mol/L). B, NO release was measured from BAECs as described. The relative expression of eNOS and Nox5 (HA) was deter-mined via Western blotting (lower panels). GAPDH was used as a loading control. The data are presented as mean�SE (n�5–6).*P�0.05 vs control or DPI. C, HAECs were transduced with control or Nox5 adenovirus and 36 hours posttransduction, NO releasewas determined via cGMP accumulation in HASMCs reporter cells. Cells were stimulated with ionomycin (1 �mol/L) and cGMP accu-mulation in SMC reporter cells was measured in the presence of SOD (100U/mL). The data are presented as mean�SE (n�4–6).*P�0.05 vs the untreated control, # in the presence of L-NAME and �P�0.05 vs �-gal.
Zhang et al Interaction of NADPH Oxidase With eNOS 1629
by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from
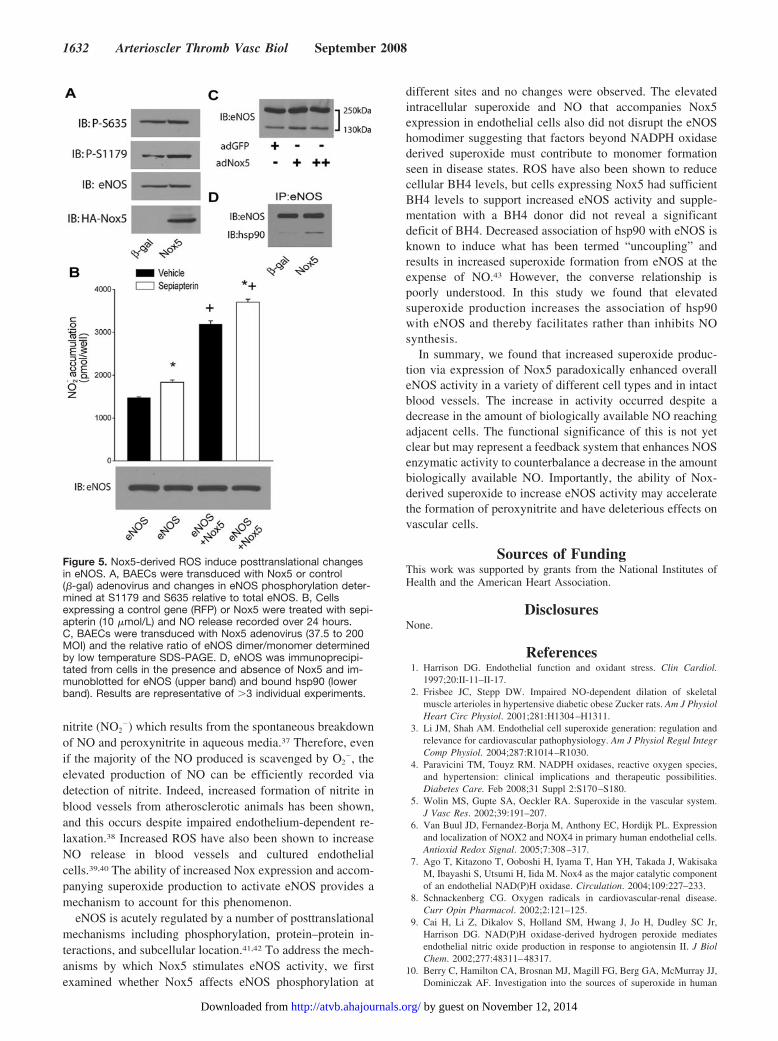
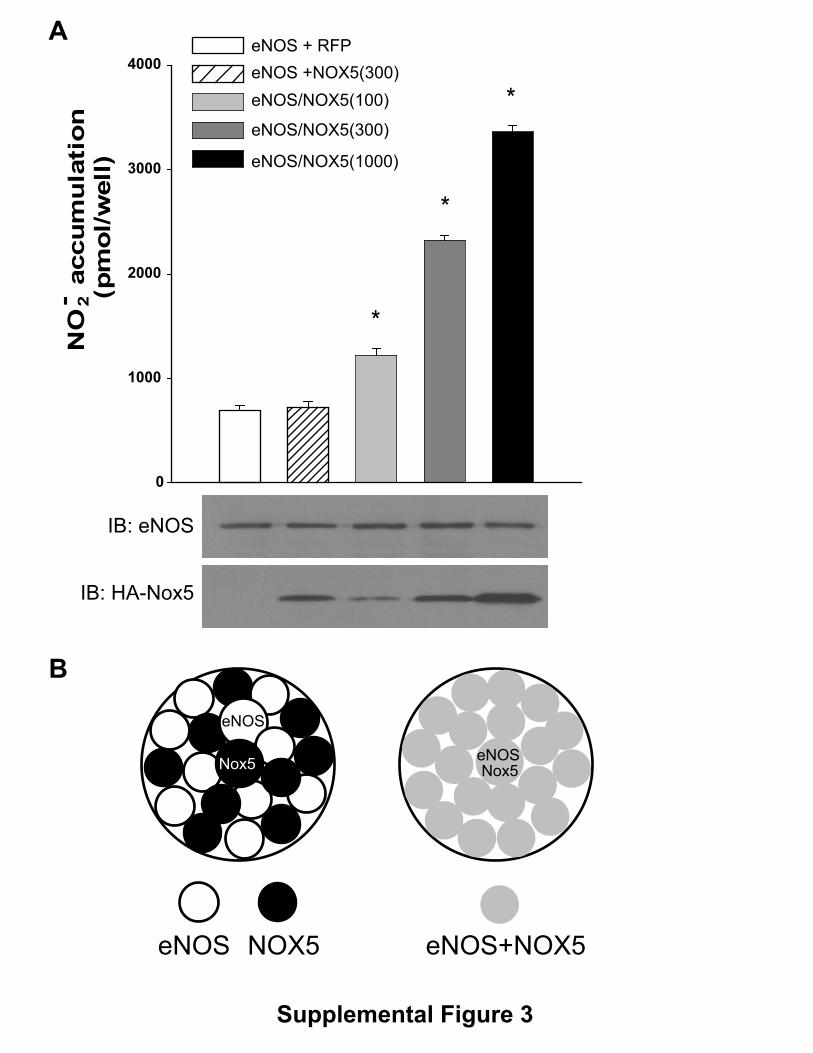
calcium-sensitivity. To further our understanding of themechanisms by which Nox5 activates eNOS, we first exam-ined whether the level of eNOS phosphorylation is modifiedby coexpression of Nox5. There was no significant differencein the level of phosphorylated bovine eNOS (S116, T-497,S-617, S-635, and S-1179) between Nox5 and control cDNA(GFP) expressing COS-7 cells (data not shown). Similarly, asshown in Figure 5A, Nox5 did not modify eNOS expressionor phosphorylation versus control (�-gal) virus in BAECs.Increased superoxide levels have been proposed to reducetetrahydrobiopterin (BH4) levels and uncouple eNOS. Toassess whether depletion of BH4 further restrains the abilityof Nox5 to activate eNOS, we administered the BH4 donorsepiapterin to cells expressing Nox5 and measured NOrelease. There was no significant difference in the ability ofsepiapterin to enhance NO release from cells regardless ofROS production (Figure 5B). We next investigated whetherthe ability of Nox5 to activate eNOS derives from extra orintracellular superoxide production. NO release was mea-sured from populations of COS cells expressing either eNOSor Nox5 in separate cells or both proteins coexpressed withinthe same cell. Cells expressing both Nox5 and eNOS pro-duced more NO compared to cells expressing equivalentamounts of eNOS only or those in which Nox5 and eNOS areexpressed in separate cells. These findings suggest that theextracellular release of superoxide is insufficient to activateeNOS (supplemental Figure III).
To determine whether increased ROS uncouples eNOS wenext measured the eNOS monomer/dimer ratio and therelative association of hsp90 in endothelial cells expressingNox5. Increased production of ROS did not influence theratio of eNOS monomer/dimer, suggesting that the low levelROS production from NADPH oxidase is not sufficient initself to disrupt the eNOS dimer (Figure 5C). We nextimmunoprecipitated eNOS from cells expressing Nox5 or acontrol gene and the relative amount of hsp90 bound to eNOSwas determined by immunoblotting. As shown in Figure 5D,increased expression of Nox5 and attendant ROS productionresulted in an increased association of eNOS with hsp90.These data argue in favor of increased coupling of eNOS andaccordingly, increased NO production.
DiscussionThe goal of the present study was to directly assess thecontribution of elevated superoxide anion to endothelial cellfunction in vitro and in isolated blood vessels. Elevated ROSresulting from increased expression or activity of Nox en-zymes has been identified as a key mechanism contributing toendothelial and vascular dysfunction.2,27–30 However, becausedisease states result in many accompanying changes to theextracellular milieu, the direct causative effects of Nox-derived superoxide on vascular cells are poorly understood. Acommon approach in the past has been to acutely apply
Figure 2. Endothelial dysfunction in blood vessels expressing Nox5. The endothelium of mouse aorta was transduced via luminal deliv-ery of control (�-gal/GFP) or Nox5 adenovirus (6�1011 particles/mL). A, En face presentation of a �-gal transduced aorta showingextent of endothelial cell uptake. B, Transduction of mouse aorta with Nox5 adenovirus increases superoxide production. C, The rela-tive expression of eNOS, phosphorylated eNOS, and Nox5 (HA) in transduced vessels. D, Contractile responses to phenylephrine(PE, 10�9 to 10�5 mol/L) in blood vessels expressing Nox5 versus GFP. E, Vasorelaxant responses to acetylcholine (ACh, 10�9 to 10�5
mol/L). F, Vasorelaxation to sodium nitroprusside (SNP, 10�9–10�5 mol/L). Data are presented as mean force generated (2D) and as themeans�SE of the percentage of PE-induced tone (2E–F). *P�0.05 vs control vessels (n�5).
1630 Arterioscler Thromb Vasc Biol September 2008
by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from
supraphysiological concentrations of ROS outside the celland assess the cellular response.31–33 Other approaches usingredox cycling agents that elevate intracellular superoxide maynot faithfully mimic the amount or location of Nox derivedsuperoxide.34 Therefore, we took the approach of expressinga novel Nox isoform, Nox5, in the vascular endothelium. Thisapproach allows for the production of more biologicallyrelevant concentrations of superoxide from a defined intra-cellular area. We found that in contrast to what we hadexpected, elevated Nox expression and ROS productiondose-dependently increased eNOS activity. However, theincreased production of superoxide from Nox5 consumed NOand abrogated the ability of both cultured endothelial cellsand blood vessel to deliver biologically active NO. These
findings suggest that increased intracellular production ofsuperoxide-derived from NADPH oxidase does not inhibiteNOS activity directly, but instead prevents the extracellularactions of NO.
We originally hypothesized that Nox5 would decreaseeNOS activity because numerous studies have shown thatROS decrease eNOS activity by reducing BH4 bioavailabil-ity,30,35 uncoupling eNOS36 and disrupting the eNOS ho-modimer.33 However, we found increased activation of eNOSin a range of cells in vitro and also in intact blood vessels.Many of these previous studies may not have detected theincreased activity of eNOS as they directly measured intactNO (which is consumed by superoxide) or vascular function.However, in the current study, we measured the level of
Figure 3. Endothelial dysfunction in blood vessels expressing Nox5 is dose-dependent and SOD sensitive. Murine aortae were trans-duced with 3-fold progressively lower (1.8�1011 to 2�1010) concentrations of Nox5 or control (GFP) adenovirus (A–C). Blood vesselswere precontracted with a submaximal concentration of PE (10�6 mol/L) and endothelium-dependent relaxation initiated with acetylcho-line (Ach, 10�9 to 10�5 mol/L). Data are presented as the means�SE of the percentage of PE-induced tone. *P�0.05 vs the control(n�4). A summary of the degree of inhibition is shown in D. E–F, Mouse aortae were transduced with control or Nox5 adenovirus(1.8�1011 particles/mL). Aortic rings were exposed to SOD 100U/mL before preconstriction with PE (10�6 mol/L). Endothelium-dependent relaxation was recorded in response to acetylcholine (Ach, 10�9 to 10�5 mol/L) as described. Data are presented as themeans�SE of the percentage of PE-induced tone. *P�0.05 vs the control (n�4).
Figure 4. Acute exposure of extracellular SOD reveals eNOS activation in blood vessels expressing Nox5. Aortic endothelium wastransduced with control or Nox5 adenovirus (6�1011 particles/mL). 36 hours later, aortic rings were precontracted with PE and acutelyexposed to 100U/mL SOD and the degree of relaxation recorded. Representative traces of the changes in isometric tension are shownin A–B. C, Data are summarized as the percentage of PE-induced tone in response to SOD in control and Nox5 transduced blood ves-sels. *P�0.05 vs the control (n�5).
Zhang et al Interaction of NADPH Oxidase With eNOS 1631
by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from
nitrite (NO2�) which results from the spontaneous breakdown
of NO and peroxynitrite in aqueous media.37 Therefore, evenif the majority of the NO produced is scavenged by O2
�, theelevated production of NO can be efficiently recorded viadetection of nitrite. Indeed, increased formation of nitrite inblood vessels from atherosclerotic animals has been shown,and this occurs despite impaired endothelium-dependent re-laxation.38 Increased ROS have also been shown to increaseNO release in blood vessels and cultured endothelialcells.39,40 The ability of increased Nox expression and accom-panying superoxide production to activate eNOS provides amechanism to account for this phenomenon.
eNOS is acutely regulated by a number of posttranslationalmechanisms including phosphorylation, protein–protein in-teractions, and subcellular location.41,42 To address the mech-anisms by which Nox5 stimulates eNOS activity, we firstexamined whether Nox5 affects eNOS phosphorylation at
different sites and no changes were observed. The elevatedintracellular superoxide and NO that accompanies Nox5expression in endothelial cells also did not disrupt the eNOShomodimer suggesting that factors beyond NADPH oxidasederived superoxide must contribute to monomer formationseen in disease states. ROS have also been shown to reducecellular BH4 levels, but cells expressing Nox5 had sufficientBH4 levels to support increased eNOS activity and supple-mentation with a BH4 donor did not reveal a significantdeficit of BH4. Decreased association of hsp90 with eNOS isknown to induce what has been termed “uncoupling” andresults in increased superoxide formation from eNOS at theexpense of NO.43 However, the converse relationship ispoorly understood. In this study we found that elevatedsuperoxide production increases the association of hsp90with eNOS and thereby facilitates rather than inhibits NOsynthesis.
In summary, we found that increased superoxide produc-tion via expression of Nox5 paradoxically enhanced overalleNOS activity in a variety of different cell types and in intactblood vessels. The increase in activity occurred despite adecrease in the amount of biologically available NO reachingadjacent cells. The functional significance of this is not yetclear but may represent a feedback system that enhances NOSenzymatic activity to counterbalance a decrease in the amountbiologically available NO. Importantly, the ability of Nox-derived superoxide to increase eNOS activity may acceleratethe formation of peroxynitrite and have deleterious effects onvascular cells.
Sources of FundingThis work was supported by grants from the National Institutes ofHealth and the American Heart Association.
DisclosuresNone.
References1. Harrison DG. Endothelial function and oxidant stress. Clin Cardiol.
1997;20:II-11–II-17.2. Frisbee JC, Stepp DW. Impaired NO-dependent dilation of skeletal
muscle arterioles in hypertensive diabetic obese Zucker rats. Am J PhysiolHeart Circ Physiol. 2001;281:H1304–H1311.
3. Li JM, Shah AM. Endothelial cell superoxide generation: regulation andrelevance for cardiovascular pathophysiology. Am J Physiol Regul IntegrComp Physiol. 2004;287:R1014–R1030.
4. Paravicini TM, Touyz RM. NADPH oxidases, reactive oxygen species,and hypertension: clinical implications and therapeutic possibilities.Diabetes Care. Feb 2008;31 Suppl 2:S170–S180.
5. Wolin MS, Gupte SA, Oeckler RA. Superoxide in the vascular system.J Vasc Res. 2002;39:191–207.
6. Van Buul JD, Fernandez-Borja M, Anthony EC, Hordijk PL. Expressionand localization of NOX2 and NOX4 in primary human endothelial cells.Antioxid Redox Signal. 2005;7:308–317.
7. Ago T, Kitazono T, Ooboshi H, Iyama T, Han YH, Takada J, WakisakaM, Ibayashi S, Utsumi H, Iida M. Nox4 as the major catalytic componentof an endothelial NAD(P)H oxidase. Circulation. 2004;109:227–233.
8. Schnackenberg CG. Oxygen radicals in cardiovascular-renal disease.Curr Opin Pharmacol. 2002;2:121–125.
9. Cai H, Li Z, Dikalov S, Holland SM, Hwang J, Jo H, Dudley SC Jr,Harrison DG. NAD(P)H oxidase-derived hydrogen peroxide mediatesendothelial nitric oxide production in response to angiotensin II. J BiolChem. 2002;277:48311–48317.
10. Berry C, Hamilton CA, Brosnan MJ, Magill FG, Berg GA, McMurray JJ,Dominiczak AF. Investigation into the sources of superoxide in human
Figure 5. Nox5-derived ROS induce posttranslational changesin eNOS. A, BAECs were transduced with Nox5 or control(�-gal) adenovirus and changes in eNOS phosphorylation deter-mined at S1179 and S635 relative to total eNOS. B, Cellsexpressing a control gene (RFP) or Nox5 were treated with sepi-apterin (10 �mol/L) and NO release recorded over 24 hours.C, BAECs were transduced with Nox5 adenovirus (37.5 to 200MOI) and the relative ratio of eNOS dimer/monomer determinedby low temperature SDS-PAGE. D, eNOS was immunoprecipi-tated from cells in the presence and absence of Nox5 and im-munoblotted for eNOS (upper band) and bound hsp90 (lowerband). Results are representative of �3 individual experiments.
1632 Arterioscler Thromb Vasc Biol September 2008
by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from

blood vessels: angiotensin II increases superoxide production in humaninternal mammary arteries. Circulation. 2000;101:2206–2212.
11. Pagano PJ, Clark JK, Cifuentes-Pagano ME, Clark SM, Callis GM, QuinnMT. Localization of a constitutively active, phagocyte-like NADPHoxidase in rabbit aortic adventitia: enhancement by angiotensin II. ProcNatl Acad Sci U S A. 1997;94:14483–14488.
12. Hanna IR, Taniyama Y, Szocs K, Rocic P, Griendling KK. NAD(P)Hoxidase-derived reactive oxygen species as mediators of angiotensin IIsignaling. Antioxid Redox Signal. 2002;4:899–914.
13. Dikalova A, Clempus R, Lassegue B, Cheng G, McCoy J, Dikalov S, SanMartin A, Lyle A, Weber DS, Weiss D, Taylor WR, Schmidt HH, OwensGK, Lambeth JD, Griendling KK. Nox1 overexpression potentiates an-giotensin II-induced hypertension and vascular smooth muscle hypertro-phy in transgenic mice. Circulation. 2005;112:2668–2676.
14. Bedard K, Krause KH. The NOX family of ROS-generating NADPHoxidases: physiology and pathophysiology. Physiol Rev. 2007;87:245–313.
15. Ushio-Fukai M, Zafari AM, Fukui T, Ishizaka N, Griendling KK.p22phox is a critical component of the superoxide-generatingNADH/NADPH oxidase system and regulates angiotensin II-inducedhypertrophy in vascular smooth muscle cells. J Biol Chem. 1996;271:23317–23321.
16. Zafari AM, Ushio-Fukai M, Akers M, Yin Q, Shah A, Harrison DG,Taylor WR, Griendling KK. Role of NADH/NADPH oxidase-derivedH2O2 in angiotensin II-induced vascular hypertrophy. Hypertension.1998;32:488–495.
17. Heumuller S, Wind S, Barbosa-Sicard E, Schmidt HH, Busse R, SchroderK, Brandes RP. Apocynin is not an inhibitor of vascular NADPHoxidases but an antioxidant. Hypertension. 2008;51:211–217.
18. Banfi B, Malgrange B, Knisz J, Steger K, Dubois-Dauphin M, KrauseKH. NOX3, a superoxide-generating NADPH oxidase of the inner ear.J Biol Chem. 2004;279:46065–46072.
19. Jagnandan D, Church JE, Banfi B, Stuehr DJ, Marrero MB, Fulton DJNovel mechanism of activation of NADPH oxidase 5(NOX5): Calcium-sensitization via phosphorylation. J Biol Chem. 2007;282:6494–6507.
20. Petry A, Djordjevic T, Weitnauer M, Kietzmann T, Hess J, Gorlach A.NOX2 and NOX4 mediate proliferative response in endothelial cells.Antioxid Redox Signal. 2006;8:1473–1484.
21. Belaiba RS, Djordjevic T, Petry A, Diemer K, Bonello S, Banfi B, HessJ, Pogrebniak A, Bickel C, Gorlach A. NOX5 variants are functionallyactive in endothelial cells. Free Radic Biol Med. 2007;42:446–459.
22. Bayraktutan U, Blayney L, Shah AM. Molecular characterization andlocalization of the NAD(P)H oxidase components gp91-phox andp22-phox in endothelial cells. Arterioscler Thromb Vasc Biol. 2000;20:1903–1911.
23. Zhang Q, Church JE, Jagnandan D, Catravas JD, Sessa WC, Fulton D.Functional relevance of Golgi- and plasma membrane-localized endothe-lial NO synthase in reconstituted endothelial cells. Arterioscler ThrombVasc Biol. 2006;26:1015–1021.
24. Scotland RS, Morales-Ruiz M, Chen Y, Yu J, Rudic RD, Fulton D,Gratton JP, Sessa WC. Functional reconstitution of endothelial nitricoxide synthase reveals the importance of serine 1179 in endothelium-dependent vasomotion. Circ Res. 2002;90:904–910.
25. Fulton D, Babbitt R, Zoellner S, Fontana J, Acevedo L, McCabe TJ,Iwakiri Y, Sessa WC. Targeting of endothelial nitric-oxide synthase to thecytoplasmic face of the Golgi complex or plasma membrane regulates
Akt- versus calcium-dependent mechanisms for nitric oxide release.J Biol Chem. 2004;279:30349–30357.
26. Romanko OP, Stepp DW. Reduced constrictor reactivity balancesimpaired vasodilation in the mesenteric circulation of the obese Zuckerrat. Am J Physiol Heart Circ Physiol. 2005;289:H2097–H2102.
27. Papaharalambus CA, Griendling KK. Basic mechanisms of oxidativestress and reactive oxygen species in cardiovascular injury. Trends Car-diovasc Med. 2007;17:48–54.
28. Tesfamariam B, Cohen RA. Free radicals mediate endothelial cell dys-function caused by elevated glucose. Am J Physiol. 1992;263:H321–H326.
29. Jung O, Schreiber JG, Geiger H, Pedrazzini T, Busse R, Brandes RP.gp91phox-containing NADPH oxidase mediates endothelial dysfunctionin renovascular hypertension. Circulation. 2004;109:1795–1801.
30. Landmesser U, Dikalov S, Price SR, McCann L, Fukai T, Holland SM,Mitch WE, Harrison DG. Oxidation of tetrahydrobiopterin leads touncoupling of endothelial cell nitric oxide synthase in hypertension.J Clin Invest. 2003;111:1201–1209.
31. Rubanyi GM, Vanhoutte PM. Oxygen-derived free radicals, endothelium,and responsiveness of vascular smooth muscle. Am J Physiol. 1986;250:H815–H821.
32. Shah KA, Samson SE, Grover AK. Effects of peroxide on endothelialnitric oxide synthase in coronary arteries. Mol Cell Biochem. 1998;183:147–152.
33. Zou MH, Shi C, Cohen RA. Oxidation of the zinc-thiolate complex anduncoupling of endothelial nitric oxide synthase by peroxynitrite. J ClinInvest. 2002;109:817–826.
34. Thomas DD, Ridnour LA, Espey MG, Donzelli S, Ambs S, Hussain SP,Harris CC, DeGraff W, Roberts DD, Mitchell JB, Wink DA. Superoxidefluxes limit nitric oxide-induced signaling. J Biol Chem. 2006;281:25984–25993.
35. Channon KM. Tetrahydrobiopterin: regulator of endothelial nitric oxidesynthase in vascular disease. Trends Cardiovasc Med. 2004;14:323–327.
36. Stepp DW, Ou J, Ackerman AW, Welak S, Klick D, Pritchard KA Jr.Native LDL and minimally oxidized LDL differentially regulatesuperoxide anion in vascular endothelium in situ. Am J Physiol HeartCirc Physiol. 2002;283:H750–H759.
37. Lewis RS, Deen WM. Kinetics of the reaction of nitric oxide with oxygenin aqueous solutions. Chem Res Toxicol. 1994;7:568–574.
38. Minor RL Jr, Myers PR, Guerra R Jr, Bates JN, Harrison DG. Diet-induced atherosclerosis increases the release of nitrogen oxides fromrabbit aorta. J Clin Invest. 1990;86:2109–2116.
39. Pagano PJ, Griswold MC, Najibi S, Marklund SL, Cohen RA. Resistanceof endothelium-dependent relaxation to elevation of O(-)(2) levels inrabbit carotid artery. Am J Physiol. 1999;277:H2109–2114.
40. Cosentino F, Hishikawa K, Katusic ZS, Luscher TF. High glucoseincreases nitric oxide synthase expression and superoxide anion gen-eration in human aortic endothelial cells. Circulation. 1997;96:25–28.
41. Sessa WC. eNOS at a glance. J Cell Sci. 2004;117:2427–2429.42. Oess S, Icking A, Fulton D, Govers R, Muller-Esterl W. Subcellular
targeting and trafficking of nitric oxide synthases. Biochem J. 2006;396:401–409.
43. Xu H, Shi Y, Wang J, Jones D, Weilrauch D, Ying R, Wakim B, PritchardKA Jr. A heat shock protein 90 binding domain in endothelial nitric-oxidesynthase influences enzyme function. J Biol Chem. 2007;282:37567–37574.
Zhang et al Interaction of NADPH Oxidase With eNOS 1633
by guest on November 12, 2014http://atvb.ahajournals.org/Downloaded from

Figure Legends for Supplemental data Supplemental Figure 1. Effect of inhibitors on Nox5-induced
chemiluminescence. BAEC were transduced with Nox5 adenovirus (100MOI).
36hrs post transduction, ROS production was monitored by chemiluminescence
in the presence and absence of (A) SOD (100U/ml), (B) L-NAME (1mM), (C)
Allopurinol (1mM) and (D) Catalase (200U/ml). The data are presented as
mean±SE (n=5). *P<0.05 vs control.
Supplemental Figure 2. Nox1 increases eNOS activity. COS-7 cells were
transfected with cDNAs encoding eNOS with or without RFP(control) and
Nox1/NoxO1, NoxA1. Basal NO release was measured using NO-specific
chemiluminescence and the data were presented as mean ± SE (n=5). * P < 0.05 vs
control.
Supplemental Figure 3. Extracellular superoxide does not increase eNOS
activity. COS-7 cells were transfected in 10cm dishes with cDNAs encoding
eNOS, RFP, Nox5, or eNOS together with Nox5 at ratios of (1000:100, 1000:300
and 1000:1000). Where needed, RFP was used a control to balance total DNA
transfected. 24hrs after transfection, the cells were split and replated into 12well
plates for NO release. Cells transfected individually with eNOS, RFP or Nox5
were combined to maintain equivalent eNOS expression. Basal NO release was
measured using NO-specific chemiluminescence and the data were presented as mean
± SE (n=6). * P < 0.05 vs control.
0
2000
4000
6000
8000
0
5000
10000
15000
20000
*0
2000
4000
6000
8000
0
1000
3000
5000
7000
Rel
ativ
e Li
ght U
nits
Rel
ativ
e Li
ght U
nits
Rel
ativ
e Li
ght U
nits
Rel
ativ
e Li
ght U
nits
Control SOD Control L-NAME
AllopurinolControl Control Catalase
A B
C D
Supplemental Figure 1
0
500
1000
1500
2000
2500
3000
eNOSeNOS+Nox1
*N
O- a
ccum
ulat
ion
(pm
ol/w
ell)
2
IB:eNOS
IB:GAPDH
*
Supplemental Figure 2
eNOS + RFP eNOS +NOX5(300)eNOS/NOX5(100)
eNOS/NOX5(300)
eNOS/NOX5(1000)
IB: eNOS
IB: HA-Nox5
eNOS NOX5 eNOS+NOX5
Nox5eNOS
eNOS
Nox5
0
1000
2000
3000
4000
A
B
NO
- acc
um
ula
tio
n(p
mo
l/w
ell)
2
*
*
*
Supplemental Figure 3