important role of nox4 type nadph oxidase in angiogenic responses in human microvascular endothelial...
TRANSCRIPT

ISSN: 1524-4636 Copyright © 2007 American Heart Association. All rights reserved. Print ISSN: 1079-5642. Online
7272 Greenville Avenue, Dallas, TX 72514Arteriosclerosis, Thrombosis, and Vascular Biology is published by the American Heart Association.
DOI: 10.1161/ATVBAHA.107.149450 published online Aug 23, 2007; Arterioscler Thromb Vasc Biol
Srinivasa Raju Datla, Hitesh Peshavariya, Gregory J. Dusting and Fan Jiang Human Microvascular Endothelial Cells In Vitro
Important Role of Nox4 Type NADPH Oxidase in Angiogenic Responses in
http://atvb.ahajournals.org/cgi/content/full/ATVBAHA.107.149450/DC1Data Supplement (unedited) at:
http://atvb.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://atvb.ahajournals.org/subscriptions/Biology is online at Subscriptions: Information about subscribing to Arteriosclerosis, Thrombosis, and Vascular
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

Important Role of Nox4 Type NADPH Oxidase inAngiogenic Responses in Human Microvascular
Endothelial Cells In VitroSrinivasa Raju Datla, Hitesh Peshavariya, Gregory J. Dusting, Fan Jiang
Objective—Redox signaling mediated by Nox2-containing NADPH oxidase has been implicated in angiogenic responsesboth in vitro and in vivo. Because Nox4 type NADPH oxidase is also highly expressed in endothelial cells, we studiedthe role of Nox4 in angiogenic responses in human endothelial cells in culture.
Methods and Results—Inhibition of Nox4 expression by small interfering RNA reduced angiogenic responses as assessedby the tube formation and wound healing assays, in both human microvascular and umbilical vein endothelial cells.Overexpression of wild-type Nox4 enhanced, whereas expression of a dominant negative form of Nox4 suppressed theangiogenic responses in endothelial cells. These effects were mimicked by exogenous H2O2 and the antioxidantcompound ebselen, respectively. Overexpression of Nox4 enhanced receptor tyrosine kinase phosphorylation and theactivation of extracellular signal-regulated kinase (Erk). Inhibition of the Erk pathway reduced the endothelialangiogenic responses. Nox4 expression also promotes proliferation and migration of endothelial cells, and reducedserum deprivation–induced apoptosis.
Conclusions—Nox4 type NADPH oxidase promotes endothelial angiogenic responses, at least partly, via enhancedactivation of receptor tyrosine kinases and the downstream Erk pathway. (Arterioscler Thromb Vasc Biol. 2007;27:000-000.)
Key Words: angiogenesis � endothelial cell � NADPH oxidase � Nox4 � redox signaling
Angiogenesis is a fundamental developmental and adultphysiological process, requiring the coordinated action
of a variety of growth factors and cell-adhesion molecules inendothelial and mural cells. Understanding the molecular andcellular mechanisms of angiogenesis has profound clinicalimplications. Inhibiting angiogenesis is a promising strategyfor treatment of cancer and several other disorders, includingage-related macular degeneration, whereas therapeutic angio-genesis (promoting new vessel growth to treat ischemicdisorders) is an exciting frontier of cardiovascular medicine.1
In both cases, a thorough understanding of the signalingpathways involved in angiogenic process is needed.
Vascular endothelial growth factor (VEGF) is one of themost important angiogenic stimuli of angiogenic responses invascular endothelial cells.2 It is shown that VEGF stimulatesgeneration of reactive oxygen species (ROS) in endothelialcells via activation of the reduced �-nicotinamide adeninedinucleotide phosphate (NADPH) oxidase,3 which is in-volved in modulating the angiogenic activities of endothelialcells.4 Endogenously generated ROS are known to serve assecond messengers activating multiple intracellular signalingpathways that have key roles in endothelial cell biology.5
There is evidence that the Nox2 type NADPH oxidase has an
important role in mediating angiogenic responses both invitro6 and in vivo.7 In Nox2-deficient animals, both VEGF-and tissue ischemia-induced new blood vessel formation isblunted,6,7 indicating that Nox2-dependent redox signaling isimportant in modulating angiogenesis.
We have previously shown that Nox4 is important for ROSgeneration in human endothelial cells,8 which is consistentwith the findings by others.9,10 Moreover, there is evidencethat Nox4 is involved in modulating endothelial cell prolif-eration.11 Together with the finding that the level of Nox4expression in endothelial cells in vivo is high as comparedwith other Nox isoforms,12 these data strongly suggest thatNox4 also has a critical role in mediating endothelial redoxsignaling and is functionally important in modulating endo-thelial cell physiology. In this study we examined the role ofNox4 in angiogenic responses in human endothelial cells invitro.
MethodsAn expanded Methods section has been published online (please seehttp://atvb.ahajournals.org).
Cell CultureHuman microvascular endothelial cells (HMECs) are a gift fromProfessor Philip Hogg (University of New South Wales, Sydney,
Original received February 12, 2007; final version accepted August 13, 2007.From the Bernard O’Brien Institute of Microsurgery, University of Melbourne, Victoria, Australia.Correspondence to Fan Jiang, Bernard O’Brien Institute of Microsurgery, 42 Fitzroy Street, Fitzroy, Victoria 3065, Australia. E-mail fjiang@
unimelb.edu.au© 2007 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol. is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.107.149450
1 by on May 18, 2011 atvb.ahajournals.orgDownloaded from

Australia). Human umbilical vein endothelial cells (HUVECs) wereobtained from ATCC. Cells were cultured in EGM-2 BulletKit(Cambrex Corporation) containing 5% FCS in a CO2/O2 incubator at37°C.
siRNA TransfectionTransfection was performed as described previously.8 The SilencerPredesigned small interfering RNA (siRNA) sequence targeting theexon 2 of human Nox4 was obtained from Ambion (Catalogue No.AM16706, sequence ID #118807). The Negative Control siRNA #1(Catalogue No. AM4611, Ambion) was used as control.
Western BlotWestern blot analysis was performed as described previously.8
Cell Migration AssayCell migration was examined in a modified Boyden chamber system(Neuro Probe Inc) as described previously.13
Preparation of Adenoviral VectorsThe replication-deficient adenoviral vectors encoding�-galactosidase (Ad-LacZ), human wild-type Nox4 (Ad-Nox4WT), ora truncated form of Nox4 lacking the NADPH binding domain(Ad-Nox4�NADPH) were kindly provided by Dr Barry Goldstein(Thomas Jefferson University, Philadelphia, Pa).14 Cell transductionwas performed by incubating the cells with the virus preparation for48 hour in the absence (for Ad-Nox4WT) or presence (for Ad-Nox4�NADPH) of 5% serum.
Real-Time Polymerase Chain ReactionQuantitative real-time polymerase chain reaction (PCR) analysis wasperformed as described previously.8
ApoptosisEndothelial cell apoptosis was measured by caspase 3/7 activationusing the Caspase-Glo 3/7 Luminescent Assay kit (Promega) as permanufacturer’s instruction.
ROS MeasurementIntracellular ROS generation was measured by 2�,7�-dichlorofluorescin diacetate (DCFH-DA) fluorescence as describedpreviously.15
Data and Statistical AnalysisData are expressed as mean�SEM. The mean data were analyzedwith Student t test or 1-way ANOVA followed by Newman-Keuls ttest as appropriate. A value of P�0.05 was regarded as statisticallysignificant.
ResultsNox4 Is Important for ROS Generation inEndothelial CellsTo demonstrate that the Nox4 subunit is important in cata-lyzing ROS generation in endothelial cells, we used Nox4siRNA-mediated gene silencing to modulate Nox4 expres-sion. In HMECs, Nox4 siRNA (100 nmol/L) significantlyreduced the expression of Nox4, whereas Nox2 was un-changed (Figure 1a and the supplemental Figure Ia), which isconsistent with our previous results.8 The Nox4 siRNAsignificantly reduced the ROS production from HMECs byabout 40% (supplemental Figure Ic).
To further explore the role of Nox4 in endothelial ROSgeneration, we used adenoviral vector-mediated gene trans-fer. To determine the efficiency of cell transduction, weinfected the HMECs with different concentrations of Ad-LacZ for 48 hours, and the transgene expression was exam-ined by X-gal staining. We found that at a concentration of3�105 pfu/mL, � 95% of the cells were stained positive(blue) (supplemental Figure II). Using the same viral concen-tration, we then examined the effects of wild-type Nox4(AdNox4WT) overexpression. We found that AdNox4WT treat-ment significantly increased the Nox4 protein level, withoutany effect on Nox2 expression (Figure 1b and supplementalFigure Ib). Wild-type Nox4 overexpression significantlyincreased endothelial ROS production (supplemental FigureId). Moreover, expression of a dominant negative form ofNox4 (Ad-Nox4�NADPH)14 significantly reduced ROS produc-tion in HMECs (supplemental Figure Id). However, the Nox4antibody used in our study does not react with the truncatedform of Nox4, thus it is difficult to directly analyze theexpression level of the dominant negative Nox4 protein.
Angiogenic Responses Were Inhibited byNox4 Gene SilencingTo clarify the role of Nox4 in angiogenic responses ofendothelial cells, we examined the effects of Nox4 genesilencing on the tube formation and wound healing responsesin HMECs. As shown in Figure 2a and 2b, knocking down ofNox4 expression significantly reduced both of the tubeformation and wound healing responses. To further confirmthese findings, we repeated the experiments in HUVECs and
Figure 1. Western blots (n�3 to 4) show-ing that Nox4 expression in HMECs wasmodulated by (a) Nox4 siRNA and (b)Ad-Nox4WT. The level of Nox2 wasunchanged by these treatments. Thedensitometry data are shown in supple-mental Figure I.
2 Arterioscler Thromb Vasc Biol. November 2007
by on May 18, 2011 atvb.ahajournals.orgDownloaded from
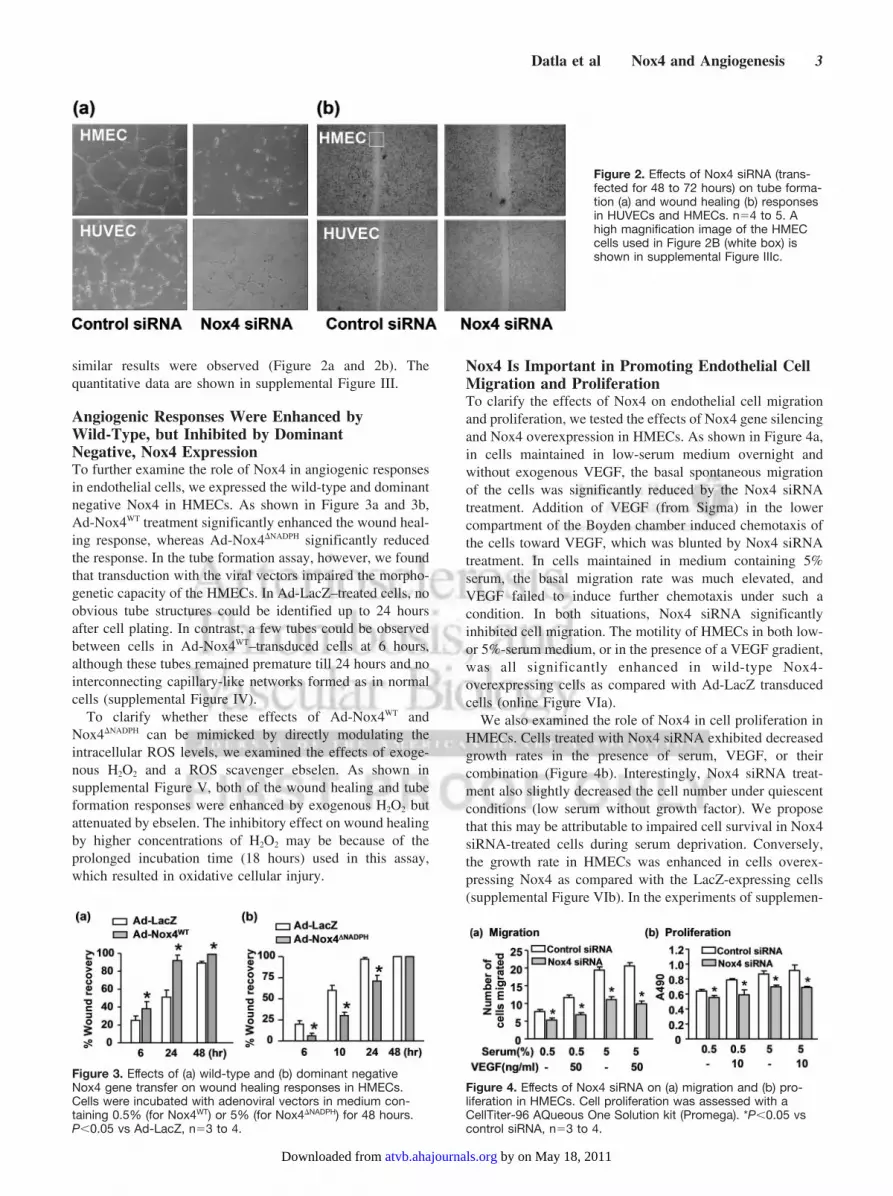
similar results were observed (Figure 2a and 2b). Thequantitative data are shown in supplemental Figure III.
Angiogenic Responses Were Enhanced byWild-Type, but Inhibited by DominantNegative, Nox4 ExpressionTo further examine the role of Nox4 in angiogenic responsesin endothelial cells, we expressed the wild-type and dominantnegative Nox4 in HMECs. As shown in Figure 3a and 3b,Ad-Nox4WT treatment significantly enhanced the wound heal-ing response, whereas Ad-Nox4�NADPH significantly reducedthe response. In the tube formation assay, however, we foundthat transduction with the viral vectors impaired the morpho-genetic capacity of the HMECs. In Ad-LacZ–treated cells, noobvious tube structures could be identified up to 24 hoursafter cell plating. In contrast, a few tubes could be observedbetween cells in Ad-Nox4WT–transduced cells at 6 hours,although these tubes remained premature till 24 hours and nointerconnecting capillary-like networks formed as in normalcells (supplemental Figure IV).
To clarify whether these effects of Ad-Nox4WT andNox4�NADPH can be mimicked by directly modulating theintracellular ROS levels, we examined the effects of exoge-nous H2O2 and a ROS scavenger ebselen. As shown insupplemental Figure V, both of the wound healing and tubeformation responses were enhanced by exogenous H2O2 butattenuated by ebselen. The inhibitory effect on wound healingby higher concentrations of H2O2 may be because of theprolonged incubation time (18 hours) used in this assay,which resulted in oxidative cellular injury.
Nox4 Is Important in Promoting Endothelial CellMigration and ProliferationTo clarify the effects of Nox4 on endothelial cell migrationand proliferation, we tested the effects of Nox4 gene silencingand Nox4 overexpression in HMECs. As shown in Figure 4a,in cells maintained in low-serum medium overnight andwithout exogenous VEGF, the basal spontaneous migrationof the cells was significantly reduced by the Nox4 siRNAtreatment. Addition of VEGF (from Sigma) in the lowercompartment of the Boyden chamber induced chemotaxis ofthe cells toward VEGF, which was blunted by Nox4 siRNAtreatment. In cells maintained in medium containing 5%serum, the basal migration rate was much elevated, andVEGF failed to induce further chemotaxis under such acondition. In both situations, Nox4 siRNA significantlyinhibited cell migration. The motility of HMECs in both low-or 5%-serum medium, or in the presence of a VEGF gradient,was all significantly enhanced in wild-type Nox4-overexpressing cells as compared with Ad-LacZ transducedcells (online Figure VIa).
We also examined the role of Nox4 in cell proliferation inHMECs. Cells treated with Nox4 siRNA exhibited decreasedgrowth rates in the presence of serum, VEGF, or theircombination (Figure 4b). Interestingly, Nox4 siRNA treat-ment also slightly decreased the cell number under quiescentconditions (low serum without growth factor). We proposethat this may be attributable to impaired cell survival in Nox4siRNA-treated cells during serum deprivation. Conversely,the growth rate in HMECs was enhanced in cells overex-pressing Nox4 as compared with the LacZ-expressing cells(supplemental Figure VIb). In the experiments of supplemen-
Figure 2. Effects of Nox4 siRNA (trans-fected for 48 to 72 hours) on tube forma-tion (a) and wound healing (b) responsesin HUVECs and HMECs. n�4 to 5. Ahigh magnification image of the HMECcells used in Figure 2B (white box) isshown in supplemental Figure IIIc.
Figure 3. Effects of (a) wild-type and (b) dominant negativeNox4 gene transfer on wound healing responses in HMECs.Cells were incubated with adenoviral vectors in medium con-taining 0.5% (for Nox4WT) or 5% (for Nox4�NADPH) for 48 hours.P�0.05 vs Ad-LacZ, n�3 to 4.
Figure 4. Effects of Nox4 siRNA on (a) migration and (b) pro-liferation in HMECs. Cell proliferation was assessed with aCellTiter-96 AQueous One Solution kit (Promega). *P�0.05 vscontrol siRNA, n�3 to 4.
Datla et al Nox4 and Angiogenesis 3
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

tal Figure VIb, the lack of stimulatory effects of VEGF andserum on cell growth was likely because of the prolongedserum starvation (48 hours) during viral vector infection.
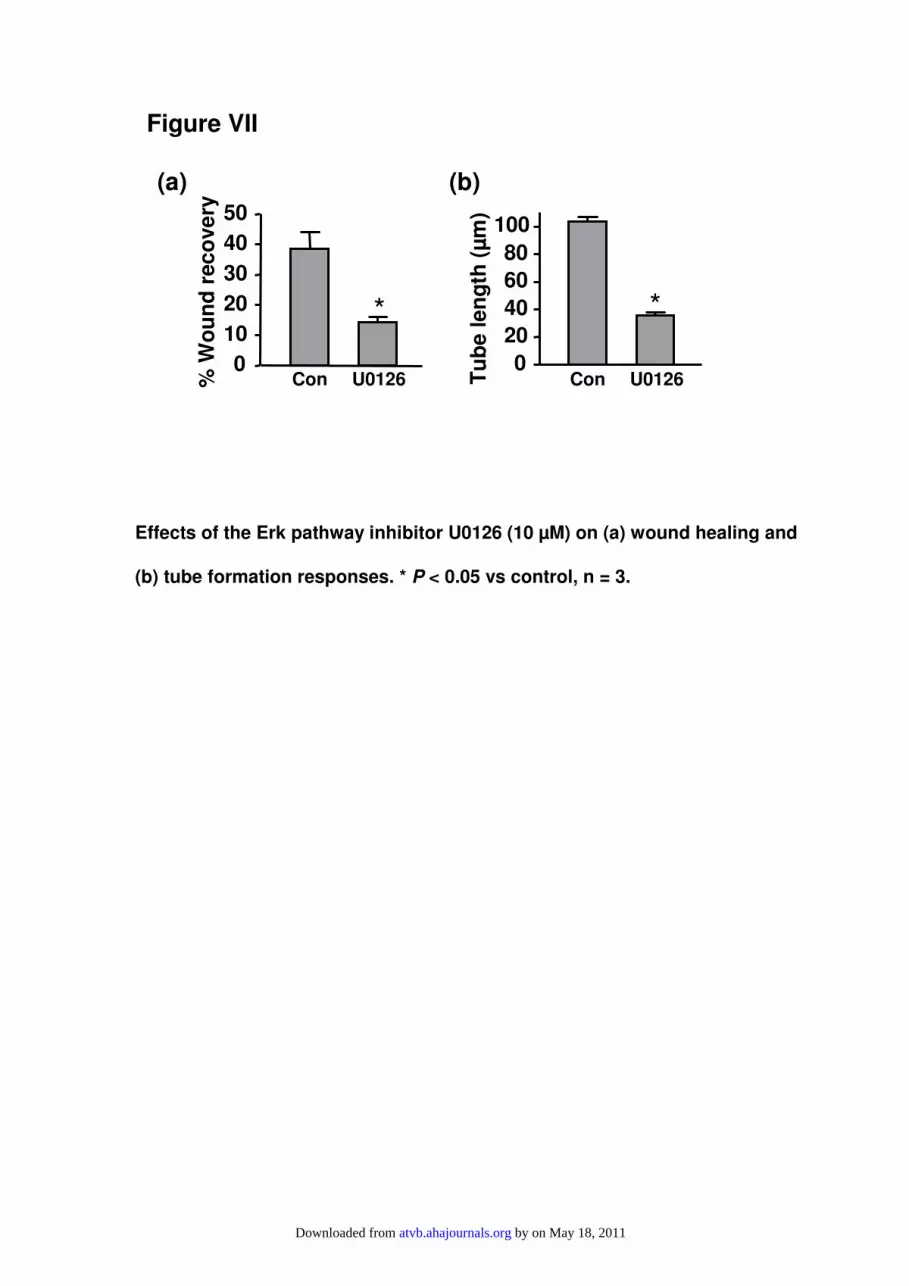
Nox4 Modulates Receptor TyrosineKinase-Mediated SignalingThere is evidence that, in adipocytes, Nox4-dependent ROSproduction may modulate intracellular signaling by inactivat-ing protein tyrosine phosphatases (PTPs),14 resulting in en-hanced tyrosine phosphorylation events. To elucidate whetherthis mechanism is also involved in endothelial cells, we firstlyexamined whether Nox4 expression affects receptor tyrosinekinase activation. We analyzed VEGF receptor-2 (VEGFR-2/KDR) phosphorylation. However, Western blot could notdetect VEGFR-2 in HMECs. Unexpectedly, we found that theplatelet-derived growth factor (PDGF) receptor-� could bereadily detected. Using a phospho-specific antibody (fromSigma), we found that the level of phosphorylation ofPDGFR-� was enhanced in Ad-Nox4WT-transduced cells, butdecreased in Ad-Nox4�NADPH-transduced cells (Figure 5a).Interestingly, these PDGF receptors were phosphorylatedwithout PDGF stimulation under the current experimentalconditions, whereas PDGF did not further increase thephosphorylation status of the receptors (data not shown). Tofurther confirm that Nox4 is important in modulating growthfactor–induced downstream signaling, we analyzed the ef-fects of Nox4 overexpression on Erk1/2 activation. As shownin Figure 5b, VEGF stimulated Erk1/2 phosphorylation,which peaked at 5 to 10 minutes and remained elevated up to20 minutes. Nox4 overexpression increased the basal level ofErk1/2 phosphorylation, and enhanced that after VEGF stim-ulation at all time points examined. To clarify whether Nox4may modulate endothelial angiogenic responses via the Erkpathway, we treated the cells with the MEK (MAPK/ERKkinase) inhibitor U0126. As shown in supplemental FigureVII, U0126 significantly reduced the wound healing and tubeformation responses, indicating that the Erk pathway isdirectly involved in endothelial angiogenic responses. This isconsistent with the findings by other groups.16,17 Finally, weexamined whether VEGF affects Nox4 expression. Real-timePCR experiments demonstrated that VEGF did not signifi-cantly change Nox4 mRNA levels from 2 to 24 hours(supplemental Figure VIII).
The Effects of Nox4 on Angiogenesis IsIndependent of Nitric Oxide FunctionsIt is known that endothelial nitric oxide (NO) has an impor-tant role in modulating angiogenesis.18 To clarify whether theeffects of Nox4 on angiogenic responses in endothelial cellsinvolve modulation of the endothelial NO function, weexamined the endothelial nitric oxide synthase (eNOS) ex-pression and NO availability in HMECs. As compared withprimary HUVECs, the expression level of eNOS in themicrovascular endothelial cells is low (supplemental FigureIXa). Then we measured the intracellular NO level with theNO-sensitive probe 4-amino-5-methylamino-2�,7�-difluorofluorescein diacetate (DAF-FM, Invitrogen) and fluo-rescent microscopy. As shown in supplemental Figure IXb,treatment with the calcium ionophore A23187 failed toinduce DAF-FM fluorescence in HMECs, whereas the NOdonor S-nitroso-N-acetylpenicillamine (SNAP) increased theintracellular fluorescence above the background. Althoughwe found that in wild-type Nox4-expressing cells, the NOlevel after loading with SNAP was significantly lower that inAd-LacZ–treated cells (supplemental Figure IXc), in separateexperiments we observed that treatment with the NOS inhib-itor NG-nitro-L-arginine methyl ester (L-NAME) did notchange the wound healing response (supplemental FigureIXd). Together with the low eNOS expression in HMECs,these results indicate that the endogenous NO is unlikely tohave a major role in modulating angiogenesis in these cells.However, we could not exclude the involvement of NO inother endothelial cells that express higher amounts of eNOS.
Nox4 Is AntiapoptoticFinally, we studied how Nox4 modulates endothelial cellapoptosis in HMECs. Apoptosis was induced by serumdeprivation for 48 hours. Overexpression of wild-type Nox4inhibited serum deprivation-induced activation of Caspase3/7 by about 40% (supplemental Figure X). The NOSinhibitor L-NAME did not modify the apoptotic response toserum deprivation (data not shown), suggesting that per-oxynitrite formation by NO and superoxide is unlikely to beinvolved.
DiscussionPrevious studies have shown that the Nox4 type NADPHoxidase is abundantly expressed in endothelial cells,10,12 andthis subunit is important in mediating NADPH oxidase-dependent superoxide generation in these cells because Nox4gene silencing induced by either antisense oligonucleotide10
or siRNA8 suppressed endothelial superoxide release. Ourprevious study has demonstrated that in vascular smoothmuscle cells, Nox4 expression is increased after cytokinestimulation and is responsible for the resultant increase inROS production, indicating that Nox4 may be involved inmediating vascular oxidative stress under inflammatory con-ditions.19 However, the role of constitutively expressed Nox4under physiological conditions in vascular endothelial cells,remains to be defined. In this study, we provided evidencethat Nox4 has an important role in modulating angiogenicresponses of human endothelial cells in culture. To assess theangiogenic activity of endothelial cells in vitro, we carried out
Figure 5. Effects of wild-type and dominant negative Nox4 on(a) tyrosine phosphorylation of PDGFR-� (*P�0.05 vs Ad-LacZ,n�4), and (b) Erk1/2 phosphorylation under basal conditionsand after VEGF (10 ng/mL) stimulation (example from 2 inde-pendent experiments).
4 Arterioscler Thromb Vasc Biol. November 2007
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

a variety of assays, including proliferation, migration, tubeformation, and wound healing assays. In all these experi-ments, we found that the angiogenic responses were consis-tently inhibited by Nox4 siRNA or overexpression of adominant negative Nox4. Moreover, overexpression of thewild-type Nox4 enhanced the angiogenic responses in endo-thelial cells, and these effects were mimicked by exogenousH2O2. The observation that inhibition of Nox4 expressionsuppressed endothelial cell proliferation is consistent with theresults reported recently.11 All these data suggest that Nox4 isan important modulator of the regenerative function ofendothelial cells, and Nox4 expression in these cells isrequired to support an efficient angiogenic process. Our studywarrants further investigation of the role of Nox4 in angio-genesis in vivo.
Our results complement the previous finding that the Nox2NADPH oxidase is essential for mediating angiogenic re-sponses both in vitro and in vivo.6,7 In endothelial cells, bothNox2 and Nox4 are involved in endothelial ROS genera-tion.10,20 Moreover, inhibition of the expression of eitherNox2 or Nox4 suppressed endothelial cell proliferation sim-ilarly.11 Several groups have studied the intracellular local-ization of Nox2 and Nox4 in endothelial cells and haveunequivocally shown that these Nox subunits have a similarpattern of intracellular distribution, predominantly existing inthe cytoplasm in association with the endoplasmic reticulumor cytoskeleton,11,21–23 in contrast to the membrane localiza-tion of the neutrophil Nox2. All these results indicate thatboth Nox2 and Nox4 are equivalently important in modulat-ing endothelial cell functions. Whether they have differentroles in endothelial cell biology is unclear, although there isevidence that in endothelial cells, the mRNA transcripts ofNox4 are expressed more highly than that of Nox2.10,23 Giventhe results by Ushio-Fukai et al,6 our study suggests thatNox2 and Nox4 have nonredundant effects on modulatingangiogenic process, as separate inhibition of each isoformsimilarly decreased the angiogenic activities in ECs. There-fore it would be interesting to study whether simultaneousinhibition of both isoforms has synergistic effects on theendothelial angiogenic response.
The involvement of Nox4-dependent redox mechanisms inregulating intracellular signaling and cellular functions hasbeen studied in nonendothelial cells. For example, in insulin-sensitive adipocytes, expression of the dominant negativeNox4 attenuated insulin-stimulated ROS generation, insulinreceptor tyrosine phosphorylation, activation of downstreamserine kinases, and glucose uptake, indicating that Nox4 isinvolved in promoting insulin-induced signaling by intracel-lular ROS generation.14 One well established target of intra-cellular ROS-mediated signaling is the thiol-containing PTPs,which terminate growth factor-induced signals by dephos-phorylation of the receptor tyrosine kinases. Inactivation ofPTPs may prolong the activation of the receptor tyrosinekinases and thereby enhance the downstream signaling cas-cades.24 Consistent with this notion, we demonstrated thatoverexpression of wild-type Nox4 enhanced receptor tyrosinekinase phosphorylation and enhanced the activation of Erk.To our knowledge, this is the first evidence that Nox4 isinvolved in modulating receptor tyrosine kinase activation in
endothelial cells. Our results support the idea that Nox4 hasprimary physiological roles in maintaining normal cell sig-naling and functions, although exaggerated expression of thissubunit may be involved in cellular oxidant stress.
We also found that Nox4 overexpression prevents endo-thelial cell apoptosis. This observation is consistent withprevious reports in pancreatic cancer cells that ROS producedby Nox4 NADPH oxidase inhibited apoptosis, whereas inhi-bition of Nox4 expression activated apoptosis in thesecells.25,26 The mechanisms by which Nox4 produces antiapo-ptotic effects are not totally understood. In pancreatic cancercells, it has been shown that Nox4 gene silencing or blockadeof NADPH oxidase by diphenyleneiodonium suppressedphosphorylation and activation of the cell survival kinaseAkt, suggesting that Nox4-induced cell survival involves thephosphatidylinositol 3-kinase (PI3-kinase)/Akt pathway. Thisargument is further supported by a recent study in the mouseheart.27 Whether this pathway is also responsible for theantiapoptotic actions of Nox4 in endothelial cells has notbeen examined. Nonetheless, our observation that Nox4expression enhanced VEGF-induced signaling supports thisnotion, because PI3-kinase/Akt is downstream of receptortyrosine kinase activation. On the other hand, it is known thathigh levels of ROS may induce cell death. Therefore theregulatory effects of Nox4 on cell apoptosis and survival mayvary under different conditions, for there is evidence thatNox4 is involved in promoting apoptosis of human aorticsmooth muscle cells during endoplasmic reticulum stressinduced by oxidized lipids.28
In summary, our results suggest that the Nox4 typeNADPH oxidase-dependent redox signaling is an importantpositive modulator of angiogenic responses in cultured endo-thelial cells, and this effect is, at least partly, mediated byenhanced VEGF-induced intracellular signaling.
AcknowledgmentsThe authors thank Nancy Guo for technical assistance with thereal-time PCR.
Sources of FundingThis study is supported by project grants from the AustralianNational Health and Medical Research Council and Grants-in-Aidfrom the National Heart Foundation. G.J.D. is a NHMRC PrincipalResearch Fellow.
DisclosuresNone.
References1. Ferrara N, Kerbel RS. Angiogenesis as a therapeutic target. Nature. 2005;
438:967–974.2. Simons M. Integrative signaling in angiogenesis. Mol Cell Biochem.
2004;264:99–102.3. Griendling KK, Sorescu D, Ushio-Fukai M. NAD(P)H oxidase: role in
cardiovascular biology and disease. Circ Res. 2000;86:494–501.4. Ushio-Fukai M, Alexander RW. Reactive oxygen species as mediators of
angiogenesis signaling: role of NAD(P)H oxidase. Mol Cell Biochem.2004;264:85–97.
5. Kunsch C, Medford RM. Oxidative stress as a regulator of geneexpression in the vasculature. Circ Res. 1999;85:753–766.
6. Ushio-Fukai M, Tang Y, Fukai T, Dikalov SI, Ma Y, Fujimoto M, QuinnMT, Pagano PJ, Johnson C, Alexander RW. Novel role of gp91(phox)-
Datla et al Nox4 and Angiogenesis 5
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

containing NAD(P)H oxidase in vascular endothelial growth factor-induced signaling and angiogenesis. Circ Res. 2002;91:1160–1167.
7. Tojo T, Ushio-Fukai M, Yamaoka-Tojo M, Ikeda S, Patrushev N,Alexander RW. Role of gp91phox (Nox2)-containing NAD(P)H oxidasein angiogenesis in response to hindlimb ischemia. Circulation. 2005;111:2347–2355.
8. Jiang F, Roberts SJ, raju Datla S, Dusting GJ. Nitric oxide modulatesNADPH oxidase function via heme oxygenase-1 in human endothelialcells. Hypertension. 2006;48:950–957.
9. Thum T, Borlak J. Mechanistic role of cytochrome P450 monooxygen-ases in oxidized low-density lipoprotein-induced vascular injury: therapythrough LOX-1 receptor antagonism? Circ Res. 2004;94:e1–e13.
10. Ago T, Kitazono T, Ooboshi H, Iyama T, Han YH, Takada J, WakisakaM, Ibayashi S, Utsumi H, Iida M. Nox4 as the major catalytic componentof an endothelial NAD(P)H oxidase. Circulation. 2004;109:227–233.
11. Petry A, Djordjevic T, Weitnauer M, Kietzmann T, Hess J, Gorlach A.NOX2 and NOX4 mediate proliferative response in endothelial cells.Antioxid Redox Signal. 2006;8:1473–1484.
12. Sorescu D, Weiss D, Lassegue B, Clempus RE, Szocs K, Sorescu GP,Valppu L, Quinn MT, Lambeth JD, Vega JD, Taylor WR, Griendling KK.Superoxide production and expression of nox family proteins in humanatherosclerosis. Circulation. 2002;105:1429–1435.
13. Abid MR, Kachra Z, Spokes KC, Aird WC. NADPH oxidase activity isrequired for endothelial cell proliferation and migration. FEBS Lett.2000;486:252–256.
14. Mahadev K, Motoshima H, Wu X, Ruddy JM, Arnold RS, Cheng G,Lambeth JD, Goldstein BJ. The NAD(P)H oxidase homolog Nox4 mod-ulates insulin-stimulated generation of H2O2 and plays an integral role ininsulin signal transduction. Mol Cell Biol. 2004;24:1844–1854.
15. Peshavariya HM, Dusting GJ, Selemidis S. Analysis of dihydroethidiumfluorescence for the detection of intracellular and extracellular superoxideproduced by NADPH oxidase. Free Radic Res. 2007;41:699–712.
16. Meadows KN, Bryant P, Pumiglia K. Vascular endothelial growth factorinduction of the angiogenic phenotype requires Ras activation. J BiolChem. 2001;276:49289–49298.
17. Bhagwat SV, Petrovic N, Okamoto Y, Shapiro LH. The angiogenicregulator CD13/APN is a transcriptional target of Ras signaling pathwaysin endothelial morphogenesis. Blood. 2003;101:1818–1826.
18. Morbidelli L, Donnini S, Ziche M. Role of nitric oxide in the modulationof angiogenesis. Curr Pharm Des. 2003;9:521–530.
19. Moe KT, Aulia S, Jiang F, Chua YL, Koh TH, Wong MC, Dusting GJ.Differential upregulation of Nox homologues of NADPH oxidase bytumor necrosis factor-alpha in human aortic smooth muscle andembryonic kidney cells. J Cell Mol Med. 2006;10:231–239.
20. Jones SA, O’Donnell VB, Wood JD, Broughton JP, Hughes EJ, JonesOT. Expression of phagocyte NADPH oxidase components in humanendothelial cells. Am J Physiol. 1996;271:H1626–H1634.
21. Bayraktutan U, Blayney L, Shah AM. Molecular characterization andlocalization of the NAD(P)H oxidase components gp91-phox andp22-phox in endothelial cells. Arterioscler Thromb Vasc Biol. 2000;20:1903–1911.
22. Li JM, Shah AM. Intracellular localization and preassembly of theNADPH oxidase complex in cultured endothelial cells. J Biol Chem.2002;277:19952–19960.
23. Van Buul JD, Fernandez-Borja M, Anthony EC, Hordijk PL. Expressionand localization of NOX2 and NOX4 in primary human endothelial cells.Antioxid Redox Signal. 2005;7:308–317.
24. Chiarugi P, Cirri P. Redox regulation of protein tyrosine phosphatasesduring receptor tyrosine kinase signal transduction. Trends Biochem Sci.2003;28:509–514.
25. Mochizuki T, Furuta S, Mitsushita J, Shang WH, Ito M, Yokoo Y,Yamaura M, Ishizone S, Nakayama J, Konagai A, Hirose K, Kiyosawa K,Kamata T. Inhibition of NADPH oxidase 4 activates apoptosis via theAKT/apoptosis signal-regulating kinase 1 pathway in pancreatic cancerPANC-1 cells. Oncogene. 2006;25:3699–3707.
26. Vaquero EC, Edderkaoui M, Pandol SJ, Gukovsky I, Gukovskaya AS.Reactive oxygen species produced by NAD(P)H oxidase inhibit apoptosisin pancreatic cancer cells. J Biol Chem. 2004;279:34643–34654.
27. Chen JX, Zeng H, Tuo QH, Yu H, Meyrick B, Aschner JL. NADPHoxidase modulates myocardial Akt, ERK1/2 activation, and angiogenesisafter hypoxia-reoxygenation. Am J Physiol Heart Circ Physiol. 2007;292:H1664–H1674.
28. Pedruzzi E, Guichard C, Ollivier V, Driss F, Fay M, Prunet C, Marie JC,Pouzet C, Samadi M, Elbim C, O’Dowd Y, Bens M, Vandewalle A,Gougerot-Pocidalo MA, Lizard G, Ogier-Denis E. NAD(P)H oxidaseNox-4 mediates 7-ketocholesterol-induced endoplasmic reticulum stressand apoptosis in human aortic smooth muscle cells. Mol Cell Biol.2004;24:10703–10717.
6 Arterioscler Thromb Vasc Biol. November 2007
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

1
Methods
Cell culture
Human microvascular endothelial cells (HMECs) are a gift from Professor Philip Hogg
(University of New South Wales, Sydney, Australia). Human umbilical vein endothelial cells
(HUVECs) were obtained from ATCC. Cells were cultured in EGM-2 BulletKit (Cambrex
Corporation) containing 5% FCS in a CO2/O2 incubator at 37°C.
siRNA transfection
To knock down the Nox4 expression in endothelial cells, we tested three Silencer® Pre-
designed small interfering RNA (siRNA) sequences obtained from Ambion (Austin, TX,
USA) (Catalogue No AM16706), with the sequence IDs being #117539 (targeting exon 2 of
the human Nox4 gene), #117540 (targeting exon 4), and #118807 (targeting exon 2)
respectively. The Silencer® Negative Control #1 siRNA (Catalogue No. AM4611, Ambion)
was used as control. We found that only the sequence #118807 consistently reduced Nox4
expression in HMEC cells, thus this siRNA was used in the following experiments.
Transfection was performed as described previously.1 Cells were subcultured in 6-well plates
24 hr before transfection in antibiotic-free medium. Control or Nox4 siRNA (100 nmol/L
final concentration) mixed with 5 µL of Lipofectamin 2000 (Invitrogen) in Opti-MEM
Reduced Serum Medium (Invitrogen) was used to transfect each well. Assays were
performed 48 hr after transfection.
Western blot
Western blot analysis was performed as described previously.1 Autophosphorylation
of receptor tyrosine kinases and phosphorylation of the extracellular signal-regulated kinase
(Erk1/2) were analyzed by western blot using phospho-specific antibodies.
Cell proliferation assay
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

2
Cells were seeded in 24-well plates (4 × 104 per well), treated with various conditions
(see Results), and the cell number in each well was determined 48 hr after seeding. A
tetrazolium-based non-radioactive proliferation assay kit (CellTiter 96 AQueous, Promega)
was used to determine the cell number according to the manufacturer’s protocol.
Cell migration assay
Cell migration was examined in a modified Boyden chamber system (Neuro Probe,
Inc., MD, USA) as described previously.2 The top well was loaded with 7500 cells and
VEGF of 50 ng/ml was added in the lower chamber to induce chemotaxis. After 6 hr of
incubation, the cells were fixed and stained with a Quick Dip staining kit (Fronine, NSW,
Australia). The number of cells migrated to the bottom side was counted under a light
microscope.
Tube formation assay
Cells were plated on Matrigel (BD Biosciences) at a density of 5 × 104 cells per well in
a 24-well plate. The capillary-like tube structures were examined using an inverted phase-
contrast microscope (Olympus CK40), and the tube formation index was determined by
measuring the average length of tubes in five random fields from each well using an
Axiovision software.
Wound healing assay
Endothelial cells were cultured in 6-well plates as confluent monolayers. A wound was
made by scratching a line across the monolayer with a pipette tip. The width of the cell-free
gap was measured continuously. The wound healing response was measured by the
percentage recovery of the width of the initial scratch. The edge between the cell monolayer
and the scratch was determined manually by an independent observer in a blind manner in at
least 5 randomly selected fields.
Preparation of adenoviral vectors
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

3
The replication-deficient adenoviral vectors encoding β-galactosidase (Ad-LacZ),
human wild type Nox4 (Ad-Nox4WT), or a truncated form of Nox4 lacking the NADPH
binding domain (Ad-Nox4∆NADPH) were kindly provided by Dr Barry Goldstein (Thomas
Jefferson University, Philadelphia, Pennsylvania, USA).3 The viruses were amplified in
HEK293 cells, and the crude cell lysates were used for cell infection. The titer of the virus
stocks was determined by the agarose overlay plaque assay. Cell transduction was performed
by incubating the cells with the virus preparation for 48 hr in the absence (for Ad-Nox4WT) or
presence (for Ad-Nox4∆NADPH) of 5% serum.
Real-time PCR
Quantitative real-time PCR analysis was performed as described previously.1
Apoptosis
Endothelial cell apoptosis was measured by caspase 3/7 activation using the Caspase-
Glo 3/7 Luminescent Assay kit (Promega) as per manufacture’s instruction. Cells were
grown in 24-well plates until confluent and apoptosis was induced by serum deprivation for
48 hr. Chemiluminescence was measured by a PolarStar microplate reader (BMG
LABTECH, Germany), and the results were normalized with protein concentrations.
ROS measurement
Intracellular ROS generation was measured by 2',7'-dichlorofluorescin diacetate
(DCFH-DA) fluorescence as described previously.4
Data and statistical analysis
Data are expressed as mean ± standard error of the mean (SEM). The mean data were
analyzed with Student’s t-test or one-way analysis of the variance (one-way ANOVA)
followed by Newman-Keuls t-test as appropriate. A value of P < 0.05 was regarded as
statistically significant.
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

4
References
1. Jiang F, Roberts SJ, raju Datla S, Dusting GJ. Nitric oxide modulates NADPH
oxidase function via heme oxygenase-1 in human endothelial cells. Hypertension.
2006;48:950-957.
2. Abid MR, Kachra Z, Spokes KC, Aird WC. NADPH oxidase activity is required for
endothelial cell proliferation and migration. FEBS Lett. 2000;486:252-256.
3. Mahadev K, Motoshima H, Wu X, Ruddy JM, Arnold RS, Cheng G, Lambeth JD,
Goldstein BJ. The NAD(P)H oxidase homolog Nox4 modulates insulin-stimulated
generation of H2O2 and plays an integral role in insulin signal transduction. Mol Cell
Biol. 2004;24:1844-1854.
4. Peshavariya HM, Dusting GJ, Selemidis S. Analysis of dihydroethidium fluorescence
for the detection of intracellular and extracellular superoxide produced by NADPH
oxidase. Free Radic Res. 2007;41:699-712.
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

Figure I
Densitometry analyses of western blots showing that (a) siRNA-induced Nox4
gene silencing reduced expression of Nox4, but not Nox2 (* P < 0.05 vs control, n
= 4); and (b) Ad-Nox4WT increased expression of Nox4 but not Nox2. Ad-LacZ was
used as control (*P < 0.05 vs control, n = 3). (c) effects of Nox4 siRNA on ROS
generation measured by DCFH-DA fluorescence (*P < 0.05 vs control, n = 3). (d)
effects of expression of wild type and the dominant negative Nox4 (∆∆∆∆Nox4) on
ROS generation (*P < 0.05 vs control, n = 3).
fold
of c
ontr
ol
00.20.40.60.81.0
Nox4Con
*
00.250.500.751.00
Nox4Con
Nox4 Nox2
012345
fold
of c
ontr
ol
Nox4
00.250.500.751.001.25
LacZ Nox4 LacZ Nox4
Nox2
*
(c)
Con LacZ ∆∆∆∆Nox4
RO
S (
% C
ontr
ol)
0255075
100125150
*
(d)
RO
S (
% c
ontr
ol)
*
Con siRNA0
20
40
60
80
100
0
50
100
150
LacZ Nox4WT
*
(b)(a)
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

Control 3××××105
Ad-LacZ infection in HMECs for 48 hr
1.5××××1057.5××××104
Ad-LacZ (pfu/ml)
Figure II
The efficiency of adenovirus infection was determined by incubation
with different concentrations of Ad-LacZ for 48 hr followed by X-gal
staining. Positively transduced cells were stained blue. Example of 2
independent experiments.
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

Figure III
Effects of Nox4 siRNA on tube formation (a) and wound healing (b) responses in
HMECs and HUVECs. Cells were transfected 48 – 72 hr before experimentation. The
total length of the tubes formed (observed 6 hr after plating on Matrigel) per high-
power field and the percentage recovery of the scratch (% wound healing) were
measured and averaged in 5 - 6 randomly selected fields. * P < 0.05 vs control (open
bars), n = 4 - 5. (c) A high magnification (100 x) image showing the morphology of the
HMEC cells used in the wound healing assay (Figure 2b). The two white lines outline
the border of the wound.
01020304050
*
Leng
th (µ
m)
01020304050
*
Leng
th (µ
m)
Con Nox4
% r
ecov
ery
0255075
100
**
0255075
100
**% r
ecov
ery
HMEC
HUVEC
6 10 24 hr
HMEC
HUVEC
(c)
(b)(a)
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

Effects of wild type and dominant negative Nox4 gene transfer on tube
formation responses in HMECs. Unlike normal HMEC cells, the
morphogenetic capacity of virus-transduced cells was impaired, as no
obvious capillary network structures could be identified up to 24 hr after
cell plating. A few premature tubes could be observed between cells in Ad-
Nox4WT-transduced cells at 6 hr (arrows), which were absent in Ad-LacZ or
Ad-Nox4∆∆∆∆NADPH treated cells.
Figure IV
Ad-LacZ Ad-Nox4WT Ad-Nox4∆∆∆∆NADPH
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

(a)
0 0.1 0.3 3 10 0
102030405060 * *
*
% W
ound
rec
over
y
H2O2 (µmol/L)
0 100
102030405060
*
ebselen0 3 10
0306090
120150
H2O2 (µmol/L)
**
Tube
leng
th (µ
m)(b)
0306090
120
0 10
ebselen
*
(µmol/L) (µmol/L)
Figure V
Effects of exogenous H2O2 and ebselen on (a) wound healing and (b) tube
formation responses in HMECs. Both compounds were added at the time
of cell plating or wound scratching. * P < 0.05 vs control, n = 3.
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

Ad-LacZAd-Nox4WT
* **
% C
ontr
ol
1050.5 0.5
- -
(b). Proliferation
*
**
Num
ber
ofce
lls m
igra
ted
0
50
100
150
0
5
10
15
Serum(%)50
50.5VEGF(ng/ml)
0.5- -
(a). MigrationAd-LacZAd-Nox4WT
Figure VI
Effects of wild type Nox4 overexpression on (a) migration and (b) proliferation
of HMECs. The migration of cells was measured as either the basal mobility
(0.5% serum) or the chemotaxic response stimulated by VEGF, serum or their
combination as indicated. Cell proliferation under resting condition (0.5%
serum) or after stimulation with VEGF or serum for 48 hr was assessed with a
CellTiter-96 AQueous One Solution kit (Promega). * P < 0.05 vs Ad-LacZ, n = 3 –
4.
by on May 18, 2011 atvb.ahajournals.orgDownloaded from
(a)
Con U01260
1020304050
020406080
100%
Wou
nd r
ecov
ery
Tube
leng
th (µ
m)
Con U0126
(b)
* *
Figure VII
Effects of the Erk pathway inhibitor U0126 (10 µM) on (a) wound healing and
(b) tube formation responses. * P < 0.05 vs control, n = 3.
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

con 2 4 6 16 240
0.5
1.0
1.5
2.0
2.5
3.0
VEGF (hr)
Fold
of c
ontr
olFigure VIII
Effects of VEGF (10 ng/ml) stimulation in serum-starved HMEC
cells on the Nox4 mRNA level measured by real-time PCR (n = 4).
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

A231870
2.5
5.0
7.5
0
10
20
30
02.55.07.5
10.012.515.017.5
Control L-NAME0
102030405060
Figure IX
(a)
(d)(c)
(b)
B/G SNAPB/G
Nox4WTLacZ
% W
ound
hea
ling
*
Inte
nsity
(arb
itrar
y un
it)
∆∆ ∆∆Int
ensi
ty(-
back
grou
nd)
Role of NO in the modulation of angiogenic responses in HMECs. (a)
western blot showing that the eNOS expression is very low in HMECs as
compared to primary HUVECs; (b) A23187 (100 µM) failed to increase
intracellular fluorescence above the background (B/G) in HMECs loaded
with 1 µM of the NO-sensitive probe DAF-FM, whereas the NO donor
SNAP (0.1 µM) increased the fluorescence intensity (averaged data from
quadruplicate measurements) ; (c) overexpression of wild type Nox4
decreased the NO availability in cells loaded with SNAP. * P < 0.05, n =
4; (d) effects of the NOS inhibitor L-NAME (100 µM) on the wound
healing response (n = 4).
HMEC HUVEC
eNOSββββ-actin
by on May 18, 2011 atvb.ahajournals.orgDownloaded from

Figure X
LacZNox4WT
+ Serum
*A
rbitr
ary
Uni
t (x
100
0)
- Serum0
2
4
6
8
10
* †
Effects of wild type Nox4 overexpression on apoptosis in HMECs
induced by serum deprivation for 48 hr. Apoptosis was measured by
Caspase 3/7 activation. * P < 0.05 vs cells without serum deprivation;
† P < 0.05 vs Ad-LacZ, n = 3.
by on May 18, 2011 atvb.ahajournals.orgDownloaded from