overexpression of an adenosine diphosphate-ribosylation factor gene from the halophytic grass...
TRANSCRIPT

ORIGINAL PAPER
Overexpression of an adenosine diphosphate-ribosylation factorgene from the halophytic grass Spartina alterniflora conferssalinity and drought tolerance in transgenic Arabidopsis
Ratna Karan • Prasanta K. Subudhi
Received: 18 September 2013 / Revised: 21 October 2013 / Accepted: 2 November 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract
Key message Isolation, cloning, and expression char-
acterization of an ARF gene from S. alterniflora, dem-
onstrating its involvement in abiotic stress tolerance.
Abstract Adenosine diphosphate-ribosylation factors
(ARFs) are small guanine nucleotide-binding proteins that
play an important role in intracellular protein trafficking
necessary for undertaking multiple physiological functions
in plant growth and developmental processes. However,
little is known about the mechanism of ARF functioning at
the molecular level, as well as its involvement in abiotic
stress tolerance. In this study, we demonstrated the direct
involvement of an ARF gene SaARF from a grass halo-
phyte Spartina alterniflora in abiotic stress adaptation for
the first time. SaARF, which encodes a protein with pre-
dicted molecular mass of 21 kDa, revealed highest identity
with ARF of Oryza sativa. The SaARF gene is transcrip-
tionally regulated by salt, drought, cold, and ABA in the
leaves and roots of S. alterniflora. Arabidopsis plants
overexpressing SaARF showed improved seed germination
and survival of seedlings under salinity stress. Similarly,
SaARF transgenic Arabidopsis plants were more tolerant to
drought stress, compared to wild-type plants, by main-
taining chlorophyll synthesis, increasing osmolyte synthe-
sis, and stabilizing membrane integrity. Oxidative damage
due to moisture stress in transgenic Arabidopsis was also
reduced possibly by activating antioxidant genes, AtSOD1
and AtCAT. Our results suggest that enhanced drought and
salinity tolerance conferred by the SaARF gene may be due
to its role in mediating multiple abiotic stress tolerance
mechanisms.
Keywords Abiotic stress �ADP-ribosylation factor �Gene expression � GTP-binding protein � Halophyte �Transgenic Arabidopsis
Introduction
Adenosine diphosphate (ADP)-ribosylation factors (ARFs)
are small guanosine triphosphate (GTP) binding proteins
belonging to the Ras-like GTPase superfamily, which is
divided into several major subfamilies, including Ras, Rab,
Rho, Ran, Arf, and others (Zerial and Huber 1995). ARF is
a protein with a molecular mass of 21 kDa and was first
identified as a cofactor required for cholera toxin-mediated
ADP ribosylation of a trimeric G protein a-chain from the
cholate extracts of rabbit liver membranes (Kahn and
Gilman 1984). Extensive studies conducted in mammalian
and yeast systems led to the elucidation of the role of ARF
in regulating a diverse array of cellular and physiological
functions, such as the regulation of intracellular membrane
traffic (Balch et al. 1992), the actin cytoskeleton (D’Souza-
Schorey and Chavrier 2006), and organelle structure
(Donaldson and Jackson 2000).
Communicated by J. S. Shin.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00299-013-1537-8) contains supplementarymaterial, which is available to authorized users.
Present Address:
R. Karan
Agronomy Department, Institute of Food and Agricultural
Sciences, University of Florida, Gainesville, FL 32611, USA
P. K. Subudhi (&)
School of Plant, Environmental, and Soil Sciences, Louisiana
State University Agricultural Center, 104 Sturgis Hall, Baton
Rouge, LA 70803, USA
e-mail: [email protected]
123
Plant Cell Rep
DOI 10.1007/s00299-013-1537-8

ARFs are best known as GTP-dependent switches for
the assembly/disassembly of the coat proteins driving
vesicle budding. Guanosine diphosphate (GDP)-bound
ARF is inactive and remains in cytosol, whereas GTP-
bound ARF is activated by specific guanine nucleotide
exchange factor (GEF) and then interacts with downstream
effector proteins to regulate various cellular processes
(Gillingham and Munro 2007). Subsequently, the GTP
bound to ARF is hydrolyzed to GDP by GTPase-activating
protein (GAP), and ARF-GDP is released from the mem-
branes to facilitate docking and fusion of the vesicles with
target membranes (Donaldson and Jackson 2000; Inoue and
Randazzo 2007). GTP-ARF recruits coat proteins from the
cytosol and helps in the budding of specific membrane-
bound receptors and cargo molecules to facilitate vesicle
trafficking (Pimpl et al. 2000; Yahara et al. 2001). ARF
proteins act as regulators of vesicle-mediated protein traf-
ficking and activators of phospholipase D (Jones et al.
1999).
Studies in higher plants have shown that ARFs play a
role in regulating intracellular transport (Ritzenthaler et al.
2002), mitosis, and the cell cycle (McElver et al. 2000).
ARFs are also implicated in many physiological functions,
such as seed development (Tzafrir et al. 2002), auxin
transport (Geldner et al. 2003), epidermal cell polarity (Xu
and Scheres 2005), endocytosis (Naramoto et al. 2010),
flowering, and apical dominance (Gebbie et al. 2005). ARF
genes have been isolated and characterized in many crop
plants (potato—Szopa and Muller-Rober 1994; rice—Higo
et al. 1994; carrot—Asakura et al. 2007; maize—Verwoert
et al. 1995; wheat—Kobayashi-Uehara et al. 2001). The
preferential expression of ARF has been reported in dif-
ferent organs of carrot, wheat, and physic nut (Asakura
et al. 2007; Yao et al. 2009; Qin et al. 2011). Its increased
expression has been observed in postharvest ripening fruit
of banana (Wang et al. 2010), potato tubers (Liu et al.
2012), and early stages of endosperm development in
maize and rice (Liu et al. 2010; Zhou et al. 2010). ARFs
have been also reported to be involved in disease response
(Lee et al. 2003; Lee and Sano 2007; Coemans et al. 2008).
In carrot and physic nut, differential expression of the ARF
gene in response to environmental stresses has been
reported (Asakura et al. 2007; Qin et al. 2011). However,
there is no report till date that demonstrates the direct
involvement of the ARF gene in conferring tolerance to
abiotic stresses.
Salinity and drought are two major abiotic stresses that
pose serious threats to world food security. Plant growth,
development, and productivity are adversely affected due
to dysfunctions in cellular machinery caused by these
disturbances (Knight and Knight 2001). Plants have
developed complex adaptive genetic mechanisms to
respond to these environmental disturbances (Zhu 2002;
Nakashima et al. 2009). Compared with crop plants, which
are vulnerable to abiotic stresses, halophytes adapt well
under extreme environmental conditions due to their
superior stress tolerance mechanisms (Bressan et al. 2013).
A halophyte grass Spartina alterniflora, commonly known
as smooth cordgrass and possessing all known mechanisms
of salt tolerance, has been used as a model to mine useful
salt stress-responsive genes (Subudhi and Baisakh 2011).
Several studies have demonstrated the utility of genes from
this halophyte to improve abiotic stress tolerance (Baisakh
et al. 2012; Karan and Subudhi 2012a, b). In the present
investigation, we characterized ‘SaARF’, an ADP-ribosyl-
ation factor gene from this grass halophyte and studied the
effect of abiotic stress on its transcriptional regulation, as
well as abiotic stress tolerance in Arabidopsis. The possible
role in various abiotic stress tolerance mechanisms is
discussed.
Materials and methods
Spartina alterniflora plants and stress treatment
Young (3- to 4-leaf stage), uniform, clonally propagated
plants of S. alterniflora cv. ‘Vermilion’ were grown in
sand-filled plastic pots under normal growth conditions
inside a greenhouse with 14 h light and 10 h dark at
26/18 �C day/night temperatures and used for stress-related
experiments (Baisakh et al. 2008). Pots were supplied with
Hoagland nutrient solution (Hoagland and Arnon 1950).
Stress treatments were provided as reported earlier by
Karan and Subudhi (2012a). For salinity stress, a 5 %
(w/v) solution of commercial synthetic sea salts (Instant
Ocean, Aquarium Systems, Mentor, OH, USA) dissolved
in Hoagland solution was used. For drought stress, uniform
Spartina plants were uprooted and kept on Whatman paper
under normal growth conditions in a greenhouse. Cold
stress was imposed by keeping pots containing plants at
4 �C under dim light. For ABA treatment, plants were
supplied with 100 lM ABA (Sigma, USA). Leaf and root
tissues collected from at least three different plants at
different time points of stress (8, 24 and 48 h) were thor-
oughly washed, wiped with tissue paper, immediately
frozen in liquid nitrogen, and stored at -80 �C until further
use. Tissues harvested from plants without providing stress
were used as control.
Sequence analysis
An expressed sequence tag (accession number
#EH277338) of S. alterniflora isolated from a salt stressed
EST library referred to as ‘SaARF’ was characterized in
this study. Deduced amino acid sequence was used for
Plant Cell Rep
123

multiple sequence alignment with orthologs from different
organisms using the ClustalW program (www2.ebi.ac.uk/
clustalw); phylogenetic analyses were performed in MEGA
4 (Tamura et al. 2007). The phylogenetic tree of these
sequences was inferred using the neighbor-joining method
(Saitou and Nei 1987). The bootstrap consensus tree
inferred from 1,000 replicates was used to represent the
evolutionary history of the selected eukaryotic species.
RNA isolation and cDNA synthesis
The total RNA from the leaves and roots of harvested
samples was isolated using the RNeasy plant mini kit
(Qiagen, USA), and on-column DNAse I digestion was
carried out to avoid the possible contamination of genomic
DNA following the manufacturer’s instruction (New Eng-
land Biolab, USA). Quality of total RNA was checked in a
1.2 % formamide-denaturing agarose gel, and quantifica-
tion was carried out using ND-1000 spectrophotometer
(Nanodrop Technologies, USA). First strand cDNA was
synthesized using iScriptTM first strand cDNA synthesis kit
(Bio-Rad, USA) for expression study.
Quantitative real-time polymerase chain reaction
(qRT-PCR)
Quantitative RT-PCR experiments were performed fol-
lowing the protocol of Karan et al. (2009). cDNAs syn-
thesized from unstressed and stressed samples of S.
alterniflora were used for qRT-PCR. Each 10 ll of PCR
sample contained 5 ll of 29 SYBR Green mix (Quanta
Bioscience, USA), diluted cDNA, and 0.4 lM of each
primer—SaARFRTF and SaARFRTR primers specific to
the SaARF gene (Table 1). The S. alterniflora tubulin gene,
amplified by gene-specific primers (SaTUBRTF and SaT-
UBRTR) (Table 1), was used as an internal control for
expression normalization in different cDNA samples. Melt
curve analysis was performed to check the specificity of the
amplified product, and relative gene expression levels were
determined using the 2-DDCT method (Livak and
Schmittgen 2001). The CT (cycle threshold) values for
both target and internal control genes were means of at
least three technical replicates.
The same protocol was used to analyze the expression
patterns of six abiotic stress-related genes AtSod1 (super-
oxide dismutase), AtCat (catalase), AtNhx1 (vacuolar
Na?/H? antiporter), AtSos1 (plasma membrane Na?/H?
antiporter), AtP5cs (delta-1-pyrroline-5-carboxylate syn-
thase) and AtRd22 (responsive to dehydration) (primers
listed in Table 2) in 3-week-old T3 homozygous transgenic
Arabidopsis and wild-type Columbia ecotype plants grown
in potting medium PM-15-13 (Lehle seeds, USA) under
normal growth conditions. Arabidopsis tubulin gene-spe-
cific primers, AtTUBRTF and AtTUBRTR (Table 2), were
used to normalize the variation in initial cDNA templates
used for the expression analysis.
Generation of transgenic plants
The complete ORF of SaARF was amplified by PCR using
a forward primer, SaARFNcoIF, and a reverse primer, Sa-
ARFSpeIR (Table 1), containing the NcoI and SpeI
restriction endonuclease sites, respectively, and using Pfu
DNA polymerase (New England Biolab, USA). The PCR
product was digested with NcoI and SpeI, and the NcoI–
SpeI fragment of SaARF was cloned into the pCAM-
BIA1304 vector (CAMBIA, Australia) to generate the
binary vector 35S-SaARF. The identity and orientation for
the directional cloning of SaARF into pCAMBIA1304
vector was further confirmed by DNA sequencing. 35S-
SaARF construct was introduced into Agrobacterium strain
LBA4404 by the freeze–thaw method, and the transfor-
mation of wild-type Columbia ecotype of Arabidopsis
Table 1 Primers used for cloning and qRT-PCR of SaARF gene in S.
alterniflora
Name Sequence (50–30)
SaARFNcoIF CCCATGGGGCTCGCGTTTGGGAAGCTC
SaARFSpeIR GGACTAGTTCAAGCCTTGCTTGCAATGTTG
SaARFRTF AACTGCGTGATGCTGTGCTGC
SaARFRTR TGTACCAATGCCGCTGGCGC
pCAMF GGAGAGAACACGGGGGACTCTTG
SaTUBRTF GAAGGTGATGAGGGTGATGAGT
SaTUBRTR TTCAAGCAAACAAGCCTTCATA
Table 2 Stress-related gene primers used for qRT-PCR in SaARF
transgenic Arabidopsis
Primer name Sequence (50–30)
AtTUBRTF ATAACCGTTTCAAATTCTCTCTCTC
AtTUBRTR TGCAAATCGTTCTCTCCTTG
AtRD22F GATTCGTCTTCCTCTGATCTG
AtRD22R TGGGTGTTAACGTTTACTCCG
AtP5CSF GAGGGGGTATGACTGCAAAA
AtP5CSR AACAGGAACGCCACCATAAG
AtNHX1F CCGTGCATTACTACTGGAGACAAT
AtNHX1R GTACAAAGCCACGACCTCCAA
AtSOS1F TCGTTTCAGCCAAATCAGAAAGT
AtSOS1R TTTGCCTTGTGCTGCTTTCC
AtSOD1F TCAACTGGAAATATGCAAGCGAGGT
AtSOD1R ACCACACAGCTGAGTTGAGCAAA
AtCATF AGCGCTTTCGGAGCCTCGTG
AtCATR GGCCTCACGTTAAGACGAGTTGC
Plant Cell Rep
123

(Lehle seeds, USA) was carried out by floral dip method
(Clough and Bent 1998). Positive transgenic lines were
screened on 40 mg/L hygromycin containing MS medium
(Karan and Subudhi 2012a), and integration of transgene
was confirmed by PCR using a vector-specific forward
primer, pCAMF, and SaARF-specific reverse primer, Sa-
ARFSpeIR (Table 1). The expression of the SaARF trans-
gene in transgenic SaARF plants was verified by RT-PCR
using cDNA made from total RNA isolated from positive
SaARF transgenic plants. The SaARF transgenic plants of
T3 generation were used for salinity and drought stress
experiments.
Stress tolerance assays of transgenic plants
Seeds of wild-type and SaARF transgenic Arabidopsis
plants (T3 generation) were directly sown on the potting
medium PM-15-13 (Lehle seeds, USA) and kept at 4 �C for
4 days before proceeding to stress-related experiments in a
growth chamber containing white fluorescent light of
100 lmol m-2 s-1 under 16 h light/8 h dark photoperiod
at 23 ± 1 �C.
Seed germination assays under salinity stress were per-
formed by placing WT and transgenic seeds on MS media
containing 150 mM NaCl, and the number of germinated
seeds was counted on the third day. Germination was defined
as the complete protrusion of the radicle. For seedling sur-
vival assay, uniformly germinated seeds (for 3 days) under
normal growth conditions were further transferred to
200 mM NaCl containing MS media, and the number of
surviving seedlings was counted after 7 days of stress.
For the drought tolerance assay, 4-week-old normally
grown WT and SaARF transgenic plants were kept without
irrigation for 14 days. Rosette leaves harvested at different
time points were used for various physiological and bio-
chemical analyses. At least three independent experiments
with three replicates for each WT and SaARF transgenic
lines were analyzed.
Measurement of electrolyte leakage (EL), total
chlorophyll content, and proline content
Rosette leaves of 4-week-old WT and SaARF transgenic
Arabidopsis, grown under non-stress and stress conditions
for 1 week, were harvested and used for physiological and
biochemical measurements. For EL measurement, protocol
of Bajji et al. (2004) was used. Briefly, 100 mg leaves were
placed in 25 ml distilled water and shaken on a gyratory
shaker (200 rpm) at room temperature for 2 h, and the
initial conductivity (C1) was measured with a VWR
Traceable� Expanded Range Conductivity Meter (VWR,
USA). Samples were then boiled for 10 min to induce
maximum leakage. After the samples were cooled down to
room temperature, electrolyte conductivity (C2) was mea-
sured, and the relative electrical conductivity (C %) was
calculated using the formula: (C1/C2) 9 100.
To estimate total chlorophyll content in WT and SaARF
lines, protocol suggested by Arnon (1949) was followed. One
hundred milligrams of finely powdered leaf tissue was
homogenized in 1 ml of 80 % acetone and kept for 15 min at
room temperature in dark. The crude extract was centrifuged
for 20 min at 10,000 rpm (rotation per minute) at room tem-
perature, and the resultant supernatant was used for assessing
absorbance at 663 and 645 nm with a spectrophotometer
(Shimadzu UV-1600, Japan). Total chlorophyll content was
measured using the fresh weight (FW) of samples.
For the free proline estimation of WT and SaARF
transgenic plants, fresh leaf tissues were used following the
standard protocol of Bates et al. (1973). One hundred
milligrams of leaf tissue was used and extracted in 5 mL of
3 % sulphosalicylic acid at 95 �C for 15 min. After filtra-
tion, 2 mL of supernatant was transferred to a new tube
containing 2 mL of acetic acid and 2 mL of acidified
ninhydrin reagent. After 30 min of incubation at 95 �C,
samples were kept at room temperature for an additional
30 min, and 5 mL of toluene was added to the tube with
shaking at 150 rpm to extract red products. The absorbance
of the toluene layer was determined at 532 nm using
spectrophotometer (Shimadzu UV-1600, Japan). The
standard curve was prepared using different concentrations
of proline by the same method, which was then used for
measuring free proline content in experimental samples.
The experiment was repeated at least three times.
In situ histochemical localization of reactive oxygen
species
For detection of reactive oxygen species (ROS), histo-
chemical staining with nitroblue tetrazolium (NBT) was
followed according to Dong et al. (2009) with minor
modification. Leaves detached from 4-week-old WT and
the SaARF Arabidopsis plants grown under non-stress or
drought stress for next 7 days were vacuum-infiltrated in
1 mg/ml fresh NBT solution (prepared in 10 mM phos-
phate buffer, pH 7.8) and incubated at an ambient tem-
perature until the appearance of dark spots. The stained
leaves were then bleached in concentrated ethanol, kept in
70 % ethanol, and photographed. Images were opened in
Adobe Photoshop version 7 (Adobe Systems Incorporated,
San Jose, CA, USA), and stained areas of leaves were
quantified following Lehr et al. (1997).
Statistical analysis
Mean values, standard errors, and t tests were performed
with the help of pre-loaded software in Excel, available for
Plant Cell Rep
123

statistical calculations (http://www.Physics.csbsju.edu/
stats/t-test.html). At least three biological samples with
three replicates for each WT and SaARF transgenic lines
were analyzed.
Results
SaARF is a highly conserved ortholog of ADP-
ribosylation factor
A full-length cDNA fragment with high similarity to the
ARF gene was obtained from a salt stressed cDNA library
of S. alterniflora (Baisakh et al. 2008) and designated as
SaARF. Sequence analysis of SaARF revealed the presence
of an ORF that encoded a single peptide of 181 amino acid
residues (Supplementary Fig. 1). The molecular mass of
the native SaARF protein was estimated to be 20.6 kDa.
SaARF gene sequence alignment with the ARFs of Arabi-
dopsis thaliana varied from 66 % identity (AT2G24765) to
84 % identity (AT1G70490) (Supplementary Fig. 2).
Deduced amino acid sequence comparisons of SaARF with
its orthologs from other species showed 97–99 % identity
with ARFs from Oryza sativa, A. thaliana, Zea mays,
Triticum aestivum, and Nicotiana benthamiana, while it
was 87 and 77 % identical with Homo sapiens and Sac-
charomyces cerevisiae, respectively. The SaARF protein
has four characteristic motifs unique for GTP binding, a
potential glycine-myristoylation site at position 2, and
conserved residues from 35 to 94 responsible for activating
phospholipase D (Fig. 1a). The structure of ARF proteins
was highly conserved across different species.
Fig. 1 Multiple sequence alignment and phylogenetic analysis of
SaARF protein. a Multiple sequence alignment of SaARF with ARF
proteins from various organisms. Conserved amino acids residues in
different accessions are shown in the same color. Four conserved
regions responsible for guanosine triphosphate (GTP) binding are
shown by horizontal arrows (Kahn et al. 1995); a site of myristoyla-
tion is indicated by a star (Antonny et al. 1997). Residues 35–94
(large box) are required for binding phospholipase D and adaptor
protein AP-1 (Liang et al. 1997). b Phylogenetic tree of SaARF. The
amino acid sequences of ARFs were subjected to a bootstrap test of
phylogeny by the MEGA 4.0 program using a neighbor-joining
method with 1,000 replicates. Accession numbers of ARF sequences
are: AAA32729 (A. thaliana), AAH10487 (Mus musculus),
AAO62347 (Gossypium hirsutum), AAP73857 (O. sativa),
AAU82112 (T. aestivum), ABB16972 (Solanum tuberosum),
ABY76246 (Brassica napus), BAF34209 (Nicotiana tabacum),
BAJ87426 (H. vulgare), CAA56351(Z. mays), EAL04312 (Candida
albicans), NP_001019399 (H. sapiens), XP_002318935 (Populus
trichocarpa), XP_002466220 (Sorghum bicolor), XP_003557369
(Brachypodium distachyon), and YDL137W (S. cerevisiae). Monocot
ARFs are shown in group ‘‘A’’, dicot ARFs in group ‘‘B’’, and
mammalian ARFs in group ‘‘C’’. The only exception is the underlined
BAJ87426 (Hordeum vulgare), which is clustered in the dicot group
Plant Cell Rep
123

Phylogenetic analysis revealed the clustering of monocot
SaARFs in one group, whereas dicots formed a distinct
group with a clear separation from mammalian and yeast
proteins (Fig. 1b). The only exception was Hordeum
vulgare, which was grouped with dicots. This analysis
suggests an evolutionary conserved function and possible
evolution from a common ancestor.
SaARF gene is transcriptionally regulated by abiotic
stresses in the leaf and root of S. alterniflora
To understand the possible regulation of the SaARF gene
by salinity, drought, cold, and ABA in leaves and roots of
S. alterniflora, a qRT-PCR study was carried out. In
leaves, salt and drought stress continuously increased the
expression of SaARF gene and increased fourfold by
48 h of stress, while cold and ABA increased the
expression of SaARF up to 24 h and then decreased to a
basal level by 48 h (Fig. 2a). In roots, SaARF expression
increased up to fourfold by 8 h salinity stress and
induced up to fivefold by 48 h of stress (Fig. 2b).
Expression of SaARF under drought stress in root was
highly conspicuous, increased gradually, and increased
eightfold during 48 h of stress. In roots, cold stress
increased the expression of SaARF by around twofold in
8 h and started decreasing afterwards, while ABA
increased its expression by 2.5-fold during 24 h of
exposure, and then decreased and reached to a basal level
by 48 h. Induced expression of SaARF transcripts with
8 h of salinity, drought, cold, and ABA indicated its
possible involvement in abiotic stress mechanisms
operating in S. alterniflora.
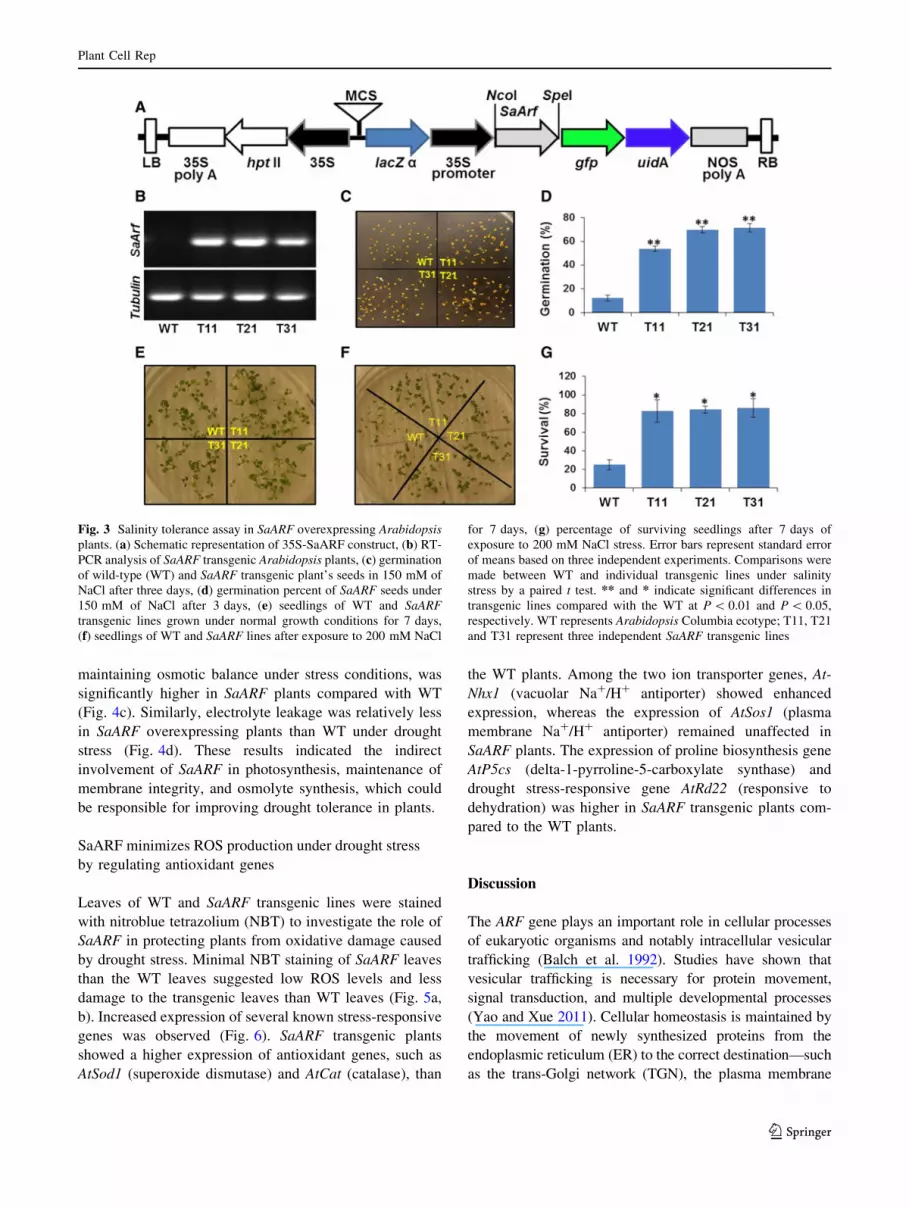
Overexpression of SaARF improved seed germination
and seedling survival under salinity stress
To identify the possible role of SaARF toward salinity
tolerance in plants, SaARF was transformed into Arabi-
dopsis ecotype Columbia, and the expression of the trans-
gene was confirmed by RT-PCR (Fig. 3b). Seeds of three
independent homozygous SaARF transgenic lines in T3
generation were analyzed for salinity tolerance. Seed ger-
mination under 150 mM NaCl stress was only 10 % in
wild-type (WT) Columbia ecotype, whereas it increased
significantly in all three SaARF overexpressing transgenic
lines with germination ranging from 50 to 70 % (Fig. 3c,
d). Similarly, there was fourfold increase in seedling sur-
vival under salinity stress in SaARF overexpressing
Arabidopsis plants compared to WT plants (Fig. 3f, g).
This analysis clearly revealed the probable involvement of
the SaARF gene in salinity stress tolerance mechanisms in
plants.
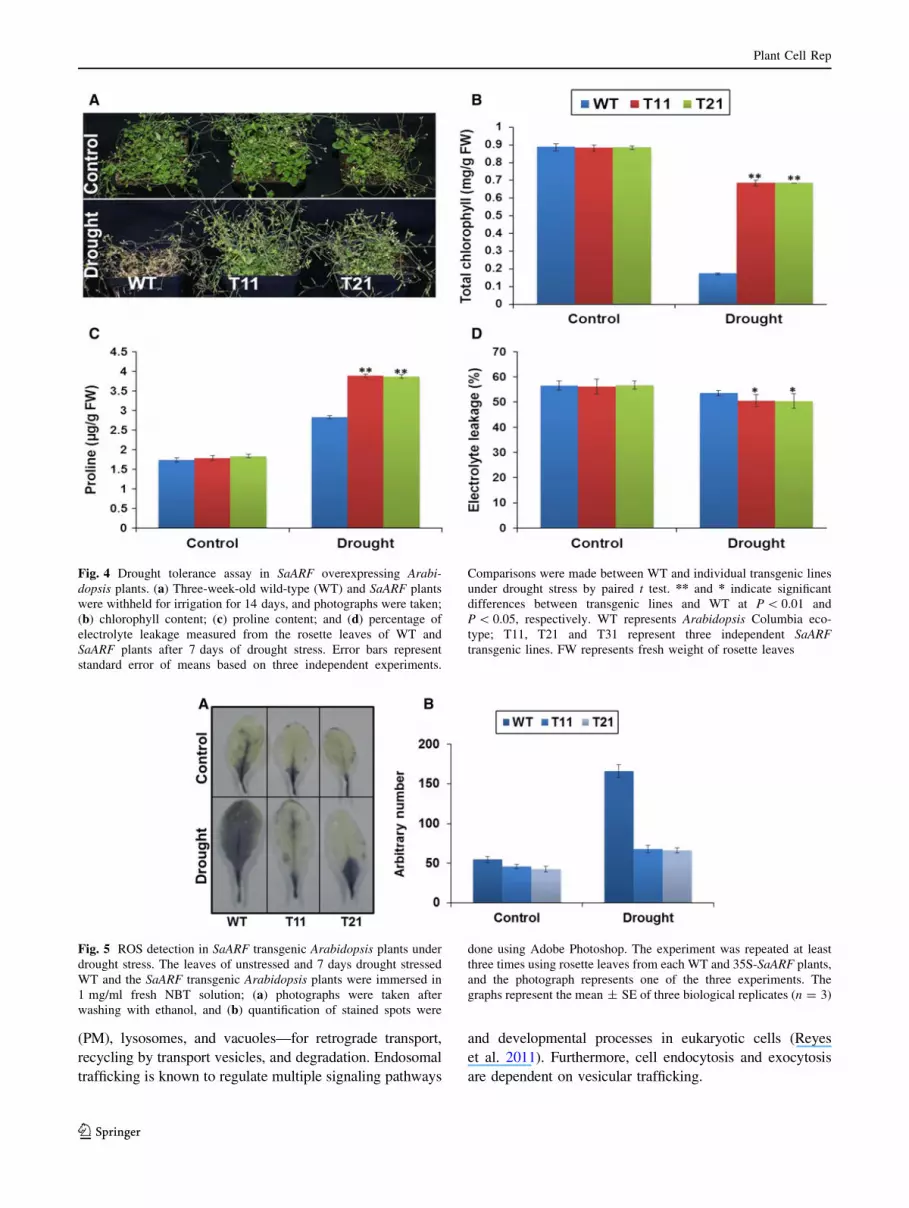
Overexpression of SaARF enhanced drought tolerance
by altering osmolyte synthesis
Wild-type and SaARF transgenic plants were compared for
their performance under drought stress to further investi-
gate the role of SaARF in conferring drought tolerance.
Under drought stress, most of the WT plants appeared
dehydrated and weak, and eventually died, while SaARF
overexpressing Arabidopsis plants grew normally and set
seeds (Fig. 4a). Total chlorophyll content was significantly
higher in SaARF transgenic plants than WT under drought
stress (Fig. 4b). Proline, an osmolyte responsible for
Fig. 2 Expression kinetics of SaARF in leaves and roots of S.
alterniflora under different abiotic stresses. Expression patterns of
SaARF at different time intervals in leaves (a) and roots (b) under
salinity stress (5 % sea salt), drought (kept on Whatman paper), cold
(at 4 �C), and ABA (100 lM). Samples were harvested at different
time intervals: 8, 24, and 48 h. Samples harvested before stress were
used as control (c). The tubulin gene of S. alterniflora was used as an
internal control for the normalization of different cDNA samples.
Error bars represent standard error of means based on three
independent reactions
Plant Cell Rep
123
maintaining osmotic balance under stress conditions, was
significantly higher in SaARF plants compared with WT
(Fig. 4c). Similarly, electrolyte leakage was relatively less
in SaARF overexpressing plants than WT under drought
stress (Fig. 4d). These results indicated the indirect
involvement of SaARF in photosynthesis, maintenance of
membrane integrity, and osmolyte synthesis, which could
be responsible for improving drought tolerance in plants.
SaARF minimizes ROS production under drought stress
by regulating antioxidant genes
Leaves of WT and SaARF transgenic lines were stained
with nitroblue tetrazolium (NBT) to investigate the role of
SaARF in protecting plants from oxidative damage caused
by drought stress. Minimal NBT staining of SaARF leaves
than the WT leaves suggested low ROS levels and less
damage to the transgenic leaves than WT leaves (Fig. 5a,
b). Increased expression of several known stress-responsive
genes was observed (Fig. 6). SaARF transgenic plants
showed a higher expression of antioxidant genes, such as
AtSod1 (superoxide dismutase) and AtCat (catalase), than
the WT plants. Among the two ion transporter genes, At-
Nhx1 (vacuolar Na?/H? antiporter) showed enhanced
expression, whereas the expression of AtSos1 (plasma
membrane Na?/H? antiporter) remained unaffected in
SaARF plants. The expression of proline biosynthesis gene
AtP5cs (delta-1-pyrroline-5-carboxylate synthase) and
drought stress-responsive gene AtRd22 (responsive to
dehydration) was higher in SaARF transgenic plants com-
pared to the WT plants.
Discussion
The ARF gene plays an important role in cellular processes
of eukaryotic organisms and notably intracellular vesicular
trafficking (Balch et al. 1992). Studies have shown that
vesicular trafficking is necessary for protein movement,
signal transduction, and multiple developmental processes
(Yao and Xue 2011). Cellular homeostasis is maintained by
the movement of newly synthesized proteins from the
endoplasmic reticulum (ER) to the correct destination—such
as the trans-Golgi network (TGN), the plasma membrane
Fig. 3 Salinity tolerance assay in SaARF overexpressing Arabidopsis
plants. (a) Schematic representation of 35S-SaARF construct, (b) RT-
PCR analysis of SaARF transgenic Arabidopsis plants, (c) germination
of wild-type (WT) and SaARF transgenic plant’s seeds in 150 mM of
NaCl after three days, (d) germination percent of SaARF seeds under
150 mM of NaCl after 3 days, (e) seedlings of WT and SaARF
transgenic lines grown under normal growth conditions for 7 days,
(f) seedlings of WT and SaARF lines after exposure to 200 mM NaCl
for 7 days, (g) percentage of surviving seedlings after 7 days of
exposure to 200 mM NaCl stress. Error bars represent standard error
of means based on three independent experiments. Comparisons were
made between WT and individual transgenic lines under salinity
stress by a paired t test. ** and * indicate significant differences in
transgenic lines compared with the WT at P \ 0.01 and P \ 0.05,
respectively. WT represents Arabidopsis Columbia ecotype; T11, T21
and T31 represent three independent SaARF transgenic lines
Plant Cell Rep
123
(PM), lysosomes, and vacuoles—for retrograde transport,
recycling by transport vesicles, and degradation. Endosomal
trafficking is known to regulate multiple signaling pathways
and developmental processes in eukaryotic cells (Reyes
et al. 2011). Furthermore, cell endocytosis and exocytosis
are dependent on vesicular trafficking.
Fig. 4 Drought tolerance assay in SaARF overexpressing Arabi-
dopsis plants. (a) Three-week-old wild-type (WT) and SaARF plants
were withheld for irrigation for 14 days, and photographs were taken;
(b) chlorophyll content; (c) proline content; and (d) percentage of
electrolyte leakage measured from the rosette leaves of WT and
SaARF plants after 7 days of drought stress. Error bars represent
standard error of means based on three independent experiments.
Comparisons were made between WT and individual transgenic lines
under drought stress by paired t test. ** and * indicate significant
differences between transgenic lines and WT at P \ 0.01 and
P \ 0.05, respectively. WT represents Arabidopsis Columbia eco-
type; T11, T21 and T31 represent three independent SaARF
transgenic lines. FW represents fresh weight of rosette leaves
Fig. 5 ROS detection in SaARF transgenic Arabidopsis plants under
drought stress. The leaves of unstressed and 7 days drought stressed
WT and the SaARF transgenic Arabidopsis plants were immersed in
1 mg/ml fresh NBT solution; (a) photographs were taken after
washing with ethanol, and (b) quantification of stained spots were
done using Adobe Photoshop. The experiment was repeated at least
three times using rosette leaves from each WT and 35S-SaARF plants,
and the photograph represents one of the three experiments. The
graphs represent the mean ± SE of three biological replicates (n = 3)
Plant Cell Rep
123

In this study, an ARF gene of the halophytic plant S.
alterniflora, ‘‘SaARF,’’ was investigated for its role in the
plant’s adaptation under salinity and drought stresses. To
date, a number of ARF genes have been isolated from
various eukaryotes, and their amino acid sequences have
shown highly conserved motifs for the binding of GTP
(Kahn et al. 1995), GAPs (Amor et al. 1994), and guanine
nucleotide exchange factors (GEFs) Sec7 (Mossessova
et al. 1998). It has a potential myristoylation site at Gly-2
(Antonny et al. 1997) and at amino acid residues 35–94,
which activates phospholipase D (Liang et al. 1997). Pre-
sence of all the above motifs in SaARF protein indicates
potentially conserved structures of the SaARF protein such
as those of other organisms (Fig. 1a). The SaARF protein is
highly identical to ARF of monocot plants, such as O.
sativa, Z. mays, and T. aestivum, and seems to have
evolved from a common ancestor (Fig. 1b). Furthermore,
SaARF was found to be 84 % identical with ARF of
Arabidopsis with accession number AT1G70490 (Supple-
mentary Fig. 2) of 15 ARF members of Arabidopsis
(Gebbie et al. 2005).
Plants are constantly challenged with environmentally
extreme conditions and therefore have developed mecha-
nisms to sense and transduce signals to the appropriate
genetic machinery to prepare plants for the unfavorable
impending changes in the surrounding environment. Plants
respond to such stresses by changing its pattern of protein
expression. Plants have to maintain its cellular activity to
continue its growth under stress environments, which
requires the transportation of protein molecules to various
cellular compartments inside the cell. Since the direct role
of ARF in vesicular trafficking in plants is well established
(Ritzenthaler et al. 2002), it is tempting to speculate how
intracellular transportation is regulated and integrated into
signaling pathways and cellular responses in achieving
tolerance to multiple abiotic stresses (Levine 2002).
The ARF gene has been reported to be preferentially
expressed in different organs of wheat (Yao et al. 2009)
and in caryopses at early developmental stages in maize
(Liu et al. 2010) and rice (Zhou et al. 2010). ARF has been
reported to be induced by ethylene in regulating posthar-
vest ripening of banana (Wang et al. 2010), and its
expression was enhanced in flowers and buds in cotton
(Ren et al. 2004) and potato tubers (Liu et al. 2012).
Although organ-specific transcriptional regulation of ARF
in plants was indicated in numerous studies, as discussed
above, the role of ARF in abiotic stress response was only
studied in physic nuts (Jatropha curcas) (Qin et al. 2011).
In this plant, drought, salinity, ABA, ethephon, heat, and
cold influenced the expression of ARF gene in the leaf,
stem, and root. In our study, the SaARF gene was consti-
tutively expressed in the leaves and roots of S. alterniflora,
which was further upregulated by salinity, drought, cold,
and ABA (Fig. 2), indicating the stress-responsive nature
of SaARF.
Previous studies have provided evidence for the
involvement of this gene in plants’ response to diseases
(Lee et al. 2003; Coemans et al. 2008), growth, and
development (Yao et al. 2009; Zhou et al. 2010). Gebbie
et al. (2005) provided evidence that ARF plays an important
role in new cell wall formation since many of the compo-
nent processes, such as cell division, cell expansion, and
cellulose production, are dependent on vesicle trafficking
regulated by the ARF gene. Further, influences on hormonal
and signaling pathways in their study using the antisense
approach were concluded due to changes in flowering pat-
tern and apical dominance. The overexpression of TaARF in
Arabidopsis resulted in increased leaf area, increased
Fig. 6 Expression of stress-responsive genes in SaARF Arabidopsis
plants. Relative mRNA levels of stress-responsive genes were
determined by quantitative RT-PCR using cDNA synthesized from
total RNAs isolated from the shoots of 3-week-old WT and SaARF
transgenic Arabidopsis plants grown under normal conditions in
potting medium. The Arabidopsis tubulin gene was used as an internal
control for the normalization of different cDNA samples. Error bars
represent standard error of means based on three independent
reactions
Plant Cell Rep
123

growth rate, and earlier transition to flowering (Yao et al.
2009). Data from our study suggest that SaARF may be
involved in the trafficking of protein molecules related to
abiotic stress tolerance mechanisms. Since ARF over-
expressing plants were able to grow and maintain a sig-
nificantly higher level of photosynthesis and membrane
stability under drought stress compared to WT plants, it
could be possible by maintaining the cell division and cell
expansion under stress conditions (Gebbie et al. 2005). This
is further supported by the fact that ARF is a crucial factor
for root and root hair growth (Song et al. 2006).
Considering the stress inducible nature of SaARF in S.
alterniflora, the SaARF gene was further tested for its role
toward abiotic stress adaptations in plants. Overexpressing
SaARF Arabidopsis plants showed increased seed germi-
nation and seedling survival than WT plants under salinity
stress, suggesting the involvement of SaARF in salinity
stress adaptation of plants (Fig. 3). Under drought stress,
SaARF Arabidopsis plants were healthy and green, while
WT plants lost their vigor and growth. SaARF plants accu-
mulated more chlorophyll and proline under drought stress
and showed improved cell membrane stability as its elec-
trolyte leakage was significantly lower than the WT (Fig. 4).
SaARF overexpression minimized oxidative damage
caused by drought stress-induced accumulation of ROS, as
shown in Fig. 5. Plants have evolved a complex antioxi-
dant system to detoxify stress-induced ROS by enzymes,
such as superoxide dismutase (Sod) (Alscher et al. 2002)
and catalase (Cat) (Havir and McHale 1989). ARF has
been previously documented to regulate metabolism and
antioxidant capacity in transgenic potato tubers (Zuk et al.
2003). Increased expression of AtSod1 and AtCat in SaARF
Arabidopsis plants (Fig. 6) revealed further evidence for
indirect involvement of SaARF in improving osmotic stress
tolerance by scavenging ROS produced by drought stress.
Proline is an important osmolyte for adjusting osmosis,
stabilizing cellular structures, and providing osmotic stress
tolerance to plants (Verbruggen and Hermans 2008; Karan
and Subudhi 2012a, b). Enhanced expression of the proline
biosynthesis gene AtP5cs (Yoshiba et al. 1995) leads to the
production of additional proline in SaARF Arabidopsis
plants and suggests the influence of ARF on abiotic stress
tolerance via proline biosynthesis pathways. In addition,
the enhanced expressions of the ion transporter gene At-
Nhx1 (Apse et al. 2003) and the dehydration responsive
gene AtRd22 (Yamaguchi-Shinozaki and Shinozaki 1993)
in SaARF Arabidopsis plants suggested for the first time
the importance of the ARF gene in regulating abiotic stress
responses in plants, possibly by the cross-talking of dif-
ferent abiotic stress signaling pathways (Knight and Knight
2001), which are dependent on intracellular trafficking and
are influenced by various plant hormones such as ABA
(Levine 2002).
The demonstration of the defensive role played by the
small GTP-binding protein SaARF in plant cells against
environmental disturbances in this study suggest that Sa-
ARF would be a potential candidate gene in developing
plants tolerant to multiple abiotic stresses using the trans-
genic approach. However, how the ARF gene regulates
vesicle formation and trafficking and influences multiple
tolerance mechanisms should be explored.
Acknowledgments This work was supported by Hatch and special
grant funds from the United States Department of Agriculture—
National Institute of Food and Agriculture. The manuscript was
approved for publication by the Director of Louisiana Agricultural
Experiment Station, USA as manuscript number 2013-306-11800.
References
Alscher RG, Erturk N, Heath LS (2002) Role of superoxide
dismutases (SODs) in controlling oxidative stress in plants.
J Exp Bot 53:1331–1341
Amor JC, Harrison DH, Kahn RA, Ringe D (1994) Structure of the
human ADP-ribosylation factor 1 complexed with GDP. Nature
372:704–708
Antonny B, BeraudDufour S, Chardin P, Chabre M (1997) N-terminal
hydrophobic residues of the G-protein ADP-ribosylation factor-1
insert into membrane phospholipids upon GDP to GTP
exchange. Biochemistry 36:4675–4684
Apse MP, Sottosanto JB, Blumwald E (2003) Vacuolar cation/H
exchange, ion homeostasis, and leaf development are altered in a
T-DNA insertional mutant of AtNHX1, the Arabidopsis vacuolar
Na?/H? antiporter. Plant J 36:229–239
Arnon D (1949) Copper enzymes in isolated chloroplasts. Polyphe-
noloxidase in Beta vulgaris. Plant Physiol 24:1–15
Asakura Y, Ishigaki E, Sugiyama R, Kurosaki F (2007) Cloning and
expression of cDNAs encoding ADP-ribosylation factor in carrot
seedling. Plant Sci 172:189–195
Baisakh N, Subudhi PK, Varadwaj P (2008) Primary responses to salt
stress in a halophyte, smooth cordgrass (Spartina alterniflora
Loisel.). Funct Integr Genomics 8:287–300
Baisakh N, RamanaRao MV, Rajasekaran K, Subudhi P, Janda J,
Galbraith D, Vanier C, Pereira A (2012) Enhanced salt stress
tolerance of rice plants expressing a vacuolar H? -ATPase
subunit c1 (SaVHAc1) gene from the halophyte grass Spartina
alterniflora Loisel. Plant Biotech J 10:453–464
Bajji M, Bertin P, Lutts S, Kinet JM (2004) Evaluation of drought
resistance-related traits in durum wheat somaclonal lines
selected in vitro. Aust J Exp Agric 44:27–35
Balch WE, Kahn RA, Schwaninger R (1992) ADP-ribosylation factor
required for vesicular trafficking between the endoplasmic
reticulum and the cis-Golgi compartment. J Biol Chem
267:13053–13061
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free
proline for water stress studies. Plant Soil 39:205–207
Bressan RA, Park HC, Orsini F, Oh D, Dassanayake M, Inan G, Yun
G, Bohnert HJ, Maggio A (2013) Biotechnology for mechanisms
that counteract salt stress in extremophile species: a genome-
based view. Plant Biotech Rep 7:27–37
Clough SJ, Bent AF (1998) Floral dip: a simplified method for
Agrobacterium mediated transformation of Arabidopsis thaliana.
Plant J 16:735–743
Plant Cell Rep
123

Coemans B, Takahashi Y, Berberich T, Ito A, Kanzaki H, Matsumura
H, Saitoh H, Tsuda S, Kamoun S, Sagi L, Swennen R, Terauchi
R (2008) High-throughput in planta expression screening
identifies an ADP-ribosylation factor (ARF1) involved in non-
host resistance and R gene-mediated resistance. Mol Plant Pathol
9:25–36
D’Souza-Schorey C, Chavrier P (2006) ARF proteins: roles in
membrane traffic and beyond. Nat Rev Mol Cell Biol 7:347–358
Donaldson JG, Jackson CL (2000) Regulators and effectors of the
ARF GTPases. Curr Opin Cell Biol 12:475–482
Dong CH, Zolman BK, Bartel B, Lee BH, Stevenson B, Agarwal M,
Zhu JK (2009) Disruption of Arabidopsis CHY1 reveals an
important role of metabolic status in plant cold stress signaling.
Mol Plant 2:59–72
Gebbie LK, Burn JE, Hocart CH, Williamson RE (2005) Genes
encoding ADP-ribosylation factors in Arabidopsis thaliana L.
Heyn.; genome analysis and antisense suppression. J Exp Bot
56:1079–1091
Geldner N, Anders N, Wolters H, Keicher J, Kornberger W, Muller P,
Delbarre A, Ueda T, Nakano A, Jurgens G (2003) The
Arabidopsis GNOM ARF-GEF mediates endosomal recycling,
auxin transport, and auxin-dependent plant growth. Cell
112:219–230
Gillingham AK, Munro S (2007) The small G proteins of the Arf
family and their regulators. Annu Rev Cell Dev Biol 23:579–611
Havir EA, McHale NA (1989) Enhanced peroxidatic activity in
specific catalase isozymes of tobacco, barley, and maize. Plant
Physiol 91:812–815
Higo H, Kishimoto N, Saito A, Higo K (1994) Molecular cloning and
characterization of a cDNA encoding a small GTP-binding
protein related to mammalian ADP ribosylation factor from rice.
Plant Sci 100:41–49
Hoagland DR, Arnon DI (1950) The water-culture method for
growing plants without soil. Calif Agr Expt Sta Circ 347:1–32
Inoue H, Randazzo PA (2007) Arf GAPs and their interacting
proteins. Traffic 8:1465–1475
Jones DH, Bex B, Fensome A, Cockcroft S (1999) ADP ribosylation
factor 1 mutants identify a phospholipase D effector region and
reveal that phospholipase D participates in lysosomal secretion
but is not sufficient for recruitment of coatomer I. Biochem J
341:185–192
Kahn RA, Gilman AG (1984) Purification of a protein cofactor
required for ADP-ribosylation of the stimulatory regulatory
component of adenylate cyclase by cholera toxin. J Biol Chem
259:6228–6234
Kahn RA, Clark J, Rulka C, Stearns T, Zhang CJ, Randazzo PA, Terui
T, Cavenagh M (1995) Mutational analysis of Saccharomyces
cerevisiae ARF1. J Biol Chem 270:143–150
Karan R, Subudhi PK (2012a) A stress inducible SUMO conjugating
enzyme gene (SaSce9) from a grass halophyte Spartina alter-
niflora enhances salinity and drought stress tolerance in Arabi-
dopsis. BMC Plant Biol 12:187
Karan R, Subudhi PK (2012b) Overexpression of a nascent polypep-
tide associated complex gene (SabNAC) of Spartina alterniflora
improves tolerance to salinity and drought in transgenic Arabi-
dopsis. Biochem Biophys Res Commun 424:747–752
Karan R, Singla-Pareek SL, Pareek A (2009) Histidine kinase and
response regulator genes as they relate to salinity tolerance in
rice. Funct Integr Genomics 9:411–417
Knight H, Knight MR (2001) Abiotic stress signalling pathways:
specificity and cross-talk. Trends Plant Sci 6:262–267
Kobayashi-Uehara A, Shimosaka E, Handa H (2001) Cloning and
expression analysis of cDNA encoding an ADP-ribosylation
factor from wheat: tissue-specific expression of wheat ARF.
Plant Sci 160:535–542
Lee MH, Sano H (2007) Attenuation of the hypersensitive response
by an ATPase associated with various cellular activities (AAA)
protein through suppression of a small GTPase, ADP ribosyl-
ation factor, in tobacco plants. Plant J 51:127–139
Lee WY, Hong JK, Kim CY, Chun HJ, Park HC, Kim JC, Yun DJ,
Chung WS, Lee SH, Lee SY, Cho MJ, Lim CO (2003) Over-
expressed rice ADP-ribosylation factor 1 (RARF1) induces
pathogenesis-related genes and pathogen resistance in tobacco
plants. Physiol Plant 119:573–581
Lehr HA, Mankoff DA, Corwin D, Santeusanio G, Gown AM (1997)
Application of Photoshop-based image analysis to quantification
of hormone receptor expression in breast cancer. J Histochem
Cytochem 45:1559–1565
Levine A (2002) Regulation of stress responses by intracellular
vesicle trafficking? Plant Physiol Biochem 40:531–535
Liang JO, Sung TC, Morris AJ, Frohman MA, Kornfeld S (1997)
Different domains of mammalian ADP-ribosylation factor 1
mediate interaction with selected target proteins. J Biol Chem
272:33001–33008
Liu Y, Li J, Li Y, Wei M, Cui Q, Wang Q (2010) Molecular cloning,
sequence and expression analysis of ZmArf2, a maize ADP-
ribosylation factor. Mol Biol Rep 37:755–761
Liu B, Si H, Zhang N, Wang D (2012) Identification of differentiallyexpressed genes in potato associated with tuber dormancy
release. Mol Biol Rep 39:11277–11287
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression
data using real-time quantitative PCR and the 2-DDCT method.
Methods 25:402–408
McElver J, Patton D, Rumbaugh M, Liu CM, Yang LJ, Meinke D
(2000) The TITAN5 gene of Arabidopsis encodes a protein
related to the ADP-ribosylation factor family of GTP binding
proteins. Plant Cell 12:1379–1392
Mossessova E, Gulbis JM, Goldberg J (1998) Structure of the guanine
nucleotide exchange factor Sec7 domain of human Arno and
analysis of the interaction with ARF GTPase. Cell 92:415–423
Nakashima K, Ito Y, Yamaguchi-Shinozaki K (2009) Transcriptional
regulatory networks in response to abiotic stresses in Arabi-
dopsis and grasses. Plant Physiol 149:88–95
Naramoto S, Kleine-Vehn J, Robert S, Fujimoto M, Dainobu T,
Paciorek T, Ueda T, Nakano A, Van Montagu MC, Fukuda H,
Friml J (2010) ADP-ribosylation factor machinery mediates
endocytosis in plant cells. Proc Natl Acad Sci USA
107:21890–21895
Pimpl P, Movafeghi A, Coughlan S, Denecke J, Hillmer S, Robinson
DG (2000) In situ localization and in vitro induction of plant
COPI-coated vesicles. Plant Cell 12:2219–2236
Qin X, Lin F, Lii Y, Gou C, Chen F (2011) Molecular analysis of
ARF1 expression profiles during development of physic nut
(Jatropha curcas L.). Mol Biol Rep 38:1681–1686
Ren MZ, Chen QJ, Zhang R, Guo SD (2004) The structural
characteristics, alternative splicing and genetic expression ana-
lysis of ADP-ribosylation factor 1 (arf1) in cotton. Acta Genet
Sin 31:850–857
Reyes FC, Buono R, Otegui MS (2011) Plant endosomal trafficking
pathways. Curr Opin Plant Biol 14:666–673
Ritzenthaler C, Nebenfuhr A, Movafeghi A, Stussi-Garauda C,
Behniac L, Pimplc P, Staehelin LA, Robinson DG (2002)
Reevaluation of the effects of brefeldin A on plant cells using
tobacco Bright Yellow 2 cells expressing Golgi-targeted green
fluorescent protein and COPI antisera. Plant Cell 14:237–261
Saitou N, Nei M (1987) The neighbor-joining method: a new method
for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Song XF, Yang CY, Liu J, Yang WC (2006) RPA, a class II ARFGAP
protein, activates ARF1 and U5 and plays a role in root hair
development in Arabidopsis. Plant Physiol 141:966–976
Plant Cell Rep
123

Subudhi PK, Baisakh N (2011) Spartina alterniflora Loisel., a
halophyte grass model to dissect salt stress tolerance. In Vitro
Cell Dev Biol Plant 47:441–457
Szopa J, Muller-Rober B (1994) Cloning and expression analysis of
an ADP-ribosylation factor from Solanum tuberosum L. Plant
Cell Rep 14:180–183
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular
evolutionary genetics analysis (MEGA) software version 4.0.
Mol Biol Evol 24:1596–1599
Tzafrir I, McElver JA, Liu CM, Yang LJ, Wu JQ, Martinez A, Patton
DA, Meinke DW (2002) Diversity of TITAN functions in
Arabidopsis seed development. Plant Physiol 128:38–51
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a
review. Amino Acids 35:753–759
Verwoert IIGS, Brown A, Slabas AR, Stuitje AR (1995) A Zea mays
GTP-binding protein of the ARF family complements an
Escherichia coli mutant with a temperature-sensitive malonyl-
coenzyme A: acyl carrier protein transacylase. Plant Mol Biol
27:629–633
Wang Y, Wu J, Xu BY, Liu JH, Zhang JB, Jia CH, Jin ZQ (2010)
Cloning of an ADP-ribosylation factor gene from banana (Musa
acuminata) and its expression patterns in postharvest ripening
fruit. J Plant Physiol 167:989–995
Xu J, Scheres B (2005) Dissection of Arabidopsis ADP-RIBOSYL-
ATION FACTOR 1 function in epidermal cell polarity. Plant
Cell 17:525–536
Yahara N, Ueda T, Sato K, Nakano A (2001) Multiple roles of Arf1
GTPase in the yeast exocytic and endocytic pathways. Mol Biol
Cell 12:221–238
Yamaguchi-Shinozaki K, Shinozaki K (1993) The plant hormone
abscisic acid mediates the drought-induced expression but not
the seed-specific expression of rd22, a gene responsive to
dehydration stress in Arabidopsis thaliana. Mol Gen Genet
238:17–25
Yao HY, Xue HW (2011) Signals and mechanisms affecting vesicular
trafficking during root growth. Curr Opin Plant Biol 14:571–579
Yao Y, Ni Z, Du J, Han Z, Chen Y, Zhang Q, Sun Q (2009) Ectopic
overexpression of wheat adenosine diphosphate-ribosylation
factor, TaARF, increases growth rate in Arabidopsis. J Integr
Plant Biol 51:35–44
Yoshiba Y, Kiyosue T, Katagiri T, Ueda H, Mizoguchi T, Yamag-
uchi-Shinozaki K, Wada K, Harada Y, Shinozaki K (1995)
Correlation between the induction of a gene for D1-pyrroline-5-
carboxylate synthetase and the accumulation of proline in
Arabidopsis thaliana under osmotic stress. Plant J 7:751–760
Zerial M, Huber LA (1995) Guidebook to the small GTPases. Oxford
University Press, Oxford
Zhou X, Li J, Cheng W, Liu H, Li M, Zhang Y, Li W, Han S, Wan Y
(2010) Gene structure analysis of rice ADP-ribosylation factors
(OsARFs) and their mRNA expression in developing rice plants.
Plant Mol Biol Rep 28:692–703
Zhu JK (2002) Salt and drought stress signal transduction in plants.
Annu Rev Plant Biol 53:247–273
Zuk M, Prescha A, Kepczynski J, Szopa J (2003) ADP ribosylation
factor regulates metabolism and antioxidant capacity of trans-
genic potato tubers. J Agric Food Chem 51:288–294
Plant Cell Rep
123