effect of growth factors on nuclear and mitochondrial adp-ribosylation processes during astroglial...
TRANSCRIPT

Mechanisms of Ageing and Development
123 (2002) 511–520
Effect of growth factors on nuclear and mitochondrialADP-ribosylation processes during astroglial cell
development and aging in culture
Vittoria Spina Purrello a, Gianfrancesco Cormaci a, Luca Denaro b,Salvatore Reale a, Antonino Costa a, Calogera Lalicata a, Maurizio Sabbatini c,
Bianca Marchetti d, Roberto Avola a,*a Department of Chemical Sciences, Section of Biochemistry and Molecular Biology, Viale Andrea Doria, 6, Uni�ersity of Catania,
95125 Catania, Italyb Neurosurgery Institute, Uni�ersita Cattolica del S. Cuore, 00167 Rome, Italy
c Department of Pharmacological Sciences and Experimental Medicine, Section of Human Anatomy, Uni�ersity of Camerino,62032 Camerino, Italy
d OASI Institute for Research and Care (IRCCS) on Mental Retardation and Brain Aging, Troina,Department of Pharmacology and Ginecology, Uni�ersity of Sassari, 07100 Sassari, Italy
Received 2 February 2001; received in revised form 11 June 2001; accepted 27 July 2001
Abstract
Epidermal growth factor (EGF), basic fibroblast growth factor (bFGF), insulin-like growth factor-I (IGF-I) andinsulin (INS) are powerful mitogens and may regulate gene expression in cultured astrocytes by ADP-ribosylationprocess. Nuclear poly-ADP ribose polymerase (PARP) and mitochondrial monoADP-ribosyltransferase (ADPRT)are the key enzymes involved in poly-ADP-ribosylation and mono ADP-ribosylation, respectively. In this investiga-tion the effect of EGF, bFGF, IGF-I or INS on nuclear PARP and mitochondrial ADPRT activities were assessedin nuclei and mitochondria purified from developing (30 DIV) or aging (90 and 190 DIV) primary rat astrocytecultures. A marked increase of PARP activity in bFGF or IGF-I treated astroglial cell cultures at 30 DIV was found.Nuclear PARP and mitochondrial ADPRT activities were greatly stimulated by treatment with EGF or INS alone ortogether in astrocyte cultures at 30 DIV. Nuclear PARP and mitochondrial ADPRT activities showed a moreremarkable increase in control untreated astrocyte cultures at 190 DIV than at 90 DIV. These findings suggest thatADP-ribosylation process is involved in DNA damage and repair during cell differentiation and aging in culture.Twelve hours treatment with EGF, INS or bFGF significantly stimulated nuclear PARP and mitochondrial ADPRTactivities in 190 DIV aging astrocyte cultures. The above results indicate that EGF, INS and bFGF may play acrucial role in the post-translational modification of chromosomal proteins including ADP-ribosylation process in invitro models. This suggests that growth factors regulate genomic stability in glial cells during development and
www.elsevier.com/locate/mechagedev
* Corresponding author. Tel.: +39-095-738-4062; fax: +39-095-336-990.E-mail address: [email protected] (R. Avola).
0047-6374/02/$ - see front matter © 2002 Published by Elsevier Science Ireland Ltd.
PII: S 0 0 47 -6374 (01 )00354 -2

V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520512
maturation, stimulating nuclear and mitochondrial ADP-ribosylation processes in developing or aging astrocytecultures. © 2002 Published by Elsevier Science Ireland Ltd.
Keywords: Astroglial cell cultures; Development; Aging; Growth factors; ADP-ribosylation processes
1. Introduction
Different types of astrocytes were identified inthe central nervous system. They have a func-tional and regional heterogeneity that couldreflect differences on scheduled brain development(Garcia-Abreu et al., 1996). Astrocyte differentia-tion is switched on by a transition program in-volving the vimentin-glial fibrillary acidic protein(GFAP) binome. GFAP is the main marker ofmature cultured astrocytes (Bignami et al., 1972),whereas vimentin is expressed only in develop-mental stages (Dahl, 1981). Increased expressionof GFAP and down-regulation of vimentin, indi-cate astrocyte maturation (Dahl et al., 1981).
Astroglial cells release peptide growth factors insurrounding environment during developmentand maturation. They act as stimulants of cellgrowth and differentiation (Morrison et al., 1982)and play an important role in glia–neuron cross-talk (Avola et al., 2000). Epidermal growth factor(EGF), basic fibroblast growth factor (bFGF),insulin-like growth factor-I (IGF-I) and insulin(INS), are powerful mitogens and neurotrophicfactors in several cell types (Morrison et al., 1986;Rotwein et al., 1988; Enfors et al., 1990; Torres-Aleman et al., 1990; Casper et al., 1991). EGFstimulates mitogenesis of astroglial cells in pri-mary culture (Avola et al., 1988a,b), whereasbFGF and IGF-I are crucial factors during braindevelopment (Rotwein et al., 1988; Han et al.,1992), and influence neurite sprouting and out-growth of neurons derived from different brainareas (Morrison et al., 1986; Walicke et al., 1986).These functions are exerted through signaling cas-cades triggered by their binding on cognate recep-tors (Carpenter and Cohen, 1990; Fantl et al.,1993). Activated receptors undergo autophospho-rylation on specific tyrosine residues. They coupleto downstream adaptors such as Grb-2, Shc, Gab-1, Nck and the p85 subunit of phosphatidyl-inosi-tol (PI) 3-kinase and leading mainly to smallG-protein RasGTP loading and activation. This,
in turn, activates the extracellular signal-regulatedkinases 1 and 2 (ERK1 and 2) cascade, the phos-pholipase A2-mediated arachidonic acid release,the phospholipase C�-mediated phosphoinositidehydrolysis, classical protein kinases C (PKC)membrane translocation and inositol-1,4,5-trisphosphate (IP3)-mediated calcium mobilizationfrom endoplasmic reticulum stores (Berridge,1987).
The above biochemical events activate severaltranscription factors, such as fos/jun activatorprotein-1 (AP-1), serum response factor (SRF),activation transcription factor 2 (ATF-2), earlygrowth response 1 (Egr-1) and polyADP-ribosepolymerase (PARP) (Homburg et al., 2000).These events in sequence modulate nuclear geneexpression activities linked to proliferation or dif-ferentiation (Denhardt, 1996). PARP enzymepolyADP-ribosylates a great number of nuclearproteins involved not only in DNA repair andgenome stabilization (e.g. p53, topoisomerases Iand II, DNA lygase III), but also in cell cycle(c-fos, DNA polymerases � and �) and differenti-ation such as TATA box binding protein and highmobility group proteins (HMGs) 1, 2, 14 and 17(D’Amours et al., 1999). It also plays a pivotalrole in DNA transcription and replication role(De Lucia et al., 1996; Simbulan-Rosenthal et al.,1996). An association of PARP with transcrip-tionally active chromatin regions was found.PARP also regulates the expression of c-Myc,protein-kinase C and DNA methyl transferasegenes (Bauer et al., 1996). However, at present, nodirect role of PARP in astrocyte differentiationwas demonstrated.
A causal relationship between ADP-ribosyla-tion and astroglial cell proliferation including his-tone H1 poly ADP-ribosylation during intenseproliferative early postnatal events (18 DIV),higher in cerebellum of rat embryos slices than incerebral cortex was shown (Spina-Purrello et al.,1990; Avola et al., 1998). A significant increase ofPARP in bFGF or IGF-I treated fetal rat brain

V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520 513
slices was also observed. Moreover, IGF-I or INStreatment resulted in a dose and time-dependentincrease in PARP activity in 18-day-old rat brainembryo slices (Avola et al., 1998). On the otherhand, mono ADP-ribosylation is likely involvedin mitochondrial metabolism, as demonstrated intestis mitochondria (Burzio et al., 1981). It wasalso suggested that ADP ribosyltransferase (AD-PRT) activation might be linked to mitochondrialCa2+ release induced by chemical oxidants(Richter and Frei, 1988).
The present study was designed to assess theeffects of the growth factors EGF, bFGF, IGF-Iand INS on nuclear PARP and mitochondrialADPRT enzymatic activities in developing (30DIV) or aging (90 and 190 DIV) primary ratastrocyte cultures. This is to clarify mitogenic ordifferentiative roles played by these neurotrophicfactors during postnatal development, maturationand aging in in vitro models.
2. Materials and methods
2.1. Astroglial cell cultures
Astroglial cell cultures were obtained from cere-bral hemispheres of newborn rats according to theprotocol detailed in earlier studies of our group(Avola et al., 1988a,b). Cultures were grown inFalcon plastic Petri dishes (35, 60 or 100 mmdiameter) at a plating density of 0.5–1×105 cells/cm2 and incubated at 37 °C in a humidified 5%CO2–95% air atmosphere. Culture medium waschanged after 7 days and then twice a week. Themorphology of this astroglial cell culture systemwas described previously (Avola et al., 1988a,b).
2.2. Immunofluorescence
Cells were fixed in 5% acetic acid in ethanol for10 min and then incubated with rabbit anti-GFAP serum (1:50) for 40 min, followed by rho-damine-labeled affinity-purified goat antibody torabbit IgG (1:40) for 40 min. Stained cultureswere mounted in 50% glycerol and phosphate-buffered saline (PBS) and examined. Immunos-taining was analyzed as reported elsewhere (Avolaet al., 1988a,b).
2.3. PARP and ADPRT enzymatic acti�ities
Astrocyte cultures were grown in serum-supple-mented medium (SSM) up to 28 or 88 or 188 DIVand then switched in serum-free medium (SFM)containing bovine serum albumin (BSA, 1 mg/ml)for a 24 h starvation period before 12 h treatmentwith EGF (10 ng/ml) or bFGF (5 ng/ml), IGF-I(10 ng/ml) or INS (10 �g/ml). Nuclei and mito-chondria were purified as previously described(Spina-Purrello et al., 1990). PARP activity wasdetermined by incubating purified nuclei (100 �g/125 �l) in a lysis buffer (10 mM Tris–HCl, pH 8,4 mM MgCl2, 0.4 mM DTT) with 2,8-H3-NAD(0.4 �Ci/ml) for 10 min in an oscillating bath at37 °C (Masmoudi et al., 1988). Reaction was thenstopped adding 2 ml ice-cold 20% TCA contain-ing 1% sodium pyrophosphate (Na2P4O7). Cellu-lar pellets were collected on Whatman filters(GF/B glass fiber) washed three times with 5%TCA in Na2P4O7 20 mM and once with a 1:1ethanol/ethylether mixture. Filters were dried at100 °C for 15 min, transferred in vials and addedwith Instagel scintillation fluid. Radioactivity wasmeasured with a Wallace �-counter and resultsexpressed as dpm/filter. For the assay of ADP-ri-bosyltransferase standard reaction mixture con-tained 100 mM Tris–HCl (pH 8) 8 mM MgCl2,0.4 mM DTT, and 100 �M [3H]NAD in a totalvolume of 125 �l. The reaction was carried out for10 min at 37 °C and stopped by addition of 10%trichloroacetic acid containing 0.02 M sodiumpyrophosphate. The precipitate was collected andwashed by filtration on Whatman GF/B glass-fiber filters. Radioactivity was measured by liquidscintillation spectrometry and results were ex-pressed as dpm/filter.
Mitochondrial total protein content was deter-mined according to Lowry et al. (1951) against astandard of BSA. Assay of ADP-ribosyltrans-ferase enzymatic activity from purified mitochon-dria was performed incubating 200 �g protein in atotal volume of 125 �l (Masmoudi et al., 1988).
2.4. Statistics
Data are expressed as means�standard errorof the mean (SEM) of two independent experi-

V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520514
ments. Each experiment consisted in the assay offive different dishes. Analysis of variance(ANOVA) was used to assess significance of dif-ferences between different treatments, takinggroup and time of treatment as independentvariables.
2.5. Chemicals
EGF, bFGF, IGF-I and INS, transferrin BSAand anti-GFAP antibodies were purchased fromSigma Chemical Co. (St Louis, MO, USA). 2,8-H3-NAD was purchased from NEN Products(Boston, MA, USA). Dulbecco’s modified Eagle’smedium (DMEM), penicillin G, streptomycin sul-fate, and fetal calf serum (FCS) were obtainedfrom GIBCO (Renfrewshire, Scotland). PlasticPetri dishes for cell cultures were purchased fromFalcon (Oxford, England), whereas rhodamine-la-beled affinity-purified goat antibody raisedagainst rabbit IgG was purchased from Kirkegardand Perry Laboratories Inc. (Faithersburg, MD,USA).
3. Results
The results of nuclear PARP activity in purifiednuclei from primary rat astrocyte cultures at 30DIV treated with IGF-I and bFGF are shown inFig. 1. PARP activity significantly increased inIGF-I- and bFGF-treated astroglial cell cultures(by about 100% for IGF-I and 140% for bFGF)compared to control untreated cultures. The ef-fects of INS or EGF or both together (INS+EGF) on PARP and ADPRT activities, in nucleiand mitochondria respectively purified from 30DIV-developing astrocyte culture, are shown inFig. 2 (panels A and B). A remarkable increase ofboth nuclear (Fig. 2, panel A) and mitochondrial(Fig. 2, panel B) enzyme activities in 30 DIVastrocyte cultures treated for 12 h with INS orEGF or both together was observed.
Nuclear PARP and mitochondrial ADPRT ac-tivities were remarkably increased in control un-treated cultures at 190 DIV, compared withcultures at 90 DIV, suggesting an active involve-ment of polyADP-ribosylation in repair of nu-
clear DNA damage and in genomic integritymaintenance during cell aging in culture (Fig. 3,panels A and B). Treatment with EGF (10 ng/ml)or INS (10 �g/ml) or bFGF (5 ng/ml) for 12 hsignificantly stimulated nuclear PARP and mito-chondrial ADPRT in 190 DIV late-aging cultures(Fig. 3, panels A and B).
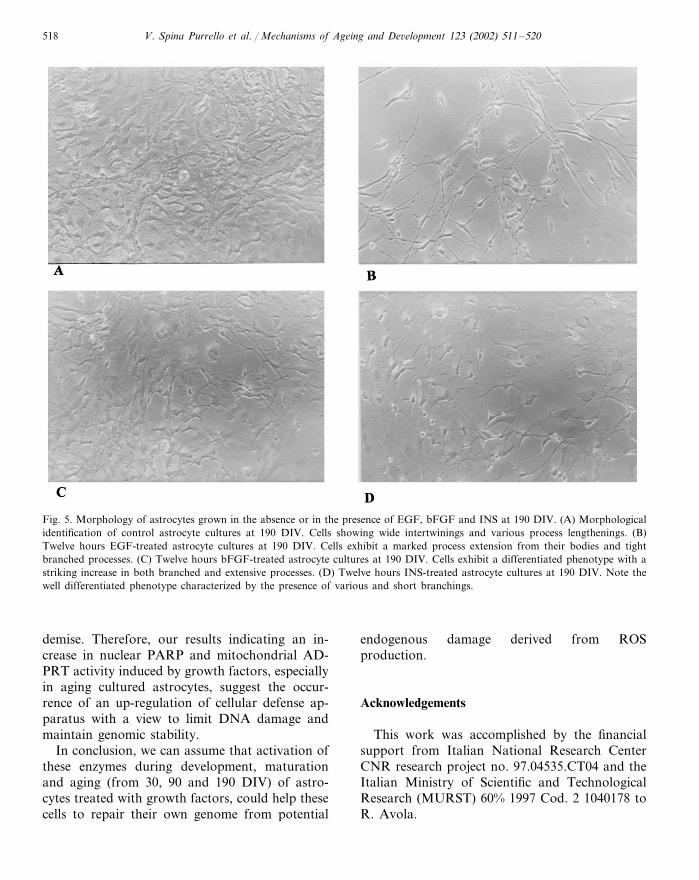
No changes of the above nuclear and mito-chondrial enzymes were found in 12 h IGF-I-treated astrocytes at 90 DIV compared to controluntreated ones (Fig. 3, panels A and B). Morpho-logical appearance of treated or untreated astro-cytes culture at 90 and 190 DIV is shown in Figs.4 and 5, respectively.
4. Discussion
The above data have shown that bFGF, IGF-I,EGF or INS, or the two last together (EGF+INS) greatly stimulate PARP and ADPRT activi-
Fig. 1. Effects of 12 h treatment with growth factors (IGF-I orbFGF) on PARP activity in nuclei purified from astrocytecultures at 30 DIV. Each value is the average�SEM of twoindependent experiments. � P�0.001 vs. controls.

V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520 515
Fig. 2. Effects of 12 h treatment of growth factors on PARP activity in nuclei (panel A) and mitochondria (panel B) from astrocytecultures at 30 DIV. Each value is the average�SEM of two independent experiments. *P�0.01 vs. controls.
ties in 30 DIV developing astroglial cell cultures.This effect can be explained by a synergistic po-tentiation of effects of these growth factors on cellproliferation and differentiation, through a crosstalk between multiple signaling cascades triggeredby them (Avola et al., 1988a,b). These mecha-nisms are known to up-regulate cell cycle proteinapparatus and aid mitogenesis and cell growth(Kazlauscas, 1994; Rudge et al., 1994).
PARP activity was found to be tightly andpreferentially associated with regions of chro-
matin that are transcriptionally active. It is alsoan integral constituent of the multiprotein DNAreplication complex (DNA synthesome) contain-ing DNA polymerase � as one of the many acces-sory proteins (Simbulan-Rosenthal et al., 1996).Moreover, it is also functionally active duringbrain aging as demonstrated by some authors(Strosznajder et al., 2000) and also in the presentstudy. In fact, in aging 190 DIV astrocyte cul-tures, we have observed a marked stimulation ofboth PARP and ADPRT activities induced by

V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520516
EGF, bFGF or INS. It is interesting to notethat bFGF apparently stimulated mainlyADPRT rather than PARP activity, which con-versely was more sensitive to EGF. This sug-gests that a preferential compartmentation oftransducing cascades, triggered by these growthfactors, may exist. Differential influences on theenzyme activities could reside not only in thesignaling cascades triggered by EGF or bFGF,but also in their redistribution and sub cellularlocalization.
Several cytosolic constituents of receptorial sig-nal transduction pathways were discovered totranslocate toward mitochondria once activated,both for survival signals such as c-Raf 1 withBcl-2 and c-Akt (Majewski et al., 1999), Raf-A(Yuryev et al., 2000), and protein kinase C �(Ruvolo et al., 1999) or for apoptotic inductionsuch as protein kinase C � (Majumder et al.,2000), the stress-activated protein kinases p38(Zhuang et al., 2000) and Jun N-terminal Kinase(JNK) (Kharbanda et al., 2000). Therefore, the
Fig. 3. Effects of 12 h treatment of growth factors on PARP activity in nuclei (panel A) and mitochondria (panel B) from agingastrocyte cultures at 90 or 190 DIV. Each value is the average�SEM of two independent experiments. *P�0.05 vs. controls at 190DIV, vs. control at 90 DIV **P�0.001.
V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520 517
Fig. 4. Cell morphology of astrocytes grown in the absence or in the presence of IGF-I at 90 DIV. (A) Morphological identificationof control untreated astrocyte cultures at 90 DIV. Note the flat polygonal shaped morphology with not branched processes. (B)Twelve hours IGF-I-treated astrocyte cultures at 90 DIV. No significant morphological modifications vs. untreated controls wereshown.
ability of the two growth factors to direct specificmessages toward the mitochondrial compartmentmay account for the above results. Another possi-ble explanation of our findings, may be that ef-fects on astrocyte PARP or ADPRT depend byspecific recruitment of fibroblast growth factorreceptor-1 (FGFR-1) docking protein FRS-2upon stimulation with bFGF (and also with NGFvia TrkA), that is unresponsive to INS and EGFand has a weak response to PDGF and IGF-I(Kaplan and Miller, 1997).
Our data are consistent with the hypothesis of akey role played by growth factors on ADP-ribosy-lation-linked events and cross talk between mito-chondria and nucleus. It is known that duringaging endogenous reactive oxygen species (ROS)production is capable of inducing both nuclearand mitochondrial DNA single- or double-strandbreaks, a typical damage involved not only incarcinogenesis but also in neurodegenerative dis-orders and tissue aging (Croteau and Bohr, 1997).Growth factor-induced activation of PARP andADPRT may therefore represent an attempt tolimit genomic damage and risk of cellular trans-formation or cell death, albeit with differentstrength exerted by the tested factors.
A protective role of PARP to ionizing radia-tion-, alkylating agents- and oxidative stress-in-duced damage was largely demonstrated (for areview see Chatterjee et al., 1999), and a role formitochondrial ADPRT in regulating calcium re-lease induced by several pro-oxidants was hypoth-esized as well (Richter and Frei, 1988). On theother hand, ATP production is impaired suppress-ing thus apoptotic program, since ATP is abso-lutely required for this event (Marton et al., 1997).The increase of mitochondrial Ca2+ release prob-ably triggers a new nitrogen- and oxygen-freeradicals production and also nuclear PARP acti-vation, as this enzyme requires Ca2+ ions foroptimal activity (Niedergang et al., 1979). Thesephenomena could result as main actors in thesuccessive cell death (Szabo, 1996). It should bealso noted that ADPRT is strongly inhibited byATP itself (Moser et al., 1983). This strengthens apossible cross talk between mitochondrial energyproduction and nuclear apoptotic program duringthe physiological aging (for a review see Cromp-ton, 1999). This metabolic evidence and the factthat PARP is cleaved and inactivated by caspasesduring apoptosis (Kaufmann et al., 1993), suggestthat cells have developed a mechanism to protectthemselves from PARP-induced switching in cell
V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520518
Fig. 5. Morphology of astrocytes grown in the absence or in the presence of EGF, bFGF and INS at 190 DIV. (A) Morphologicalidentification of control astrocyte cultures at 190 DIV. Cells showing wide intertwinings and various process lengthenings. (B)Twelve hours EGF-treated astrocyte cultures at 190 DIV. Cells exhibit a marked process extension from their bodies and tightbranched processes. (C) Twelve hours bFGF-treated astrocyte cultures at 190 DIV. Cells exhibit a differentiated phenotype with astriking increase in both branched and extensive processes. (D) Twelve hours INS-treated astrocyte cultures at 190 DIV. Note thewell differentiated phenotype characterized by the presence of various and short branchings.
demise. Therefore, our results indicating an in-crease in nuclear PARP and mitochondrial AD-PRT activity induced by growth factors, especiallyin aging cultured astrocytes, suggest the occur-rence of an up-regulation of cellular defense ap-paratus with a view to limit DNA damage andmaintain genomic stability.
In conclusion, we can assume that activation ofthese enzymes during development, maturationand aging (from 30, 90 and 190 DIV) of astro-cytes treated with growth factors, could help thesecells to repair their own genome from potential
endogenous damage derived from ROSproduction.
Acknowledgements
This work was accomplished by the financialsupport from Italian National Research CenterCNR research project no. 97.04535.CT04 and theItalian Ministry of Scientific and TechnologicalResearch (MURST) 60% 1997 Cod. 2 1040178 toR. Avola.

V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520 519
References
Avola, R., Condorelli, D.F., Surrentino, S., Turpeenoja, L.,Costa, A., Giuffrida Stella, A.M., 1988a. Effect of epider-mal growth factor and insulin on DNA, RNA and cy-toskeletal protein labeling in primary rat astroglial cellcultures. Neurochem. Res. 11, 789–800.
Avola, R., Condorelli, D.F., Turpeenoja, A., Ingrao, F.,Reale, S., Ragusa, N., Giuffrida Stella, A.M., 1988b. Ef-fect of epidermal growth factor on the labeling of thevarious species of RNA and of nuclear proteins in primaryrat astroglial cell cultures. J. Neurosci. Res. 20, 54–63.
Avola, R., Spina Purrello, V., Morale, M.C., Gallo, F.,Farinella, Z., Costa, A., Reale, S., Marletta, N., Ragusa,N., Marchetti, B., 1998. Insulin-like growth factor-I effectson ADP-ribosylation processes and interactions with glu-cocorticoids during maturation and differentiation of as-troglial cells in primary culture. In: Muller, E.E. (Ed.),IGFs in the Nervous System. Springer Verlag, Milan, pp.127–134.
Avola, R., Spina-Purrello, V., Gallo, F., Morale, M.C., Mar-letta, N., Costa, A., Tirolo, C., Testa, N., Reale, S.,Marchetti, B., 2000. Immortalized hypothalamic luteiniz-ing hormone-releasing hormone (LHRH) neurons induce afunctional switch in the growth factor responsiveness inastroglia: involvement of basic fibroblast growth factor.Int. J. Dev. Neurosci. 18, 743–763.
Bauer, P.I., Kirsten, E., Young, L.J.T., Varadi, G., Csonka,E., Buki, K.G., Mikala, G., Hu, R., Comstock, J.A.,Mendeleyev, J., Hakam, A., Kun, E., 1996. Modificationof growth related enzymatic pathways and apparent loss oftumorigenicity of a ras-transformed bovine endothelial cellline by treatment with 5-iodo-6-amino-1,2-pyrone. Int. J.Oncol. 8, 239–252.
Berridge, M.J., 1987. Inositoltrisphosphate and diacylglicerol:two interacting second messengers. Annu. Rev. Biochem.56, 159–193.
Bignami, A., Eng, L.F., Dahl, D., Uyeda, C.T., 1972. Local-ization of glial fibrillary acidic protein in astrocytes byimmunofluorescence. Brain Res. 43, 429–435.
Burzio, L.O., Saez, L., Cornejo, R., 1981. Poly(ADP-ribose)synthetase activity in rat testis mitochondria. Biochem.Biophys. Res. Commun. 103, 369–375.
Carpenter, G., Cohen, E., 1990. Epidermal growth factor. J.Biol. Chem. 265, 7709–7712.
Casper, D., Mytilineau, C., Blum, M., 1991. EGF enhancesthe survival of dopamine neurons in rat embryonic mesen-cephalon primary cell cultures. J. Neurosci. Res. 30, 372–381.
Chatterjee, S., Berger, S.J., Berger, N.A., 1999. PolyADP-ri-bose polymerase: a guardian of the genome that facilitatesDNA repair by protecting against DNA recombination.Mol. Cell. Biochem. 193, 23–30.
Crompton, M., 1999. The mitochondrial permeability transi-tion pore. Biochem. J. 341, 233–249.
Croteau, D.L., Bohr, V.A., 1997. Repair of oxidative damageto nuclear and mitochondrial DNA in mammalian cells. J.Biol. Chem. 272, 25409–25412.
Dahl, D., 1981. The vimentin-GFA protein transition in ratneuroglia cytoskeleton occurs at the time of myelination. J.Neurosci. Res. 6, 741–748.
Dahl, D., Rueger, D.C., Bignami, A., Weber, K., Osborn, M.,1981. Vimentin, the 57,000 dalton protein of fibroblastfilaments, is the major cytoskeletal component in immatureglia. Eur. J. Cell. Biol. 24, 191–196.
D’Amours, D., Desnoyers, S., D’Silva, I., Poirer, G.G., 1999.Poly (ADP-ribosyl)ation reaction in the regulation of nu-clear function. Biochem. J. 342, 249–268.
De Lucia, F., Mennella, M.R., Quesada, P.P., Farina, B.,1996. Poly (ADPribosil)ation system in transcriptionallyactive rat testis chromatine fractions. J. Cell. Biochem. 63,334–341.
Denhardt, D.T., 1996. Signal transducing protein phosphory-lation cascades mediated by Ras/Rho proteins in the mam-malian cell: the potential role for multiplex signalling.Biochem. J. 318, 729–747.
Enfors, P., Lonnerberg, P., Ayer Le Lievre, C., Persson, H.,1990. Developmental and regional expression of bFGFmRNA in the rat central nervous system. J. Neurosci. Res.27, 10–15.
Fantl, W.J., Johnson, D.E., Williams, L.T., 1993. Signallingby receptor tyrosine kinases. Annu. Rev. Biochem. 62,453–481.
Garcia-Abreu, J., Cavalcante, L.A., Silva, L.G., Moura Neto,V., 1996. The extracellular matrix of the midline andnon-midline midbrain glia: correlations with neuritegrowth-supporting abilities. Braz. J. Med. Biol. Res. 29,1179–1187.
Han, V.K., Smith, A., Myint, W., Mygard, K., Bradshaw, S.,1992. Mitogenic activity of EGF on newborn rat astroglia:interactions with IGF-I. Endocrinology 131, 1134–1142.
Homburg, S., Visochek, L., Moran, L., Dantzer, F., Priel, E.,Asculai, E., Schwartz, D., Rotter, V., Dekel, N., Cohen-Armon, M., 2000. A fast signal-induced activation of poly(ADP-ribose) polymerase: a novel downstream target ofphospholipase C. J. Cell. Biol. 150, 293–307.
Kaufmann, S.H., Desnoyers, S., Ottaviano, Y., Davidson,N.E, Poirer, G.G., 1993. Specific proteolytic cleavage ofpoly ADP-ribose polymerase: an early marker of chemio-therapy-induced apoptosis. Cancer Res. 53, 3976–3985.
Kaplan, D.R., Miller, F.D., 1997. Signal transduction by theneurotrophin receptors. Curr. Opin. Cell. Biol. 9, 213–221.
Kazlauscas, A., 1994. Receptor tyrosine kinases and theirtargets. Curr. Opin. Genet. Dev. 4, 5–14.
Kharbanda, S., Saxena, S., Yoshida, K., Pandley, P., Kaneki,M., Wang, Q., Cheng, K., Chen, Y., Campbell, A., Sudha,T., Yuan, Z., Narula, J., Weichselbaum, R., Nalin, C.,Kufe, D., 2000. Translocation of SAPK/JNK to mitochon-dria and interaction with Bcl-XL in response to DNAdamage. J. Biol. Chem. 275, 322–327.
Lowry, O.H., Rosebrought, N.J., Fau, A., Randall, L.J., 1951.Protein measurement with the Folin phenol reagent. J.Biol. Chem. 193, 265–275.
Marton, A., Mihalik, R., Bratincsak, A., Adleff, V., Petak, I.,Vegh, M., Bauer, P.I., Krajcsi, P., 1997. Apoptotic cell

V. Spina Purrello et al. / Mechanisms of Ageing and De�elopment 123 (2002) 511–520520
death induced by inhibitors of energy conservation. Bcl2inhibits apoptosis downstream of a fall of ATP levels. Eur.J. Biochem. 238, 1857–1863.
Masmoudi, A., Islam, F., Mandel, P., 1988. ADP-ribosylationof highly purified rat brain mitochondria. J. Neurochem.51, 188–193.
Majewski, M., Niebowska-Skorska, M., Salomoni, P., Slupi-anek, A., Reiss, K., Trotta, R., Calabretta, B., Skorski, T.,1999. Activation of mitochondrial Raf-1 is involved in theantiapoptotic effect of Akt. Cancer Res. 59, 2815–2819.
Majumder, P.K., Pandley, P., Sun, X., Cheng, K., Datta, R.,Saxena, S., Kharbanda, S., Kufe, D., 2000. Mitochondrialtranslocation of protein kinase C delta in phorbol ester-in-duced cytochrome c release and apoptosis. J. Biol. Chem.275, 21793–21796.
Morrison, R.S., Saneto, R.P., de Vellis, J., 1982. Developmen-tal expression of rat brain mitogens for cultured astrocytes.J. Neurosci. Res. 8, 435–442.
Morrison, R.S., Sharma, A., deVellis, J., Bradshaw, R.A.,1986. Basic fibroblast growth factor supports the survivalof cerebral cortical neurons in culture. Proc. Natl. Acad.Sci. USA 83, 7537–7541.
Moser, B., Wimterhalter, K.H., Richter, C., 1983. Purificationand properties of a mitochondrial NAD+ glycohydrolase.Arch. Biochem. Biophys. 224, 358–364.
Niedergang, H., Okazaki, H., Mandel, P., 1979. Properties ofpurified calf thymus poly (adenosine diphosphate ribose)polymerase. Eur. J. Biochem. 102, 43–57.
Richter, C., Frei, B., 1988. Ca2+ release from mitochondriainduced by pro-oxidants. Free Radic. Biol. Med. 4, 365–375.
Rotwein, P., Burgess, S.K., Millbrandt, J.D., Krause, J.E.,1988. Differential expression of insulin-like growth factorgenes in tar central nervous system. Proc. Natl. Acad. Sci.USA 85, 265–269.
Rudge, J.S., Li, Y., Pasnikowski, E.M., Mattson, K., Pan, L.,Yancopoulos, G.D., Niegand, S.J., Lindsay, K.M., Ip, N.,1994. Neurotrophic factor receptors and their signalling
transduction capabilities in rat astrocytes. Eur. J. Neurosci.6, 693–705.
Ruvolo, P.P., Deng, X., Carr, B.K., May, W.S., 1999. Afunctional role for mitochondrial protein kinase C� inBcl-2 phosphorylation and suppression of apoptosis. J.Biol. Chem. 274, 25436–25442.
Simbulan-Rosenthal, C.M., Rosenthal, D.S., Hilz, H., Hickey,R., Malkas, L, Applegren, N., Wu, Y., Bers, G., Smulson,M., 1996. The expression of poly (ADP-ribose) polymeraseduring differentiation-linked DNA replication reveals thatit is a component of the multiprotein DNA replicationcomplex. Biochemistry 35, 11622–11633.
Spina-Purrello, V., Avola, R., Condorelli, D.F., Nicoletti,V.G., Insirello, L., Reale, S., Costa, A., Ragusa, N., Giuf-frida Stella, A.M., 1990. ADP-ribosylation of proteins inbrain regions of rats during postnatal development. Int. J.Neurosci. 8, 167–174.
Strosznajder, J.B., Jesco, H., Strosznajder, R.P., 2000. Age-re-lated alteration of poly(ADP-ribose) polymerase in differ-ent part of the brain. Acta Biochim. Pol. 47 (2), 331–337.
Szabo, C., 1996. DNA strand breakage and activation ofpolyADP-ribosyltransferase: a cytotoxic pathway triggeredby peroxynitrite. Free Radic. Biol. Med. 21, 855–869.
Torres-Aleman, I., Naftolin, F., Robbins, R.J., 1990. Trophiceffects of insulin-like growth factor I on fetal rat hypotha-lamic cells in culture. Neuroscience 35, 601–608.
Yuryev, A., Ono, M., Goff, S.A., Macaluso, F., Wennogle,L.P., 2000. Isoform-specific localization of A-Raf in mito-chondria. Mol. Cell. Biol. 20, 4870–4878.
Walicke, P., Cowan, W.M., Ueno, N., Baird, A., Guillelmin,R., 1986. Fibroblast growth factor promotes survival ofdissociated hippocampal neurons and enhances neurite ex-tension. Proc. Natl. Acad. Sci. USA 83, 3012–3016.
Zhuang, S., Demirs, J.T., Kockevar, I.E., 2000. p38 mitogen-activated protein kinase mediates Bid cleavage, mitochon-drial dysfunction and caspase 3 activation during apoptosisinduced by singlet oxygen but not by hydrogen peroxide. J.Biol. Chem. 275, 25933–25948.