interaction among sediment anaerobiosis, nitrogen uptake and photosynthesis of spartina alterniflora
TRANSCRIPT

! HYSIOLOGIA PLANTARUM 74: 561-565. Copenhagen 1988
Interaction among sediment anaerobiosis, nitrogen uptake andphotosynthesis of Spartina aUemiflora
S. R. Pezeshki, R. D. DeLaune and C. W. Lindau
Pezeshki, S. R., DeLaune, R. D. and Lindau, C. W. 1988. Interaction amongsediment anaerobiosis, nitrogen uptake and photosynthesis of Spartina alterniflora. -Physiol. Plant. 74: 561-565.
Spartina alterniflora Lois, plants were grown under controlled sediment-pH-redoxconditions. Uptake of added "NH4-nitrogen and subsequent photosynthetic activityunder different redox conditions in the plant root rhizosphere were measured. Datafor total plant nitrogen and ' N concentration indicated that nitrogen allocation wasnot altered by anaerobic conditions of the sediment. However, average net photosyn-thesis was reduced by up to 35% for plants under anaerobiosis. The results indicatethat anaerobiosis in the root rhizosphere, rather than limiting nitrogen uptake,influences photosynthesis and growth of 5. alterniflora under anaerobic conditions.
Key words - Anaerobiosis, leaf conductance, nitrogen uptake, nutrient allocation,photosynthesis, sediment, Spartina alterniflora.
S. R. Pezeshki {corresponding author), et al.. Lab. for Wetland Soils and Sediments,Center for Wetland Resources, Louisiana State Univ., Baton Rouge, LA 70803, USA.
Introduction
Spartina alterniflora Lois, is a dominant C4 grass grow-ing in brackish and saline marshes in Louisiana. Re-markable gradients in height and productivity occurwithin a short distance across these stands. A taller,greener, more vigorous and more productive form isfound in the slightly elevated areas bordering streamsand a shorter, less productive form is found in lowerinland areas (DeLaune et al. 1983). Similar differencesin productivity have been reported for this species inother coastal regions (Valiela et al. 1978, Mendelssohn1979a).
The difference in productivity of S. alterniflora hasbeen attributed to many environmental features includ-ing salinity (Nestler 1977, Smart and Barko 1978, Va-liela et al, 1978), ion toxicity (Linthurst 1979), excesssulfide concentration (DeLaune et al. 1983), sedimentredox potential (Howes et al. 1981, DeLaune et al,1983) and nitrogen availability (Gallagher 1975, Valielaet al, 1978, Mendelssohn 1979a, Cavalieri and Huang1981). Anaerobiosis in the rhizosphere is a dominantfactor in wetland areas, causing physiological stressesA'hich can result in limitation of active uptake of essen-
tial elements such as nitrogen. The nitrogen status inturn can affect photosynthetic activity in plants. Highorganic nitrogen content in leaves is necessary for CO2fixation (Makino et al. 1984, Hak and Natr 1987a, Sageand Pearcy 1987), A close relationship between leafnitrogen content and activity of ribulose 1,5-bisphos-phate carboxylase/oxygenase (Rubisco, EC 4.1,1,39),an important enzyme in photosynthesis has been dem-onstrated by Friedrich and Huffaker (1980), Evans(1983), Makino et al. (1984), Hak and Natr (1987b) andSeemann et al, (1987),
Linthurst (1979) found that uptake of several nutri-ents by 5. alterniflora was correlated with soil aeration.Several workers have emphasized the potential limitingeffect of reduced nitrogen uptake on growth of S. al-terniflora under anaerobiosis (Gallagher 1975, Mendels-sohn 1979b, Morris 1984). Despite these reports, verylittle published evidence exists to show that anaero-biosis alone can result in nitrogen deficiency in S. al-terniflora exposed to long-term anaerobic conditions.The present study was conducted to examine the effectsof low sdediment redox potential on nitrogen uptakeand allocation in S. alterniflora. The hypotheses testedwere (1) nitrogen uptake is inhibited by anaerobiosis,
iecieved 17 May, 1988; revised 17 August, 1988
'hysiol. Planl. 74, 1988 561

(2) nitrogen allocation patterns within the plant arealtered as a result of anaerobic conditions, and (3) lownitrogen in leaf tissue, resulting from changes in thenitrogen allocation pattern, reduces net photosynthesisin 5. alterniflora.
Abbreviations - E^, redox potential in mV; PPFD, photosyn-thetic phot fl d i t i l ^ ' R b i ib
, pthetic photon flux density inl 1 5 b i h h
; pys~'; Rubisco, ribu-p nsity in nmol m s
lose- 1,5-bisphosphate carboxylase/oxygenase.
Materials and methods
Plant materiat
Spartina alterniflora Lois, were obtained from a saltmarsh stand located in Barataria Basin, LA, USA. Theplants were initially grown in sand in a greenhouse andfertilized with half-strength Hoagland solution contain-ing ammonium nitrate (Johnson et al. 1957). The fertil-ization was on a biweekly schedule in which 50 ml ofsolution were added to each 4-1 pot. Plantlets regener-ated from rhizomes of these plants; that is, newly germi-nated plantlets and the attached rhizome and roots wereused for different experiments.
Soil solution
Sediment was collected from a salt marsh area. Air-plane Lake in Barataria Basin, LA. In the laboratory,225 g of dry sediment and 1600 ml of half-strengthHoagland solution (with nitrogen component elimi-nated) were mixed in 2-1 desiccator bases and werecontinuously stirred with a magnetic stirrer.
Experimentat procedures
Plantlets 3-5 cm in height and uniform in size andappearance were placed through holes in plexiglassplates designed to fit on top of the desiccator bases (3-4plantlets/each desiccator base). Plants mounted in aplexiglass plate were transferred onto the desiccatorbase containing the soil suspension and were grownunder controlled root rhizosphere E^ conditions inside agrowth chamber. No shoot die-back was noted for thetransplanted plantlets. The growth chamber was main-tained at days of 14 h (25°C, 70% relative humidity) andnights of 10 h (20°C, 85% relative humidity). Photosyn-thetic photon flux density (PPFD) was 800-1000 M-molm ' s"' at the top of the plants. The PPFD was reducedto 200 |xmol m - s~' during the first and last hour of eachphotoperiod.
The system for controlling the redox potential in thesediment in which the plants were grown was modifiedfrom that described by Reddy et al. (1976). It consistedof a desiccator plexiglass plate (containing plants asdescribed earlier) and a small impeller attached to aminiature motor to keep the soil in suspension. To avoidroot damage by the motor shaft, the shaft was shieldedusing 10 mm diameter plexiglass tubes attached to the
upper plate. The system also contained a meter relay,platinum electrodes, a calomel-half cell connected tothe soil suspension with a saturated KCl-agar salt bridgeas well as other associated equipment. The desiccatortop was covered with a plexiglass plate and sealed withblack plastic rubber. Two platinum electrodes, one sahbridge, serum cap and glass tubings were fitted into theholes in the plexiglass plate. Nitrogen gas was bubbledcontinuously through the soil suspension at a low flowrate (15-25 ml min ") to remove oxygen and to promoteanaerobic conditions in the suspension. The reductionprocesses in the soil suspension caused the E^ to de-crease. When the E^ set on the meter relay was reached,the air pump was activated to pump air (oxygen) intothe soil suspension to prevent the E,, from falling belowthe set value.
At the beginning of each experiment, the soil E^ was>400 mV for all desiccators. E^ levels then were main-tained for control (aerated) system while E^ levels werereduced in other systems. To impose extremely negativeEh conditions (< -200 mV), 2 g sucrose were added in0.5 g increments to each desiccator. After addition ofsucrose and continuous N, injection, the low E^ (anaer-obic) systems were maintained at < -200 mV through-out the experiment. When E^ became stable for allsystems, i.e. >400 for high E,, (aerated) systems (Treat-ment I) and < -200 mV for anaerobic systems (Treat-ment II), 0.724 g of •5NH4CI (99.3 atom %) was addedto each desiccator. "N-labelled NH^ was used since thisis the dominant inorganic nitrogen form found in anaer-obic marsh sediments (DeLaume et al. 1983). Through-out the experiment, deionized water was added twiceper week to compensate for transpirational waterlosses.
Measurement of leaf conductance and net photosyn-thesis was conducted during the last week of each exper-iment when leaf blades were well developed. Net photo-synthesis was measured on intact attached leaves ofplants under different treatment. An open gas-exchangesystem was used for photosynthetic measurements. Thissystem included a multicuvette system supplied withtemperature and humidity-conditioned air. The temper-ature in each cuvette was controlled by water-cooledconvection fins, and air was stirred by an internal fan.An infrared gas analyser (IRGA), ADC Model LCA-2was used for differential CO2 measurements. Other im-portant plant and environmental factors which wererecorded throughout the experiment included: (1)PPFD levels measured with LI-COR Ouantum Sensorsand LI-185 light meter, and (2) leaf and air temper-atures measured with fine-wire copper constantan ther-mocouples. A multichannel recorder was used to recordthese variables throughout the experiment.
Both leaf conductance and net photosynthesis werecalculated per unit leaf area (single surface) determinedwith a surface area meter (Model SI701, SKYE In-struments, Inc., Buckingham, PA, USA). Leaf con-ductance was calculated from leaf transpiration rates
562 Physiol. Plant. 74. 1988
letermined from the water vapor mol-fraction differ-ence across the leaf chamber. Net photosynthetic rates,vere calculated from the flow rate of air and from theCOj partial pressure differences between incoming andoutgoing air through the chamber as outlined by Caem-merer and Farquhar (1981).
At the conclusion of each experiment, i.e. 28 daysafter each experiment was initiated, plant nitrogen con-tent was measured by conducting tissue analyses onharvested biomass components (foliages and roots).Plant samples were dried at 60°C for 48 h and ground topass through a 250 [im mesh sieve. Nitrogen uptake andallocation in plant samples were determined for total Nby a semimicro-Kjeldahl method and analyzed for '""Ncontent using a Dupont 21-621 isotope ratio mass spec-trometer (Buresh et al. 1982, Hauck 1982). The entireexperiment was replicated twice using similar materialsand techniques.
Redox levels
The level of sediment E^ was maintained close to pre-setvalues (-220 ± 25 mV) in both treatments.
ResultsNitrogen content and allocation
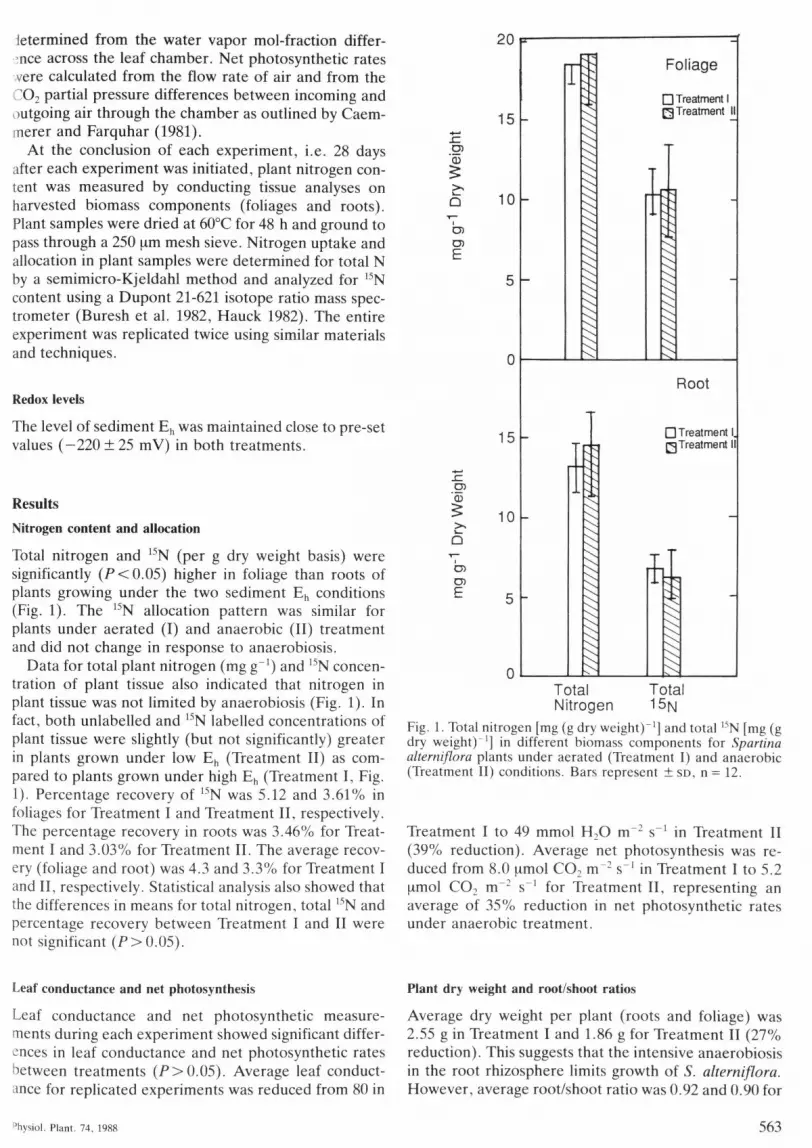
Total nitrogen and '""N (per g dry weight basis) weresignificantly (P<0.05) higher in foliage than roots ofplants growing under the two sediment E,, conditions(Fig. 1). The '" N allocation pattern was similar forplants under aerated (I) and anaerobic (II) treatmentand did not change in response to anaerobiosis.
Data for total plant nitrogen (mg g"') and "N concen-tration of plant tissue also indicated that nitrogen inplant tissue was not limited by anaerobiosis (Fig. 1). Infact, both unlabelled and "N labelled concentrations ofplant tissue were slightly (but not significantly) greaterin plants grown under low E^ (Treatment II) as com-pared to plants grown under high E,, (Treatment I, Fig.1). Percentage recovery of '"'N was 5.12 and 3.61% infoliages for Treatment I and Treatment II, respectively.The percentage recovery in roots was 3.46% for Treat-ment I and 3.03% for Treatment II. The average recov-ery (foliage and root) was 4.3 and 3.3% for Treatment Iand II, respectively. Statistical analysis also showed thatthe differences in means for total nitrogen, total ''N andpercentage recovery between Treatment I and II werenot significant (P>0.05).
Q
I
D)
20
15 -
10 -
5 -
'03
b)
15 -
10 -
5 -
IT
11 r
1
1 f
Foliage
]3 Treatment 13 Treatment II
\\\
\~\" \"^\• ~ \
Root
• Treatment 1.Q Treatment 11
1TotalNitrogen
Total
Fig. 1. Total nitrogen [mg (g dry weight) '] and total "N [mg (gdry weight)^'] in different biomass components for Spartinaalterniflora plants under aerated (Treatment I) and anaerobic(Treatment II) conditions. Bars represent ± SD, n = 12.
Treatment I to 49 mmol HjO m ^ s ' in Treatment II(39% reduction). Average net photosynthesis was re-duced from 8.0 \imo\ CO, m ' s~' in Treatment I to 5.2|j,mol CO2 m"- s"' for Treatment II, representing anaverage of 35% reduction in net photosynthetic ratesunder anaerobic treatment.
Leaf conductance and net photosynthesis
Leaf conductance and net photosynthetic measure-ments during each experiment showed significant differ-ences in leaf conductance and net photosynthetic ratesbetween treatments (P>0.05). Average leaf conduct-ance for replicated experiments was reduced from 80 in
Plant dry weight and root/shoot ratios
Average dry weight per plant (roots and foliage) was2.55 g in Treatment I and 1.86 g for Treatment II (27%reduction). This suggests that the intensive anaerobiosisin the root rhizosphere limits growth of S. alterniflora.However, average root/shoot ratio was 0.92 and 0.90 for
"hysiol. Plant. 74, 1988 563

Treatment I and II, respectively. Based on dry weightdata and tissue nitrogen concentrations, total nitrogenuptake was less under anaerobic treatment.
Discussion
Many workers have implicated nitrogen as a major fac-tor contributing to the restricted growth and low pro-ductivity of short 5, alterniflora stands (Gallagher 1975,Valiela et al, 1978, Cavalieri and Huang 1981). Howev-er, as pointed out by DeLaune et al. (1983), the conclu-sions were primarily based on indirect evidence includ-ing increases in growth of the short form of S. alterni-flora in response to addition of nitrogen fertilizer suchas those reported by Patrick and DeLaune (1976) andCavalieri (1983), There is direct evidence indicatingadverse effects of low E , (anaerobiosis) on rates ofnitrogen uptake in short-term laboratory experiments.For example, Morris and Dacey (1984) found that nitro-gen uptake by S. alterniflora was 4 times greater forroots under high E^ (aerobic) sediment compared toroots under anaerobic conditions. However, the samestudy showed that plants continued to absorb NH^ fromanaerobic sediments at a reduced rate. This finding wasattributed to the ability of 5. alterniflora to transportoxygen from aerial parts to the roots despite the lowoxygen concentration in the rhizosphere. Previous stud-ies have shown that the oxygen diffusion system pro-vides a potential advantage which may enhance nutrientuptake from anaerobic sediments (Teal and Kanwisher1966, Gleason and Dunn 1982 and references therein).
There was no reduction in foliage and root nitrogenconcentration of S. alterniflora grown under anaerobicconditions. In fact, the nitrogen concentration of planttissues was slightly higher in plants grown under anaero-bic conditions compared to aerobic plants. Althoughprevious studies have shown that nitrogen uptake in 5,alterniflora is reduced by anaerobiosis, the nitrogenconcentration of plant tissue as shown in this experi-ment was not changed under anaerobic conditions.These findings are in agreement with results of Lin-thurst (1979) who showed that nitrogen was not a signif-icant factor in the observed differences in productivityof S. alterniflora under different anaerobic treatments.
Net photosynthetic rates were lowest for plants grownunder anaerobiosis. The 27% reduction in growth ob-served under anaerobic treatment may have reflectedthe decline in photosynthetic activity. The significantreductions in net photosynthesis resulted from severalphysiological effects of anaerobiosis. For example, thesubstantial decline in leaf conductance found in thepresent study is a reflection of partial stomatal closurein response to anaerobiosis. Similar responses havebeen reported for numerous species (Bradford andYang 1981). Partial stomatal closure can impose diffu-sional limitation of photosynthesis (Pezeshki and Sund-strom 1988). The diffusional limitation may impose lim-itations for gas exchange, including restriction of ox-
ygen transport to the roots. Despite the existence of anextensive aerenchyma system for oxygen diffusion tcthe roots from aerial parts in S. alterniflora, the systemmay not supply sufficient oxygen to the roots undercertain conditions where there is a large oxygen demandin sediment. The possibility of limited capacity for rootoxygenation in this species was pointed out by Mendels-sohn and McKee (1987), Photosynthetic processes mayalso be adversely affected by metabolic consequences ofanaerobiosis (Bradford 1983a,b).
Under field conditions, Linthurst and Seneca (1980,1981) and DeLaune et al. (1983) reported decreasedgrowth in 5, alterniflora in response to low E^. There isincreasing evidence to suggest that nitrogen is unlikelyto be the growth-limiting factor in short forms of S.alterniflora. High NH4-N concentrations have beenfound in sediment waters in short 5. alterniflora standsin North Carolina (Mendelssohn 1979b) and in GulfCoast areas (Casselman et al, 1981, DeLaune et al.1983). In all these cases, the concentrations of nitrogenin these stands were higher than those of adjacentstands where tall forms were growing. In the field,reduction in E^ is accompanied by changes in severalsoil parameters, including an increase in free sulfide(Linthurst and Seneca 1980). The primary factor limit-ing growth of 5, alterniflora in Louisiana salt marsh isthe reduced sediment E^ which can result in excess ionavailability and potentially cause ion toxicity. Underreduced E^ conditions, excess sulfide is a major growth-limiting factor at the inland sites (DeLaune et al, 1983,Pezeshki et al. 1988),
Acknowledgement- Funding for this research was provided bythe National Science Foundation (Grant #BSR 8414006),
References
Bradford, K. J, t983a. Effects of soil flooding on leaf gasexchange of tomato plants, - Plant Physiol, 73: 475^79,
- 1983b. Involvement of plant growth substances in the alter-ation of leaf gas exchange of flooded tomato plants, - PlantPhysiol, 73: 480-483,
- & Yang, S, F, 1981, Physiological responses of plants towaterlogging. - HortScience 16: 25-30,
Buresh, R, J,, Austin, E, R, & Craswell, E, T, 1982, Analyt-ical methods in ' N research, - Fert, Res, 3: 37-62,
Casselman, M. E,, Patrick, W, H,, Jr, & DeLaune, R, D.1981, Nitrogen fixation in a Gulf Coast salt marsh. - SoilSci, Soc, Am, J, 45: 51-56,
Cavalieri, A, J, 1983. Proline and glycinebetaine accumulationby Spartina alterniflora in response to NaCl and nitrogen ina controlled environment. - Oecologia 57: 20-24,
- & Huang, A. H. C, 1981, Accumulation of prohne andglycinebetaine in Spartina alterniflora in response to NaCIand nitrogen in the marsh, - Oecologia 49: 224-228,
DeLaune, R. D., Smith, C. J. & Patrick, W, H,, Jr, 1983.Relationship of marsh evaluation, redox potential, and sul-fide to Spartina alterniflora productivity, - Soil Sci, Soc.Am, J, 47: 930-935,
Evans, J, R, 1983, Nitrogen and photosynthesis in the flag lea!of wheat {Triticum aestivum L,), - Plant Physiol. 72: 297-302,
Friedrich, J, W, & Huffaker, R, C, 1980, Photosynthesis, learesistance, and ribulose 1,5-bisphosphate earboxylase deg
564 Physiol. Planl. 74, 198!

radation in senescing barley leaves; - Plant Physiol. 65:1103-1107.
( allagher, J. L. 1975. Effect of an ammonium pulse on growthand elemental composition of natural stands of Spartinaalterniflora and Juncus roemerianus. - Am. J. Bot. 62:644-648.
Gleason, M. L. & Dunn, E. L. 1982. Effects of hypoxia on rootand shoot respiration of Spartina alterniflora. - In EstuarineComparisons (V. S. Kennedy, ed.), pp. 243-253. AcademicPress, New York, N.Y. ISBN 0-12-404070-5.
Hak, R. & Natr, L. 1987a. Effect of nitrogen starvation andrecovery on gas exchange characteristics of young barleyleaves. - Photosynthetica 21: 9-14.
- & Natr, L. 1987b. Effect of nitrogen starvation and recov-ery on carbon fluxes in photosynthetic carbon reductionand oxidation cycles in barley leaves. - Photosynthetica 21:15-22.
Hauck, R. D. 1982. Nitrogen-isotope-ratio analysis. - In Meth-ods of Soil Analysis, Part 2, 2nd Ed. (A. L. Page, R. H.Miller and D. R. Keeney, eds), pp. 735-779. Am. Soc.Agron., Madison, WI. ISBN 0-89118-072-9.
Howes, B. L., Howarth, R. W., Teal, J. M. & Valiela, I. 1981.Oxidation-reduction potentials in a salt marsh: spatial pat-terns and interactions with primary production. - Limnol.Qceanogr. 26: 350-360.
Johnson, C. M., Stout, P R., Broyer, T. C. & Carlton, A. B.1957. Comparative chlorine requirements of different plantspecies. - Plant Soil. 8: 337-353.
Linthurst, R. A. 1979. The effect of aeration on the growth ofSpartina alterniflora. - Am. J. Bot. 66: 685-691.
- & Seneca, E. D. 1980. The effects of water and drainagepotential on the Spartina alterniflora substrate complex inNorth Carolina salt marsh. - Est. Coast. Mar. Sci. 11:41-42.
- & Seneca, E. D. 1981. Aeration, nitrogen and salinity asdeterminants of Spartina alterniflora growth response. -Estuaries 4: 53-63.
Makino, A., Mae, T. & Qhira, A. K. 1984. Effect of nitrogen,phosphorus or potasium on the photosynthesis rate andribulose-1,5-bisphosphate carboxylase content in riceleaves during expansion. - Soil Sci. Plant Nutr. 30: 63-70.
Mendelssohn, I. A. 1979a. Nitrogen metabolism in the heightforms of Spartina alterniflora in North Carolina. - Ecology60: 574-584.
- 1979b. Influence of nitrogen level, form, and applicationmethod on the growth response of Spartina alterniflora inNorth Carolina. Estuaries 2: 106-111.
- & McKee, K. L. 1987. Root metabolic response of Spartinaalterniflora to hypoxia. - In Plant Life in Aquatic andAmphibious Habitats (R. M. M. Crawford, ed.), pp. 239-253. British Ecol. Soc. Special Sym. Proc. Publ. No. 5.
Morris, J. T. 1984. Effects of oxygen and salinity on ammo-nium uptake by Spartina alterniflora and Spartina patens. -J. Exp. Mar. Biol. Ecol. 70: 87-98.
- & Dacey, J. W. 1984. Effects of Q2 on ammonium uptakeand root respiration by Spartina alterniflora. - Am. J. Bot.71: 979-985.
Nestler, J. 1977. Interstitial salinity as a cause of ecophenicvariation in Spartina alterniflora. - Est. Coast. Mar. Sci. 5:707-714.
Patrick, W. H., Jr. & DeLaune, R. D. 1976. Nitrogen andphosphorous utilization by Spartina alterniflora in a saltmarsh in Barataria Bay, Louisiana. - Est. Coast. Mar. Sci.4: 59-64.
Pezeshki, S. R. & Sundstrom, F. J. 1988. Effect of soil anaero-biosis on photosynthesis of Capsicum annuum L. - Sci.Hort. 35: 27-35.
- , Pan, S. Z., DeLaune, R. D. & Patrick, W. H., Jr. 1988.Sulfide-induced toxicity: inhibition of carbon assimilationin Spartina alterniflora. - Photosynthetica 22 (In press).
Reddy, C. N., Jugsujinda, A. & Patrick, W. H., Jr. 1976.System for growing plants under controlled redox poten-tial-pH conditions. - Agron. J. 68: 987-989.
Sage, R. F. & Pearcy, R. W. 1987. The nitrogen use efficiencyof C3 and C4 plants. II. Leaf nitrogen effects on the gasexchange characteristics of Chenopodium album and Am-aranthus retroflexus. - Plant Physiol. 84: 959-963.
Seeman, J. R., Sharkey, T. D., Wang, J. & Qsmond, C. B.1987. Environmental effects on photosynthesis, nitrogen-use efficiency, and metabolite pools in leaves of sun andshade plants. - Plant Physiol. 84: 796-802.
Smart, R. M. & Barko, J. W. 1978. Influence of sedimentsalinity and nutrients in the physiological ecology of se-lected salt marsh plants. - Est. Coast. Mar. Sci. 7: 487-495.
Teal, J. M. & Kanwisher, W. 1966. Gas transport in the marshgrass, Spartina alterniflora- J. Exp. Bot. 17: 355-361.
Valiela, I., Teal, J. M. & Deuser, W. G. 1978. The nature ofgrowth forms in the salt marsh grass Spartina alterniflora. -Am. Nat. 112: 461-470.
von Caemmerer, S. & Farquhar, G. D. 1981. Some relation-ships between the biochemistry of photosynthesis and thegas exchange of leaves. - Planta 153: 376-387.
dited by P. Nissen
siol. Plant. 74, 1988 565
