ecophysiology of xerophytic and halophytic vegetation of a coastal alluvial plain in northern...
TRANSCRIPT

New Phytol. (1989), Ill, 245-251
Ecophysiology of xerophytic and halophytic vegetation of a coastal alluvial plain In northern Venezuela
I I . Cactaceae
By U. LUTTGE1:JI:, E. MEDINA2 , W. J. CRAM3 , H. S. J. LEE1,3, M. POPP4,5 AND J. A. C. SMITH4,6
1 Institut fur Botanik, Technische Hochschule Darmstadt, D-6100 Darmstadt, FRG 2 Centro de Ecologia y Ciencias Ambientales, Instituto Venezolano de Investigaciones Cientificas, Caracas 1020-A, Venezuela 3 Department of Biology, The University, Newcastle upon Tyne, NEI 7RU, UK 4 Institut fur Pjianzenphysiologie der Universitiit, A-I091 Wien, Austria 5 Institut fur Angewandte Botanik, Westfiilische Wilhelms- Universitiit, D-4400 Munster, FRG 6 Department of Botany, University of Edinburgh, Edinburgh, EH93JH, UK
(Received 8 March 1988; accepted 22 July 1988)
SUMMARY
In an ecophysiological field investigation of plant communities of vegetation islands of an alluvial plain in northern Venezuela the members of the Cactaceae, Pereskiaguamacho, and a columnar ceroid cactus, Subpilosocereus ottonis, were studied. The alluvial plain was flooded by fresh water in the rainy season in November IDecember 1985 and was dry and saline in the dry season in Marchi April 1986. The highly succulent leaves of P. guamacho were shed in the dry season. They performed Ca photosynthesis in the wet season and did not show signs of considerable salt accumulation. P. guamacho avoids stress due to drought and salinity by leaf-shedding. The columnar stem succulent ceroids are salt-excluding plants with crassulacean acid metabolism (CAM). Rapid die back and regeneration of absorptive roots, water parenchyma and CAM, with the possibility of nocturnal stomatal closure and CO2 recycling, are traits of adaptive value under fluctuating conditions between rainy and dry seasons on the alluvial plain.
Key words: Cactaceae, crassulacean acid metabolism (CAM), drought, Pereskia guamacho, Subpilosocereus ottonis, salinity.
INTRODUCTION
From studies of CO2 uptake patterns in leafy Cactaceae and considering the relevant literature, Nobel & Hartsock (1987) arrived at a comprehensive survey of drought responses in the three subfamilies of the Cactaceae. The Pereskioideae are droughtdeciduous plants. Leaves are formed in rainy and shed in dry seasons. Thus, these plants are largely avoiders of drought stress. The gas exchange pattern is the Ca type. However, small CAM-like day-night changes of tissue acidity in leaves of Pereskia species
"To whom all correspondence should be addressed.
submitted to drought, indicate that the enzymology and other factors necessary for CAM are expressed notwithstanding the absence of CAM-like gas exchange. In the Opuntioideae there are also leafy species, where drought induces C 3 to CAM shifts or young cladodes may show day time stomatal opening with Ca-type CO2 fixation. The stem succulent forms of the Cereoideae nearly all have constitutive CAM.
In the alluvial plain of Chichiriviche members of all of the three subfamilies of the Cactaceae are found, namely Pereskia guamacho Weber (Pereskioideae), Opuntia wentiana Britton & Rose and other Opuntia spp. (Opuntioideae) and columnar

246 U. Liittge and others
ceroid forms of the species Subpilosocereus oltonis Backeberg and some Acanthocereus leJ.ragonus (L.) Humik (Cereoideae). They occur mainly in the deciduous woodland (vegetation unit 5 of Medina et al., 1989). The columnar ceroid forms may also prevail on the vegetation islands (vegetation unit 4 of Medina el al., 1989) and some A. tetragonus and Opuntia spp. may he found hoth there and in the grassland (vegetation unit 3 of Medina et al., 1989).
When small vegetation islands in the Cienega el Ostional (such as that shown in figure 6 of Medina el
01., 1989) with Sesuvium portulacaslrum L., a few rosettes of Bromelia humilis jacq. and some plants of Opuntia and Subpilosocereus ouonis are flooded in the rainy season, the cacti grow with their roots and stem bases in 'fresh' water. For cacti this is a rather extraordinary habitat. The large salinity changes between rainy season and dry season of 37- 203 mequiv. Cl- kg- I air-dried soil observed in the upper layers of the alluvial plain in the Batis- Sesuvium community (Table I of Medina et al., 1989) apply here. This poses a remarkable ecophysiological challenge to seedlings of cacti for establishment and survival on the primordial islands. Conversely, on the older islands, where the top soil is elevated above the alluvial plain, smaller changes of salinity between the seasons are found, i.e. 11- 36 mequiv. CI- kg- 1 air dried soil in the rainy season and 46- 114 mequiv. CI kg- 1 in the dry season. In the woodland community the range is from 11-28 or 44 mequiv. CI- kg- I (Table I in Medina et al., 1989). This is the situation for the larger cacti and for establishment of seedlings on older islands.
In the present study we concentrated on S. ottonis, since these plants often contribute considerably to the total biomass of the island vegetation (see also Figure 2 a below). Autecological investigations of these plants may be valuable for synecological
understanding of the dynamics of the islands. However, a few measurements were also made on P. guamacho for comparison.
:vtATERIALS AND METHODS
Samples for analyses, taken in the field in the Cienega el Ostional of Chichiriviche, were immediately sealed in plastic vials and stored on ice or dry ice in the dark. Within a maximum of 8 h, samples were transferred to the deep-freeze compartment of a refrigerator. For measurements of cell-sap osmotic pressure, 17, and titratable acidity the sap was pressed out of freeze-thawed material using a garlic press. An Osmomat 030 freezing-point osmometer (Gonotec, GmbH, West Berlin) was used to obtain 17, expressed in osmol kg- 1 fresh weight, or converted to Pa using the van't Hoff equation. Titratable acidity was determined by titration with 2'5 mol m - 3 NaOH using phenolphthalein as an indicator as described in detail by Smith et al. (1986).
For analyses of inorganic ions, material was kept on dry ice after sampling in the field and subsequently freeze-dried. This also gave comparisons of fresh weights (f. wt) and dry weights (d. wt). Hotwater extracts prepared from the freeze-dried as well as tissue saps, were analysed for cations and CI- by atomic absorption spectrophotometry (PerkinElmer) and Buchi chloridometer, respectively. The data of the hot-water extracts, calculated on a plant water basis, were in good agreement with the values obtained from the tissue saps. Thus hot-water extracts and tissue saps obtained from the same material were treated as parallel samples.
Due to the mucilage in stem tissue extracts of the S. ottonis only limited amounts of sap could be obtained, sufficient for the determination of 7T but not for analyses of inorganic ions.
Xylem tension was measured using an SME
Figure 1. Pereskia guamacho with succulent leaves in the rainy season (a), and the same plant without leaves in the dry season (b).

Ecophysiology of xerophytic and halophytic vegetation in Venez uela 247
pressure chamber (Plant Water Status Console Model 3005: Soilmoisture Equipment Corp ., Santa Barbara, Califo rnia, USA) as described by Smith et
al. ( 1986). Values presented in the text and tables are means
± SD (number of samples) unless stated otherwise .
RESULTS (1) Leaf tissue sap osmotic pressures and ion contents of leaves of Pereskia guamacho
The leaves of the drought-deciduous Pereskia guamacho in the C ienega el Ostional (Fig.l) are very succulent , the d . wt: f. wt ratio was about 1: 25. Leaf-sap osm otic pressure, 7T, in the rainy season was about 1·0 MPa. Contents of CI-, Na+ and K + were 120-164 mol m-" 64-76 mol m - 3 and 13- 22 mol m - 3 , respecti vely. Leaves of P. guamacho did not perform CAM. They had titratable acidities of 12 mol m -3 both at dawn and at dusk in the rainy season .
(2) Stem tissue sap osmotic pressures and xylem tensiollS of Su bpi losocereus ottonis
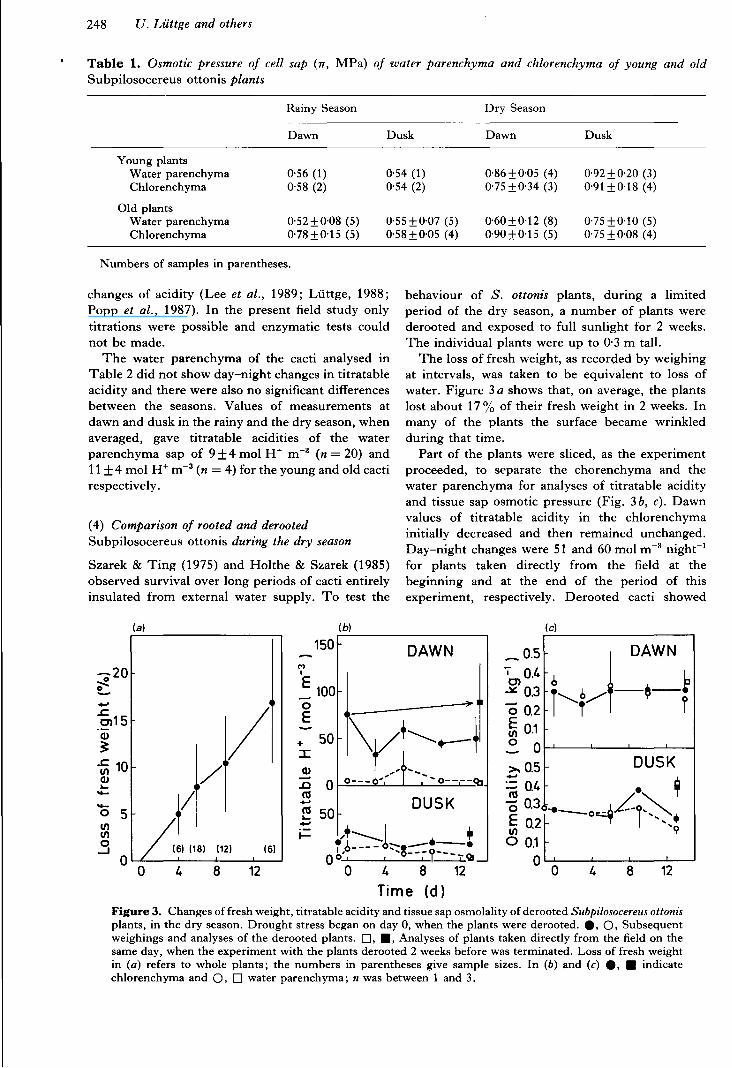
Stem tissue sap osmotic pressures were measured in the water parenchyma and in the chlorenchyma of young plants of S. ottonis, which were up to 0'1 m tall, and in old plants several metres tall (Fig. 2, Table t) . Measurements were made at dawn and dusk during the rain y and the dry seasons. In no case was osmotic pressure high enough to suggest any accumu lation o f salt by these cacti .
In the older plants, but not in the young ones, osmotic pressures were somewhat greater in the chlo renchyma as compared to the water parenchyma. I n the young plants, osmotic pressures on average were higher in the dry season by a factor of t ·55. This is thought to be due to lower water content rather than accumulation of salt (see below and Fig. 3). I n the large plants this seasonal change of 7T was much smalle r , i.e. by a factor of 1·15- 1·36 depending on the time of day and the tissue (Table 1).
Occasional values for xylem tensions were obtained from a few small plants of S. otton;s mounted with thei r cut stem bases in the SME pressure chamber (see Luttge & Nobel, 1984, for the performance of such measurements). Values recorded in the rainy season were 0·26 ±0· t6 MPa (n = 4) and in the dry season 0'52 ±0·25 Mra (n = 2). These values were not corrected for xylem sap osmotic pressure.
(3) Day- night changes of tit ratable acidity in stem tissue sap of S ubpilosoce reus ottonis
For technical reasons measurements of CO 2 exchange could not be performed on the cacti in the
Figure 2. Cacti of the Cienega el Ostional. (a) Large plants of Subpilosocerells ottollis on a vegetat ion island with Sporobo/lls v irgillicus in the foreground, Opllfltia rve fltiafla, Bromelia Illlmilis and shrubs. (b) Sm all plant showing the root system. (c) A large fallen plant (6'5 m tall) with its root system partially exposed. (d) Young plant with adventitious roots above a necrotic part of the s tem.
present fi eld project. However, titratable acidity was determined (Table 2). It is normall y assumed that day- night changes in tit ratable protons are related to changes in malic ac id levels by a factor of 2: t . This is la rgely based on experiments with Kalanchoe' spp . (Luttge, Ball & Tromballa, 1975 ; Luttge & Ball , 1979, 1980; Luttge, Smith & Marigo, 1982) where changes of malate levels and titratabl e acidity were measured simultaneousl y. It does not apply to all conditions and all species, when citric acid in addition to malic acid is involved in day- night
248 U. Liittge and others
Table 1. Osmotic pressure of cell sap (7T, MPa) of water parenchyma and chlorenchyma of young and old Subpilosocereus ottonis plants
Rainy Season Dry Season
Dawn Dusk Dawn Dusk
Young plants Water parenchyma 0·56 (1) 0·54 (1) 0·86±0·05 (4) 0·92 ± 0·20 (3) Chlorenchyma 0·58 (2) 0·54 (2) 0·75 ±0·34 (3) 0·91 ±0·18 (4)
Old plants Water parenchyma 0·52±0·08 (5) 0·55 ±0·07 (5) 0·60±0·12 (8) 0·75 ±0·1O (5) Chlorenchyma 0·78±0·15 (5) 0·58±0·05 (4) 0·90±0·15 (5) 0·75 ±0·08 (4)
Numbers of samples in parentheses.
changes of acidity (Lee et al., 1989; Liittge, 1988; Popp et al., 1987). In the present field study only tit rations were possible and enzymatic tests could not be made.
The water parenchyma of the cacti analysed in Table 2 did not show day-night changes in tit ratable acidity and there were also no significant differences between the seasons. Values of measurements at dawn and dusk in the rainy and the dry season, when averaged, gave tit ratable acidities of the water parenchyma sap of 9±4 mol H+ m-3 (n = 20) and 11 ± 4 mol H+ m -3 (n = 4) for the young and old cacti respectively.
(4) Comparison of rooted and derooted Subpilosocereus ottonis during the dry season
Szarek & Ting (1975) and Holthe & Szarek (1985) observed survival over long periods of cacti entirely insulated from external water supply. To test the
(a) (b)
150
-20 (') I
~ : 100 e.--
/ 0
~15 E Q) 50 ~ +
:::r: .J::.
behaviour of S. ottonis plants, during a limited period of the dry season, a number of plants were derooted and exposed to full sunlight for 2 weeks. The individual plants were up to 0·3 m tall.
The loss of fresh weight, as recorded by weighing at intervals, was taken to be equivalent to loss of water. Figure 3 a shows that, on average, the plants lost about 17 % of their fresh weight in 2 weeks. In many of the plants the surface became wrinkled during that time.
Part of the plants were sliced, as the experiment proceeded, to separate the chorenchyma and the water parenchyma for analyses of titratable acidity and tissue sap osmotic pressure (Fig. 3 b, c). Dawn values of tit ratable acidity in the chlorenchyma initially decreased and then remained unchanged. Day-night changes were 51 and 60 mol m-3 nighC1
for plants taken directly from the field at the beginning and at the end of the period of this experiment, respectively. Derooted cacti showed
(e)
DAWN _0.5 DAWN ., 0.4
r • f-$-~ 0)
~ 0.3 00.2 '1'",-,- r E III 0.1 0 0
DUSK , " 10 / >. 0.5 III Q) Q)
.0 0 o---t,.,,' ........ o----~ = 04 ~ ~ ...... ...... 0 III III 0
.....J
r CIS CIS • e - DUSK <5 0.30.e_o=~/-i~ 5 CIS 50 ~ E 02 ",+ -/:11181 i= .,+'--1 • + III . ~
(12) (6) 16---- ::"'-8- -e o 0.1 00', ! .. --~~---iQl 0 0
0 4 8 12 0 4 8 12 0 4 8 12 Time (d)
Figure 3. Changes of fresh weight, titratable acidity and tissue sap osmolality of derooted Subpilosocereus ottonis plants, in the dry season. Drought stress began on day 0, when the plants were derooted .• , 0, Subsequent weighings and analyses of the derooted plants. D, ., Analyses of plants taken directly from the field on the same day, when the experiment with the plants derooted 2 weeks before was terminated. Loss of fresh weight in (a) refers to whole plants; the numbers in parentheses give sample sizes. In (b) and (c) ., • indicate chlorenchyma and 0, D water parenchyma; n was between 1 and 3.

Ecophysiology of xerophytic and halophytic vegetation in Venezuela 249
Table 2. Day-night changes of titratable acidity (mol H+ m-3) In the chlorenchyma of young and old
Subpilosocereus ottonis plants
Rainy Season Dry Season
Dawn Dusk Dawn Dusk
Young plants 214 (1) 54 (2) 82±40 (6) 27±7 (9) ~H+ 160 55
Old plants 124±56 (4) 30± 10 (4) 121±38 (8) 32±8 (9) ~H+ 94
~H+, dawn values minus dusk values.
day-night changes of 40 mol m-3 nighC1 on the 6th day, 25 mol m-3 nighC1 on the 9th day and 33 mol m-3 nighC1 on the 14th day. Thus the decline of acid accumulation from both external and internal sources of CO 2 , which cannot be distinguished here, was 35-45 % in 2 weeks.
DISCUSSION
The absence of clear day-night changes of tit ratable acidity in the leaves of Pereskia guamacho in the rainy season agrees with observations of Nobel & Hartsock (1986)' that, under well-watered conditions, species of the subfamily Pereskioideae behave like C 3 plants and all the net shoot CO2 uptake occurs via the leaves and during the daytime. Pereskia does have a latent potential for CAM. It is important to note that day-night oscillations with nocturnal increases of tit ratable acidity of up to 100 mmol H+ kg-1 f. wt were observed in P. grandifolia and P. aculeata while stomata were closed (Rayder & Ting, .1981): This can be explained by internal CO2 recycling, i.e. nocturnal malic acid synthesis by dark fixation of respiratory CO2 ,
During the present project all plants of P. guamacho had shed their leaves at the peak of the dry season so that no analyses could be made. A more detailed investigation of P. guamacho in the field in the state of Falcon in Venezuela was presented by Diaz (1983) and Diaz & Medina (1984) and shows that under conditions of slight drought stress, before and after the major rainy season, leaves of P. guamacho may perform weak CAM with some nocturnal recycling of respiratory CO2 in addition to C 3-type photosynthesis, while this drought-deciduous plant sheds leaves in the major dry season.
In the woodland community, where P. guamacho occurs in the Cienega el Ostional, topsoil salinity is only 11-28 mequiv. kg-1 air-dried soil. Cell-sap osmotic pressure of P. guamacho leaves in the rainy season was only 1 to ! of that of the small perennial halophytes Sesuvium portulacastrum and 'Batis maritima but similar to that of Portulaca rubricaulis which also sheds its leaves in the dry season (Liittge et al., 1989). Cl- contents are! to land! to l compared
18
89
with the small perennial halophytes above and the shrub-like halophytes (Conocarpus erectus and Avicennia germinans, Smith et al., 1989), respectively. For Na+ the comparisons are l to ! and 1 to l, respectively. Hence, the drought-deciduous P. guamacho avoids high levels of salt in its leaves by leaf-shedding, and there are no traits of halophilism in this shrub or tree-like plant.
Young plants of S. ottonis, growing in the sand on primordial islands of the alluvial plain were subject to increase in soil salinity in the dry season. The observation that none of these plants accumulated salt is consistent with results of an earlier study in the laboratory where roots died off due to the impact of salinity (Nobel et al., 1984).
Such observations partially confirm Walter's hypothesis on the mechanism of survival of the cacti in salinas during the dry season (Walter & Breckle, 1984). Walter suggested that the absorptive roots of the cacti die off during the dry season, so that plants are insulated from soil salinity. Indeed the root system, even of young cacti in the sand plain, shows an extensive vertical development of thick tap roots while the thinner absorptive roots die (Fig. 2b). Moreover, young cacti can be found in the field, with new adventitious ,roots .formed aboye necrotic parts of the stem (Fig. 2d).
However, Walter's hypothesis needs to be modified and extended. On the larger vegetation islands, and in the community of cacti and trees, salinity stress in the top soil is much milder. Figure 2c shows the upper part of the soil profile dug near a 6· 5 m tall fallen S. ottonis, with its strongly branched root system partially visible, at the fringe of a cactus and tree community (see also closed circles in Fig. 10 of Medina et al., 1989). The major tap root of the S. ottonis plant seen in Figure 2c was only 0·45 m long, i.e. it did not extend below the level of 150 mequiv. Cl- kg-1 air-dried soil. These tap roots ~ever extend into the deeper layer of the substratum, where the salt load is high during the dry season. Hence, a great part of the cactus population of the Cienega el Ostional may not be subject to much salinity stress. This is the case for most of the taller individuals of S. ottonis where only in very exceptional dry seasons,
ANP III

250 U. Liittge and others
like that of 1983 (see Fig. 11 in Medina et ai., 1989), the salt exclusion mechanism, with absorptive roots dying and large tap roots protected by suberization, actually operates.
For those cacti which overcome the dry season without functional absorptive roots, i.e. without the opportunity to take up water during the dry season even when there is some occasional rain, CAM is an additional important trait in the survival strategy. The plants may keep stomata closed both day and night and perform intracellular recycling of CO2 ,
evolved during respiration, into malate (Szarek, Johnson & Ting, 1973; Nobel, 1977; Ting & Rayder, 1982; Holthe & Szarek, 1985; Brulfert et ai., 1987). It is known that the proportion of such recycling of CO2 in CAM plants may vary, and that this is modulated by environmental conditions. Recycling increases with increasing drought stress (Osmond, Nott & Firth, 1979; Sale & Neales, 1980; Martin, Christensen & Strain, 1981 ; Nobel & Hartsock, 1983, 1984; Nobel et ai., 1984; Griffiths et ai., 1986; Winter, Schr6ppel-Meier & Caldwell, 1986; Lee et ai., 1989; Griffiths et ai., 1989). In the experiments of Nobel et ai. (1984) with young seedlings of Cereus vaiidus mentioned above, recycling was '" 20 % in the controls and '" 50 % in NaCl-treated plants. Recycling, of course, reduces carbon gain. In the case of continuous stomatal closure, without uptake of atmospheric CO2 , recycling alone allows no productivity at all but prevents loss of carbon.
The data of Table 2 and Figure 3 show that the dawn-dusk differences in titratable acidity in the chlorenchyma of young cacti decreased considerably after the transition from the rainy to the dry season. No such decline was observed in older cacti. It is important to recall in this context that stem sap osmotic pressure, 7T, increased in the young cacti much more than in the old cacti (Table 1). In the chlorenchyma of the old plants osmotic pressures were higher at dawn than at dusk, a phenomenon frequently observed in relation to nocturnal accumulation of malic acid in CAM (Liittge & Ball, 1977; Liittge & Nobel, 1984; Smith & Liittge, 1985; Smith et ai., 1986). The large water reserves in the water parenchyma of the large bodies of the older cacti and their rooting in less saline soil, allowing replenishment of water reserves via the roots during occasional rain in the dry season, mitigate the effects on stem sap osmotic pressure.
The results of Figure 3 confirm that the water parenchyma of S. ottonis plants does not participate in the acid rhythm of CAM. Up to now this has generally been accepted for water parenchyma of CAM plants (Kluge et aI., 1979) although Earnshaw, Carver & Charlton (1987) recently found small day-night variations of tit ratable acidity in water parenchyma of some CAM plants.
It is evidently extremely difficult for young cacti to become established under the environmental con-
ditions of the Cienega el Ostional. Water supply is ample for seed germination in the rainy season. Young seedlings were frequently found in the tanks of Bromeiia humiiis. Few seedlings survive, however. Possible increase of wind-blown soil around the bases would allow salt damaged plants to reroot as shown in Figure 2d. Jordan & Nobel (1979, 1982) have shown that young CAM plants, like species of Agave and cacti, need a certain biomass to withstand drought periods and that seedling establishment is infrequent and depends on the intensity of the rainy and dry seasons.
ACKNOWLEDGEMENT
This work was supported by the Deutsche Forschungsgemeinschaft. For detailed acknowledgements see the accompanying paper by Medina et at. (1989).
REFERENCES
BRULFERT, J., KLUGE, M., GUERRIER, D. & QUEIROZ, O. (1987). Characterization of carbon metabolism in Opuntia ficus-indica Mill. exhibiting the idling mode of crassulacean acid metabolism. Planta 170, 92-98.
DIAZ, M. (1983). Estudios fisioecologicos de 4 especies de cactaceas en condiciones naturales. Trabajo de Grado de Magister Scientiarum en Biologia mencion Ecologia. Centro de Estudios Avanzados, IVIC, Caracas.
DIAZ, M. & MEDINA, E. (1984). Actividad CAM de cactaceas en condiciones naturales. In: Eco-Fisiologia de Plantas CAM (Ed. by E. Medina), pp. 98-113. CIET (lVIC-UNESCO), Caracas.
EARNSHAW, M. J., CARVER, K A. & CHARLTON, W. A. (1987). Leaf anatomy, water relations and crassulacean acid metabolism in the chlorenchyma and colourless internal water-storage tissue of Carpobrotus edulis and Senecio mandraliscae. Planta 170, 421-432.
GRIFFITHS, H., LUTTGE, U., STIMMEL, K.-H., CROOK, C. E., GRIFFITHS, N. M. & SMITH, J. A. C. (1986). Comparative ecophysiology of CAM and Ca bromeliads. III. Environmental influences on CO. assimilation and transpiration. Plant, Cell and Environment 9, 385-393.
GRIFFITHS, H., SMITH, J. A. C., LUTTGE, U., Popp, M., CRAM, W. J., DIAZ, M., LEE, H. S. J., MEDINA, E., SCHAFER, C. & STIMMEL, K-H. (1989). Ecophysiology of xerophytic and halophytic vegetation of a coastal alluvial plain in northern Venezuela. IV. Tillandsia fiexuosa Sw. and Schomburgkia humboldtiana Reichb., epiphytic CAM plants. New Phytologist 111, 273-282.
HOLTHE, P. A. & SZAREK, S. R. (1985). Physiological potential for survival of propagules of crassulacean acid metabolism species. Plant Physiology 79, 219-224.
JORDAN, P. W. & NOBEL, P. S. (1979). Infrequent establishment of seedlings of Agave deserti (Agavaceae) in the northwestern Sonoran desert. American Journal of Botany 66, 1079-1084.
JORDAN, P. W. & NOBEL, P. S. (1982). Height distributions of two species of cacti in relation to rainfall, seedling establishment, and growth. Botanical Gazette 143, 511-517.
KLUGE, M., KNAPP, I., KRAMER, D., SCHWERTNER, I. & RITTER, H. (1979). Crassulacean Acid Metabolism (CAM) in leaves of Aloe arborescens Mill. Comparative studies of the carbon metabolism of chlorenchym and central hydrenchym. Planta 145, 357-363.
LEE, H. S. J., LUTTGE, U., MEDINA, E., SMITH, J. A. C., CRAM, W. J., DIAZ, M., GRIFFITHS, H., Popp, M., SCHAFER, C., STIMMEL, K-H. & THONKE, B. (1989). Ecophysiology of xerophytic and halophytic vegetation of a co~stal alluvial plain in northern Venezuela. III. Bromelia humilis Jacq., a terrestrial CAM bromeliad. New Phytologist 111, 253-271.
LUTTGE, U. (1988). Day-night changes of citric-acid levels in crassulacean acid metabolism: Phenomenon and ecophysiological significance. Plant, Cell and En~'ironment 11, 445-45t.

Ecophysiology of xerophytic and halophytic vegetation in Venezuela 251
LUTTGE, U. & BALL, E. (1977). Water relation parameters of the CAM plant Kalanchoe' daigremontiana in relation to diurnal malate oscillations. Oecologia 31, 85-94.
LUTTGE, U. & BALL, E. (1979). Electrochemical investigation of active malic acid transport at the tonoplast into the vacuoles of the CAM plant Kalanchoe' daigremontiana. Journal of Membrane Biology 47, 401-422.
LUTTGE, U. & BALL, E. (1980). 2H+: 1 malate2- stoichiometry during Crassulacean Acid Metabolism is unaffected by lipophilic cations. Plant, Cell and Environment 3, 195-200.
LUTTGE, U., BALL, E. & TROMBALLA, H.-W. (1975). Potassium independence of osmoregulated oscillations of malate2- levels in the cells of CAM-leaves. Biochemie und Physiologie der Pflanzen 167, 267-283.
LUTTGE, U. & NOBEL, P. S. (1984). Day-night variations in malate concentration, osmotic pressure, and hydrostatic pressure in Cereus validus. Plant Physiology 75, 804-807.
LUTTGE, U., Popp, M., MEDINA, E., CRAM, W. J., DIAZ M., GRIFFITHS, H., LEE, H. S. ]., SCHAFER, C., SMITH, ]. A. C. & STIMMEL, K-H. (1989). Ecophysiology of xerophytic and halophytic vegetation of a coastal alluvial plain in northern Venezuela. V. The Batis maritima-Sesuvium portulacastrum vegetation unit. New Phytologist 111, 283-291.
LUTTGE, U., SMITH, J. A. C. & MARIGO G. (1982). Membrane transport, osmoregulation, and the control of CAM. In: Crassulacean Acid Metabolism (Ed. by I. P. Ting & M. Gibbs), pp. 69-91. American Society of Plant Physiologists, Rockville, Maryland.
MARTIN, C. E., CHRISTENSEN, N. L. & STRAIN, B. R. (1981). Seasonal patterns of growth, tissue acid fluctuations and 14C02 uptake in the crassulacean acid metabolism epiphyte Tillandsia usneoides L. (Spanish moss). Oecologia 49, 322-328.
MEDINA, E., CRAM, W. J., LEE, H. S. J., LUTTGE, U., Popp, M., SMITH, ]. A. C. & DIAZ, M. (1989). Ecophysiology of xerophytic and halophytic vegetation of a coastal alluvial plain in northern Venezuela. I. Site description and plant communities. New Phytologist 111, 233-243.
NOBEL, P. S. (1977). Water relations and photosynthesis of a barrel cactus, Ferocactus acanthodes, in the Colorado desert. Oecologia 27, 117-133.
NOBEL, P. S. & HARTSOCK, T. L. (1983). Relationships between photosynthetically active radiation, nocturnal acid accumulation, and CO2 uptake from a crassulacean acid metabolism plant, Opuntia ficus-indica. Plant Physiology 71, 71-75.
NOBEL, P. S. & HARTSOCK, T. L. (1984). Physiological responses of Opuntia ficus-indica to growth temperature. Physiologia Plantarum 60,98-105.
NOBEL, P. S. & HARTSOCK, T. L. (1986). Leaf and stem CO2 uptake in three subfamilies of the Cactaceae. Plant Physiology 80,913-917.
NOBEL, P. S. & HARTSOCK, T. L. (1987). Drought-induced shifts in daily CO2 uptake patterns for leafy cacti. Physiologia Plantarum 70, 114-118.
NOBEL, P. S., LUTTGE, U., HEUER, S. & BALL, E. (1984). Influence of applied NaCI on crassulacean acid metabolism and ionic levels in a cactus, Cereus validus. Plant Physiology 75,799-803.
OSMOND, C. B., NOTT, D. L. & FIRTH, P. M. (1979). Carbon assimilation patterns and growth of the introduced CAM plant Opuntia inermis in Eastern Australia. Oecologia 40, 331-350.
Popp, M., KRAMER, D., LEE, H., DIAZ, M., ZIEGLER, H. & LUTTGE, U. (1987). Crassulacean acid metabolism in tropical dicotyledonous trees of the genus Clusia. Trees 1, 238-247.
RAYDER, L. & TING, I. P. (1981). Carbon metabolism in two species of Pereskia (Cactaceae). Plant Physiology 68, 139-142.
SALE, P. J. M. & NEALES, T. F. (1980). Carbon dioxide assimilation by pineapple plants. Ananas comosus (L.) Merr. I. Effects of daily irradiance. Australian Journal of Plant Physiology 7, 363-373.
SMITH, ]. A. C., GRIFFITHS, H., LUTTGE, U., CROOK, C. E., GRIFFITHS, N. M. & STIMMEL, K-H. (1986). Comparative ecophysiology of CAM and Ca bromeliads. IV. Plant water relations. Plant, Cell and Environment 9, 395-410.
SMITH, J. A. C. & LUTTGE, U. (1985). Day-night changes in leaf water relations associated with the rhythm of crassulacean acid metabolism in Kalanchoe daigremontiana. Planta 163, 272-283.
SMITH, J. A. C., Popp, M., LUTTGE, U., CRAM, W.]., DIAZ, M., GRIFFITHS, H., LEE, H. S. J., MEDINA, E., SCHAFER, C., STIMMEL, K-H. & THONKE, B. (1989). Ecophysiology of xerophytic and halophytic vegetation of a coastal alluvial plain in northern Venezuela. VI. Water relations and gas exchange of mangroves. New Phytologist 111, 293-307.
SZAREK, S. R., JOHNSON, H. B. & TING, I. P. (1973). Drought adaptation in Opuntia basilaris. Significance of recycling carbon through crassulacean acid metabolism. Plant Physiology 52, 539-541.
SZAREK, S. R. & TING, I. P. (1975). Physiological responses to rainfall in Opuntia basilaris (Cactaceae). American Journal of Botany 62, 602-609.
TING, I. P. & RAYDER, L. (1982). Regulation of Ca to CAM shifts. In: Crassulacean Acid Metabolism (Ed. by I. P. Ting & M. Gibbs), pp. 193-207. American Society of Plant Physiologists, Rockville, Maryland.
WALTER, H. & BRECKLE, S. W. (1984). Okologie der Erde. Band 2. Spezielle Okologie der tropischen und subtropischen Zone. G. Fischer, Stuttgart.
WINTER, K, SCHROPPEL-MEIER, G. & CALDWELL, M. M. (1986). Respiratory CO2 as carbon source for nocturnal acid synthesis at high temperatures in three species exhibiting crassulacean acid metabolism. Plant Physiology 8, 390-394.
18-2