ontogenesis of leptin receptor in rat leydig cells
TRANSCRIPT

1199
BIOLOGY OF REPRODUCTION 68, 1199–1207 (2003)Published online before print 30 October 2002.DOI 10.1095/biolreprod.102.007831
Ontogenesis of Leptin Receptor in Rat Leydig Cells1
Massimiliano Caprio,3,4 Elisa Fabbrini,5 Giulia Ricci,6 Sabrina Basciani,5 Lucio Gnessi,5 Mario Arizzi,5Anna R. Carta,7 Massimo U. De Martino,4 Andrea M. Isidori,4 Giovanni V. Frajese,3 and Andrea Fabbri2,3
Cattedra di Endocrinologia,3 Dipartimento di Medicina Interna, Universita ‘‘Tor Vergata,’’ 00133 Rome, ItalyCattedra di Andrologia,4 Dipartimento di Fisiopatologia Medica, Universita ‘‘La Sapienza,’’ 00161 Rome, ItalyCattedra di Medicina Interna,5 Dipartimento di Fisiopatologia Medica, Universita di Roma ‘‘La Sapienza,’’00161 Rome, ItalyDipartimento di Medicina Sperimentale,6 Laboratorio di Istologia ed Embriologia, Seconda Universita di Napoli,80138 Naples, ItalyDipartimento di Tossicologia,7 Universita di Cagliari, 09124 Cagliari, Italy
ABSTRACT
There are still many controversies about the role of leptin inreproductive function and sexual development. We recentlydemonstrated that leptin receptors are expressed in rodent Ley-dig cells and that leptin has inhibitory effects on hCG-stimulatedtestosterone production by adult rat Leydig cells in culture. Inthis study, we evaluated the expression of leptin receptor (Ob-R) in rat testes from gestational to adult age in comparison withthe pattern of expression of relaxin-like factor (RLF), a specificmarker of Leydig cell differentiation status. Immunohistochem-ical analysis showed that, in prenatal life, Ob-R immunoreactiv-ity was absent at early embryonic ages (E14.5) and appeared ata late embryonic age (E19.5); in postnatal life, immunoreactivitywas evident only after sexual maturation (35-, 60-, and 90-daysold), whereas it was absent in testes from sexually immature rats(7-, 14-, and 21-days old). Immunoreaction was always confinedto Leydig cells and no signal of Ob-R was detected within thetubules. The pattern of expression of Ob-R during testicular de-velopment was similar with that of RLF immunoreactivity, whichwas present in mature fetal as well as adult-type Leydig cells. Incontrast with the findings in the testis, in the hypothalamus, theimmunohistochemical pattern of Ob-R was very similar betweenpre- and postpubertal life. Reverse transcription-polymerasechain reaction studies showed that Ob-R expression was presentin embryonic, prepubertal, and adult rat testes; semiquantitativeanalysis showed that mRNA levels were much higher in late ver-sus early embryonic testes, as well as in mature adults versussexually immature testes, with a gradual increase from youngerto older ages. Functional studies showed that, while leptin (150ng/ml) significantly inhibited hCG-stimulated testosterone pro-duction in adult rat Leydig cells (46% reduction; P . 0.01), itdid not modify prepubertal rat Leydig cells steroidogenic func-tion in vitro. In conclusion, we showed that, in rat testis, Ob-Rexpression is characteristic of mature Leydig cells (fetal andadult type) and it is functional in adult but not prepubertal life.
1Presented in part at the 84th annual meeting of the Endocrine Society,San Francisco, CA, 2002, P3-59. This work was supported by a grant fromthe Universities of Rome ‘‘La Sapienza’’ and ‘‘Tor Vergata,’’ Departmentof Medical Pathophysiology, and Department of Internal Medicine (Pro-getti 60%).2Correspondence: Andrea Fabbri, Endocrinology Unit, Department of In-ternal Medicine, University Tor Vergata, Via di Tor Vergata 135, 00133Rome, Italy. FAX: 39 06 72596663; e-mail: a [email protected]
Received: 28 May 2002.First decision: 14 June 2002.Accepted: 16 October 2002.Q 2003 by the Society for the Study of Reproduction, Inc.ISSN: 0006-3363. http://www.biolreprod.org
leptin, leptin receptor, Leydig cells, testis, testosterone
INTRODUCTION
Leptin is the hormonal link between energy stores andseveral vital functions, such as food intake, energy homeo-stasis, and reproductive function [1–3]. Such a wide rangeof biological actions is exerted through a receptor that be-longs to the cytokine receptor family [4]. There are severalisoforms of leptin receptor (Ob-Ra to Ob-Re), which derivefrom alternative splicing of Ob-R mRNA [5]. All Ob-Risoforms have an identical ligand binding domain but differat the C-terminus, and only Ob-Rb contains both proteindomains capable of signaling through the Jak-Stat pathway[6]. The major site of action of leptin is the hypothalamus,where the concentration of the functional isoform of Ob-Ris maximal [7]; however, leptin receptors have been re-cently identified in several endocrine peripheral tissues,such as ovary [8–10], adrenals [11], pancreas [12], and tes-tis [13]. We recently found that the functional isoform ofleptin receptor is present in isolated rodent Leydig cells andshowed that leptin has a direct inhibitory effect on hCG-induced testosterone production by purified adult rat Leydigcells in culture. Furthermore, leptin, upon binding with itsreceptor, was able to magnify hCG-induced intracellularcAMP production in the studied system [14]. Similar find-ings were obtained by other authors with rodent and bovineovarian [8, 15] and adrenal cells [11, 16], where leptin hasbeen shown to significantly inhibit the FSH-stimulated es-tradiol and ACTH-stimulated cortisol production, respec-tively.
It is known that animal [17] and human [18] models ofleptin resistance and deficiency show a severe impairmentof the reproductive function. Homozygote mutations of lep-tin receptor (db/db mutants) have low levels of circulatingtestosterone, fail to undergo normal sexual maturation, andare infertile. These alterations are due to the absence ofleptin stimulatory actions on GnRH neurons in the hypo-thalamus and a consequent hypogonadotropic hypogonad-ism [19]. In men, it has been shown that leptin levels riseby ;50% before the onset of puberty and decrease to base-line after its initiation [20], whereas in male monkeys, noc-turnal leptin levels rise about 10–20 days before the pre-pubertal increase in pulsatile LH release [21]. These find-ings follow previous negative studies [22] and suggest thatmaturation of the HPG axis requires the integrity of theleptin-leptin receptor system. It is likely that, in the hypo-thalamus, leptin concentrations above a minimal threshold

1200 CAPRIO ET AL.
are necessary to trigger sexual maturation and maintain re-productive function [19].
In the present study, we investigated the developmentalpattern of Ob-R gene expression and protein localization inrat testis from embryonic life to adulthood to gain moreinsight into the delicate cross talk between adipose tissueand testicular function during the entire life span. Our aimwas to study whether critical events such as Leydig celldifferentiation, birth, sexual maturation, and aging may af-fect leptin receptor expression and lead to significant mod-ifications in the interaction between leptin and Leydig cellfunction. In order to better characterize Leydig cell speci-ficity and differentiation status, we used an anti-relaxin likefactor (RLF) antibody as a control in the immunohisto-chemical studies because RLF is considered to be a specificmarker for the mature fetal as well as the adult Leydig cells[23, 24]. Finally, the effects of leptin on hCG-stimulatedtestosterone and intracellular cAMP production by Leydigcells isolated from prepubertal and adult rat testes werestudied.
MATERIALS AND METHODS
Animals and Tissues PreparationMale Sprague-Dawley rats (at least three per age group) (Charles River
Laboratories, Wilmington, MA) of different embryonic (E14.5 and E19.5)and postnatal ages (7-, 14-, 21-, 35-, 60-, 90-day-old rats) were used. Theanimals were provided with a standard laboratory diet and normal lighthours (14L:10D) and temperature (21–228C) and were killed by oxygendeprivation. At early and late gestational ages (see Results), pregnant ratswere killed and fetuses were removed. For the determination of the ageof the embryos, the morning after vaginal plug formation was consideredas Day 0.5 of embryonic development. Testes were rapidly removed andfixed overnight in Bouin solution for immunohistochemical experiments,frozen in liquid nitrogen for mRNA extraction, or placed in ice-cold PBSand decapsulated for collagenase digestion and subsequent Leydig cellspurification and extraction. In separate experiments, sexually immaturejuvenile (21-day-old) and adult (60-day-old) male rats [25] were anesthe-tized with chloral hydrate and perfused transcardially with PBS followedby 4% paraformaldehyde in PBS, and brains were extracted after decapi-tation.
ImmunohistochemistryBouin-fixed testes were washed twice in PBS, dehydrated, treated with
xylene, embedded in paraffin, and sectioned at a thickness of 5 mm. Sec-tions were dewaxed, hydrated, and rinsed with PBS. Endogenous peroxi-dases were blocked by incubation with 3% hydrogen peroxide in distilledwater for 20 min at room temperature. Leydig cells were detected usinga polyclonal anti-rat RLF antibody raised in rabbit (generously providedby Professor Richard Ivell, Institute of Hormone and Fertility Research,Hamburg, Germany) at 1:1000 dilution. The primary antibody against lep-tin receptor (rabbit anti-human Ob-R polyclonal antibody [H-300; SantaCruz Biotechnology, Santa Cruz, CA]), specific for all the isoforms of thereceptor, was used at 1:150 dilution, and sections were incubated overnightat 48C. The following steps were performed according to the manufac-turer’s instructions (Histostain-Plus kit; Zymed Laboratories). The avidin-biotin immunoperoxidase system with 3-amino-9-ethylcarbazole as chro-mogen was used to visualize bound antibodies. The preparations werecounterstained with hematoxylin, dehydrated, cleared, and mounted, aspreviously described [26].
Rat brains were postfixed in the same solution of perfusion and cutcoronally on a vibratome (40 mm) 2 days later. Free-floating sections werewashed in 0.1 M glycine in PBS, washed twice in PBS, and treated with0.5% H2O2 in PBS, washed three times in PBS, and preincubated in PBScontaining 10% normal goat serum and 0.25% Triton X-100 for 1 h at48C. Free-floating hypothalamic sections were incubated for 48 h with theprimary Ob-R antibody (H-300; Santa Cruz Biotechnology) at a dilutionof 1:500. The reaction was visualized using biotinylated secondary antiseraand by standard avidin-biotin-horseradish peroxidase technique as de-scribed by Dragunow et al. [27]. After stopping the reaction by severalwashes in PBS, sections were mounted on chrome-alume gelatin-coatedslides, dehydrated, and coverslipped with mounting medium. Slides were
analyzed with a Zeiss Axioskope 2 light microscope (Oberkochen, Ger-many) equipped with high-resolution digital camera.
In both testes and brain experiments, negative control sections wereprocessed in the absence of the primary Ob-R antibody and after prein-cubation of the Ob-R antibody with oversaturating concentrations of Ob-R (541–840) peptide (Santa Cruz Biotechnology) overnight at 48C.
RNA Extraction and Reverse Transcription-PolymeraseChain Reaction Analysis
Tissue mRNAs were extracted by using a commercial kit (Micro-Fast-Track Kit; Invitrogen, San Diego, CA). First-strand cDNA was synthesizedfrom 0.5 mg of mRNA using an annealing temperature of 658C in a finalvolume of 25 ml containing 250 mM Tris-HCl, 375 mM KCl, 15 mMMgCl2, 50 mM dithiothreitol, 0.5 mM dNTPs, 0.5 mg random hexameroligonucleotide, 200 U M-MLV-RT, 26 U ribonuclease inhibitor (Promega,Madison, WI). b-Actin was used as a constitutive expressed gene productfor comparison of Ob-R abundances in different samples. A 0.5-ml volumeof the reverse transcription (RT) products was amplified with 2.5 units ofTaq DNA polymerase (Promega) and 20 mM specific rat b-actin primers(59-ATTGGCAATGAGCGGTTCCGC-39 [sense, nucleotides 2413–2437]and 59-CTCCTGCTTGCTGATCCACATC-39 [antisense, nucleotides2749–2727], GenBank accession no. J00691) in 50 ml of reaction mixcontaining 50 mM KCl, 10 mM Tris-HCl, 1.5 mM MgCl2, and 10 mMeach of the dNTPs. Thermocycling conditions were 1 min of denaturationat 948C, 1 min of annealing at 588C, and 1 min of extension at 728C. Toensure amplification in the exponential phase of the polymerase chainreaction (PCR), reactions were temporarily halted at 20, 25, 30, 35, and40 cycles and 12 ml of reaction mix were removed from each tube. Allproducts were analyzed by 1.5% agarose gel electrophoresis (data notshown), and 25 cycles was chosen for further analysis. Quantitation of thesignals was performed by densitometric analysis using densitometric com-puter software (Kodak Digital Sciences ID Image Analysis software; East-man Kodak, Rochester, NY). Dilution of RT products was made whennecessary and the amplification procedure was repeated until all sampleswere standardized for b-actin content. After standardization, PCR was per-formed using appropriately diluted RT products in 50 ml of the reactionmix by utilizing 20 mM of each rat Ob-R primer (59-ATGCTGTGCAGT-CACTCAGTG-39 [sense, nucleotides 2284–2303] and 59-CA-ACTCCTTCCATAAATACTGGG-39 [antisense, nucleotides 2526–2503],GenBank accession no. U52966). The used set of primers generated a 242-base pair (bp) PCR product corresponding to the extracellular domaincommon to all Ob-R isoforms. In order to study the developmental patternof the different isoforms of leptin receptor, we used two more sets ofprimers (59-GATATTTGGTCCTCTTCTTCTGG-39 [sense, nucleotides2786–2809] and 59AGTTGTGGTGAAATCACATTGG-39 [antisense, nu-cleotides 3223–3201], GenBank accession no. U52966), generating a 437-bp PCR product corresponding to the intracellular domain of Ob-R, spe-cific for the long isoform of the receptor (Ob-Rb); and (59-ATGCTGTGCAGTCACTCAGTG-39 [sense, nucleotides 2284–2303] and59-ACTTCAAAGAGTGTCCGCTCT-39 [antisense, nucleotides 2668–2689], GenBank accession no. D84125), chosen to generate a fragment of479 bp, specific for the short Ob-Ra isoform. For each gene examined, allprimers were derived from separate exons and spanned at least one intronof genomic sequence, thus excluding the possibility of genomic DNAcontamination. No PCR product was obtained with any of the set of prim-ers in the absence of cDNA template (data not shown). Thermocyclingconditions were 30 sec of denaturation at 948C, 30 sec of annealing at558C, and 60 sec of extension at 728C, with an additional 2-min extensionstep at 728C. Again, reactions were temporarily halted at 25, 30, 33, and35 cycles to ensure amplification in the exponential phase of PCR; 30cycles was chosen for densitometric analysis of Ob-R and 33 cycles waschosen for Ob-Ra and Ob-Rb because levels of PCR products increasedin a linear fashion for up to 35 cycles. The identities of the generated PCRproducts were confirmed by cycle sequencing (Thermo Sequenase Radio-labeled Terminator Cycle Sequencing Kit; Amersham Italia, Milan, Italy),and the size was compared with molecular weight markers (100-bp ladder,Promega, Madison, WI). Quantitations of Ob-R, Ob-Ra, and Ob-RbmRNA were evaluated by densitometric analysis as previously describedafter normalization with b-actin.
Leydig Cells StudiesImmature and adult Leydig cells were isolated from Sprague-Dawley
rats that were, respectively, 21 (10 rats per experiment) and 60 days old(6 rats per experiments) on the day of isolation. Leydig cells were obtainedby collagenase digestion of decapsulated testes as already described [28].

1201LEPTIN RECEPTOR IN THE TESTIS
FIG. 1. Immunohistochemistry of Ob-Rin rat testes from animals of different ages:early gestational (E14.5, A), late gestation-al (E19.5, B), postnatal prepubertal (7days, C; 14 days, D; 22 days, E), pubertal(35 days, F), and adult postpubertal (60days, G; 90 days, H). The results shownare representative of three independent ex-periments using different tissues in eachexperimental setting. Ob-R-specific immu-nostaining was observed in the interstitiumat E19.5 (B) but not at E14.5 (A), while inpostnatal life, it was evident only after sex-ual maturation (35, 60, 90 days old) andwas confined to Leydig cells. No immuno-reaction was present in the tubules. A, B,F–H) Bar 5 80 mm. C–E) Bar 5 160 mm.
Crude cell suspension was washed and then pelleted at 200 3 g for 10min. The cell pellet was then resuspended in regular Medium 199 (Whit-taker M.A. Bioproducts, Walkerville, MD) with Hanks salts and L-gluta-mine containing 1.4 g/L NaHCO3, 0.5% BSA, 1 mM EDTA, 50 U/mlheparin, 12.5 mg/L deoxyribonuclease, 50 mg/L gentamicin, pH 7.4) andthen centrifuged in a discontinuous Percoll density gradient at 800 3 gfor 30 min at 208C without rotor brake. Bands between Percoll 68% and43% were carefully taken, washed twice, and then pelleted and resus-pended in Medium 199 containing 0.1% BSA, 50 mg/L gentamicin, and0.125 mM 3-methylisobutyl xanthine (Aldrich Chemical, Milwaukee, WI).Nitro-blue-tetrazolium staining showed a purity of approximately 90% forimmature Leydig cell and 95% for adult Leydig cell fractions. The purifiedimmature and adult rat Leydig cells were plated in 24-well cell culturedishes (0.75 3 106 cells/well) and then incubated at 378C under 95% O2and 5% CO2 for 30 and 90 min in the presence or absence of saturating
doses of hCG (1 ng/ml; CR 121; preparation kindly provided by the centerfor Population Research, NICHD, Bethesda, MD) with or without recom-binant murine leptin (R & D Systems, Minneapolis, MN) at a final con-centration of 150 ng/ml, which in other studies has already been shownto determine the maximal inhibitory effect on hCG-induced testosteroneproduction by adult Leydig cells in culture [14]. At the end of incubation,media were removed and centrifuged at 250 3 g for 10 min, and thesupernatants were saved for the assay of extracellular testosterone (T). Allplated cells were also processed for the analysis of intracellular cAMP, asdescribed elsewhere [14, 29]. The measurements of testosterone andcAMP were performed by RIA, as previously described [14, 30, 31]. ForcAMP, the sensitivity of the assay was 1 fmol/ml; the inter- and intraassaycoefficients of variation were 6.4% and 4%, respectively. For T, the sen-sitivity of the method was 10 pg/assay tube; the mean inter- and intraassaycoefficients of variation were 10% and 8%, respectively.

1202 CAPRIO ET AL.
FIG. 2. Immunohistochemistry of relaxin-like factor (RLF) in rat testes from animalsof the same ages studied in Figure 1. Theresults shown are representative of threeindependent experiments using differenttissues in each experimental setting. Anti-RLF staining was positive in the intersti-tium at E19.5 (B), identifying mature fetalLeydig cells. Mature fetal Leydig cellswere still present in the interstitium at Day7 postpartum (C; see arrows), while nostaining was evident at later ages beforesexual maturation (D, E). A clear punctateRLF staining appeared again in the intersti-tium on Day 35 (F) and a more homoge-neous signal identified mature adult-typeLeydig cells on Days 60 (G) and 90 (H).A, B, F–H) Bar 5 80 mm. C–E) Bar 5 160mm.
Every treatment was run in triplicate and every experiments was per-formed at least three times. Results are the mean 6 SEM unless otherwisespecified. Statistical significance was determined by ANOVA test.
RESULTS
Immunohistochemical localization of Ob-R and RLF inrat testis at different ages is shown in Figures 1 and 2,respectively. Ob-R immunoreactivity was undetectable atearly embryonic age (E14.5; Fig. 1A) but became evidentat late embryonic age (E19.5), where it was localized inseveral interstitial cells (Fig. 1B). No specific signal wasdetected in the tubular compartment. Immunostaining of
serial testicular sections with an antibody against RLF(kindly provided by Dr. Richard Ivell, University of Ham-burg, Hamburg, Germany), a specific marker for the maturefetal as well as adult rodent Leydig cells [23, 24], con-firmed the presence of mature fetal Leydig cells in E19.5rat testis (Fig. 2B). Furthermore, the pattern of RLF im-munostaining strongly resembled that of Ob-R, suggestingthat Ob-R protein is expressed in rat embryonic testis bymature fetal Leydig cells. After birth, Ob-R immunoreac-tivity was absent in 7-, 14-, and 21-day-old rat testes (Fig.1, C–E) either in the interstitial compartment or in the sem-iniferous tubule, indicating that Ob-R protein expression in

1203LEPTIN RECEPTOR IN THE TESTIS
FIG. 3. Immunohistochemistry of Ob-Rin rat hypothalamus dissected from prepu-bertal (14 days, A–C) and postpubertal rats(60 days, D–F). The results shown are rep-resentative of three independent experi-ments using different tissues in each. Im-munoreaction was evident at both studiedages, with similar intensity and a wholecell pattern of staining, and it was mainlylocalized in the arcuate nucleus and cho-roid plexus (see arrows). A, D) Bar 5 320mm. B, E) Bar 5 160 mm. C, F) Bar 5 80mm.
the testis is switched off after birth and is negligible beforesexual maturation either in residual mature fetal Leydigcells or in immature adult-type Leydig cells. Anti-RLFstaining identified mature fetal Leydig cells in the intersti-tium of 7-day-old rat testes (Fig. 2C; see arrows), whereasRLF immunoreactivity was absent in later prepubertal ages(14- and 21-day-old rats; Fig. 2, D and E), concomitantlywith the involution of fetal Leydig cells and the appearanceof the adult Leydig cell progenitors. Ob-R immunoreactiv-ity was again evident in postnatal testes after sexual mat-uration (35-, 60- and 90-day-old rats; see Fig. 1, F–H).Again, immunoreaction was exclusively confined to Leydigcells and no signal was detected in Sertoli and germ cells.RLF immunoreactivity pattern after sexual maturation par-alleled that of Ob-R, with a clear punctate RLF-specificstaining in the interstitium of rat testis on Day 35 (Fig. 2F)and a more homogeneous and intense RLF signal on Days60 and 90 (Fig. 2, G and H).
Figure 3 shows the immunolocalization of Ob-R in thehypothalamus of rats at different pubertal status (14 daysold versus 60 days old). Different from what was observedin the testis, Ob-R immunopositivity was clearly presentboth before (Fig. 3, A–C) and after (Fig. 3, D–F) sexualmaturation with a similar intensity, and it was mainly lo-calized in the choroid plexus and arcuate nucleus (see ar-rows), indicating a different regulatory mechanism of Ob-R expression in the testis and brain and suggesting thatsexual maturation does not alter the intensity and the pat-tern of Ob-R expression in the hypothalamus. In both testisand brain experiments, control sections processed in theabsence of primary Ob-R antibody and after preincubationof the Ob-R antibody with oversaturating concentrations ofOb-R protein showed no staining.
Figures 4 and 5 show the semiquantitative RT-PCR anal-ysis of mRNA extracted from rat testes at different ages by
using a set of primers specific for the common extracellulardomain (Fig. 4) or the short (Ob-Ra) and long (Ob-Rb)isoforms (Fig. 5) of leptin receptor. Ob-R mRNA expres-sion was present in testes from each age group, but semi-quantitative analysis showed a 3.5-fold higher transcriptconcentration in late gestational (E19.5) versus early ges-tational (E14.5) testes. After birth (7- and 14-day-old rats),Ob-R mRNA was present at lower levels, similar to thoseexpressed on Day E14.5, with a 2-fold increase on Day 21and significantly higher mRNA concentrations in sexuallymature testes, with gradual increases from younger to olderages (35- [3.5-fold], 60- [4.4-fold], and 90-day-old rats[6.7-fold] versus E14.5). The developmental pattern of Ob-Ra mRNA expression was similar to that of Ob-R: at DayE19.5, mRNA concentrations were three times higher thanat Day E14.5; at Days 7 and 14, they were relatively low,similar to those present at E14.5, with a subsequent gradualincrease from younger to older postnatal ages (21- [2.8-fold], 35- [3.1-fold], 60- [3.5-fold], and 90-day-old [5.15-fold] rats versus E14.5). In line with this pattern of expres-sion, also Ob-Rb mRNA concentrations were relativelyhigher at E19.5 (4.2-fold versus E14.5) and at adult ages(60- [6.26-fold] and 90-day-old rats [9.5-fold versusE14.5]), whereas they were less abundant at 7 (2.4), 14(2.6), 21 (3.3), and 35 (3.2-fold versus E14.5) days old.
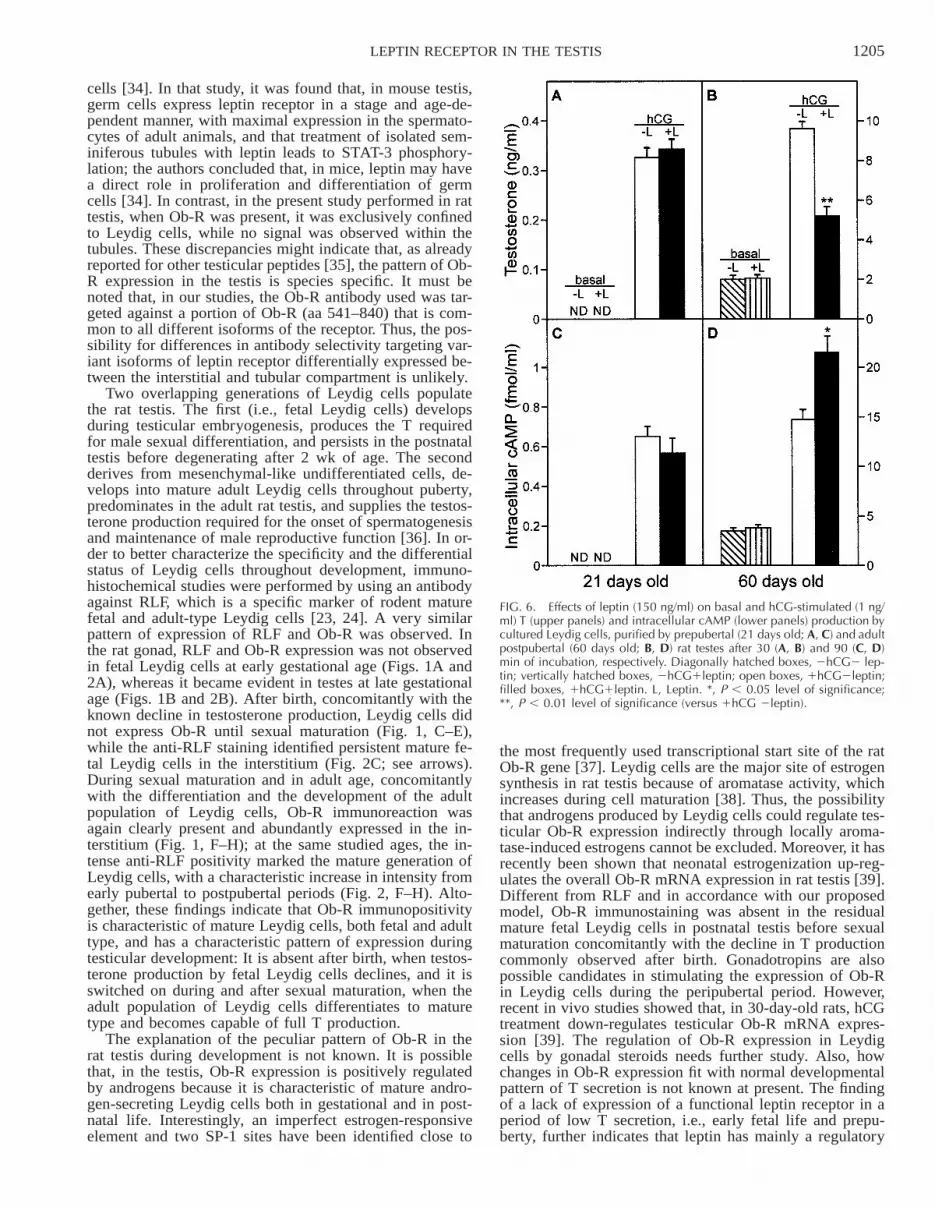
In order to establish whether leptin receptor has a func-tional role in rat prepubertal testis, in vitro studies wereperformed using isolated Leydig cells. Figure 6, A and B,shows basal and hCG-stimulated (1 ng/ml) T secretion bycultured Leydig cells, isolated from 21-day-old rats (sexu-ally immature) and 60-day-old rats (mature adults), respec-tively, after 30 min of incubation with leptin (150 ng/ml).As already found in previous studies [14, 32], leptin sig-nificantly inhibited hCG-stimulated T production after 30min of incubation in adult rat Leydig cells in culture (5.2

1204 CAPRIO ET AL.
FIG. 4. RT-PCR expression analysis of Ob-R in rat testes from animals ofdifferent ages. PCR products were subjected to electrophoresis in 1.5% aga-rose gel, visualized by ethidium bromide staining (upper), and quantified bydensitometric computer software after normalization for b-actin (lower). Thesizes of the generated amplification products were confirmed by comparisonwith a molecular weight marker and are indicated on the right side. Valuesare the mean 6 SEM of three separate experiments. A representative exper-iment is provided in the upper part of the figure. *, P , 0.05 level of signif-icance; **, P , 0.01 level of significance (versus E14.5).
FIG. 5. RT-PCR expression analysis of the short (Ob-Ra) and long iso-form (Ob-Rb) of leptin receptor in rat testes from animals of different ages.Values are the mean 6 SEM of three separate experiments. A represen-tative experiment is provided in the upper part of the figure. *, P , 0.05level of significance; **, P , 0.01 level of significance (versus E14.5).
6 0.46 ng/ml versus 9.6 6 0.36 ng/ml, 46% reduction; P, 0.01), whereas it had no effect on hCG-stimulated Tproduction in prepubertal Leydig cells. In both examinedsystems, leptin did not modify T production in the absenceof hCG stimulus. In Figure 6, C and D, are shown basaland hCG-stimulated intracellular cAMP production fromthe same experiment as in Figure 6, A and B, after 90 minof incubation with leptin (150 ng/ml). In line with previ-ously published results [14], leptin significantly increasedintracellular cAMP production in adult Leydig cells (21.496 1.68 fmol/ml versus 14.73 6 0.99 fmol/L, 46% increase;P , 0.05), whereas it did not affect cAMP production inimmature Leydig cells. Again, leptin alone did not affectcAMP production in both 21- and 60-day-old rat Leydigcells. These results indicate that, in rat Leydig cells, theleptin receptor is not functional before sexual maturation.
DISCUSSION
In this study, we investigated the developmental expres-sion of leptin receptor in rat testis from early gestational toadult age and found a peculiar pattern of Ob-R expressionin Leydig cells: in the testis, Ob-R immunoreactivity waspresent in late embryonic life, was absent in prepubertallife, and appeared again in adulthood. The developmentalpattern of Ob-R immunoreactivity in the testis was differentfrom that observed in the hypothalamus, where Ob-R im-munoreactivity was constantly evident throughout devel-opment and was consistent with functional studies. In fact,while leptin inhibited hCG-induced testosterone productionand amplified hCG-stimulated intracellular cAMP produc-tion by adult Leydig cells, it did not modify hCG-induced
steroidogenesis or hCG-stimulated intracellular cAMP pro-duction by prepubertal Leydig cells in vitro.
In our study at any studied age, Ob-R immunoreactionwas exclusively confined to Leydig cells, whereas no signalwas detected in the tubules, in Sertoli cells, or in germ cells.Others, by using in situ hybridization techniques, showedthat Ob-R mRNA is expressed in Sertoli cells of adult rats[33]. Thus, the lack of immunoreactivity inside the tubulessuggests that Ob-R protein is not translated in Sertoli cellsenough to be detectable by immunohistochemistry. Inter-estingly, in contrast with rat testis, in neonatal and adultmouse testis, it has been found that Ob-R immunoreactionis mainly present in the tubules and is localized to germ
1205LEPTIN RECEPTOR IN THE TESTIS
FIG. 6. Effects of leptin (150 ng/ml) on basal and hCG-stimulated (1 ng/ml) T (upper panels) and intracellular cAMP (lower panels) production bycultured Leydig cells, purified by prepubertal (21 days old; A, C) and adultpostpubertal (60 days old; B, D) rat testes after 30 (A, B) and 90 (C, D)min of incubation, respectively. Diagonally hatched boxes, 2hCG2 lep-tin; vertically hatched boxes, 2hCG1leptin; open boxes, 1hCG2leptin;filled boxes, 1hCG1leptin. L, Leptin. *, P , 0.05 level of significance;**, P , 0.01 level of significance (versus 1hCG 2leptin).
cells [34]. In that study, it was found that, in mouse testis,germ cells express leptin receptor in a stage and age-de-pendent manner, with maximal expression in the spermato-cytes of adult animals, and that treatment of isolated sem-iniferous tubules with leptin leads to STAT-3 phosphory-lation; the authors concluded that, in mice, leptin may havea direct role in proliferation and differentiation of germcells [34]. In contrast, in the present study performed in rattestis, when Ob-R was present, it was exclusively confinedto Leydig cells, while no signal was observed within thetubules. These discrepancies might indicate that, as alreadyreported for other testicular peptides [35], the pattern of Ob-R expression in the testis is species specific. It must benoted that, in our studies, the Ob-R antibody used was tar-geted against a portion of Ob-R (aa 541–840) that is com-mon to all different isoforms of the receptor. Thus, the pos-sibility for differences in antibody selectivity targeting var-iant isoforms of leptin receptor differentially expressed be-tween the interstitial and tubular compartment is unlikely.
Two overlapping generations of Leydig cells populatethe rat testis. The first (i.e., fetal Leydig cells) developsduring testicular embryogenesis, produces the T requiredfor male sexual differentiation, and persists in the postnataltestis before degenerating after 2 wk of age. The secondderives from mesenchymal-like undifferentiated cells, de-velops into mature adult Leydig cells throughout puberty,predominates in the adult rat testis, and supplies the testos-terone production required for the onset of spermatogenesisand maintenance of male reproductive function [36]. In or-der to better characterize the specificity and the differentialstatus of Leydig cells throughout development, immuno-histochemical studies were performed by using an antibodyagainst RLF, which is a specific marker of rodent maturefetal and adult-type Leydig cells [23, 24]. A very similarpattern of expression of RLF and Ob-R was observed. Inthe rat gonad, RLF and Ob-R expression was not observedin fetal Leydig cells at early gestational age (Figs. 1A and2A), whereas it became evident in testes at late gestationalage (Figs. 1B and 2B). After birth, concomitantly with theknown decline in testosterone production, Leydig cells didnot express Ob-R until sexual maturation (Fig. 1, C–E),while the anti-RLF staining identified persistent mature fe-tal Leydig cells in the interstitium (Fig. 2C; see arrows).During sexual maturation and in adult age, concomitantlywith the differentiation and the development of the adultpopulation of Leydig cells, Ob-R immunoreaction wasagain clearly present and abundantly expressed in the in-terstitium (Fig. 1, F–H); at the same studied ages, the in-tense anti-RLF positivity marked the mature generation ofLeydig cells, with a characteristic increase in intensity fromearly pubertal to postpubertal periods (Fig. 2, F–H). Alto-gether, these findings indicate that Ob-R immunopositivityis characteristic of mature Leydig cells, both fetal and adulttype, and has a characteristic pattern of expression duringtesticular development: It is absent after birth, when testos-terone production by fetal Leydig cells declines, and it isswitched on during and after sexual maturation, when theadult population of Leydig cells differentiates to maturetype and becomes capable of full T production.
The explanation of the peculiar pattern of Ob-R in therat testis during development is not known. It is possiblethat, in the testis, Ob-R expression is positively regulatedby androgens because it is characteristic of mature andro-gen-secreting Leydig cells both in gestational and in post-natal life. Interestingly, an imperfect estrogen-responsiveelement and two SP-1 sites have been identified close to
the most frequently used transcriptional start site of the ratOb-R gene [37]. Leydig cells are the major site of estrogensynthesis in rat testis because of aromatase activity, whichincreases during cell maturation [38]. Thus, the possibilitythat androgens produced by Leydig cells could regulate tes-ticular Ob-R expression indirectly through locally aroma-tase-induced estrogens cannot be excluded. Moreover, it hasrecently been shown that neonatal estrogenization up-reg-ulates the overall Ob-R mRNA expression in rat testis [39].Different from RLF and in accordance with our proposedmodel, Ob-R immunostaining was absent in the residualmature fetal Leydig cells in postnatal testis before sexualmaturation concomitantly with the decline in T productioncommonly observed after birth. Gonadotropins are alsopossible candidates in stimulating the expression of Ob-Rin Leydig cells during the peripubertal period. However,recent in vivo studies showed that, in 30-day-old rats, hCGtreatment down-regulates testicular Ob-R mRNA expres-sion [39]. The regulation of Ob-R expression in Leydigcells by gonadal steroids needs further study. Also, howchanges in Ob-R expression fit with normal developmentalpattern of T secretion is not known at present. The findingof a lack of expression of a functional leptin receptor in aperiod of low T secretion, i.e., early fetal life and prepu-berty, further indicates that leptin has mainly a regulatory

1206 CAPRIO ET AL.
role on gonadotropin control of androgen production [14],the same as has been found for many other regulators oftesticular function [35].
The pattern of expression of Ob-R in the testis was dif-ferent from that observed in rat hypothalamus, where amarked Ob-R immunoreaction was evident both before andafter sexual maturation. Ob-R was mainly localized in thearcuate nucleus and choroid plexus (Fig. 3; see arrows),with no significant difference in the intensity of Ob-R im-munostaining before and after sexual maturation. These ob-servations are in line with previous studies showing thatOb-R immunopositivity is present in the paraventricular nu-cleus at the late embryonic age, in the newborn, and duringthe suckling period, even if at a weak level compared withthe adult age [40]. Therefore, it appears that different mech-anisms control Ob-R expression in the testis and in thehypothalamus during development, probably through a tis-sue-specific different promoter [41] and/or a different localhormonal milieu [35].
Semiquantitative RT-PCR results obtained on testes fromrats of different ages were in line with the immunohisto-chemical data (Figs. 4 and 5): Ob-R mRNA expression wassignificantly higher at late gestational (E19.5) versus earlygestational (E14.5) age. After birth (7 and 14 days old),Ob-R mRNA levels were similar to those found on dayE14.5, showed a slight increase on Day 21, and were muchhigher in mature testes, with a gradual increase from youn-ger to older ages. Also, the developmental pattern ofmRNA concentrations of the short (Ob-Ra) and long (Ob-Rb) isoforms of Ob-R was very similar to that of overalltesticular Ob-R splice variants. Ob-R protein was markedlyswitched on at the time of sexual maturation, and absolutelyno protein signal was detected at all studied prepubertalages. These observations suggest that Ob-R expression istranscriptionally regulated, even if a posttranscriptional lev-el of regulation cannot be excluded.
These latter data are partially in contrast with evidencefrom other authors [39], who detected Ob-R mRNA in tes-tes from 15-, 30-, 45-, and 75-day-old rats at rather constantrelative levels, while the Ob-Rb isoform mRNA concentra-tion was found to be higher in pubertal testes (30 days old),with a subsequent decline in adulthood (75 days old). Wecannot explain the discrepancies between these and ourfindings. However, in contrast with the report by Tena-Sem-pere et al. [39], we performed all RT-PCR studies on mes-senger RNA rather than total RNA, which allows a morespecific and accurate quantification of transcripts. In addi-tion, we used Sprague-Dawley rats, while the others usedWistar rats. Finally, in our study, we found a valuable con-sistency between immunohistochemical data, mRNA ex-pression analysis, and functional findings. In order to es-tablish whether immunohistochemical and RT-PCR resultshad functional relevance, studies on Leydig cells isolatedbefore and after sexual maturation were performed. We re-cently showed that leptin is able to suppress hCG-inducedT production and increase cAMP production by adult iso-lated Leydig cells in vitro [14]. Also, leptin levels in obesehuman subjects are the best hormonal predictor of the obe-sity-related reduction in androgen response to hCG tests invivo [42]. In line with the morphological data and RT-PCRresults, leptin did not modify either hCG-stimulated T orintracellular cAMP production by cultured prepubertal Ley-dig cells (Fig. 6), where Ob-R immunostaining was nega-tive and the relative abundance of Ob-R mRNA was low.Similar results were reported by other authors using slicesof prepubertal testicular tissue incubated in the presence of
hCG and leptin [32]. Taken together, these results indicatethat, in rat Leydig cells, the Ob → Ob-R system is notfunctional before sexual maturation.
In conclusion our results underline how complex are theleptin-leptin receptor interactions at different anatomic lev-els, and indicate that this system may regulate Leydig cellfunction in a concentration and age-dependent manner. Itappears that the Ob → Ob-R system in the testis may serveto negatively regulate T production by Leydig cells duringadult but not prepubertal life and that it could have a roleduring testicular embryogenesis and Leydig cell maturationin prenatal life.
REFERENCES
1. Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM.Positional cloning of the mouse obese gene and its human homologue.Nature 1994; 372:425–432.
2. Halaas JL, Gajiwala KS, Maffei M, Cohen SL, Chait BT, RabinowitzD, Lallone RL, Burley SK, Friedman JM. Weight-reducing effect ofthe plasma protein encoded by the obese gene. Science 1995; 269:543–546.
3. Chehab FF, Lim ME, Lu R. Correction of the sterility defect in ho-mozygous obese female mice by treatment with human recombinantleptin. Nat Genet 1996; 12:318–320.
4. Tartaglia LA, Dembski M, Weng X, Deng N, Culpepper J, Devos R,Richards GJ, Campfield LA, Clark FT, Deeds J, Muir C, Sanker S,Moriarty A, Moore KJ, Smutko JS, Mays GG, Woolf EA, Tepper RL.Identification and expression cloning of a leptin receptor, OB-R. Cell1995; 83:1263–1271.
5. Tartaglia LA. The leptin receptor. J Biol Chem 1997; 272:6092–6096.6. Lee GH, Proenca R, Montez JM, Carroll KM, Darvishzadeh JG, Lee
JI, Friedman JM. Abnormal splicing of leptin receptor in diabeticmice. Nature 1996; 379:632–635.
7. Fei H, Okano HJ, Li C, Lee GH, Zhao C, Darnell R, Friedman JM.Anatomic localization of alternatively spliced leptin receptors (Ob-R)in mouse brain and other tissues. Proc Natl Acad Sci U S A 1997;94:7001–7005.
8. Spicer JL, Francisco CC. The adipose obese gene product, leptin:evidence of a direct inhibitory role in ovarian function. Endocrinology1997; 138:3374–3379.
9. Karlsson C, Lindell K, Svensson E, Bergh C, Lind P, Billig H, Carls-son LM, Carlsson B. Expression of functional leptin receptors in hu-man ovary. J Clin Endocr Metab 1997; 82:4114–4148.
10. Ryan NK, Woodhouse CM, Van der Hoek KH, Gilchrist RB, Arm-strong DT, Norman RJ. Expression of leptin and its receptor in themurine ovary: possible role in the regulation of oocyte maturation.Biol Reprod 2002; 66:1548–1554.
11. Bornstein SR, Uhlmann K, Haidan A, Ehrhart-Bornstein M, Scher-baum WA. Evidence for a novel peripheral action of leptin as a met-abolic signal to the adrenal gland: leptin inhibits cortisol release di-rectly. Diabetes 1997; 46:1235–1238.
12. Kieffer TJ, Heller RS, Habener JF. Leptin receptors expressed on pan-creatic beta-cells. Biochem Biophys Res Commun 1996; 224:522–527.
13. Zamorano PL, Mahesh VB, De Sevilla LM, Chorich LP, Bhat GK,Brann DW. Expression and localization of the leptin receptor in en-docrine and neuroendocrine tissues of the rat. Neuroendocrinology1997; 65:223–238.
14. Caprio M, Isidori AM, Carta AR, Moretti C, Dufau ML, Fabbri A.Expression of functional leptin receptors in rodent Leydig cells. En-docrinology 1999; 140:4939–4947.
15. Zachow RJ, Magoffin DA. Direct intraovarian effects of leptin: im-pairment of the synergistic action of insulin-like growth factor-1 onFSH-dependent 17b-estradiol production by rat ovarian granulosacells. Endocrinology 1999; 137:847–850.
16. Pralong FP, Roduit R, Waeber G, Castillo E, Mosimann F, Thorens B,Gaillard RC. Leptin inhibits directly glucocorticoid secretion by nor-mal human and rat adrenal gland. Endocrinology 1998; 139:4264–4268.
17. Swerdloff RS, Batt RA, Bray GA. Reproductive hormonal function inthe genetically obese (ob/ob) mouse. Endocrinology 1975; 98:1359–1364.
18. Farooqi IS, Jebb SA, Langmack G, Lawrence E, Cheetham CH, Pren-tice AM, Hughes IA, McCamish MA, O’Rahilly S. Effects of recom-

1207LEPTIN RECEPTOR IN THE TESTIS
binant leptin therapy in a child with congenital leptin deficiency. NEngl J Med 1999; 341:879–884.
19. Caprio M, Fabbrini E, Isidori AM, Aversa A, Fabbri A. Leptin inreproduction. Trends Endocrinol Metab 2001; 12:65–72.
20. Mantzorozs CS, Flier JS, Rogol AD. A longitudinal assessment ofhormonal and physical alteration during normal puberty in boys. Ris-ing leptin levels may signal the onset of puberty. J Clin EndocrinolMetab 1997; 82:1066–1070.
21. Suter KJ, Pohl CR, Wilson ME. Circulating concentration of nocturnalleptin, growth hormone and insulin-like growth factor-1 increase be-fore the onset of puberty in agonadal male monkey: potential signalfor the initiation of puberty. J Clin Endocrinol Metab 2000; 85:808–814.
22. Plant TM, Durrant AR. Circulating leptin does not appear to providea signal for triggering the initiation of puberty in the male rhesusmonkey (Macaca mulatta). Endocrinology 1997; 138:4505–4508.
23. Balvers M, Spiess AN, Domagalski R, Hunt N, Kilic E, Mukhopa-dhyay AK, Hanks E, Charlton HM, Ivell R. Relaxin-like factor ex-pression as a marker of differentiation in the mouse testis and ovary.Endocrinology 1998; 139:2960–2970.
24. Spiess AN, Balvers M, Tena-Sempere M, Huhtaniemi I, Parry L, IvellR. Structure and expression of the rat relaxin-like factor (RLF) gene.Mol Reprod Dev 1999; 54:319–325.
25. Ojeda SR, Andrews WW, Advis JP, Smith-White S. Recent advancesin the endocrinology of puberty. Endocr Rev 1980; 1:228–257.
26. Claesson-Welsh L, Hammacher A, Westermark B, Heldin CH, NisterM. Identification and structural analysis of the type A receptor for theplatelet-derived growth factor: similarity with the type B receptor. JBiol Chem 1989; 264:1742–1747.
27. Dragunow M, Peterson MR, Robertson HA. Presence of c-fos-likeimmunoreactivity in the adult brain. Eur J Pharmacol 1987; 135:113–114.
28. Dufau ML, Mendelson CR, Catt KJ. A highly sensitive in vitro bio-assay for luteinizing hormone and chorionic gonadotropin: testoster-one production by dispersed Leydig cells. J Clin Endocrinol Metab1974; 39:610–613.
29. Khanum A, Dufau ML. A cAMP-independent inhibitory action ofhigh doses of forskolin in rat Leydig cells. J Steroid Biochem MolBiol 1990; 37:669–674.
30. Dufau ML, Horner KA, Hayashi K. Action of choleragen and gonad-otropin in isolated Leydig cells. J Biol Chem 1978; 253:3721–3729.
31. Ulisse S, Fabbri A, Dufau ML. Corticotropin-releasing factor recep-tors and actions in rat Leydig cells. J Biol Chem 1989; 264:2156–2163.
32. Tena-Sempere M, Pinilla L, Gonzalez LC, Dieguez C, Casanueva FF,Aguilar E. Leptin inhibits testosterone secretion from adult rat testisin vitro. J Endocrinol 1999; 161:211–218.
33. Tena-Sempere M, Manna PR, Zhang FP, Pinilla L, Gonzalez LC, Die-guez C, Huhtaniemi I, Aguilar E. Molecular mechanisms of leptinaction in adult rat testis: potential targets for leptin-induced inhibitionof steroidogenesis and pattern of leptin receptor messenger ribonucleicacid expression. J Endocrinol 2001; 170:413–423.
34. El-Hefnawy T, Ioffe S, Dym M. Expression of the leptin receptorduring germ cell development in the mouse testis. Endocrinology2000; 141:2624–2630.
35. Gnessi L, Fabbri A, Spera G. Gonadal peptides as mediators of de-velopment and functional control of the testis: an integrated systemwith hormones and local environment. Endocr Rev 1997; 18:541–609.
36. Benton L, Shan LX, Hardy MP. Differentiation of adult Leydig cells.J Steroid Biochem Mol Biol 1995; 53:61–68.
37. Lindell K, Bennett PA, Itoh Y, Robinson IC, Carlsson LM, CarlssonB. Leptin receptor 59 untranslated regions in the rat: relative abun-dance, genomic organization and relation to putative response ele-ments. Mol Cell Endocrinol 2001; 172:37–45.
38. Tsai-Morris CH, Aquilano DR, Dufau ML. Cellular localization of rattesticular aromatase activity during development. Endocrinology1985; 116:38–46.
39. Tena-Sempere M, Pinilla L, Zhang FP, Gonzalez LC, Huhtaniemi I,Casanueva FF, Dieguez C, Aguilar E. Developmental and hormonalregulation of leptin receptor (Ob-R) messenger ribonucleic acid ex-pression in rat testis. Biol Reprod 2001; 64:634–643.
40. Matsuda J, Yokota I, Tsuruo Y, Murakami T, Ishimura K, Shima K,Kuroda Y. Developmental changes in long-form leptin receptor ex-pression and localization in rat brain. Endocrinology 1999; 140:5233–5238.
41. Mercer JG, Moar KM, Hoggard N, Strosberg AD, Froguel P, BailleulB. B219/OB-R 59-UTR and leptin receptor gene-related protein geneexpression in mouse brain and placenta: tissue-specific leptin receptorpromoter activity. J Neuroendocrinol 2000; 12:649–655.
42. Isidori AM, Caprio M, Strollo F, Moretti C, Frajese G, Isidori A,Fabbri A. Leptin and androgens in male obesity: evidence for leptincontribution to reduced androgen levels. J Clin Endocrinol Metab1999; 84:3673–3680.