expression of leptin and leptin receptor isoforms in the rat and human carotid body
TRANSCRIPT

B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Expression of leptin and leptin receptor isoforms in the rat andhuman carotid body
Andrea Porzionatoa, Marcin Rucinskib, Veronica Macchia, Carla Steccoa,Ignazio Castagliuoloc, Ludwik K. Malendowiczb, Raffaele De Caroa,⁎aSection of Anatomy, Department of Human Anatomy and Physiology, University of Padova, 35121 Padova, ItalybDepartment of Histology and Embryology, Poznan University of Medical Sciences, 6 Swiecicki St., Poznan PL-60781, PolandcDepartment of Histology, Microbiology and Medical Biotechnologies, University of Padua, Padua, Italy
A R T I C L E I N F O
⁎ Corresponding author. Fax: +39 049 8272328E-mail address: [email protected] (R. De C
0006-8993/$ – see front matter © 2011 Elsevidoi:10.1016/j.brainres.2011.02.028
A B S T R A C T
Article history:Accepted 11 February 2011Available online 18 February 2011
Leptin is known to play a role in themodulation ofmetabolism and control of breathing actingmainly on central nervous structures, although additional actions on peripheral arterialchemoreceptors have also been suggested in the literature. We therefore examined bymeansof immunohistochemistry the expression of leptin and leptin receptors in the carotid bodies ofrats and humans. Leptin expression and relative expression of leptin receptor isoforms werealso studied in rats by real-time PCR. No leptin or leptin receptor immunoreactivities werevisible in the type II cells of either series. In rat carotid bodies, diffuse positive stainings forleptin and leptin receptors (bothwith antibody recognizing all receptor isoforms and antibodyspecific for Ob-Rb)were observed in type I cells. In human carotid bodies, themeanpercentage(±standard error) of leptin immunoreactive type I cells was 39.4%±5.1% and the percentages ofleptin receptor immunoreactive type I cells were 57.3%±3.9% with antibody recognizing allreceptor isoforms and 33.3%±4.2% with antibody specific for isoform Ob-Rb. Doubleimmunofluorescences with anti-tyrosine hydroxylase (type I cell marker) and anti-glialfibrillary acidic protein (type II cell markers) confirmed the selective location of leptin and Ob-Rb in type I cells. Real-time PCR showed the expression of leptin and Ob-Ra, Ob-Rb, Ob-Rc andOb-Rf isoform mRNA in the rat carotid body, levels of expression being Ob-Rf>Ob-Rc>>Ob-Ra>>Ob-Rb. Ob-RemRNAwas not detected. The above findings suggest a role of circulating orlocally produced leptin in the regulation of chemoreceptor discharge and/ormetabolic sensingfunction, by means of direct action on type I cells.
© 2011 Elsevier B.V. All rights reserved.
Keywords:LeptinLeptin receptorImmunohistochemistryReal-time PCRCarotid body
1. Introduction
Leptin is a 167-amino acid non-glycosylated peptide which isencoded in rats andmice by the obese (ob) gene and in humansby the homolog gene LEP (Zhang et al., 1994). It is highlyconserved and shows a bundle of four α-helices, characteristicof the class I cytokine superfamily (Zhang et al., 1997). Leptin is
.aro).
er B.V. All rights reserved
predominantly synthesized by adipocytes, plasma levels beingproportional to body fat content (Frederich et al., 1995; Maffeiet al., 1995), and regulates food intake andenergy consumptionby hypothalamic action (Ahima et al., 1996; Jacob et al., 1997).More recent studies have revealedmany other roles, includingthe ability to modulate breathing, neuroendocrine, reproduc-tive, inflammatory and immune functions, angiogenesis, fatty
.

Fig. 1 – Ethidium bromide-stained 2% agarose gel, showingPCR product amplified with rat specific primer from cDNA of
57B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
acid metabolism, glucose homeostasis, sensory nerve inputand autonomic outflow, neuronal plasticity, survival andproliferation (Bjørbaek and Kahn, 2004; Lago et al., 2007;Malendowicz et al., 2007; O'Donnell et al., 1999, 2000; Ribattiet al., 2007; Tankersley et al., 1998; Tang, 2008).
Leptin receptors are encoded by the gene diabetes (db) andbelong to the class I cytokine receptor superfamily, includinggp130 and receptors for interleukin-6, leukemia inhibitoryfactor and granulocyte colony-stimulating factor (Lee et al.,1996; Tartaglia et al., 1995). Alternative splicing of the db genegenerates 6 leptin receptor isoforms, termed from Ob-Ra toOb-Rf, which have an identical extracellular N-terminalportion of more than 800 amino acids. Ob-Re is the onlysoluble receptor form, probably binding circulating leptin andaffecting its stability and availability (Gavrilova et al., 1997;Yang et al., 2004). The other isoforms show a 34-amino acidtransmembrane region and short or long C-terminal domainsof 30–40 (isoforms Ob-Ra, Ob-Rc, Ob-Rd, Ob-Rf) or 302 (isoformOb-Rb) amino acids, respectively (Lee et al., 1996; Wang et al.,1996). Short isoforms are considered to be mainly involved inendocytosis and transport of leptin across the blood-brainbarrier (Banks et al., 1996), although a possible transport rolehas also been recently hypothesized for Ob-Rb (Tu et al., 2007).
Leptin modulatory actions on metabolism and breathingmainly involve central nervous structures. However, addition-al actions on peripheral arterial chemoreceptors have alsobeen suggested in the literature (Groeben et al., 2004) although,to the best of our knowledge, there are no data regarding theexpression of leptin and its receptors in these structures. Thecarotid body is the main peripheral arterial chemoreceptorwhich induces increases in ventilatory volume and frequencyin response to hypoxia, hypercapnia or reduction of blood pH.It also shows glucosensing properties and intervenes inregulation of metabolism (Alvarez-Buylla and de Alvarez-Buylla, 1988; Fitzgerald et al., 2009; Koyama et al., 2000; Pardaland López-Barneo, 2002). It is organized in lobules of cellsbelonging to two different populations: type I (or chief) cells,roundish in shape and considered to be the true chemorecep-tor elements of the carotid body, releasing various neuro-transmitters and neuromodulators in response to stimulation(Nurse, 2005; Porzionato et al., 2008a,b); type II (or sustenta-cular) cells, fusiform in shape, which show astrocytic markersand play a supportive role, although these cells have alsorecently been observed to behave as stem cell precursors fortype I cells when exposed to hypoxic stimuli (Pardal et al.,2007). Chemosensory impulses are mainly conveyed throughglossopharyngeal afferent fibers going to thepetrosal ganglion.
The aim of the present study was to examine by means ofimmunohistochemistry and immunofluorescence the expres-sion and distribution of leptin and leptin receptors in the ratand human carotid body, with particular reference to thedifferent cellular populations. Leptin expression and relativeexpression of leptin receptor isoforms were also investigatedin rats by real-time polymerase chain reaction (real-time PCR).
carotid body. Size marker: O'Range Ruler 50-bp DNA Ladder;MBI Fermentas, Lithuania. Two carotid body lanes representPCR products from two independent reverse transcriptions(RT) from one pool. Reaction products of expected size areseen: Ob, 195 bp; Ob-Ra, 235 bp; Ob-Rb, 171 bp; Ob-Rc, 161 bp,Ob-Rf, 151 bp.
2. Results
PCR amplification of tyrosine hydroxylase mRNA confirmedthe correspondence of RNA assayed with carotid body RNA.
The classic RT-PCR method demonstrated the presence ofleptin and Ob-Ra, Ob-Rb, Ob-Rc and Ob-Rf isoform mRNA inthe rat carotid body. With specific primers, RNA amplificationby RT-PCR revealed the presence of reaction products ofexpected size (Fig. 1). There were no PCR products in negativecontrol reactions.

Fig. 2 – QPCR analyses of Ob and Ob-R gene expression in ratcarotid body. All samples were amplified in triplicate, andPBGD gene expression was used as reference to normalizedata. Means±S.E.M. are shown.
Fig. 3 –Anti-leptin immunohistochemistry in rat (A, B) and humanrat (A) and selective immunostaining of some type I cells in humsurrounding lobules of type I cells and the absence of immunostaarrowheads: negative type I cells; asterisks: negative type II cells
58 B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
Semiquantitative PCR revealed the highest expression levelfor the leptin gene and lower levels of expression for allreceptors. Among the receptors, Ob-Rf and Ob-Rc genesshowed the highest expression levels. The expression levelof Ob-Ra gene was approximately 10 times lower than that ofOb-Rf and Ob-Rc, but approximately 10 times higher than Ob-Rb. Ob-Rb was the lowest detectable isoform of leptin receptorgenes. There was no expression of Ob-Re gene in the carotidbody (Fig. 2).
Immunohistochemical analysis revealed the expression ofleptin (Fig. 3) and leptin receptors (Figs. 4–5) both in humanand rat presumptive type I cells, i.e., large and roundish cellsorganized in clusters. Immunostaining of these cells wasobserved, both with an antibody recognizing all receptorisoforms and one specific for Ob-Rb. In rat carotid bodies,cytoplasmic stainings for leptin (Fig. 3A) and leptin receptors(Fig. 4B–C, E–F) were diffusely positive in almost all glomic typeI cells. In human carotid bodies, anti-leptin (Fig. 3C) and –leptin receptor (Fig. 5B–C, E–F) immunohistochemistriesrevealed the coexistence of both positive and negativepresumptive type I cells in all specimens examined. Whenpresent, the immunoreactivity of both leptin and leptinreceptors was intense and widely distributed in the cyto-plasm. Immunostained cells were found in both the centerand periphery of lobules. In human carotid bodies, the meanpercentage (±standard error) of leptin immunoreactive type Icells was 39.4%±5.1% and the percentages of leptin receptorimmunoreactive type I cells were 57.3%±3.9% with antibodyrecognizing all receptor isoforms and 33.3%±4.2% with
(C, D) carotid bodies. Diffuse positivity of glomic type I cells inan (C) are visible. Note the negativity of elongated type II cellsining in negative controls (B, D). Arrows: positive type I cells;. Scale bars=25 μm (A, C); 38 μm (B, D).
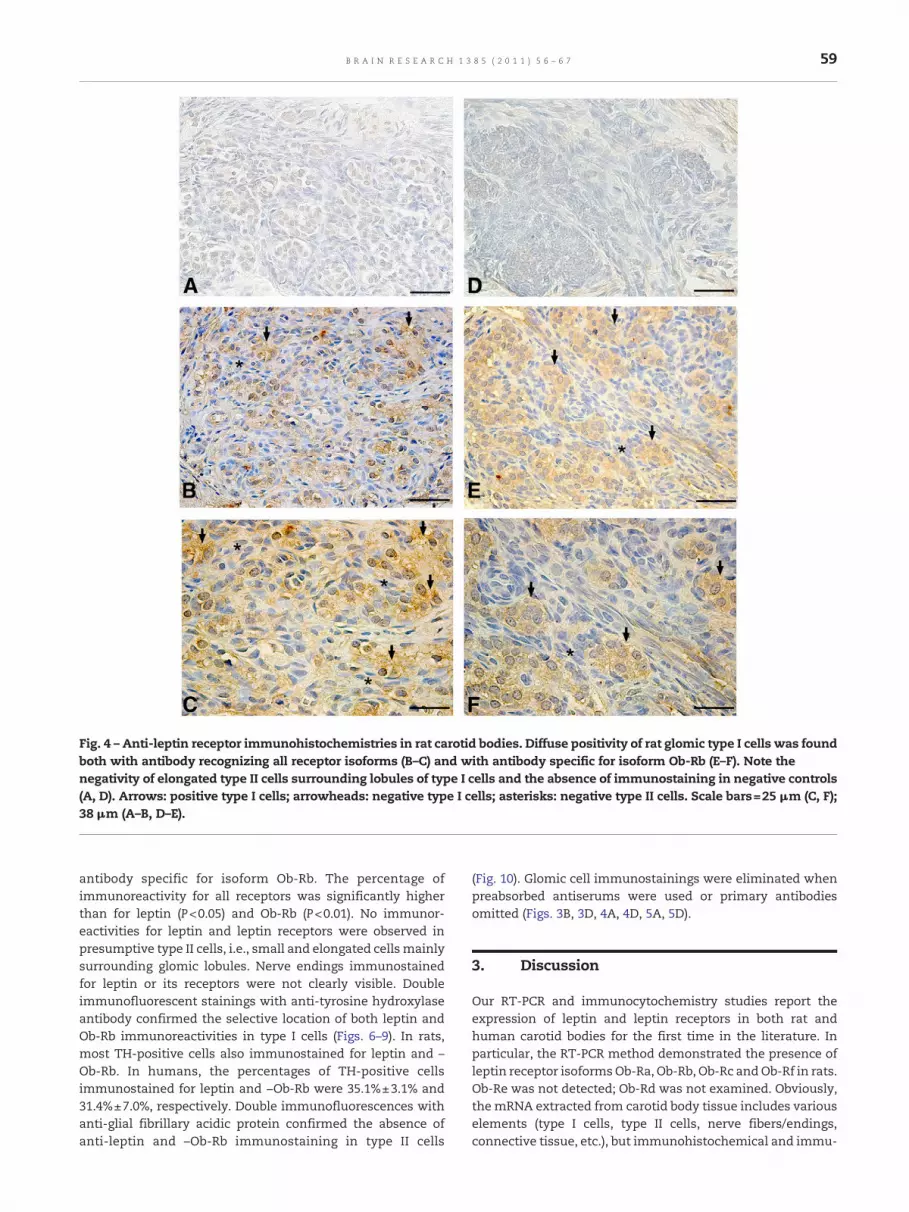
Fig. 4 – Anti-leptin receptor immunohistochemistries in rat carotid bodies. Diffuse positivity of rat glomic type I cells was foundboth with antibody recognizing all receptor isoforms (B–C) and with antibody specific for isoform Ob-Rb (E–F). Note thenegativity of elongated type II cells surrounding lobules of type I cells and the absence of immunostaining in negative controls(A, D). Arrows: positive type I cells; arrowheads: negative type I cells; asterisks: negative type II cells. Scale bars=25 μm (C, F);38 μm (A–B, D–E).
59B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
antibody specific for isoform Ob-Rb. The percentage ofimmunoreactivity for all receptors was significantly higherthan for leptin (P<0.05) and Ob-Rb (P<0.01). No immunor-eactivities for leptin and leptin receptors were observed inpresumptive type II cells, i.e., small and elongated cells mainlysurrounding glomic lobules. Nerve endings immunostainedfor leptin or its receptors were not clearly visible. Doubleimmunofluorescent stainings with anti-tyrosine hydroxylaseantibody confirmed the selective location of both leptin andOb-Rb immunoreactivities in type I cells (Figs. 6–9). In rats,most TH-positive cells also immunostained for leptin and –Ob-Rb. In humans, the percentages of TH-positive cellsimmunostained for leptin and −Ob-Rb were 35.1%±3.1% and31.4%±7.0%, respectively. Double immunofluorescences withanti-glial fibrillary acidic protein confirmed the absence ofanti-leptin and –Ob-Rb immunostaining in type II cells
(Fig. 10). Glomic cell immunostainings were eliminated whenpreabsorbed antiserums were used or primary antibodiesomitted (Figs. 3B, 3D, 4A, 4D, 5A, 5D).
3. Discussion
Our RT-PCR and immunocytochemistry studies report theexpression of leptin and leptin receptors in both rat andhuman carotid bodies for the first time in the literature. Inparticular, the RT-PCR method demonstrated the presence ofleptin receptor isoformsOb-Ra, Ob-Rb, Ob-Rc andOb-Rf in rats.Ob-Re was not detected; Ob-Rd was not examined. Obviously,the mRNA extracted from carotid body tissue includes variouselements (type I cells, type II cells, nerve fibers/endings,connective tissue, etc.), but immunohistochemical and immu-

Fig. 5 – Anti-leptin receptor immunohistochemistries in human carotid bodies. Positivity of glomic type I cells was found bothwith antibody recognizing all receptor isoforms (B–C) and with antibody specific for isoform Ob-Rb (E–F). Also in humans notethe negativity of elongated type II cells surrounding lobules of type I cells and the absence of immunostaining in negativecontrols (A, D). Arrows: positive type I cells; arrowheads: negative type I cells; asterisks: negative type II cells. Scale bars=25 μm(C, F); 38 μm (A–B, D–E).
60 B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
nofluorescence studies allowed us to locate leptin and itsreceptors in type I cells selectively. In particular, Ob-Rb isconsidered the main receptor involved in signal transductionand, although it was found to be expressed at the lowest levelamong various isoforms by semiquantitative PCR, immuno-histochemical and immunofluorescence analysis involving anantibody specific for this isoform confirmed its expression inboth human and rat type I cells.
Leptin has been reported to play a role in the control ofbreathing. Genetically obese ob/ob mice exhibit rapid breath-ing frequency, depressed hypercapnic ventilatory responseand elevated PaCO2
(Tankersley et al., 1996), and both chronicand acute leptin replacements in ob/ob mice have been foundto increase baseline ventilation and ventilatory response toCO2 (O'Donnell et al., 1999, 2000; Tankersley et al., 1998). Therole of circulating leptin in respiratory regulation is less clearin humans. Patients with obesity hypoventilation syndrome
show a higher serum leptin level than patients with obstruc-tive sleep apnea syndrome, suggesting resistance to leptinrespiratory action, perhaps due to a different penetration rateinto the cerebrospinal fluid (Caro et al., 1996; Phipps et al.,2002; Shimura et al., 2005; Yee et al., 2006). Conversely, serumleptin levels have been found to be lower in patients withobesity hypoventilation syndrome without obstructive sleepapnea than in matched obese subjects, suggesting that theblunted response to hypercapniamay derive from lower leptinlevels (Redolfi et al., 2007). The absence of depression of thehypoxic ventilatory response in ob/ob mice suggests a specificeffect of leptin on central CO2-sensing neurons more than thecarotid body, which is considered to be mainly involved inhypoxic response (Polotsky et al., 2004). However, the possibleaction of leptin on the carotid body has not yet been excludedin the literature, as it has been reported that the response tohyperoxia, which is also mainly mediated by peripheral

Fig. 6 –Double immunofluorescence for tyrosine hydroxylase(TH) and leptin in rat carotid body. TH was demonstrated inred (A) and leptin in green (B). The combination of theprevious images (C) shows colocalization of TH and leptin,demonstrating the presence of leptin in type I cells (yellow).Scale bar=12 μm.
Fig. 7 –Double immunofluorescence for tyrosine hydroxylase(TH) and leptin in human carotid body. THwas demonstratedin red (A) and leptin in green (B). The combination of theprevious images (C) shows colocalization of TH and leptin,demonstrating the expression of leptin by type I cells. Scalebar=10 μm.
61B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
chemoreceptors, is blunted by leptin deficiency and restoredby leptin replacement (Groeben et al., 2004). Our findingsabout the expression of both leptin and leptin receptors intype I cells, which represents the true chemoreceptiveelement of the carotid body, suggests that leptin plays amodulator role in peripheral chemoreception. In the carotidbody, glomic type I cells releasemany neuromodulators whichplay a role in regulating chemoreceptor discharge (Iturriagaand Alcayaga, 2004; Nurse, 2005; Porzionato et al., 2006, 2008a,2009a): on the basis of our findings, leptin could be added tothe neuromodulators expressed in type I cells, althoughexperimental data demonstrating the effects of leptin orleptin receptor agonists on chemosensory process will benecessary. Moreover, circulating factors have also beenreported to act on carotid body type I cells, and possibleaction on the carotid body by blood-borne leptin may behypothesized. In rats, leptin and leptin receptor expressions
were diffused to almost all type I cells whereas in humans onlya sub-population of type I cells were positive for leptin and/orits receptors, although it showed stronger reactions. On thebasis of this finding we could hypothesize major involvementof leptin in the chemosensory function of rats, although thismust be experimentally verified.
The control of energy homeostasis is mediated by anetwork of glucosensing neurons located in the central andperipheral nervous systems. In ventromedial and arcuatehypothalamic nuclei, glucosensing neurons may also directly

Fig. 8 –Double immunofluorescence for tyrosine hydroxylase(TH) and leptin receptor isoform b (Ob-Rb) in rat carotid body.TH was demonstrated in red (A) and Ob-Rb in green (B). Thecombination of the previous images (C) shows colocalizationof TH and Ob-Rb, demonstrating the presence of the receptorin type I cells (yellow). Scale bar=17 μm.
Fig. 9 – Double immunofluorescence for tyrosine hydroxylase(TH) and leptin receptor isoform b (Ob-Rb) in human carotidbody. TH was demonstrated in red (A) and Ob-Rb in green (B).The combination of the previous images (C) shows colocali-zation of TH and Ob-Rb in some cells (yellow), demonstratingthe presence of the receptor in type I cells. Scale bar=10 μm.
62 B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
respond to metabolites from the periphery and peptidesinvolved in energy homeostasis, such as insulin and leptin.For this reason, these neurons are now considered not onlyglucosensing but also metabolic sensing (Levin, 2001, 2006). Ithas recently been emphasized that the carotid body can alsodetect low glucose concentrations and may contribute to theproduction of counterregulatory responses to hypoglycemia(Alvarez-Buylla and de Alvarez-Buylla, 1988, 1994; Koyama etal., 2000; reviewed in López-Barneo, 2003; López-Barneo et al.,2008; Pardal and López-Barneo, 2002). However, although therole of the carotid body in glucose homeostasis is generallyaccepted, some recent works suggest that some othermetabolically derived blood-borne factors, more than glucoseper se, are involved in mediated transduction of hypoglycemicstimuli to the carotid body (Bin-Jaliah et al., 2004). Theselective expression of leptin receptors in type I cells gives
rise to the intriguing idea that leptin may modulate metabo-lism not only through central action but also throughperipheral binding to its receptors in glomic type I cells. Thecarotid body is also known to express receptors for some otherblood-borne peptides, such as vasopressin (Montero et al.,2003) and galanin (Porzionato et al., 2010a), which are involvedin metabolism regulation. Thus, we may hypothesize that thecarotid body is involved not only in glucoregulation throughsensing of short-term alterations in glucose availability(glucosensing function) but also in energy homeostasisthrough sensing of long-term signals mediated by circulatingpeptides, such as leptin, vasopressin, galanin and probablyothers (metabolic sensing function).
Apart from the possible roles played by leptin on carotidbody chemoreception and energy homeostasis, other actions

Fig. 10 – Double immunofluorescences for glial fibrillary acidic protein (GFAP) and leptin in human carotid body (A–C), and forGFAP and leptin receptor isoform b (Ob-Rb) in rat carotid body. GFAP was demonstrated in red (A, D), leptin (B) and Ob-Rb (E) ingreen. The combinations of the previous images (C, F) show the absence of anti-leptin and –Ob-Rb positivities in GFAP-positivetype II cells. Scale bars=12 μm (C), 17 μm (F).
63B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
of leptin on the carotid body may be hypothesized. Leptin hasbeen reported to modulate the survival (Dicou et al., 2001;Doherty et al., 2008) and synaptic plasticity (reviewed inHarvey et al., 2006) of various types of central and peripheralneurons. The carotid body undergoes many morphological,cellular and biochemical changes during its perinatal devel-opment and/or in response to environmental stimuli such ashypoxic, hyperoxic or inflammatory noxae (Porzionato et al.,2005, 2008a, 2008b 2009b, 2009c, 2010b). The above changes aremediated by a large series of trophic factors, to which leptincould be added.
In conclusion, identification of leptin and leptin receptorsin glomic type I cells poses new questions about the possible
peripheral modulatory roles of leptin on breathing, glucosen-sing and carotid body plasticity mechanisms, which must befurther addressed in the future.
4. Experimental procedures
4.1. Tissue sampling and preparation
Materials consisted of carotid bodies obtained at autopsy from10 adult human subjects (6 male, 4 female; mean age 46 years,standard deviation (SD)±3.6), clinically negative for chronicpulmonary or cardiovascular diseases, and carotid bodies

Table 1 – PCR primers and PCR products.
Primers used in RT-PCR and QPCR
Gene Genbank accession number Primer Sequence of primer Position PCR product length (bp)
Leptin (Ob) NM_013076 L GAC ATT TCA CAC ACG CAG TC 190–209 195R GAG GAG GTC TCG CAG GTT 366–384
Ob-Ra AF304191 L CAC TGT TAA TTT CAC ACC AGA G 110–131 235R GTC ATT CAA ACC ATA GTT TAG G 323–344
Ob-Rb U52966 L TGC TCG GAA CAC TGT TAA T 2635–2653 171R GAA GAA GAG CAA ATA TCA 2785–2805
Ob-Rc AF007818 L TGC TCG GAA CAC TGT TAA T 35–53 161R ATA GAG TAT CTA ACC TGC ACC CTT 172–195
Ob-Re AF007819 L TCC TGG ACA CTG TCA CCT AA 595–614 184R ATC AGG ATT GCC AAT TTA CA 759–778
Ob-Rf D84125 L GCT GCT CGG AAC ACT GTT AAT 2676–2696 151R ACG GCA TCC ACT CTA TAT CCT 2806–2826
PBGD NM_013168 L GAA AGA CCC TGG AAA CCT TG 522–541 148R TGC TCA TCC AGC TTC CGT A 651–669
PBGD: porphobilinogen deaminase.
64 B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
sampled from 10 adult Sprague–Dawley rats. Autopsies wereperformed between 24 and 30 h after death. Sampling fromratswas performed soon after sacrifice, in accordancewith theEuropean Communities Council Directive of 24 November1986 (86/609/EEC). For immunohistochemistry, the rightcarotid bifurcations from both humans and rats were fixedin Bouin solution and embedded in paraffin wax. For real-timePCR, the left carotid bifurcations of rats were dissected andcarotid bodies cleaned from the surrounding tissues under adissecting microscope, frozen immediately on dry ice andstored at −80 °C. One pool containing all the carotid bodiessampled for real-time PCR was constituted.
4.2. RT-PCR
Conventional RT-PCR and QPCRApplied methods have been described earlier (Rucinski et al.,2005, 2008; Ziolkowska et al., 2007). Briefly, from freshlysampled carotid bodies, total RNA was extracted by a methodusing TRI Reagent (Sigma) and purified on columns (RNeasyMini Kit, Qiagen). mRNA was extracted from total RNA by thePolyATract® mRNA Isolation System III (Promega, Madison,WI), the amount of total mRNAwas determined by measuringoptical density at 260 nm, and purity was estimated by the260/280 nm absorption ratio, which was consistently higherthan 1.8. A NanoDrop (Thermo Scientific) spectrophotometerwas used for this measurement. Reverse transcription wasperformed with AMV reverse transcriptase (Promega) withOligo dT (PE Biosystems, Warrington, UK) as primers. Reac-tions were performed at 42 °C for 60 min (Thermocycler UNOII, Biometra). Primers used for expression analysis experi-ments were designed by Primer3 software (Whitehead Insti-tute for Biomedical Research, Cambridge, MA) and amplifiedfragments spanning introns. Primerswere purchased from theLaboratory of DNA Sequencing and Oligonucleotide Synthesis,Institute of Biochemistry and Biophysics, Polish Academy ofSciences, Warsaw. The primer sequence and predicted size ofthe PCR products are listed in Table 1. For all tested genes, 3cycles of the following sequences were applied: denaturation94 °C, 30 s; annealing 58 °C, 60 s; extension 68 °C, 60 s. In
control reactions, water (instead of cDNA) was used, andreverse transcriptase (−RT) was omitted. Reaction productswere separated on ethidium bromide containing 2% agarosegel and photographed in a transilluminator.
QPCR was performed by means of the LightCycler 2.0instrument (ROCHE) with software version 4.05. The SYBRgreen detection system was used with the above-mentionedprimers. Each 20-μl reaction mixture contained 4 μl oftemplate cDNA (or standards, or controls) and 0.5 μM of eachgene-specific primer at previously determined optimal MgCl2concentration (3.5 μM for one reaction). LightCycler FastStartDNA Master SYBR Green I mix (ROCHE) was used. The real-time PCR program included a 10-min denaturation step toactivate the Taq DNA Polymerase, followed by a 3-stepamplification program: denaturation at 95 °C for 10 s, anneal-ing at 56 °C for 5 s, and extension at 72 °C for 10 s. Thespecificity of reaction products was checked by determinationof melting points (0.1 °C/s transition rate). The expression ofall tested genes was normalized to the porphobilinogendeaminase (PBGD) reference gene. Amplification of tyrosinehydroxylase mRNA was also performed, in order to confirmthe correspondence of RNA assayed with carotid body RNA.
PCR efficiency was assessed by a serial dilution method.Products of traditional RT-PCR reaction were separated on 2%agarose gel and specific bands were extracted by the DNA gelextraction kit (Millipore). The amount of extracted DNA wasestimated spectrophotometrically. Extracted DNA was diluted(10-fold serial dilutions) in order to obtain standard curves forefficiency calculation. The applied version of LightCyclersoftware (4.05) can evaluate amplification efficiency plots.Two independent reverse transcriptions (RT) were preparedfrom the RNA pool. Then two PCR were performed from eachRT, giving a total of four PCR reactions for each amplified gene.
4.3. Immunohistochemical and immunofluorescenceanalysis
Immunostaining for leptin was performed with a rabbitpolyclonal antibody (A-20: sc-842, Santa Cruz BiotechnologyInc., Santa Cruz, CA, USA) raised against a peptide mapping at

65B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
the C-terminus of leptin of human origin. Immunohistochem-istry for the leptin receptors was performedwith two differentantibodies: a goat polyclonal antibody (M-18: sc-1834, SantaCruz Biotechnology) raised against a peptide mapping at theC-terminus of the short form of Ob-R of mouse origin, whichcan detect both short and long forms of Ob-R; and a goatpolyclonal antibody (C-20: sc-1832, Santa Cruz Biotechnology)raised against a peptide mapping near the C-terminus of Ob-Rof human origin, which was specific for the long receptorisoform (Ob-Rb).
Immunohistochemical examinations were carried out on3-μm thick sections. For anti-leptin receptor immunohisto-chemistries, unmasking was performed with 10 mM sodiumcitrate buffer, pH 6.0, at 90 °C for 30 min. For anti-leptinimmunohistochemistry, antigen unmasking was not neces-sary. Sections were incubated in 0.3% hydrogen peroxide for10 min at room temperature, to remove endogenous peroxi-dase activity, and then in blocking serum for 30 min at roomtemperature. As regards primary antibody incubations, bothanti-leptin antibody and anti-leptin receptor antibodies werediluted 1:100 in blocking serum and incubated for 1 h at roomtemperature. Sectionswere thenwashed three times for 5 minin PBS. For anti-leptin immunohistochemistry, sections wererevealed with anti-rabbit serum (DAKO® EnVision+TM Per-oxidase, Rabbit, Dako Corporation, Carpinteria, CA, USA) for30 min at room temperature. For anti-leptin receptor immu-nohistochemistries, sections were revealed with anti-goatserum (Peroxidase-conjugated AffiniPure Rabbit Anti-GoatIgG, Fc Fragment Specific, Jackson ImmunoResearch Labora-tories Inc., Code N 305-035-008, West Grove, PA, USA) for30 min at room temperature. Lastly, sections were developedin 3,3′-diaminobenzidine (DAB, Sigma-Aldrich, Milan, Italy)and counterstained with hematoxylin.
In order to confirm and validate the cellular location ofleptin and leptin receptors in the carotid body, doubleimmunofluorescent stainings were performed in 4 frozencarotid bodies taken at autopsy from 4 adult human subjects(3 male, 1 female; mean age 49.3 years, SD±1.2) and 4 carotidbodies sampled from 4 adult Sprague–Dawley rats. In order tomark type I and type II cells, sections were first incubatedwithmouse anti-tyrosine hydroxylase antibody (Clone 1B5, Cat. No.MONX10786, Monosan) diluted 1:50 in PBS for 2 h at roomtemperature or mouse anti-glial fibrillary acidic proteinmonoclonal antibody (Clone GA5, Cat. No. MAB360, Chemicon)diluted 1:800 in PBS for 1 h at room temperature, respectively,and then with the above primary antibodies for leptin andleptin receptor isoform b at the same dilutions as theimmunohistochemical protocol. Sectionswere then incubatedwith Alexa fluor 594 anti-mouse (Molecular Probes, Leiden, theNetherlands) andAlexa fluor 488 anti-rabbit (Molecular Probes)or Alexa fluor 488 anti-goat (Molecular Probes). Both incuba-tionswere performed for 30 min at a concentration of 1:1000 inPBS. After thorough washing, the slides were mounted andobserved with a Leica TCSNT/SP2 confocal microscope.
Negative controls were performed by omission of primaryantibody and through overnight absorption tests with anexcess of the corresponding immunizing peptides (sc-842 P forleptin, sc-1834 P for the antibody detecting both short and longforms of Ob-R, and sc-1832 P for the antibody specific for Ob-Rb; Santa Cruz Biotechnology). Anti-leptin and –leptin recep-
tor immunoreactions detected in human mammary glandcarcinoma and rat brain were used as positive controls.
The percentages of type I and II positive cells wereevaluated on a field of 180×134 μm. Five sections and 3 fieldsper section were examined. In rat and human carotid bodies,about 40 and 25–30 type I cells were counted for each field,respectively. The mean percentages of positive cells werecalculated for each case and for the entire series.
4.4. Statistics
Percentages obtained were compared with the Kruskal–Wallistest and Dunn's multiple comparison test. A P value of 0.05was considered significant.
Acknowledgments
The authors are grateful to Gloria Sarasin and Anna Rambaldofor skillful technical assistance.
R E F E R E N C E S
Ahima, R.S., Prabakaran, D., Mantzoros, C., Qu, D., Lowell, B.,Maratos-Flier, E., Flier, J.S., 1996. Role of leptin in theneuroendocrine response to fasting. Nature 382, 250–252.
Alvarez-Buylla, R., de Alvarez-Buylla, E.R., 1988. Carotid sinusreceptors participate in glucose homeostasis. Respir. Physiol.72, 347–359.
Alvarez-Buylla, R., de Alvarez-Buylla, E.R., 1994. Changes in bloodglucose concentration in the carotid body-sinus modify brainglucose retention. Brain Res. 654, 167–170.
Banks, W.A., Kastin, A.J., Huang, W., Jaspan, J.B., Maness, L.M.,1996. Leptin enters the brain by a saturable systemindependent of insulin. Peptides 17, 305–311.
Bin-Jaliah, I., Maskell, P.D., Kumar, P., 2004. Indirect sensing ofinsulin-induced hypoglycaemia by the carotid body in the rat.J. Physiol. 556, 255–266.
Bjørbaek, C., Kahn, B.B., 2004. Leptin signaling in the centralnervous system and the periphery. Recent Prog. Horm. Res. 59,305–331.
Caro, J.F., Kolaczynski, J.W., Nyce, M.R., Ohannesian, J.P.,Opentanova, I., Goldman, W.H., Lynn, R.B., Zhang, P.L., Sinha,M.K., Considine, R.V., 1996. Decreased cerebrospinal-fluid/serum leptin ratio in obesity: a possible mechanism for leptinresistance. Lancet 348, 159–161.
Dicou, E., Attoub, S., Gressens, P., 2001. Neuroprotective effects ofleptin in vivo and in vitro. NeuroReport 12, 3947–3951.
Doherty, G.H., Oldreive, C., Harvey, J., 2008. Neuroprotectiveactions of leptin on central and peripheral neurons in vitro.Neuroscience 154, 1297–1307.
Fitzgerald, R.S., Shirahata, M., Chang, I., Kostuk, E., 2009. Theimpact of hypoxia and low glucose on the release ofacetylcholine and ATP from the incubated cat carotid body.Brain Res. 1270, 39–44.
Frederich, R.C., Hamann, A., Anderson, S., Löllmann, B., Lowell,B.B., Flier, J.S., 1995. Leptin levels reflect body lipid content inmice: evidence for diet-induced resistance to leptin action. Nat.Med. 1, 1311–1314.
Gavrilova, O., Barr, V., Marcus-Samuels, B., Reitman, M., 1997.Hyperleptinemia of pregnancy associated with the appearanceof a circulating form of the leptin receptor. J. Biol. Chem. 272,30546–30551.

66 B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
Groeben, H., Meier, S., Brown, R.H., O'Donnell, C.P., Mitzner, W.,Tankersley, C.G., 2004. The effect of leptin on the ventilatoryresponse to hyperoxia. Exp. Lung Res. 30, 559–570.
Harvey, J., Solovyova, N., Irving, A., 2006. Leptin and its role inhippocampal synaptic plasticity. Prog. Lipid Res. 45, 369–378.
Jacob, R.J., Dziura, J., Medwick, M.B., Leone, P., Caprio, S., During,M., Shulman, G.I., Sherwin, R.S., 1997. The effect of leptin isenhanced by microinjection into the ventromedialhypothalamus. Diabetes 46, 150–152.
Iturriaga, R., Alcayaga, J., 2004. Neurotransmission in the carotidbody: transmitters and modulators between glomus cells andpetrosal ganglion nerve terminals. Brain Res. Brain Res. Rev. 47,46–53.
Koyama, Y., Coker, R.H., Stone, E.E., Lacy, D.B., Jabbour, K.,Williams, P.E., Wasserman, D.H., 2000. Evidence that carotidbodies play an important role in glucoregulation in vivo.Diabetes 49, 1434–1442.
Lago, F., Dieguez, C., Gómez-Reino, J., Gualillo, O., 2007. Theemerging role of adipokines as mediators of inflammationand immune responses. Cytokine Growth Factor Rev. 18,313–325.
Lee, G.H., Proenca, R.,Montez, J.M., Carroll, K.M., Darvishzadeh, J.G.,Lee, J.I., Friedman, J.M., 1996. Abnormal splicing of the leptinreceptor in diabetic mice. Nature 379, 632–635.
Levin, B.E., 2001. Glucosensing neurons do more than just senseglucose. Int. J. Obes. Relat. Metab. Disord. 25 (Suppl 5),S68–S72.
Levin, B.E., 2006. Metabolic sensing neurons and the control ofenergy homeostasis. Physiol. Behav. 89, 486–489.
López-Barneo, J., 2003. Oxygen and glucose sensing by carotidbody glomus cells. Curr. Opin. Neurobiol. 13, 493–499.
López-Barneo, J., Ortega-Sáenz, P., Pardal, R., Pascual, A., Piruat, J.I.,2008. Carotid body oxygen sensing. Eur. Respir. J. 32, 1386–1398.
Maffei, M., Halaas, J., Ravussin, E., Pratley, R.E., Lee, G.H., Zhang, Y.,Fei, H., Kim, S., Lallone, R., Ranganathan, S., Kern, P.A.,Friedman, J.M., 1995. Leptin levels in human and rodent:measurement of plasma leptin and ob RNA in obese andweight-reduced subjects. Nat. Med. 1, 1155–1161.
Malendowicz, L.K., Rucinski, M., Belloni, A.S., Ziolkowska, A.,Nussdorfer, G.G., 2007. Leptin and the regulation of thehypothalamic-pituitary-adrenal axis. Int. Rev. Cytol. 263, 63–102.
Montero, S.A., Yarkov, A., Lemus, M., Mendoza, H., Valles, V.,de Alvarez-Buylla, E.R., Alvarez-Buylla, R., 2003. Enhancingeffect of vasopressin on the hyperglycemic response to carotidbody chemoreceptor stimulation. Role of metabolic variables.Adv. Exp. Med. Biol. 536, 95–107.
Nurse, C.A., 2005. Neurotransmission and neuromodulation in thechemosensory carotid body. Auton. Neurosci. 120, 1–9.
O'Donnell, C.P., Schaub, C.D., Haines, A.S., Berkowitz, D.E.,Tankersley, C.G., Schwartz, A.R., Smith, P.L., 1999. Leptinprevents respiratory depression in obesity. Am. J. Respir. Crit.Care Med. 159, 1477–1484.
O'Donnell, C.P., Tankersley, C.G., Polotsky, V.P., Schwartz, A.R.,Smith, P.L., 2000. Leptin, obesity, and respiratory function.Respir. Physiol. 119, 163–170.
Pardal, R., López-Barneo, J., 2002. Low glucose-sensing cells in thecarotid body. Nat. Neurosci. 5, 197–198.
Pardal, R., Ortega-Sáenz, P., Durán, R., López-Barneo, J., 2007.Glia-like stem cells sustain physiologic neurogenesis in theadult mammalian carotid body. Cell 131, 364–377.
Phipps, P.R., Starritt, E., Caterson, I., Grunstein, R.R., 2002.Association of serum leptin with hypoventilation in humanobesity. Thorax 57, 75–76.
Polotsky, V.Y., Smaldone, M.C., Scharf, M.T., Li, J., Tankersley, C.G.,Smith, P.L., Schwartz, A.R., O'Donnell, C.P., 2004. Impact ofinterrupted leptin pathways on ventilatory control. J. Appl.Physiol. 96, 991–998.
Porzionato, A., Macchi, V., Guidolin, D., Parenti, A., Ferrara, S.D.,De Caro, R., 2005. Histopathology of carotid body in heroin
addiction. Possible chemosensitive impairment. Histopathology46, 296–306.
Porzionato, A., Macchi, V., Belloni, A.S., Parenti, A., De Caro, R.,2006. Adrenomedullin immunoreactivity in the human carotidbody. Peptides 27, 69–73.
Porzionato, A., Macchi, V., Parenti, A., De Caro, R., 2008a. Trophicfactors in the carotid body. Int. Rev. Cell Mol. Biol. 269, 1–58.
Porzionato, A., Macchi, V., Parenti, A., Matturri, L., De Caro, R.,2008b. Peripheral chemoreceptors: postnatal development andcytochemical findings in sudden infant death syndrome.Histol. Histopathol. 23, 351–365.
Porzionato, A., Macchi, V., Amagliani, A., Castagliuolo, I., Parenti,A., De Caro, R., 2009a. Neurotensin receptor 1 immunoreactivityin the peripheral ganglia and carotid body. Eur. J. Histochem. 53,135–142.
Porzionato, A., Macchi, V., Parenti, A., De Caro, R., 2009b. Chroniccarotid glomitis in heroin addiction. Histol. Histopathol. 24,707–715.
Porzionato, A., Macchi, V., De Caro, R., 2009c. Sudden infant deathsyndrome. N. Engl. J. Med. 361, 2580.
Porzionato, A., Macchi, V., Barzon, L., Masi, G., Belloni, A., Parenti,A., Palù, G., De Caro, R., 2010a. Expression and distribution ofgalanin receptor subtypes in the rat carotid body. Mol. Med.Rep. 3, 37–42.
Porzionato, A., Macchi, V., Parenti, A., De Caro, R., 2010b.Extracellular signal-regulated kinase andphosphatidylinositol-3-kinase/AKT signalling pathways inthe human carotid body and peripheral ganglia. ActaHistochem. 112, 305–316.
Redolfi, S., Corda, L., La Piana, G., Spandrio, S., Prometti, P., Tantucci,C., 2007. Long-term non-invasive ventilation increaseschemosensitivity and leptin in obesity-hypoventilationsyndrome. Respir. Med. 101, 1191–1195.
Ribatti, D., Conconi, M.T., Nussdorfer, G.G., 2007. Nonclassicendogenous novel regulators of angiogenesis. Pharmacol. Rev.59, 185–205.
Rucinski, M., Andreis, P.G., Ziolkowska, A., Nussdorfer, G.G.,Malendowicz, L.K., 2005. Differential expression and functionof beacon in the rat adrenal cortex and medulla. Int. J. Mol.Med. 16, 35–40.
Rucinski, M., Zok, A., Guidolin, D., De Caro, R., Malendowicz, L.K.,2008. Expression of precerebellins in cultured rat calvariaosteoblast-like cells. Int. J. Mol. Med. 22, 553–558.
Shimura, R., Tatsumi, K., Nakamura, A., Kasahara, Y., Tanabe, N.,Takiguchi, Y., Kuriyama, T., 2005. Fat accumulation, leptin, andhypercapnia in obstructive sleep apnea–hypopnea syndrome.Chest 127, 543–549.
Tang, B.L., 2008. Leptin as a neuroprotective agent. Biochem.Biophys. Res. Commun. 368, 181–185.
Tankersley, C., Kleeberger, S., Russ, B., Schwartz, A., Smith, P.,1996. Modified control of breathing in genetically obese (ob/ob)mice. J. Appl. Physiol. 81, 716–723.
Tankersley, C.G., O'Donnell, C., Daood, M.J., Watchko, J.F., Mitzner,W., Schwartz, A., Smith, P., 1998. Leptin attenuates respiratorycomplications associated with the obese phenotype. J. Appl.Physiol. 85, 2261–2269.
Tartaglia, L.A., Dembski, M., Weng, X., Deng, N., Culpepper, J., Devos,R., Richards, G.J., Campfield, L.A., Clark, F.T., Deeds, J., Muir, C.,Sanker, S., Moriarty, A., Moore, K.J., Smutko, J.S., Mays, G.G., Wool,E.A., Monroe, C.A., Tepper, R.I., 1995. Identification and expressioncloning of a leptin receptor, OB-R. Cell 83, 1263–1271.
Tu, H., Pan, W., Feucht, L., Kastin, A.J., 2007. Convergent traffickingpattern of leptin after endocytosis mediated by ObRa-ObRd.J. Cell. Physiol. 212, 215–222.
Wang, M.Y., Zhou, Y.T., Newgard, C.B., Unger, R.H., 1996. A novelleptin receptor isoform in rat. FEBS Lett. 392, 87–90.
Yang, G., Ge, H., Boucher, A., Yu, X., Li, C., 2004. Modulation ofdirect leptin signaling by soluble leptin receptor. Mol.Endocrinol. 18, 1354–1362.

67B R A I N R E S E A R C H 1 3 8 5 ( 2 0 1 1 ) 5 6 – 6 7
Yee, B.J., Cheung, J., Phipps, P., Banerjee, D., Piper, A.J., Grunstein,R.R., 2006. Treatment of obesity hypoventilation syndrome andserum leptin. Respiration 73, 209–212.
Zhang, F., Basinski, M.B., Beals, J.M., Briggs, S.L., Churgay, L.M.,Clawson, D.K., DiMarchi, R.D., Furman, T.C., Hale, J.E., Hsiung,H.M., Schoner, B.E., Smith, D.P., Zhang, X.Y., Wery, J.P.,Schevitz, R.W., 1997. Crystal structure of the obese proteinleptin-E100. Nature 387, 206–209.
Zhang, Y., Proenca, R., Maffei, M., Barone, M., Leopold, L.,Friedman, J.M., 1994. Positional cloning of the mouseobese gene and its human homologue. Nature 372,425–432.
Ziolkowska, A., Rucinski, M., Tortorella, C., Tyczewska, M.,Nussdorfer, G.G., Malendowicz, L.K., 2007. Cultured ratcalvarial osteoblast-like cells are provided with orexin type 1receptors. Int. J. Mol. Med. 20, 779–782.