myeloid-derived micrornas, mir-223, mir27a, and mir-652, are dominant players in myeloid regulation
TRANSCRIPT

Review ArticleMyeloid-Derived microRNAs miR-223 miR27a and miR-652Are Dominant Players in Myeloid Regulation
Anna B Gilicze1 Zoltaacuten Wiener1 Saacutera Toacuteth1 Edit Buzaacutes1 Eacuteva Paacutellinger1
Franco H Falcone2 and Andraacutes Falus1
1 Department of Genetics Cell- and Immunobiology Semmelweis University Nagyvarad ter 4 Budapest 1089 Hungary2Division of Molecular amp Cellular Science The School of Pharmacy University Park Science Road Nottingham NG7 2RD UK
Correspondence should be addressed to Andras Falus falusandrasmedsemmelweis-univhu
Received 17 February 2014 Revised 29 June 2014 Accepted 16 July 2014 Published 11 August 2014
Academic Editor Ming D Li
Copyright copy 2014 Anna B Gilicze et alThis is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
In the past few years expanding knowledge has been accumulated about the role ofmicroRNAs (miRNAs) not only in hematopoiesisand cancer but also in inflammatory and infectious diseases Regarding myeloid cells our knowledge is relatively insufficienttherefore we intended to collect the available data of miRNA profiles of myeloid cells In addition to a rather general myeloidregulator miR-223 two other miRNAs seem to be useful subjects in understanding of myeloid miRNA biology miR-27a and miR-652 We review functions of these three miRNAs and other myeloid miRNAs focusing on their roles in monocytes neutrophilseosinophils basophils and mast cells
1 Introduction
MicroRNAs (miRNAs) are a family of small evolutionaryconserved and noncoding RNAs that regulate gene expres-sion negatively at the posttranscriptional levelmdashbindingcomplementary sequences within the 31015840 untranslated regionsof their target mRNA and inducing their degradation ortranslation inhibition miRNAs are generated via a special-ized pathway that involves the RNase Dicer that producesRNA duplexes of 19ndash23 nucleotides in length the maturemiRNAs [1]
miRNAs were first described as factors involved in celldevelopment and differentiation but later their importancewas verified in many other biological processes such as cellfate determination apoptosis signal transduction and organdevelopment and in various human diseases (Table 1) suchas developmental abnormalities cancer (including haemato-logical malignancies) inflammatory diseases [2 3] asthma[4] and infection [4] It is now well established that miRNAsplay a role both in the differentiation and the functionof peripheral blood cells Data are rapidly accumulatingregarding leukocytes and their miRNA signatures Accordingto a microarray study from selected immune blood cell pop-ulations there are more miRNAs in myeloid lineages than in
lymphoid lineages Although itmeans a relatively lownumberof few miRNAs they may exert significant secondary effectson the transcription of other genes by directly repressing theexpression of genes involved in translational regulation [5](Figure 1)
Innate immunity provides the first line of protectionupon pathogen invasion and it mainly involves cells ofmyeloid lineages These cells are capable of mediating rapidclearance of invading pathogens by phagocytosis and theyare rapidly recruited to local tissues at the site of infection viavarious chemokine receptors leading to a complex sequenceof inflammatory responses including vasodilatation andincreased vascular permeability [6] Abnormalities in thedevelopment ofmyeloid cellsmay cause an abnormal increasein mature myeloid or blast cells resulting in chronic or acutemyelogenous leukemia Acute myeloid leukemia (AML) isdivided into 8 subtypes based on abnormal cell types anddegree of maturity [7] Myeloid cells can facilitate angiogene-sis and tumor growth by both VEGF-dependent and VEGF-independent manners and moreover by their capability toinduce immunosuppressive effects [8]
There are a few miRNA microarray studies investigatingmiRNA patterns of blood cells Regarding miRNA pattern of
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 870267 9 pageshttpdxdoiorg1011552014870267
2 BioMed Research International
Table 1 Dominant miRNAs in human diseases
miRNA Disease ReferencemiR-223 miR-16 rheumatoid arthritis [3]miR-146 rheumatoid arthritis [9]miR-223 glioblastoma [4]miR-223 breast cancer [10]miR-27 breast cancer [11]miR-21 colorectal cancer [12]miR-223 acut myeloid leukemia [13 14]miR-146b -223 -29b-29c -483 -574-5p-672 and -690
chronic asthma [15]
miR-21 allergic airway inflammation [16]miR-223 tuberculosis [17]miR-571miR-652 liver cirrhosis [18]
miR-155 many myeloproliferativedisorders [19]
miR-155 multiple sclerosis [20]mi-R146A systemic lupus erythematosus [21]miR-7 Alzheimer disease [22]miR-132 miR-212 Alzheimer disease [23]miR-132 inflammatory bowel diseases [24]
myeloid cells there are some information about neutrophilsand monocytes a few about myeloid dendritic cells (DCs)and very few about eosinophils and mast cells The informa-tion about basophils is still lacking
There are data on the miRNA profile of 9 types ofleukocytes (including neutrophils eosinophils monocytesand mDCs) by array The authors found that nine miRNAswere specific to myeloid lineage cells miR-223 miR-143miR-145 miR-25 miR-27 miR-425 miR-17 miR-652 andmiR-191 Also an mRNA array was performed on the sameset of samples in order to determine if cell-type specificmiRNAs downregulated the expression levels of their targetgenes It was found that C-Abl oncogene 2 nonreceptortyrosine kinase (ABL2 it regulates cytoskeleton remodellingduring cell differentiation cell division and cell adhesion)was targeted by the largest number of myeloid specificmiRNAs These miRNAs may promote differentiation andgranulopoiesis by repressing ABL2 levels disrupting main-tenance of the undifferentiated myeloid precursor state [5]In another miRNA microarray study the role of miRNAswas investigated on gene expression of neutrophils afterphysical exercise Many of the exercise-affected miRNAsin neutrophils are known to regulate genes involved inimmune processes and apoptosis including miR-17 miR-18a and miR-20a (all part of the miR-17-92 cluster whichhad decreased expression level after exercise) In case ofmiR-223 the authors found elevated expression immediatelyafter exercise The authors found three major pathways inneutrophils in which both exercise-influenced miRNAs andmRNAs are involved the ubiquitin-mediated proteolysis
pathway the Jak-STATmdashand the Hedgehog signaling path-ways Each of these pathways plays a role in key mechanismsof inflammation [25] A further neutrophil study wheremiRNA expression of freshly isolated human neutrophilswas analysed by microarray found that the most abundantmiRNA in all samples at any time point was miR-223 [26]In the case of mast cells (MCs) there is only one murinemiRNA microarray study that analysed miRNA patternsduring differentiation and activation of bone-marrowderivedmast cells MiR-132 showed the highest upregulation uponactivation by IgEantigen which regulates heparin-bindingepidermal-like growth factor (HB-EGF) expression at thetranslational level Expression of miR-132 was confirmed inhuman cord-blood derived mast cells [27] In a study byTserel et al the miRNA pattern of monocytes DCs andmacrophages was analysed by microarray where miR-511miR-99b andmiR-212 were detected as the three most highlyupregulatedmiRNAs in bothDCs andmacrophages Accord-ing to the miRNA patterns monocytes are more similar tomacrophages than dendritic cells as shown by hierarchicalclustering analysis MiR-146a and miR-132 were detected asdifferentially regulated miRNAs during cell differentiation[28]
After collecting data about miRNA patterns of myeloidcells based on miRNA microarray analysis from above stud-ies there were 2 miRNAs (miR-27a and miR-652) whichseemed to be common in myeloid cells as they are expressedby at least 4 of the myeloid cells in our focus (Table 2) Inthe case of mast cells only murine data are available andwe presented miRNAs which were significantly upregulatedduring MC differentiation or could be activated by Fc120576RIcrosslinking in the case of MCs in Table 2 MiR-223 is signif-icantly downregulated during MC differentiation RegardingmiR-27a in an earlier study its expression was also observedin mature murine bone marrow-derived MCs [29] It is wellknown that miR-223 has a key role in myeloid developmentand function (Figure 1) but in the case of miR-27a and miR-652 there is no evidence for this and our knowledge aboutthe role of these miRNAs in myeloid cells is rather poorOur findings presented in this paper suggest that basophilsalso express miR-223 MiRNAs have been demonstrated tobe present in serum or plasma where they are resistant toRNase digestion and their expression is consistent amongindividuals There are specific expression patterns of serummiRNAs in certain diseases therefore serum or plasmamiRNAs can be used as fingerprints of various diseases [30]
After reviewing miR-223 miR-27a and miR-652 wedetail some other miRNAs which are known to be importantplayers in myeloid biology
2 A Dominant Player miR-223
miR-223 is an essential and central modulator of myeloiddifferentiation and may control granulocytic differentiationin humans [31]
Using the miR-223 gene targeted (KO) murine modelit was shown that miR-223 is a negative regulator of granu-locytic differentiation and activation in neutrophils MiR-223
BioMed Research International 3
CMP GMP
MDP
EoP
BasP
MCP
DCs
Monocytes Macrophages
Neutrophils
Eosinophils
Basophils
MCs
Activation ReferencesDifferentiation
Jurkin J et al J Immunol 184(9)4955-65 2010
Turner ML et al J Immunol 187(8)3911-17 2011
Tserel et al (2011)
Lindsay MA Trends Immunol 29(7)343-51 2008
Vasilatou D Eur J Haematol 84(1)1-16 2010
Johnnidis et al (2008)
Lu et al (2013) Lu et al (2013)Thomas XL et al J Allergy Clin Immunol 129(4)1064-75 2013
Own results
Molnaacuter et al (2012)
Mayoral RJ al J Immunol 182433-445 2009
miR-21
miR-17-5pmiR-20a
miR-16miR-103miR-107miR-29
miR-223miR-27
miR-223
miR-223miR-21
miR-223
miR-106amiR-155miR-424
miR-26amiR-24miR-27amiR-223
miR-34a
miR-155miR-146a
miR-155miR-146a
miR-223miR-146a
miR-223
miR-146b
miR-132miR-221miR-222
miR-223
miR-511
Figure 1 An overview on micro RNA regulation in myeloid development
100120583m
(a)
100120583m
(b)
Figure 2 Basophils purified from human peripheral blood (gt99 pure data not shown) before (a) and (b) after treatment by 10 ngmL hIL-3for 100 minutes
null (miR-223 minusY chimaeric) mice had increased numbersof granulocyte progenitors in the bone marrow and showeda hypermature and hypersensitive circulating neutrophil-likephenotype (nuclear hypersegmentation and blebbing) withan aberrant pattern of lineage-specific marker expressionThese mice also had inflammatory lung lesions and demon-strated enhanced tissue destruction following bacterial endo-toxin challenge [32] In an miRNA microarray study it wasshown that miR-223 is the most abundant and likely the mostimportant miRNA in human peripheral blood neutrophilgranulocytes [26]
In the case of eosinophils miR-223 ablation was shownto cause increased proliferation of eosinophil progenitorsdelayed differentiation and upregulation of miR-223 targetedgene insulin-like growth factor receptor (IGF1R) in them[33]
In basophils purified from human peripheral blood(gt99 pure data not shown) as shown in Figures 2(a) and2(b) our data show that after 100minutes of cell treatment byhuman interleukin-3 (hIL-3) a pronounced clustering of cellscan be observed Basophils cultured in the presence of hIL-3 formed clusters while control cells remained evenly dis-tributed in the culture As shown in Figure 3 (representativesample) both miR-16 and miR-223 were highly expressedwhile miR-155 expression was lower albeit pronounced in ourhealthy untreated human basophil samples Table 3 shows thefold changes of hIL-3 treated samples All of the investigatedmiRNAs showed reduced expression upon hIL-3 treatmentMiR-233 expression showed the most pronounced decreasein expression followed by miR-155 and miR-16
It seems that the importance of miR-223 is not confinedto granulocytic differentiation In monocytemacrophage
4 BioMed Research International
Table 2 Published miRNAs of myeloid cells
Mo Neu Eo muc MCs DCsmir-17 h [5] h [26]mir-21 h [26] m [27]mir-25 h [5] h [26] h [5]mir-24-1lowast m [27]mir-24-2lowast m [27]mir-27a h [5] h [5] h [26] h [5] m [27]mir-34a h [26] m [27] h [28]mir-93 h [5] h [5] h [26] h [5]mir-99a h [28]mir-99b h [28]mir-106a h [5]mir-106b h [5] h [5] h [26] h [5]mir-125a-5p h [5] h [28]mir-125a-3p m [27]mir-125b h [28]mir-132 m [27] h [28]mir-143 h [5]mir-145 h [5]mir-146a h [28]mir-155 h [28]mir-191 h [5] h [5] h [5]mir-193b m [27] h [28]mir-212 h [28]mir-223 h [5] h [5] h [26] h [5]mir-362-5p h [5] h [5]mir-362-3p m [27]mir-378 h [5]mir-422a h [5]mir-425 h [5] h [5] h [26] h [5]mir-494 m [27]mir-500-star h [5] m [27] h [5]mir-511 h [28]mir-532-5p h [5] m [27] h [5]mir-652 h [5] h [5] h [5] m [27]mir-720 h [26]mir-935 h [5]Momonocyte Neu neutrophil Eo eosinophil MucMCmucosal mast cellDC dendritic cellh human m murineFor references see numbers in brackets
differentiation miR-233 modulates NF-120581B activation andtargets IKK-120572 kinase a member of NF-120581B pathway Duringmonocytemacrophage differentiationmiR-223 miR-16 andmiR-15a are all decreased and IKK-120572 is increased which thencontributes to relBp52 production and repression of bothcanonical and noncanonical NF-120581B pathways Therefore itprevents macrophages from overactivation but possibly sen-sitizes them for futureNF-120581B signalling events It is suggestedthat the decrease in certain miRNAs during differentiationleads to a condition where macrophages can adequately
Table 3 The effect (fold change) of hIL-3 on miRNA expression inhuman basophils
miRNA untreated IL-3sect 119875
miR-155 137 1084 lt005miR-223 402 347 lt001miR-16 317 301 NSsect10 ngmL hIL-3 100min 119899 = 15NS Non significant
Log (relative expression to RNU48) of nontreated basophilmiRNA expression
miR-1551
10
miR-223 miR-16Lo
g ex
pres
sion
relat
ive t
o RN
U48
Figure 3 Expressions of miR-155 miR-16 and miR-223 inbasophils of healthy untreated human subjects (standardized toRNU48-GATGACCCCAGGTAACTCTGAGTGTGTCGCTGAT-GCCATCACCGCAGCGCTCTGACC) (Life Technologies) 119899 = 17
respond to infection but where additional tissue damages areprevented [34]
MiR-223 antagonises angiogenesis as its overexpressioninhibits both the vascular endothelia growth factor- (VEGF-) and basic fibroblast growth factor- (bFGF-) induced phos-phorylation of the respective receptors and activation ofprotein kinase B (PKBAkt) This phenomenon correlateswith downregulation of miR-223 target 1205731-integrin [35]
MiR-223 targets an aniridia type II protein (AN2PAX6)or oculorhombin a transcription factor present duringembryonic development and acts as a tumor suppres-sor in glioblastoma as well MiR-223 overexpression andAN2PAX6 knockdown caused growth and invasion ofglioblastoma cells and increased matrix metalloproteinasesMMP-2 MMP-9 and VEGF-A expression [4] In breastcancer miR-223 suppresses the proliferation and invasion ofcancer cells by targeting cytoplasmic activationproliferation-associated protein 1 (Caprin-1) [10]
MiR-223 expression rises incrementally during successivestages of granulocytic differentiation In this process miR-223targets the transcription factor nuclear factor 1 A (NFI-A)which is required for granulocyteneutrophil differentiationand which competes with myeloid specific factor CEBP120572 forbinding miR-223 promoter These miRNAs induce a regula-tory loop in myeloid progenitor development (commitment)and differentiation where NFI-A is downregulated by miR-223 and miR-223 is upregulated by CCAATenhancer bind-ing protein (CEBP120572) The increase in miR-223 expressionresults from the displacement of the inhibitor NFI-A by
BioMed Research International 5
CEBP120572 [31 36] Based on other data the situation seemsto be even more complex MiR-223 inhibits the transcriptionfactor myocyte enhancer factor-2 (Mef2c) which is involvedin promoting myeloid progenitor differentiation and whichseems to be responsible for the changes of the amount and thephenotype of neutrophils in gene targeted miR-223-deficient(KO) mice mentioned earlier [32] Beside CEBP myeloidtranscription factor PU1 a key regulator of haematopoi-etic differentiation also induces miR-223 expression whileerythroid transcription factor globin transcription factor1 (GATA1) represses it [37] CEBP120572 downregulates cell-cycle regulator E2F1 protein during granulopoiesis and E2F1inhibits miR-223 transcription while miR-223 targets E2F1and blocks cell cycle progression in AML cells forming anegative autoregulatory loop [13] In AML it was shownthat CEBP120572 contributes directly to the development of thedisease [38]
It was also shown that miR-223 is downregulatedin human AML [13] Runt-related transcription factor 1(RUNX1) mutations have been associated with poor clinicaloutcome in younger patients with cytogenetically normalAML In RUNX1-mutated AML patients CEBP120572 is down-regulated [39] CEBP120572 mediated upregulation of miR-223could be a possible explanation of the phenomenon thatmiR-223 is largely suppressed in cells from AML patients [14]
Interestingly in a recent study by Filkova et al [3] itwas suggested that bymonitoringmiR-16 andmiR-223 levelsthe disease outcome in early phase rheumatoid arthritis(ERA) patients could be predicted The authors found lowercirculating miR-16 miR-155 and miR-146 levels in ERAand healthy controlsrsquo samples than in those of patients withrheumatoid arthritis (RA) Furthermore a relationship wasshown between themiR-223 level and peripheral lymphocytenumber changes in ERA patients Moreover it seemed that ahigher circulating miR-223 baseline level could signify a bet-ter improvement in disease activity in ERA after therapeuticintervention [3]
In a murine model of asthma a miRNA microarrayanalysis had been used from allergen-exposed lung at 3 time-points (short- intermediate- and long term) of allergen expo-sure It was found that miR-223 was significantly upregulatedat short-time and was downregulated at intermediate-timeof aerosol exposure Moreover mRNA potentially targetedby miR-223 (Arid4b Il-6 and Lpin2) has undergone adownregulation at short-term exposure [15]
MiR-223 is abundantly expressed during active tubercu-losis (TB) in humans as it was shown by Dorhoi et al [17]based on a microarray from active pulmonary TB patientsand latently infected healthy individuals The authors foundthat similar to human TB miR-223 was upregulated inMycobacterium tuberculosis infected mice Moreover miR-223 deletion rendered resistant mice susceptible to TBand it seemed that miR-223 regulated NF120581B activity inmacrophages and cytokine release in myeloid cells duringTB As it was shown by another microarray experimentfrom lung tissue on days 14 and 21 during Mtb infectionlung influx of innate cells during TB was modulated bymiR-223 through the regulation of chemoattractants Cxcl2Ccl3 and IL-6 were identified as new targets of miR-223
Mortality in infected miR-223 KO animals was 80ndash100while it was only 10 in ldquowild-typerdquo littermates Neutrophildepletion prolonged survival of the animals therefore themajor cause of morbidity in the absence of miR-223 wasconsidered to be neutrophilic inflammation and subsequenttissue destruction It was suggested that the miR-223 targetedchemokines CXCL2 and CCL3 and the proinflammatorycytokine IL-6 played an important role in these processes Inthe long run miR-223 induced negative feedback control ofneutrophil chemotaxis [17]
Despite the general assumption of miRNA duplexes botharms of miR-223 duplex have an active role as it was shownby Kuchenbaueret al The authors found that miR-223lowast had aregulatory role in myeloid progenitor cells In a murine miR-223 KOmodel where the seed region of themiR-223 armwasinactivated the colony-forming cell output was significantlyreduced It was also shown by target predictions and a miR-223 KO-miR-223 WT microarray that miR-223 and miR-223lowast both target insulin-like growth factor (IGF1r)Therefore
they might have cooperative role on IGF1RPI3 K axis a keypathway for developmental and malignant processes Theauthors found that miR-223lowast had a higher expression inhealthy donors than in AML patients and a superior overallsurvival was significantly associated with the high miR-223lowastexpression level [40]
Beside miR-223 two other miRNAs are included inTable 2 as miRNAs which are present in 4 out of 6 typesof myeloid cells miR-27a and miR-652 Likely these threemiRNAs could be key modulators in myeloid cells as well
21 miR-27a miR-27a plays a role in granulocytic differ-entiation as it was shown that the amount of pri-miR-27ain myeloblasts was low while it was high in granulocytesMoreover it targeted the 31015840 UTR of RUNX1 transcriptionfactor a master regulator in hematopoietic development andmiR-27a inhibited the Runx1 protein during differentiation ofmyeloblasts [41] It is possible that themiR-27a-Runx1 systeminvolves a feedback loop [41 42] Interestingly a microarrayanalysis of RUNX1 mutation suggested the importance ofmiR-223 [39] It was shown in a murine asthma model (inthe case of a long-term aerosol exposure) that miR-27a wassignificantly downregulated [15]
MiR-27a promotes angiogenesis [11] and can alterthe activity of NF-120581B mitogen-activated protein kinases(MAPKs) p38 c-Jun N-terminal kinase (JNK) and extra-cellular-signal-regulated kinase (ERK) and therefore influ-ence the production of certain proinflammatory cytokines[43]
22 miR-652 Our knowledge onmiR-652 is rather poor It isspecifically regulated in serum of liver disease and is down-regulated in circulating monocytes isolated from patientscompared to controls As it was shown that stimulation ofmonocyticU937 cells with bacterial lipopolysaccharide (LPS)led to a significant downregulation of miR-652 thereforeit seems that this miRNA is involved in the mediation ofinflammatory signals in immune cells [18]
6 BioMed Research International
3 Further Myeloid miRNAs with WellCharacterized Effects
31 miR-155 MiR-155 plays a critical role during hematopoi-esis and regulates lymphocyte homeostasis and tolerance[44] In a recent study about miRNA expression profilesin leukocytes it was observed that miR-155 was specificallyexpressed in lymphoid lineage cells but it was downregulatedin neutrophils eosinophils and mDCs [5] On the otherhand it was shown that a sustained expression of miR-155 in hematopoietic stem cells could increase immaturegranulocyte numbers in vivo [19] MiR-155 was shown tobe required for normal immune function as gene targetedmiR-155-deficient mice were highly susceptible to Salmonellatyphimurium infection Rodriguezet al [45] also suggestedthat IL-4 promoter binding of c-Maf transcription factor canbe inhibited by loss-of-miR-155 This raises the intriguingpossibility that miRNAs may have an important effect evenonTh2-related humoral immune responses
MiR-155 is the most commonly overexpressed miRNA invarious malignancies It is related to cancers of breast lungliver and the lymphatic system therefore agents that effec-tively block miR-155 may have an outstanding importance incancer therapy [46]
According to a murine asthma model miR-155 is sig-nificantly upregulated at intermediate aerosol exposure [15]It is also important to note that the chromosomal regionin which the miR-155 gene is located (chromosome 21q21)has been previously shown to be associated with increasedasthma susceptibility inHutterites in SouthDakota a founderpopulation of European ancestry [47]
In recent studies it has been reported that miR-155 is ableto modulate myeloid cells in some pathological conditionsrelated to the central nervous system MiR-155 expressionis significantly increased in circulating CD14+ monocytesof multiple sclerosis (MS) patients compared to healthycontrols Transfection of miR-155 increased tumor necrosisfactor (TNF)120572 IL-6 CD80 and CCR7 expression whilethe expression of the miR-155 targeted gene suppressorof cytokine signaling protein (SOCS1) was decreased inhuman microglia On the other hand interferon (IFN)120574 wassignificantly increased in supernatants derived from CD4+ Tcells cocultured with miR-155-transfected monocyte-derivedmacrophages [20]
32 miR-146a LPS-induced elevation of miR-146a miR-155and miR-132 was observed in a microarray study of themonocytic leukemia cell line THP-1 after LPS challengeTherefore miR-146a seemed to be a negative regulator ofinflammation Toll-like receptors (TLRs) that recognize bac-terial constituents (TLR2 TLR4 and TLR5) could triggermiR-146a induction while TLRs that recognize viral nucleicacids (TLR3 TLR7 and TLR9) had little effect on miR-146aexpression [48]
In the absence of miR-146a hematopoietic stem cellhomeostasis is disrupted under aging and periodic bacterialencounters as indicated by declines of hematopoietic stemcell number and quality dysregulated hematopoietic stem
and progenitor cell proliferation and differentiation Chroni-cally it can lead to myeloproliferative disease The molecularpathway miR-146aTRAF6NF-120581BIL-6 seems to be involvedin these processes [49]
Rusca et al [50] identified a new molecular network thatcomprises specifically of NF120581B1 and miR-146a The authorshave shown that the same activation-induced miRNA canbe similarly regulated in different cell types in mast cells itcontributes to the regulation of cell homeostasis and survivalwhile in T lymphocytes it modulates T cell memory function
MiR-146a has a role in inflammatory diseases such aspsoriasis and rheumatoid arthritis and in cancers (such aspapillary thyroid carcinoma cervical cancer breast cancerpancreatic cancer and prostate cancer) [51] MiR-146a isupregulated in RA [9] it is downregulated in systemic lupuserythematosus (SLE) where it is a negative regulator of IFN120574pathway [21]
ElevatedmiR-146a expressionwas observed in circulatingCD14+ cells fromuntreatedMS patients [20]MiR-146amiR-125b miR-9 and miR-155 are upregulated in Alzheimerrsquosdisease in which this relatively small number of miRNAsmayimpact several key pathological features [22]
The multilevel effect of the micro-RNA networks is con-vincingly demonstrated by miR-146A since the complexity ofmiRNA and transcription factors are illustrated by the factthat not only the inhibition but also the overexpression ofmiR-146a leads to an inflammatory phenotype
33 miR-132 MiR-132 is expressed in mDCs [28] and inmurine MCs and it targets HB-EGF [27]
In human inflammatory bowel disease miR-132 wasfound to be a modulator of cholinergic signalling [24] InMS miR-132 expression was significantly decreased in circu-lating CD14+ monocytes [20] The miR-132miR-212PTENFOXO3a signaling pathway contributes to neuronal apoptosisin Alzheimerrsquos disease [23]
34 miR-21 miR-21 is a regulator of eosinophil progenitorgrowth MiR21++ and miR-21minusminus bone-marrow derivedeosinophils were morphologically indistinguishable fromeach other butmiR21minusminus cells had decreased growth capacitywith an increased level of apoptosis in vitro A whole genomemicroarray study of wild type- and gene targeted cellsidentified regulation of cell growth cell cycle and immuneresponse as the pathways were affected most significantlyby miR-21 [52] MiR-21 is upregulated in allergic airwayinflammation [16] MiR-21 suppresses TLR2 signaling andIL-13 can upregulate miR-21 expression in human airwayepithelial cells [53] The upregulation of miR-21 miR-142-3pmiR-142-5p and miR-223 was observed both in human andmouse allergic contact dermatitis [54] Esophageal miRNAexpression profile was investigated by microarray fromhuman esophageal tissues of eosinophilic esophagitis (EoE)patients and controls where the most upregulated miRNAswere miR-21 and miR-223 and the most downregulated onewas miR-375 In the plasma of EoE patients miR-146a miR-146b and miR-223 were upregulated [55]
BioMed Research International 7
MiR-21 has a dual role in CD4+ T cells activation-induced miR-21 supports survival of memory T cells whilemodulation of the potential homing by downregulation ofCCR7 protein expression was identified in active naive Tcells [56] One may consider that in vivo silencing of miR-21reversed cardinalmanifestations of autoimmunity in SLE andother diseases [12] suggesting thatmiRNAs have the potentialto become an effective therapeutic target in the future
In a recent study serum level of miR-21 proved to be aprognosticmarker for colorectal cancer as serummiR-21 levelcorrelated with the recurrence and mortality of colorectalcancer patients [57] In a further meta-analysis study earlierresults regarding circulating miR-21 level in cancer weregeneralized [58]
4 Conclusions and Future Perspectives
By now several studies certified that abnormal miRNAexpression is a common feature of various human diseasesand that the number of diseases in which the role of miRNAshas been confirmed increases continuously In some cancertypes certain circulating miRNAs were identified as potentialprognostic markers
In order to find out how inflammation-induced tumori-genesis works and in order to determine new therapeutic tar-gets identification and further investigation of key miRNAsinvolved in inflammation and cancer could be an expedientstrategy In the case of miR-155 it was reported that miR-155 and miR-155-related proinflammatory environment havemutatory-activity and play a role in inflammation-inducedcancers [59] MiR-146a could be one of the missing linksbetween inflammation and tumorigenesis as suggested by anincreasing number of publications that show its importanceboth in inflammatory diseases and tumors
Myeloid lineage development and differentiation path-ways are rather complex as miRNAs and transcriptional fac-tors form a complex network that is lineage-specific negativeand positive regulatory loops By these loops miRNAs areable to regulate normal hematopoietic lineage developmentMoreover the imbalance or disturbance can evoke diseasesby aberrant expression [36]
There is still a limited set of information about miRNApatterns and networks of monocytes mast cells eosinophilsand basophils in the literature while there is growing evi-dence that inflammation and infections can influence malig-nancies Better knowledge of this area of myeloid biology andpathophysiology would be cordially suggested
In this review we intended to overview information onmyeloid miRNAs based on available microarray analysisinvestigating miRNA pattern of myeloid cells We focusedon monocytes neutrophils eosinophils basophils and mastcells There are very few data about myeloid DCs in thisreview as DCs are rather different regarding their miRNAexpression We also did not discuss the miRNA regulation ofnewly identified myeloid-derived suppressor cells
We could reconfirm the key importance of miR-223 inmyeloid cells and provide novel information about miR-223expression in basophils
Even though there are only a few miRNA microarraystudies of myeloid cells in the literature based on them itseems that beside the myeloid key miRNA miR-223 miR-27a and miR-652 and some others could be also interestingregulators with the potential to become therapeutic targets inmyeloid pathology as well
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] D P Bartel ldquoMicroRNAs genomics biogenesis mechanismand functionrdquo Cell vol 116 no 2 pp 281ndash297 2004
[2] E Sonkoly M Stahle and A Pivarcsi ldquoMicroRNAs andimmunity novel players in the regulation of normal immunefunction and inflammationrdquo Seminars in Cancer Biology vol 18no 2 pp 131ndash140 2008
[3] M Filkova B Aradi L Senolt et al ldquoAssociation of circulatingmiR-223 and miR-16 with disease activity in patients with earlyrheumatoid arthritisrdquo Annals of the Rheumatic Diseases 2013
[4] B S Huang Q Z Luo Y Han X B Li L J Cao and LX Wu ldquomicroRNA-223 promotes the growth and invasionof glioblastoma cells by targeting tumor suppressor PAX6rdquoOncology Reports vol 30 no 5 pp 2263ndash2269 2013
[5] F Allantaz D T Cheng T Bergauer et al ldquoExpressionprofiling of human immune cell subsets identifies miRNA-mRNAregulatory relationships correlatedwith cell type specificexpressionrdquo PLoS ONE vol 7 no 1 Article ID e29979 2012
[6] H Kawamoto and N Minato ldquoMyeloid cellsrdquoThe InternationalJournal of Biochemistry amp Cell Biology vol 36 no 8 pp 1374ndash1379 2004
[7] A Tefferi JThiele and JWVardiman ldquoThe 2008WorldHealthOrganization classification system for myeloproliferative neo-plasms order out of chaosrdquo Cancer vol 115 no 17 pp 3842ndash3847 2009
[8] F Shojaei C Zhong X Wu L Yu and N Ferrara ldquoRole ofmyeloid cells in tumor angiogenesis and growthrdquo Trends in CellBiology vol 18 no 8 pp 372ndash378 2008
[9] T Nakasa S Miyaki A Okubo et al ldquoExpression ofMicroRNA-146 in rheumatoid arthritis synovial tissuerdquo Arthri-tis amp Rheumatism vol 58 no 5 pp 1284ndash1292 2008
[10] B Gong H Hu J Chen et al ldquoCaprin-1 is a novel microRNA-223 target for regulating the proliferation and invasion ofhuman breast cancer cellsrdquo Biomedicine amp Pharmacotherapyvol 67 no 7 pp 629ndash636 2013
[11] W Tang F Yu H Yao et al ldquomiR-27a regulates endothelialdifferentiation of breast cancer stem like cellsrdquo Oncogene vol33 no 20 pp 2629ndash2638 2013
[12] B G Garchow O B Encinas Y T Leung et al ldquoSilencing ofmicroR6-21 in vivo ameliorates autoimmune splenomegaly inlupus micerdquo EMBOMolecular Medicine vol 3 no 10 pp 605ndash615 2011
[13] J A Pulikkan V Dengler P S Peramangalam et al ldquoCell-cycleregulator E2F1 and microRNA-223 comprise an autoregulatorynegative feedback loop in acute myeloid leukemiardquo Blood vol115 no 9 pp 1768ndash1778 2010
8 BioMed Research International
[14] M Eyholzer S Schmid J A Schardt S Haefliger B UMuellerand T Pabst ldquoComplexity of miR-223 regulation by CEBPA inhuman AMLrdquo Leukemia Research vol 34 no 5 pp 672ndash6762010
[15] NGarbacki E diValentinVAHuynh-Thuet al ldquoMicroRNAsprofiling in murine models of acute and chronic asthma arelationship with mRNAs targetsrdquo PLoS ONE vol 6 no 1Article ID e16509 2011
[16] T X Lu A Munitz and M E Rothenberg ldquoMicroRNA-21 isup-regulated in allergic airway inflammation and regulates IL-12p35 expressionrdquo Journal of Immunology vol 182 no 8 pp4994ndash5002 2009
[17] A Dorhoi M Iannaccone M Farinacci et al ldquoMicroRNA-223 controls susceptibility to tuberculosis by regulating lungneutrophil recruitmentrdquo The Journal of Clinical Investigationvol 123 no 11 pp 4836ndash4848 2013
[18] C Roderburg T Mollnow B Bongaerts et al ldquoMicro-RNAprofiling in human serum reveals compartment-specific rolesof miR-571 and miR-652 in liver cirrhosisrdquo PLoS ONE vol 7no 3 Article ID e32999 2012
[19] R M OrsquoConnell D S Rao A A Chaudhuri et al ldquoSustainedexpression of microRNA-155 in hematopoietic stem cells causesa myeloproliferative disorderrdquo The Journal of ExperimentalMedicine vol 205 no 3 pp 585ndash594 2008
[20] C S Moore V T Rao B A Durafourt et al ldquomiR-155 as a mul-tiple sclerosis-relevant regulator of myeloid cell polarizationrdquoAnnals of Neurology vol 74 no 5 pp 709ndash720 2013
[21] Y Tang X Luo H Cui et al ldquoMicroRNA-146a contributes toabnormal activation of the type I interferon pathway in humanlupus by targeting the key signaling proteinsrdquo Arthritis andRheumatism vol 60 no 4 pp 1065ndash1075 2009
[22] W J Lukiw T V Andreeva A P Grigorenko and E I RogaevldquoStudying micro RNA function and dysfunction in Alzheimerrsquosdiseaserdquo Frontiers in Genetics vol 3 article 327 2013
[23] H A Wong T Veremeyko N Patel et al ldquoDe-repression ofFOXO3a death axis by microRNA-132 and -212 causes neuronalapoptosis in Alzheimers diseaserdquo Human Molecular Geneticsvol 22 no 15 pp 3077ndash3092 2013
[24] NMaharshak S Shenhar-TsarfatyNAroyo et al ldquoMicroRNA-132 modulates cholinergic signaling and inflammation inhuman inflammatory bowel diseaserdquo Inflammatory Bowel Dis-eases vol 19 no 7 pp 1346ndash1353 2013
[25] S Radom-Aizik F Zaldivar Jr S Oliver P Galassetti and DM Cooper ldquoEvidence for microRNA involvement in exercise-associated neutrophil gene expression changesrdquo Journal ofApplied Physiology vol 109 no 1 pp 252ndash261 2010
[26] J R Ward P R Heath J W Catto M K B Whyte M Miloand S A Renshaw ldquoRegulation of neutrophil senescence bymicroRNAsrdquo PLoS ONE vol 6 no 1 Article ID e15810 2011
[27] V Molnar B E Rsek Z Wiener et al ldquoMicroRNA-132 targetsHB-EGF upon IgE-mediated activation in murine and humanmast cellsrdquo Cellular and Molecular Life Sciences vol 69 no 5pp 793ndash808 2012
[28] L Tserel T Runnel K Kisand et al ldquoMicroRNA expressionprofiles of human blood monocyte-derived dendritic cells andmacrophages reveal miR-511 as putative positive regulator oftoll-like receptor 4rdquo Journal of Biological Chemistry vol 286 no30 pp 26487ndash26495 2011
[29] S Monticelli K M Ansel C Xiao et al ldquoMicroRNA profilingof the murine hematopoietic systemrdquo Genome Biology vol 6no 8 p R71 2005
[30] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[31] F Fazi A Rosa A Fatica et al ldquoA minicircuitry com-prised of microRNA-223 and transcription factors NFI-A andCEBPalpha regulates human granulopoiesisrdquo Cell vol 123 no5 pp 819ndash831 2005
[32] J B Johnnidis M H Harris R T Wheeler et al ldquoRegulationof progenitor cell proliferation and granulocyte function bymicroRNA-223rdquo Nature vol 451 no 7182 pp 1125ndash1129 2008
[33] T X Lu E Lim J A Besse et al ldquoMiR-223 deficiency increaseseosinophil progenitor proliferationrdquo Journal of Immunologyvol 190 no 4 pp 1576ndash1582 2013
[34] T Li M J Morgan S Choksi Y Zhang Y Kim and Z LiuldquoMicroRNAs modulate the noncanonical transcription factorNF-120581B pathway by regulating expression of the kinase IKK120572duringmacrophage differentiationrdquoNature Immunology vol 11no 9 pp 799ndash805 2010
[35] L Shi B Fisslthaler N Zippel et al ldquoMicroRNAs-223 antag-onises angiogenesis by targeting 1205731 integrin and prevent-ing growth factor signaling in endothelial cellsrdquo CirculationResearch 2013
[36] M El Gazzar and C E McCall ldquoMicroRNAs regulatorynetworks in myeloid lineage development and differentiationregulators of the regulatorsrdquo Immunology and Cell Biology vol90 no 6 pp 587ndash593 2012
[37] T Fukao Y Fukuda K Kiga et al ldquoAn Evolutionarily Con-served Mechanism for MicroRNA-223 Expression Revealed byMicroRNA Gene Profilingrdquo Cell vol 129 no 3 pp 617ndash6312007
[38] B U Mueller and T Pabst ldquoCEBP120572 and the pathophysiologyof acutemyeloid leukemiardquoCurrentOpinion inHematology vol13 no 1 pp 7ndash14 2006
[39] J H Mendler K Maharry M D Radmacher et al ldquoRUNX1mutations are associated with poor outcome in younger andolder patients with cytogenetically normal acute myeloidleukemia and with distinct gene and microRNA expressionsignaturesrdquo Journal of Clinical Oncology vol 30 no 25 pp3109ndash3118 2012
[40] F Kuchenbauer S M Mah M Heuser et al ldquoComprehensiveanalysis of mammalian miRNAlowastspecies and their role inmyeloid cellsrdquo Blood vol 118 no 12 pp 3350ndash3358 2011
[41] J Feng A Iwama M Satake and K Kohu ldquoMicroRNA-27 enhances differentiation of myeloblasts into granulocytesby post-transcriptionally downregulating Runx1rdquo The BritishJournal of Haematology vol 145 no 3 pp 412ndash423 2009
[42] O Ben-AmiN Pencovich J LotemD Levanon andYGronerldquoA regulatory interplay between miR-27a and Runx1 duringmegakaryopoiesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 106 no 1 pp 238ndash243 2009
[43] S Min L Li M Zhang et al ldquoTGF-120573-associated miR-27ainhibits dendritic cell-mediated differentiation of Th1 andTh17cells by TAB3 p38 MAPK MAP2K4 and MAP2K7rdquo Genes andImmunity vol 13 no 8 pp 621ndash631 2012
[44] E Tili C M Croce and J J Michaille ldquomiR-155 on thecrosstalk between inflammation and cancerrdquo InternationalReviews of Immunology vol 28 no 5 pp 264ndash284 2009
[45] A Rodriguez E Vigorito S Clare et al ldquoRequirement ofbicmicroRNA-155 for normal immune functionrdquo Science vol316 no 5824 pp 608ndash611 2007
BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 BioMed Research International
Table 1 Dominant miRNAs in human diseases
miRNA Disease ReferencemiR-223 miR-16 rheumatoid arthritis [3]miR-146 rheumatoid arthritis [9]miR-223 glioblastoma [4]miR-223 breast cancer [10]miR-27 breast cancer [11]miR-21 colorectal cancer [12]miR-223 acut myeloid leukemia [13 14]miR-146b -223 -29b-29c -483 -574-5p-672 and -690
chronic asthma [15]
miR-21 allergic airway inflammation [16]miR-223 tuberculosis [17]miR-571miR-652 liver cirrhosis [18]
miR-155 many myeloproliferativedisorders [19]
miR-155 multiple sclerosis [20]mi-R146A systemic lupus erythematosus [21]miR-7 Alzheimer disease [22]miR-132 miR-212 Alzheimer disease [23]miR-132 inflammatory bowel diseases [24]
myeloid cells there are some information about neutrophilsand monocytes a few about myeloid dendritic cells (DCs)and very few about eosinophils and mast cells The informa-tion about basophils is still lacking
There are data on the miRNA profile of 9 types ofleukocytes (including neutrophils eosinophils monocytesand mDCs) by array The authors found that nine miRNAswere specific to myeloid lineage cells miR-223 miR-143miR-145 miR-25 miR-27 miR-425 miR-17 miR-652 andmiR-191 Also an mRNA array was performed on the sameset of samples in order to determine if cell-type specificmiRNAs downregulated the expression levels of their targetgenes It was found that C-Abl oncogene 2 nonreceptortyrosine kinase (ABL2 it regulates cytoskeleton remodellingduring cell differentiation cell division and cell adhesion)was targeted by the largest number of myeloid specificmiRNAs These miRNAs may promote differentiation andgranulopoiesis by repressing ABL2 levels disrupting main-tenance of the undifferentiated myeloid precursor state [5]In another miRNA microarray study the role of miRNAswas investigated on gene expression of neutrophils afterphysical exercise Many of the exercise-affected miRNAsin neutrophils are known to regulate genes involved inimmune processes and apoptosis including miR-17 miR-18a and miR-20a (all part of the miR-17-92 cluster whichhad decreased expression level after exercise) In case ofmiR-223 the authors found elevated expression immediatelyafter exercise The authors found three major pathways inneutrophils in which both exercise-influenced miRNAs andmRNAs are involved the ubiquitin-mediated proteolysis
pathway the Jak-STATmdashand the Hedgehog signaling path-ways Each of these pathways plays a role in key mechanismsof inflammation [25] A further neutrophil study wheremiRNA expression of freshly isolated human neutrophilswas analysed by microarray found that the most abundantmiRNA in all samples at any time point was miR-223 [26]In the case of mast cells (MCs) there is only one murinemiRNA microarray study that analysed miRNA patternsduring differentiation and activation of bone-marrowderivedmast cells MiR-132 showed the highest upregulation uponactivation by IgEantigen which regulates heparin-bindingepidermal-like growth factor (HB-EGF) expression at thetranslational level Expression of miR-132 was confirmed inhuman cord-blood derived mast cells [27] In a study byTserel et al the miRNA pattern of monocytes DCs andmacrophages was analysed by microarray where miR-511miR-99b andmiR-212 were detected as the three most highlyupregulatedmiRNAs in bothDCs andmacrophages Accord-ing to the miRNA patterns monocytes are more similar tomacrophages than dendritic cells as shown by hierarchicalclustering analysis MiR-146a and miR-132 were detected asdifferentially regulated miRNAs during cell differentiation[28]
After collecting data about miRNA patterns of myeloidcells based on miRNA microarray analysis from above stud-ies there were 2 miRNAs (miR-27a and miR-652) whichseemed to be common in myeloid cells as they are expressedby at least 4 of the myeloid cells in our focus (Table 2) Inthe case of mast cells only murine data are available andwe presented miRNAs which were significantly upregulatedduring MC differentiation or could be activated by Fc120576RIcrosslinking in the case of MCs in Table 2 MiR-223 is signif-icantly downregulated during MC differentiation RegardingmiR-27a in an earlier study its expression was also observedin mature murine bone marrow-derived MCs [29] It is wellknown that miR-223 has a key role in myeloid developmentand function (Figure 1) but in the case of miR-27a and miR-652 there is no evidence for this and our knowledge aboutthe role of these miRNAs in myeloid cells is rather poorOur findings presented in this paper suggest that basophilsalso express miR-223 MiRNAs have been demonstrated tobe present in serum or plasma where they are resistant toRNase digestion and their expression is consistent amongindividuals There are specific expression patterns of serummiRNAs in certain diseases therefore serum or plasmamiRNAs can be used as fingerprints of various diseases [30]
After reviewing miR-223 miR-27a and miR-652 wedetail some other miRNAs which are known to be importantplayers in myeloid biology
2 A Dominant Player miR-223
miR-223 is an essential and central modulator of myeloiddifferentiation and may control granulocytic differentiationin humans [31]
Using the miR-223 gene targeted (KO) murine modelit was shown that miR-223 is a negative regulator of granu-locytic differentiation and activation in neutrophils MiR-223
BioMed Research International 3
CMP GMP
MDP
EoP
BasP
MCP
DCs
Monocytes Macrophages
Neutrophils
Eosinophils
Basophils
MCs
Activation ReferencesDifferentiation
Jurkin J et al J Immunol 184(9)4955-65 2010
Turner ML et al J Immunol 187(8)3911-17 2011
Tserel et al (2011)
Lindsay MA Trends Immunol 29(7)343-51 2008
Vasilatou D Eur J Haematol 84(1)1-16 2010
Johnnidis et al (2008)
Lu et al (2013) Lu et al (2013)Thomas XL et al J Allergy Clin Immunol 129(4)1064-75 2013
Own results
Molnaacuter et al (2012)
Mayoral RJ al J Immunol 182433-445 2009
miR-21
miR-17-5pmiR-20a
miR-16miR-103miR-107miR-29
miR-223miR-27
miR-223
miR-223miR-21
miR-223
miR-106amiR-155miR-424
miR-26amiR-24miR-27amiR-223
miR-34a
miR-155miR-146a
miR-155miR-146a
miR-223miR-146a
miR-223
miR-146b
miR-132miR-221miR-222
miR-223
miR-511
Figure 1 An overview on micro RNA regulation in myeloid development
100120583m
(a)
100120583m
(b)
Figure 2 Basophils purified from human peripheral blood (gt99 pure data not shown) before (a) and (b) after treatment by 10 ngmL hIL-3for 100 minutes
null (miR-223 minusY chimaeric) mice had increased numbersof granulocyte progenitors in the bone marrow and showeda hypermature and hypersensitive circulating neutrophil-likephenotype (nuclear hypersegmentation and blebbing) withan aberrant pattern of lineage-specific marker expressionThese mice also had inflammatory lung lesions and demon-strated enhanced tissue destruction following bacterial endo-toxin challenge [32] In an miRNA microarray study it wasshown that miR-223 is the most abundant and likely the mostimportant miRNA in human peripheral blood neutrophilgranulocytes [26]
In the case of eosinophils miR-223 ablation was shownto cause increased proliferation of eosinophil progenitorsdelayed differentiation and upregulation of miR-223 targetedgene insulin-like growth factor receptor (IGF1R) in them[33]
In basophils purified from human peripheral blood(gt99 pure data not shown) as shown in Figures 2(a) and2(b) our data show that after 100minutes of cell treatment byhuman interleukin-3 (hIL-3) a pronounced clustering of cellscan be observed Basophils cultured in the presence of hIL-3 formed clusters while control cells remained evenly dis-tributed in the culture As shown in Figure 3 (representativesample) both miR-16 and miR-223 were highly expressedwhile miR-155 expression was lower albeit pronounced in ourhealthy untreated human basophil samples Table 3 shows thefold changes of hIL-3 treated samples All of the investigatedmiRNAs showed reduced expression upon hIL-3 treatmentMiR-233 expression showed the most pronounced decreasein expression followed by miR-155 and miR-16
It seems that the importance of miR-223 is not confinedto granulocytic differentiation In monocytemacrophage
4 BioMed Research International
Table 2 Published miRNAs of myeloid cells
Mo Neu Eo muc MCs DCsmir-17 h [5] h [26]mir-21 h [26] m [27]mir-25 h [5] h [26] h [5]mir-24-1lowast m [27]mir-24-2lowast m [27]mir-27a h [5] h [5] h [26] h [5] m [27]mir-34a h [26] m [27] h [28]mir-93 h [5] h [5] h [26] h [5]mir-99a h [28]mir-99b h [28]mir-106a h [5]mir-106b h [5] h [5] h [26] h [5]mir-125a-5p h [5] h [28]mir-125a-3p m [27]mir-125b h [28]mir-132 m [27] h [28]mir-143 h [5]mir-145 h [5]mir-146a h [28]mir-155 h [28]mir-191 h [5] h [5] h [5]mir-193b m [27] h [28]mir-212 h [28]mir-223 h [5] h [5] h [26] h [5]mir-362-5p h [5] h [5]mir-362-3p m [27]mir-378 h [5]mir-422a h [5]mir-425 h [5] h [5] h [26] h [5]mir-494 m [27]mir-500-star h [5] m [27] h [5]mir-511 h [28]mir-532-5p h [5] m [27] h [5]mir-652 h [5] h [5] h [5] m [27]mir-720 h [26]mir-935 h [5]Momonocyte Neu neutrophil Eo eosinophil MucMCmucosal mast cellDC dendritic cellh human m murineFor references see numbers in brackets
differentiation miR-233 modulates NF-120581B activation andtargets IKK-120572 kinase a member of NF-120581B pathway Duringmonocytemacrophage differentiationmiR-223 miR-16 andmiR-15a are all decreased and IKK-120572 is increased which thencontributes to relBp52 production and repression of bothcanonical and noncanonical NF-120581B pathways Therefore itprevents macrophages from overactivation but possibly sen-sitizes them for futureNF-120581B signalling events It is suggestedthat the decrease in certain miRNAs during differentiationleads to a condition where macrophages can adequately
Table 3 The effect (fold change) of hIL-3 on miRNA expression inhuman basophils
miRNA untreated IL-3sect 119875
miR-155 137 1084 lt005miR-223 402 347 lt001miR-16 317 301 NSsect10 ngmL hIL-3 100min 119899 = 15NS Non significant
Log (relative expression to RNU48) of nontreated basophilmiRNA expression
miR-1551
10
miR-223 miR-16Lo
g ex
pres
sion
relat
ive t
o RN
U48
Figure 3 Expressions of miR-155 miR-16 and miR-223 inbasophils of healthy untreated human subjects (standardized toRNU48-GATGACCCCAGGTAACTCTGAGTGTGTCGCTGAT-GCCATCACCGCAGCGCTCTGACC) (Life Technologies) 119899 = 17
respond to infection but where additional tissue damages areprevented [34]
MiR-223 antagonises angiogenesis as its overexpressioninhibits both the vascular endothelia growth factor- (VEGF-) and basic fibroblast growth factor- (bFGF-) induced phos-phorylation of the respective receptors and activation ofprotein kinase B (PKBAkt) This phenomenon correlateswith downregulation of miR-223 target 1205731-integrin [35]
MiR-223 targets an aniridia type II protein (AN2PAX6)or oculorhombin a transcription factor present duringembryonic development and acts as a tumor suppres-sor in glioblastoma as well MiR-223 overexpression andAN2PAX6 knockdown caused growth and invasion ofglioblastoma cells and increased matrix metalloproteinasesMMP-2 MMP-9 and VEGF-A expression [4] In breastcancer miR-223 suppresses the proliferation and invasion ofcancer cells by targeting cytoplasmic activationproliferation-associated protein 1 (Caprin-1) [10]
MiR-223 expression rises incrementally during successivestages of granulocytic differentiation In this process miR-223targets the transcription factor nuclear factor 1 A (NFI-A)which is required for granulocyteneutrophil differentiationand which competes with myeloid specific factor CEBP120572 forbinding miR-223 promoter These miRNAs induce a regula-tory loop in myeloid progenitor development (commitment)and differentiation where NFI-A is downregulated by miR-223 and miR-223 is upregulated by CCAATenhancer bind-ing protein (CEBP120572) The increase in miR-223 expressionresults from the displacement of the inhibitor NFI-A by
BioMed Research International 5
CEBP120572 [31 36] Based on other data the situation seemsto be even more complex MiR-223 inhibits the transcriptionfactor myocyte enhancer factor-2 (Mef2c) which is involvedin promoting myeloid progenitor differentiation and whichseems to be responsible for the changes of the amount and thephenotype of neutrophils in gene targeted miR-223-deficient(KO) mice mentioned earlier [32] Beside CEBP myeloidtranscription factor PU1 a key regulator of haematopoi-etic differentiation also induces miR-223 expression whileerythroid transcription factor globin transcription factor1 (GATA1) represses it [37] CEBP120572 downregulates cell-cycle regulator E2F1 protein during granulopoiesis and E2F1inhibits miR-223 transcription while miR-223 targets E2F1and blocks cell cycle progression in AML cells forming anegative autoregulatory loop [13] In AML it was shownthat CEBP120572 contributes directly to the development of thedisease [38]
It was also shown that miR-223 is downregulatedin human AML [13] Runt-related transcription factor 1(RUNX1) mutations have been associated with poor clinicaloutcome in younger patients with cytogenetically normalAML In RUNX1-mutated AML patients CEBP120572 is down-regulated [39] CEBP120572 mediated upregulation of miR-223could be a possible explanation of the phenomenon thatmiR-223 is largely suppressed in cells from AML patients [14]
Interestingly in a recent study by Filkova et al [3] itwas suggested that bymonitoringmiR-16 andmiR-223 levelsthe disease outcome in early phase rheumatoid arthritis(ERA) patients could be predicted The authors found lowercirculating miR-16 miR-155 and miR-146 levels in ERAand healthy controlsrsquo samples than in those of patients withrheumatoid arthritis (RA) Furthermore a relationship wasshown between themiR-223 level and peripheral lymphocytenumber changes in ERA patients Moreover it seemed that ahigher circulating miR-223 baseline level could signify a bet-ter improvement in disease activity in ERA after therapeuticintervention [3]
In a murine model of asthma a miRNA microarrayanalysis had been used from allergen-exposed lung at 3 time-points (short- intermediate- and long term) of allergen expo-sure It was found that miR-223 was significantly upregulatedat short-time and was downregulated at intermediate-timeof aerosol exposure Moreover mRNA potentially targetedby miR-223 (Arid4b Il-6 and Lpin2) has undergone adownregulation at short-term exposure [15]
MiR-223 is abundantly expressed during active tubercu-losis (TB) in humans as it was shown by Dorhoi et al [17]based on a microarray from active pulmonary TB patientsand latently infected healthy individuals The authors foundthat similar to human TB miR-223 was upregulated inMycobacterium tuberculosis infected mice Moreover miR-223 deletion rendered resistant mice susceptible to TBand it seemed that miR-223 regulated NF120581B activity inmacrophages and cytokine release in myeloid cells duringTB As it was shown by another microarray experimentfrom lung tissue on days 14 and 21 during Mtb infectionlung influx of innate cells during TB was modulated bymiR-223 through the regulation of chemoattractants Cxcl2Ccl3 and IL-6 were identified as new targets of miR-223
Mortality in infected miR-223 KO animals was 80ndash100while it was only 10 in ldquowild-typerdquo littermates Neutrophildepletion prolonged survival of the animals therefore themajor cause of morbidity in the absence of miR-223 wasconsidered to be neutrophilic inflammation and subsequenttissue destruction It was suggested that the miR-223 targetedchemokines CXCL2 and CCL3 and the proinflammatorycytokine IL-6 played an important role in these processes Inthe long run miR-223 induced negative feedback control ofneutrophil chemotaxis [17]
Despite the general assumption of miRNA duplexes botharms of miR-223 duplex have an active role as it was shownby Kuchenbaueret al The authors found that miR-223lowast had aregulatory role in myeloid progenitor cells In a murine miR-223 KOmodel where the seed region of themiR-223 armwasinactivated the colony-forming cell output was significantlyreduced It was also shown by target predictions and a miR-223 KO-miR-223 WT microarray that miR-223 and miR-223lowast both target insulin-like growth factor (IGF1r)Therefore
they might have cooperative role on IGF1RPI3 K axis a keypathway for developmental and malignant processes Theauthors found that miR-223lowast had a higher expression inhealthy donors than in AML patients and a superior overallsurvival was significantly associated with the high miR-223lowastexpression level [40]
Beside miR-223 two other miRNAs are included inTable 2 as miRNAs which are present in 4 out of 6 typesof myeloid cells miR-27a and miR-652 Likely these threemiRNAs could be key modulators in myeloid cells as well
21 miR-27a miR-27a plays a role in granulocytic differ-entiation as it was shown that the amount of pri-miR-27ain myeloblasts was low while it was high in granulocytesMoreover it targeted the 31015840 UTR of RUNX1 transcriptionfactor a master regulator in hematopoietic development andmiR-27a inhibited the Runx1 protein during differentiation ofmyeloblasts [41] It is possible that themiR-27a-Runx1 systeminvolves a feedback loop [41 42] Interestingly a microarrayanalysis of RUNX1 mutation suggested the importance ofmiR-223 [39] It was shown in a murine asthma model (inthe case of a long-term aerosol exposure) that miR-27a wassignificantly downregulated [15]
MiR-27a promotes angiogenesis [11] and can alterthe activity of NF-120581B mitogen-activated protein kinases(MAPKs) p38 c-Jun N-terminal kinase (JNK) and extra-cellular-signal-regulated kinase (ERK) and therefore influ-ence the production of certain proinflammatory cytokines[43]
22 miR-652 Our knowledge onmiR-652 is rather poor It isspecifically regulated in serum of liver disease and is down-regulated in circulating monocytes isolated from patientscompared to controls As it was shown that stimulation ofmonocyticU937 cells with bacterial lipopolysaccharide (LPS)led to a significant downregulation of miR-652 thereforeit seems that this miRNA is involved in the mediation ofinflammatory signals in immune cells [18]
6 BioMed Research International
3 Further Myeloid miRNAs with WellCharacterized Effects
31 miR-155 MiR-155 plays a critical role during hematopoi-esis and regulates lymphocyte homeostasis and tolerance[44] In a recent study about miRNA expression profilesin leukocytes it was observed that miR-155 was specificallyexpressed in lymphoid lineage cells but it was downregulatedin neutrophils eosinophils and mDCs [5] On the otherhand it was shown that a sustained expression of miR-155 in hematopoietic stem cells could increase immaturegranulocyte numbers in vivo [19] MiR-155 was shown tobe required for normal immune function as gene targetedmiR-155-deficient mice were highly susceptible to Salmonellatyphimurium infection Rodriguezet al [45] also suggestedthat IL-4 promoter binding of c-Maf transcription factor canbe inhibited by loss-of-miR-155 This raises the intriguingpossibility that miRNAs may have an important effect evenonTh2-related humoral immune responses
MiR-155 is the most commonly overexpressed miRNA invarious malignancies It is related to cancers of breast lungliver and the lymphatic system therefore agents that effec-tively block miR-155 may have an outstanding importance incancer therapy [46]
According to a murine asthma model miR-155 is sig-nificantly upregulated at intermediate aerosol exposure [15]It is also important to note that the chromosomal regionin which the miR-155 gene is located (chromosome 21q21)has been previously shown to be associated with increasedasthma susceptibility inHutterites in SouthDakota a founderpopulation of European ancestry [47]
In recent studies it has been reported that miR-155 is ableto modulate myeloid cells in some pathological conditionsrelated to the central nervous system MiR-155 expressionis significantly increased in circulating CD14+ monocytesof multiple sclerosis (MS) patients compared to healthycontrols Transfection of miR-155 increased tumor necrosisfactor (TNF)120572 IL-6 CD80 and CCR7 expression whilethe expression of the miR-155 targeted gene suppressorof cytokine signaling protein (SOCS1) was decreased inhuman microglia On the other hand interferon (IFN)120574 wassignificantly increased in supernatants derived from CD4+ Tcells cocultured with miR-155-transfected monocyte-derivedmacrophages [20]
32 miR-146a LPS-induced elevation of miR-146a miR-155and miR-132 was observed in a microarray study of themonocytic leukemia cell line THP-1 after LPS challengeTherefore miR-146a seemed to be a negative regulator ofinflammation Toll-like receptors (TLRs) that recognize bac-terial constituents (TLR2 TLR4 and TLR5) could triggermiR-146a induction while TLRs that recognize viral nucleicacids (TLR3 TLR7 and TLR9) had little effect on miR-146aexpression [48]
In the absence of miR-146a hematopoietic stem cellhomeostasis is disrupted under aging and periodic bacterialencounters as indicated by declines of hematopoietic stemcell number and quality dysregulated hematopoietic stem
and progenitor cell proliferation and differentiation Chroni-cally it can lead to myeloproliferative disease The molecularpathway miR-146aTRAF6NF-120581BIL-6 seems to be involvedin these processes [49]
Rusca et al [50] identified a new molecular network thatcomprises specifically of NF120581B1 and miR-146a The authorshave shown that the same activation-induced miRNA canbe similarly regulated in different cell types in mast cells itcontributes to the regulation of cell homeostasis and survivalwhile in T lymphocytes it modulates T cell memory function
MiR-146a has a role in inflammatory diseases such aspsoriasis and rheumatoid arthritis and in cancers (such aspapillary thyroid carcinoma cervical cancer breast cancerpancreatic cancer and prostate cancer) [51] MiR-146a isupregulated in RA [9] it is downregulated in systemic lupuserythematosus (SLE) where it is a negative regulator of IFN120574pathway [21]
ElevatedmiR-146a expressionwas observed in circulatingCD14+ cells fromuntreatedMS patients [20]MiR-146amiR-125b miR-9 and miR-155 are upregulated in Alzheimerrsquosdisease in which this relatively small number of miRNAsmayimpact several key pathological features [22]
The multilevel effect of the micro-RNA networks is con-vincingly demonstrated by miR-146A since the complexity ofmiRNA and transcription factors are illustrated by the factthat not only the inhibition but also the overexpression ofmiR-146a leads to an inflammatory phenotype
33 miR-132 MiR-132 is expressed in mDCs [28] and inmurine MCs and it targets HB-EGF [27]
In human inflammatory bowel disease miR-132 wasfound to be a modulator of cholinergic signalling [24] InMS miR-132 expression was significantly decreased in circu-lating CD14+ monocytes [20] The miR-132miR-212PTENFOXO3a signaling pathway contributes to neuronal apoptosisin Alzheimerrsquos disease [23]
34 miR-21 miR-21 is a regulator of eosinophil progenitorgrowth MiR21++ and miR-21minusminus bone-marrow derivedeosinophils were morphologically indistinguishable fromeach other butmiR21minusminus cells had decreased growth capacitywith an increased level of apoptosis in vitro A whole genomemicroarray study of wild type- and gene targeted cellsidentified regulation of cell growth cell cycle and immuneresponse as the pathways were affected most significantlyby miR-21 [52] MiR-21 is upregulated in allergic airwayinflammation [16] MiR-21 suppresses TLR2 signaling andIL-13 can upregulate miR-21 expression in human airwayepithelial cells [53] The upregulation of miR-21 miR-142-3pmiR-142-5p and miR-223 was observed both in human andmouse allergic contact dermatitis [54] Esophageal miRNAexpression profile was investigated by microarray fromhuman esophageal tissues of eosinophilic esophagitis (EoE)patients and controls where the most upregulated miRNAswere miR-21 and miR-223 and the most downregulated onewas miR-375 In the plasma of EoE patients miR-146a miR-146b and miR-223 were upregulated [55]
BioMed Research International 7
MiR-21 has a dual role in CD4+ T cells activation-induced miR-21 supports survival of memory T cells whilemodulation of the potential homing by downregulation ofCCR7 protein expression was identified in active naive Tcells [56] One may consider that in vivo silencing of miR-21reversed cardinalmanifestations of autoimmunity in SLE andother diseases [12] suggesting thatmiRNAs have the potentialto become an effective therapeutic target in the future
In a recent study serum level of miR-21 proved to be aprognosticmarker for colorectal cancer as serummiR-21 levelcorrelated with the recurrence and mortality of colorectalcancer patients [57] In a further meta-analysis study earlierresults regarding circulating miR-21 level in cancer weregeneralized [58]
4 Conclusions and Future Perspectives
By now several studies certified that abnormal miRNAexpression is a common feature of various human diseasesand that the number of diseases in which the role of miRNAshas been confirmed increases continuously In some cancertypes certain circulating miRNAs were identified as potentialprognostic markers
In order to find out how inflammation-induced tumori-genesis works and in order to determine new therapeutic tar-gets identification and further investigation of key miRNAsinvolved in inflammation and cancer could be an expedientstrategy In the case of miR-155 it was reported that miR-155 and miR-155-related proinflammatory environment havemutatory-activity and play a role in inflammation-inducedcancers [59] MiR-146a could be one of the missing linksbetween inflammation and tumorigenesis as suggested by anincreasing number of publications that show its importanceboth in inflammatory diseases and tumors
Myeloid lineage development and differentiation path-ways are rather complex as miRNAs and transcriptional fac-tors form a complex network that is lineage-specific negativeand positive regulatory loops By these loops miRNAs areable to regulate normal hematopoietic lineage developmentMoreover the imbalance or disturbance can evoke diseasesby aberrant expression [36]
There is still a limited set of information about miRNApatterns and networks of monocytes mast cells eosinophilsand basophils in the literature while there is growing evi-dence that inflammation and infections can influence malig-nancies Better knowledge of this area of myeloid biology andpathophysiology would be cordially suggested
In this review we intended to overview information onmyeloid miRNAs based on available microarray analysisinvestigating miRNA pattern of myeloid cells We focusedon monocytes neutrophils eosinophils basophils and mastcells There are very few data about myeloid DCs in thisreview as DCs are rather different regarding their miRNAexpression We also did not discuss the miRNA regulation ofnewly identified myeloid-derived suppressor cells
We could reconfirm the key importance of miR-223 inmyeloid cells and provide novel information about miR-223expression in basophils
Even though there are only a few miRNA microarraystudies of myeloid cells in the literature based on them itseems that beside the myeloid key miRNA miR-223 miR-27a and miR-652 and some others could be also interestingregulators with the potential to become therapeutic targets inmyeloid pathology as well
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] D P Bartel ldquoMicroRNAs genomics biogenesis mechanismand functionrdquo Cell vol 116 no 2 pp 281ndash297 2004
[2] E Sonkoly M Stahle and A Pivarcsi ldquoMicroRNAs andimmunity novel players in the regulation of normal immunefunction and inflammationrdquo Seminars in Cancer Biology vol 18no 2 pp 131ndash140 2008
[3] M Filkova B Aradi L Senolt et al ldquoAssociation of circulatingmiR-223 and miR-16 with disease activity in patients with earlyrheumatoid arthritisrdquo Annals of the Rheumatic Diseases 2013
[4] B S Huang Q Z Luo Y Han X B Li L J Cao and LX Wu ldquomicroRNA-223 promotes the growth and invasionof glioblastoma cells by targeting tumor suppressor PAX6rdquoOncology Reports vol 30 no 5 pp 2263ndash2269 2013
[5] F Allantaz D T Cheng T Bergauer et al ldquoExpressionprofiling of human immune cell subsets identifies miRNA-mRNAregulatory relationships correlatedwith cell type specificexpressionrdquo PLoS ONE vol 7 no 1 Article ID e29979 2012
[6] H Kawamoto and N Minato ldquoMyeloid cellsrdquoThe InternationalJournal of Biochemistry amp Cell Biology vol 36 no 8 pp 1374ndash1379 2004
[7] A Tefferi JThiele and JWVardiman ldquoThe 2008WorldHealthOrganization classification system for myeloproliferative neo-plasms order out of chaosrdquo Cancer vol 115 no 17 pp 3842ndash3847 2009
[8] F Shojaei C Zhong X Wu L Yu and N Ferrara ldquoRole ofmyeloid cells in tumor angiogenesis and growthrdquo Trends in CellBiology vol 18 no 8 pp 372ndash378 2008
[9] T Nakasa S Miyaki A Okubo et al ldquoExpression ofMicroRNA-146 in rheumatoid arthritis synovial tissuerdquo Arthri-tis amp Rheumatism vol 58 no 5 pp 1284ndash1292 2008
[10] B Gong H Hu J Chen et al ldquoCaprin-1 is a novel microRNA-223 target for regulating the proliferation and invasion ofhuman breast cancer cellsrdquo Biomedicine amp Pharmacotherapyvol 67 no 7 pp 629ndash636 2013
[11] W Tang F Yu H Yao et al ldquomiR-27a regulates endothelialdifferentiation of breast cancer stem like cellsrdquo Oncogene vol33 no 20 pp 2629ndash2638 2013
[12] B G Garchow O B Encinas Y T Leung et al ldquoSilencing ofmicroR6-21 in vivo ameliorates autoimmune splenomegaly inlupus micerdquo EMBOMolecular Medicine vol 3 no 10 pp 605ndash615 2011
[13] J A Pulikkan V Dengler P S Peramangalam et al ldquoCell-cycleregulator E2F1 and microRNA-223 comprise an autoregulatorynegative feedback loop in acute myeloid leukemiardquo Blood vol115 no 9 pp 1768ndash1778 2010
8 BioMed Research International
[14] M Eyholzer S Schmid J A Schardt S Haefliger B UMuellerand T Pabst ldquoComplexity of miR-223 regulation by CEBPA inhuman AMLrdquo Leukemia Research vol 34 no 5 pp 672ndash6762010
[15] NGarbacki E diValentinVAHuynh-Thuet al ldquoMicroRNAsprofiling in murine models of acute and chronic asthma arelationship with mRNAs targetsrdquo PLoS ONE vol 6 no 1Article ID e16509 2011
[16] T X Lu A Munitz and M E Rothenberg ldquoMicroRNA-21 isup-regulated in allergic airway inflammation and regulates IL-12p35 expressionrdquo Journal of Immunology vol 182 no 8 pp4994ndash5002 2009
[17] A Dorhoi M Iannaccone M Farinacci et al ldquoMicroRNA-223 controls susceptibility to tuberculosis by regulating lungneutrophil recruitmentrdquo The Journal of Clinical Investigationvol 123 no 11 pp 4836ndash4848 2013
[18] C Roderburg T Mollnow B Bongaerts et al ldquoMicro-RNAprofiling in human serum reveals compartment-specific rolesof miR-571 and miR-652 in liver cirrhosisrdquo PLoS ONE vol 7no 3 Article ID e32999 2012
[19] R M OrsquoConnell D S Rao A A Chaudhuri et al ldquoSustainedexpression of microRNA-155 in hematopoietic stem cells causesa myeloproliferative disorderrdquo The Journal of ExperimentalMedicine vol 205 no 3 pp 585ndash594 2008
[20] C S Moore V T Rao B A Durafourt et al ldquomiR-155 as a mul-tiple sclerosis-relevant regulator of myeloid cell polarizationrdquoAnnals of Neurology vol 74 no 5 pp 709ndash720 2013
[21] Y Tang X Luo H Cui et al ldquoMicroRNA-146a contributes toabnormal activation of the type I interferon pathway in humanlupus by targeting the key signaling proteinsrdquo Arthritis andRheumatism vol 60 no 4 pp 1065ndash1075 2009
[22] W J Lukiw T V Andreeva A P Grigorenko and E I RogaevldquoStudying micro RNA function and dysfunction in Alzheimerrsquosdiseaserdquo Frontiers in Genetics vol 3 article 327 2013
[23] H A Wong T Veremeyko N Patel et al ldquoDe-repression ofFOXO3a death axis by microRNA-132 and -212 causes neuronalapoptosis in Alzheimers diseaserdquo Human Molecular Geneticsvol 22 no 15 pp 3077ndash3092 2013
[24] NMaharshak S Shenhar-TsarfatyNAroyo et al ldquoMicroRNA-132 modulates cholinergic signaling and inflammation inhuman inflammatory bowel diseaserdquo Inflammatory Bowel Dis-eases vol 19 no 7 pp 1346ndash1353 2013
[25] S Radom-Aizik F Zaldivar Jr S Oliver P Galassetti and DM Cooper ldquoEvidence for microRNA involvement in exercise-associated neutrophil gene expression changesrdquo Journal ofApplied Physiology vol 109 no 1 pp 252ndash261 2010
[26] J R Ward P R Heath J W Catto M K B Whyte M Miloand S A Renshaw ldquoRegulation of neutrophil senescence bymicroRNAsrdquo PLoS ONE vol 6 no 1 Article ID e15810 2011
[27] V Molnar B E Rsek Z Wiener et al ldquoMicroRNA-132 targetsHB-EGF upon IgE-mediated activation in murine and humanmast cellsrdquo Cellular and Molecular Life Sciences vol 69 no 5pp 793ndash808 2012
[28] L Tserel T Runnel K Kisand et al ldquoMicroRNA expressionprofiles of human blood monocyte-derived dendritic cells andmacrophages reveal miR-511 as putative positive regulator oftoll-like receptor 4rdquo Journal of Biological Chemistry vol 286 no30 pp 26487ndash26495 2011
[29] S Monticelli K M Ansel C Xiao et al ldquoMicroRNA profilingof the murine hematopoietic systemrdquo Genome Biology vol 6no 8 p R71 2005
[30] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[31] F Fazi A Rosa A Fatica et al ldquoA minicircuitry com-prised of microRNA-223 and transcription factors NFI-A andCEBPalpha regulates human granulopoiesisrdquo Cell vol 123 no5 pp 819ndash831 2005
[32] J B Johnnidis M H Harris R T Wheeler et al ldquoRegulationof progenitor cell proliferation and granulocyte function bymicroRNA-223rdquo Nature vol 451 no 7182 pp 1125ndash1129 2008
[33] T X Lu E Lim J A Besse et al ldquoMiR-223 deficiency increaseseosinophil progenitor proliferationrdquo Journal of Immunologyvol 190 no 4 pp 1576ndash1582 2013
[34] T Li M J Morgan S Choksi Y Zhang Y Kim and Z LiuldquoMicroRNAs modulate the noncanonical transcription factorNF-120581B pathway by regulating expression of the kinase IKK120572duringmacrophage differentiationrdquoNature Immunology vol 11no 9 pp 799ndash805 2010
[35] L Shi B Fisslthaler N Zippel et al ldquoMicroRNAs-223 antag-onises angiogenesis by targeting 1205731 integrin and prevent-ing growth factor signaling in endothelial cellsrdquo CirculationResearch 2013
[36] M El Gazzar and C E McCall ldquoMicroRNAs regulatorynetworks in myeloid lineage development and differentiationregulators of the regulatorsrdquo Immunology and Cell Biology vol90 no 6 pp 587ndash593 2012
[37] T Fukao Y Fukuda K Kiga et al ldquoAn Evolutionarily Con-served Mechanism for MicroRNA-223 Expression Revealed byMicroRNA Gene Profilingrdquo Cell vol 129 no 3 pp 617ndash6312007
[38] B U Mueller and T Pabst ldquoCEBP120572 and the pathophysiologyof acutemyeloid leukemiardquoCurrentOpinion inHematology vol13 no 1 pp 7ndash14 2006
[39] J H Mendler K Maharry M D Radmacher et al ldquoRUNX1mutations are associated with poor outcome in younger andolder patients with cytogenetically normal acute myeloidleukemia and with distinct gene and microRNA expressionsignaturesrdquo Journal of Clinical Oncology vol 30 no 25 pp3109ndash3118 2012
[40] F Kuchenbauer S M Mah M Heuser et al ldquoComprehensiveanalysis of mammalian miRNAlowastspecies and their role inmyeloid cellsrdquo Blood vol 118 no 12 pp 3350ndash3358 2011
[41] J Feng A Iwama M Satake and K Kohu ldquoMicroRNA-27 enhances differentiation of myeloblasts into granulocytesby post-transcriptionally downregulating Runx1rdquo The BritishJournal of Haematology vol 145 no 3 pp 412ndash423 2009
[42] O Ben-AmiN Pencovich J LotemD Levanon andYGronerldquoA regulatory interplay between miR-27a and Runx1 duringmegakaryopoiesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 106 no 1 pp 238ndash243 2009
[43] S Min L Li M Zhang et al ldquoTGF-120573-associated miR-27ainhibits dendritic cell-mediated differentiation of Th1 andTh17cells by TAB3 p38 MAPK MAP2K4 and MAP2K7rdquo Genes andImmunity vol 13 no 8 pp 621ndash631 2012
[44] E Tili C M Croce and J J Michaille ldquomiR-155 on thecrosstalk between inflammation and cancerrdquo InternationalReviews of Immunology vol 28 no 5 pp 264ndash284 2009
[45] A Rodriguez E Vigorito S Clare et al ldquoRequirement ofbicmicroRNA-155 for normal immune functionrdquo Science vol316 no 5824 pp 608ndash611 2007
BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
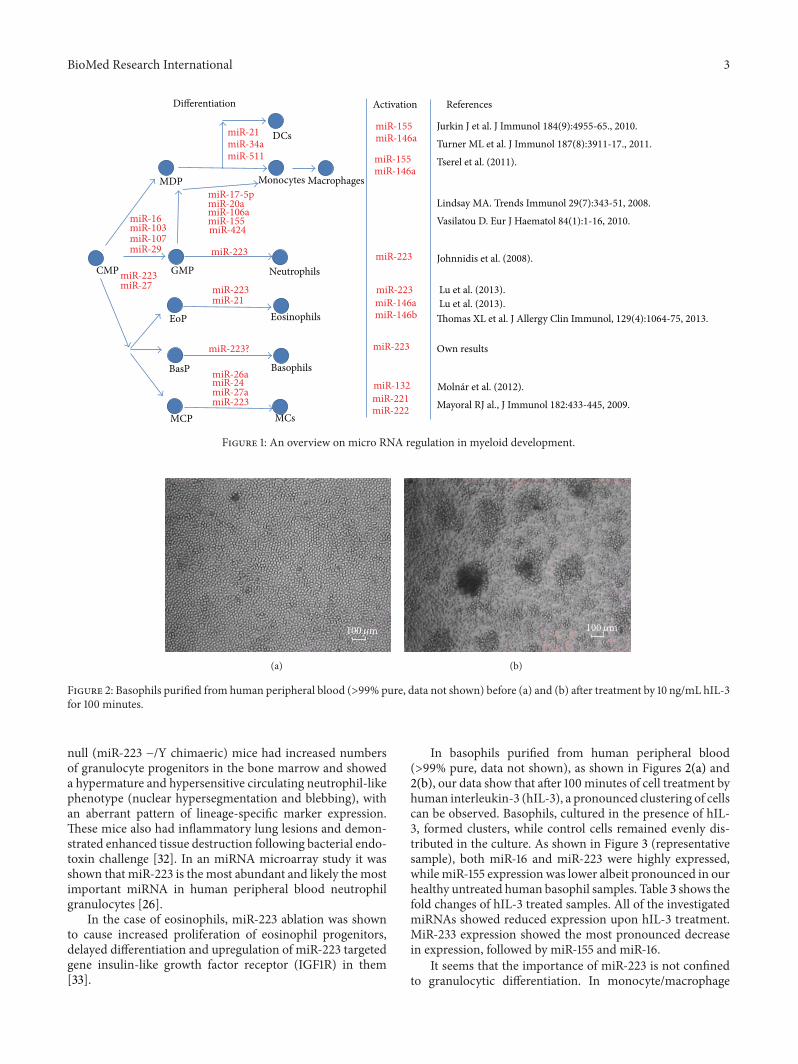
BioMed Research International 3
CMP GMP
MDP
EoP
BasP
MCP
DCs
Monocytes Macrophages
Neutrophils
Eosinophils
Basophils
MCs
Activation ReferencesDifferentiation
Jurkin J et al J Immunol 184(9)4955-65 2010
Turner ML et al J Immunol 187(8)3911-17 2011
Tserel et al (2011)
Lindsay MA Trends Immunol 29(7)343-51 2008
Vasilatou D Eur J Haematol 84(1)1-16 2010
Johnnidis et al (2008)
Lu et al (2013) Lu et al (2013)Thomas XL et al J Allergy Clin Immunol 129(4)1064-75 2013
Own results
Molnaacuter et al (2012)
Mayoral RJ al J Immunol 182433-445 2009
miR-21
miR-17-5pmiR-20a
miR-16miR-103miR-107miR-29
miR-223miR-27
miR-223
miR-223miR-21
miR-223
miR-106amiR-155miR-424
miR-26amiR-24miR-27amiR-223
miR-34a
miR-155miR-146a
miR-155miR-146a
miR-223miR-146a
miR-223
miR-146b
miR-132miR-221miR-222
miR-223
miR-511
Figure 1 An overview on micro RNA regulation in myeloid development
100120583m
(a)
100120583m
(b)
Figure 2 Basophils purified from human peripheral blood (gt99 pure data not shown) before (a) and (b) after treatment by 10 ngmL hIL-3for 100 minutes
null (miR-223 minusY chimaeric) mice had increased numbersof granulocyte progenitors in the bone marrow and showeda hypermature and hypersensitive circulating neutrophil-likephenotype (nuclear hypersegmentation and blebbing) withan aberrant pattern of lineage-specific marker expressionThese mice also had inflammatory lung lesions and demon-strated enhanced tissue destruction following bacterial endo-toxin challenge [32] In an miRNA microarray study it wasshown that miR-223 is the most abundant and likely the mostimportant miRNA in human peripheral blood neutrophilgranulocytes [26]
In the case of eosinophils miR-223 ablation was shownto cause increased proliferation of eosinophil progenitorsdelayed differentiation and upregulation of miR-223 targetedgene insulin-like growth factor receptor (IGF1R) in them[33]
In basophils purified from human peripheral blood(gt99 pure data not shown) as shown in Figures 2(a) and2(b) our data show that after 100minutes of cell treatment byhuman interleukin-3 (hIL-3) a pronounced clustering of cellscan be observed Basophils cultured in the presence of hIL-3 formed clusters while control cells remained evenly dis-tributed in the culture As shown in Figure 3 (representativesample) both miR-16 and miR-223 were highly expressedwhile miR-155 expression was lower albeit pronounced in ourhealthy untreated human basophil samples Table 3 shows thefold changes of hIL-3 treated samples All of the investigatedmiRNAs showed reduced expression upon hIL-3 treatmentMiR-233 expression showed the most pronounced decreasein expression followed by miR-155 and miR-16
It seems that the importance of miR-223 is not confinedto granulocytic differentiation In monocytemacrophage
4 BioMed Research International
Table 2 Published miRNAs of myeloid cells
Mo Neu Eo muc MCs DCsmir-17 h [5] h [26]mir-21 h [26] m [27]mir-25 h [5] h [26] h [5]mir-24-1lowast m [27]mir-24-2lowast m [27]mir-27a h [5] h [5] h [26] h [5] m [27]mir-34a h [26] m [27] h [28]mir-93 h [5] h [5] h [26] h [5]mir-99a h [28]mir-99b h [28]mir-106a h [5]mir-106b h [5] h [5] h [26] h [5]mir-125a-5p h [5] h [28]mir-125a-3p m [27]mir-125b h [28]mir-132 m [27] h [28]mir-143 h [5]mir-145 h [5]mir-146a h [28]mir-155 h [28]mir-191 h [5] h [5] h [5]mir-193b m [27] h [28]mir-212 h [28]mir-223 h [5] h [5] h [26] h [5]mir-362-5p h [5] h [5]mir-362-3p m [27]mir-378 h [5]mir-422a h [5]mir-425 h [5] h [5] h [26] h [5]mir-494 m [27]mir-500-star h [5] m [27] h [5]mir-511 h [28]mir-532-5p h [5] m [27] h [5]mir-652 h [5] h [5] h [5] m [27]mir-720 h [26]mir-935 h [5]Momonocyte Neu neutrophil Eo eosinophil MucMCmucosal mast cellDC dendritic cellh human m murineFor references see numbers in brackets
differentiation miR-233 modulates NF-120581B activation andtargets IKK-120572 kinase a member of NF-120581B pathway Duringmonocytemacrophage differentiationmiR-223 miR-16 andmiR-15a are all decreased and IKK-120572 is increased which thencontributes to relBp52 production and repression of bothcanonical and noncanonical NF-120581B pathways Therefore itprevents macrophages from overactivation but possibly sen-sitizes them for futureNF-120581B signalling events It is suggestedthat the decrease in certain miRNAs during differentiationleads to a condition where macrophages can adequately
Table 3 The effect (fold change) of hIL-3 on miRNA expression inhuman basophils
miRNA untreated IL-3sect 119875
miR-155 137 1084 lt005miR-223 402 347 lt001miR-16 317 301 NSsect10 ngmL hIL-3 100min 119899 = 15NS Non significant
Log (relative expression to RNU48) of nontreated basophilmiRNA expression
miR-1551
10
miR-223 miR-16Lo
g ex
pres
sion
relat
ive t
o RN
U48
Figure 3 Expressions of miR-155 miR-16 and miR-223 inbasophils of healthy untreated human subjects (standardized toRNU48-GATGACCCCAGGTAACTCTGAGTGTGTCGCTGAT-GCCATCACCGCAGCGCTCTGACC) (Life Technologies) 119899 = 17
respond to infection but where additional tissue damages areprevented [34]
MiR-223 antagonises angiogenesis as its overexpressioninhibits both the vascular endothelia growth factor- (VEGF-) and basic fibroblast growth factor- (bFGF-) induced phos-phorylation of the respective receptors and activation ofprotein kinase B (PKBAkt) This phenomenon correlateswith downregulation of miR-223 target 1205731-integrin [35]
MiR-223 targets an aniridia type II protein (AN2PAX6)or oculorhombin a transcription factor present duringembryonic development and acts as a tumor suppres-sor in glioblastoma as well MiR-223 overexpression andAN2PAX6 knockdown caused growth and invasion ofglioblastoma cells and increased matrix metalloproteinasesMMP-2 MMP-9 and VEGF-A expression [4] In breastcancer miR-223 suppresses the proliferation and invasion ofcancer cells by targeting cytoplasmic activationproliferation-associated protein 1 (Caprin-1) [10]
MiR-223 expression rises incrementally during successivestages of granulocytic differentiation In this process miR-223targets the transcription factor nuclear factor 1 A (NFI-A)which is required for granulocyteneutrophil differentiationand which competes with myeloid specific factor CEBP120572 forbinding miR-223 promoter These miRNAs induce a regula-tory loop in myeloid progenitor development (commitment)and differentiation where NFI-A is downregulated by miR-223 and miR-223 is upregulated by CCAATenhancer bind-ing protein (CEBP120572) The increase in miR-223 expressionresults from the displacement of the inhibitor NFI-A by
BioMed Research International 5
CEBP120572 [31 36] Based on other data the situation seemsto be even more complex MiR-223 inhibits the transcriptionfactor myocyte enhancer factor-2 (Mef2c) which is involvedin promoting myeloid progenitor differentiation and whichseems to be responsible for the changes of the amount and thephenotype of neutrophils in gene targeted miR-223-deficient(KO) mice mentioned earlier [32] Beside CEBP myeloidtranscription factor PU1 a key regulator of haematopoi-etic differentiation also induces miR-223 expression whileerythroid transcription factor globin transcription factor1 (GATA1) represses it [37] CEBP120572 downregulates cell-cycle regulator E2F1 protein during granulopoiesis and E2F1inhibits miR-223 transcription while miR-223 targets E2F1and blocks cell cycle progression in AML cells forming anegative autoregulatory loop [13] In AML it was shownthat CEBP120572 contributes directly to the development of thedisease [38]
It was also shown that miR-223 is downregulatedin human AML [13] Runt-related transcription factor 1(RUNX1) mutations have been associated with poor clinicaloutcome in younger patients with cytogenetically normalAML In RUNX1-mutated AML patients CEBP120572 is down-regulated [39] CEBP120572 mediated upregulation of miR-223could be a possible explanation of the phenomenon thatmiR-223 is largely suppressed in cells from AML patients [14]
Interestingly in a recent study by Filkova et al [3] itwas suggested that bymonitoringmiR-16 andmiR-223 levelsthe disease outcome in early phase rheumatoid arthritis(ERA) patients could be predicted The authors found lowercirculating miR-16 miR-155 and miR-146 levels in ERAand healthy controlsrsquo samples than in those of patients withrheumatoid arthritis (RA) Furthermore a relationship wasshown between themiR-223 level and peripheral lymphocytenumber changes in ERA patients Moreover it seemed that ahigher circulating miR-223 baseline level could signify a bet-ter improvement in disease activity in ERA after therapeuticintervention [3]
In a murine model of asthma a miRNA microarrayanalysis had been used from allergen-exposed lung at 3 time-points (short- intermediate- and long term) of allergen expo-sure It was found that miR-223 was significantly upregulatedat short-time and was downregulated at intermediate-timeof aerosol exposure Moreover mRNA potentially targetedby miR-223 (Arid4b Il-6 and Lpin2) has undergone adownregulation at short-term exposure [15]
MiR-223 is abundantly expressed during active tubercu-losis (TB) in humans as it was shown by Dorhoi et al [17]based on a microarray from active pulmonary TB patientsand latently infected healthy individuals The authors foundthat similar to human TB miR-223 was upregulated inMycobacterium tuberculosis infected mice Moreover miR-223 deletion rendered resistant mice susceptible to TBand it seemed that miR-223 regulated NF120581B activity inmacrophages and cytokine release in myeloid cells duringTB As it was shown by another microarray experimentfrom lung tissue on days 14 and 21 during Mtb infectionlung influx of innate cells during TB was modulated bymiR-223 through the regulation of chemoattractants Cxcl2Ccl3 and IL-6 were identified as new targets of miR-223
Mortality in infected miR-223 KO animals was 80ndash100while it was only 10 in ldquowild-typerdquo littermates Neutrophildepletion prolonged survival of the animals therefore themajor cause of morbidity in the absence of miR-223 wasconsidered to be neutrophilic inflammation and subsequenttissue destruction It was suggested that the miR-223 targetedchemokines CXCL2 and CCL3 and the proinflammatorycytokine IL-6 played an important role in these processes Inthe long run miR-223 induced negative feedback control ofneutrophil chemotaxis [17]
Despite the general assumption of miRNA duplexes botharms of miR-223 duplex have an active role as it was shownby Kuchenbaueret al The authors found that miR-223lowast had aregulatory role in myeloid progenitor cells In a murine miR-223 KOmodel where the seed region of themiR-223 armwasinactivated the colony-forming cell output was significantlyreduced It was also shown by target predictions and a miR-223 KO-miR-223 WT microarray that miR-223 and miR-223lowast both target insulin-like growth factor (IGF1r)Therefore
they might have cooperative role on IGF1RPI3 K axis a keypathway for developmental and malignant processes Theauthors found that miR-223lowast had a higher expression inhealthy donors than in AML patients and a superior overallsurvival was significantly associated with the high miR-223lowastexpression level [40]
Beside miR-223 two other miRNAs are included inTable 2 as miRNAs which are present in 4 out of 6 typesof myeloid cells miR-27a and miR-652 Likely these threemiRNAs could be key modulators in myeloid cells as well
21 miR-27a miR-27a plays a role in granulocytic differ-entiation as it was shown that the amount of pri-miR-27ain myeloblasts was low while it was high in granulocytesMoreover it targeted the 31015840 UTR of RUNX1 transcriptionfactor a master regulator in hematopoietic development andmiR-27a inhibited the Runx1 protein during differentiation ofmyeloblasts [41] It is possible that themiR-27a-Runx1 systeminvolves a feedback loop [41 42] Interestingly a microarrayanalysis of RUNX1 mutation suggested the importance ofmiR-223 [39] It was shown in a murine asthma model (inthe case of a long-term aerosol exposure) that miR-27a wassignificantly downregulated [15]
MiR-27a promotes angiogenesis [11] and can alterthe activity of NF-120581B mitogen-activated protein kinases(MAPKs) p38 c-Jun N-terminal kinase (JNK) and extra-cellular-signal-regulated kinase (ERK) and therefore influ-ence the production of certain proinflammatory cytokines[43]
22 miR-652 Our knowledge onmiR-652 is rather poor It isspecifically regulated in serum of liver disease and is down-regulated in circulating monocytes isolated from patientscompared to controls As it was shown that stimulation ofmonocyticU937 cells with bacterial lipopolysaccharide (LPS)led to a significant downregulation of miR-652 thereforeit seems that this miRNA is involved in the mediation ofinflammatory signals in immune cells [18]
6 BioMed Research International
3 Further Myeloid miRNAs with WellCharacterized Effects
31 miR-155 MiR-155 plays a critical role during hematopoi-esis and regulates lymphocyte homeostasis and tolerance[44] In a recent study about miRNA expression profilesin leukocytes it was observed that miR-155 was specificallyexpressed in lymphoid lineage cells but it was downregulatedin neutrophils eosinophils and mDCs [5] On the otherhand it was shown that a sustained expression of miR-155 in hematopoietic stem cells could increase immaturegranulocyte numbers in vivo [19] MiR-155 was shown tobe required for normal immune function as gene targetedmiR-155-deficient mice were highly susceptible to Salmonellatyphimurium infection Rodriguezet al [45] also suggestedthat IL-4 promoter binding of c-Maf transcription factor canbe inhibited by loss-of-miR-155 This raises the intriguingpossibility that miRNAs may have an important effect evenonTh2-related humoral immune responses
MiR-155 is the most commonly overexpressed miRNA invarious malignancies It is related to cancers of breast lungliver and the lymphatic system therefore agents that effec-tively block miR-155 may have an outstanding importance incancer therapy [46]
According to a murine asthma model miR-155 is sig-nificantly upregulated at intermediate aerosol exposure [15]It is also important to note that the chromosomal regionin which the miR-155 gene is located (chromosome 21q21)has been previously shown to be associated with increasedasthma susceptibility inHutterites in SouthDakota a founderpopulation of European ancestry [47]
In recent studies it has been reported that miR-155 is ableto modulate myeloid cells in some pathological conditionsrelated to the central nervous system MiR-155 expressionis significantly increased in circulating CD14+ monocytesof multiple sclerosis (MS) patients compared to healthycontrols Transfection of miR-155 increased tumor necrosisfactor (TNF)120572 IL-6 CD80 and CCR7 expression whilethe expression of the miR-155 targeted gene suppressorof cytokine signaling protein (SOCS1) was decreased inhuman microglia On the other hand interferon (IFN)120574 wassignificantly increased in supernatants derived from CD4+ Tcells cocultured with miR-155-transfected monocyte-derivedmacrophages [20]
32 miR-146a LPS-induced elevation of miR-146a miR-155and miR-132 was observed in a microarray study of themonocytic leukemia cell line THP-1 after LPS challengeTherefore miR-146a seemed to be a negative regulator ofinflammation Toll-like receptors (TLRs) that recognize bac-terial constituents (TLR2 TLR4 and TLR5) could triggermiR-146a induction while TLRs that recognize viral nucleicacids (TLR3 TLR7 and TLR9) had little effect on miR-146aexpression [48]
In the absence of miR-146a hematopoietic stem cellhomeostasis is disrupted under aging and periodic bacterialencounters as indicated by declines of hematopoietic stemcell number and quality dysregulated hematopoietic stem
and progenitor cell proliferation and differentiation Chroni-cally it can lead to myeloproliferative disease The molecularpathway miR-146aTRAF6NF-120581BIL-6 seems to be involvedin these processes [49]
Rusca et al [50] identified a new molecular network thatcomprises specifically of NF120581B1 and miR-146a The authorshave shown that the same activation-induced miRNA canbe similarly regulated in different cell types in mast cells itcontributes to the regulation of cell homeostasis and survivalwhile in T lymphocytes it modulates T cell memory function
MiR-146a has a role in inflammatory diseases such aspsoriasis and rheumatoid arthritis and in cancers (such aspapillary thyroid carcinoma cervical cancer breast cancerpancreatic cancer and prostate cancer) [51] MiR-146a isupregulated in RA [9] it is downregulated in systemic lupuserythematosus (SLE) where it is a negative regulator of IFN120574pathway [21]
ElevatedmiR-146a expressionwas observed in circulatingCD14+ cells fromuntreatedMS patients [20]MiR-146amiR-125b miR-9 and miR-155 are upregulated in Alzheimerrsquosdisease in which this relatively small number of miRNAsmayimpact several key pathological features [22]
The multilevel effect of the micro-RNA networks is con-vincingly demonstrated by miR-146A since the complexity ofmiRNA and transcription factors are illustrated by the factthat not only the inhibition but also the overexpression ofmiR-146a leads to an inflammatory phenotype
33 miR-132 MiR-132 is expressed in mDCs [28] and inmurine MCs and it targets HB-EGF [27]
In human inflammatory bowel disease miR-132 wasfound to be a modulator of cholinergic signalling [24] InMS miR-132 expression was significantly decreased in circu-lating CD14+ monocytes [20] The miR-132miR-212PTENFOXO3a signaling pathway contributes to neuronal apoptosisin Alzheimerrsquos disease [23]
34 miR-21 miR-21 is a regulator of eosinophil progenitorgrowth MiR21++ and miR-21minusminus bone-marrow derivedeosinophils were morphologically indistinguishable fromeach other butmiR21minusminus cells had decreased growth capacitywith an increased level of apoptosis in vitro A whole genomemicroarray study of wild type- and gene targeted cellsidentified regulation of cell growth cell cycle and immuneresponse as the pathways were affected most significantlyby miR-21 [52] MiR-21 is upregulated in allergic airwayinflammation [16] MiR-21 suppresses TLR2 signaling andIL-13 can upregulate miR-21 expression in human airwayepithelial cells [53] The upregulation of miR-21 miR-142-3pmiR-142-5p and miR-223 was observed both in human andmouse allergic contact dermatitis [54] Esophageal miRNAexpression profile was investigated by microarray fromhuman esophageal tissues of eosinophilic esophagitis (EoE)patients and controls where the most upregulated miRNAswere miR-21 and miR-223 and the most downregulated onewas miR-375 In the plasma of EoE patients miR-146a miR-146b and miR-223 were upregulated [55]
BioMed Research International 7
MiR-21 has a dual role in CD4+ T cells activation-induced miR-21 supports survival of memory T cells whilemodulation of the potential homing by downregulation ofCCR7 protein expression was identified in active naive Tcells [56] One may consider that in vivo silencing of miR-21reversed cardinalmanifestations of autoimmunity in SLE andother diseases [12] suggesting thatmiRNAs have the potentialto become an effective therapeutic target in the future
In a recent study serum level of miR-21 proved to be aprognosticmarker for colorectal cancer as serummiR-21 levelcorrelated with the recurrence and mortality of colorectalcancer patients [57] In a further meta-analysis study earlierresults regarding circulating miR-21 level in cancer weregeneralized [58]
4 Conclusions and Future Perspectives
By now several studies certified that abnormal miRNAexpression is a common feature of various human diseasesand that the number of diseases in which the role of miRNAshas been confirmed increases continuously In some cancertypes certain circulating miRNAs were identified as potentialprognostic markers
In order to find out how inflammation-induced tumori-genesis works and in order to determine new therapeutic tar-gets identification and further investigation of key miRNAsinvolved in inflammation and cancer could be an expedientstrategy In the case of miR-155 it was reported that miR-155 and miR-155-related proinflammatory environment havemutatory-activity and play a role in inflammation-inducedcancers [59] MiR-146a could be one of the missing linksbetween inflammation and tumorigenesis as suggested by anincreasing number of publications that show its importanceboth in inflammatory diseases and tumors
Myeloid lineage development and differentiation path-ways are rather complex as miRNAs and transcriptional fac-tors form a complex network that is lineage-specific negativeand positive regulatory loops By these loops miRNAs areable to regulate normal hematopoietic lineage developmentMoreover the imbalance or disturbance can evoke diseasesby aberrant expression [36]
There is still a limited set of information about miRNApatterns and networks of monocytes mast cells eosinophilsand basophils in the literature while there is growing evi-dence that inflammation and infections can influence malig-nancies Better knowledge of this area of myeloid biology andpathophysiology would be cordially suggested
In this review we intended to overview information onmyeloid miRNAs based on available microarray analysisinvestigating miRNA pattern of myeloid cells We focusedon monocytes neutrophils eosinophils basophils and mastcells There are very few data about myeloid DCs in thisreview as DCs are rather different regarding their miRNAexpression We also did not discuss the miRNA regulation ofnewly identified myeloid-derived suppressor cells
We could reconfirm the key importance of miR-223 inmyeloid cells and provide novel information about miR-223expression in basophils
Even though there are only a few miRNA microarraystudies of myeloid cells in the literature based on them itseems that beside the myeloid key miRNA miR-223 miR-27a and miR-652 and some others could be also interestingregulators with the potential to become therapeutic targets inmyeloid pathology as well
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] D P Bartel ldquoMicroRNAs genomics biogenesis mechanismand functionrdquo Cell vol 116 no 2 pp 281ndash297 2004
[2] E Sonkoly M Stahle and A Pivarcsi ldquoMicroRNAs andimmunity novel players in the regulation of normal immunefunction and inflammationrdquo Seminars in Cancer Biology vol 18no 2 pp 131ndash140 2008
[3] M Filkova B Aradi L Senolt et al ldquoAssociation of circulatingmiR-223 and miR-16 with disease activity in patients with earlyrheumatoid arthritisrdquo Annals of the Rheumatic Diseases 2013
[4] B S Huang Q Z Luo Y Han X B Li L J Cao and LX Wu ldquomicroRNA-223 promotes the growth and invasionof glioblastoma cells by targeting tumor suppressor PAX6rdquoOncology Reports vol 30 no 5 pp 2263ndash2269 2013
[5] F Allantaz D T Cheng T Bergauer et al ldquoExpressionprofiling of human immune cell subsets identifies miRNA-mRNAregulatory relationships correlatedwith cell type specificexpressionrdquo PLoS ONE vol 7 no 1 Article ID e29979 2012
[6] H Kawamoto and N Minato ldquoMyeloid cellsrdquoThe InternationalJournal of Biochemistry amp Cell Biology vol 36 no 8 pp 1374ndash1379 2004
[7] A Tefferi JThiele and JWVardiman ldquoThe 2008WorldHealthOrganization classification system for myeloproliferative neo-plasms order out of chaosrdquo Cancer vol 115 no 17 pp 3842ndash3847 2009
[8] F Shojaei C Zhong X Wu L Yu and N Ferrara ldquoRole ofmyeloid cells in tumor angiogenesis and growthrdquo Trends in CellBiology vol 18 no 8 pp 372ndash378 2008
[9] T Nakasa S Miyaki A Okubo et al ldquoExpression ofMicroRNA-146 in rheumatoid arthritis synovial tissuerdquo Arthri-tis amp Rheumatism vol 58 no 5 pp 1284ndash1292 2008
[10] B Gong H Hu J Chen et al ldquoCaprin-1 is a novel microRNA-223 target for regulating the proliferation and invasion ofhuman breast cancer cellsrdquo Biomedicine amp Pharmacotherapyvol 67 no 7 pp 629ndash636 2013
[11] W Tang F Yu H Yao et al ldquomiR-27a regulates endothelialdifferentiation of breast cancer stem like cellsrdquo Oncogene vol33 no 20 pp 2629ndash2638 2013
[12] B G Garchow O B Encinas Y T Leung et al ldquoSilencing ofmicroR6-21 in vivo ameliorates autoimmune splenomegaly inlupus micerdquo EMBOMolecular Medicine vol 3 no 10 pp 605ndash615 2011
[13] J A Pulikkan V Dengler P S Peramangalam et al ldquoCell-cycleregulator E2F1 and microRNA-223 comprise an autoregulatorynegative feedback loop in acute myeloid leukemiardquo Blood vol115 no 9 pp 1768ndash1778 2010
8 BioMed Research International
[14] M Eyholzer S Schmid J A Schardt S Haefliger B UMuellerand T Pabst ldquoComplexity of miR-223 regulation by CEBPA inhuman AMLrdquo Leukemia Research vol 34 no 5 pp 672ndash6762010
[15] NGarbacki E diValentinVAHuynh-Thuet al ldquoMicroRNAsprofiling in murine models of acute and chronic asthma arelationship with mRNAs targetsrdquo PLoS ONE vol 6 no 1Article ID e16509 2011
[16] T X Lu A Munitz and M E Rothenberg ldquoMicroRNA-21 isup-regulated in allergic airway inflammation and regulates IL-12p35 expressionrdquo Journal of Immunology vol 182 no 8 pp4994ndash5002 2009
[17] A Dorhoi M Iannaccone M Farinacci et al ldquoMicroRNA-223 controls susceptibility to tuberculosis by regulating lungneutrophil recruitmentrdquo The Journal of Clinical Investigationvol 123 no 11 pp 4836ndash4848 2013
[18] C Roderburg T Mollnow B Bongaerts et al ldquoMicro-RNAprofiling in human serum reveals compartment-specific rolesof miR-571 and miR-652 in liver cirrhosisrdquo PLoS ONE vol 7no 3 Article ID e32999 2012
[19] R M OrsquoConnell D S Rao A A Chaudhuri et al ldquoSustainedexpression of microRNA-155 in hematopoietic stem cells causesa myeloproliferative disorderrdquo The Journal of ExperimentalMedicine vol 205 no 3 pp 585ndash594 2008
[20] C S Moore V T Rao B A Durafourt et al ldquomiR-155 as a mul-tiple sclerosis-relevant regulator of myeloid cell polarizationrdquoAnnals of Neurology vol 74 no 5 pp 709ndash720 2013
[21] Y Tang X Luo H Cui et al ldquoMicroRNA-146a contributes toabnormal activation of the type I interferon pathway in humanlupus by targeting the key signaling proteinsrdquo Arthritis andRheumatism vol 60 no 4 pp 1065ndash1075 2009
[22] W J Lukiw T V Andreeva A P Grigorenko and E I RogaevldquoStudying micro RNA function and dysfunction in Alzheimerrsquosdiseaserdquo Frontiers in Genetics vol 3 article 327 2013
[23] H A Wong T Veremeyko N Patel et al ldquoDe-repression ofFOXO3a death axis by microRNA-132 and -212 causes neuronalapoptosis in Alzheimers diseaserdquo Human Molecular Geneticsvol 22 no 15 pp 3077ndash3092 2013
[24] NMaharshak S Shenhar-TsarfatyNAroyo et al ldquoMicroRNA-132 modulates cholinergic signaling and inflammation inhuman inflammatory bowel diseaserdquo Inflammatory Bowel Dis-eases vol 19 no 7 pp 1346ndash1353 2013
[25] S Radom-Aizik F Zaldivar Jr S Oliver P Galassetti and DM Cooper ldquoEvidence for microRNA involvement in exercise-associated neutrophil gene expression changesrdquo Journal ofApplied Physiology vol 109 no 1 pp 252ndash261 2010
[26] J R Ward P R Heath J W Catto M K B Whyte M Miloand S A Renshaw ldquoRegulation of neutrophil senescence bymicroRNAsrdquo PLoS ONE vol 6 no 1 Article ID e15810 2011
[27] V Molnar B E Rsek Z Wiener et al ldquoMicroRNA-132 targetsHB-EGF upon IgE-mediated activation in murine and humanmast cellsrdquo Cellular and Molecular Life Sciences vol 69 no 5pp 793ndash808 2012
[28] L Tserel T Runnel K Kisand et al ldquoMicroRNA expressionprofiles of human blood monocyte-derived dendritic cells andmacrophages reveal miR-511 as putative positive regulator oftoll-like receptor 4rdquo Journal of Biological Chemistry vol 286 no30 pp 26487ndash26495 2011
[29] S Monticelli K M Ansel C Xiao et al ldquoMicroRNA profilingof the murine hematopoietic systemrdquo Genome Biology vol 6no 8 p R71 2005
[30] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[31] F Fazi A Rosa A Fatica et al ldquoA minicircuitry com-prised of microRNA-223 and transcription factors NFI-A andCEBPalpha regulates human granulopoiesisrdquo Cell vol 123 no5 pp 819ndash831 2005
[32] J B Johnnidis M H Harris R T Wheeler et al ldquoRegulationof progenitor cell proliferation and granulocyte function bymicroRNA-223rdquo Nature vol 451 no 7182 pp 1125ndash1129 2008
[33] T X Lu E Lim J A Besse et al ldquoMiR-223 deficiency increaseseosinophil progenitor proliferationrdquo Journal of Immunologyvol 190 no 4 pp 1576ndash1582 2013
[34] T Li M J Morgan S Choksi Y Zhang Y Kim and Z LiuldquoMicroRNAs modulate the noncanonical transcription factorNF-120581B pathway by regulating expression of the kinase IKK120572duringmacrophage differentiationrdquoNature Immunology vol 11no 9 pp 799ndash805 2010
[35] L Shi B Fisslthaler N Zippel et al ldquoMicroRNAs-223 antag-onises angiogenesis by targeting 1205731 integrin and prevent-ing growth factor signaling in endothelial cellsrdquo CirculationResearch 2013
[36] M El Gazzar and C E McCall ldquoMicroRNAs regulatorynetworks in myeloid lineage development and differentiationregulators of the regulatorsrdquo Immunology and Cell Biology vol90 no 6 pp 587ndash593 2012
[37] T Fukao Y Fukuda K Kiga et al ldquoAn Evolutionarily Con-served Mechanism for MicroRNA-223 Expression Revealed byMicroRNA Gene Profilingrdquo Cell vol 129 no 3 pp 617ndash6312007
[38] B U Mueller and T Pabst ldquoCEBP120572 and the pathophysiologyof acutemyeloid leukemiardquoCurrentOpinion inHematology vol13 no 1 pp 7ndash14 2006
[39] J H Mendler K Maharry M D Radmacher et al ldquoRUNX1mutations are associated with poor outcome in younger andolder patients with cytogenetically normal acute myeloidleukemia and with distinct gene and microRNA expressionsignaturesrdquo Journal of Clinical Oncology vol 30 no 25 pp3109ndash3118 2012
[40] F Kuchenbauer S M Mah M Heuser et al ldquoComprehensiveanalysis of mammalian miRNAlowastspecies and their role inmyeloid cellsrdquo Blood vol 118 no 12 pp 3350ndash3358 2011
[41] J Feng A Iwama M Satake and K Kohu ldquoMicroRNA-27 enhances differentiation of myeloblasts into granulocytesby post-transcriptionally downregulating Runx1rdquo The BritishJournal of Haematology vol 145 no 3 pp 412ndash423 2009
[42] O Ben-AmiN Pencovich J LotemD Levanon andYGronerldquoA regulatory interplay between miR-27a and Runx1 duringmegakaryopoiesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 106 no 1 pp 238ndash243 2009
[43] S Min L Li M Zhang et al ldquoTGF-120573-associated miR-27ainhibits dendritic cell-mediated differentiation of Th1 andTh17cells by TAB3 p38 MAPK MAP2K4 and MAP2K7rdquo Genes andImmunity vol 13 no 8 pp 621ndash631 2012
[44] E Tili C M Croce and J J Michaille ldquomiR-155 on thecrosstalk between inflammation and cancerrdquo InternationalReviews of Immunology vol 28 no 5 pp 264ndash284 2009
[45] A Rodriguez E Vigorito S Clare et al ldquoRequirement ofbicmicroRNA-155 for normal immune functionrdquo Science vol316 no 5824 pp 608ndash611 2007
BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
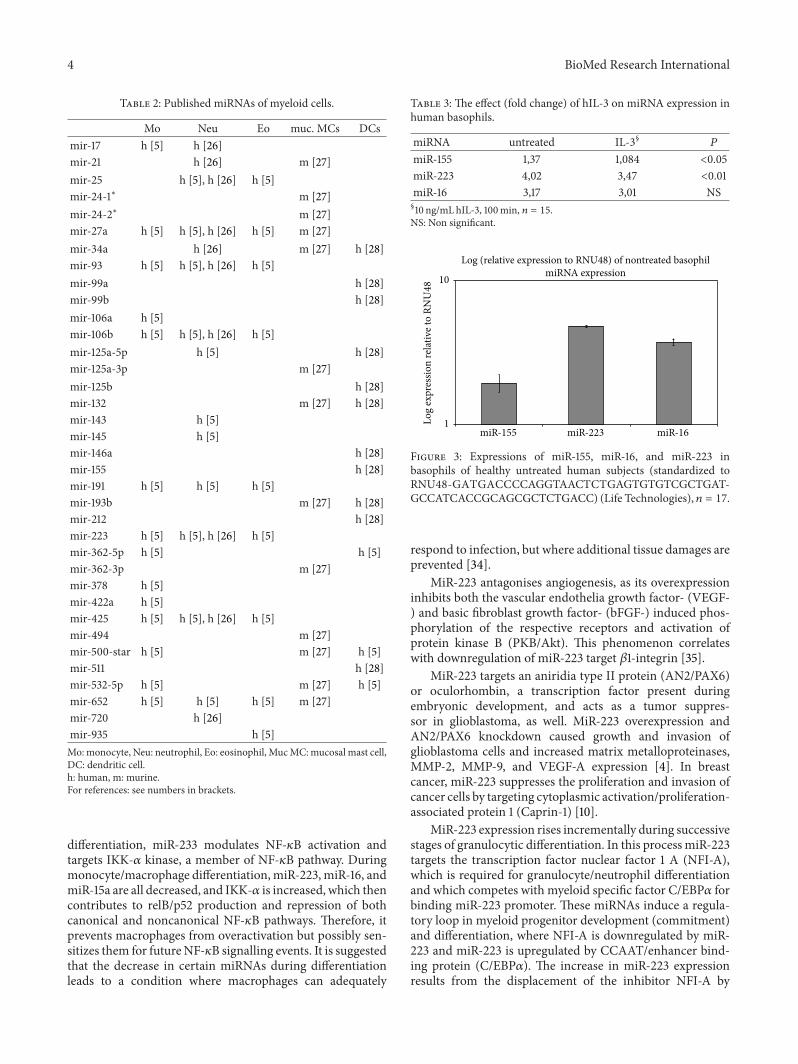
4 BioMed Research International
Table 2 Published miRNAs of myeloid cells
Mo Neu Eo muc MCs DCsmir-17 h [5] h [26]mir-21 h [26] m [27]mir-25 h [5] h [26] h [5]mir-24-1lowast m [27]mir-24-2lowast m [27]mir-27a h [5] h [5] h [26] h [5] m [27]mir-34a h [26] m [27] h [28]mir-93 h [5] h [5] h [26] h [5]mir-99a h [28]mir-99b h [28]mir-106a h [5]mir-106b h [5] h [5] h [26] h [5]mir-125a-5p h [5] h [28]mir-125a-3p m [27]mir-125b h [28]mir-132 m [27] h [28]mir-143 h [5]mir-145 h [5]mir-146a h [28]mir-155 h [28]mir-191 h [5] h [5] h [5]mir-193b m [27] h [28]mir-212 h [28]mir-223 h [5] h [5] h [26] h [5]mir-362-5p h [5] h [5]mir-362-3p m [27]mir-378 h [5]mir-422a h [5]mir-425 h [5] h [5] h [26] h [5]mir-494 m [27]mir-500-star h [5] m [27] h [5]mir-511 h [28]mir-532-5p h [5] m [27] h [5]mir-652 h [5] h [5] h [5] m [27]mir-720 h [26]mir-935 h [5]Momonocyte Neu neutrophil Eo eosinophil MucMCmucosal mast cellDC dendritic cellh human m murineFor references see numbers in brackets
differentiation miR-233 modulates NF-120581B activation andtargets IKK-120572 kinase a member of NF-120581B pathway Duringmonocytemacrophage differentiationmiR-223 miR-16 andmiR-15a are all decreased and IKK-120572 is increased which thencontributes to relBp52 production and repression of bothcanonical and noncanonical NF-120581B pathways Therefore itprevents macrophages from overactivation but possibly sen-sitizes them for futureNF-120581B signalling events It is suggestedthat the decrease in certain miRNAs during differentiationleads to a condition where macrophages can adequately
Table 3 The effect (fold change) of hIL-3 on miRNA expression inhuman basophils
miRNA untreated IL-3sect 119875
miR-155 137 1084 lt005miR-223 402 347 lt001miR-16 317 301 NSsect10 ngmL hIL-3 100min 119899 = 15NS Non significant
Log (relative expression to RNU48) of nontreated basophilmiRNA expression
miR-1551
10
miR-223 miR-16Lo
g ex
pres
sion
relat
ive t
o RN
U48
Figure 3 Expressions of miR-155 miR-16 and miR-223 inbasophils of healthy untreated human subjects (standardized toRNU48-GATGACCCCAGGTAACTCTGAGTGTGTCGCTGAT-GCCATCACCGCAGCGCTCTGACC) (Life Technologies) 119899 = 17
respond to infection but where additional tissue damages areprevented [34]
MiR-223 antagonises angiogenesis as its overexpressioninhibits both the vascular endothelia growth factor- (VEGF-) and basic fibroblast growth factor- (bFGF-) induced phos-phorylation of the respective receptors and activation ofprotein kinase B (PKBAkt) This phenomenon correlateswith downregulation of miR-223 target 1205731-integrin [35]
MiR-223 targets an aniridia type II protein (AN2PAX6)or oculorhombin a transcription factor present duringembryonic development and acts as a tumor suppres-sor in glioblastoma as well MiR-223 overexpression andAN2PAX6 knockdown caused growth and invasion ofglioblastoma cells and increased matrix metalloproteinasesMMP-2 MMP-9 and VEGF-A expression [4] In breastcancer miR-223 suppresses the proliferation and invasion ofcancer cells by targeting cytoplasmic activationproliferation-associated protein 1 (Caprin-1) [10]
MiR-223 expression rises incrementally during successivestages of granulocytic differentiation In this process miR-223targets the transcription factor nuclear factor 1 A (NFI-A)which is required for granulocyteneutrophil differentiationand which competes with myeloid specific factor CEBP120572 forbinding miR-223 promoter These miRNAs induce a regula-tory loop in myeloid progenitor development (commitment)and differentiation where NFI-A is downregulated by miR-223 and miR-223 is upregulated by CCAATenhancer bind-ing protein (CEBP120572) The increase in miR-223 expressionresults from the displacement of the inhibitor NFI-A by
BioMed Research International 5
CEBP120572 [31 36] Based on other data the situation seemsto be even more complex MiR-223 inhibits the transcriptionfactor myocyte enhancer factor-2 (Mef2c) which is involvedin promoting myeloid progenitor differentiation and whichseems to be responsible for the changes of the amount and thephenotype of neutrophils in gene targeted miR-223-deficient(KO) mice mentioned earlier [32] Beside CEBP myeloidtranscription factor PU1 a key regulator of haematopoi-etic differentiation also induces miR-223 expression whileerythroid transcription factor globin transcription factor1 (GATA1) represses it [37] CEBP120572 downregulates cell-cycle regulator E2F1 protein during granulopoiesis and E2F1inhibits miR-223 transcription while miR-223 targets E2F1and blocks cell cycle progression in AML cells forming anegative autoregulatory loop [13] In AML it was shownthat CEBP120572 contributes directly to the development of thedisease [38]
It was also shown that miR-223 is downregulatedin human AML [13] Runt-related transcription factor 1(RUNX1) mutations have been associated with poor clinicaloutcome in younger patients with cytogenetically normalAML In RUNX1-mutated AML patients CEBP120572 is down-regulated [39] CEBP120572 mediated upregulation of miR-223could be a possible explanation of the phenomenon thatmiR-223 is largely suppressed in cells from AML patients [14]
Interestingly in a recent study by Filkova et al [3] itwas suggested that bymonitoringmiR-16 andmiR-223 levelsthe disease outcome in early phase rheumatoid arthritis(ERA) patients could be predicted The authors found lowercirculating miR-16 miR-155 and miR-146 levels in ERAand healthy controlsrsquo samples than in those of patients withrheumatoid arthritis (RA) Furthermore a relationship wasshown between themiR-223 level and peripheral lymphocytenumber changes in ERA patients Moreover it seemed that ahigher circulating miR-223 baseline level could signify a bet-ter improvement in disease activity in ERA after therapeuticintervention [3]
In a murine model of asthma a miRNA microarrayanalysis had been used from allergen-exposed lung at 3 time-points (short- intermediate- and long term) of allergen expo-sure It was found that miR-223 was significantly upregulatedat short-time and was downregulated at intermediate-timeof aerosol exposure Moreover mRNA potentially targetedby miR-223 (Arid4b Il-6 and Lpin2) has undergone adownregulation at short-term exposure [15]
MiR-223 is abundantly expressed during active tubercu-losis (TB) in humans as it was shown by Dorhoi et al [17]based on a microarray from active pulmonary TB patientsand latently infected healthy individuals The authors foundthat similar to human TB miR-223 was upregulated inMycobacterium tuberculosis infected mice Moreover miR-223 deletion rendered resistant mice susceptible to TBand it seemed that miR-223 regulated NF120581B activity inmacrophages and cytokine release in myeloid cells duringTB As it was shown by another microarray experimentfrom lung tissue on days 14 and 21 during Mtb infectionlung influx of innate cells during TB was modulated bymiR-223 through the regulation of chemoattractants Cxcl2Ccl3 and IL-6 were identified as new targets of miR-223
Mortality in infected miR-223 KO animals was 80ndash100while it was only 10 in ldquowild-typerdquo littermates Neutrophildepletion prolonged survival of the animals therefore themajor cause of morbidity in the absence of miR-223 wasconsidered to be neutrophilic inflammation and subsequenttissue destruction It was suggested that the miR-223 targetedchemokines CXCL2 and CCL3 and the proinflammatorycytokine IL-6 played an important role in these processes Inthe long run miR-223 induced negative feedback control ofneutrophil chemotaxis [17]
Despite the general assumption of miRNA duplexes botharms of miR-223 duplex have an active role as it was shownby Kuchenbaueret al The authors found that miR-223lowast had aregulatory role in myeloid progenitor cells In a murine miR-223 KOmodel where the seed region of themiR-223 armwasinactivated the colony-forming cell output was significantlyreduced It was also shown by target predictions and a miR-223 KO-miR-223 WT microarray that miR-223 and miR-223lowast both target insulin-like growth factor (IGF1r)Therefore
they might have cooperative role on IGF1RPI3 K axis a keypathway for developmental and malignant processes Theauthors found that miR-223lowast had a higher expression inhealthy donors than in AML patients and a superior overallsurvival was significantly associated with the high miR-223lowastexpression level [40]
Beside miR-223 two other miRNAs are included inTable 2 as miRNAs which are present in 4 out of 6 typesof myeloid cells miR-27a and miR-652 Likely these threemiRNAs could be key modulators in myeloid cells as well
21 miR-27a miR-27a plays a role in granulocytic differ-entiation as it was shown that the amount of pri-miR-27ain myeloblasts was low while it was high in granulocytesMoreover it targeted the 31015840 UTR of RUNX1 transcriptionfactor a master regulator in hematopoietic development andmiR-27a inhibited the Runx1 protein during differentiation ofmyeloblasts [41] It is possible that themiR-27a-Runx1 systeminvolves a feedback loop [41 42] Interestingly a microarrayanalysis of RUNX1 mutation suggested the importance ofmiR-223 [39] It was shown in a murine asthma model (inthe case of a long-term aerosol exposure) that miR-27a wassignificantly downregulated [15]
MiR-27a promotes angiogenesis [11] and can alterthe activity of NF-120581B mitogen-activated protein kinases(MAPKs) p38 c-Jun N-terminal kinase (JNK) and extra-cellular-signal-regulated kinase (ERK) and therefore influ-ence the production of certain proinflammatory cytokines[43]
22 miR-652 Our knowledge onmiR-652 is rather poor It isspecifically regulated in serum of liver disease and is down-regulated in circulating monocytes isolated from patientscompared to controls As it was shown that stimulation ofmonocyticU937 cells with bacterial lipopolysaccharide (LPS)led to a significant downregulation of miR-652 thereforeit seems that this miRNA is involved in the mediation ofinflammatory signals in immune cells [18]
6 BioMed Research International
3 Further Myeloid miRNAs with WellCharacterized Effects
31 miR-155 MiR-155 plays a critical role during hematopoi-esis and regulates lymphocyte homeostasis and tolerance[44] In a recent study about miRNA expression profilesin leukocytes it was observed that miR-155 was specificallyexpressed in lymphoid lineage cells but it was downregulatedin neutrophils eosinophils and mDCs [5] On the otherhand it was shown that a sustained expression of miR-155 in hematopoietic stem cells could increase immaturegranulocyte numbers in vivo [19] MiR-155 was shown tobe required for normal immune function as gene targetedmiR-155-deficient mice were highly susceptible to Salmonellatyphimurium infection Rodriguezet al [45] also suggestedthat IL-4 promoter binding of c-Maf transcription factor canbe inhibited by loss-of-miR-155 This raises the intriguingpossibility that miRNAs may have an important effect evenonTh2-related humoral immune responses
MiR-155 is the most commonly overexpressed miRNA invarious malignancies It is related to cancers of breast lungliver and the lymphatic system therefore agents that effec-tively block miR-155 may have an outstanding importance incancer therapy [46]
According to a murine asthma model miR-155 is sig-nificantly upregulated at intermediate aerosol exposure [15]It is also important to note that the chromosomal regionin which the miR-155 gene is located (chromosome 21q21)has been previously shown to be associated with increasedasthma susceptibility inHutterites in SouthDakota a founderpopulation of European ancestry [47]
In recent studies it has been reported that miR-155 is ableto modulate myeloid cells in some pathological conditionsrelated to the central nervous system MiR-155 expressionis significantly increased in circulating CD14+ monocytesof multiple sclerosis (MS) patients compared to healthycontrols Transfection of miR-155 increased tumor necrosisfactor (TNF)120572 IL-6 CD80 and CCR7 expression whilethe expression of the miR-155 targeted gene suppressorof cytokine signaling protein (SOCS1) was decreased inhuman microglia On the other hand interferon (IFN)120574 wassignificantly increased in supernatants derived from CD4+ Tcells cocultured with miR-155-transfected monocyte-derivedmacrophages [20]
32 miR-146a LPS-induced elevation of miR-146a miR-155and miR-132 was observed in a microarray study of themonocytic leukemia cell line THP-1 after LPS challengeTherefore miR-146a seemed to be a negative regulator ofinflammation Toll-like receptors (TLRs) that recognize bac-terial constituents (TLR2 TLR4 and TLR5) could triggermiR-146a induction while TLRs that recognize viral nucleicacids (TLR3 TLR7 and TLR9) had little effect on miR-146aexpression [48]
In the absence of miR-146a hematopoietic stem cellhomeostasis is disrupted under aging and periodic bacterialencounters as indicated by declines of hematopoietic stemcell number and quality dysregulated hematopoietic stem
and progenitor cell proliferation and differentiation Chroni-cally it can lead to myeloproliferative disease The molecularpathway miR-146aTRAF6NF-120581BIL-6 seems to be involvedin these processes [49]
Rusca et al [50] identified a new molecular network thatcomprises specifically of NF120581B1 and miR-146a The authorshave shown that the same activation-induced miRNA canbe similarly regulated in different cell types in mast cells itcontributes to the regulation of cell homeostasis and survivalwhile in T lymphocytes it modulates T cell memory function
MiR-146a has a role in inflammatory diseases such aspsoriasis and rheumatoid arthritis and in cancers (such aspapillary thyroid carcinoma cervical cancer breast cancerpancreatic cancer and prostate cancer) [51] MiR-146a isupregulated in RA [9] it is downregulated in systemic lupuserythematosus (SLE) where it is a negative regulator of IFN120574pathway [21]
ElevatedmiR-146a expressionwas observed in circulatingCD14+ cells fromuntreatedMS patients [20]MiR-146amiR-125b miR-9 and miR-155 are upregulated in Alzheimerrsquosdisease in which this relatively small number of miRNAsmayimpact several key pathological features [22]
The multilevel effect of the micro-RNA networks is con-vincingly demonstrated by miR-146A since the complexity ofmiRNA and transcription factors are illustrated by the factthat not only the inhibition but also the overexpression ofmiR-146a leads to an inflammatory phenotype
33 miR-132 MiR-132 is expressed in mDCs [28] and inmurine MCs and it targets HB-EGF [27]
In human inflammatory bowel disease miR-132 wasfound to be a modulator of cholinergic signalling [24] InMS miR-132 expression was significantly decreased in circu-lating CD14+ monocytes [20] The miR-132miR-212PTENFOXO3a signaling pathway contributes to neuronal apoptosisin Alzheimerrsquos disease [23]
34 miR-21 miR-21 is a regulator of eosinophil progenitorgrowth MiR21++ and miR-21minusminus bone-marrow derivedeosinophils were morphologically indistinguishable fromeach other butmiR21minusminus cells had decreased growth capacitywith an increased level of apoptosis in vitro A whole genomemicroarray study of wild type- and gene targeted cellsidentified regulation of cell growth cell cycle and immuneresponse as the pathways were affected most significantlyby miR-21 [52] MiR-21 is upregulated in allergic airwayinflammation [16] MiR-21 suppresses TLR2 signaling andIL-13 can upregulate miR-21 expression in human airwayepithelial cells [53] The upregulation of miR-21 miR-142-3pmiR-142-5p and miR-223 was observed both in human andmouse allergic contact dermatitis [54] Esophageal miRNAexpression profile was investigated by microarray fromhuman esophageal tissues of eosinophilic esophagitis (EoE)patients and controls where the most upregulated miRNAswere miR-21 and miR-223 and the most downregulated onewas miR-375 In the plasma of EoE patients miR-146a miR-146b and miR-223 were upregulated [55]
BioMed Research International 7
MiR-21 has a dual role in CD4+ T cells activation-induced miR-21 supports survival of memory T cells whilemodulation of the potential homing by downregulation ofCCR7 protein expression was identified in active naive Tcells [56] One may consider that in vivo silencing of miR-21reversed cardinalmanifestations of autoimmunity in SLE andother diseases [12] suggesting thatmiRNAs have the potentialto become an effective therapeutic target in the future
In a recent study serum level of miR-21 proved to be aprognosticmarker for colorectal cancer as serummiR-21 levelcorrelated with the recurrence and mortality of colorectalcancer patients [57] In a further meta-analysis study earlierresults regarding circulating miR-21 level in cancer weregeneralized [58]
4 Conclusions and Future Perspectives
By now several studies certified that abnormal miRNAexpression is a common feature of various human diseasesand that the number of diseases in which the role of miRNAshas been confirmed increases continuously In some cancertypes certain circulating miRNAs were identified as potentialprognostic markers
In order to find out how inflammation-induced tumori-genesis works and in order to determine new therapeutic tar-gets identification and further investigation of key miRNAsinvolved in inflammation and cancer could be an expedientstrategy In the case of miR-155 it was reported that miR-155 and miR-155-related proinflammatory environment havemutatory-activity and play a role in inflammation-inducedcancers [59] MiR-146a could be one of the missing linksbetween inflammation and tumorigenesis as suggested by anincreasing number of publications that show its importanceboth in inflammatory diseases and tumors
Myeloid lineage development and differentiation path-ways are rather complex as miRNAs and transcriptional fac-tors form a complex network that is lineage-specific negativeand positive regulatory loops By these loops miRNAs areable to regulate normal hematopoietic lineage developmentMoreover the imbalance or disturbance can evoke diseasesby aberrant expression [36]
There is still a limited set of information about miRNApatterns and networks of monocytes mast cells eosinophilsand basophils in the literature while there is growing evi-dence that inflammation and infections can influence malig-nancies Better knowledge of this area of myeloid biology andpathophysiology would be cordially suggested
In this review we intended to overview information onmyeloid miRNAs based on available microarray analysisinvestigating miRNA pattern of myeloid cells We focusedon monocytes neutrophils eosinophils basophils and mastcells There are very few data about myeloid DCs in thisreview as DCs are rather different regarding their miRNAexpression We also did not discuss the miRNA regulation ofnewly identified myeloid-derived suppressor cells
We could reconfirm the key importance of miR-223 inmyeloid cells and provide novel information about miR-223expression in basophils
Even though there are only a few miRNA microarraystudies of myeloid cells in the literature based on them itseems that beside the myeloid key miRNA miR-223 miR-27a and miR-652 and some others could be also interestingregulators with the potential to become therapeutic targets inmyeloid pathology as well
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] D P Bartel ldquoMicroRNAs genomics biogenesis mechanismand functionrdquo Cell vol 116 no 2 pp 281ndash297 2004
[2] E Sonkoly M Stahle and A Pivarcsi ldquoMicroRNAs andimmunity novel players in the regulation of normal immunefunction and inflammationrdquo Seminars in Cancer Biology vol 18no 2 pp 131ndash140 2008
[3] M Filkova B Aradi L Senolt et al ldquoAssociation of circulatingmiR-223 and miR-16 with disease activity in patients with earlyrheumatoid arthritisrdquo Annals of the Rheumatic Diseases 2013
[4] B S Huang Q Z Luo Y Han X B Li L J Cao and LX Wu ldquomicroRNA-223 promotes the growth and invasionof glioblastoma cells by targeting tumor suppressor PAX6rdquoOncology Reports vol 30 no 5 pp 2263ndash2269 2013
[5] F Allantaz D T Cheng T Bergauer et al ldquoExpressionprofiling of human immune cell subsets identifies miRNA-mRNAregulatory relationships correlatedwith cell type specificexpressionrdquo PLoS ONE vol 7 no 1 Article ID e29979 2012
[6] H Kawamoto and N Minato ldquoMyeloid cellsrdquoThe InternationalJournal of Biochemistry amp Cell Biology vol 36 no 8 pp 1374ndash1379 2004
[7] A Tefferi JThiele and JWVardiman ldquoThe 2008WorldHealthOrganization classification system for myeloproliferative neo-plasms order out of chaosrdquo Cancer vol 115 no 17 pp 3842ndash3847 2009
[8] F Shojaei C Zhong X Wu L Yu and N Ferrara ldquoRole ofmyeloid cells in tumor angiogenesis and growthrdquo Trends in CellBiology vol 18 no 8 pp 372ndash378 2008
[9] T Nakasa S Miyaki A Okubo et al ldquoExpression ofMicroRNA-146 in rheumatoid arthritis synovial tissuerdquo Arthri-tis amp Rheumatism vol 58 no 5 pp 1284ndash1292 2008
[10] B Gong H Hu J Chen et al ldquoCaprin-1 is a novel microRNA-223 target for regulating the proliferation and invasion ofhuman breast cancer cellsrdquo Biomedicine amp Pharmacotherapyvol 67 no 7 pp 629ndash636 2013
[11] W Tang F Yu H Yao et al ldquomiR-27a regulates endothelialdifferentiation of breast cancer stem like cellsrdquo Oncogene vol33 no 20 pp 2629ndash2638 2013
[12] B G Garchow O B Encinas Y T Leung et al ldquoSilencing ofmicroR6-21 in vivo ameliorates autoimmune splenomegaly inlupus micerdquo EMBOMolecular Medicine vol 3 no 10 pp 605ndash615 2011
[13] J A Pulikkan V Dengler P S Peramangalam et al ldquoCell-cycleregulator E2F1 and microRNA-223 comprise an autoregulatorynegative feedback loop in acute myeloid leukemiardquo Blood vol115 no 9 pp 1768ndash1778 2010
8 BioMed Research International
[14] M Eyholzer S Schmid J A Schardt S Haefliger B UMuellerand T Pabst ldquoComplexity of miR-223 regulation by CEBPA inhuman AMLrdquo Leukemia Research vol 34 no 5 pp 672ndash6762010
[15] NGarbacki E diValentinVAHuynh-Thuet al ldquoMicroRNAsprofiling in murine models of acute and chronic asthma arelationship with mRNAs targetsrdquo PLoS ONE vol 6 no 1Article ID e16509 2011
[16] T X Lu A Munitz and M E Rothenberg ldquoMicroRNA-21 isup-regulated in allergic airway inflammation and regulates IL-12p35 expressionrdquo Journal of Immunology vol 182 no 8 pp4994ndash5002 2009
[17] A Dorhoi M Iannaccone M Farinacci et al ldquoMicroRNA-223 controls susceptibility to tuberculosis by regulating lungneutrophil recruitmentrdquo The Journal of Clinical Investigationvol 123 no 11 pp 4836ndash4848 2013
[18] C Roderburg T Mollnow B Bongaerts et al ldquoMicro-RNAprofiling in human serum reveals compartment-specific rolesof miR-571 and miR-652 in liver cirrhosisrdquo PLoS ONE vol 7no 3 Article ID e32999 2012
[19] R M OrsquoConnell D S Rao A A Chaudhuri et al ldquoSustainedexpression of microRNA-155 in hematopoietic stem cells causesa myeloproliferative disorderrdquo The Journal of ExperimentalMedicine vol 205 no 3 pp 585ndash594 2008
[20] C S Moore V T Rao B A Durafourt et al ldquomiR-155 as a mul-tiple sclerosis-relevant regulator of myeloid cell polarizationrdquoAnnals of Neurology vol 74 no 5 pp 709ndash720 2013
[21] Y Tang X Luo H Cui et al ldquoMicroRNA-146a contributes toabnormal activation of the type I interferon pathway in humanlupus by targeting the key signaling proteinsrdquo Arthritis andRheumatism vol 60 no 4 pp 1065ndash1075 2009
[22] W J Lukiw T V Andreeva A P Grigorenko and E I RogaevldquoStudying micro RNA function and dysfunction in Alzheimerrsquosdiseaserdquo Frontiers in Genetics vol 3 article 327 2013
[23] H A Wong T Veremeyko N Patel et al ldquoDe-repression ofFOXO3a death axis by microRNA-132 and -212 causes neuronalapoptosis in Alzheimers diseaserdquo Human Molecular Geneticsvol 22 no 15 pp 3077ndash3092 2013
[24] NMaharshak S Shenhar-TsarfatyNAroyo et al ldquoMicroRNA-132 modulates cholinergic signaling and inflammation inhuman inflammatory bowel diseaserdquo Inflammatory Bowel Dis-eases vol 19 no 7 pp 1346ndash1353 2013
[25] S Radom-Aizik F Zaldivar Jr S Oliver P Galassetti and DM Cooper ldquoEvidence for microRNA involvement in exercise-associated neutrophil gene expression changesrdquo Journal ofApplied Physiology vol 109 no 1 pp 252ndash261 2010
[26] J R Ward P R Heath J W Catto M K B Whyte M Miloand S A Renshaw ldquoRegulation of neutrophil senescence bymicroRNAsrdquo PLoS ONE vol 6 no 1 Article ID e15810 2011
[27] V Molnar B E Rsek Z Wiener et al ldquoMicroRNA-132 targetsHB-EGF upon IgE-mediated activation in murine and humanmast cellsrdquo Cellular and Molecular Life Sciences vol 69 no 5pp 793ndash808 2012
[28] L Tserel T Runnel K Kisand et al ldquoMicroRNA expressionprofiles of human blood monocyte-derived dendritic cells andmacrophages reveal miR-511 as putative positive regulator oftoll-like receptor 4rdquo Journal of Biological Chemistry vol 286 no30 pp 26487ndash26495 2011
[29] S Monticelli K M Ansel C Xiao et al ldquoMicroRNA profilingof the murine hematopoietic systemrdquo Genome Biology vol 6no 8 p R71 2005
[30] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[31] F Fazi A Rosa A Fatica et al ldquoA minicircuitry com-prised of microRNA-223 and transcription factors NFI-A andCEBPalpha regulates human granulopoiesisrdquo Cell vol 123 no5 pp 819ndash831 2005
[32] J B Johnnidis M H Harris R T Wheeler et al ldquoRegulationof progenitor cell proliferation and granulocyte function bymicroRNA-223rdquo Nature vol 451 no 7182 pp 1125ndash1129 2008
[33] T X Lu E Lim J A Besse et al ldquoMiR-223 deficiency increaseseosinophil progenitor proliferationrdquo Journal of Immunologyvol 190 no 4 pp 1576ndash1582 2013
[34] T Li M J Morgan S Choksi Y Zhang Y Kim and Z LiuldquoMicroRNAs modulate the noncanonical transcription factorNF-120581B pathway by regulating expression of the kinase IKK120572duringmacrophage differentiationrdquoNature Immunology vol 11no 9 pp 799ndash805 2010
[35] L Shi B Fisslthaler N Zippel et al ldquoMicroRNAs-223 antag-onises angiogenesis by targeting 1205731 integrin and prevent-ing growth factor signaling in endothelial cellsrdquo CirculationResearch 2013
[36] M El Gazzar and C E McCall ldquoMicroRNAs regulatorynetworks in myeloid lineage development and differentiationregulators of the regulatorsrdquo Immunology and Cell Biology vol90 no 6 pp 587ndash593 2012
[37] T Fukao Y Fukuda K Kiga et al ldquoAn Evolutionarily Con-served Mechanism for MicroRNA-223 Expression Revealed byMicroRNA Gene Profilingrdquo Cell vol 129 no 3 pp 617ndash6312007
[38] B U Mueller and T Pabst ldquoCEBP120572 and the pathophysiologyof acutemyeloid leukemiardquoCurrentOpinion inHematology vol13 no 1 pp 7ndash14 2006
[39] J H Mendler K Maharry M D Radmacher et al ldquoRUNX1mutations are associated with poor outcome in younger andolder patients with cytogenetically normal acute myeloidleukemia and with distinct gene and microRNA expressionsignaturesrdquo Journal of Clinical Oncology vol 30 no 25 pp3109ndash3118 2012
[40] F Kuchenbauer S M Mah M Heuser et al ldquoComprehensiveanalysis of mammalian miRNAlowastspecies and their role inmyeloid cellsrdquo Blood vol 118 no 12 pp 3350ndash3358 2011
[41] J Feng A Iwama M Satake and K Kohu ldquoMicroRNA-27 enhances differentiation of myeloblasts into granulocytesby post-transcriptionally downregulating Runx1rdquo The BritishJournal of Haematology vol 145 no 3 pp 412ndash423 2009
[42] O Ben-AmiN Pencovich J LotemD Levanon andYGronerldquoA regulatory interplay between miR-27a and Runx1 duringmegakaryopoiesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 106 no 1 pp 238ndash243 2009
[43] S Min L Li M Zhang et al ldquoTGF-120573-associated miR-27ainhibits dendritic cell-mediated differentiation of Th1 andTh17cells by TAB3 p38 MAPK MAP2K4 and MAP2K7rdquo Genes andImmunity vol 13 no 8 pp 621ndash631 2012
[44] E Tili C M Croce and J J Michaille ldquomiR-155 on thecrosstalk between inflammation and cancerrdquo InternationalReviews of Immunology vol 28 no 5 pp 264ndash284 2009
[45] A Rodriguez E Vigorito S Clare et al ldquoRequirement ofbicmicroRNA-155 for normal immune functionrdquo Science vol316 no 5824 pp 608ndash611 2007
BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 5
CEBP120572 [31 36] Based on other data the situation seemsto be even more complex MiR-223 inhibits the transcriptionfactor myocyte enhancer factor-2 (Mef2c) which is involvedin promoting myeloid progenitor differentiation and whichseems to be responsible for the changes of the amount and thephenotype of neutrophils in gene targeted miR-223-deficient(KO) mice mentioned earlier [32] Beside CEBP myeloidtranscription factor PU1 a key regulator of haematopoi-etic differentiation also induces miR-223 expression whileerythroid transcription factor globin transcription factor1 (GATA1) represses it [37] CEBP120572 downregulates cell-cycle regulator E2F1 protein during granulopoiesis and E2F1inhibits miR-223 transcription while miR-223 targets E2F1and blocks cell cycle progression in AML cells forming anegative autoregulatory loop [13] In AML it was shownthat CEBP120572 contributes directly to the development of thedisease [38]
It was also shown that miR-223 is downregulatedin human AML [13] Runt-related transcription factor 1(RUNX1) mutations have been associated with poor clinicaloutcome in younger patients with cytogenetically normalAML In RUNX1-mutated AML patients CEBP120572 is down-regulated [39] CEBP120572 mediated upregulation of miR-223could be a possible explanation of the phenomenon thatmiR-223 is largely suppressed in cells from AML patients [14]
Interestingly in a recent study by Filkova et al [3] itwas suggested that bymonitoringmiR-16 andmiR-223 levelsthe disease outcome in early phase rheumatoid arthritis(ERA) patients could be predicted The authors found lowercirculating miR-16 miR-155 and miR-146 levels in ERAand healthy controlsrsquo samples than in those of patients withrheumatoid arthritis (RA) Furthermore a relationship wasshown between themiR-223 level and peripheral lymphocytenumber changes in ERA patients Moreover it seemed that ahigher circulating miR-223 baseline level could signify a bet-ter improvement in disease activity in ERA after therapeuticintervention [3]
In a murine model of asthma a miRNA microarrayanalysis had been used from allergen-exposed lung at 3 time-points (short- intermediate- and long term) of allergen expo-sure It was found that miR-223 was significantly upregulatedat short-time and was downregulated at intermediate-timeof aerosol exposure Moreover mRNA potentially targetedby miR-223 (Arid4b Il-6 and Lpin2) has undergone adownregulation at short-term exposure [15]
MiR-223 is abundantly expressed during active tubercu-losis (TB) in humans as it was shown by Dorhoi et al [17]based on a microarray from active pulmonary TB patientsand latently infected healthy individuals The authors foundthat similar to human TB miR-223 was upregulated inMycobacterium tuberculosis infected mice Moreover miR-223 deletion rendered resistant mice susceptible to TBand it seemed that miR-223 regulated NF120581B activity inmacrophages and cytokine release in myeloid cells duringTB As it was shown by another microarray experimentfrom lung tissue on days 14 and 21 during Mtb infectionlung influx of innate cells during TB was modulated bymiR-223 through the regulation of chemoattractants Cxcl2Ccl3 and IL-6 were identified as new targets of miR-223
Mortality in infected miR-223 KO animals was 80ndash100while it was only 10 in ldquowild-typerdquo littermates Neutrophildepletion prolonged survival of the animals therefore themajor cause of morbidity in the absence of miR-223 wasconsidered to be neutrophilic inflammation and subsequenttissue destruction It was suggested that the miR-223 targetedchemokines CXCL2 and CCL3 and the proinflammatorycytokine IL-6 played an important role in these processes Inthe long run miR-223 induced negative feedback control ofneutrophil chemotaxis [17]
Despite the general assumption of miRNA duplexes botharms of miR-223 duplex have an active role as it was shownby Kuchenbaueret al The authors found that miR-223lowast had aregulatory role in myeloid progenitor cells In a murine miR-223 KOmodel where the seed region of themiR-223 armwasinactivated the colony-forming cell output was significantlyreduced It was also shown by target predictions and a miR-223 KO-miR-223 WT microarray that miR-223 and miR-223lowast both target insulin-like growth factor (IGF1r)Therefore
they might have cooperative role on IGF1RPI3 K axis a keypathway for developmental and malignant processes Theauthors found that miR-223lowast had a higher expression inhealthy donors than in AML patients and a superior overallsurvival was significantly associated with the high miR-223lowastexpression level [40]
Beside miR-223 two other miRNAs are included inTable 2 as miRNAs which are present in 4 out of 6 typesof myeloid cells miR-27a and miR-652 Likely these threemiRNAs could be key modulators in myeloid cells as well
21 miR-27a miR-27a plays a role in granulocytic differ-entiation as it was shown that the amount of pri-miR-27ain myeloblasts was low while it was high in granulocytesMoreover it targeted the 31015840 UTR of RUNX1 transcriptionfactor a master regulator in hematopoietic development andmiR-27a inhibited the Runx1 protein during differentiation ofmyeloblasts [41] It is possible that themiR-27a-Runx1 systeminvolves a feedback loop [41 42] Interestingly a microarrayanalysis of RUNX1 mutation suggested the importance ofmiR-223 [39] It was shown in a murine asthma model (inthe case of a long-term aerosol exposure) that miR-27a wassignificantly downregulated [15]
MiR-27a promotes angiogenesis [11] and can alterthe activity of NF-120581B mitogen-activated protein kinases(MAPKs) p38 c-Jun N-terminal kinase (JNK) and extra-cellular-signal-regulated kinase (ERK) and therefore influ-ence the production of certain proinflammatory cytokines[43]
22 miR-652 Our knowledge onmiR-652 is rather poor It isspecifically regulated in serum of liver disease and is down-regulated in circulating monocytes isolated from patientscompared to controls As it was shown that stimulation ofmonocyticU937 cells with bacterial lipopolysaccharide (LPS)led to a significant downregulation of miR-652 thereforeit seems that this miRNA is involved in the mediation ofinflammatory signals in immune cells [18]
6 BioMed Research International
3 Further Myeloid miRNAs with WellCharacterized Effects
31 miR-155 MiR-155 plays a critical role during hematopoi-esis and regulates lymphocyte homeostasis and tolerance[44] In a recent study about miRNA expression profilesin leukocytes it was observed that miR-155 was specificallyexpressed in lymphoid lineage cells but it was downregulatedin neutrophils eosinophils and mDCs [5] On the otherhand it was shown that a sustained expression of miR-155 in hematopoietic stem cells could increase immaturegranulocyte numbers in vivo [19] MiR-155 was shown tobe required for normal immune function as gene targetedmiR-155-deficient mice were highly susceptible to Salmonellatyphimurium infection Rodriguezet al [45] also suggestedthat IL-4 promoter binding of c-Maf transcription factor canbe inhibited by loss-of-miR-155 This raises the intriguingpossibility that miRNAs may have an important effect evenonTh2-related humoral immune responses
MiR-155 is the most commonly overexpressed miRNA invarious malignancies It is related to cancers of breast lungliver and the lymphatic system therefore agents that effec-tively block miR-155 may have an outstanding importance incancer therapy [46]
According to a murine asthma model miR-155 is sig-nificantly upregulated at intermediate aerosol exposure [15]It is also important to note that the chromosomal regionin which the miR-155 gene is located (chromosome 21q21)has been previously shown to be associated with increasedasthma susceptibility inHutterites in SouthDakota a founderpopulation of European ancestry [47]
In recent studies it has been reported that miR-155 is ableto modulate myeloid cells in some pathological conditionsrelated to the central nervous system MiR-155 expressionis significantly increased in circulating CD14+ monocytesof multiple sclerosis (MS) patients compared to healthycontrols Transfection of miR-155 increased tumor necrosisfactor (TNF)120572 IL-6 CD80 and CCR7 expression whilethe expression of the miR-155 targeted gene suppressorof cytokine signaling protein (SOCS1) was decreased inhuman microglia On the other hand interferon (IFN)120574 wassignificantly increased in supernatants derived from CD4+ Tcells cocultured with miR-155-transfected monocyte-derivedmacrophages [20]
32 miR-146a LPS-induced elevation of miR-146a miR-155and miR-132 was observed in a microarray study of themonocytic leukemia cell line THP-1 after LPS challengeTherefore miR-146a seemed to be a negative regulator ofinflammation Toll-like receptors (TLRs) that recognize bac-terial constituents (TLR2 TLR4 and TLR5) could triggermiR-146a induction while TLRs that recognize viral nucleicacids (TLR3 TLR7 and TLR9) had little effect on miR-146aexpression [48]
In the absence of miR-146a hematopoietic stem cellhomeostasis is disrupted under aging and periodic bacterialencounters as indicated by declines of hematopoietic stemcell number and quality dysregulated hematopoietic stem
and progenitor cell proliferation and differentiation Chroni-cally it can lead to myeloproliferative disease The molecularpathway miR-146aTRAF6NF-120581BIL-6 seems to be involvedin these processes [49]
Rusca et al [50] identified a new molecular network thatcomprises specifically of NF120581B1 and miR-146a The authorshave shown that the same activation-induced miRNA canbe similarly regulated in different cell types in mast cells itcontributes to the regulation of cell homeostasis and survivalwhile in T lymphocytes it modulates T cell memory function
MiR-146a has a role in inflammatory diseases such aspsoriasis and rheumatoid arthritis and in cancers (such aspapillary thyroid carcinoma cervical cancer breast cancerpancreatic cancer and prostate cancer) [51] MiR-146a isupregulated in RA [9] it is downregulated in systemic lupuserythematosus (SLE) where it is a negative regulator of IFN120574pathway [21]
ElevatedmiR-146a expressionwas observed in circulatingCD14+ cells fromuntreatedMS patients [20]MiR-146amiR-125b miR-9 and miR-155 are upregulated in Alzheimerrsquosdisease in which this relatively small number of miRNAsmayimpact several key pathological features [22]
The multilevel effect of the micro-RNA networks is con-vincingly demonstrated by miR-146A since the complexity ofmiRNA and transcription factors are illustrated by the factthat not only the inhibition but also the overexpression ofmiR-146a leads to an inflammatory phenotype
33 miR-132 MiR-132 is expressed in mDCs [28] and inmurine MCs and it targets HB-EGF [27]
In human inflammatory bowel disease miR-132 wasfound to be a modulator of cholinergic signalling [24] InMS miR-132 expression was significantly decreased in circu-lating CD14+ monocytes [20] The miR-132miR-212PTENFOXO3a signaling pathway contributes to neuronal apoptosisin Alzheimerrsquos disease [23]
34 miR-21 miR-21 is a regulator of eosinophil progenitorgrowth MiR21++ and miR-21minusminus bone-marrow derivedeosinophils were morphologically indistinguishable fromeach other butmiR21minusminus cells had decreased growth capacitywith an increased level of apoptosis in vitro A whole genomemicroarray study of wild type- and gene targeted cellsidentified regulation of cell growth cell cycle and immuneresponse as the pathways were affected most significantlyby miR-21 [52] MiR-21 is upregulated in allergic airwayinflammation [16] MiR-21 suppresses TLR2 signaling andIL-13 can upregulate miR-21 expression in human airwayepithelial cells [53] The upregulation of miR-21 miR-142-3pmiR-142-5p and miR-223 was observed both in human andmouse allergic contact dermatitis [54] Esophageal miRNAexpression profile was investigated by microarray fromhuman esophageal tissues of eosinophilic esophagitis (EoE)patients and controls where the most upregulated miRNAswere miR-21 and miR-223 and the most downregulated onewas miR-375 In the plasma of EoE patients miR-146a miR-146b and miR-223 were upregulated [55]
BioMed Research International 7
MiR-21 has a dual role in CD4+ T cells activation-induced miR-21 supports survival of memory T cells whilemodulation of the potential homing by downregulation ofCCR7 protein expression was identified in active naive Tcells [56] One may consider that in vivo silencing of miR-21reversed cardinalmanifestations of autoimmunity in SLE andother diseases [12] suggesting thatmiRNAs have the potentialto become an effective therapeutic target in the future
In a recent study serum level of miR-21 proved to be aprognosticmarker for colorectal cancer as serummiR-21 levelcorrelated with the recurrence and mortality of colorectalcancer patients [57] In a further meta-analysis study earlierresults regarding circulating miR-21 level in cancer weregeneralized [58]
4 Conclusions and Future Perspectives
By now several studies certified that abnormal miRNAexpression is a common feature of various human diseasesand that the number of diseases in which the role of miRNAshas been confirmed increases continuously In some cancertypes certain circulating miRNAs were identified as potentialprognostic markers
In order to find out how inflammation-induced tumori-genesis works and in order to determine new therapeutic tar-gets identification and further investigation of key miRNAsinvolved in inflammation and cancer could be an expedientstrategy In the case of miR-155 it was reported that miR-155 and miR-155-related proinflammatory environment havemutatory-activity and play a role in inflammation-inducedcancers [59] MiR-146a could be one of the missing linksbetween inflammation and tumorigenesis as suggested by anincreasing number of publications that show its importanceboth in inflammatory diseases and tumors
Myeloid lineage development and differentiation path-ways are rather complex as miRNAs and transcriptional fac-tors form a complex network that is lineage-specific negativeand positive regulatory loops By these loops miRNAs areable to regulate normal hematopoietic lineage developmentMoreover the imbalance or disturbance can evoke diseasesby aberrant expression [36]
There is still a limited set of information about miRNApatterns and networks of monocytes mast cells eosinophilsand basophils in the literature while there is growing evi-dence that inflammation and infections can influence malig-nancies Better knowledge of this area of myeloid biology andpathophysiology would be cordially suggested
In this review we intended to overview information onmyeloid miRNAs based on available microarray analysisinvestigating miRNA pattern of myeloid cells We focusedon monocytes neutrophils eosinophils basophils and mastcells There are very few data about myeloid DCs in thisreview as DCs are rather different regarding their miRNAexpression We also did not discuss the miRNA regulation ofnewly identified myeloid-derived suppressor cells
We could reconfirm the key importance of miR-223 inmyeloid cells and provide novel information about miR-223expression in basophils
Even though there are only a few miRNA microarraystudies of myeloid cells in the literature based on them itseems that beside the myeloid key miRNA miR-223 miR-27a and miR-652 and some others could be also interestingregulators with the potential to become therapeutic targets inmyeloid pathology as well
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] D P Bartel ldquoMicroRNAs genomics biogenesis mechanismand functionrdquo Cell vol 116 no 2 pp 281ndash297 2004
[2] E Sonkoly M Stahle and A Pivarcsi ldquoMicroRNAs andimmunity novel players in the regulation of normal immunefunction and inflammationrdquo Seminars in Cancer Biology vol 18no 2 pp 131ndash140 2008
[3] M Filkova B Aradi L Senolt et al ldquoAssociation of circulatingmiR-223 and miR-16 with disease activity in patients with earlyrheumatoid arthritisrdquo Annals of the Rheumatic Diseases 2013
[4] B S Huang Q Z Luo Y Han X B Li L J Cao and LX Wu ldquomicroRNA-223 promotes the growth and invasionof glioblastoma cells by targeting tumor suppressor PAX6rdquoOncology Reports vol 30 no 5 pp 2263ndash2269 2013
[5] F Allantaz D T Cheng T Bergauer et al ldquoExpressionprofiling of human immune cell subsets identifies miRNA-mRNAregulatory relationships correlatedwith cell type specificexpressionrdquo PLoS ONE vol 7 no 1 Article ID e29979 2012
[6] H Kawamoto and N Minato ldquoMyeloid cellsrdquoThe InternationalJournal of Biochemistry amp Cell Biology vol 36 no 8 pp 1374ndash1379 2004
[7] A Tefferi JThiele and JWVardiman ldquoThe 2008WorldHealthOrganization classification system for myeloproliferative neo-plasms order out of chaosrdquo Cancer vol 115 no 17 pp 3842ndash3847 2009
[8] F Shojaei C Zhong X Wu L Yu and N Ferrara ldquoRole ofmyeloid cells in tumor angiogenesis and growthrdquo Trends in CellBiology vol 18 no 8 pp 372ndash378 2008
[9] T Nakasa S Miyaki A Okubo et al ldquoExpression ofMicroRNA-146 in rheumatoid arthritis synovial tissuerdquo Arthri-tis amp Rheumatism vol 58 no 5 pp 1284ndash1292 2008
[10] B Gong H Hu J Chen et al ldquoCaprin-1 is a novel microRNA-223 target for regulating the proliferation and invasion ofhuman breast cancer cellsrdquo Biomedicine amp Pharmacotherapyvol 67 no 7 pp 629ndash636 2013
[11] W Tang F Yu H Yao et al ldquomiR-27a regulates endothelialdifferentiation of breast cancer stem like cellsrdquo Oncogene vol33 no 20 pp 2629ndash2638 2013
[12] B G Garchow O B Encinas Y T Leung et al ldquoSilencing ofmicroR6-21 in vivo ameliorates autoimmune splenomegaly inlupus micerdquo EMBOMolecular Medicine vol 3 no 10 pp 605ndash615 2011
[13] J A Pulikkan V Dengler P S Peramangalam et al ldquoCell-cycleregulator E2F1 and microRNA-223 comprise an autoregulatorynegative feedback loop in acute myeloid leukemiardquo Blood vol115 no 9 pp 1768ndash1778 2010
8 BioMed Research International
[14] M Eyholzer S Schmid J A Schardt S Haefliger B UMuellerand T Pabst ldquoComplexity of miR-223 regulation by CEBPA inhuman AMLrdquo Leukemia Research vol 34 no 5 pp 672ndash6762010
[15] NGarbacki E diValentinVAHuynh-Thuet al ldquoMicroRNAsprofiling in murine models of acute and chronic asthma arelationship with mRNAs targetsrdquo PLoS ONE vol 6 no 1Article ID e16509 2011
[16] T X Lu A Munitz and M E Rothenberg ldquoMicroRNA-21 isup-regulated in allergic airway inflammation and regulates IL-12p35 expressionrdquo Journal of Immunology vol 182 no 8 pp4994ndash5002 2009
[17] A Dorhoi M Iannaccone M Farinacci et al ldquoMicroRNA-223 controls susceptibility to tuberculosis by regulating lungneutrophil recruitmentrdquo The Journal of Clinical Investigationvol 123 no 11 pp 4836ndash4848 2013
[18] C Roderburg T Mollnow B Bongaerts et al ldquoMicro-RNAprofiling in human serum reveals compartment-specific rolesof miR-571 and miR-652 in liver cirrhosisrdquo PLoS ONE vol 7no 3 Article ID e32999 2012
[19] R M OrsquoConnell D S Rao A A Chaudhuri et al ldquoSustainedexpression of microRNA-155 in hematopoietic stem cells causesa myeloproliferative disorderrdquo The Journal of ExperimentalMedicine vol 205 no 3 pp 585ndash594 2008
[20] C S Moore V T Rao B A Durafourt et al ldquomiR-155 as a mul-tiple sclerosis-relevant regulator of myeloid cell polarizationrdquoAnnals of Neurology vol 74 no 5 pp 709ndash720 2013
[21] Y Tang X Luo H Cui et al ldquoMicroRNA-146a contributes toabnormal activation of the type I interferon pathway in humanlupus by targeting the key signaling proteinsrdquo Arthritis andRheumatism vol 60 no 4 pp 1065ndash1075 2009
[22] W J Lukiw T V Andreeva A P Grigorenko and E I RogaevldquoStudying micro RNA function and dysfunction in Alzheimerrsquosdiseaserdquo Frontiers in Genetics vol 3 article 327 2013
[23] H A Wong T Veremeyko N Patel et al ldquoDe-repression ofFOXO3a death axis by microRNA-132 and -212 causes neuronalapoptosis in Alzheimers diseaserdquo Human Molecular Geneticsvol 22 no 15 pp 3077ndash3092 2013
[24] NMaharshak S Shenhar-TsarfatyNAroyo et al ldquoMicroRNA-132 modulates cholinergic signaling and inflammation inhuman inflammatory bowel diseaserdquo Inflammatory Bowel Dis-eases vol 19 no 7 pp 1346ndash1353 2013
[25] S Radom-Aizik F Zaldivar Jr S Oliver P Galassetti and DM Cooper ldquoEvidence for microRNA involvement in exercise-associated neutrophil gene expression changesrdquo Journal ofApplied Physiology vol 109 no 1 pp 252ndash261 2010
[26] J R Ward P R Heath J W Catto M K B Whyte M Miloand S A Renshaw ldquoRegulation of neutrophil senescence bymicroRNAsrdquo PLoS ONE vol 6 no 1 Article ID e15810 2011
[27] V Molnar B E Rsek Z Wiener et al ldquoMicroRNA-132 targetsHB-EGF upon IgE-mediated activation in murine and humanmast cellsrdquo Cellular and Molecular Life Sciences vol 69 no 5pp 793ndash808 2012
[28] L Tserel T Runnel K Kisand et al ldquoMicroRNA expressionprofiles of human blood monocyte-derived dendritic cells andmacrophages reveal miR-511 as putative positive regulator oftoll-like receptor 4rdquo Journal of Biological Chemistry vol 286 no30 pp 26487ndash26495 2011
[29] S Monticelli K M Ansel C Xiao et al ldquoMicroRNA profilingof the murine hematopoietic systemrdquo Genome Biology vol 6no 8 p R71 2005
[30] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[31] F Fazi A Rosa A Fatica et al ldquoA minicircuitry com-prised of microRNA-223 and transcription factors NFI-A andCEBPalpha regulates human granulopoiesisrdquo Cell vol 123 no5 pp 819ndash831 2005
[32] J B Johnnidis M H Harris R T Wheeler et al ldquoRegulationof progenitor cell proliferation and granulocyte function bymicroRNA-223rdquo Nature vol 451 no 7182 pp 1125ndash1129 2008
[33] T X Lu E Lim J A Besse et al ldquoMiR-223 deficiency increaseseosinophil progenitor proliferationrdquo Journal of Immunologyvol 190 no 4 pp 1576ndash1582 2013
[34] T Li M J Morgan S Choksi Y Zhang Y Kim and Z LiuldquoMicroRNAs modulate the noncanonical transcription factorNF-120581B pathway by regulating expression of the kinase IKK120572duringmacrophage differentiationrdquoNature Immunology vol 11no 9 pp 799ndash805 2010
[35] L Shi B Fisslthaler N Zippel et al ldquoMicroRNAs-223 antag-onises angiogenesis by targeting 1205731 integrin and prevent-ing growth factor signaling in endothelial cellsrdquo CirculationResearch 2013
[36] M El Gazzar and C E McCall ldquoMicroRNAs regulatorynetworks in myeloid lineage development and differentiationregulators of the regulatorsrdquo Immunology and Cell Biology vol90 no 6 pp 587ndash593 2012
[37] T Fukao Y Fukuda K Kiga et al ldquoAn Evolutionarily Con-served Mechanism for MicroRNA-223 Expression Revealed byMicroRNA Gene Profilingrdquo Cell vol 129 no 3 pp 617ndash6312007
[38] B U Mueller and T Pabst ldquoCEBP120572 and the pathophysiologyof acutemyeloid leukemiardquoCurrentOpinion inHematology vol13 no 1 pp 7ndash14 2006
[39] J H Mendler K Maharry M D Radmacher et al ldquoRUNX1mutations are associated with poor outcome in younger andolder patients with cytogenetically normal acute myeloidleukemia and with distinct gene and microRNA expressionsignaturesrdquo Journal of Clinical Oncology vol 30 no 25 pp3109ndash3118 2012
[40] F Kuchenbauer S M Mah M Heuser et al ldquoComprehensiveanalysis of mammalian miRNAlowastspecies and their role inmyeloid cellsrdquo Blood vol 118 no 12 pp 3350ndash3358 2011
[41] J Feng A Iwama M Satake and K Kohu ldquoMicroRNA-27 enhances differentiation of myeloblasts into granulocytesby post-transcriptionally downregulating Runx1rdquo The BritishJournal of Haematology vol 145 no 3 pp 412ndash423 2009
[42] O Ben-AmiN Pencovich J LotemD Levanon andYGronerldquoA regulatory interplay between miR-27a and Runx1 duringmegakaryopoiesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 106 no 1 pp 238ndash243 2009
[43] S Min L Li M Zhang et al ldquoTGF-120573-associated miR-27ainhibits dendritic cell-mediated differentiation of Th1 andTh17cells by TAB3 p38 MAPK MAP2K4 and MAP2K7rdquo Genes andImmunity vol 13 no 8 pp 621ndash631 2012
[44] E Tili C M Croce and J J Michaille ldquomiR-155 on thecrosstalk between inflammation and cancerrdquo InternationalReviews of Immunology vol 28 no 5 pp 264ndash284 2009
[45] A Rodriguez E Vigorito S Clare et al ldquoRequirement ofbicmicroRNA-155 for normal immune functionrdquo Science vol316 no 5824 pp 608ndash611 2007
BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 BioMed Research International
3 Further Myeloid miRNAs with WellCharacterized Effects
31 miR-155 MiR-155 plays a critical role during hematopoi-esis and regulates lymphocyte homeostasis and tolerance[44] In a recent study about miRNA expression profilesin leukocytes it was observed that miR-155 was specificallyexpressed in lymphoid lineage cells but it was downregulatedin neutrophils eosinophils and mDCs [5] On the otherhand it was shown that a sustained expression of miR-155 in hematopoietic stem cells could increase immaturegranulocyte numbers in vivo [19] MiR-155 was shown tobe required for normal immune function as gene targetedmiR-155-deficient mice were highly susceptible to Salmonellatyphimurium infection Rodriguezet al [45] also suggestedthat IL-4 promoter binding of c-Maf transcription factor canbe inhibited by loss-of-miR-155 This raises the intriguingpossibility that miRNAs may have an important effect evenonTh2-related humoral immune responses
MiR-155 is the most commonly overexpressed miRNA invarious malignancies It is related to cancers of breast lungliver and the lymphatic system therefore agents that effec-tively block miR-155 may have an outstanding importance incancer therapy [46]
According to a murine asthma model miR-155 is sig-nificantly upregulated at intermediate aerosol exposure [15]It is also important to note that the chromosomal regionin which the miR-155 gene is located (chromosome 21q21)has been previously shown to be associated with increasedasthma susceptibility inHutterites in SouthDakota a founderpopulation of European ancestry [47]
In recent studies it has been reported that miR-155 is ableto modulate myeloid cells in some pathological conditionsrelated to the central nervous system MiR-155 expressionis significantly increased in circulating CD14+ monocytesof multiple sclerosis (MS) patients compared to healthycontrols Transfection of miR-155 increased tumor necrosisfactor (TNF)120572 IL-6 CD80 and CCR7 expression whilethe expression of the miR-155 targeted gene suppressorof cytokine signaling protein (SOCS1) was decreased inhuman microglia On the other hand interferon (IFN)120574 wassignificantly increased in supernatants derived from CD4+ Tcells cocultured with miR-155-transfected monocyte-derivedmacrophages [20]
32 miR-146a LPS-induced elevation of miR-146a miR-155and miR-132 was observed in a microarray study of themonocytic leukemia cell line THP-1 after LPS challengeTherefore miR-146a seemed to be a negative regulator ofinflammation Toll-like receptors (TLRs) that recognize bac-terial constituents (TLR2 TLR4 and TLR5) could triggermiR-146a induction while TLRs that recognize viral nucleicacids (TLR3 TLR7 and TLR9) had little effect on miR-146aexpression [48]
In the absence of miR-146a hematopoietic stem cellhomeostasis is disrupted under aging and periodic bacterialencounters as indicated by declines of hematopoietic stemcell number and quality dysregulated hematopoietic stem
and progenitor cell proliferation and differentiation Chroni-cally it can lead to myeloproliferative disease The molecularpathway miR-146aTRAF6NF-120581BIL-6 seems to be involvedin these processes [49]
Rusca et al [50] identified a new molecular network thatcomprises specifically of NF120581B1 and miR-146a The authorshave shown that the same activation-induced miRNA canbe similarly regulated in different cell types in mast cells itcontributes to the regulation of cell homeostasis and survivalwhile in T lymphocytes it modulates T cell memory function
MiR-146a has a role in inflammatory diseases such aspsoriasis and rheumatoid arthritis and in cancers (such aspapillary thyroid carcinoma cervical cancer breast cancerpancreatic cancer and prostate cancer) [51] MiR-146a isupregulated in RA [9] it is downregulated in systemic lupuserythematosus (SLE) where it is a negative regulator of IFN120574pathway [21]
ElevatedmiR-146a expressionwas observed in circulatingCD14+ cells fromuntreatedMS patients [20]MiR-146amiR-125b miR-9 and miR-155 are upregulated in Alzheimerrsquosdisease in which this relatively small number of miRNAsmayimpact several key pathological features [22]
The multilevel effect of the micro-RNA networks is con-vincingly demonstrated by miR-146A since the complexity ofmiRNA and transcription factors are illustrated by the factthat not only the inhibition but also the overexpression ofmiR-146a leads to an inflammatory phenotype
33 miR-132 MiR-132 is expressed in mDCs [28] and inmurine MCs and it targets HB-EGF [27]
In human inflammatory bowel disease miR-132 wasfound to be a modulator of cholinergic signalling [24] InMS miR-132 expression was significantly decreased in circu-lating CD14+ monocytes [20] The miR-132miR-212PTENFOXO3a signaling pathway contributes to neuronal apoptosisin Alzheimerrsquos disease [23]
34 miR-21 miR-21 is a regulator of eosinophil progenitorgrowth MiR21++ and miR-21minusminus bone-marrow derivedeosinophils were morphologically indistinguishable fromeach other butmiR21minusminus cells had decreased growth capacitywith an increased level of apoptosis in vitro A whole genomemicroarray study of wild type- and gene targeted cellsidentified regulation of cell growth cell cycle and immuneresponse as the pathways were affected most significantlyby miR-21 [52] MiR-21 is upregulated in allergic airwayinflammation [16] MiR-21 suppresses TLR2 signaling andIL-13 can upregulate miR-21 expression in human airwayepithelial cells [53] The upregulation of miR-21 miR-142-3pmiR-142-5p and miR-223 was observed both in human andmouse allergic contact dermatitis [54] Esophageal miRNAexpression profile was investigated by microarray fromhuman esophageal tissues of eosinophilic esophagitis (EoE)patients and controls where the most upregulated miRNAswere miR-21 and miR-223 and the most downregulated onewas miR-375 In the plasma of EoE patients miR-146a miR-146b and miR-223 were upregulated [55]
BioMed Research International 7
MiR-21 has a dual role in CD4+ T cells activation-induced miR-21 supports survival of memory T cells whilemodulation of the potential homing by downregulation ofCCR7 protein expression was identified in active naive Tcells [56] One may consider that in vivo silencing of miR-21reversed cardinalmanifestations of autoimmunity in SLE andother diseases [12] suggesting thatmiRNAs have the potentialto become an effective therapeutic target in the future
In a recent study serum level of miR-21 proved to be aprognosticmarker for colorectal cancer as serummiR-21 levelcorrelated with the recurrence and mortality of colorectalcancer patients [57] In a further meta-analysis study earlierresults regarding circulating miR-21 level in cancer weregeneralized [58]
4 Conclusions and Future Perspectives
By now several studies certified that abnormal miRNAexpression is a common feature of various human diseasesand that the number of diseases in which the role of miRNAshas been confirmed increases continuously In some cancertypes certain circulating miRNAs were identified as potentialprognostic markers
In order to find out how inflammation-induced tumori-genesis works and in order to determine new therapeutic tar-gets identification and further investigation of key miRNAsinvolved in inflammation and cancer could be an expedientstrategy In the case of miR-155 it was reported that miR-155 and miR-155-related proinflammatory environment havemutatory-activity and play a role in inflammation-inducedcancers [59] MiR-146a could be one of the missing linksbetween inflammation and tumorigenesis as suggested by anincreasing number of publications that show its importanceboth in inflammatory diseases and tumors
Myeloid lineage development and differentiation path-ways are rather complex as miRNAs and transcriptional fac-tors form a complex network that is lineage-specific negativeand positive regulatory loops By these loops miRNAs areable to regulate normal hematopoietic lineage developmentMoreover the imbalance or disturbance can evoke diseasesby aberrant expression [36]
There is still a limited set of information about miRNApatterns and networks of monocytes mast cells eosinophilsand basophils in the literature while there is growing evi-dence that inflammation and infections can influence malig-nancies Better knowledge of this area of myeloid biology andpathophysiology would be cordially suggested
In this review we intended to overview information onmyeloid miRNAs based on available microarray analysisinvestigating miRNA pattern of myeloid cells We focusedon monocytes neutrophils eosinophils basophils and mastcells There are very few data about myeloid DCs in thisreview as DCs are rather different regarding their miRNAexpression We also did not discuss the miRNA regulation ofnewly identified myeloid-derived suppressor cells
We could reconfirm the key importance of miR-223 inmyeloid cells and provide novel information about miR-223expression in basophils
Even though there are only a few miRNA microarraystudies of myeloid cells in the literature based on them itseems that beside the myeloid key miRNA miR-223 miR-27a and miR-652 and some others could be also interestingregulators with the potential to become therapeutic targets inmyeloid pathology as well
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] D P Bartel ldquoMicroRNAs genomics biogenesis mechanismand functionrdquo Cell vol 116 no 2 pp 281ndash297 2004
[2] E Sonkoly M Stahle and A Pivarcsi ldquoMicroRNAs andimmunity novel players in the regulation of normal immunefunction and inflammationrdquo Seminars in Cancer Biology vol 18no 2 pp 131ndash140 2008
[3] M Filkova B Aradi L Senolt et al ldquoAssociation of circulatingmiR-223 and miR-16 with disease activity in patients with earlyrheumatoid arthritisrdquo Annals of the Rheumatic Diseases 2013
[4] B S Huang Q Z Luo Y Han X B Li L J Cao and LX Wu ldquomicroRNA-223 promotes the growth and invasionof glioblastoma cells by targeting tumor suppressor PAX6rdquoOncology Reports vol 30 no 5 pp 2263ndash2269 2013
[5] F Allantaz D T Cheng T Bergauer et al ldquoExpressionprofiling of human immune cell subsets identifies miRNA-mRNAregulatory relationships correlatedwith cell type specificexpressionrdquo PLoS ONE vol 7 no 1 Article ID e29979 2012
[6] H Kawamoto and N Minato ldquoMyeloid cellsrdquoThe InternationalJournal of Biochemistry amp Cell Biology vol 36 no 8 pp 1374ndash1379 2004
[7] A Tefferi JThiele and JWVardiman ldquoThe 2008WorldHealthOrganization classification system for myeloproliferative neo-plasms order out of chaosrdquo Cancer vol 115 no 17 pp 3842ndash3847 2009
[8] F Shojaei C Zhong X Wu L Yu and N Ferrara ldquoRole ofmyeloid cells in tumor angiogenesis and growthrdquo Trends in CellBiology vol 18 no 8 pp 372ndash378 2008
[9] T Nakasa S Miyaki A Okubo et al ldquoExpression ofMicroRNA-146 in rheumatoid arthritis synovial tissuerdquo Arthri-tis amp Rheumatism vol 58 no 5 pp 1284ndash1292 2008
[10] B Gong H Hu J Chen et al ldquoCaprin-1 is a novel microRNA-223 target for regulating the proliferation and invasion ofhuman breast cancer cellsrdquo Biomedicine amp Pharmacotherapyvol 67 no 7 pp 629ndash636 2013
[11] W Tang F Yu H Yao et al ldquomiR-27a regulates endothelialdifferentiation of breast cancer stem like cellsrdquo Oncogene vol33 no 20 pp 2629ndash2638 2013
[12] B G Garchow O B Encinas Y T Leung et al ldquoSilencing ofmicroR6-21 in vivo ameliorates autoimmune splenomegaly inlupus micerdquo EMBOMolecular Medicine vol 3 no 10 pp 605ndash615 2011
[13] J A Pulikkan V Dengler P S Peramangalam et al ldquoCell-cycleregulator E2F1 and microRNA-223 comprise an autoregulatorynegative feedback loop in acute myeloid leukemiardquo Blood vol115 no 9 pp 1768ndash1778 2010
8 BioMed Research International
[14] M Eyholzer S Schmid J A Schardt S Haefliger B UMuellerand T Pabst ldquoComplexity of miR-223 regulation by CEBPA inhuman AMLrdquo Leukemia Research vol 34 no 5 pp 672ndash6762010
[15] NGarbacki E diValentinVAHuynh-Thuet al ldquoMicroRNAsprofiling in murine models of acute and chronic asthma arelationship with mRNAs targetsrdquo PLoS ONE vol 6 no 1Article ID e16509 2011
[16] T X Lu A Munitz and M E Rothenberg ldquoMicroRNA-21 isup-regulated in allergic airway inflammation and regulates IL-12p35 expressionrdquo Journal of Immunology vol 182 no 8 pp4994ndash5002 2009
[17] A Dorhoi M Iannaccone M Farinacci et al ldquoMicroRNA-223 controls susceptibility to tuberculosis by regulating lungneutrophil recruitmentrdquo The Journal of Clinical Investigationvol 123 no 11 pp 4836ndash4848 2013
[18] C Roderburg T Mollnow B Bongaerts et al ldquoMicro-RNAprofiling in human serum reveals compartment-specific rolesof miR-571 and miR-652 in liver cirrhosisrdquo PLoS ONE vol 7no 3 Article ID e32999 2012
[19] R M OrsquoConnell D S Rao A A Chaudhuri et al ldquoSustainedexpression of microRNA-155 in hematopoietic stem cells causesa myeloproliferative disorderrdquo The Journal of ExperimentalMedicine vol 205 no 3 pp 585ndash594 2008
[20] C S Moore V T Rao B A Durafourt et al ldquomiR-155 as a mul-tiple sclerosis-relevant regulator of myeloid cell polarizationrdquoAnnals of Neurology vol 74 no 5 pp 709ndash720 2013
[21] Y Tang X Luo H Cui et al ldquoMicroRNA-146a contributes toabnormal activation of the type I interferon pathway in humanlupus by targeting the key signaling proteinsrdquo Arthritis andRheumatism vol 60 no 4 pp 1065ndash1075 2009
[22] W J Lukiw T V Andreeva A P Grigorenko and E I RogaevldquoStudying micro RNA function and dysfunction in Alzheimerrsquosdiseaserdquo Frontiers in Genetics vol 3 article 327 2013
[23] H A Wong T Veremeyko N Patel et al ldquoDe-repression ofFOXO3a death axis by microRNA-132 and -212 causes neuronalapoptosis in Alzheimers diseaserdquo Human Molecular Geneticsvol 22 no 15 pp 3077ndash3092 2013
[24] NMaharshak S Shenhar-TsarfatyNAroyo et al ldquoMicroRNA-132 modulates cholinergic signaling and inflammation inhuman inflammatory bowel diseaserdquo Inflammatory Bowel Dis-eases vol 19 no 7 pp 1346ndash1353 2013
[25] S Radom-Aizik F Zaldivar Jr S Oliver P Galassetti and DM Cooper ldquoEvidence for microRNA involvement in exercise-associated neutrophil gene expression changesrdquo Journal ofApplied Physiology vol 109 no 1 pp 252ndash261 2010
[26] J R Ward P R Heath J W Catto M K B Whyte M Miloand S A Renshaw ldquoRegulation of neutrophil senescence bymicroRNAsrdquo PLoS ONE vol 6 no 1 Article ID e15810 2011
[27] V Molnar B E Rsek Z Wiener et al ldquoMicroRNA-132 targetsHB-EGF upon IgE-mediated activation in murine and humanmast cellsrdquo Cellular and Molecular Life Sciences vol 69 no 5pp 793ndash808 2012
[28] L Tserel T Runnel K Kisand et al ldquoMicroRNA expressionprofiles of human blood monocyte-derived dendritic cells andmacrophages reveal miR-511 as putative positive regulator oftoll-like receptor 4rdquo Journal of Biological Chemistry vol 286 no30 pp 26487ndash26495 2011
[29] S Monticelli K M Ansel C Xiao et al ldquoMicroRNA profilingof the murine hematopoietic systemrdquo Genome Biology vol 6no 8 p R71 2005
[30] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[31] F Fazi A Rosa A Fatica et al ldquoA minicircuitry com-prised of microRNA-223 and transcription factors NFI-A andCEBPalpha regulates human granulopoiesisrdquo Cell vol 123 no5 pp 819ndash831 2005
[32] J B Johnnidis M H Harris R T Wheeler et al ldquoRegulationof progenitor cell proliferation and granulocyte function bymicroRNA-223rdquo Nature vol 451 no 7182 pp 1125ndash1129 2008
[33] T X Lu E Lim J A Besse et al ldquoMiR-223 deficiency increaseseosinophil progenitor proliferationrdquo Journal of Immunologyvol 190 no 4 pp 1576ndash1582 2013
[34] T Li M J Morgan S Choksi Y Zhang Y Kim and Z LiuldquoMicroRNAs modulate the noncanonical transcription factorNF-120581B pathway by regulating expression of the kinase IKK120572duringmacrophage differentiationrdquoNature Immunology vol 11no 9 pp 799ndash805 2010
[35] L Shi B Fisslthaler N Zippel et al ldquoMicroRNAs-223 antag-onises angiogenesis by targeting 1205731 integrin and prevent-ing growth factor signaling in endothelial cellsrdquo CirculationResearch 2013
[36] M El Gazzar and C E McCall ldquoMicroRNAs regulatorynetworks in myeloid lineage development and differentiationregulators of the regulatorsrdquo Immunology and Cell Biology vol90 no 6 pp 587ndash593 2012
[37] T Fukao Y Fukuda K Kiga et al ldquoAn Evolutionarily Con-served Mechanism for MicroRNA-223 Expression Revealed byMicroRNA Gene Profilingrdquo Cell vol 129 no 3 pp 617ndash6312007
[38] B U Mueller and T Pabst ldquoCEBP120572 and the pathophysiologyof acutemyeloid leukemiardquoCurrentOpinion inHematology vol13 no 1 pp 7ndash14 2006
[39] J H Mendler K Maharry M D Radmacher et al ldquoRUNX1mutations are associated with poor outcome in younger andolder patients with cytogenetically normal acute myeloidleukemia and with distinct gene and microRNA expressionsignaturesrdquo Journal of Clinical Oncology vol 30 no 25 pp3109ndash3118 2012
[40] F Kuchenbauer S M Mah M Heuser et al ldquoComprehensiveanalysis of mammalian miRNAlowastspecies and their role inmyeloid cellsrdquo Blood vol 118 no 12 pp 3350ndash3358 2011
[41] J Feng A Iwama M Satake and K Kohu ldquoMicroRNA-27 enhances differentiation of myeloblasts into granulocytesby post-transcriptionally downregulating Runx1rdquo The BritishJournal of Haematology vol 145 no 3 pp 412ndash423 2009
[42] O Ben-AmiN Pencovich J LotemD Levanon andYGronerldquoA regulatory interplay between miR-27a and Runx1 duringmegakaryopoiesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 106 no 1 pp 238ndash243 2009
[43] S Min L Li M Zhang et al ldquoTGF-120573-associated miR-27ainhibits dendritic cell-mediated differentiation of Th1 andTh17cells by TAB3 p38 MAPK MAP2K4 and MAP2K7rdquo Genes andImmunity vol 13 no 8 pp 621ndash631 2012
[44] E Tili C M Croce and J J Michaille ldquomiR-155 on thecrosstalk between inflammation and cancerrdquo InternationalReviews of Immunology vol 28 no 5 pp 264ndash284 2009
[45] A Rodriguez E Vigorito S Clare et al ldquoRequirement ofbicmicroRNA-155 for normal immune functionrdquo Science vol316 no 5824 pp 608ndash611 2007
BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 7
MiR-21 has a dual role in CD4+ T cells activation-induced miR-21 supports survival of memory T cells whilemodulation of the potential homing by downregulation ofCCR7 protein expression was identified in active naive Tcells [56] One may consider that in vivo silencing of miR-21reversed cardinalmanifestations of autoimmunity in SLE andother diseases [12] suggesting thatmiRNAs have the potentialto become an effective therapeutic target in the future
In a recent study serum level of miR-21 proved to be aprognosticmarker for colorectal cancer as serummiR-21 levelcorrelated with the recurrence and mortality of colorectalcancer patients [57] In a further meta-analysis study earlierresults regarding circulating miR-21 level in cancer weregeneralized [58]
4 Conclusions and Future Perspectives
By now several studies certified that abnormal miRNAexpression is a common feature of various human diseasesand that the number of diseases in which the role of miRNAshas been confirmed increases continuously In some cancertypes certain circulating miRNAs were identified as potentialprognostic markers
In order to find out how inflammation-induced tumori-genesis works and in order to determine new therapeutic tar-gets identification and further investigation of key miRNAsinvolved in inflammation and cancer could be an expedientstrategy In the case of miR-155 it was reported that miR-155 and miR-155-related proinflammatory environment havemutatory-activity and play a role in inflammation-inducedcancers [59] MiR-146a could be one of the missing linksbetween inflammation and tumorigenesis as suggested by anincreasing number of publications that show its importanceboth in inflammatory diseases and tumors
Myeloid lineage development and differentiation path-ways are rather complex as miRNAs and transcriptional fac-tors form a complex network that is lineage-specific negativeand positive regulatory loops By these loops miRNAs areable to regulate normal hematopoietic lineage developmentMoreover the imbalance or disturbance can evoke diseasesby aberrant expression [36]
There is still a limited set of information about miRNApatterns and networks of monocytes mast cells eosinophilsand basophils in the literature while there is growing evi-dence that inflammation and infections can influence malig-nancies Better knowledge of this area of myeloid biology andpathophysiology would be cordially suggested
In this review we intended to overview information onmyeloid miRNAs based on available microarray analysisinvestigating miRNA pattern of myeloid cells We focusedon monocytes neutrophils eosinophils basophils and mastcells There are very few data about myeloid DCs in thisreview as DCs are rather different regarding their miRNAexpression We also did not discuss the miRNA regulation ofnewly identified myeloid-derived suppressor cells
We could reconfirm the key importance of miR-223 inmyeloid cells and provide novel information about miR-223expression in basophils
Even though there are only a few miRNA microarraystudies of myeloid cells in the literature based on them itseems that beside the myeloid key miRNA miR-223 miR-27a and miR-652 and some others could be also interestingregulators with the potential to become therapeutic targets inmyeloid pathology as well
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] D P Bartel ldquoMicroRNAs genomics biogenesis mechanismand functionrdquo Cell vol 116 no 2 pp 281ndash297 2004
[2] E Sonkoly M Stahle and A Pivarcsi ldquoMicroRNAs andimmunity novel players in the regulation of normal immunefunction and inflammationrdquo Seminars in Cancer Biology vol 18no 2 pp 131ndash140 2008
[3] M Filkova B Aradi L Senolt et al ldquoAssociation of circulatingmiR-223 and miR-16 with disease activity in patients with earlyrheumatoid arthritisrdquo Annals of the Rheumatic Diseases 2013
[4] B S Huang Q Z Luo Y Han X B Li L J Cao and LX Wu ldquomicroRNA-223 promotes the growth and invasionof glioblastoma cells by targeting tumor suppressor PAX6rdquoOncology Reports vol 30 no 5 pp 2263ndash2269 2013
[5] F Allantaz D T Cheng T Bergauer et al ldquoExpressionprofiling of human immune cell subsets identifies miRNA-mRNAregulatory relationships correlatedwith cell type specificexpressionrdquo PLoS ONE vol 7 no 1 Article ID e29979 2012
[6] H Kawamoto and N Minato ldquoMyeloid cellsrdquoThe InternationalJournal of Biochemistry amp Cell Biology vol 36 no 8 pp 1374ndash1379 2004
[7] A Tefferi JThiele and JWVardiman ldquoThe 2008WorldHealthOrganization classification system for myeloproliferative neo-plasms order out of chaosrdquo Cancer vol 115 no 17 pp 3842ndash3847 2009
[8] F Shojaei C Zhong X Wu L Yu and N Ferrara ldquoRole ofmyeloid cells in tumor angiogenesis and growthrdquo Trends in CellBiology vol 18 no 8 pp 372ndash378 2008
[9] T Nakasa S Miyaki A Okubo et al ldquoExpression ofMicroRNA-146 in rheumatoid arthritis synovial tissuerdquo Arthri-tis amp Rheumatism vol 58 no 5 pp 1284ndash1292 2008
[10] B Gong H Hu J Chen et al ldquoCaprin-1 is a novel microRNA-223 target for regulating the proliferation and invasion ofhuman breast cancer cellsrdquo Biomedicine amp Pharmacotherapyvol 67 no 7 pp 629ndash636 2013
[11] W Tang F Yu H Yao et al ldquomiR-27a regulates endothelialdifferentiation of breast cancer stem like cellsrdquo Oncogene vol33 no 20 pp 2629ndash2638 2013
[12] B G Garchow O B Encinas Y T Leung et al ldquoSilencing ofmicroR6-21 in vivo ameliorates autoimmune splenomegaly inlupus micerdquo EMBOMolecular Medicine vol 3 no 10 pp 605ndash615 2011
[13] J A Pulikkan V Dengler P S Peramangalam et al ldquoCell-cycleregulator E2F1 and microRNA-223 comprise an autoregulatorynegative feedback loop in acute myeloid leukemiardquo Blood vol115 no 9 pp 1768ndash1778 2010
8 BioMed Research International
[14] M Eyholzer S Schmid J A Schardt S Haefliger B UMuellerand T Pabst ldquoComplexity of miR-223 regulation by CEBPA inhuman AMLrdquo Leukemia Research vol 34 no 5 pp 672ndash6762010
[15] NGarbacki E diValentinVAHuynh-Thuet al ldquoMicroRNAsprofiling in murine models of acute and chronic asthma arelationship with mRNAs targetsrdquo PLoS ONE vol 6 no 1Article ID e16509 2011
[16] T X Lu A Munitz and M E Rothenberg ldquoMicroRNA-21 isup-regulated in allergic airway inflammation and regulates IL-12p35 expressionrdquo Journal of Immunology vol 182 no 8 pp4994ndash5002 2009
[17] A Dorhoi M Iannaccone M Farinacci et al ldquoMicroRNA-223 controls susceptibility to tuberculosis by regulating lungneutrophil recruitmentrdquo The Journal of Clinical Investigationvol 123 no 11 pp 4836ndash4848 2013
[18] C Roderburg T Mollnow B Bongaerts et al ldquoMicro-RNAprofiling in human serum reveals compartment-specific rolesof miR-571 and miR-652 in liver cirrhosisrdquo PLoS ONE vol 7no 3 Article ID e32999 2012
[19] R M OrsquoConnell D S Rao A A Chaudhuri et al ldquoSustainedexpression of microRNA-155 in hematopoietic stem cells causesa myeloproliferative disorderrdquo The Journal of ExperimentalMedicine vol 205 no 3 pp 585ndash594 2008
[20] C S Moore V T Rao B A Durafourt et al ldquomiR-155 as a mul-tiple sclerosis-relevant regulator of myeloid cell polarizationrdquoAnnals of Neurology vol 74 no 5 pp 709ndash720 2013
[21] Y Tang X Luo H Cui et al ldquoMicroRNA-146a contributes toabnormal activation of the type I interferon pathway in humanlupus by targeting the key signaling proteinsrdquo Arthritis andRheumatism vol 60 no 4 pp 1065ndash1075 2009
[22] W J Lukiw T V Andreeva A P Grigorenko and E I RogaevldquoStudying micro RNA function and dysfunction in Alzheimerrsquosdiseaserdquo Frontiers in Genetics vol 3 article 327 2013
[23] H A Wong T Veremeyko N Patel et al ldquoDe-repression ofFOXO3a death axis by microRNA-132 and -212 causes neuronalapoptosis in Alzheimers diseaserdquo Human Molecular Geneticsvol 22 no 15 pp 3077ndash3092 2013
[24] NMaharshak S Shenhar-TsarfatyNAroyo et al ldquoMicroRNA-132 modulates cholinergic signaling and inflammation inhuman inflammatory bowel diseaserdquo Inflammatory Bowel Dis-eases vol 19 no 7 pp 1346ndash1353 2013
[25] S Radom-Aizik F Zaldivar Jr S Oliver P Galassetti and DM Cooper ldquoEvidence for microRNA involvement in exercise-associated neutrophil gene expression changesrdquo Journal ofApplied Physiology vol 109 no 1 pp 252ndash261 2010
[26] J R Ward P R Heath J W Catto M K B Whyte M Miloand S A Renshaw ldquoRegulation of neutrophil senescence bymicroRNAsrdquo PLoS ONE vol 6 no 1 Article ID e15810 2011
[27] V Molnar B E Rsek Z Wiener et al ldquoMicroRNA-132 targetsHB-EGF upon IgE-mediated activation in murine and humanmast cellsrdquo Cellular and Molecular Life Sciences vol 69 no 5pp 793ndash808 2012
[28] L Tserel T Runnel K Kisand et al ldquoMicroRNA expressionprofiles of human blood monocyte-derived dendritic cells andmacrophages reveal miR-511 as putative positive regulator oftoll-like receptor 4rdquo Journal of Biological Chemistry vol 286 no30 pp 26487ndash26495 2011
[29] S Monticelli K M Ansel C Xiao et al ldquoMicroRNA profilingof the murine hematopoietic systemrdquo Genome Biology vol 6no 8 p R71 2005
[30] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[31] F Fazi A Rosa A Fatica et al ldquoA minicircuitry com-prised of microRNA-223 and transcription factors NFI-A andCEBPalpha regulates human granulopoiesisrdquo Cell vol 123 no5 pp 819ndash831 2005
[32] J B Johnnidis M H Harris R T Wheeler et al ldquoRegulationof progenitor cell proliferation and granulocyte function bymicroRNA-223rdquo Nature vol 451 no 7182 pp 1125ndash1129 2008
[33] T X Lu E Lim J A Besse et al ldquoMiR-223 deficiency increaseseosinophil progenitor proliferationrdquo Journal of Immunologyvol 190 no 4 pp 1576ndash1582 2013
[34] T Li M J Morgan S Choksi Y Zhang Y Kim and Z LiuldquoMicroRNAs modulate the noncanonical transcription factorNF-120581B pathway by regulating expression of the kinase IKK120572duringmacrophage differentiationrdquoNature Immunology vol 11no 9 pp 799ndash805 2010
[35] L Shi B Fisslthaler N Zippel et al ldquoMicroRNAs-223 antag-onises angiogenesis by targeting 1205731 integrin and prevent-ing growth factor signaling in endothelial cellsrdquo CirculationResearch 2013
[36] M El Gazzar and C E McCall ldquoMicroRNAs regulatorynetworks in myeloid lineage development and differentiationregulators of the regulatorsrdquo Immunology and Cell Biology vol90 no 6 pp 587ndash593 2012
[37] T Fukao Y Fukuda K Kiga et al ldquoAn Evolutionarily Con-served Mechanism for MicroRNA-223 Expression Revealed byMicroRNA Gene Profilingrdquo Cell vol 129 no 3 pp 617ndash6312007
[38] B U Mueller and T Pabst ldquoCEBP120572 and the pathophysiologyof acutemyeloid leukemiardquoCurrentOpinion inHematology vol13 no 1 pp 7ndash14 2006
[39] J H Mendler K Maharry M D Radmacher et al ldquoRUNX1mutations are associated with poor outcome in younger andolder patients with cytogenetically normal acute myeloidleukemia and with distinct gene and microRNA expressionsignaturesrdquo Journal of Clinical Oncology vol 30 no 25 pp3109ndash3118 2012
[40] F Kuchenbauer S M Mah M Heuser et al ldquoComprehensiveanalysis of mammalian miRNAlowastspecies and their role inmyeloid cellsrdquo Blood vol 118 no 12 pp 3350ndash3358 2011
[41] J Feng A Iwama M Satake and K Kohu ldquoMicroRNA-27 enhances differentiation of myeloblasts into granulocytesby post-transcriptionally downregulating Runx1rdquo The BritishJournal of Haematology vol 145 no 3 pp 412ndash423 2009
[42] O Ben-AmiN Pencovich J LotemD Levanon andYGronerldquoA regulatory interplay between miR-27a and Runx1 duringmegakaryopoiesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 106 no 1 pp 238ndash243 2009
[43] S Min L Li M Zhang et al ldquoTGF-120573-associated miR-27ainhibits dendritic cell-mediated differentiation of Th1 andTh17cells by TAB3 p38 MAPK MAP2K4 and MAP2K7rdquo Genes andImmunity vol 13 no 8 pp 621ndash631 2012
[44] E Tili C M Croce and J J Michaille ldquomiR-155 on thecrosstalk between inflammation and cancerrdquo InternationalReviews of Immunology vol 28 no 5 pp 264ndash284 2009
[45] A Rodriguez E Vigorito S Clare et al ldquoRequirement ofbicmicroRNA-155 for normal immune functionrdquo Science vol316 no 5824 pp 608ndash611 2007
BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 BioMed Research International
[14] M Eyholzer S Schmid J A Schardt S Haefliger B UMuellerand T Pabst ldquoComplexity of miR-223 regulation by CEBPA inhuman AMLrdquo Leukemia Research vol 34 no 5 pp 672ndash6762010
[15] NGarbacki E diValentinVAHuynh-Thuet al ldquoMicroRNAsprofiling in murine models of acute and chronic asthma arelationship with mRNAs targetsrdquo PLoS ONE vol 6 no 1Article ID e16509 2011
[16] T X Lu A Munitz and M E Rothenberg ldquoMicroRNA-21 isup-regulated in allergic airway inflammation and regulates IL-12p35 expressionrdquo Journal of Immunology vol 182 no 8 pp4994ndash5002 2009
[17] A Dorhoi M Iannaccone M Farinacci et al ldquoMicroRNA-223 controls susceptibility to tuberculosis by regulating lungneutrophil recruitmentrdquo The Journal of Clinical Investigationvol 123 no 11 pp 4836ndash4848 2013
[18] C Roderburg T Mollnow B Bongaerts et al ldquoMicro-RNAprofiling in human serum reveals compartment-specific rolesof miR-571 and miR-652 in liver cirrhosisrdquo PLoS ONE vol 7no 3 Article ID e32999 2012
[19] R M OrsquoConnell D S Rao A A Chaudhuri et al ldquoSustainedexpression of microRNA-155 in hematopoietic stem cells causesa myeloproliferative disorderrdquo The Journal of ExperimentalMedicine vol 205 no 3 pp 585ndash594 2008
[20] C S Moore V T Rao B A Durafourt et al ldquomiR-155 as a mul-tiple sclerosis-relevant regulator of myeloid cell polarizationrdquoAnnals of Neurology vol 74 no 5 pp 709ndash720 2013
[21] Y Tang X Luo H Cui et al ldquoMicroRNA-146a contributes toabnormal activation of the type I interferon pathway in humanlupus by targeting the key signaling proteinsrdquo Arthritis andRheumatism vol 60 no 4 pp 1065ndash1075 2009
[22] W J Lukiw T V Andreeva A P Grigorenko and E I RogaevldquoStudying micro RNA function and dysfunction in Alzheimerrsquosdiseaserdquo Frontiers in Genetics vol 3 article 327 2013
[23] H A Wong T Veremeyko N Patel et al ldquoDe-repression ofFOXO3a death axis by microRNA-132 and -212 causes neuronalapoptosis in Alzheimers diseaserdquo Human Molecular Geneticsvol 22 no 15 pp 3077ndash3092 2013
[24] NMaharshak S Shenhar-TsarfatyNAroyo et al ldquoMicroRNA-132 modulates cholinergic signaling and inflammation inhuman inflammatory bowel diseaserdquo Inflammatory Bowel Dis-eases vol 19 no 7 pp 1346ndash1353 2013
[25] S Radom-Aizik F Zaldivar Jr S Oliver P Galassetti and DM Cooper ldquoEvidence for microRNA involvement in exercise-associated neutrophil gene expression changesrdquo Journal ofApplied Physiology vol 109 no 1 pp 252ndash261 2010
[26] J R Ward P R Heath J W Catto M K B Whyte M Miloand S A Renshaw ldquoRegulation of neutrophil senescence bymicroRNAsrdquo PLoS ONE vol 6 no 1 Article ID e15810 2011
[27] V Molnar B E Rsek Z Wiener et al ldquoMicroRNA-132 targetsHB-EGF upon IgE-mediated activation in murine and humanmast cellsrdquo Cellular and Molecular Life Sciences vol 69 no 5pp 793ndash808 2012
[28] L Tserel T Runnel K Kisand et al ldquoMicroRNA expressionprofiles of human blood monocyte-derived dendritic cells andmacrophages reveal miR-511 as putative positive regulator oftoll-like receptor 4rdquo Journal of Biological Chemistry vol 286 no30 pp 26487ndash26495 2011
[29] S Monticelli K M Ansel C Xiao et al ldquoMicroRNA profilingof the murine hematopoietic systemrdquo Genome Biology vol 6no 8 p R71 2005
[30] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[31] F Fazi A Rosa A Fatica et al ldquoA minicircuitry com-prised of microRNA-223 and transcription factors NFI-A andCEBPalpha regulates human granulopoiesisrdquo Cell vol 123 no5 pp 819ndash831 2005
[32] J B Johnnidis M H Harris R T Wheeler et al ldquoRegulationof progenitor cell proliferation and granulocyte function bymicroRNA-223rdquo Nature vol 451 no 7182 pp 1125ndash1129 2008
[33] T X Lu E Lim J A Besse et al ldquoMiR-223 deficiency increaseseosinophil progenitor proliferationrdquo Journal of Immunologyvol 190 no 4 pp 1576ndash1582 2013
[34] T Li M J Morgan S Choksi Y Zhang Y Kim and Z LiuldquoMicroRNAs modulate the noncanonical transcription factorNF-120581B pathway by regulating expression of the kinase IKK120572duringmacrophage differentiationrdquoNature Immunology vol 11no 9 pp 799ndash805 2010
[35] L Shi B Fisslthaler N Zippel et al ldquoMicroRNAs-223 antag-onises angiogenesis by targeting 1205731 integrin and prevent-ing growth factor signaling in endothelial cellsrdquo CirculationResearch 2013
[36] M El Gazzar and C E McCall ldquoMicroRNAs regulatorynetworks in myeloid lineage development and differentiationregulators of the regulatorsrdquo Immunology and Cell Biology vol90 no 6 pp 587ndash593 2012
[37] T Fukao Y Fukuda K Kiga et al ldquoAn Evolutionarily Con-served Mechanism for MicroRNA-223 Expression Revealed byMicroRNA Gene Profilingrdquo Cell vol 129 no 3 pp 617ndash6312007
[38] B U Mueller and T Pabst ldquoCEBP120572 and the pathophysiologyof acutemyeloid leukemiardquoCurrentOpinion inHematology vol13 no 1 pp 7ndash14 2006
[39] J H Mendler K Maharry M D Radmacher et al ldquoRUNX1mutations are associated with poor outcome in younger andolder patients with cytogenetically normal acute myeloidleukemia and with distinct gene and microRNA expressionsignaturesrdquo Journal of Clinical Oncology vol 30 no 25 pp3109ndash3118 2012
[40] F Kuchenbauer S M Mah M Heuser et al ldquoComprehensiveanalysis of mammalian miRNAlowastspecies and their role inmyeloid cellsrdquo Blood vol 118 no 12 pp 3350ndash3358 2011
[41] J Feng A Iwama M Satake and K Kohu ldquoMicroRNA-27 enhances differentiation of myeloblasts into granulocytesby post-transcriptionally downregulating Runx1rdquo The BritishJournal of Haematology vol 145 no 3 pp 412ndash423 2009
[42] O Ben-AmiN Pencovich J LotemD Levanon andYGronerldquoA regulatory interplay between miR-27a and Runx1 duringmegakaryopoiesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 106 no 1 pp 238ndash243 2009
[43] S Min L Li M Zhang et al ldquoTGF-120573-associated miR-27ainhibits dendritic cell-mediated differentiation of Th1 andTh17cells by TAB3 p38 MAPK MAP2K4 and MAP2K7rdquo Genes andImmunity vol 13 no 8 pp 621ndash631 2012
[44] E Tili C M Croce and J J Michaille ldquomiR-155 on thecrosstalk between inflammation and cancerrdquo InternationalReviews of Immunology vol 28 no 5 pp 264ndash284 2009
[45] A Rodriguez E Vigorito S Clare et al ldquoRequirement ofbicmicroRNA-155 for normal immune functionrdquo Science vol316 no 5824 pp 608ndash611 2007
BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 9
[46] G Higgs and F Slack ldquoThe multiple roles of microRNA-155 inoncogenesisrdquo Journal of Clinical Bioinformatics vol 3 no 1 p17 2013
[47] C Ober N J Cox M Abney et al ldquoGenome-wide search forasthma susceptibility loci in a founder population The Collab-orative Study on the Genetics of Asthmardquo Human MolecularGenetics vol 7 no 9 pp 1393ndash1398 1998
[48] K D Taganov M P Boldin K Chang and D Baltimore ldquoNF-120581B-dependent induction of microRNA miR-146 an inhibitortargeted to signaling proteins of innate immune responsesrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 103 no 33 pp 12481ndash12486 2006
[49] J L Zhao D S Rao R M OrsquoConnell Y Garcia-Flores and DBaltimore ldquoMicroRNA-146a acts as a guardian of the qualityand longevity of hematopoietic stem cells in micerdquo eLife vol2013 no 2 Article ID e00537 2013
[50] N Rusca L Deho S Montagner et al ldquomiR-146a and NF-120581B1regulate mast cell survival and T lymphocyte differentiationrdquoMolecular and Cellular Biology vol 32 no 21 pp 4432ndash44442012
[51] A E Williams M M Perry S A Moschos H M Larner-Svensson andM A Lindsay ldquoRole of miRNA-146a in the regu-lation of the innate immune response and cancerrdquo BiochemicalSociety Transactions vol 36 part 6 pp 1211ndash1215 2008
[52] T X Lu E Lim S Itskovich et al ldquoTargeted ablation of miR-21 decreases murine eosinophil progenitor cell growthrdquo PLoSONE vol 8 no 3 Article ID e59397 2013
[53] S R Case R J Martin D Jiang M N Minor and H W ChuldquoMicroRNA-21 inhibits toll-like receptor 2 agonistinduced lunginflammation in micerdquo Experimental Lung Research vol 37 no8 pp 500ndash508 2011
[54] M T Vennegaard C M Bonefeld P H Hagedorn et alldquoAllergic contact dermatitis induces upregulation of identicalmicroRNAs in humans and micerdquo Contact Dermatitis vol 67no 5 pp 298ndash305 2012
[55] T X Lu J D Sherrill T Wen et al ldquoMicroRNA signaturein patients with eosinophilic esophagitis reversibility withglucocorticoids and assessment as disease biomarkersrdquo TheJournal of Allergy and Clinical Immunology vol 129 no 4 pp1064ndash1075 2012
[56] K Smigielska-Czepiel A van den Berg P Jellema et alldquoDual role of miR-21 in CD4+ T-cells activation-induced miR-21 supports survival of memory T-cells and regulates CCR7expression in naive T-cellsrdquo PloS ONE vol 8 no 10 Article IDe76217 2013
[57] P Menendez D Padilla P Villarejo et al ldquoPrognostic impli-cations of serum microRNA-21 in colorectal cancerrdquo Journal ofSurgical Oncology vol 108 no 6 pp 369ndash373 2013
[58] Y Wang X Gao F Wei et al ldquoDiagnostic and prognostic valueof circulating miR-21 for cancer a systematic review and meta-analysisrdquo Gene vol 533 no 1 pp 389ndash397 2014
[59] E Tili J J Michaille D Wernicke et al ldquoMutator activityinduced by microRNA-155 (miR-155) links inflammation andcancerrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 108 no 12 pp 4908ndash4913 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom