influence of maternal antibodies on active pertussis toxoid immunization of neonatal mice and...
TRANSCRIPT

In
MAa
b
c
d
a
ARRAA
KPIMN
1
iidloe2erisb[
e5
0d
Vaccine 29 (2011) 7718– 7726
Contents lists available at ScienceDirect
Vaccine
j ourna l ho me pag e: www.elsev ier .com/ locate /vacc ine
nfluence of maternal antibodies on active pertussis toxoid immunization ofeonatal mice and piglets
onika. Polewicza, Aleksandra Graciaa, Rachelle Buchanana, Stacy Stroma, Scott A. Halperinc,ndrew A. Pottera, Lorne A. Babiukd, Volker Gerdtsa,b,∗
Vaccine and Infectious Disease Organization (VIDO), University of Saskatchewan S7N 5E3, SK, CanadaDepartment of Veterinary Microbiology, Western College of Veterinary Medicine, University of Saskatchewan, SK, CanadaDepartment of Pediatrics, Faculty of Medicine, Dalhousie University, Halifax, NS, CanadaUniversity of Alberta, Edmonton, Alberta, Canada
r t i c l e i n f o
rticle history:eceived 1 June 2011eceived in revised form 27 July 2011ccepted 28 July 2011vailable online 19 August 2011
eywords:ertussis
a b s t r a c t
Whooping cough caused by infection with Bordetella pertussis, is a serious illness in infants and youngchildren. Mortality due to whooping cough is being reported in infants too young to be immunized aswell as those who have not completed their series of vaccinations. One of the major factors that interfereswith successful active immunization in early life is the presence of maternal antibodies (MatAbs). Usingthe mouse and pig models, we evaluated the effect of maternal antibodies on active immunization withpertussis toxoid (PTd) and explored strategies to overcome this interference. Our results indicate that pas-sively transferred maternal antibodies interfered with active immunization using pertussis toxoid. The
nterference with maternal antibodiesaternal antibodieseonatal vaccines
level of passively transferred antibodies directly correlated with the level of interference observed. How-ever, this interference could be overcome by using a second booster immunization or by co-formulatingthe toxoid with novel adjuvants. These results support the need for novel vaccine formulations thatare optimized for the neonate and that can be used not only to modulate the inherently biased neona-tal immune system but also to prime the response in the presence of passively transferred maternalantibodies.
. Introduction
Bordetella pertussis, the causative agent of pertussis (whoop-ng cough), is a Gram-negative bacillus. This serious respiratorynfection is easily transmitted by close contact, mainly throughroplets [1]. Although vaccination with whole cell (wP) and acellu-
ar pertussis (aP) vaccines has significantly reduced the incidencef the disease, some 20–40 million cases of pertussis occur globallyach year, 90% of which are found in developing countries. About00,000–400,000 of those infected, mostly infants, die from the dis-ase [2,3]. Numerous countries, especially developing nations, haveeported an increase in infant morbidity due to pertussis [4]. A risen pertussis cases has also been observed in developed countriesuch as the United States [5,6] and Canada [7]. Recently there has
een a dramatic increase in disease in older children and adults6,8–10], who can transmit the infection to young infants who are∗ Corresponding author at: Vaccine and Infectious Disease Organization, 120 Vet-rinary Road, University of Saskatchewan, Saskatoon, Saskatchewan, Canada, S7NE3. Tel.: +1 306 966 1513; fax: +1 306 966 7478.
E-mail address: [email protected] (V. Gerdts).
264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2011.07.135
© 2011 Elsevier Ltd. All rights reserved.
most susceptible to the disease [11]. This trend is due, in part, towaning immunity over time in vaccinated individuals [12–14].
Whooping cough is most severe in neonates who are too youngto have been immunized and in infants who have not been com-pletely immunized [15,16]. Most deaths occur in the first 3 monthsof life [17], before administration of the first dose of the pertus-sis vaccine [17], which is given at two months of age. In mostEuropean and North American countries, pertussis vaccines are cur-rently administered at two, four and six months of age, at a timewhen MatAbs are declining [8,18,19].
Multiple doses of the vaccine are needed for protection asMatAbs pose an important challenge for infant vaccination [20,21].MatAbs result from the mother’s previous exposure to disease orvaccination and are transferred from mother to infant transpla-centally or through the colostrum and breast milk. The mostefficiently transferred immunoglobulin subclass is IgG1 [17]. Pas-sive immunity provides short-lived protection against disease, butalso interferes with vaccination of neonates [22]. Thus, passiveimmunity is a double-edged sword; it protects the young from
disease, but also interferes with immunization of the infant. Forexample, important inhibitory effects on infant antibody responseshave been observed following immunization with vaccines againstmeasles [23,24], Haemophilus influenzae type b conjugate vaccine
Vaccin
[z
ipactoaorpmorsactcp
ird(mFsmptvaLpotpb
itosPewPwcima
2
2
Rsbwa
Monika. Polewicz et al. /
25], hepatitis A [26], influenza A [27], tetanus [28], and varicella-oster [21].
Numerous studies have addressed the issue of MatAbs follow-ng whooping cough vaccination [29–32]. Transplacental MatAbs toertussis antigens can interfere with the infant’s ability to mountntibody responses following immunization with either the wholeell or acellular pertussis pediatric vaccines [30,31,33–35]. MatAbshat possess such inhibitory effects are thought to be a resultf the formation of immune complexes between antibodies andntigen and include the neutralization of antigen, phagocytosisf MatAb-coated antigen, inhibition of B cell activation by Fc�-eceptor mediated signals, and epitope masking by MatAbs thusreventing antigen binding to infant B cells [33,36]. However, theajor determinant of infant antibody responses is found in the ratio
f the antigen concentration to the MatAb concentration. Antibodyesponses may only be obtained when MatAbs have fallen below apecific threshold, and secondary responses may be elicited as soons this threshold is reached [24,33,36]. MatAbs influence infant Bell responses but leave T cell responses unaffected [33,34,37]. Inhe presence of MatAbs, the neonate’s antibody response to the spe-ific antigen is repressed. Consequently, successful vaccines againstertussis must be able to overcome the interference of MatAbs.
Mice are the most commonly used animal model for study-ng pertussis [38–40]. Murine immunization followed by aespiratory challenge has been used to evaluate efficacy ofiphtheria–tetanus–pertussis acellular (DTaP) and whole cellDTwP) vaccines [41,42]. When infected with B. pertussis, neonatal
ice show symptoms such as weight loss and hypoglycemia [43].urthermore, the mouse model is not ideal as it is limited in whichamples can be collected. We developed a new pertussis diseaseodel in newborn piglets [44,45]. Pigs and humans share many
hysiological characteristics [46], including specific functions ofhe innate and adaptive immune response. The model also pro-ides access to various immune compartments and samples suchs serum, colostrum, milk and bronchoalveolar lavage (BAL) fluid.arge amounts of IgG and IgA antibodies are transferred throughorcine colostrum and milk, which allows us to study the effectsf MatAbs. In humans and rodents, maternal IgG is transferredhrough the placenta [47]. Since immunoglobulins cannot cross thelacenta in pregnant sows [48,49], we are able to exchange pigletsetween sows to eliminate the mother effect.
In the present study two animal species were used to exam-ne the effect of MatAb on infant PTd vaccination. We investigatedhe interference of MatAbs on PTd vaccination as well as the rolef a booster dose and adjuvants as a means of trying to overcomeuch interference. Pregnant mice and sows were vaccinated withTd and the offspring were immunized within the first week of lifeither in the presence or absence of MatAbs. While interferenceith vaccination was observed after a single immunization with
Td, co-formulation with adjuvants and a booster immunizationas able to overcome this interference and resulted in the suc-
essful immunization of both neonatal mice and pigs. The level ofnterference was dependent on the immunization regimen for the
other and directly correlated with the amount of MatAbs presentt the time of neonatal vaccination.
. Materials and methods
.1. Animals
Male and female BALB/c mice were purchased from Charlesiver (Montreal, Quebec, Canada). All female mice were housed in
eparate cages. Breeding cages were checked on a daily basis andirths were recorded. Offspring were kept with their mothers untileaned at about four weeks. At that time, the pups were separatedccording to sex.
e 29 (2011) 7718– 7726 7719
Sows were pre-screened for cross-reactive antibodies to Borde-tella bronchiseptica. Pregnant Laundrace sows were purchased fromthe Saskatoon Prairie Swine Centre, University of Saskatchewan.The animals were stimulated to farrow by intramuscular (i.m.)injection of 2 ml prostaglandin (Planate; Schering-Plough CanadaInc., Pointe-Claire, Quebec, Canada) at 113th day of gestation andanother 1 ml at 114th day. Piglets were born on day 114 and 115 ofgestation. Nursing piglets were kept in the same isolation roomsin separate pens. All experiments were performed according tothe guidelines of the University of Saskatchewan and the CanadianCouncil for Animal Care.
2.2. Vaccination of mice and pigs
Both mice and sows and their offspring were immunized withgenetically detoxified pertussis toxoid (PTd) kindly provided byNovartis Vaccines and Diagnostics (Siena, Italy). This antigen wasshown to be completely safe with an antigenic profile compara-ble to wild-type PT, and an immunogenicity that is greater thanchemically detoxified PTd [50]. Dams were immunized one weekprior to becoming pregnant with a subcutaneous injection of 1 �gPTd in phosphate buffered saline (PBS, pH = 7.2, 1.54 mM KH2PO4,155.17 mM NaCl, 2.71 mM Na2HPO4·7 H2O) (Gibco). One week intothe pregnancy (two weeks before delivery) dams were boosted inthe same manner. The control animals were subcutaneously treatedwith 100 �l PBS prior to pregnancy. Pregnant sows were vaccinatedintramuscularly on each side of the neck (trapezius muscle) behindthe ear with 5 �g of PTd in 1 ml of PBS and 30% Emulsigen (MVP Lab-oratories, Ralston, NE; oil-in-water emulsion) resulting in a totaldose of 10 �g/per sow. The control sow received the same volumeof PBS and Emulsigen in each side of the neck. Neonatal mice wererandomly assigned and vaccinated at 7 days of age. At that time,the development of the immune system is comparable to that of anewborn human [51,52]. Pups were either vaccinated with 1 �g ofPTd diluted in PBS or were injected with PBS. All vaccinations were50 �l in volume and injected subcutaneously. Neonatal piglets wererandomly assigned and immunized at 3 days of age with 10 �g PTdplus 150 �g CpG ODN 10101 (Pfizer Canada, Kirkland, Quebec) in1 ml of PBS or treated with 1 ml of PBS. The animals were boostedtwo weeks later in the same manner. All piglets were vaccinatedi.m. in the side of the neck.
2.3. Sample collection
Blood samples were collected from mouse dams before vaccina-tion and at the time of vaccination of their pups. In order to evaluateimmunity in the offspring following vaccination, serum samples ofneonatal pups were collected 2, 4, 6 and 8 weeks post vaccination.All blood samples were centrifuged (4547 × g) and serum stored at−20 ◦C.
Sows were bled before priming, boosting and after farrowing.Sow colostrum and milk samples were collected. Rennet tablets(Sigma–Aldrich) were added to samples and incubated overnightat 37 ◦C. Once clots were formed, the samples were centrifugedat 1349 × g for 20 min. Centrifugation resulted in the formation ofthree layers. The middle layer (whey) was removed and stored in−20 ◦C. Newborn piglets were bled before vaccination as well as1–4 weeks past vaccination.
2.4. ELISA
Polystyrene microtiter plates were coated with 0.25 �g/ml
(100 �l per well) PTd for analysis of murine serum samples. Theantigen was diluted in coating buffer (sodium carbonate buffer,15 mM Na2CO3 and 35 mM NaHCO3, pH 9.6). The plates were incu-bated overnight at 4 ◦C. Sera were diluted four-fold, starting with
7720 Monika. Polewicz et al. / Vaccin
Fig. 1. Induction of IgG antibodies in the serum of dams subcutaneously primedand boosted with 1 �g PTd (�), primed with 1 �g PTd (�) or PBS (�). Dams (six pergroup) were primed four weeks prior to delivery and one group (�) boosted twoweeks later. Serum samples were collected prior to prime (week 0), at the time pupswere born (week 4) as well as at the time the pups were weaned (week 8). Serumsi
1uuosi
When experiments involved more than two groups, data were rank
Fbv
amples were analysed using an ELISA assay. Data sets with differing subscriptsndicate statistical difference (p < 0.05).
:100 dilution. Biotin-conjugated goat anti-mouse immunoglob-lin G (IgG; 1:10,000 dilution; Invitrogen, Camarillo, CA) wassed for the detection of PTd specific IgG. Detection was carried
ut by addition of streptavidin peroxidase (1:5000 dilution; Jack-on Laboratories). Serum samples and antibodies were dilutedn TBST gelatin (Tris buffered saline pH 7.3) containing 0.5%ig. 2. Induction of IgG antibodies in the serum of mouse pups subcutaneously primed woosted with 1 �g PTd (A), primed with 1 �g PTd (B) or PBS vaccinated (C). Pups were praccination and analysed using an ELISA assay (p < 0.05).
e 29 (2011) 7718– 7726
Tween and 0.5% gelatin (Sigma). The reaction was visualized by p-nitrophenylphosphate. No anti-PTd IgA was detected in pup serum.
Polystyrene microtiter plates were coated with 0.5 �g/ml(100 �l per well) PTd for analysis of porcine serum and colostrumsamples. The antigen was diluted in coating buffer. The plates wereincubated overnight at 4 ◦C. The plates were washed and blockedwith TBST gelatin. Sera, colostrum and milk were diluted four-fold, starting with 1:40 dilution. Alkaline-phosphatase-conjugatedgoat anti-pig immunoglobulin G (IgG; 1:5000 dilution; Kirkegaad& Perry Laboratories, Gaithersburg, MD) was used for thedetection of PTd specific IgG. The reaction was visualized by p-nitrophenylphosphate (Sigma–Aldrich). The plates were read at450 nm with an iMark Microplate Reader (Bio-Rad Laboratories).PTd-specific antibody titers were calculated by Microplate Man-ager 6.0 (Bio-Rad Laboratories). To assess B. bronchiseptica specificantibodies, polystyrene microtiter plates (Immulon 2HB; DynexTechnologies, Chantilly, VA) were coated with 2 �g/ml (100 �l perwell) sonicated, heat-inactivated B. bronchiseptica. The ELISA wasperformed as described above.
2.5. Statistical analysis
All statistical analyses were carried out using GraphPad Prismsoftware, version 5.0b. The data from the experiments were notnormally distributed and therefore, the Mann–Whitney test wasused to examine differences between two experimental groups.
transformed and then analysed by one-way analysis of variance(ANOVA). In those instances where the F ratio was significant, dif-ferences among the means of the ranks of the experimental groups
ith 1 �g PTd (�) or PBS (�). Pups were born to three groups of mothers: primed andimed at seven days of age. Serum samples were collected 2, 4, 6 and 8 weeks post
Monika. Polewicz et al. / Vaccine 29 (2011) 7718– 7726 7721
F ed wb inateda
ws
3
3
lg
Ftv
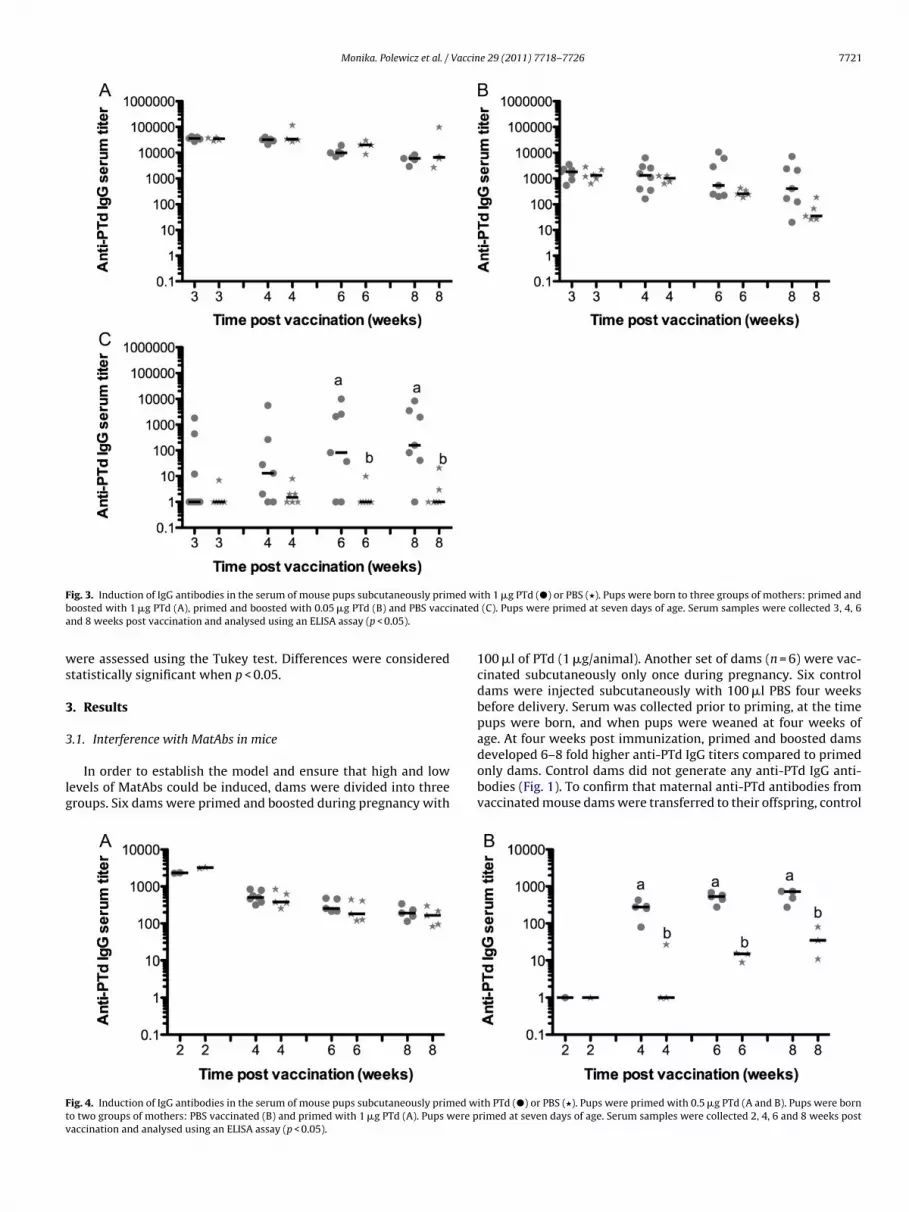
ig. 3. Induction of IgG antibodies in the serum of mouse pups subcutaneously primoosted with 1 �g PTd (A), primed and boosted with 0.05 �g PTd (B) and PBS vaccnd 8 weeks post vaccination and analysed using an ELISA assay (p < 0.05).
ere assessed using the Tukey test. Differences were consideredtatistically significant when p < 0.05.
. Results
.1. Interference with MatAbs in mice
In order to establish the model and ensure that high and lowevels of MatAbs could be induced, dams were divided into threeroups. Six dams were primed and boosted during pregnancy with
ig. 4. Induction of IgG antibodies in the serum of mouse pups subcutaneously primed wo two groups of mothers: PBS vaccinated (B) and primed with 1 �g PTd (A). Pups were paccination and analysed using an ELISA assay (p < 0.05).
ith 1 �g PTd (�) or PBS (�). Pups were born to three groups of mothers: primed and (C). Pups were primed at seven days of age. Serum samples were collected 3, 4, 6
100 �l of PTd (1 �g/animal). Another set of dams (n = 6) were vac-cinated subcutaneously only once during pregnancy. Six controldams were injected subcutaneously with 100 �l PBS four weeksbefore delivery. Serum was collected prior to priming, at the timepups were born, and when pups were weaned at four weeks ofage. At four weeks post immunization, primed and boosted dams
developed 6–8 fold higher anti-PTd IgG titers compared to primedonly dams. Control dams did not generate any anti-PTd IgG anti-bodies (Fig. 1). To confirm that maternal anti-PTd antibodies fromvaccinated mouse dams were transferred to their offspring, controlith PTd (�) or PBS (�). Pups were primed with 0.5 �g PTd (A and B). Pups were bornrimed at seven days of age. Serum samples were collected 2, 4, 6 and 8 weeks post
7722 Monika. Polewicz et al. / Vaccine 29 (2011) 7718– 7726
F ed ant g PTdfi and a
pPimedcfvtorin(rpd
3n
v1eti
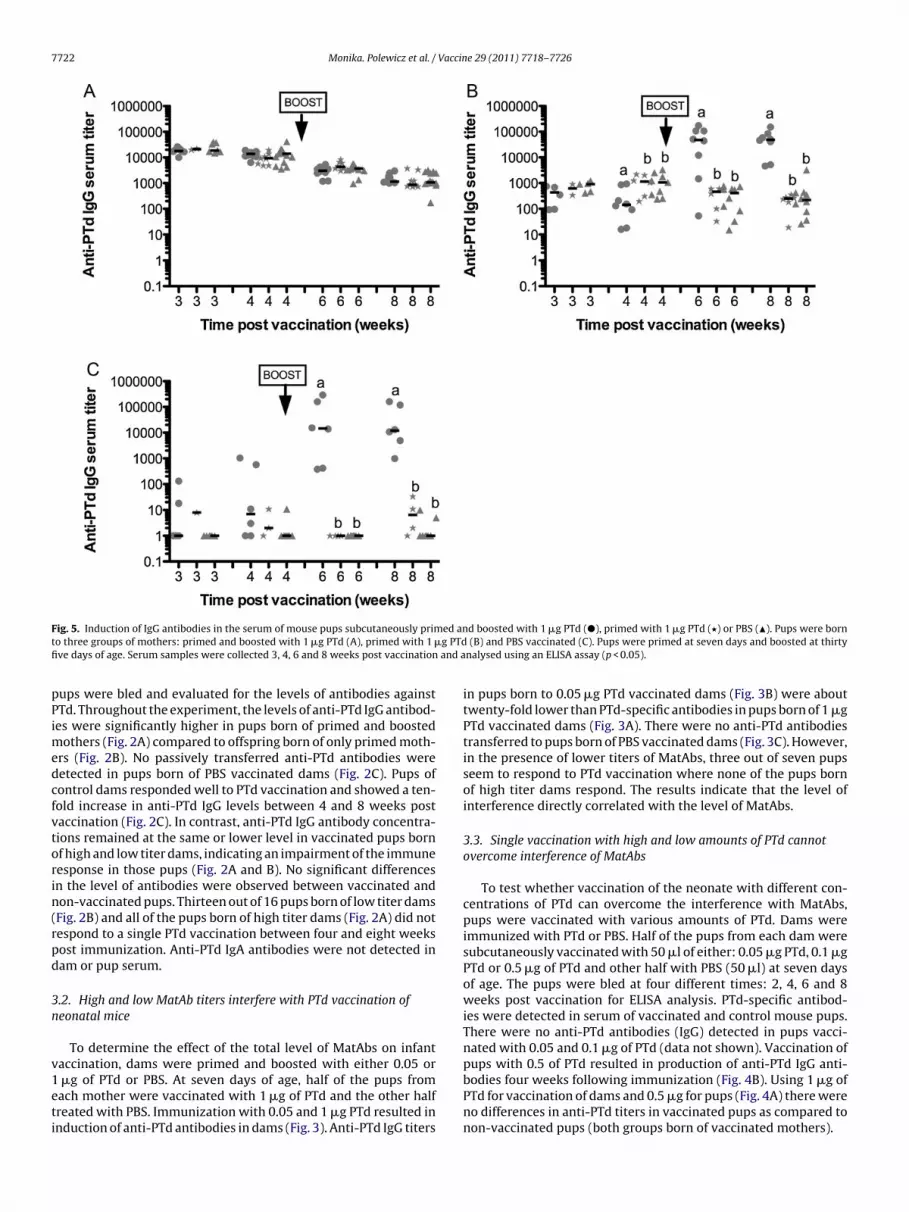
ig. 5. Induction of IgG antibodies in the serum of mouse pups subcutaneously primo three groups of mothers: primed and boosted with 1 �g PTd (A), primed with 1 �ve days of age. Serum samples were collected 3, 4, 6 and 8 weeks post vaccination
ups were bled and evaluated for the levels of antibodies againstTd. Throughout the experiment, the levels of anti-PTd IgG antibod-es were significantly higher in pups born of primed and boosted
others (Fig. 2A) compared to offspring born of only primed moth-rs (Fig. 2B). No passively transferred anti-PTd antibodies wereetected in pups born of PBS vaccinated dams (Fig. 2C). Pups ofontrol dams responded well to PTd vaccination and showed a ten-old increase in anti-PTd IgG levels between 4 and 8 weeks postaccination (Fig. 2C). In contrast, anti-PTd IgG antibody concentra-ions remained at the same or lower level in vaccinated pups bornf high and low titer dams, indicating an impairment of the immuneesponse in those pups (Fig. 2A and B). No significant differencesn the level of antibodies were observed between vaccinated andon-vaccinated pups. Thirteen out of 16 pups born of low titer damsFig. 2B) and all of the pups born of high titer dams (Fig. 2A) did notespond to a single PTd vaccination between four and eight weeksost immunization. Anti-PTd IgA antibodies were not detected inam or pup serum.
.2. High and low MatAb titers interfere with PTd vaccination ofeonatal mice
To determine the effect of the total level of MatAbs on infantaccination, dams were primed and boosted with either 0.05 or
�g of PTd or PBS. At seven days of age, half of the pups fromach mother were vaccinated with 1 �g of PTd and the other halfreated with PBS. Immunization with 0.05 and 1 �g PTd resulted innduction of anti-PTd antibodies in dams (Fig. 3). Anti-PTd IgG titers
d boosted with 1 �g PTd (�), primed with 1 �g PTd (�) or PBS (�). Pups were born (B) and PBS vaccinated (C). Pups were primed at seven days and boosted at thirtynalysed using an ELISA assay (p < 0.05).
in pups born to 0.05 �g PTd vaccinated dams (Fig. 3B) were abouttwenty-fold lower than PTd-specific antibodies in pups born of 1 �gPTd vaccinated dams (Fig. 3A). There were no anti-PTd antibodiestransferred to pups born of PBS vaccinated dams (Fig. 3C). However,in the presence of lower titers of MatAbs, three out of seven pupsseem to respond to PTd vaccination where none of the pups bornof high titer dams respond. The results indicate that the level ofinterference directly correlated with the level of MatAbs.
3.3. Single vaccination with high and low amounts of PTd cannotovercome interference of MatAbs
To test whether vaccination of the neonate with different con-centrations of PTd can overcome the interference with MatAbs,pups were vaccinated with various amounts of PTd. Dams wereimmunized with PTd or PBS. Half of the pups from each dam weresubcutaneously vaccinated with 50 �l of either: 0.05 �g PTd, 0.1 �gPTd or 0.5 �g of PTd and other half with PBS (50 �l) at seven daysof age. The pups were bled at four different times: 2, 4, 6 and 8weeks post vaccination for ELISA analysis. PTd-specific antibod-ies were detected in serum of vaccinated and control mouse pups.There were no anti-PTd antibodies (IgG) detected in pups vacci-nated with 0.05 and 0.1 �g of PTd (data not shown). Vaccination ofpups with 0.5 of PTd resulted in production of anti-PTd IgG anti-
bodies four weeks following immunization (Fig. 4B). Using 1 �g ofPTd for vaccination of dams and 0.5 �g for pups (Fig. 4A) there wereno differences in anti-PTd titers in vaccinated pups as compared tonon-vaccinated pups (both groups born of vaccinated mothers).
Monika. Polewicz et al. / Vaccine 29 (2011) 7718– 7726 7723
Fig. 6. Induction of IgG antibodies in the serum of mouse pups subcutaneously primed as neonates and boosted with 1 �g PTd once MatAbs waned (�), primed with 1 �gP primeP ples ww
3th
wiiMo3
(s(bdmhebtwTtiinap
Td once MatAbs waned (�) or PBS (�). Pups were born to three groups of mothers:ups were primed at seven days and boosted at thirty five days of age. Serum samere analysed using an ELISA assay (p < 0.05).
.4. A booster immunization can overcome MatAb interference inhe presence of low titers of MatAbs but not in the presence ofigh titers of MatAbs
Previous experiments showed that vaccination of neonatal miceith 1 �g of PTd without adjuvants was not able to overcome the
nterference of MatAbs. We introduced a booster shot of PTd to seef this could overcome the interference. Neonatal pups born of high
atAb titer dams and low MatAb titer dams were either immunizednce or twice. Control pups were injected with PBS. Pups were bled, 4, 6 and 8 weeks post vaccination.
Passive transfer of antibodies to pups born to vaccinated damsFig. 5B) was about 10-fold lower than the passive transfer of PTdpecific antibodies to pups born of vaccinated and boosted damsFig. 5A). There were no anti-PTd antibodies transferred to pupsorn of PBS vaccinated dams (Fig. 5C). Waning of MatAbs was evi-ent in both groups at eight weeks at which time the levels ofaternal anti-PTd antibodies were still high in offspring born to
igh titer dams. Vaccinating and boosting pups born to naïve moth-rs resulted in superior antibody production one week after theoost. All of the six animals induced anti-PTd antibodies followinghe boost (Fig. 5C). It was evident that a boost immunization of pupsas not able to overcome interference of high titers of MatAbs.
wice immunized pups had similar anti-PTd IgG serum concen-ration as compared to PBS vaccinated pups (Fig. 5A). Boostermmunization of pups with PTd was able to overcome interference
n the presence of lower concentrations of MatAbs (single vacci-ated dams). Following the boost, anti-PTd antibody titers werebout 100-fold higher than in single vaccinated pups or controlups (Fig. 5B).d and boosted with 1 �g PTd (A), primed with 1 �g PTd (B) and PBS vaccinated (C).ere collected every two weeks. Pre boost, 16, 18 and 20 weeks post boost bleeds
3.5. Priming occurred in the presence of MatAbs
To test if it would be possible to prime an immune responsein the presence of MatAbs, neonatal pups were vaccinated in thepresence of MatAbs and boosted once MatAbs waned. Dams werevaccinated once or twice with PTd or PBS during pregnancy. Atseven days of age, a third of the pups of each dam were subcu-taneously vaccinated with 1 �g of PTd and a third of all pups withPBS. Throughout the experiment pup serum was analysed for thepresence of maternal anti-PTd antibodies. Once MatAbs were nolonger detectable in pup serum (16 weeks), animals were boosted.Pups not primed as neonates were vaccinated for the first timeonce MatAbs were no longer detectable. Pups born to naïve damshad high anti-PTd IgG titers following a single vaccination. Anti-body titers increased significantly following the boost (Fig. 6C), asexpected for a secondary response. Similarly, titers in pups bornof vaccinated dams and boosted at 16 weeks of age significantlyincreased following the boost (Fig. 6A and B). Primed and boostedpups had much higher antibody titers at 18 and 20 weeks of agethan singly vaccinated pups, demonstrating that this was a resultof a secondary response.
3.6. Porcine model of MatAb interference
To study MatAb interference in pigs, sows were vaccinatedintramuscularly with PTd (10 �g) and emulsigen four weeks prior
to farrowing, and boosted after two weeks. The control sow wastreated at the same time points with the same volume of PBS. Sowserum samples were taken before each vaccination and before far-rowing. After farrowing, sow colostrum and milk samples were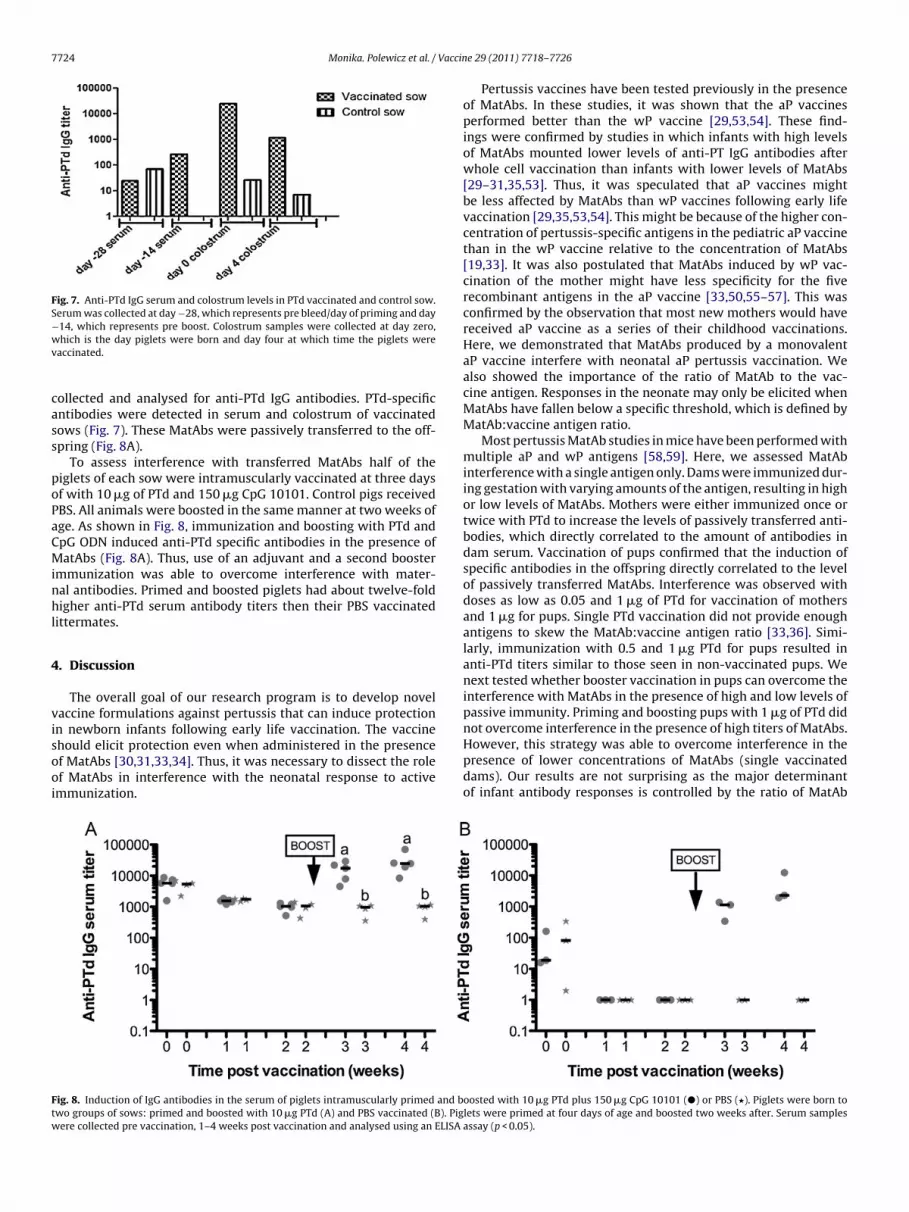
7724 Monika. Polewicz et al. / Vaccin
Fig. 7. Anti-PTd IgG serum and colostrum levels in PTd vaccinated and control sow.Serum was collected at day −28, which represents pre bleed/day of priming and day−14, which represents pre boost. Colostrum samples were collected at day zero,wv
cass
poPaCMinhl
4
visooi
However, this strategy was able to overcome interference in the
Ftw
hich is the day piglets were born and day four at which time the piglets wereaccinated.
ollected and analysed for anti-PTd IgG antibodies. PTd-specificntibodies were detected in serum and colostrum of vaccinatedows (Fig. 7). These MatAbs were passively transferred to the off-pring (Fig. 8A).
To assess interference with transferred MatAbs half of theiglets of each sow were intramuscularly vaccinated at three daysf with 10 �g of PTd and 150 �g CpG 10101. Control pigs receivedBS. All animals were boosted in the same manner at two weeks ofge. As shown in Fig. 8, immunization and boosting with PTd andpG ODN induced anti-PTd specific antibodies in the presence ofatAbs (Fig. 8A). Thus, use of an adjuvant and a second booster
mmunization was able to overcome interference with mater-al antibodies. Primed and boosted piglets had about twelve-foldigher anti-PTd serum antibody titers then their PBS vaccinated
ittermates.
. Discussion
The overall goal of our research program is to develop novelaccine formulations against pertussis that can induce protectionn newborn infants following early life vaccination. The vaccinehould elicit protection even when administered in the presence
f MatAbs [30,31,33,34]. Thus, it was necessary to dissect the rolef MatAbs in interference with the neonatal response to activemmunization.ig. 8. Induction of IgG antibodies in the serum of piglets intramuscularly primed and bwo groups of sows: primed and boosted with 10 �g PTd (A) and PBS vaccinated (B). Pigere collected pre vaccination, 1–4 weeks post vaccination and analysed using an ELISA
e 29 (2011) 7718– 7726
Pertussis vaccines have been tested previously in the presenceof MatAbs. In these studies, it was shown that the aP vaccinesperformed better than the wP vaccine [29,53,54]. These find-ings were confirmed by studies in which infants with high levelsof MatAbs mounted lower levels of anti-PT IgG antibodies afterwhole cell vaccination than infants with lower levels of MatAbs[29–31,35,53]. Thus, it was speculated that aP vaccines mightbe less affected by MatAbs than wP vaccines following early lifevaccination [29,35,53,54]. This might be because of the higher con-centration of pertussis-specific antigens in the pediatric aP vaccinethan in the wP vaccine relative to the concentration of MatAbs[19,33]. It was also postulated that MatAbs induced by wP vac-cination of the mother might have less specificity for the fiverecombinant antigens in the aP vaccine [33,50,55–57]. This wasconfirmed by the observation that most new mothers would havereceived aP vaccine as a series of their childhood vaccinations.Here, we demonstrated that MatAbs produced by a monovalentaP vaccine interfere with neonatal aP pertussis vaccination. Wealso showed the importance of the ratio of MatAb to the vac-cine antigen. Responses in the neonate may only be elicited whenMatAbs have fallen below a specific threshold, which is defined byMatAb:vaccine antigen ratio.
Most pertussis MatAb studies in mice have been performed withmultiple aP and wP antigens [58,59]. Here, we assessed MatAbinterference with a single antigen only. Dams were immunized dur-ing gestation with varying amounts of the antigen, resulting in highor low levels of MatAbs. Mothers were either immunized once ortwice with PTd to increase the levels of passively transferred anti-bodies, which directly correlated to the amount of antibodies indam serum. Vaccination of pups confirmed that the induction ofspecific antibodies in the offspring directly correlated to the levelof passively transferred MatAbs. Interference was observed withdoses as low as 0.05 and 1 �g of PTd for vaccination of mothersand 1 �g for pups. Single PTd vaccination did not provide enoughantigens to skew the MatAb:vaccine antigen ratio [33,36]. Simi-larly, immunization with 0.5 and 1 �g PTd for pups resulted inanti-PTd titers similar to those seen in non-vaccinated pups. Wenext tested whether booster vaccination in pups can overcome theinterference with MatAbs in the presence of high and low levels ofpassive immunity. Priming and boosting pups with 1 �g of PTd didnot overcome interference in the presence of high titers of MatAbs.
presence of lower concentrations of MatAbs (single vaccinateddams). Our results are not surprising as the major determinantof infant antibody responses is controlled by the ratio of MatAb
oosted with 10 �g PTd plus 150 �g CpG 10101 (�) or PBS (�). Piglets were born tolets were primed at four days of age and boosted two weeks after. Serum samplesassay (p < 0.05).

Vaccin
lwMawPt
wbvaai
bMcfaapefmpi
A
GCdD
rafe
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
Monika. Polewicz et al. /
evels to the concentration of vaccine antigen [33,36]. Importantly,e showed that it is possible to prime in the presence of MatAbs.ice primed as neonates had higher anti-PTd IgG titers following
second immunization at 16 weeks of age than their littermatesho were immunized once as adults. We did not detect any anti-
Td IgA antibodies in the dam and pup serum. This might be due tohe systemic route of vaccination of dams.
In order to enhance anti-PTd antibody induction in piglets,e included an adjuvant in our vaccine formulation and added a
ooster shot. At three days of age, half of the piglets per sow wereaccinated and boosted with 10 �g of PTd and 150 �g CpG 10101nd another half with PBS. Use of 10 �g PTd and CpG 10101 as andjuvant for priming and boosting piglets was able to overcomenterference by MatAbs.
The current study was performed to examine the interactionetween PTd and MatAbs. We established the negative impact ofatAbs and their interference on neonatal murine and porcine vac-
ination. We determined that including an adjuvant in our vaccineormulations or adding a booster vaccine can overcome the neg-tive effects of passive immunity. Our next goal is to use noveldjuvants such as CpG ODN, innate defense regulator peptides andolyphosphazenes in our vaccine formulations as well as new deliv-ry mechanisms that could potentially protect the vaccine antigenrom neutralizing MatAbs. In subsequent experiments we plan to
easure the priming of the immune response in neonates in theresence of MatAbs and the degree of protection against challenge
nfection.
cknowledgements
This work is funded by a grant through: the Grand Challenges inlobal Health Initiative by the Bill and Melinda Gates Foundation,anadian Institutes of Health Research (CIHR), the Krembil Foun-ation and the Research Alliance for the Prevention of Infectiousiseases (RAPID).
We would like to thank VIDO Animal Care, especially Barry Car-ol and Sherry Tetland for performing the animal breeding, bleedingnd vaccination. Stacy Strom, Jill Van Kessel and Rachelle Buchananor assistance in laboratory work. We would also like to acknowl-dge Dr. Hugh Townsend for his help with the statistical analysis.
eferences
[1] Mattoo S, Cherry JD. Molecular pathogenesis, epidemiology, and clinical man-ifestations of respiratory infections due to Bordetella pertussis and otherBordetella subspecies. Clin Microbiol Rev 2005;18(April (2)):326–82.
[2] Pertussis vaccines. Wkly Epidemiol Rec 1999;74(May (18)):137–43.[3] Ivanoff B, Robertson SE. Pertussis: a worldwide problem. Dev Biol Stand
1997;89:3–13.[4] Crowcroft NS, Pebody RG. Recent developments in pertussis. Lancet
2006;367(June (9526)):1926–36.[5] Vitek CR, Pascual FB, Baughman AL, Murphy TV. Increase in deaths from per-
tussis among young infants in the United States in the 1990s. Pediatr Infect DisJ 2003;22(July (7)):628–34.
[6] Tanaka M, Vitek CR, Pascual FB, Bisgard KM, Tate JE, Murphy TV.Trends in pertussis among infants in the United States, 1980–1999. JAMA2003;290(December (22)):2968–75.
[7] Ntezayabo B, De Serres G, Duval B. Pertussis resurgence in Canada largelycaused by a cohort effect. Pediatr Infect Dis J 2003;22(January (1)):22–7.
[8] Healy CM, Munoz FM, Rench MA, Halasa NB, Edwards KM, Baker CJ. Prevalenceof pertussis antibodies in maternal delivery, cord, and infant serum. J Infect Dis2004;190(July (2)):335–40.
[9] Forsyth KD, Campins-Marti M, Caro J, Cherry JD, Greenberg D, Guiso N, et al.New pertussis vaccination strategies beyond infancy: recommendations by theglobal pertussis initiative. Clin Infect Dis 2004;39(December (12)):1802–9.
10] Skowronski DM, De Serres G, MacDonald D, Wu W, Shaw C, Macnabb J, et al.The changing age and seasonal profile of pertussis in Canada. J Infect Dis2002;185(May (10)):1448–53.
11] Wendelboe AM, Njamkepo E, Bourillon A, Floret DD, Gaudelus J, Gerber M,et al. Transmission of Bordetella pertussis to young infants. Pediatr Infect Dis J2007;26(April (4)):293–9.
12] Theilen U, Johnston ED, Robinson PA. Rapidly fatal invasive pertussis in younginfants—how can we change the outcome? BMJ 2008;337:a343.
[
e 29 (2011) 7718– 7726 7725
13] Wood N, McIntyre P. Pertussis: review of epidemiology, diagnosis, manage-ment and prevention. Paediatr Respir Rev 2008;9(September (3)):201–11, quiz11–2.
14] Tan T, Halperin S, Cherry JD, Edwards K, Englund JA, Glezen P, et al. Pertussisimmunization in the global pertussis initiative North American region: rec-ommended strategies and implementation considerations. Pediatr Infect Dis J2005;24(May (5 Suppl.)):S83–6.
15] Pertussis deaths—United States, 2000. MMWR Morb Mortal Wkly Rep2002;51(July (28)):616–8.
16] Pertussis—United States, 1997–2000. MMWR Morb Mortal Wkly Rep2002;51(February (4)):73–6.
17] Van Rie A, Wendelboe AM, Englund JA. Role of maternal pertussis antibodiesin infants. Pediatr Infect Dis J 2005;24(May (5 Suppl.)):S62–5.
18] Edwards KM. Pertussis: an important target for maternal immunization. Vac-cine 2003;21(July (24)):3483–6.
19] Mooi FR, de Greeff SC. The case for maternal vaccination against pertussis.Lancet Infect Dis 2007;7(September (9)):614–24.
20] Osborn JJ, Dancis J, Julia JF. Studies of the immunology of the newborn infant.II. Interference with active immunization by passive transplacental circulatingantibody. Pediatrics 1952;10(September (3)):328–34.
21] Glezen WP. Effect of maternal antibodies on the infant immune response. Vac-cine 2003;21(July (24)):3389–92.
22] Siegrist CA, Cordova M, Brandt C, Barrios C, Berney M, Tougne C, et al. Deter-minants of infant responses to vaccines in presence of maternal antibodies.Vaccine 1998;16(August–September (14–15)):1409–14.
23] Albrecht P, Ennis FA, Saltzman EJ, Krugman S. Persistence of maternal antibodyin infants beyond 12 months: mechanism of measles vaccine failure. J Pediatr1977;91(November (5)):715–8.
24] Crowe Jr JE. Influence of maternal antibodies on neonatal immunization againstrespiratory viruses. Clin Infect Dis 2001;33(November (10)):1720–7.
25] Daum RS, Siber GR, Ballanco GA, Sood SK. Serum anticapsular antibody responsein the first week after immunization of adults and infants with the Haemophilusinfluenzae type b-Neisseria meningitidis outer membrane protein complex con-jugate vaccine. J Infect Dis 1991;164(December (6)):1154–9.
26] Kanra G, Yalcin SS, Ceyhan M, Yurdakok K. Clinical trial to evaluateimmunogenicity and safety of inactivated hepatitis A vaccination starting at2-month-old children. Turk J Pediatr 2000;42(April–June (2)):105–8.
27] Karron RA, Steinhoff MC, Subbarao EK, Wilson MH, Macleod K, Clements ML,et al. Safety and immunogenicity of a cold-adapted influenza A (H1N1) reassor-tant virus vaccine administered to infants less than six months of age. PediatrInfect Dis J 1995;14(January (1)):10–6.
28] Sarvas H, Kurikka S, Seppala IJ, Makela PH, Makela O. Maternal antibodies partlyinhibit an active antibody response to routine tetanus toxoid immunization ininfants. J Infect Dis 1992;165(May (5)):977–9.
29] Englund JA, Anderson EL, Reed GF, Decker MD, Edwards KM, Pichichero ME,et al. The effect of maternal antibody on the serologic response and the inci-dence of adverse reactions after primary immunization with acellular andwhole-cell pertussis vaccines combined with diphtheria and tetanus toxoids.Pediatrics 1995;96(September (3 Pt. 2)):580–4.
30] Burstyn DG, Baraff LJ, Peppler MS, Leake RD, St Geme Jr J, ManclarkCR. Serological response to filamentous hemagglutinin and lymphocytosis-promoting toxin of Bordetella pertussis. Infect Immun 1983;41(September (3)):1150–6.
31] Baraff LJ, Leake RD, Burstyn DG, Payne T, Cody CL, Manclark CR, et al. Immuno-logic response to early and routine DTP immunization in infants. Pediatrics1984;73(January (1)):37–42.
32] Provenzano RW, Wetterlow LH, Sullivan CL. Immunization and antibodyresponse in the newborn infant. I. Pertussis inoculation within twenty-fourhours of birth. N Engl J Med 1965;273(October (18)):959–65.
33] Siegrist CA. Mechanisms by which maternal antibodies influence infant vaccineresponses: review of hypotheses and definition of main determinants. Vaccine2003;21(July (24)):3406–12.
34] Siegrist CA. The challenges of vaccine responses in early life: selected examples.J Comp Pathol 2007;137(July (Suppl. 1)):S4–9.
35] Heininger U, Cherry JD, Christenson PD, Eckhardt T, Goering U, Jakob P,et al. Comparative study of Lederle/Takeda acellular and Lederle whole-cell pertussis-component diphtheria–tetanus–pertussis vaccines in infants inGermany. Vaccine 1994;12(January (1)):81–6.
36] Siegrist CA. Neonatal and early life vaccinology. Vaccine 2001;19(May(25–26)):3331–46.
37] Siegrist CA, Barrios C, Martinez X, Brandt C, Berney M, Cordova M, et al. Influenceof maternal antibodies on vaccine responses: inhibition of antibody but notT cell responses allows successful early prime-boost strategies in mice. Eur JImmunol 1998;28(December (12)):4138–48.
38] Elahi S, Holmstrom J, Gerdts V. The benefits of using diverse animal models forstudying pertussis. Trends Microbiol 2007;15(October (10)):462–8.
39] Standfast AF. The comparison between field trials and mouse protectiontests against intranasal and intracerebral challenges with Bordetella pertussis.Immunology 1958;1(April (2)):135–43.
40] Mills KH, Brady M, Ryan E, Mahon BP. A respiratory challenge model for infec-tion with Bordetella pertussis: application in the assessment of pertussis vaccine
potency and in defining the mechanism of protective immunity. Dev Biol Stand1998;95:31–41.41] Guiso N, Capiau C, Carletti G, Poolman J, Hauser P. Intranasal murine modelof Bordetella pertussis infection. I. Prediction of protection in human infants byacellular vaccines. Vaccine 1999;17(May (19)):2366–76.

7 Vaccin
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
726 Monika. Polewicz et al. /
42] Mills KH. Immunity to Bordetella pertussis. Microbes Infect 2001;3(July(8)):655–77.
43] Pittman M, Furman BL, Wardlaw AC. Bordetella pertussis respiratory tract infec-tion in the mouse: pathophysiological responses. J Infect Dis 1980;142(July(1)):56–66.
44] Elahi S, Brownlie R, Korzeniowski J, Buchanan R, O’Connor B, Peppler MS, et al.Infection of newborn piglets with Bordetella pertussis: a new model for pertus-sis. Infect Immun 2005;73(June (6)):3636–45.
45] Elahi S, Buchanan RM, Babiuk LA, Gerdts V. Maternal immunity providesprotection against pertussis in newborn piglets. Infect Immun 2006;74(May(5)):2619–27.
46] Rothkotter HJ, Sowa E, Pabst R. The pig as a model of developmen-tal immunology. Hum Exp Toxicol 2002;21(September–October (9–10)):533–6.
47] Goldman AS. Evolution of the mammary gland defense system and theontogeny of the immune system. J Mammary Gland Biol Neoplasia 2002;7(July(3)):277–89.
48] Salmon H, Berri M, Gerdts V, Meurens F. Humoral and cellular factors of mater-nal immunity in swine. Dev Comp Immunol 2009;33(March (3)):384–93.
49] Boersma WJ, Van Rooij EM, Scholten JW, Zwart RJ, Kimman TG, Bianchi A. Silentmemory induction in maternal immune young animals. Vet Q 1998;20(Suppl.
3):S89–92.50] Rappuoli R. The vaccine containing recombinant pertussis toxin induces earlyand long-lasting protection. Biologicals 1999;27(June (2)):99–102.
51] Roduit C, Bozzotti P, Mielcarek N, Lambert PH, del Giudice G, Locht C,et al. Immunogenicity and protective efficacy of neonatal vaccination against
[
e 29 (2011) 7718– 7726
Bordetella pertussis in a murine model: evidence for early control of pertussis.Infect Immun 2002;70(July (7)):3521–8.
52] Siegrist CA. Vaccination in the neonatal period and early infancy. Int RevImmunol 2000;19(2–3):195–219.
53] Van Savage J, Decker MD, Edwards KM, Sell SH, Karzon DT. Natural history ofpertussis antibody in the infant and effect on vaccine response. J Infect Dis1990;161(March (3)):487–92.
54] Belloni C, De Silvestri A, Tinelli C, Avanzini MA, Marconi M, Strano F, et al.Immunogenicity of a three-component acellular pertussis vaccine adminis-tered at birth. Pediatrics 2003;111(May (5 Pt. 1)):1042–5.
55] Heron I, Chen FM, Fusco J. DTaP vaccines from North American vaccine (NAVA):composition and critical parameters. Biologicals 1999;27(June (2)):91–6.
56] Ibsen PH. The effect of formaldehyde, hydrogen peroxide and genetic detox-ification of pertussis toxin on epitope recognition by murine monoclonalantibodies. Vaccine 1996;14(April (5)):359–68.
57] Murphy TV, Slade BA, Broder KR, Kretsinger K, Tiwari T, Joyce PM, et al.Prevention of pertussis, tetanus, and diphtheria among pregnant and postpar-tum women and their infants recommendations of the Advisory Committeeon Immunization Practices (ACIP). MMWR Recomm Rep 2008;57(May (RR-4)):1–51.
58] Oda M, Izumiya K, Sato Y, Hirayama M. Transplacental and transcolostral immu-
nity to pertussis in a mouse model using acellular pertussis vaccine. J Infect Dis1983;148(July (1)):138–45.59] Sato H, Sato Y. Protective antigens of Bordetella pertussis mouse-protectiontest against intracerebral and aerosol challenge of B. pertussis. Dev Biol Stand1985;61:461–7.