adenosine a 2a receptors interact with gabaergic pathways to modulate respiration in neonatal...
TRANSCRIPT

Respiratory Physiology & Neurobiology 141 (2004) 201–211
Adenosine A2A receptors interact with GABAergic pathwaysto modulate respiration in neonatal piglets
Christopher G. Wilsona,∗, Richard J. Martina, Marwan Jabera, Jalal Abu-Shaweesha,Anjun Jafrib, Musa A. Haxhiua,b, Syed Zaidib
a Department of Pediatrics, Case Western Reserve University School of Medicine, Division of Neonatology,Suite 3100, 11100 Euclid Avenue, Cleveland, OH 44106, USA
b Department of Physiology and Biophysics, Howard University College of Medicine, Washington, DC 20059, USA
Accepted 22 April 2004
Abstract
GABA and adenosine contribute to respiratory inhibition in early postnatal life. In this study the adenosine A2A receptoragonist CGS21680 was used to evaluate adenosine receptor specificity and the interrelation of adenosine and GABA in theinhibition of inspiratory drive. In neonatal piglets (n = 10), CGS21680 was injected into the fourth ventricle resulting in apneaand/or decreased burst area and frequency of phrenic discharge. Phrenic burst area decreased to 58.9± 8.6% (S.E.M.) afterCGS21680 injection (control= 91.8± 1.0%). Expiratory time increased 261.0± 59.9% after CGS21680 from control (87.7± 2.7%). When bicuculline was injected locally within the rostral ventrolateral medulla (n = 5), or into the fourth ventricle (n= 5), the CGS21680 induced inhibition of phrenic was abolished. To define expression of A2A receptor at the message level(mRNA), we employed in situ hybridization with a digoxigenin-coupled oligonucleotide. Adenosine A2A receptor mRNA wasexpressed in regions of the medulla oblongata known to contain GABAergic neurons. We conclude that GABAergic inputsaffecting respiratory timing and inspiratory drive are modulated by activation of A2A receptors. These findings offer new insightinto the mechanism whereby xanthine therapy diminishes apnea of prematurity.© 2004 Elsevier B.V. All rights reserved.
Keywords: Development, respiratory inhibition; Mammals, Piglet; Pharmacological agents, A2A receptor agonist CGS21680; Receptor, A2A,GABAA; Ventilation, drive, inhibition by GABA, adenosine
1. Introduction
Immaturity of respiratory neural control appears tobe a universal feature of early postnatal life, especiallyafter preterm gestation. This is manifested by char-
∗ Corresponding author. Tel.:+1 216 368 3757;fax: +1 216 844 3380.
E-mail address: [email protected] (C.G. Wilson).
acteristic responses of respiratory timing, resulting inprolongation of expiratory duration in response to hy-percapnia, hypoxia or stimulation of laryngeal affer-ents (Abu-Shaweesh et al., 2001; Miller et al., 2000;Dreshaj et al., 1999). Apnea of prematurity appears tobe a consequence of immature respiratory neural con-trol, and is a particularly troublesome clinical prob-lem in low birth weight infants (Miller et al., 2003).Therefore it is important to characterize the role of in-
1569-9048/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.resp.2004.04.012

202 C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211
hibitory neurotransmitter mediated pathways that con-tribute to inhibition in central respiratory circuits inearly development.
We have previously demonstrated that centrally me-diated �-aminobutyric acid (GABA)-ergic pathwayscontribute greatly to this inhibition of inspiration thatcharacterizes respiratory reflex responses in early post-natal life (Abu-Shaweesh et al., 2001; Miller et al.,2000; Dreshaj et al., 1999). In addition to GABA,adenosine is also known to play an important role asa central inhibitor of respiratory neural output dur-ing early maturation (Koos et al., 2001). The mecha-nism whereby adenosine inhibits inspiratory drive isnot well characterized even though xanthines, whichblock adenosine receptor subtypes, are widely usedin preterm infants exhibiting neonatal apnea. It hasbeen assumed that the respiratory stimulating effectsof caffeine are secondary to blockade of inhibitoryadenosine A1 receptors (Herlenius et al., 1997, 2002).However, excitatory adenosine A2A receptors are alsowidely distributed throughout the developing brain(Weaver, 1993) and A2A receptor activation mediateshypoxic inhibition of fetal breathing in sheep (Kooset al., 2002), by unknown mechanisms. We hypothe-sized that GABAergic mechanisms may be involvedin A2A receptor activation-induced respiratory depres-sion.
Therefore, our focus in this work is the interac-tion between GABA and adenosine, and the means bywhich they may act synergistically to modulate res-piratory neural output in early postnatal life. To ad-dress this aim we examined whether medullary regionsknown to contain GABAergic neurons express A2Areceptors, and whether activation of adenosine A2A re-ceptors in the brainstem induces inspiratory inhibitionvia GABA release and activation of GABAA receptorsignaling pathways.
2. Methods
2.1. Surgical preparation: physiological studies
We studied piglets aged 5–10 days, purchased froma local vendor for all procedures reported (N = 10,average weight= 2.2 ± 0.5 kg). Animal care and allprotocols were in accordance with the Case WesternInstitutional Animal Care and Use (IACUC) commit-
tee. All piglets were sedated with intramuscular xy-lazine (1.4 mg/kg) and ketamine (7.2 mg/kg), followedby intravenous thiopental sodium (25–30 mg/kg) ad-ministered via peripheral ear vein. Animals were thenplaced on an electric heating pad to maintain a bodytemperature of 36.5–37.5◦C. Femoral arterial andvenous catheters were inserted to allow continuousmeasurement of blood pressure, periodic measure-ment of blood gases, and for the administration ofsupplemental anesthetic, paralytic agent, and fluids. Amidline incision in the neck was made to allow accessto the trachea and vagi. The animals underwent bilat-eral vagotomy to avoid the effects of volume changeon respiratory timing. A tracheal cannula was securedand the piglet was then placed on a volume ventilator(Harvard Apparatus). Ventilator frequency and vol-ume were adjusted to maintain a PaCO2 of 35–45 Torrand a PaO2 of greater than 120 Torr (RadiometrieABL-5). Room air was supplemented by 100%O2 through a T-connection to the inlet side of theventilator.
To avoid potentially confounding effects of anes-thesia on respiration, we performed a midcolliculardecerebration as previously described (Dreshaj et al.,1999). Briefly, we made an approximately 4 cm longincision over the parietal ridge of the piglet’s skull,the skin was reflected, and a narrow line of bonewas removed parallel to the parietal ridge. Once suf-ficient bone was removed to reveal the dura overly-ing the cortex, we incised the dura on the left andright sides, avoiding cutting the dura near the supe-rior sagittal sinus. We then inserted a thin spatula,angled at approximately 70◦ vertical to the plane ofthe skull, and advanced it ventrally until the floorof the skull was reached and then swung the spat-ula gently left and right to the lateral margins of theskull, completing the decerebration. After approxi-mately 1 h, we checked for withdrawal reflexes fromhoof or tail pinches to assure recovery from the ini-tial anesthesia and then, just prior to data collection,we paralyzed the piglet by administering gallaminetriethiodide (10 mg/kg).
We isolated the phrenic nerve by extending the neckincision caudally, dissecting out the phrenic, desheath-ing the nerve, cutting the nerve and covering it withpetroleum jelly and mineral oil. After isolation of thephrenic, the piglet was placed in a stereotaxic frameand the brainstem was exposed through an incision

C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211 203
over the caudal skull extending to C2. The skin and in-tervening muscle were reflected to allow clear accessto the occipital bone and the atlanto-occipital mem-brane. Careful sectioning of the atlanto-occipital mem-brane and underlying dura allowed clear visualizationof the brainstem. After exposing the brainstem, a highspeed rotary tool (Dremel) with a cutting bit was usedto remove the caudal portion of the occipital bone andexpose the midline of the cerebellum. Bilateral holeswere drilled for subsequent injection of bicuculline.The phrenic nerve was then isolated and the centralend was placed over a bipolar hook recording elec-trode attached to a high-impedance head stage feed-ing a Grass P-511 amplifier which was then fed to ourcomputerized acquisition and analysis system.
2.2. Physiologic protocols
We used intracisternal injections of the adenosineA2A specific agonist CGS21680 (25–50�l of 10 mMstock, dissolved in DMSO, Sigma) to determine theeffects of adenosine receptor activation upon breath-ing pattern (Fig. 1). Ultimately we chose the minimalCGS dose that resulted in an apnea in all the animalstested (50�L of 10 mM CGS21680). Lower doses hadvariable effects on peak phrenic activity and respira-tory timing, but the 50�L dose repeatedly caused inhi-bition of phrenic area and timing (Fig. 2). We definedapnea as a doubling of expiratory time, when com-pared toTE in the control period before CGS injection.We performed a control experiment using 50�L in-jections of vehicle and saw no changes in respirationor blood pressure.
To block GABAA receptors we administered bicu-culline (methiodide form, 50�L of 3 mM, Sigma). Weprepared individual aliquots at the appropriate concen-tration and froze them for use on the day of the ex-periment. We used a dose of bicuculline that blockedapneas induced by superior laryngeal nerve stimula-tion in the experiments ofAbu-Shaweesh et al. (2001).Bicuculline was administered in two different ways,either by intracisternal injection (N = 3) or by mi-croinjection into the rostral ventral lateral medulla (co-ordinates: 5.0 mm rostral to obex, 5.0 mm lateral tomidline, and 2.0 mm dorsal to ventral surface of thebrainstem,N = 2). Injection of bicuculline either in-tracisternally or local to the respiratory circuitry of thebrainstem resulted in the same effect regardless of the
point of injection so we have pooled the results fromthe two different injection sites.
2.3. Data collection and statistical analysis
We used PowerLab hardware and Chart software(both from AD Instruments, CA) to capture signalsin real-time (4 kHz sampling rate). Blood pressure,raw phrenic neurogram (bandpass 100–1000 Hz), fullwave rectified and moving averaged (100 ms window)phrenic neurogram, and expired CO2 were recorded.We analyzed data on a breath-by-breath basis by mea-suring area of phrenic discharge, inspiratory, and expi-ratory times during the control period, and after drugadministration. Typically the five breaths before an in-tervention were used to calculate normalized controlvalues (all five averaged, divided by the largest valueand multiplied by 100 to give normalized percent-age). All treatments were compared to control usingone-way analysis of variance and values are reportedas normalized values (mean± S.E.M.).
2.4. Neuroanatomical studies, in situ hybridization
We studied the distribution of adenosine A2A re-ceptor mRNA in the medulla oblongata of pigletsaged 5–10 days (n = 3). Healthy piglets were deeplyanesthetized (pentobarbital, 50 mg/kg) and intracar-dially perfused with Ca2+ free Tyrode solution fol-lowed by 4% paraformaldehyde. The brainstem wasremoved and postfixed for 3 days in 4% paraformalde-hyde containing 30% sucrose. For in situ hybridiza-tion, we cut coronal sections of the medulla oblon-gata (40�m) from the pontomedullary border to C1for each animal using a cryostat (Hacker Instruments,NJ). Non-radioactive in situ hybridization immunohis-tochemistry for adenosine A2A receptor mRNA wasperformed as previously described byBrady et al.(1999).
We obtained rat adenosine A2A receptor probefrom Dr. Estelle Gauda (Johns Hopkins, Baltimore,MD, Gauda et al., 2000). The plasmid templatewas linearized by digestion with BamH1 or HindIII to generate the corresponding sense or antisensetranscripts, under T7 or T3 promoters, respectively.Post-restriction digest DNA was treated with pro-teinase K and was phenol-chloroform extracted. Wesynthesized cRNA probes using a RNA transcription
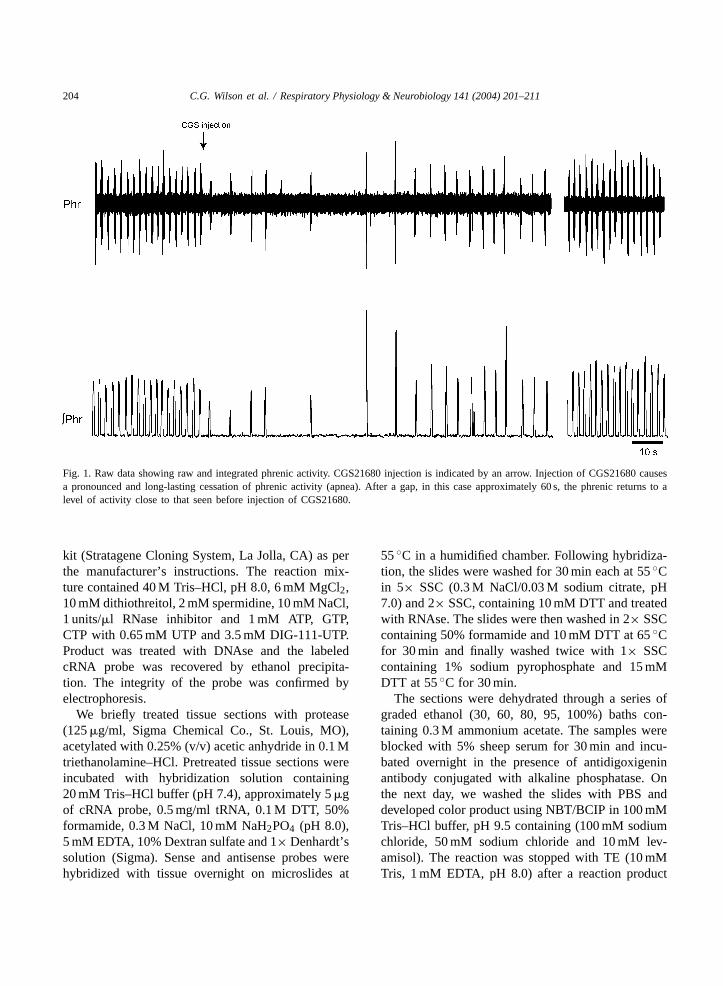
204 C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211
Fig. 1. Raw data showing raw and integrated phrenic activity. CGS21680 injection is indicated by an arrow. Injection of CGS21680 causesa pronounced and long-lasting cessation of phrenic activity (apnea). After a gap, in this case approximately 60 s, the phrenic returns to alevel of activity close to that seen before injection of CGS21680.
kit (Stratagene Cloning System, La Jolla, CA) as perthe manufacturer’s instructions. The reaction mix-ture contained 40 M Tris–HCl, pH 8.0, 6 mM MgCl2,10 mM dithiothreitol, 2 mM spermidine, 10 mM NaCl,1 units/�l RNase inhibitor and 1 mM ATP, GTP,CTP with 0.65 mM UTP and 3.5 mM DIG-111-UTP.Product was treated with DNAse and the labeledcRNA probe was recovered by ethanol precipita-tion. The integrity of the probe was confirmed byelectrophoresis.
We briefly treated tissue sections with protease(125�g/ml, Sigma Chemical Co., St. Louis, MO),acetylated with 0.25% (v/v) acetic anhydride in 0.1 Mtriethanolamine–HCl. Pretreated tissue sections wereincubated with hybridization solution containing20 mM Tris–HCl buffer (pH 7.4), approximately 5�gof cRNA probe, 0.5 mg/ml tRNA, 0.1 M DTT, 50%formamide, 0.3 M NaCl, 10 mM NaH2PO4 (pH 8.0),5 mM EDTA, 10% Dextran sulfate and 1× Denhardt’ssolution (Sigma). Sense and antisense probes werehybridized with tissue overnight on microslides at
55◦C in a humidified chamber. Following hybridiza-tion, the slides were washed for 30 min each at 55◦Cin 5× SSC (0.3 M NaCl/0.03 M sodium citrate, pH7.0) and 2× SSC, containing 10 mM DTT and treatedwith RNAse. The slides were then washed in 2× SSCcontaining 50% formamide and 10 mM DTT at 65◦Cfor 30 min and finally washed twice with 1× SSCcontaining 1% sodium pyrophosphate and 15 mMDTT at 55◦C for 30 min.
The sections were dehydrated through a series ofgraded ethanol (30, 60, 80, 95, 100%) baths con-taining 0.3 M ammonium acetate. The samples wereblocked with 5% sheep serum for 30 min and incu-bated overnight in the presence of antidigoxigeninantibody conjugated with alkaline phosphatase. Onthe next day, we washed the slides with PBS anddeveloped color product using NBT/BCIP in 100 mMTris–HCl buffer, pH 9.5 containing (100 mM sodiumchloride, 50 mM sodium chloride and 10 mM lev-amisol). The reaction was stopped with TE (10 mMTris, 1 mM EDTA, pH 8.0) after a reaction product
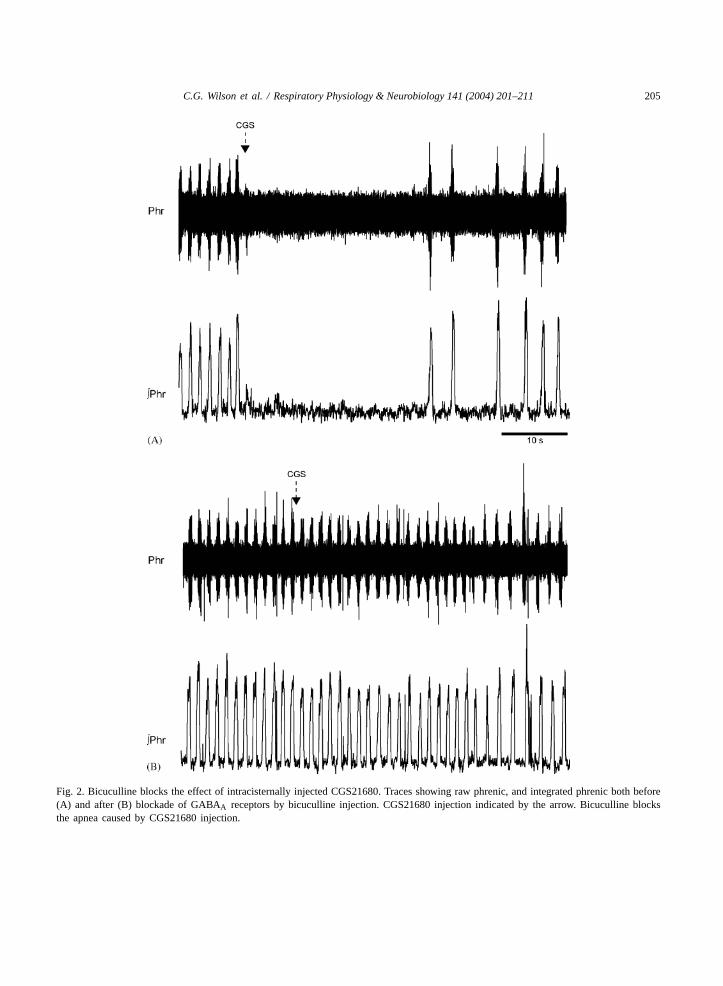
C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211 205
Fig. 2. Bicuculline blocks the effect of intracisternally injected CGS21680. Traces showing raw phrenic, and integrated phrenic both before(A) and after (B) blockade of GABAA receptors by bicuculline injection. CGS21680 injection indicated by the arrow. Bicuculline blocksthe apnea caused by CGS21680 injection.
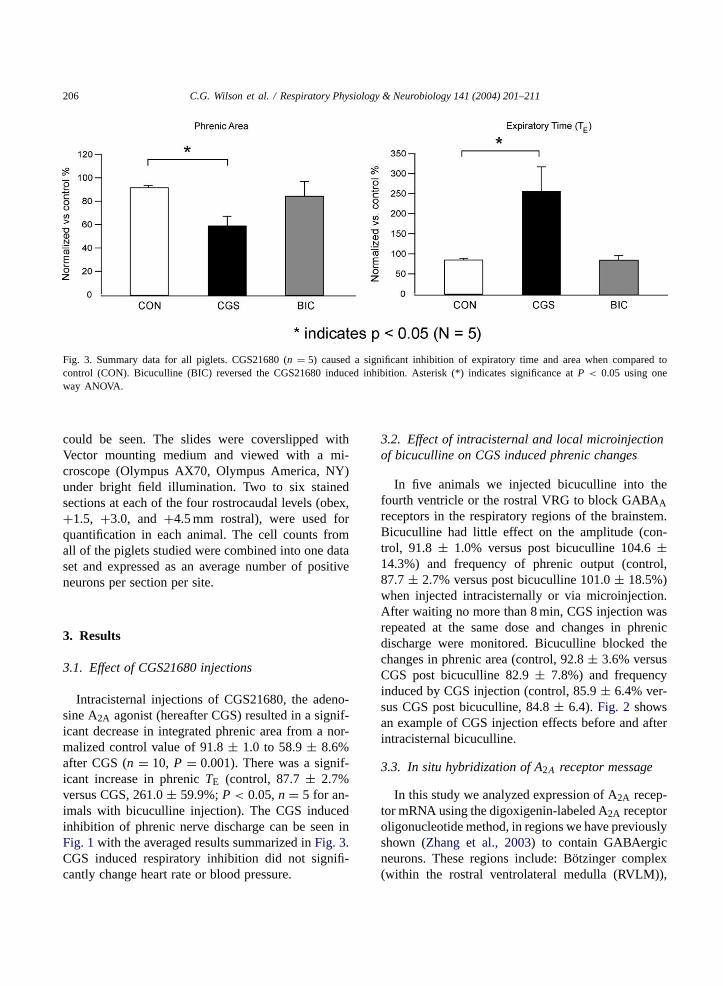
206 C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211
Fig. 3. Summary data for all piglets. CGS21680 (n = 5) caused a significant inhibition of expiratory time and area when compared tocontrol (CON). Bicuculline (BIC) reversed the CGS21680 induced inhibition. Asterisk (*) indicates significance atP < 0.05 using oneway ANOVA.
could be seen. The slides were coverslipped withVector mounting medium and viewed with a mi-croscope (Olympus AX70, Olympus America, NY)under bright field illumination. Two to six stainedsections at each of the four rostrocaudal levels (obex,+1.5, +3.0, and+4.5 mm rostral), were used forquantification in each animal. The cell counts fromall of the piglets studied were combined into one dataset and expressed as an average number of positiveneurons per section per site.
3. Results
3.1. Effect of CGS21680 injections
Intracisternal injections of CGS21680, the adeno-sine A2A agonist (hereafter CGS) resulted in a signif-icant decrease in integrated phrenic area from a nor-malized control value of 91.8± 1.0 to 58.9± 8.6%after CGS (n = 10, P = 0.001). There was a signif-icant increase in phrenicTE (control, 87.7± 2.7%versus CGS, 261.0± 59.9%;P < 0.05,n = 5 for an-imals with bicuculline injection). The CGS inducedinhibition of phrenic nerve discharge can be seen inFig. 1with the averaged results summarized inFig. 3.CGS induced respiratory inhibition did not signifi-cantly change heart rate or blood pressure.
3.2. Effect of intracisternal and local microinjectionof bicuculline on CGS induced phrenic changes
In five animals we injected bicuculline into thefourth ventricle or the rostral VRG to block GABAAreceptors in the respiratory regions of the brainstem.Bicuculline had little effect on the amplitude (con-trol, 91.8 ± 1.0% versus post bicuculline 104.6±14.3%) and frequency of phrenic output (control,87.7± 2.7% versus post bicuculline 101.0± 18.5%)when injected intracisternally or via microinjection.After waiting no more than 8 min, CGS injection wasrepeated at the same dose and changes in phrenicdischarge were monitored. Bicuculline blocked thechanges in phrenic area (control, 92.8± 3.6% versusCGS post bicuculline 82.9± 7.8%) and frequencyinduced by CGS injection (control, 85.9± 6.4% ver-sus CGS post bicuculline, 84.8± 6.4). Fig. 2 showsan example of CGS injection effects before and afterintracisternal bicuculline.
3.3. In situ hybridization of A2A receptor message
In this study we analyzed expression of A2A recep-tor mRNA using the digoxigenin-labeled A2A receptoroligonucleotide method, in regions we have previouslyshown (Zhang et al., 2003) to contain GABAergicneurons. These regions include: Bötzinger complex(within the rostral ventrolateral medulla (RVLM)),
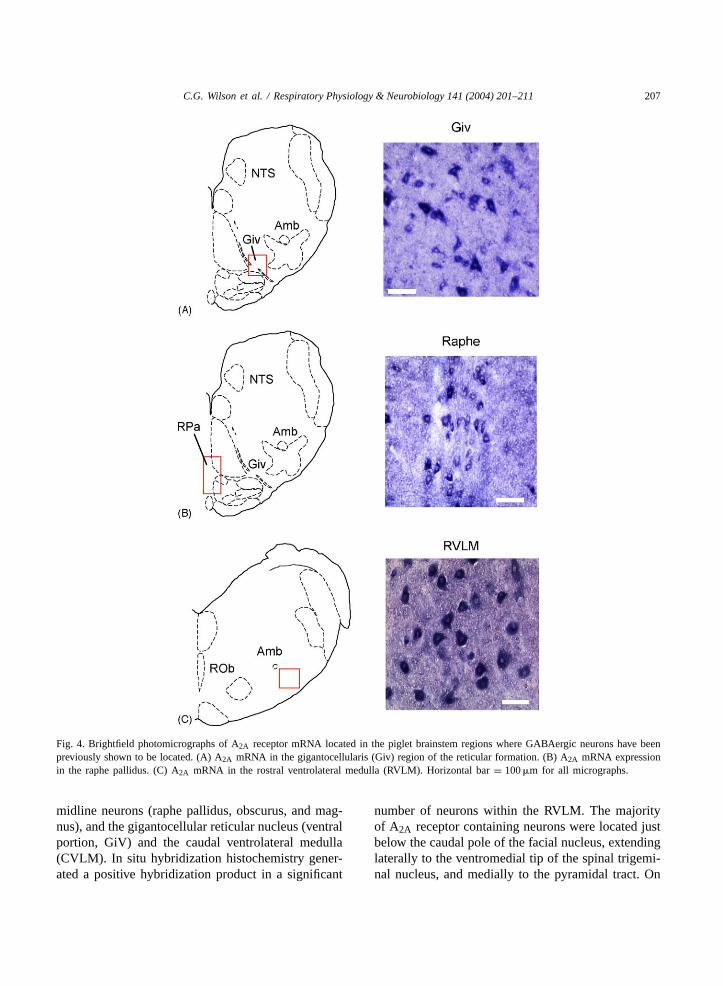
C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211 207
Fig. 4. Brightfield photomicrographs of A2A receptor mRNA located in the piglet brainstem regions where GABAergic neurons have beenpreviously shown to be located. (A) A2A mRNA in the gigantocellularis (Giv) region of the reticular formation. (B) A2A mRNA expressionin the raphe pallidus. (C) A2A mRNA in the rostral ventrolateral medulla (RVLM). Horizontal bar= 100�m for all micrographs.
midline neurons (raphe pallidus, obscurus, and mag-nus), and the gigantocellular reticular nucleus (ventralportion, GiV) and the caudal ventrolateral medulla(CVLM). In situ hybridization histochemistry gener-ated a positive hybridization product in a significant
number of neurons within the RVLM. The majorityof A2A receptor containing neurons were located justbelow the caudal pole of the facial nucleus, extendinglaterally to the ventromedial tip of the spinal trigemi-nal nucleus, and medially to the pyramidal tract. On

208 C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211
average, we counted 110 A2A receptor containingneurons per section per site within the RVLM region.Furthermore, individual as well as clusters of A2Areceptor containing neurons were observed in caudalregions of the ventrolateral medulla and within thelateral reticular nucleus, including the parvicellularsubnucleus. The number of cells containing A2A re-ceptor traits was slightly lower in the CVLM than inthe RVLM (77 neurons per section per site). A largenumber of the A2A receptor expressing neurons (83cells per section per site) was also located in the ven-tral reticular nucleus (GiV). The midline raphe nucleiof the medulla oblongata (pallidus, obscurus, andmagnus) contained a lower number of A2A receptorcontaining neurons than any of the other analyzedsites (16 neurons per site per section). Examples areshown inFig. 4.
4. Discussion
Our major findings indicate that: (1) the adeno-sine A2A agonist CGS21680 inhibits inspiratory drivewhen injected centrally, (2) blockade of GABAA re-ceptors with bicuculline abolishes the inhibitory effectof this adenosine A2A agonist, and (3) adenosine A2Areceptor mRNA is expressed by neurons located in re-gions known to contain GABA producing cells. Thesedata support our hypothesis that adenosine interactswith GABAergic pathways to inhibit respiratory driveand show, for the first time, that adenosine interactsdirectly with GABAergic pathways in modulating res-piratory neural inputs.
Adenosine is a product of nucleotides such as ATPand is formed as a consequence of metabolic and neu-ral activity at many sites including the brain, espe-cially during hypoxia (Moss et al., 1987). Adenosineacts via four known major receptor subtypes A1, A2A,A2B and A3 (Cunha, 2001; Ribeiro et al., 2002; Ochiet al., 2000). Although adenosine is well characterizedas an inhibitory neurotransmitter in the CNS (Cunha,2001; Ribeiro et al., 2002), excitatory actions have alsobeen observed and the net effect on neuronal activitydepends on a balance in activation of A1 and A2 re-ceptors. Inhibitory actions of adenosine are commonlymediated by the A1 receptor which reduces cellularcAMP levels (Ralevic and Burnstock, 1998) and ithas been presumed that the stimulatory effects of caf-
feine are secondary to A1 receptor blockade (Herleniuset al., 1997; Ralevic and Burnstock, 1998). In contrast,A2 receptors are known to mediate excitatory effectsin central neurons via P type calcium channel activa-tion in a cAMP and PKC-dependent manner (Cunha,2001).
Although the distribution of the A2A receptor sub-type was thought to be predominantly in the striatalareas of the brain, newer molecular techniques havedemonstrated a more widespread distribution of A2Areceptor mRNA in mature and developing rat brain(Dixon et al., 1996; Weaver, 1993). In this study, weshow for the first time the presence of A2A receptormRNA expression at a single cell level within regionsof the medulla oblongata that contain GABAergicneurons in piglets (Zaidi et al., 2002; Zhang et al.,2003) and GABA synthesizing neurons in adultrats (Ellenberger, 1999). We assume that inhibitoryinputs elicited by CGS administration that inducemainly prolongation of expiratory duration arisefrom GABAergic neurons in the caudal ventrolateralmedulla. It has been shown that these neurons containGABAergic neurons (Blessing, 1990) that project tothe rostral ventrolateral medulla (Stocker et al., 1997),and when stimulated produce prolongation of expira-tory duration and apnea (Tolentino-Silva et al., 2000).However, CGS-induced decreases in peak phrenic ac-tivity could be mediated via activation of GABAergicneurons within the Bötzinger region. The Bötzingerneurons synthesize GABA (Livingston and Berger,1989) which monosynaptically inhibits phrenic mo-toneurons (Merrill and Fedorko, 1984) as well asventral and dorsal bulbospinal inspiratory neurons(Fedorko et al., 1989). Previous findings also showedthat GABA terminal varicosities innervate phrenicmotoneurons in the spinal cord (Ellenberger, 1999).
Earlier studies have implicated adenosine in theregulation of breathing in late fetal or early neonatallife. Adenosine is released in response to hypoxia inpiglets (Yan et al., 1995) and respiratory inhibitioncan be induced by administration of adenosine ana-logues to fetal sheep (Bissonnette et al., 1991; Yanet al., 1995). Furthermore, adenosine A1 antago-nists diminished the respiratory depression inducedby hypoxia in piglets (Darnall, 1985; Long andLawson, 1984) and rabbit pups (Runold et al., 1989).Runold et al. (1986)also observed that the adenosineanalog, N6-l-(R-phenylisopropyl) adenosine, given

C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211 209
Fig. 5. Proposed model for GABA/Adenosine (A2A) interactions. We propose that centrally released adenosine excites A2A receptorssituated on GABA containing neurons. The resulting GABA release inhibits key inspiratory related sites that regulate respiratory neuraloutput. This simplified scheme is not intended to summarize all known connections and neruron types but rather to summarize our findingsfrom this study.
intraperitoneally elicited greater ventilatory inhibitionwhen administered earlier rather than later in the firstweek of life. Studies employing a rat in vitro brain-stem/spinal cord preparation have shown the greatestrespiratory inhibition after an A1 receptor agonistin the first hours after birth and a higher respiratoryrate after fetal caffeine exposure, neither of whichcould be attributed to change in A1 receptor numberor distribution (Herlenius et al., 2002). Furthermore,Herlenius et al. (2002)has shown that adenosine af-fects respiratory drive by activation of A1 receptors,in the late fetal and very early post-natal period,causing a decrease in cell excitability through a (pre-sumed) direct effect on the brainstem neural networkresponsible for the generation of inspiratory drive andregulation of respiratory timing.
Recent physiologic studies have focused on spe-cific A1 and A2A receptor agonists and have shownthat these adenosine receptors mediate inhibition offetal respiratory drive (Koos et al., 2001; Watsonet al., 1999; Herlenius et al., 2002). In our exper-iments, activation of brainstem adenosine A2A re-ceptors induced prolongation of expiratory durationand inspiratory depression that was blocked by thepresence of GABAA receptor antagonist. Therefore,our data suggest that activation of A2A receptors pro-duces release of GABA within the inspiratory-relatedmedullary network, which in turn, via activation ofGABAA receptors, prolongs expiratory time (TE) andinhibition of inspiratory drive. Hence, adenosine act-
ing via A2A receptors on GABA producing cells maycause depression of breathing and apneic episodes.Future studies might consider whether GABAergicmechanisms also contribute to responses elicited byA1 receptor activation.
GABA is the major inhibitory neurotransmitter inthe adult mammalian CNS, and mediates its effectsthrough both fast and slow synaptic transmission.Fast, ionotropic inhibition occurs through activationof the GABAA receptor and passage of chloride ions(Greengard, 2001). While our data clearly show thatbicuculline either completely abolished or dramati-cally attenuated the expiratory prolongation inducedby activation of A2A receptors, we acknowledge thatvarying agonist/antagonist doses have the potentialto influence our physiologic observations. Futher-more, we recognize that other important neuroin-hibitory pathways (such as those containing glycineor enkephalins) may contribute to inhibition of res-piratory drive (Russier et al., 2002; Grunstein andGrunstein, 1982). It is also possible that GABABreceptors may be involved in inhibition of phrenicdischarge, however, involvement of this mechanismwas not tested in this series of experiments due to theefficacy of GABAA blockade in eliminating apneas.
Fig. 5 provides a summary model of how we thinkadenosine is interacting with the GABAergic pro-jections to respiratory rhythm generating and patternformation components of the brainstem breathingcircuitry. Since we found A2A receptor message in re-

210 C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211
gions known to contain GABAergic neurons, as wellas within the pre-Bötzinger complex and the ventralbrainstem, it is likely that adenosine, apart from itsactions on A1 receptors, specifically activates the A2Aadenosine receptors on inhibitory interneurons, andthrough GABAA receptor dependent signaling path-ways affects phrenic discharge and duration ofTE.
We have demonstrated that adenosine, specificallythe A2A adenosine receptor, has a prominent role in theinhibition of phrenic (inspiratory) drive. This parallelsclinical evidence that adenosine antagonists, the xan-thine derivatives theophylline and caffeine, decreasethe incidence of apnea of prematurity (Miller et al.,2003). Xanthine therapy has been in widespread usefor over 25 years in preterm neonates (Aranda et al.,1976; Shannon et al., 1975; Uauy et al., 1975). Al-though xanthines presumably stimulate central neuraloutput to respiratory muscles—probably via antago-nism of adenosine receptors, the precise mechanismwhereby apneic episodes are attenuated is unknown(Miller et al., 2003). Our results indicate that xanthinetherapy may be acting by blockade of the A2A recep-tor on inhibitory interneurons within the respiratoryrhythm generating circuitry of the brainstem. The endresult is an enhancement of inspiratory drive and mod-ulation of the interplay between excitation and inhi-bition that is crucial for the maintenance of normalbreathing in the early postnatal period.
Acknowledgements
The authors would like to thank Drs. M. Miller andT.E. Dick for critical comments and stimulating dis-cussion during the course of this work. This work wassupported by the National Institutes of Health (HL62527 and IU 54 NS 39407).
References
Abu-Shaweesh, J.M., Dreshaj, I.A., Haxhiu, M.A., Martin, R.J.,2001. Central GABAergic mechanisms are involved in apneainduced by SLN stimulation in piglets. J. Appl. Physiol. 90,1570–1576.
Aranda, J.V., Sitar, D.S., Parsons, W.D., Loughnan, P.M., Neims,A.H., 1976. Pharmacokinetic aspects of theophylline inpremature newborns. N. Engl. J. Med. 295, 413–416.
Bissonnette, J.M., Hohimer, A.R., Knopp, S.J., 1991. The effect ofcentrally administered adenosine on fetal breathing movements.Respir. Physiol. 84, 273–285.
Blessing, W.W., 1990. Distribution of glutamate decarboxylase-containing neurons in rabbit medulla oblongata with attentionto intramedullary and spinal projections. Neuroscience 37,171–185.
Brady, R., Zaidi, S.I., Mayer, C., Katz, D.M., 1999. BDNF isa target-derived survival factor for arterial baroreceptor andchemoafferent primary sensory neurons. J. Neurosci. 19, 2131–2142.
Cunha, R.A., 2001. Adenosine as a neuromodulator and as ahomeostatic regulator in the nervous system: different roles,different sources and different receptors. Neurochem. Int. 38,107–125.
Darnall, R.A., 1985. Aminophylline reduces hypoxic ventilatorydepression: possible role of adenosine. Pediatr. Res. 19, 706–710.
Dixon, A.K., Gubitz, A.K., Sirinathsinghji, D.J.S., Richardson, P.J.,Freeman, T.C., 1996. Tissue distribution of adenosine receptormRNAs in the rat. Br. J. Pharmacol. 118, 1461–1468.
Dreshaj, I.A., Haxhiu, M.A., Abu-Shaweesh, J., Carey, R.E.,Martin, R.J., 1999. CO2-induced prolongation of expiratorytime during early development. Respir. Physiol. 116, 125–132.
Ellenberger, H.H., 1999. Distribution of bulbospinalgamma-aminobutyric acid-synthesizing neurons of the ventralrespiratory group of the rat. J. Comp. Neurol. 411, 130–144.
Fedorko, L., Duffin, J., England, S., 1989. Inhibition of inspiratoryneurons of the nucleus retroambigualis by expiratory neuronsof the Bötzinger complex in the cat. Exp. Neurol. 106, 74–77.
Gauda, E.B., Northington, F.J., Linden, J., Rosin, D.L.,2000. Differential expression of a(2a), A(1)-adenosine andD(2)-dopamine receptor genes in rat peripheral arterialchemoreceptors during postnatal development. Brain Res.872 (1/2), 1–10.
Greengard, P., 2001. The neurobiology of slow synaptictransmission. Science 294, 1024–1030.
Grunstein, M.M., Grunstein, J.S., 1982. Maturational effects ofenkephalin on respiratory control in newborn rabbits. J. Appl.Physiol. 53, 1063–1070.
Herlenius, E., Lagercrantz, H., Yamamoto, Y., 1997. Adenosinemodulates inspiratory neurons and the respiratory pattern inthe brainstem of neonatal rats. Pediatr. Res. 42, 46–53.
Herlenius, E., Aden, U., Tang, L.Q., Lagercrantz, H., 2002.Perinatal respiratory control and its modulation by adenosineand caffeine in the rat. Pediatr. Res. 51, 4–12.
Koos, B.J., Maeda, T., Jan, C., 2001. Adenosine A1 and A2A
receptors modulate sleep state and breathing in fetal sheep. J.Appl. Physiol. 91, 343–350.
Koos, B.J., Maeda, T., Jan, C., Lopez, G., 2002. Adenosine A(2A)receptors mediate hypoxic inhibition of fetal breathing in sheep.Am. J. Obstet. Gynecol. 186 (4), 663–668.
Livingston, C.A., Berger, A.J., 1989. Immunocytochemicallocalization of GABA in neurons projecting to the ventrolateralnucleus of the solitary tract. Brain Res. 494, 143–150.
Long, W.A., Lawson, E.E., 1984. Neurotransmitters and biphasicrespiratory response to hypoxia. J. Appl. Physiol. 57, 213–222.

C.G. Wilson et al. / Respiratory Physiology & Neurobiology 141 (2004) 201–211 211
Merrill, E.G., Fedorko, L., 1984. Monosynaptic inhibition ofphrenic motoneurons: a long descending projection fromBötzinger neurons. J. Neurosci. 4, 2350–2353.
Miller, M.J., Haxhiu, M.A., Haxhiu-Poskurica, B., Dreshaj, I.A.,DiFiore, J.M., Martin, R.J., 2000. Recurrent hypoxic exposureand reflex responses during development in the piglet. Respir.Physiol. 123, 51–61.
Miller, M.J., Haxhiu, M.A., Martin, R.J., 2003. Chemical controlof breathing from the fetus through the newborn period. In:Matthew O. (Ed.), Lung Biology in Health and Disease:Respiratory Control and its Disorders in the Newborn. MarcelDekker, NY, pp. 83–103.
Moss, I.R., Runold, M., Dahlin, I., Fredholm, B.B., Nyberg,F., Lagercrantz, H., 1987. Respiratory and neuroendocrineresponses of piglets to hypoxia during postnatal development.Acta Physiol. Scand. 131, 533–541.
Ochi, M., Koga, K., Kurokawa, M., Kase, H., Nakamura, J.,Kuwana, Y., 2000. Systemic administration of adenosine A2A
receptor antagonist reverses increased GABA release in theglobus pallidus of unilateral 6-hydroxydopamine-lesioned rats:a microdialysis study. Neuroscience 100, 53–62.
Ralevic, V., Burnstock, G., 1998. Receptors for purines andpyrimidines. Pharmacol. Rev. 50, 413–492.
Ribeiro, J.A., Sebastiao, A.M., de Mendonca, A., 2002.Adenosine receptors in the nervous system: pathophysiologicalimplications. Prog. Neurobiol. 68 (6), 377–392.
Runold, M., Lagercrantz, H., Fredholm, B.B., 1986. Ventilatoryeffect of an adenosine analogue in unanesthetized rabbits duringdevelopment. J. Appl. Physiol. 61, 255–258.
Runold, M., Lagercrantz, H., Prabhakar, N.R., Fredholm, B.B.,1989. Role of adenosine in hypoxic ventilatory depression. J.Appl. Physiol. 67, 541–546.
Russier, M., Kopysova, I.L., Ankri, N., Ferrand, N., Debanne,D., 2002. GABA and glycine co-release optimizes functional
inhibition in rat brainstem motoneurons in vitro. J. Physiol.Lond. 541.1, 123–137.
Shannon, D.C., Gotay, F., Stein, I.M., Rogers, M.C., Todres, I.D.,Moylan, F.M.B., 1975. Prevention of apnea and bradycardiain low-birth weight infants. Pediatrics 55, 589.
Stocker, S.D., Steinbacher, B.C., Jr, C.D., Yates, B.J., 1997.Connections of the caudal ventrolateral medullary reticularformation in the cat brainstem. Exp. Brain Res. 116, 270–282.
Tolentino-Silva, F.P., Haxhiu, M.A., Ernsberger, P., Waldbaum,S., Dreshaj, I.A., 2000. Differential cardiorespiratory controlelicited by activation of ventral medullary sites in mice. J.Appl. Physiol. 89, 437–444.
Uauy, R., Shapiro, D.L., Smith, B., Warshaw, J.B., 1975. Treatmentof severe apnea in prematures with orally administeredtheophylline. Pediatrics 55, 595–598.
Watson, C.S., White, S.E., Homan, J.H., Fraher, L., Brien, J.F.,Bocking, A.D., 1999. The adenosine A1-receptor antagonist8-CPT reverses ethanol-induced inhibition of fetal breathingmovements. J. Appl. Physiol. 87, 1333–1338.
Weaver, D.R., 1993. A2A adenosine receptor gene expression indeveloping rat brain. Mol. Brain Res. 20, 313–327.
Yan, S., Lafferriere, A., Zhang, C., Moss, M.R., 1995.Microdialyzed adenosine in nucleus tractus solitarii andventilatory response to hypoxia in piglets. J. Appl. Physiol.79, 405–410.
Zaidi, S.I., Jafri, A., Martin, R.J., Gauda, E., Johnson, S.,Haxhiu, M.A., 2002. Age-related changes of A2A receptorexpression within brain stem regions involved in respiratorycontrol. Program #530.13, 2002 Abstract Viewer. Society forNeuroscience, Washington, DC, Online.
Zhang, L., Wilson, C.G., Liu, S., Haxhiu, M.A., Martin, R.J.,2003. Hypercapnia-induced activation of brainstem GABAergicneurons during early development. Respir. Physiol. Neurobiol.136 (1), 25–37.