hippocampal low-frequency stimulation and chronic mild stress similarly disrupt fear extinction...
TRANSCRIPT

Available online at www.sciencedirect.com
www.elsevier.com/locate/ynlme
Neurobiology of Learning and Memory 89 (2008) 560–566
Hippocampal low-frequency stimulation and chronic mildstress similarly disrupt fear extinction memory in rats
Rene Garcia a,*, Guillaume Spennato a, Linda Nilsson-Todd a,Jean-Luc Moreau b, Olivier Deschaux a
a Laboratoire de Neurobiologie et Psychopathologie, JE2441, Universite de Nice-Sophia Antipolis, Nice, Franceb Pharma Division, Hoffmann-La Roche, Basel, Switzerland
Received 30 August 2007; revised 12 October 2007; accepted 15 October 2007Available online 26 November 2007
Abstract
Disruptions of fear extinction-related potentiation of synaptic efficacy in the connection between the hippocampus (HPC) and themedial prefrontal cortex (mPFC) have been shown to impair the recall of extinction memory. This study was undertaken to examineif chronic mild stress (CMS), which is known to alter induction of HPC–mPFC long-term potentiation, would also interfere with bothextinction-related HPC–mPFC potentiation and extinction memory. Following fear conditioning (5 tone-shock pairings), rats were sub-mitted to fear extinction (20 tone-alone presentations), which produced an increase in the amplitude of HPC–mPFC field potentials.HPC low-frequency stimulation (LFS), applied immediately after training, suppressed these changes and induced fear return duringthe retention test (5 tone-alone presentations). CMS, delivered before fear conditioning, did not interfere with fear extinction but blockedthe development of extinction-related potentiation in the HPC–mPFC pathway and impaired the recall of extinction. These findings sug-gest that HPC LFS may provoke metaplastic changes in HPC outputs that may mimic alterations associated with a history of chronicstress.� 2007 Elsevier Inc. All rights reserved.
Keywords: Extinction of conditioned freezing; Chronic stress; Hippocampus; Medial prefrontal cortex; Synaptic plasticity
1. Introduction
Following fear conditioning, in which a neutral stimu-lus such as non-aversive tone (conditioned stimulus: CS)has been paired with an aversive stimulus such as electri-cal shock (unconditioned stimulus: US), re-exposure tothe CS without the US can provoke aversive responsessuch as defensive freezing behavior (conditionedresponse: CR) (Blanchard & Blanchard, 1969). However,repeated presentations of the CS alone cause a gradualattenuation of the CR, a phenomenon called extinction(Pavlov, 1927). Extinction is not considered to be a lossof the initial ‘CS–US’ memory, but rather seems to be anew learning process based on a ‘CS–no US’ association.
1074-7427/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.nlm.2007.10.005
* Corresponding author. Fax: +33 492 07 61 62.E-mail address: [email protected] (R. Garcia).
Therefore, following fear extinction, two CS-relatedmemories (‘CS–US’ and ‘CS–no US’) co-exist and com-pete upon each presentation of the CS (e.g., Garcia,2002; Myers & Davis, 2007).
Given that symptoms of fear-related disorders, such aspost-traumatic stress disorder, can be eliminated viaexposure therapy, a clinical form of fear extinction, onecan suggest that impairment of consolidation or retrievalof the ‘CS–no US’ memory may be responsible forrelapses observed during follow-up tests (Tarrier, Som-merfield, Pilgrim, & Humphreys, 1999). This type of dys-function is believed to involve networks including thehippocampus (HPC), the medial prefrontal cortex(mPFC) and the amygdala (e.g., Bouton, Westbrook,Corcoran, & Maren, 2006; Garcia, 2002; Ji & Maren,2007; Myers & Davis, 2007; Quirk, Garcia, & Gonz-alez-Lima, 2006; Quirk & Mueller, 2007). This view is,

R. Garcia et al. / Neurobiology of Learning and Memory 89 (2008) 560–566 561
in part, supported by various recent studies. For exam-ple, hippocampal dysfunction during or after extinctiontraining has been shown to disrupt the context-dependentexpression of the ‘CS–no US’ memory (Corcoran, Des-mond, Frey, & Maren, 2005; Corcoran & Maren, 2001,2004; Ji & Maren, 2005). It has been demonstrated thatthe hippocampus plays a crucial role in the recall of fearextinction through its direct and indirect (via the mPFC)influences on the amygdala (Bruchey, Shumake, & Gonz-alez-Lima, 2007). Lesions of the mPFC (Lebron, Milad,& Quirk, 2004; Quirk, Russo, Barron, & Lebron, 2000)or pharmacological impairment of mPFC function(Hugues, Chessel, Lena, Marsault, & Garcia, 2006;Hugues, Deschaux, & Garcia, 2004; Santini, Ge, Ren,Pena de Ortiz, & Quirk, 2004; Santini, Muller, & Quirk,2001) have been reported to impair the recall of extinc-tion memory. Electrophysiological findings have revealedthat fear extinction is accompanied by increases of syn-aptic efficacy in the HPC projections to the mPFC (Far-inelli, Deschaux, Hugues, Thevenet, & Garcia, 2006;Hugues & Garcia, 2007; Hugues et al., 2006). Interest-ingly, artificial disruption of these synaptic changes byHPC low-frequency stimulation (LFS) also impairsretrieval of the ‘CS–no US’ memory (Farinelli et al.,2006; Hugues & Garcia, 2007).
Given that stressful conditions such as chronic mildstress (CMS) blocks the induction of long-term potentia-tion (LTP) in the HPC–mPFC pathway (Cerqueira, Mail-liet, Almeida, Jay, & Sousa, 2007) and that a history ofchronic stress is known to interfere with extinction memory(Miracle, Brace, Huyck, Singler, & Wellman, 2006), thepresent study examined whether CMS would also alterdevelopment of extinction-associated changes in synaptictransmission in the HPC–mPFC pathway and if such syn-aptic changes would be associated with impairment of therecall of extinction.
2. Materials and methods
2.1. Subjects
Twenty-five male albino Wistar rats (Biological Research Institute,Fullinsdorf, Switzerland) weighing approximately 280–300 g were used.They were housed individually and had free access to food and water,except when temporarily group housed or deprived of food and water aspart of the CMS protocol. Experiments were conducted in accordancewith the European Community Guidelines on the care and use of labora-tory animals (86/609/EEC) and the Swiss Federal Regulations on AnimalProtection.
Rats were divided into two initial groups, one submitted to CMS(n = 10) for 3 weeks and the other group (n = 15) submitted to a 3-weekperiod of handling (Fig. 1A). The CMS procedure has been described indetail elsewhere (Moreau, Bourson, Jenck, Martin, & Mortas, 1994).Briefly, the stress regimen was applied each week with a variety of unpre-dictable, mild stressors (periods of confinement to small cages, continuousovernight illumination, deprivation of food and water, and group housingin a soiled cage). Rats exposed to such a stress regimen are known todevelop an anhedonic state as revealed by a decreased intake/preferencefor sweet solutions or a decreased sensitivity to rewarding electrical brainstimulation (Moreau et al., 1994).
2.2. Surgery
At the end of the 3-week period, all rats (with or without CMS proce-dure) were anesthetized with sodium pentobarbital (65 mg/kg, ip), placedinto a stereotaxic frame, and unilaterally implanted (right side) with elec-trodes made of a pair of twisted silver wires (90 lm diameter; insulatedexcept at section) for field potential recording. Recording electrodes tar-geted the mPFC (2.7–3.2 mm anterior to bregma, 0.4–0.8 mm lateral tomidline, and 3.2 mm from dura) and stimulating electrodes targeted theHPC area (5.8–6.3 mm posterior to bregma, 5.2–6.0 mm lateral to midline,and 3.9–4.2 mm from dura). One of three screws on the skull served as therecording ground. In addition to electrode placement in the brain, two sil-ver wires (110 lm diameter, insulated except at the section: 0.4–0.6 mmexposed) were inserted in the right eyelid for shock delivery during fearconditioning. The entire miniature system was fixed in place on the skullwith dental cement.
2.3. Stimulating and recording procedures
Field potentials evoked in the mPFC by single-pulse stimulation (0.1-ms rectangular monophasic pulses) applied to the HPC were sent to anamplifier (gain 1000·; bandpass 0.001–1 kHz) and recorded (Spike2 soft-ware; Cambridge Electronic Design, Cambridge, UK) for off-line analy-sis. Prior to the first test recording, responses were measured as afunction of stimulus strength (input–output curves: 100–800 lA). Anintensity corresponding to 60–70% of the saturation level was chosenfor the test stimulus, which was applied every 5 s during each recordingsession.
2.4. Experimental protocol
After three days of recovery from surgery, rats were habituated overa 2-day period to being transported (from the animal house to the exper-imental room) and connected to electrophysiological cables (relayed atthe top of the experimental box by a multi-channel rotating connector,allowing free movement). Following habituation, electrophysiologicaland behavioral experiments started. Two sessions of baseline recordingsduring 2 consecutive days (7 field potentials/session/day) took place inbox A (consisting of four vertical sides, each 30 cm wide), which wascleaned with ethanol (50%) before introducing each animal. For fearconditioning, 30 min following the second baseline recording session,each rat was placed in a cylindrical cage, box B (25 cm diameter, floorcovered with animal bedding that was changed before introducing eachanimal), which was cleaned with a solution of ethanol (50%) and lemonscent. Boxes A and B were placed inside a sound-attenuating and tem-perature-regulated chamber (Imetronic, Pessac, France) during the exper-iment. Two minutes after being placed in box B, all CMS rats (CMS–FCgroup) and 10 of the unstressed rats were subjected to five tone (a 30-s/2.5–kHz/75-dB)-shock (a train of eight 100-ls pulses/5 Hz/ 3.5 mA) pair-ings (inter-trial interval: 60 s). The 5 remaining unstressed rats were alsoplaced in box B and received 5 tone presentations without shock appli-cation (NFC). For the extinction training the following day, each rat wasplaced in box A, where twenty tone-alone presentations were deliveredwith inter-trial interval of 60 s. After the last tone presentation, 5 ofthe unstressed and shocked rats were immediately submitted to HPCLFS (FC-LFS group), which consisted of a train of 100-ls pulses at2 Hz for 25 min. CMS rats (CMS–FC group), non-shocked rats (NFCgroup) and the 5 other unstressed and shocked rats (FC group) were alsoleft in box A for 25 min, but did not receive LFS. Twenty-four hourslater, each rat was returned to in box A, where five tone-alone presenta-tions were delivered (inter-trial interval: 60 s) for the retention test(Fig. 1A).
Field potentials (7/session) were recorded in box A before extinctiontraining and retention test. The behavior of each rat was continuouslymonitored and recorded via a camera–VCR–monitor system. To assessconditioned fear evoked by each tone presentation, freezing behaviorwas measured with a time-sampling procedure.

Fig. 1. (A) Experimental timeline. CMS: chronic mild stress; Ext. training: extinction training (20 CS–no US presentations); FC: fear conditioning (5 CS–US presentations); FP, field potentials (7 field potentials/recording session); NFC, no fear conditioning (5 CS–no US presentations during the conditioningphase). (B,C) Diagrams showing electrode placements (filled circles) in the CA2 area of the hippocampus for stimulation (B), and the prelimbic area (PrL)of the medial prefrontal cortex for field potential recordings (C).
562 R. Garcia et al. / Neurobiology of Learning and Memory 89 (2008) 560–566
2.5. Histology and data analyses
On completion of the experiments, rats were deeply anesthetized, andthe tips of electrode placements in the HPC and mPFC were marked bypassing 0.3- to 0.5-mA current for 20 s. The placement of the electrodeswas then verified by standard histological methods.
All data were expressed as means and standard error of the means (SE)and analyzed by analysis of variance (ANOVA). Direct between-groupanalyses of both electrophysiological and behavioral data were performedby using ANOVA with Fisher post hoc comparison of means.
3. Results
3.1. Histology
Verification of electrode placements revealed that allrats were implanted with stimulating electrodes in theCA2 region located between the dorsal and ventral HPC(Fig. 1B) and recording electrodes in the prelimbic areaof the mPFC (Fig. 1C).
3.2. Electrophysiology
Field potentials recorded in the mPFC following stim-ulation of the HPC (Fig. 2A) were similar to responses
that have been described previously (Laroche, Davis, &Jay, 2000; Izaki et al., 2001; Hugues & Garcia, 2007;Okulski et al., 2007). Three rats from the CMS–FC groupwere excluded for unstable responses over days. Allremaining rats (n = 22) exhibited stable field potentialresponses across the two days of baseline recordings.Responses remained stable following fear conditioning(Fig. 2B: Pre-Ext recordings). Twenty-four hours follow-ing extinction training, rats in the FC group exhibitedpotentiation of their HPC–mPFC synaptic efficacy,whereas rats that received HPC LFS immediately aftertraining and those in the CMS–FC group did not showthis potentiation. A two-way ANOVA performed on thesedata (4 groups, 4 recording periods) showed no effectof group and no effect of recording session. However,the interaction between group and recording sessionapproached significance (p = 0.05). Post hoc Fisher testsrevealed that fear extinction induced a significant potenti-ation of HPC–mPFC synaptic transmission only in theFC group (Pre-Ext vs. Post-Ext: P < 0.05). Furthermore,rats in the FC group differed from each of the three othergroups during the Post-Ext recording session (FC vs.NFC, FC vs. FC-LFS and FC vs. CMS–FC: allPs < 0.05).
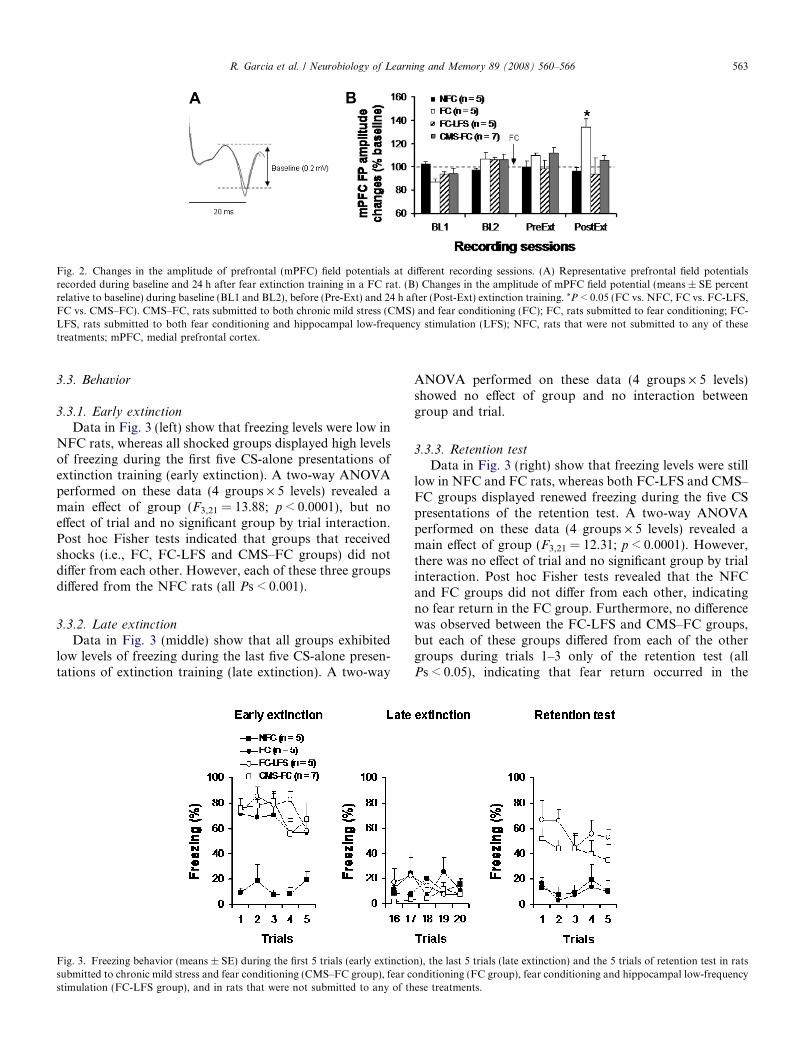
Fig. 2. Changes in the amplitude of prefrontal (mPFC) field potentials at different recording sessions. (A) Representative prefrontal field potentialsrecorded during baseline and 24 h after fear extinction training in a FC rat. (B) Changes in the amplitude of mPFC field potential (means ± SE percentrelative to baseline) during baseline (BL1 and BL2), before (Pre-Ext) and 24 h after (Post-Ext) extinction training. *P < 0.05 (FC vs. NFC, FC vs. FC-LFS,FC vs. CMS–FC). CMS–FC, rats submitted to both chronic mild stress (CMS) and fear conditioning (FC); FC, rats submitted to fear conditioning; FC-LFS, rats submitted to both fear conditioning and hippocampal low-frequency stimulation (LFS); NFC, rats that were not submitted to any of thesetreatments; mPFC, medial prefrontal cortex.
R. Garcia et al. / Neurobiology of Learning and Memory 89 (2008) 560–566 563
3.3. Behavior
3.3.1. Early extinction
Data in Fig. 3 (left) show that freezing levels were low inNFC rats, whereas all shocked groups displayed high levelsof freezing during the first five CS-alone presentations ofextinction training (early extinction). A two-way ANOVAperformed on these data (4 groups · 5 levels) revealed amain effect of group (F3,21 = 13.88; p < 0.0001), but noeffect of trial and no significant group by trial interaction.Post hoc Fisher tests indicated that groups that receivedshocks (i.e., FC, FC-LFS and CMS–FC groups) did notdiffer from each other. However, each of these three groupsdiffered from the NFC rats (all Ps < 0.001).
3.3.2. Late extinction
Data in Fig. 3 (middle) show that all groups exhibitedlow levels of freezing during the last five CS-alone presen-tations of extinction training (late extinction). A two-way
Fig. 3. Freezing behavior (means ± SE) during the first 5 trials (early extinctiosubmitted to chronic mild stress and fear conditioning (CMS–FC group), fear cstimulation (FC-LFS group), and in rats that were not submitted to any of th
ANOVA performed on these data (4 groups · 5 levels)showed no effect of group and no interaction betweengroup and trial.
3.3.3. Retention test
Data in Fig. 3 (right) show that freezing levels were stilllow in NFC and FC rats, whereas both FC-LFS and CMS–FC groups displayed renewed freezing during the five CSpresentations of the retention test. A two-way ANOVAperformed on these data (4 groups · 5 levels) revealed amain effect of group (F3,21 = 12.31; p < 0.0001). However,there was no effect of trial and no significant group by trialinteraction. Post hoc Fisher tests revealed that the NFCand FC groups did not differ from each other, indicatingno fear return in the FC group. Furthermore, no differencewas observed between the FC-LFS and CMS–FC groups,but each of these groups differed from each of the othergroups during trials 1–3 only of the retention test (allPs < 0.05), indicating that fear return occurred in the
n), the last 5 trials (late extinction) and the 5 trials of retention test in ratsonditioning (FC group), fear conditioning and hippocampal low-frequencyese treatments.

564 R. Garcia et al. / Neurobiology of Learning and Memory 89 (2008) 560–566
FC-LFS and CMS–FC groups during these three initialtrials.
4. Discussion
The results of the present study demonstrate that bothLFS applied in the HPC and CMS block the develop-ment of fear extinction-associated prefrontal synapticplasticity, causing the recovery of the extinguished fear.These data support the concept that a disturbance ofthe interaction between the HPC and the mPFC follow-ing fear extinction training interferes with extinctionmemory.
Although our lesion studies do not suggest that a suc-cessful recall of extinction memory depends on the integ-rity of the mPFC (Farinelli et al., 2006; Garcia, Chang,& Maren, 2006), one cannot categorically exclude a keyrole of the mPFC in networks modulating recall ofextinction, as such a role has been suggested by otherlesion studies as well as studies employing otherapproaches, such as electrophysiology and pharmacolog-ical manipulations. For instance, Quirk and colleagueshave shown that mPFC lesions interfere with recall offear extinction (Lebron et al., 2004; Quirk et al., 2000).The same group has also found that successful recallof fear extinction is associated with increased burst firingof mPFC neurons (Burgos-Robles, Vidal-Gonzalez, San-tini, & Quirk, 2007), which may be required for extinc-tion-related neuronal plasticity in the mPFC asobserved with single-unit recordings (Milad & Quirk,2002). Interestingly, pre- or post-extinction trainingblockade of N-methyl-D-aspartate receptors in themPFC, which impairs the recall of extinction (Santiniet al., 2001), also reduces this bursting (Burgos-Robleset al., 2007). The present data, together with our previ-ous findings (Farinelli et al., 2006; Hugues & Garcia,2007; Hugues et al., 2006), support the same idea interms of interactions between the HPC and the mPFC.Indeed, we have previously reported that extinctiontraining produces an increase in field potential amplitudein both dorsal HPC–mPFC (Farinelli et al., 2006) andventral HPC–mPFC (Hugues & Garcia, 2007; Hugueset al., 2006) pathways. In the present study, we againfound that outputs from the area including both the dor-sal and ventral HPC (with stimulating electrodes locatedin the CA2) to the mPFC display a potentiation in theirsynaptic efficacy following extinction training. However,because of (i) the absence of a direct projection fromthe CA2 to the mPFC and (ii) the proximity of theCA1 area, which does project to the mPFC (Jay & Wit-ter, 1991), it is possible that prefrontal field potentialswere evoked by stimulation of the CA1 region ratherthan the CA2 region. Furthermore, immediate post-extinction suppression of the development of the mPFCpotentiation with a pharmacological treatment (Hugueset al., 2004,2006) or LFS applied in the dorsal HPC(Farinelli et al., 2006) or the ventral HPC (Hugues &
Garcia, 2007) impairs the recall of extinction. The pres-ent study replicated this effect as well. Although manip-ulation of the dorsal and ventral HPC outputs to themPFC led to similar observations, it remains to be deter-mined whether there is a dorso–ventral segregation inmodulation of extinction memory processes within theHPC as has been shown for anxiety (Nascimento Hackl& Carobrez, 2007) and learning (Lopez-Fernandez et al.,2007; McDonald, Jones, Richards, & Hong, 2006) tasks.
It should be noted that the HPC projects not only to themPFC, but also to many other brain regions that displaysynaptic changes following HPC train stimulation (e.g.,the amygdala: Maren & Fanselow, 1995; the nucleusaccumbens: Dong, Cao, & Xu, 2007; the thalamus: Hugues& Garcia, 2007). The disruption of extinction recall byHPC LFS could also be provoked by direct changes inthese other HPC outputs rather than HPC–mPFC outputsas we have shown previously in rats with mPFC lesions(Farinelli et al., 2006). Indeed, recent findings also havesuggested that the HPC modulates recall of fear extinctionvia its direct and indirect projections to the amygdala (Bru-chey et al., 2007).
It has been demonstrated in several studies that stressinterferes with mechanisms of synaptic plasticity (Kimand Yoon, 1998; Garcia, 2001; Kim et al., 2006). For exam-ple, recent studies have revealed that both elevated platformstress (Rocher, Spedding, Munoz, & Jay, 2004) and CMS(Cerqueira et al., 2007) inhibit induction of LTP in theHPC–mPFC pathway. This impact of stress on synapticplasticity is known as metaplasticity (Abraham & Bear,1996). Our data also demonstrate that stress, in particularCMS, induced metaplasticity in the HPC–mPFC pathway.However, while there are similarities between our observa-tions and those of other authors Rocher et al., 2004; Cer-queira et al., 2007) regarding the electrophysiologicaleffects of CMS and elevated platform stress on HPC–mPFCsynaptic plasticity, there is a difference between our findingand those of Akirav and Maroun (2007) on the effect ofstress on extinction. That is Akirav and Maroun reportedthat elevated platform stress provokes a resistance to extinc-tion, whereas we did not find any CMS effect on extinctionlearning, but rather observed a CMS effect on the recall ofextinction memory. This behavioral difference may berelated to the duration of stress exposure (acute vs. chronic)rather than the nature of the stress (‘elevated platform’stressor vs. unpredictable various stressors). This hypothe-sis is supported by another recent finding showing thatchronic restraint stress does not affect extinction of condi-tioned fear, but impairs the recall of extinction memory(Miracle et al., 2006). In other words, a history of chronicstress may be one of the causes of relapse following fearextinction-based treatment of fear disorders.
Given that rats exposed to HPC LFS and CMS displayedsimilar electrophysiological and behavioral effects, we sug-gest that HPC LFS may provoke metaplasticity in HPC–mPFC that may mimic alterations associated with a historyof chronic stress. Given that we have also previously

R. Garcia et al. / Neurobiology of Learning and Memory 89 (2008) 560–566 565
reported that HPC tetanus stimulation reverses metaplasticchanges evoked by HPC LFS (Farinelli et al., 2006), we pro-pose that HPC tetanus may also block the effect of CMS.
Acknowledgments
We thank K.M. Myers for critical reading of the manu-script. This study was supported by F. Hoffmann-LaRoche (J.L.M.) and the University of Nice-Sophia Antipo-lis (O.D., G.S., L.N.T., R.G.).
References
Abraham, W. C., & Bear, M. F. (1996). Metaplasticity: the plasticity ofsynaptic plasticity. Trends in Neurosciences, 19, 126–130.
Akirav, I., & Maroun, M. (2007). The role of the medial prefrontal cortex-amygdala circuit in stress effects on the extinction of fear. NeuralPlasticity, Article ID: 30873.
Blanchard, R. J., & Blanchard, C. (1969). Crouching as an index of fear.Journal of Comparative and Physiological Psychology, 6, 370–375.
Burgos-Robles, A., Vidal-Gonzalez, I., Santini, E., & Quirk, G. J.(2007). Consolidation of fear extinction requires NMDA receptor-dependent bursting in the ventromedial prefrontal cortex. Neuron,
53, 871–880.Bouton, M. E., Westbrook, R. F., Corcoran, K. A., & Maren, S. (2006).
Contextual and temporal modulation of extinction: behavioral andbiological mechanisms. Biol Psychiatry, 60, 352–360.
Bruchey, A. K., Shumake, J., & Gonzalez-Lima, F. (2007). Networkmodel of fear extinction and renewal functional pathways. Neurosci-
ence, 145, 423–437.Cerqueira, J. J., Mailliet, F., Almeida, O. F., Jay, T. M., & Sousa, N.
(2007). The prefrontal cortex as a key target of the maladaptiveresponse to stress. Journal of Neuroscience, 27, 2781–2787.
Corcoran, K. A., & Maren, S. (2001). Hippocampal inactivation disruptscontextual retrieval of fear memory after extinction. Journal of
Neuroscience, 21, 1720–1726.Corcoran, K. A., & Maren, S. (2004). Factors regulating the effects of
hippocampal inactivation on renewal of conditional fear after extinc-tion. Learning and Memory, 11, 598–603.
Corcoran, K. A., Desmond, T. J., Frey, K. A., & Maren, S. (2005).Hippocampal inactivation disrupts the acquisition and contextualencoding of fear extinction. Journal of Neuroscience, 25, 8978–8987.
Dong, Z., Cao, J., & Xu, L. (2007). Opiate withdrawal modifies synapticplasticity in subicular-nucleus accumbens pathway in vivo. Neurosci-
ence, 144, 845–854.Farinelli, M., Deschaux, O., Hugues, S., Thevenet, A., & Garcia, R.
(2006). Hippocampal train stimulation modulates recall of fearextinction independently of prefrontal cortex synaptic plasticity andlesions. Learning and Memory, 13, 329–334.
Garcia, R. (2001). Stress, hippocampal plasticity and spatial learning.Synapse, 40, 180–183.
Garcia, R. (2002). Post-extinction of conditioned fear: between two CS-related memories. Learning and Memory, 9, 361–363.
Garcia, R., Chang, C. H., & Maren, S. (2006). Electrolytic lesions of themedial prefrontal cortex do not interfere with long-term memory ofextinction of conditioned fear. Learning and Memory, 13, 14–17.
Hugues, S., & Garcia, R. (2007). Reorganization of learning-associatedprefrontal synaptic plasticity between the recall of recent and remotefear extinction memory. Learning and Memory, 14, 520–524.
Hugues, S., Deschaux, O., & Garcia, R. (2004). Post-extinction infusion ofa mitogen-activated protein kinase inhibitor into the medial prefrontalcortex impairs memory of the extinction of conditioned fear. Learning
and Memory, 11, 540–543.Hugues, S., Chessel, A., Lena, I., Marsault, R., & Garcia, R. (2006).
Prefrontal infusion of PD098059 immediately after fear extinction
training blocks extinction-associated prefrontal synaptic plasticityand decreases prefrontal ERK2 phosphorylation. Synapse, 60,280–287.
Izaki, Y., Takita, M., Jay, T. M., Kaneko, H., Suzuki, S. S., &Nomura, M. (2001). Effect of long-term potentiation induction ongamma-band electroencephalograms in prefrontal cortex followingstimulation of rat hippocampus in vivo. Neuroscience Letters, 305,57–60.
Jay, T. M., & Witter, M. P. (1991). Distribution of hippocampal CA1 andsubicular efferents in the prefrontal cortex of the rat studied by meansof anterograde transport of Phaseolus vulgaris-leucoagglutinin. Jour-
nal of Computer Neurology, 313, 574–586.Ji, J., & Maren, S. (2005). Electrolytic lesions of the dorsal hippocampus
disrupt renewal of conditional fear after extinction. Learning and
Memory, 12, 270–276.Ji, J., & Maren, S. (2007). Hippocampal involvement in contextual
modulation of fear extinction. Hippocampus, 17, 749–758.Kim, J. J., Song, E. Y., & Kosten, T. A. (2006). Stress effects in the
hippocampus: synaptic plasticity and memory. Stress, 9, 1–11.Kim, J. J., & Yoon, K. S. (1998). Stress: metaplastic effects in the
hippocampus. Trends in Neurosciences, 21, 505–509.Laroche, S., Davis, S., & Jay, T. M. (2000). Plasticity at hippocampal to
prefrontal cortex synapses: dual roles in working memory andconsolidation. Hippocampus, 10, 438–446.
Lebron, K., Milad, M. R., & Quirk, G. J. (2004). Delayed recall of fearextinction in rats with lesions of ventral medial prefrontal cortex.Learn Mem, 11, 544–548.
Lopez-Fernandez, M. A., Montaron, M. F., Varea, E., Rougon, G.,Venero, C., Abrous, D. N., & Sandi, C. (2007). Upregulation ofpolysialylated neural cell adhesion molecule in the dorsal hippocampusafter contextual fear conditioning is involved in long-term memoryformation. Journal of Neuroscience, 27, 4552–4561.
Maren, S., & Fanselow, M. S. (1995). Synaptic plasticity in the basolateralamygdala induced by hippocampal formation stimulation in vivo. J
Neurosci, 15, 7548–7564.McDonald, R. J., Jones, J., Richards, B., & Hong, N. S. (2006). A double
dissociation of dorsal and ventral hippocampal function on a learningand memory task mediated by the dorso-lateral striatum. European
Journal of Neuroscience, 24, 1789–1801.Milad, M. R., & Quirk, G. J. (2002). Neurons in medial prefrontal cortex
signal memory for fear extinction. Nature, 420, 70–74.Miracle, A. D., Brace, M. F., Huyck, K. D., Singler, S. A., & Wellman, C.
L. (2006). Chronic stress impairs recall of extinction of conditionedfear. Neurobiology of Learning and Memory, 85, 213–218.
Moreau, J. L., Bourson, A., Jenck, F., Martin, J. R., & Mortas, P. (1994).Curative effects of the atypical antidepressant mianserin in the chronicmild stress-induced anhedonia model of depression. Journal of
Psychiatry and Neuroscience, 19, 51–56.Myers, K. M., & Davis, M. (2007). Mechanisms of fear extinction.
Molecular Psychiatry, 12, 120–150.Nascimento Hackl, L.P., & Carobrez, A.P. (2007) Distinct ventral and
dorsal hippocampus AP5 anxiolytic effects revealed in the elevatedplus-maze task in rats. Neurobiology of Learning & Memory (inpress).
Okulski, P., Jay, T. M., Jaworski, J., Duniec, K., Dzwonek, J.,Konopacki, F. A., Wilczynski, G. M., Sanchez-Capelo, A., Mallet,J., & Kaczmarek, L. (2007). TIMP-1 Abolishes MMP-9-DependentLong-lasting Long-term Potentiation in the Prefrontal Cortex. Bio-
logical Psychiatry, 62, 359–362.Pavlov, I. P. (1927). Conditioned reflexes. Oxford University Press.Quirk, G. J., Garcia, R., & Gonzalez-Lima, F. (2006). Prefrontal
mechanisms in extinction of conditioned fear. Biological Psychiatry,
60, 337–343.Quirk, G.J., & Mueller, D. (2007). Neural Mechanisms of Extinction
Learning and Retrieval. Neuropsychopharmacology (in press).Quirk, G. J., Russo, G. K., Barron, J. L., & Lebron, K. (2000). The role of
ventromedial prefrontal cortex in the recovery of extinguished fear. J
Neurosci, 20, 6225–6231.

566 R. Garcia et al. / Neurobiology of Learning and Memory 89 (2008) 560–566
Rocher, C., Spedding, M., Munoz, C., & Jay, T. M. (2004). Acute stress-induced changes in hippocampal/prefrontal circuits in rats: effects ofantidepressants. Cerebral Cortex, 14, 224–229.
Santini, E., Muller, R. U., & Quirk, G. J. (2001). Consolidation ofextinction learning involves transfer from NMDA-independent toNMDA-dependent memory. Journal of Neuroscience, 21,9009–9017.
Santini, E., Ge, H., Ren, K., Pena de Ortiz, S., & Quirk, G. J. (2004).Consolidation of fear extinction requires protein synthesis in themedial prefrontal cortex. Journal of Neuroscience, 24, 5704–5710.
Tarrier, N., Sommerfield, C., Pilgrim, H., & Humphreys, L. (1999).Cognitive therapy or imaginal exposure in the treatment of post-traumatic stress disorder. Twelve-month follow-up. British Journal of
Psychiatry, 175, 571–575.