implicit redundant-targets effect in visual extinction
TRANSCRIPT

Pergamon 0028-3932(95)00059-3
Neuropsychologia, Vol. 34, No. 1, pp. 9 22, 1996 Copyright ~ 1996 Elsevier Science Ltd
Printed in Great Britain. All rights reserved 0028-3932 / 96 $15.00 + 0.00
Implicit redundant-targets effect in visual extinction
C. A. MARZI,*] N. SMANIA,~ M. C. MARTINI, t G. GAMBINA,§ G. TOMELLERI,§ A. PALAMARA, t F. ALESSANDRINI§ and M. PRIOR¶
tDipartimento di Scienze Neurologiche e della Visione-Sezione di Fisiologia, University of Verona, Italy; $Servizio di Rieducazione Funzionale, Ospedale Policlinico, Verona, Italy; §Divisione di Neurologia, Neuroradiologia, Ospedale Civile Maggiore, Verona,
Italy; and ¶Dipartimento di Psicologia Generale, University of Padua, Italy
(Received 12 February 1995; accepted 25 April 1995)
Abstract--Patients with left visual extinction as a result of unilateral right hemisphere damage were tested on a redundant-targets effect paradigm (RTE). LED-generated brief flashes were lateralized either to the left or to the right visual hemifield or presented bilaterally. Subjects were asked to press a key as fast as possible following either unilateral or bilateral stimuli and immediately afterwards to report on the number of stimuli presented. As previously found in normal subjects, bilateral stimuli were responded to faster than unilateral ones, and this was evidence of a RTE. The main thrust of this study was that extinction patients showed a RTE not only for correctly perceived bilateral stimuli but also in trials in which they extinguished the stimulus on the field contralateral to the lesion. This result is compatible with a preserved processing of the extinguished input at least up to the stage at which it may interact with the input from the normal side to yield a speeded motor response. Interestingly, the implicit redundancy gain of extinction patients was found to fit a coactivation (i.e. neural) rather than a probabilistic model.
Key Words: extinction; redundant-target effect; interhemispheric transfer; parietal lobe; implicit processing.
Introduction
Visual extinction [4] consists in the inability to detect a target during simultaneous presentation of another similar target in the opposite hemifield, see [9] for a recent review. In contrast to hemianopic or hemineglect patients who fail to detect stimuli in the hemifield contralateral to the hemispheric side of the lesion, in extinction patients, the detection of contralesional single stimuli is unimpaired. Such a peculiar phenomenon is often seen following unilateral lesions of the parietal- temporal cortex of the right hemisphere (RH) but can also occur following left hemisphere (LH) lesions in corresponding areas. Extinction can be dissociated from unilateral neglect both clinically and anatomically [8, 17, 36], although is often considered as part of the same syndrome. It is a widely held belief that an under- standing of the neural mechanisms underlying extinc- tion can shed light on important aspects of normal perception and attention [29].
*Address for correspondence: Dipartimento di Scienze Neurologiche e della Visione, Sez. di Fisiologia, Universiti di Verona, 8 Strada Le Grazie, 37134 Verona, Italy.
The present report is concerned with a crucial aspect of extinction, namely whether the extinguished stimulus undergoes some degree of processing even though it is not consciously perceived. Evidence has been accumu- lating in the last years that the stimuli which are perceptually lost in visual hemineglect or extinction undergo a considerable extent of implicit processing [3, 5, 6, 13, 21-23, 37]. However, the stage in the visual information processing flow at which extinction occurs is still controversial [14, 15]. Therefore, what is needed is a fine-grain analysis of the perceptual stages at which implicit processing occurs, spanning from simple stimulus detection to cognitive higher-order operations.
Our strategy to tap implicit processing of the extinguished stimulus has been to use the redundant- target effect (RTE) paradigm, which consists in a speeding up of simple RT for redundant vs single targets. Such a strategy is similar to that we applied some years ago for revealing the presence of blindsight in hemianopic patients [11, 26]. In the present study we tested the RTE in patients who, as a result of RH damage, extinguished the stimulus presented to the left visual field (LVF) during bilateral presentations. The patients were asked to press a key as fast as possible following presentation of one or two brief LED-emitted

10 C .A . Marzi e t al./Implicit redundant-targets effect in visual extinction
light flashes using the h a n d ipsi la teral to the lesion. The st imuli could a p p e a r in an unpred ic tab le o rde r ei ther in the L V F or the R V F or in bo th sides s imultaneously. Wi th such p a r a d i g m in previous studies we found in a few hemianop ic pa t ien ts a R T E even when, dur ing bi la tera l p resenta t ions , one o f the st imuli fell into the hemianop ic field and therefore could no t be perceived. In the presen t exper iments we also asked the pat ients to indicate immedia t e ly fol lowing execut ion o f the m a n u a l response the n u m b e r and the loca t ion o f the st imuli they saw in tha t pa r t i cu l a r trial. This a l lowed us to compare the R T E in the cond i t i on in which the subjects saw bo th st imuli with tha t in which they showed extinction. Wi th such procedure , the presence o f impl ic i t process ing m a y be witnessed by the presence o f a R T E even in those trials in which the pa t ien ts c la imed to have seen only one target . Obvious ly , our a p p r o a c h does no t need to assume tha t R T E and pe rcep t ion o f b i la te ra l stimuli are necessari ly re la ted p h e n o m e n a (a l though they happen to go toge ther in n o r m a l subjects); it s imply takes advan tage o f the presence o f the fo rmer in the absence o f the la t ter to d r a w inferences a b o u t the fate o f the
' ex t inguished ' s t imulus. R e d u n d a n c y gain has been hypothes ized to result
e i ther f rom probab i l i s t i c s u m m a t i o n or f rom a coact iva- t ion process. The former can be expla ined by an independen t para l le l race mode l [30] which assumes tha t the st imuli to be s u m m a t e d are processed separa te ly and tha t the speed advan tage found for r e d u n d a n t targets derives f rom there being more chances for a s t imulus to undergo fast de tec t ion (win the race). A c c o r d i n g to coac t iva t ion models [27], on the o ther hand, a r edundancy gain occurs because signal s t rength is summed across targets unti l a cri t ical ac t iva t ion th resho ld is reached. F ina l ly , mixed models tha t i n c o r p o r a t e elements o f bo th the above p roposa l s have been pu t f o rwa rd recent ly [28, 33]. In the l ight o f these different accounts o f the RTE, a fur ther a im of the present s tudy was to assess which mode l might bet ter account for the R T E in our pa t ien ts with extinct ion.
Method
Subjects
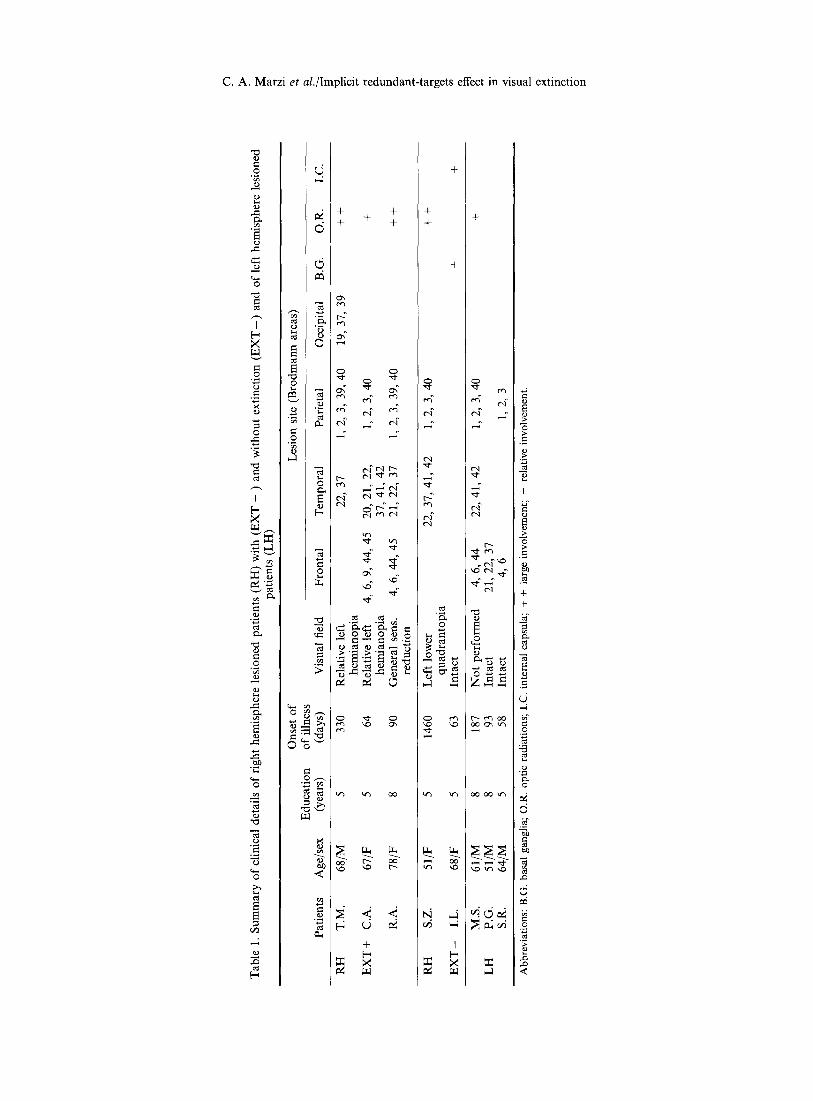
We studied a total of eight brain damaged patients who were divided into three groups on the basis of the presence and rate of extinction and of the hemispheric side of the lesion, see Table 1 for clinical details.
From a larger population of patients with damage to the RH we selected three patients with visual extinction (EXT + ), two patients with RH lesions but no extinction ( E X T - ) and finally we included in the study three patients with LH lesions and no extinction as controls. Extinction patients were included in the study only if they did not present with symptoms of spatial hemineglect at the time of testing and if they did not miss more than 30% of unilateral stimuli presented to the contralesional hemifield. Neglect was assessed by informal examination of various functional activities
including reading and, more formally, by means of the Albert line-crossing test [1] and by means of the letter 'H' cancellation test [34]. Finally, RH patients were included in the EXT + when their overall extinction rate, see Table 2, was higher than 40 %. RH patients with an extinction rate less than 5% and no spatial hemineglect were included in the E X T - group. Patients with an intermediate degree of extinction were not considered in the present study. All lesions were the consequence of ischaemic cerebral vascular accidents (see Fig. 1 for a reconstruction of the lesions). The patients were tested when they were undergoing motor or sensory rehabilitation. Particular care was taken in the evaluation of the visual field defects of the patients. All but one patient (M.S.) were tested by means of a Humphrey 630 computerized perimeter (Humphrey Instruments, San Leandro, Ca) using the 30-2 program. This program allows quantitative assessment in dB of the differential light threshold of 72 points distributed according to a regular 6 ° matrix within the central 30 °. The stimuli are white and their size is 4 mm2; they are presented in a random order with an exposure duration of 200 msec. The background luminance level is 10.0 cd-1 m 2 and the maximum luminance usable is 3180 cd -1 m 2. Two of the three EXT+ patients (T.M. and C.A.) showed an homonymous type of left hemianopia but their defect was relative rather than absolute in that they showed a reduction in sensitivity rather than a complete visual loss. The third patient (R.A.) showed an overall reduced sensitivity with a slightly more pronounced loss in the upper left quadrant.
It should be stressed that the presence of a relative left hemianopia in our EXT + is not in contrast with the relatively good detection rate shown for stimuli presented to the LVF, see Table 2, since none of the patients showed an absolute visual loss in the visual field area where the stimuli were presented in the RT task.
Apparatus, stimuli and procedure
Subjects were seated in front of a horizontal arc perimeter (57 cm radius) with the head restrained by a chin- and a front- rest. Attached to the bar were two LEDs of about 1 ° of visual angle diameter emitting a greenish light of 100 cd -1 m 2 luminance against a background of 1 cd I m 2 luminance. Activation of the LED was controlled by means of an interface connected to a portable PC which allowed to program the duration (10 msec) and the side of presentation (LVF, RVF or Bilateral) of the stimuli which were presented at an eccentricity of 10 ° from the fixation point. The alternation between conditions of stimulus presentation (hemifield side and stimulus number) was unpredictable with the constraint of having an equal number of stimuli for each condition. There were two separate sessions and in each of them there were 80 stimulus presentations for the two unilateral conditions together and 80 presentations for the bilateral condition. Therefore, the overall number of stimuli presented to each subject was 320. Subjects were asked to keep fixation steady on a white point (1 ° diameter) at the center of the perimetric bar and to respond to the presentation of a single or a double stimulus by pressing as fast as possible with the hand ipsilateral to the lesion the space bar of a PC keyboard placed in front of them. The fixation of the subject was controlled very carefully by an examiner and in the rare occurrence of an unsteady fixation, the trial was cancelled and repeated at the end of the block. In three patients, fixation was checked by means of an EOG recorded while performing the same task as above. Such recording provided evidence of an excellent match between the observational and the electro- physiological assessment of fixation. RTs shorter than 140 msec and longer than 800 msec were considered anticipations
C. A . M a r z i et al./Implicit r e d u n d a n t - t a r g e t s ef fect in v i s u a l e x t i n c t i o n
0
I b~ ×
0
0
4-
e/)
r~
4- 4- -I- 4- 4-
~ m ~ m ~ ~ m ~
r ~ ~ ~ ~
~ ~ r..~ ,.~ ~ ~ r - - I
~ ~ ~ ~ - ~
t¢~ t ~ OO
~ < <
q- [ -
4-
+
eq ~
¢-q
t 'q
tr~ tt~
06
I [ -
O
+
"6
.3
+
÷
o
o
:2 c5
~5
o
e~ <
12 C.A. Marzi et al./Implicit redundant-targets effect in visual extinction
RH
EXT+
Patients
T.M.
C.A.
R.A.
RH
EXT-
S.Z.
I.L.
LH
M.S.
P.G.
S, R.
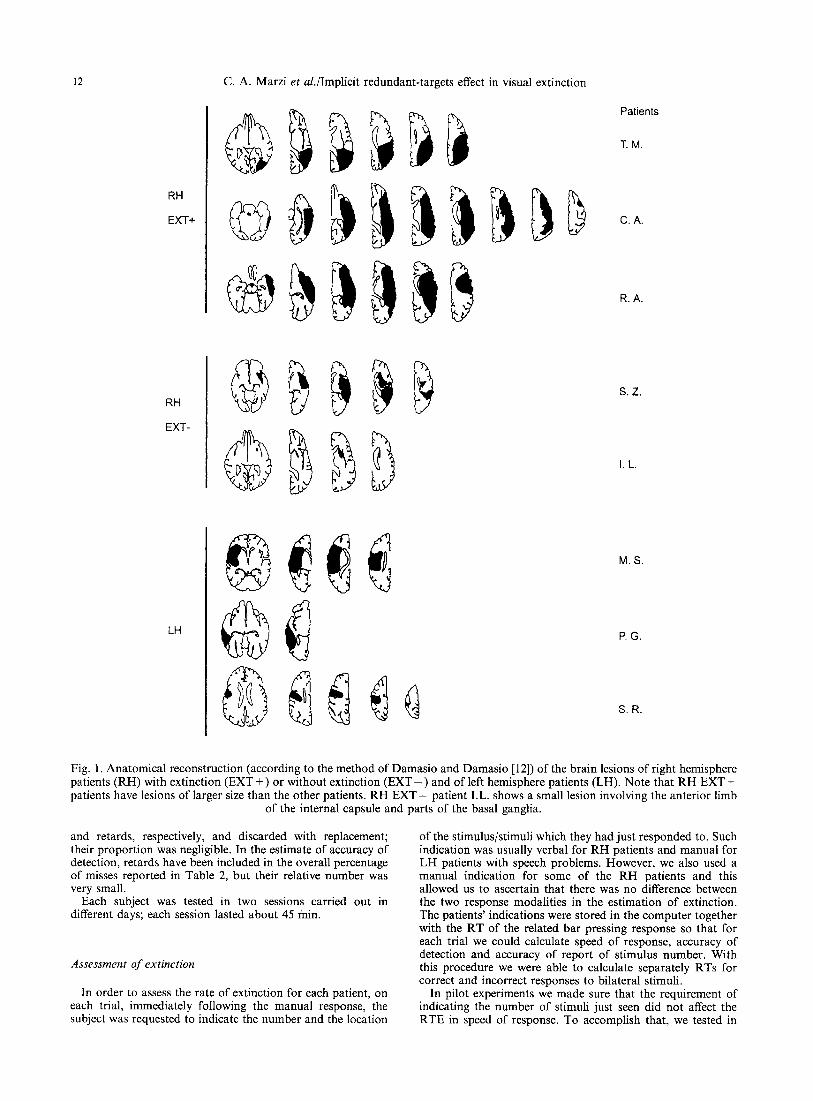
Fig. 1. Anatomical reconstruction (according to the method of Damasio and Damasio [12]) of the brain lesions of right hemisphere patients (RH) with extinction (EXT +) or without extinction ( E X T - ) and of left hemisphere patients (LH). Note that RH EXT + patients have lesions of larger size than the other patients. RH E X T - patient I.L. shows a small lesion involving the anterior limb
of the internal capsule and parts of the basal ganglia.
and retards, respectively, and discarded with replacement; their proportion was negligible. In the estimate of accuracy of detection, retards have been included in the overall percentage of misses reported in Table 2, but their relative number was very small.
Each subject was tested in two sessions carried out in different days; each session lasted about 45 rain.
Assessment of extinction
In order to assess the rate of extinction for each patient, on each trial, immediately following the manual response, the subject was requested to indicate the number and the location
of the stimulus/stimuli which they had just responded to. Such indication was usually verbal for RH patients and manual for LH patients with speech problems. However, we also used a manual indication for some of the RH patients and this allowed us to ascertain that there was no difference between the two response modalities in the estimation of extinction. The patients' indications were stored in the computer together with the RT of the related bar pressing response so that for each trial we could calculate speed of response, accuracy of detection and accuracy of report of stimulus number. With this procedure we were able to calculate separately RTs for correct and incorrect responses to bilateral stimuli.
In pilot experiments we made sure that the requirement of indicating the number of stimuli just seen did not affect the RTE in speed of response. To accomplish that, we tested in

14 C. A. Marzi et al./Implicit redundant-targets effect in visual extinction
Table 3. Speed of response
Unilateral Bilateral LVF RVF INC. CORR.
T.M. 493 432 413 426 RH C.A. 451 387 357 347 EXT + R.A. 514 401 386 - -
RH S.Z. 658 622 - - 587 E X T - I.L. 540 526 - - 504
M.S. 391 481 - - 378 LH P.G. 575 566 - - 535
S.R. 594 561 - - 568
However, it should be stressed that the crucial result here is that in the three E X T + patients, both correct and extinguished bilateral stimuli yield faster RTs than each of the two unilateral conditions of stimulus presentation. This can be taken as evidence of an implicit processing of the stimuli in the extinguished hemifield.
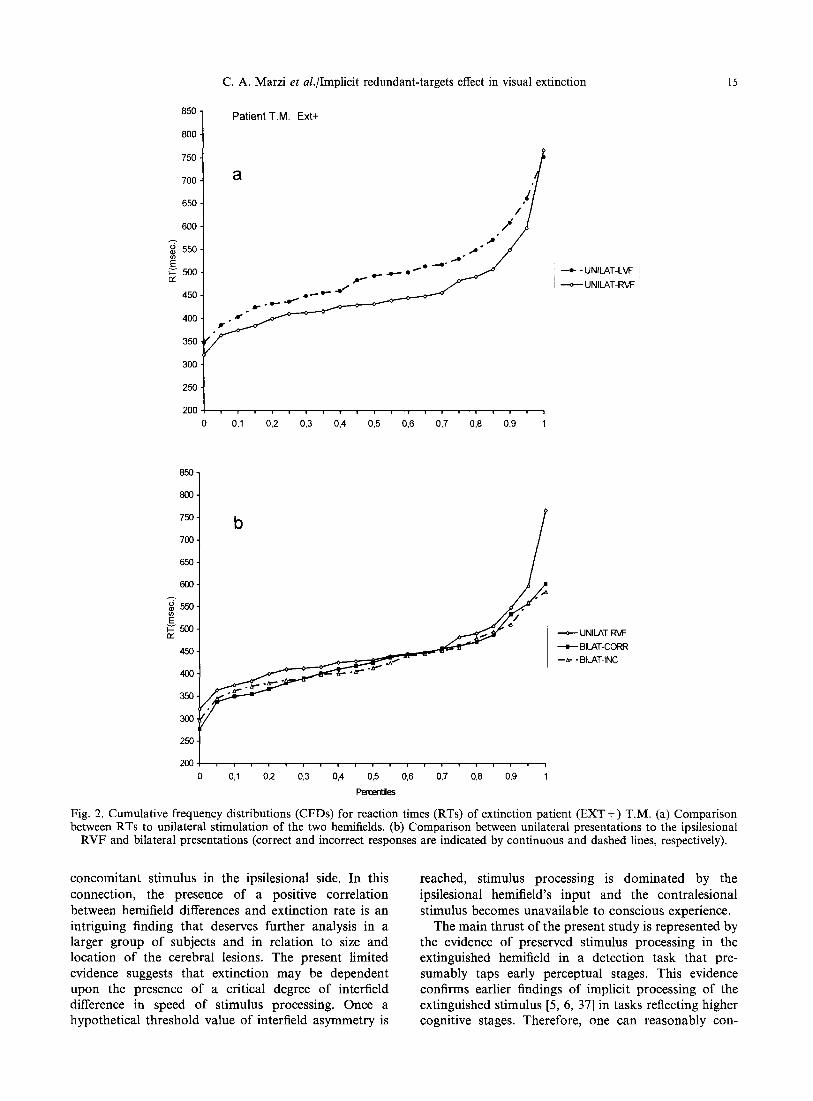
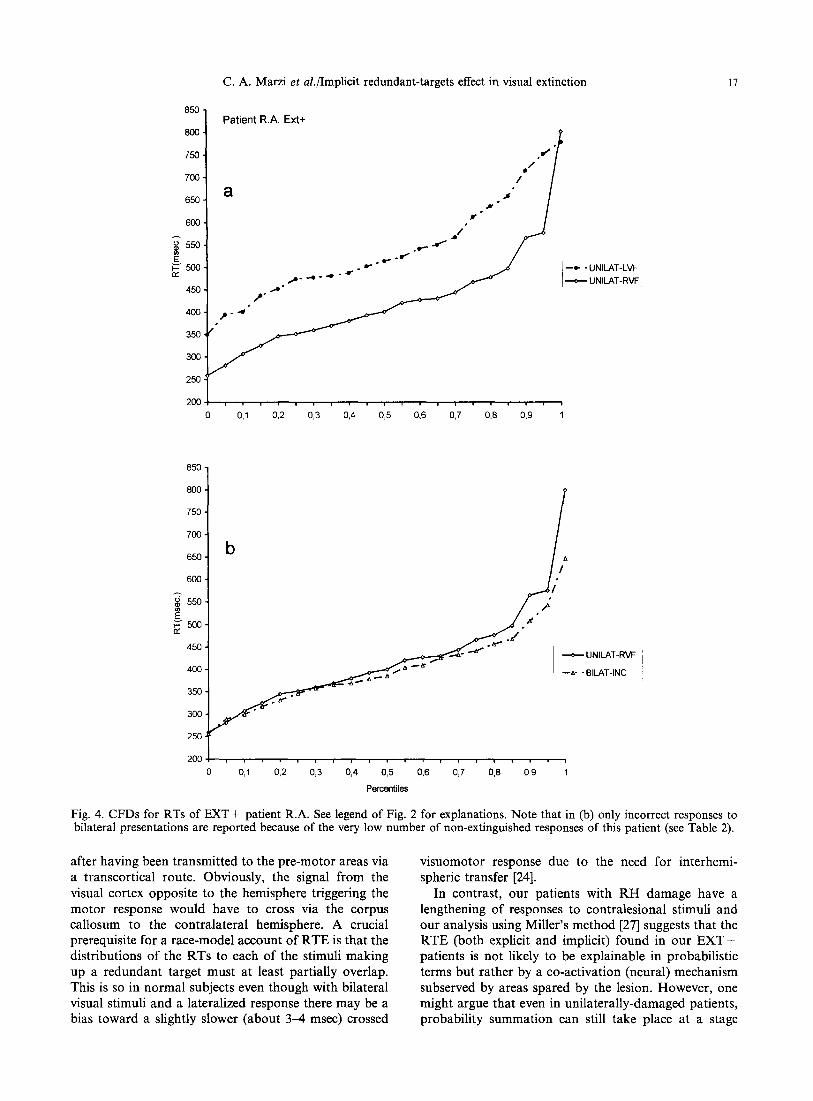
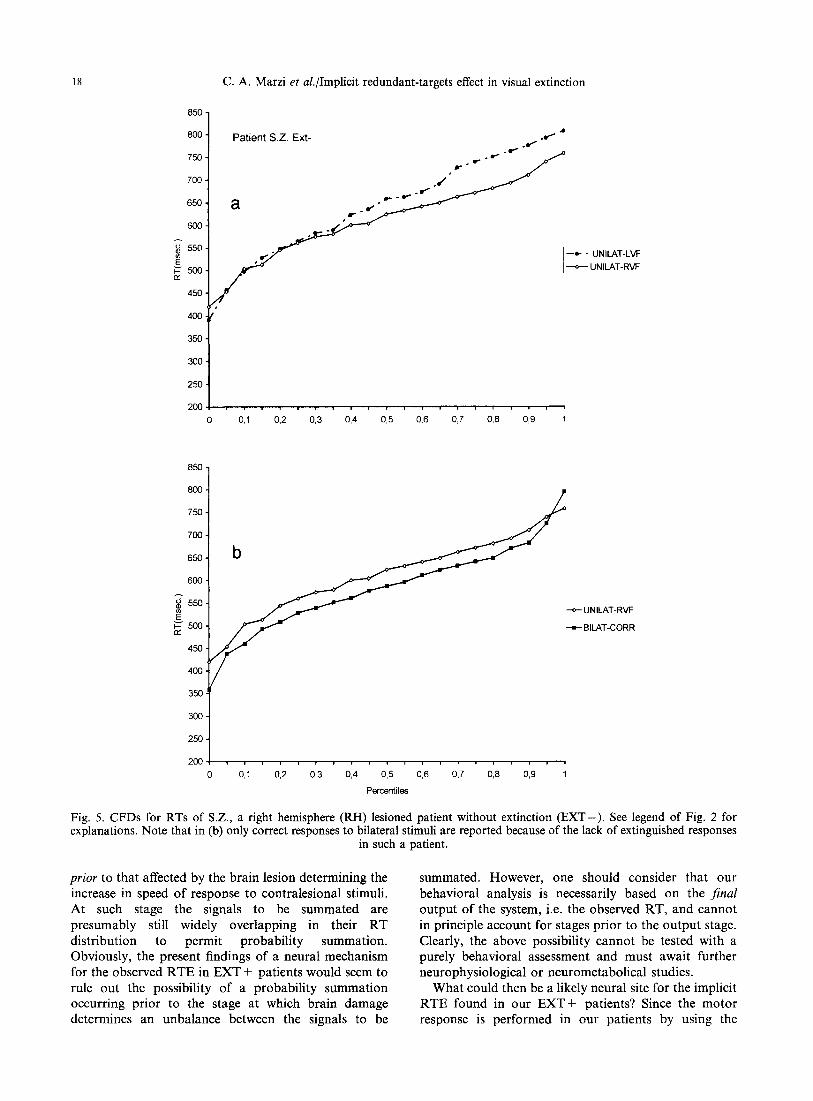
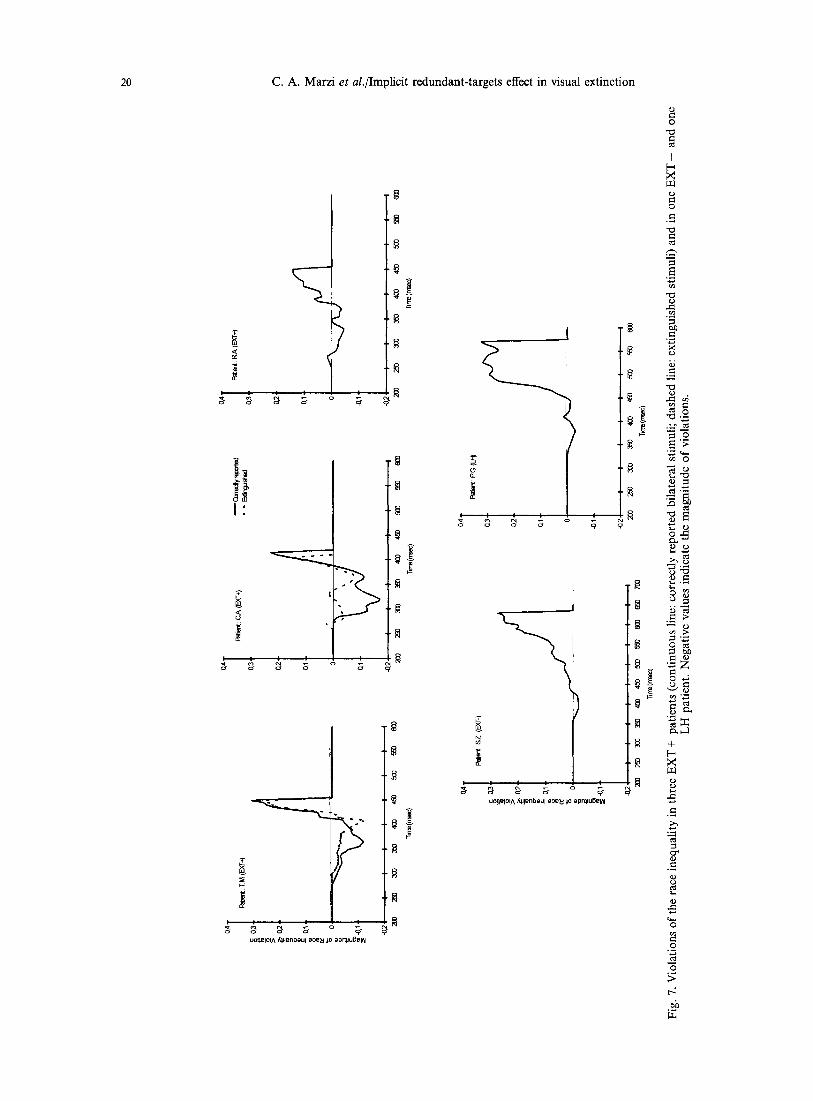
To further clarify the nature of such effect, we examined the possible presence of an implicit RTE using the method proposed by Miller [27], see above in the Method section. Figures 2(a)-4(a) show for each EXT + patient the CFDs of RTs to unilateral stimula- tion of the two hemifields: unsurprisingly, given the clearcut results reported in Table 3, the two CFDs do not cross each other, with the CFD related to the RVF dominating over the CFD related to the LVF. More interestingly for the present purposes are the results reported in Figs 2(b)--4(b) showing the CFDs for three conditions of stimulus presentation, namely, stimulation of the normal RVF, bilateral correctly reported stimuli and bilateral extinguished stimuli. The presence of a RTE is confirmed by the observation that in all the above three patients the CFDs of RTs to the correctly reported or extinguished bilateral stimuli show no or minimal crossing with the CFDs of RTs for unilateral stimulation of the faster hemifield (RVF). Finally, Fig. 7 shows the results of Miller's test of the race-inequality: the magnitude of violation is indicated as negative values below the zero line. In patients T.M. and C.A. there is a clear violation indicating a neural rather than a probabilistic mechanism of summation while in patient R.A. the violation is less conspicuous. For the former two patients this is true for both correctly reported and extinguished bilateral stimuli. A similar analysis was carried out in the patients of the two other groups: in all of them there was a clear RTE with a distribution of RTs similar to that of RH E X T + patients, see patient S.Z. as an example for the RH EXT-- group (Figs 5 and 7) and patient P.G. as an example for the LH group (Figs 6 and 7). Miller's race- inequality analysis showed unequivocally for 2/2 RH EXT-- patients and for two out of three LH patients a lack of violation, hence a presumably probabilistic explanation for the RTE.
Summing up, three differences can be noted between RH E X T + and the other two groups: first, interfield differences in RT to unilateral stimuli are much less in E X T - and LH patients [compare Figs 5(a) and 6(a) with 2(a), 3(a) and 4(a)]. Second, the amount of RTE seems to be somewhat more conspicuous in E X T - and LH patients than in EXT + patients [compare Figs 5(b) and 6(b) with Figs 2(b) and 4(b)]. Third, in E X T - and LH patients the mechanism underlying the observed RTE is presumably probabilistic while in E X T + appears to be neural (compare patients S.Z. and P.G. with patients T.M., C.A. and R.A. in Fig. 7).
Discussion
A first consideration concerns the observed length- ening of RT in the hemifield contralateral to the hemispheric lesion present in the majority of the patients tested in this study. This result has been obtained by other experiments in patients with unilateral hemi- spheric lesions [2, 20, 29]. Its nature is still unclear, but it is not unreasonable to relate it to a cortical intrahemispheric disconnection between the visually- receiving areas and those pre-motor areas that are supposed to initiate the motor response. The presence of a large hemispheric lesion in many of the above patients (see Fig. 1) is in keeping with such a possibility in that the normal cortico-cortical route subserving fast visuo- motor responses may not be viable. In addition, the presence of damage to the optic radiation in all EXT + patients, as well as in some patients of the other groups, suggests that part of the RT lengthening in the contralesional hemifield is likely to be related to an abnormally weak input to the cortical visual centers. However, independently from its nature, the presence of a larger laterality effect in RT in E X T + patients in comparison with the other two groups of the present study certainly shows that stimulus processing in the contralesional hemifield of an extinction patient is highly abnormal. This finding represents an obvious constraint for models of extinction in that they have to take into account a less efficient processing of stimuli in the contralesional hemifield even in the absence of a

C. A. Marzi et al./Implicit redundant-targets effect in visual extinction 13
normal as well as in some brain-damaged subjects whether RTE was affected by the subsequent response and we did not find any evidence for such an effect.
Data analys&
For each subject we plotted the cumulative frequency distributions (CFDs) of RTs for the three conditions of stimulus presentation. It has been shown convincingly [31] that such a detailed graphical description of the distribution is more suitable than summary statistics for examining the present data. Moreover, the analysis of the model that best explains the RTE requires assessment of the entire distribu- tions of RTs to double and single stimuli, rather than simply the means of these distributions [27, 31]. In particular, the presence of a RTE can be assessed by comparing the distribution of RTs to stimuli presented to the normal hemifield with those to bilateral stimuli. RTs to unilateral stimuli presented to the hemifield contralateral to the cerebral lesion are of no use in assessing RTE in unilaterally brain damaged patients because they are usually consistently slower than those for stimulation of the normal field (see Table 3). By the same reasoning, the presence of an implicit RTE can be revealed by comparing RTs to normal hemifield stimulation with those to extinguished bilateral stimuli. In both cases patients report having seen only a stimulus in the hemifield ipsilateral to the lesion. If there is an implicit RTE, however, they ought to have faster RTs for bilateral stimulation in spite of their conscious awareness of one stimulus.
A further goal of our study was to investigate the mechanism underlying the RTE in our patients. We used the formula proposed by Miller [27] to discriminate between a probability (race) model and a coactivation (neural) model. The race-model inequality test of Miller sets an upper limit for the cumulative probability of a response by any time t given redundant targets:
P(RT_<t[ T L and TR)<_P(RT<_tl T L) + P(RT_<tl TR),
where P(RT<tlT L and R _ T ) is the cumulative probability of a correct detection with bilateral targets, P(RT<_tIT L) is the cumulative probability of a response given a target in the LVF and nothing in the RVF, and P(RT_<tl T R) is the cumulative probability of a response given a target in the RVF and nothing in the LVF. When the upper bound is violated one is justified in speaking of neural summation; otherwise, a probabilistic explanation is more likely to apply, see [18, 32] for a similar approach and a more detailed description of the rationale.
R e s u l t s
Detection of unilateral and bilateral stimuli (extinction rate)
Table 2 shows the individual extinction score for R H and L H patients as well as the percentage o f correct detection o f unilateral stimuli. The vast majori ty o f errors for unilateral st imulation were represented by misses o f LVF flashes in R H patients, and of R V F flashes in one L H patient (S.R.). As to detection o f bilateral stimuli, R H patients were included in the extinction group ( E X T + ) when the percentage o f incorrect identifications o f bilateral stimuli was higher
Table 2. Overall extinction rate and percentage of missed unilateral stimuli in the two hemifields
Ext. rate LVF RVF
T.M. 59% 20% 7% RH C.A. 64% 15% 4% EXT + R.A. 96 % 29 % 11%
RH S.Z. 1% 24% 15% E X T - I.L. 3% 10% 5%
M.S. 1% 8% 8% LH P.G. 0% 11% 10%
S.R. 1% 8% 18%
than 40%. R H patients with few (less than 5%) or no errors in detecting bilateral stimuli were included in the E X T - group. Finally, L H patients showed practically no extinction at all.
Speed of response
The mean R T for responses to unilateral and bilateral stimulus representations are reported in Table 3.
Unilateral stimuli. For bo th E X T + and E X T - R H patients responses to LVF stimuli are clearly slower than to R V F stimuli with a mean difference o f 80 msec for the former and o f 25 msec for the latter group. Fur thermore, it might be interesting to note that each of the three E X T + patients has an overall speed of response faster than each E X T - patient. This suggests that the presence o f extinction a m o n g R H patients is not simply the result o f a larger aspecific effect o f brain damage witnessed by the larger size o f the lesion in R H + than in R H - patients (see Fig. 1). I f such would have been the case one would have expected to find longer overall RTs in E X T + than in the other R H group. Interestingly, only one o f the three L H patients (M.S.) shows a lengthening of the RTs to contralesional hemifield (RVF) stimulation and this is in line with previous reports o f a less lateralized effect o f L H lesions on R T [20].
A potentially impor tant result is represented by the positive correlation existing between interfield differ- ences in RT and extinction rate. Patient R.A. who has the highest extinction score (96%) also has the highest L V F - R V F difference in R T (113 msec); pat ient C.A. is in the middle position for both extinction rate (64%) and L V F - R V F difference (68 msec). Finally, patient T.M. has the smallest extinction rate (59%) and correspondingly the smallest L V F - R V F asymmetry (61 msec); the possible meaning o f such a correlat ion will be discussed in the Discussion section.
Bilateral stimuli. It can be seen f rom Table 3 that for all patients but one in the L H group (S.R.), bilateral responses are faster than those to the faster hemifield and this represents clear evidence that the vast majori ty o f our patients shows a R T E ( r a n g e = 6 - 3 5 msec).
C. A. Marzi et al./Implicit redundant-targets effect in visual extinction 15
850
8O0
75O
700
650
6oo A 6 550
450 -
400.
350.
300 •
250.
2O0
Patient T.M. Ext+
a / / Y
,D~ •,... -O O ~
Jr' R * ° L ~ ' ~ ~ " ~ ' ' ~ ~ "
--e- . UNILAT-LVF i i
UNILAT-R~I: '
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
850 ¸
8OO
75O
7 ~
65O
600
E ~'500
45O
4OO
350
3OO
250
200
b
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
Peroent]les
---o--- UNILAT-F~F
• BILAT-CORR
--~- - BILAT-INC
Fig. 2. Cumulative frequency distributions (CFDs) for reaction times (RTs) of extinction patient (EXT +) T.M. (a) Comparison between RTs to unilateral stimulation of the two hemifields. (b) Comparison between unilateral presentations to the ipsilesional
RVF and bilateral presentations (correct and incorrect responses are indicated by continuous and dashed lines, respectively).
concomitant stimulus in the ipsilesional side. In this connection, the presence of a positive correlation between hemifield differences and extinction rate is an intriguing finding that deserves further analysis in a larger group of subjects and in relation to size and location of the cerebral lesions. The present limited evidence suggests that extinction may be dependent upon the presence of a critical degree of interfield difference in speed of stimulus processing. Once a hypothetical threshold value of interfield asymmetry is
reached, stimulus processing is dominated by the ipsilesional hemifield's input and the contralesional stimulus becomes unavailable to conscious experience.
The main thrust of the present study is represented by the evidence of preserved stimulus processing in the extinguished hemifield in a detection task that pre- sumably taps early perceptual stages. This evidence confirms earlier findings of implicit processing of the extinguished stimulus [5, 6, 37] in tasks reflecting higher cognitive stages. Therefore, one can reasonably con-

16 C.A. Marzi et al./Implicit redundant-targets effect in visual extinction
850,
800.
750
700.
650 -
600 -
d 550.
E ~ 500. n,"
45O
400
35O
3OO
250 •
2OO
Pat ien t C.A. Ex t+
,o /
a /'/'/1 Y . I ~''
, i / "
,o~" ° ~" °'- "°/ Y
o* t"" ~' p . ~ - a,." " ~
--o- - UNILAT-LVF
UN LAT-RVF
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
850
800
750
700
650
600
o 5 5 0
~- 5 0 0
450
4 0 0
350
300.
250
2OO
/"
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9
Percentiles
UNILAT-RVF
• BILAT-CORR
--A- - BILAT-INC
Fig. 3. CFDs for RTs of EXT + patient C.A. See legend of Fig. 2 for explanations.
clude that in our E X T + patients, the concomitant presentation of a stimulus on the ipsilesional side does not suppress the stimulus on the contralesional side entirely but only at processing stages leading to conscious experience.
It would be interesting to assess which is th~ model better accounting for the implicit RTE in our E X T + patients because this would contribute to an under- standing o f its neural bases. Before attempting that, it is useful to briefly discuss what are the neural mechanisms underlying the RTE in normal subjects. We have preliminary evidence in our laboratory that visually
evoked potentials show a neurophysiological correlate of summation in the enhanced response of the P1 and N1 components. Such waveforms are believed to reflect the activity of extrastriate visual areas [10]. Therefore, it is likely that in normal subjects the RTE for the simple detection of flashes is subserved by a summation occurring at early cortical stages of visual processing but outside the primary visual cortex (V1). In previous experiments we found that such summation is presum- ably best explained by a race model [25]. According to such model, the fastest of the signals reaching the visual cortex will be the first to initiate the motor response
C. A. Marzi et al./Implicit redundant-targets effect in visual extinction ! 7
E
rY
8OO
75O
700
650.
600,
550,
500
450
400
350
300
250
20o
0
Patient R.A. Ext+
a ,"" ,,'/" /
--e- - UNILAT-LVF e"
. "
, . , ~ . . 4 . . 4 1 - ° ' e * UNILAT-RVF
,..,e ° ,p°
, , , , , , , , , , , , , , , , , , ,
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
850
8O0
750
700
650
600
550
E ~" 500 n-
45O
400
35O
300
25O
20O
b /4
.a." " a / "~ '
; .~" ~ ~ UNILAT-RVF z~ ~ Z t
~ " " ~ ~ a t --a- - BILAT-INC
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
Percentiles
Fig. 4. CFDs for RTs of EXT + patient R.A. See legend of Fig. 2 for explanations. Note that in (b) only incorrect responses to bilateral presentations are reported because of the very low number of non-extinguished responses of this patient (see Table 2).
after having been transmitted to the pre-motor areas via a transcortical route. Obviously, the signal from the visual cortex opposite to the hemisphere triggering the motor response would have to cross via the corpus callosum to the contralateral hemisphere. A crucial prerequisite for a race-model account of RTE is that the distributions of the RTs to each of the stimuli making up a redundant target must at least partially overlap. This is so in normal subjects even though with bilateral visual stimuli and a lateralized response there may be a bias toward a slightly slower (about 3 4 msec) crossed
visuomotor response due to the need for interhemi- spheric transfer [24].
In contrast, our patients with RH damage have a lengthening of responses to contralesional stimuli and our analysis using Miller's method [27] suggests that the RTE (both explicit and implicit) found in our EXT + patients is not likely to be explainable in probabilistic terms but rather by a co-activation (neural) mechanism subserved by areas spared by the lesion. However, one might argue that even in unilaterally-damaged patients, probability summation can still take place at a stage
18 C.A. Marzi et al./Implicit redundant-targets effect in visual extinction
850 •
800 •
750
700
65o
6oo
550.
g 5 0 0 .
4 5 0 •
4 0 0 -
350 •
300.
250 '
200
J Patient S.Z. Ext- . ~
o-. -o'~ . e , " ~
l --l- - UNILAT-LVF
l ~ UNILAT-RVF
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
850.
800.
750.
700.
650.
600.
550,
E ~ 500. r (
450 •
400.
350
300
250
2OO
b
UNILAT-RVF
] -~-BILAT-CORR
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
Percentiles
Fig. 5. CFDs for RTs of S.Z., a right hemisphere (RH) lesioned patient without extinction (EXT-) . See legend of Fig. 2 for explanations. Note that in (b) only correct responses to bilateral stimuli are reported because of the lack of extinguished responses
in such a patient.
prior to that affected by the brain lesion determining the increase in speed of response to contralesional stimuli. At such stage the signals to be summated are presumably still widely overlapping in their RT distribution to permit probability summation. Obviously, the present findings of a neural mechanism for the observed RTE in EXT + patients would seem to rule out the possibility of a probability summation occurring prior to the stage at which brain damage determines an unbalance between the signals to be
summated. However, one should consider that our behavioral analysis is necessarily based on the final output of the system, i.e. the observed RT, and cannot in principle account for stages prior to the output stage. Clearly, the above possibility cannot be tested with a purely behavioral assessment and must await further neurophysiological or neurometabolical studies.
What could then be a likely neural site for the implicit R T E found in our E X T + patients? Since the motor response is performed in our patients by using the

C. A. Marzi et al./Implicit redundant-targets effect in visual extinction 19
850
800
750-
700-
650-
600-
~ 550- f f l
E ~" 500- e,,
450.
400.
:350.
300.
250.
200
Pat ien t P . G LH
a /,-
l --e.- - UNILAT-L'VF
I - -~- UNILAT-~F i
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
850.
800.
750,
700,
650
600.
~55o. E ~ ' 5 0 0 rY
450
400
350
25O
200
b ~ UNILAT-RVF
i i v , i , i , , i , , , , , i , , , i
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
PercerCdles
Fig. 6. CFDs for RTs of P.G., a left hemisphere (LH) lesioned control patient. See legend of Fig. 2 for explanations. Note that in (b) only correct responses to bilateral stimuli are reported because of the lack of extinguished responses in such a patient.
ipsilesional (right) hand, it can be assumed that the convergence between the two hemispheric signals necessary for a neural summation subserving the speeded motor response occurs in the LH. Therefore, one could hypothesize that in EXT + patients exhibiting a dissociation between presence of a RTE and extinc- tion there is a selective impairment of the interhemi- spheric integration necessary to detect bilateral stimuli but that interhemispheric integration of visuo-motor responses is spared. In normal subjects, or in brain damaged patients without extinction, usually, the two
pathways of interhemispheric integration are both viable and the two processes go together. Therefore, normal subjects both perceive bilateral stimuli and show a RTE; however, such may not be the case if cortical damage affects the two interhemispheric pathways differentially. The large lesions crucially centered on parietal cortex that characterize patients with extinction are very likely to disrupt the widespread callosal connections subserving the interhemispheric integration of visual information over vast areas of the visual field [24]. On the other hand, the interhemispheric crossing
20 C.A. Marzi et al./Implicit redundant-targets effect in visual extinction
• , , , • ,
o- o- o ~ g- ~-
I '
/ I I o
/ i i i o
uop~elO~A/dl lenbeul a o e ~ JO epr~u~ei~ l
o - / i i i ~- ~ g- o ~.
I I I I
uop~e lO~ /~ lenbeu l ~ 0 e ~ Jo epn~ul~e~i
r,i
O
I
X
o
r~
~5
~D a= ,d
~.~
o ~
+
o "~_~
o

C. A. Marzi et al./Implicit redundant-targets effect in visual extinction 21
subserving the necessary visuomotor integration when visual hemifield of stimulus entry and responding hand belong to different hemispheres is likely to take place at several callosal sites in a sort of horse-race fashion [7]. It would not be surprising, therefore, that EXT + patients have difficulties in the interhemispheric integration of perceptual operations necessary to decide whether a unilateral or a bilateral stimulus has been presented while the interhemispheric visuomotor integration necessary for accessing the responding hand might be less affected.
Summing up, implicit RTE may be accounted for rather parsimoniously by assuming a differential impairment in the interhemispheric pathways subser- ving RTE vs perception of bilateral stimuli. In EXT + patients exhibiting an implicit RTE there might be a normal cross-over of information from the lesioned to the intact hemisphere initiating the motor response at several callosal sites. However, in the same patients, the interhemispheric callosal pathway important for the conscious detection of bilateral stimuli may be inter- rupted as a consequence of the lesion. Obviously, only dedicated functional neuro-imaging studies can help casting light on the neural localization of the implicit processing of the extinguished signal in our patients. However, on a more general perspective, the results of the present study add further evidence in favour of the idea (see [14, 35] for recent reviews) that the percep- tually lost information as a result of cortical damage might still be available for unconscious operations and especially for those related to motor performance [16, 19]. Needless to say, such dissociations can help us a great deal in understanding the neural substrate of conscious experience.
Acknowledgements--This research has been supported by CTB CNR 92 02686.04 granted to C.A.M. We acknowledge the precious help of Alessandra Fanini at various stages of the project and the invaluable contribution of Franco Capaldi and Silvia Pellati for the computerized reconstruction of the brain lesions. Finally, we thank Marco Veronese for the computer- ized preparation of the diagrams. We are particularly grateful to P.D. Dr. Wolfgang Schwarz for his most useful comments on a previous version of this paper.
References
1. Albert, M. L. A simple test for visual neglect. Neurology 23, 658-664, 1973.
2. Anzola, G. P. and Vignoto, L. A. Simple reaction time to lateralized visual stimuli is not related to the hemispheric side of lesion. Cortex 28, 401-409, 1992.
3. Baylis, G., Rafal, R. and Driver, J. Attentional set determines extinction following parietal lesions. J. cognit. Neurosci. 5, 453-466, 1993.
4. Bender, M. B. and Furlow, L. T. Phenomenon of visual extinction in homonymous half fields and the psychologic principles involved. Arch. Neurol. Psychiatry. 53, 29-33, 1945.
5. Berti, A., Allport, A., Driver, J., Dienes, Z., Oxbury, J. and Oxbury, S. Levels of processing for visual stimuli in an "extinguished" field. Neuropsychologia 30, 403-415, 1992.
6. Berti, A. and Rizzolatti, G. Visual processing without awareness: Evidence from unilateral neglect. J. cognit. Neurosci. 4, 345-351, 1992.
7. Bisiacchi, P., Marzi, C. A., Nicoletti, R., Carena, G., Mucignat, C. and Tomaiuolo, F. Left-right asymmetry of callosal transfer in normal human subjects. Behav. Brain Res. 64, 173-178, 1994.
8. Bisiach, E. Extinction and neglect: Same or different? In Brain and Space, J. Paillard (Editor), pp. 251-257. Oxford University Press, Oxford, 1991.
9. Bisiach, E. and Vallar, G. Hemineglect in humans. In Handbook of Neuropsychology, F. Boller and J. Grafman (Editors), Vol. I, pp. 195-222. Elsevier, Amsterdam, 1988.
10. Clark, V. P., Fan, S., and Hillyard, S. A. Identification of early visual evoked potential generators by retinotopic and topographic analyses. Human Brain Mapping 2, 170-187, 1995.
11. Corbetta, M., Marzi, C. A., Tassinari, G. and Aglioti, S. Effectiveness of different task paradigms in revealing blindsight. Brain 113, 603-616.
12. Damasio, H. and Damasio, A. R. Lesion Analysis in Neuropsychology. Oxford University Press, New York, 1989.
13. Driver, J., Bayliss, G. C. and Rafal, R. Preserved figure-ground segmentation and symmetry percep- tion in a patient with neglect. Nature 360, 73-75, 1993.
14. Farah, M. J. Visual perception and visual aware- ness after brain damage: A tutorial overview. In Attention and Performance XV. Conscious and Unconscious Information Processing, C. UmiltA and M. Moscovitch (Editors), pp. 37-76. MIT Press, Cambridge, Mass., 1994.
15. Farah, M. J., Monheit, M. A. and Wallace, M. A. Unconscious perception of "extinguished" visual stimuli: Reassessing the evidence. Neuropsychologia 29, 949-958, 1991.
16. Goodale, M. A., Meenan, J. P., Biilthoff, H. H., Nicolle, D. A., Murphy, K. J. and Racicot, C. I. Separate neural pathways for the visual analysis of object shape in perception and prehension. Curr. Biol. 4, 604-610, 1994.
17. Hier, D. B., Mondlock, J., Caplan, L. R. Recovery of behavioral abnormalities after right hemisphere stroke. Neurology 33, 345-350, 1983.
18. Hughes, H. C., Reuter-Lorenz, P. A., Fendrich, R. and Gazzaniga, M. S. Visual-auditory interactions in sensory-motor processing: Saccades versus manual responses. J. exp. Psychol.: Hum. Percept. Perform. 20, 131-153, 1994.

22 C.A. Marzi et al./Implicit redundant-targets effect in visual extinction
19. Jeannerod, M. The Neural and Behavioural Organi- zation of Goal-directed Movements. Clarendon Press, Oxford, 1988.
20. Kaizer, F., Korner-Bitenski, N., Mayo, N., Becker, R. and Coopersmith, H. Response time of stroke patients to a visual stimulus. Stroke 13, 335-339, 1988.
21. Karnath, H.-O. Deficits of attention in acute and recovered visual hemi-neglect. Neuropsychologia 26, 27-43, 1988.
22. Karnath. H.-O. and Hartje, W. Residual informa- tion processing in the neglected half-field. J. Neurol. 234, 180-184, 1987.
23. Marshall, J. C. and Halligan, P. W. Blindsight and insight in visuo-spatial neglect. Nature 336, 766- 767, 1988.
24. Marzi, C. A. Transfer of visual information after unilateral input to the brain. Brain Cognit. 5, 163- 173, 1986.
25. Marzi, C. A. Prior, M. and Martini, C. Spatial summation of widely separated stimuli: Neural or probabilistic? Paper presented at the 34th Meeting of the Psychonomic Society, abstr, n. 151, 1993.
26. Marzi, C. A., Tassinari, G., Aglioti, S. and Lutzemberger, L. Spatial summation across the vertical meridian in hemianopics: A test of blindsight. Neuropsychologia 24, 749-758, 1986.
27. Miller, J. Divided attention: Evidence for coactiva- tion with redundant signals. Cognit. Psychol. 14, 247-279, 1982.
28. Mordkoff, J. T. and Yantis, S. An interactive race model of divided attention. J. exp. Psychol.: Hum. Percept. Perform. 17, 520-538, 1991.
29. Posner, M. I., Walker, J. A., Friederich, F. J. and Rafal, R. D. Effects of parietal injury on covert orienting of attention. J. Neurosci. 4, 1863-1874, 1984.
30. Raab, D. Statistical facilitation of simple reaction time. Trans. N.Y. Acad. Sci. 43, 574-590, 1962.
31. Ratcliff, R. Group reaction time distributions and an analysis of distribution statistics. Psychol. Bull. 86, 446-461, 1979.
32. Reuter-Lorenz, P. A., Nozawa, G., Gazzaniga, M. S. and Hughes, H. C. The fate of neglected targets: A chronometric analysis of redundant target effects in the bisected brain. J. exp. Psychol.: Hum. Percept. Perform. 21, 211-230, 1995.
33. Schwarz, W. and Ischebeck, A. Coactivation and statistical facilitation in the detection of lines. Perception 23, 157-168, 1994.
34. Weinberg, J., Diller, L., Gordon, W. A., Gerstman, L. J., Lieberman, A., Lakin, P., Hodges, G. and Ezrachi, O. Visual scanning training effect on reading-related tasks in acquired right brain damage. Arch. Phys. Med. Rehabil. 58, 479-486, 1977.
35. Weiskrantz, L. Outlooks for blindsight: Explicit methodologies for implicit processes. Proc. R. Soc. London B239, 247-278, 1990.
36. Vallar, G., Rusconi, M. L., Bignamini, L., Gemi- niani, G. and Perani, D. The anatomical correlates of visual and tactile extinction in humans. A clinical CT-scan study. J. Neurol. Neurosurg. Psychiat. 57, 464-470, 1994.
37. Volpe, B. T., LeDoux, J. E. and Gazzaniga, M. S. Information processing of visual stimuli in an 'extinguished' field. Nature 282, 722-724, 1979.