genetically dependent modulation of serotonergic inactivation in the human prefrontal cortex
TRANSCRIPT

www.elsevier.com/locate/ynimg
NeuroImage 40 (2008) 1264–1273Genetically dependent modulation of serotonergic inactivationin the human prefrontal cortex
Luca Passamonti,a,b Antonio Cerasa,a,c Maria Cecilia Gioia,a Angela Magariello,a
Maria Muglia,a Aldo Quattrone,a,d and Francesco Feraa,e,⁎
aInstitute of Neurological Sciences, National Research Council, Piano Lago di Mangone, Cosenza, 87050, ItalybCognition Brain Science Unit, Medical Research Council, Cambridge, CB2 7EF, UKcDepartment of Neurosciences, Psychiatry and Anaesthesiology, University of Messina, I-98125, Messina, ItalydInstitute of Neurology, University “Magna Graecia”, Catanzaro, 88100, ItalyeNeuroradiology Unit, University “Magna Graecia”, Catanzaro, 88100, Italy
Received 12 September 2007; revised 6 December 2007; accepted 11 December 2007Available online 27 December 2007
Previous research suggests that genetic variations regulating seroto-nergic neurotransmission mediate individual differences in the neuralnetwork underlying impulsive and aggressive behaviour. Althoughwith conflicting findings, the monoamine oxidase-A (MAOA) and theserotonin transporter (5HTT) gene polymorphisms have been asso-ciated with an increased risk to develop impulsive and aggressivebehaviour. Double knock-out mice studies have also demonstrated thatMAOA and 5HTT genes strongly interact in the metabolic pathwayleading to the serotonergic inactivation; however, their potentialinteractive effect in human brain remains uninvestigated.
We used blood oxygenation level-dependent (BOLD) functionalmagnetic resonance imaging (fMRI) to assess the independent andinteractive effects of both MAOA and 5HTT polymorphisms on thebrain activity elicited by a response inhibition task in healthy volunteers.
Multivariate analysis demonstrated an individual effect of bothMAOA and 5HTT polymorphisms and a strong allele–allele interac-tion in the anterior cingulate cortex (ACC), a key region implicated incognitive control and in the pathophysiology of impulsive and aggres-sive behaviour.
These findings suggest that the MAOA×5HTT allelic interactionexerts a significant modulation on the BOLD response associated withresponse inhibition and contribute to validate haplotype models asuseful tools for a better understanding of the neurobiology underlyingcomplex cognitive functions.© 2008 Elsevier Inc. All rights reserved.
Keywords: Impulsivity; MAOA; 5HTT; Gene polymorphisms; Responseinhibition; fMRI
⁎ Corresponding author. University of Catanzaro, “Magna Graecia”,Building B, Level 0, Room 86, Viale Europa, Germaneto (CZ), 88100, Italy.
E-mail address: [email protected] (F. Fera).Available online on ScienceDirect (www.sciencedirect.com).
1053-8119/$ - see front matter © 2008 Elsevier Inc. All rights reserved.doi:10.1016/j.neuroimage.2007.12.028
Introduction
Serotonergic dysfunction within the orbitofrontal cortex (OFC)and anterior cingulate cortex (ACC) is proposed as the core deficitin the pathophysiology of several psychiatric diseases character-ized by aggressive and impulsive behaviour (Davidson et al., 2000;Frankle et al., 2005; Robbins, 2005). Reducing the concentrationof serotonin (5-HT) by dietary tryptophan depletion in humans orselectively destructing the ascending serotonergic projections inanimals strongly alters the OFC and ACC function (Park et al.,1994; Mobini et al., 2000; Clarke et al., 2004). Genetic variations,environmental factors and more complex genes by environmentinteractions have been associated with an increased risk to developaggressive behaviour and other psychiatric disorders (Noblett andCoccaro, 2005; Caspi et al., 2002, 2003). Although the underlyingneurobiological mechanisms are still poorly understood, they mayresult from individual differences in the serotonergic neurotrans-mission within the OFC and ACC due, for instance, to geneticallyregulated variations in the inactivation of 5-HT (i.e. reuptake bythe 5-HT transporter (5HTT) and breakdown by the monoamineoxidase-A (MAOA) enzyme).
Both MAOA and 5HTT genes present two well-characterizedlength-variable [i.e. variable number of tandem repeats (VNTR)]polymorphisms in their promoter regions that alter the transcriptionalactivity of the genes and hence the function of the correspondingproteins (Lesch et al., 1996; Sabol et al., 1998; Denney et al., 1999).The long variant of the MAOA polymorphism promotes a highertranscription of the gene [MAOA high-activity allele (MAOA-H)]which results in a higher activity of the enzyme when compared withthe MAOA short variant [therefore defined as MAOA-low activityallele (MAOA-L)] (Sabol et al., 1998; Denney et al., 1999). The5HTT gene presents an analogous VNTR polymorphism in the pro-moter region with short (5HTT-S) and long (5HTT-L) variants (Leschet al., 1996). The analysis of lymphoblast cells demonstrated that the

1265L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
5HTT-S allele resulted in a reduced serotonin uptake, lower serotoninreceptor binding and lower mRNA concentration compared with the5HTT-L allele (Lesch et al., 1996). Albeit different association studieshave not consistently associated MAOA-H and 5HTT-S alleles withimpulsive and aggressive behaviour or with other psychiatricdisorders (Caspi et al., 2002, 2003; Courtet et al., 2004; Baca-Garciaet al., 2004; Gerra et al., 2004), converging evidence suggests thatthese polymorphisms strongly regulate the serotonergic function invitro and in vivo (Lesch et al., 1996; Sabol et al., 1998; Denney et al.,1999; Manuck et al., 2000, 2002).
Functional imaging procedures that use the “endophenotypicapproach” have began to show how genetic variations in the sero-tonergic molecular pathway modulate the neural circuits underlyingthe impulsive and aggressive behaviour (Congdon and Canli 2005;Hariri et al., 2006a). A blood oxygenation level-dependent (BOLD)functional magnetic resonance imaging (fMRI) study showed thatMAOA-H carriers displayed a greater response of the anteriorcingulate cortex (ACC) when compared with MAOA-L carriersduring the execution of an attentional task (conflict resolution) (Fanet al., 2003). Other findings further demonstrated that the responseof orbitofrontal cortex (OFC), elicited by a response inhibitionparadigm (Go/NoGo), was increased in MAOA-H carriers (Passa-monti et al., 2006). Finally, a recent work reported the profoundimpact of the MAOA polymorphism on different brain circuitsinvolved in emotions, memory and cognitive control (Meyer-Lindenberg et al., 2006). Those authors showed a strong genderrelated difference, such that the ACC response during the executionof a cognitive control paradigm (response inhibition flanker task)was much greater in males carrying the MAOA-H allele.
Although both animal studies and imaging findings on humansalso highlighted the impact of the 5HTT gene on the modulation ofemotional circuits (Holmes et al., 2002; Hariri et al., 2002;Bertolino et al., 2005; Wellman et al., 2007), how MAOA and5HTT genes might interact in the human brain remains unexplored.Comparative research indicates that these genes present combinedand interplaying effects in regulating the metabolic pathway thatleads to 5-HT inactivation (Murphy et al., 2003).
On the basis of these previous findings, we expected thatMAOA and 5HTT genes might play an interactive role inmodulating the serotonergic function, in particular in the OFCand the ACC, two cerebral regions presenting a high expression ofthe MAOA and 5HTT proteins (Fowler et al., 1987; Varnas et al.,2004) and a dense serotonergic innervation from the raphe nuclei(Clarke et al., 2004). To directly assess the individual andinteractive contributions of the MAOA and 5HTT polymorphismson brain responses, we studied healthy male volunteers usingBOLD fMRI during the execution of a response inhibition task.
We hypothesized that individuals carrying simultaneously theallelic variants conferring a reduced serotonergic function (i.e.subjects carrying both the MAOA-H variant and at least one copy ofthe 5HTT-S allele) would display an increased BOLD response inthe OFC and ACC compared with subjects presenting a geneticallymediated higher serotonergic function (i.e. subjects carrying boththe MAOA-L variant and homozygous for the 5HTT-L allele).
Subjects and methods
Subjects
Thirty-five right-handed healthy males (Caucasian, fromsouthern Italy, age-range: 20–44 years) were recruited by local
advertisements. Fifteen of them also participated in a previousstudy (Passamonti et al., 2006). We included only men for severalreasons: (1) impulsive and aggressive behaviour is a typical malefeature (Loeber and Keenan, 1994); (2) MAOA polymorphismsmap in a region of the X chromosome suspected to escape thenormal inactivation (Carrel et al., 1999); this makes it very difficultcomparing hemizygous males (carrying either one MAOA high-activity allele or one MAOA low-activity allele) to homozygousfemales (carrying either two MAOA high-activity alleles or twoMAOA low-activity alleles) or heterozygous females (carryingboth one MAOA high-activity allele and one MAOA low-activityallele) in terms of enzymatic activity; (3) there is evidence that theeffects of MAOA-H and 5-HTT-S alleles on the serotonergicfunction in vivo varied as a function of both ethnicity and gender(Williams et al., 2003); (4) previous fMRI results showed a gender-related (males) effect of MAOA-H allele on the ACC BOLDactivity during the execution of cognitive control task (Meyer-Lindenberg et al., 2006).
Inclusion criteria were as follows: ability to give voluntaryinformed consent, absence of concurrent neurological or psychiatricdisorders according to DSM-IV criteria (American PsychiatricAssociation, 1994), no history of head trauma, alcohol or drug abuse,no history of other medical problems ormedical treatments relevant tocerebral blood flow andmetabolism, optimal visual acuity and normalMRI of the brain. Written informed consent was obtained by allsubjects participating to the study, which was approved by the localethics committee at the University of Catanzaro, in accordance withthe guidelines of the Helsinki Declaration (1983). Intelligencequotient (IQ) was estimated using the Wechsler Adult IntelligenceScale–Revised (WAIS-R) (Wechsler, 1981). Trait impulsivity wasassessedwith the Italian version ofBarratt Impulsivity Scale (BIS-11),a self-report measure that presents high internal consistency and hightest–retest reliability (Fossati et al., 2001).
All subjects were genotyped based on the MAOA-H and theMAOA-L variants, and based on the long (5HTT-L) and short(5HTT-S) alleles of the 5HTT gene. Previous pharmacologicalstudies showed a blunted serotonergic responsivity in vivo in healthyvolunteers carrying either the MAOA-H variant or at least one copyof the 5HTT-S allele (Manuck et al., 2000, 2002; Whale et al., 2000;Reist et al., 2001). Because carriers of either one or two copies of the5HTT-S allele did not differ each other in term of protein binding(Little et al., 1998; Heinz et al., 2000), we grouped them together, inaccordance also with previous imaging studies (Hariri et al., 2002;Smith et al., 2004). Hence, subjects simultaneously carrying theMAOA-H variant and at least one copy of the 5HTT-S allele weregrouped in a “hypo-serotonergic” group (presumed lowest seroto-nergic responsivity), while individuals homozygous for the 5HTT-Lallele (two long alleles) carrying also the MAOA-L variant weregrouped in a “hyper-serotonergic” group (presumed greatestserotonergic responsivity). Any other combination of MAOA and5HTTalleles [i.e. carriers of theMAOA-H variant also homozygousfor the 5HTT-L allele (5HTT-L,L) or subjects carrying theMAOA-Lvariant and at least one copy of the 5HTT-S allele] was hypothesizedto result in an intermediate serotonergic function.
Genotyping
DNAwas extracted from blood samples obtained from all subjectsaccording to standard procedures. The MAOA promoter region poly-morphism was genotyped based on previously described methods(Passamonti et al., 2006). The 5HTT regulatory gene region was
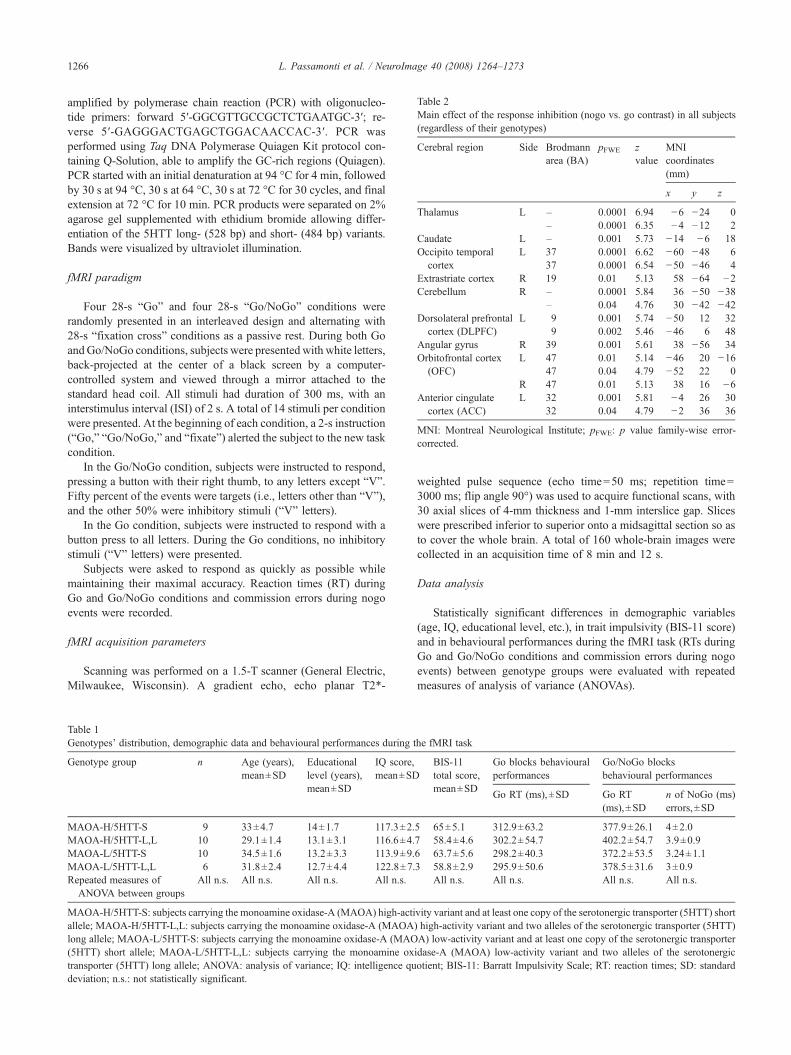
Table 2Main effect of the response inhibition (nogo vs. go contrast) in all subjects(regardless of their genotypes)
Cerebral region Side Brodmannarea (BA)
pFWE zvalue
MNIcoordinates(mm)
x y z
Thalamus L – 0.0001 6.94 −6 −24 0– 0.0001 6.35 −4 −12 2
Caudate L – 0.001 5.73 −14 −6 18Occipito temporal
cortexL 37 0.0001 6.62 −60 −48 6
37 0.0001 6.54 −50 −46 4Extrastriate cortex R 19 0.01 5.13 58 −64 −2Cerebellum R – 0.0001 5.84 36 −50 −38
– 0.04 4.76 30 −42 −42Dorsolateral prefrontal
cortex (DLPFC)L 9 0.001 5.74 −50 12 32
9 0.002 5.46 −46 6 48Angular gyrus R 39 0.001 5.61 38 −56 34Orbitofrontal cortex
(OFC)L 47 0.01 5.14 −46 20 −16
47 0.04 4.79 −52 22 0R 47 0.01 5.13 38 16 −6
Anterior cingulatecortex (ACC)
L 32 0.001 5.81 −4 26 3032 0.04 4.79 −2 36 36
MNI: Montreal Neurological Institute; pFWE: p value family-wise error-corrected.
1266 L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
amplified by polymerase chain reaction (PCR) with oligonucleo-tide primers: forward 5′-GGCGTTGCCGCTCTGAATGC-3′; re-verse 5′-GAGGGACTGAGCTGGACAACCAC-3′. PCR wasperformed using Taq DNA Polymerase Quiagen Kit protocol con-taining Q-Solution, able to amplify the GC-rich regions (Quiagen).PCR started with an initial denaturation at 94 °C for 4 min, followedby 30 s at 94 °C, 30 s at 64 °C, 30 s at 72 °C for 30 cycles, and finalextension at 72 °C for 10 min. PCR products were separated on 2%agarose gel supplemented with ethidium bromide allowing differ-entiation of the 5HTT long- (528 bp) and short- (484 bp) variants.Bands were visualized by ultraviolet illumination.
fMRI paradigm
Four 28-s “Go” and four 28-s “Go/NoGo” conditions wererandomly presented in an interleaved design and alternating with28-s “fixation cross” conditions as a passive rest. During both GoandGo/NoGo conditions, subjects were presented with white letters,back-projected at the center of a black screen by a computer-controlled system and viewed through a mirror attached to thestandard head coil. All stimuli had duration of 300 ms, with aninterstimulus interval (ISI) of 2 s. A total of 14 stimuli per conditionwere presented. At the beginning of each condition, a 2-s instruction(“Go,” “Go/NoGo,” and “fixate”) alerted the subject to the new taskcondition.
In the Go/NoGo condition, subjects were instructed to respond,pressing a button with their right thumb, to any letters except “V”.Fifty percent of the events were targets (i.e., letters other than “V”),and the other 50% were inhibitory stimuli (“V” letters).
In the Go condition, subjects were instructed to respond with abutton press to all letters. During the Go conditions, no inhibitorystimuli (“V” letters) were presented.
Subjects were asked to respond as quickly as possible whilemaintaining their maximal accuracy. Reaction times (RT) duringGo and Go/NoGo conditions and commission errors during nogoevents were recorded.
fMRI acquisition parameters
Scanning was performed on a 1.5-T scanner (General Electric,Milwaukee, Wisconsin). A gradient echo, echo planar T2⁎-
Table 1Genotypes' distribution, demographic data and behavioural performances during t
Genotype group n Age (years),mean±SD
Educationallevel (years),mean±SD
IQ score,mean±SD
MAOA-H/5HTT-S 9 33±4.7 14±1.7 117.3±2.MAOA-H/5HTT-L,L 10 29.1±1.4 13.1±3.1 116.6±4.MAOA-L/5HTT-S 10 34.5±1.6 13.2±3.3 113.9±9.MAOA-L/5HTT-L,L 6 31.8±2.4 12.7±4.4 122.8±7.Repeated measures of
ANOVA between groupsAll n.s. All n.s. All n.s. All n.s.
MAOA-H/5HTT-S: subjects carrying the monoamine oxidase-A (MAOA) high-actiallele; MAOA-H/5HTT-L,L: subjects carrying the monoamine oxidase-A (MAOAlong allele; MAOA-L/5HTT-S: subjects carrying the monoamine oxidase-A (MAO(5HTT) short allele; MAOA-L/5HTT-L,L: subjects carrying the monoamine oxtransporter (5HTT) long allele; ANOVA: analysis of variance; IQ: intelligence qudeviation; n.s.: not statistically significant.
weighted pulse sequence (echo time=50 ms; repetition time=3000 ms; flip angle 90°) was used to acquire functional scans, with30 axial slices of 4-mm thickness and 1-mm interslice gap. Sliceswere prescribed inferior to superior onto a midsagittal section so asto cover the whole brain. A total of 160 whole-brain images werecollected in an acquisition time of 8 min and 12 s.
Data analysis
Statistically significant differences in demographic variables(age, IQ, educational level, etc.), in trait impulsivity (BIS-11 score)and in behavioural performances during the fMRI task (RTs duringGo and Go/NoGo conditions and commission errors during nogoevents) between genotype groups were evaluated with repeatedmeasures of analysis of variance (ANOVAs).
he fMRI task
BIS-11total score,mean±SD
Go blocks behaviouralperformances
Go/NoGo blocksbehavioural performances
Go RT (ms),±SD Go RT(ms),±SD
n of NoGo (ms)errors,±SD
5 65±5.1 312.9±63.2 377.9±26.1 4±2.07 58.4±4.6 302.2±54.7 402.2±54.7 3.9±0.96 63.7±5.6 298.2±40.3 372.2±53.5 3.24±1.13 58.8±2.9 295.9±50.6 378.5±31.6 3±0.9
All n.s. All n.s. All n.s. All n.s.
vity variant and at least one copy of the serotonergic transporter (5HTT) short) high-activity variant and two alleles of the serotonergic transporter (5HTT)A) low-activity variant and at least one copy of the serotonergic transporteridase-A (MAOA) low-activity variant and two alleles of the serotonergicotient; BIS-11: Barratt Impulsivity Scale; RT: reaction times; SD: standard
1267L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
Analyses of fMRI data were completed with SPM2 software(www.fil.ion.ucl.ac.uk) under the assumption of the general linearmodel (GLM). Images for each subject were realigned to the firstin the time series to correct for head motion, spatially normalizedto a standard stereotactic space [Montreal Neurological Institute(MNI) template] with a 12-parameter affine model, and smoothedwith a 6-mm full-width at half-maximum (FWHM) isotropicGaussian kernel to minimize noise and residual differences in gyralanatomy and to allow valid statistical inference according toGaussian random field theory (Friston et al., 1995). Subjectsshowing head movements greater than 2 mm were excluded fromall subsequent analyses.
For each participant we used a GLM to assess regionallyspecific effects of task parameters on BOLD indices of activation.The first level model included two experimental factors (nogo andgo events) and effects of no interest (fixation cross and realignmentparameters). The individual first level images were generated using
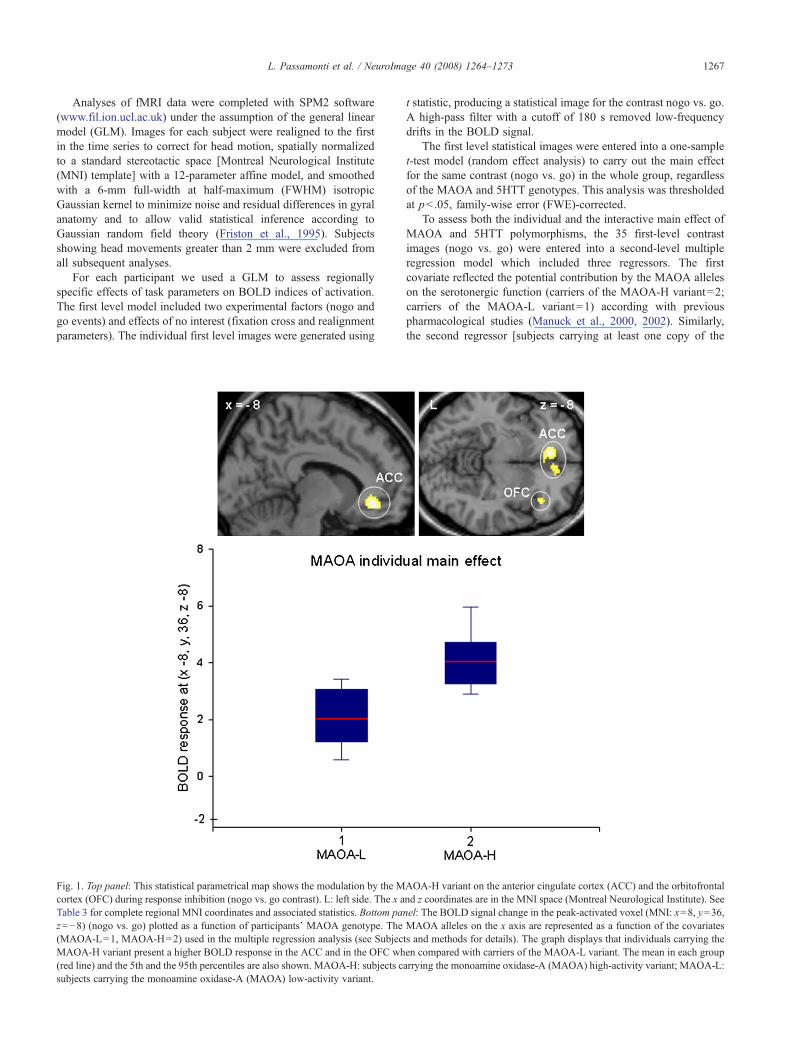
Fig. 1. Top panel: This statistical parametrical map shows the modulation by the Mcortex (OFC) during response inhibition (nogo vs. go contrast). L: left side. The x aTable 3 for complete regional MNI coordinates and associated statistics. Bottom panz=−8) (nogo vs. go) plotted as a function of participants' MAOA genotype. The(MAOA-L=1, MAOA-H=2) used in the multiple regression analysis (see SubjecMAOA-H variant present a higher BOLD response in the ACC and in the OFC wh(red line) and the 5th and the 95th percentiles are also shown. MAOA-H: subjects csubjects carrying the monoamine oxidase-A (MAOA) low-activity variant.
t statistic, producing a statistical image for the contrast nogo vs. go.A high-pass filter with a cutoff of 180 s removed low-frequencydrifts in the BOLD signal.
The first level statistical images were entered into a one-samplet-test model (random effect analysis) to carry out the main effectfor the same contrast (nogo vs. go) in the whole group, regardlessof the MAOA and 5HTT genotypes. This analysis was thresholdedat pb .05, family-wise error (FWE)-corrected.
To assess both the individual and the interactive main effect ofMAOA and 5HTT polymorphisms, the 35 first-level contrastimages (nogo vs. go) were entered into a second-level multipleregression model which included three regressors. The firstcovariate reflected the potential contribution by the MAOA alleleson the serotonergic function (carriers of the MAOA-H variant=2;carriers of the MAOA-L variant=1) according with previouspharmacological studies (Manuck et al., 2000, 2002). Similarly,the second regressor [subjects carrying at least one copy of the
AOA-H variant on the anterior cingulate cortex (ACC) and the orbitofrontalnd z coordinates are in the MNI space (Montreal Neurological Institute). Seeel: The BOLD signal change in the peak-activated voxel (MNI: x=8, y=36,MAOA alleles on the x axis are represented as a function of the covariatests and methods for details). The graph displays that individuals carrying theen compared with carriers of the MAOA-L variant. The mean in each grouparrying the monoamine oxidase-A (MAOA) high-activity variant; MAOA-L:

1268 L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
5HTT short allele (5HTT-S)=2; subjects homozygous for the5HTT long allele (5HTT-L,L)=1] predicted the potential effect bythe 5HTT polymorphism alone (Whale et al., 2000; Reist et al.,2001). Finally, to evaluate the interaction between MAOA-H and5HTT-S alleles, we used a third predictor which was computed asthe cross-product of MAOA-H and 5HTT-S values (carriers ofboth MAOA-H and 5HTT-S=4; carriers of both MAOA-H and5HTT-L,L=2; carriers of both MAOA-L and 5HTT-S=2; carriersof both MAOA-L and 5HTT-L,L=1).
Thus, subjects simultaneously carrying the MAOA-H alleleand at least one copy of the 5HTT-S allele were aggregated in a“hypo-serotonergic” group (presumed lowest serotonergic respon-sivity in vivo) while subjects homozygous for the 5HTT-L allele(carrying two long alleles) simultaneously carrying the MAOA-Lallele were grouped in a “hyper-serotonergic” group (presumedgreatest serotonergic responsivity in vivo). Any other combinationof MAOA and 5HTT alleles [i.e. carriers of the MAOA-H variantalso homozygous for the 5HTT-L allele (5HTT-L,L) or subjectscarrying the MAOA-L variant and at least one copy of the 5HTT-S allele] was hypothesized to result in an intermediate serotonergicfunction.
The multiple regression model was run and contrast images weregenerated for the individual main effect ofMAOA-H (first regressor:1 0 0), for the main effect of 5HTT-S (second regressor: 0 1 0) andfinally, for the interaction between MAOA-H and 5HTT-S (thirdregressor: 0 0 1). These analyses were thresholded at pb .05, FWE,small volume correction (svc) (Worsley et al., 1996) within regionsof interest (ROIs) (i.e. the OFC and the ACC defined by using theWake Forest University PICKATLAS templates).
An ROI approach was also used to test for any correlationbetween trait impulsivity and BOLD signal changes extracted fromthe ACC (the region in which we identified the MAOA-H×5HTT-Sallelic interaction; see Results) resulting from the contrast nogo vs.go. A regression analysis was performed (Spearman's r) to test forpositive or negative correlations between ACC response and traitimpulsivity.
Table 3Individual and interactive main effects of the MAOA-H and of the 5HTT-Salleles on the brain BOLD responses
Cerebral region Side Brodmannarea (BA)
p FWE
(svc)zvalue
MNIcoordinates(mm)
x y z
Individual main effect of the MAOA-H alleleVentral anteriorcingulate cortex (ACC)⁎
L 32 0.009 4.23 −8 36 −8R 0.037 3.83 6 40 −10
Orbitofrontal cortex(OFC)⁎
R 47 0.034 3.84 42 28 −10
Individual main effect of the 5HTT-S alleleVentral anteriorcingulate cortex (ACC)⁎
R 32 0.044 3.95 2 28 −6
Interactive main effect of the MAOA-H×5HTT-S allelesVentral anteriorcingulate cortex (ACC)⁎
R 32 0.001 4.76 4 40 −10
MAOA-H: monoamine oxidase-A high-activity variant; 5HTT-S: seroto-nergic transporter short allele (individuals carrying at least on copy of thisallele); MNI: Montreal Neurological Institute; p FWE: p value family-wiseerror, small volume correction (svc) in the regions of interest (ROI)⁎ (seeSubjects and methods for details).
Results
The number of subjects for each genotype group, demographicdata (age, educational level and IQ), trait impulsivity (as assessedby BIS-11 total score) and behavioural performances during thefMRI task are summarized in Table 1. In particular, the repeatedmeasures of ANOVA did not show any statistically significantdifference in all variables analyzed.
Consistent with previous reports (Liddle et al., 2001; Garavanet al., 2003), the main effect of the response inhibition in allsubjects, independently of their MAOA or 5HTT genotype,mapped on a distributed cortico-subcortical circuitry (Table 2).
The individual main effect of MAOA-H variant mapped bothon the OFC and on the ACC (Fig. 1 and Table 3), while the 5HTT-S allele affected only the ACC (Fig. 2 and Table 3).
The multiple regression model evaluating the allele–allele inter-action showed that subjects carrying simultaneously the MAOA-Hvariant and at least one copy of the 5HTT-S allele presented thehighest BOLD activity in the ACC (Fig. 3, top panel; Table 3).Conversely, the lowest ACC BOLD response was displayed bysubjects homozygous for the 5HTT-L allele simultaneously carryingthe MAOA-L allele (Fig. 3, bottom panel). Carriers of any othercombination ofMAOA and 5HTTalleles (MAOA-H/5HTT-L,L andMAOA-L/5HTT-S) displayed an intermediate activity (Fig. 3, bot-tom panel).
Finally, the correlation analysis between the ACC BOLDresponse and trait impulsivity did not show statistical significance,but only a negative trend (r=− .31, p=.73).
Discussion
This study provides new evidence that a significant interactionbetween MAOA-H and 5HTT-S impacts on the response ofanterior cingulate cortex (ACC), as assessed by fMRI in a sampleof healthy males. The highest BOLD activation in the ACC wasfound in individuals simultaneously carrying the MAOA high-activity variant (MAOA-H) and at least a copy of the 5HTT shortallele (5HTT-S), both responsible of reduced serotonergic functionin vivo (Manuck et al., 2000, 2002; Whale et al., 2000; Reist et al.,2001; Smith et al., 2004). On the contrary, subjects homozygousfor the 5HTT long allele (5HTT-L) also carrying the MAOA low-activity variant (MAOA-L) displayed the lowest BOLD responsein the ACC. All other subjects presented an intermediate activation(Fig. 3).
The individual main effect of the MAOA-H variant modulatedboth the orbitofrontal cortex (OFC) and the anterior cingulatecortex (ACC) responses, while subjects carrying at least one copyof the 5HTT-S allele presented an effect only in the ACC. This mayreflect the broader action of MAOA enzyme compared to the 5-HTtransporter on the inactivation of other amines (i.e. noradrenalineand, to a lesser extent, dopamine) that play a critical role incognitive control and in regulating the impulsive and aggressivebehaviour (Arnsten and Li, 2005; Congdon and Canli, 2005; vanGaalen et al., 2006). It has also been proposed (Rubia et al., 2003;Mathalon et al., 2003; Garavan et al., 2003) that OFC and ACCmediate segregated neural computations (e.g. response inhibitionby the OFC as opposed to error monitoring and conflict resolutionby the ACC) and that different neurochemical functions (i.e.serotonergic and noradrenergic) are dissociable in cognitive control(Chamberlain et al., 2006). Furthermore, while behavioural studiesfailed to find an effect of the acute tryptophan depletion (ATD) on
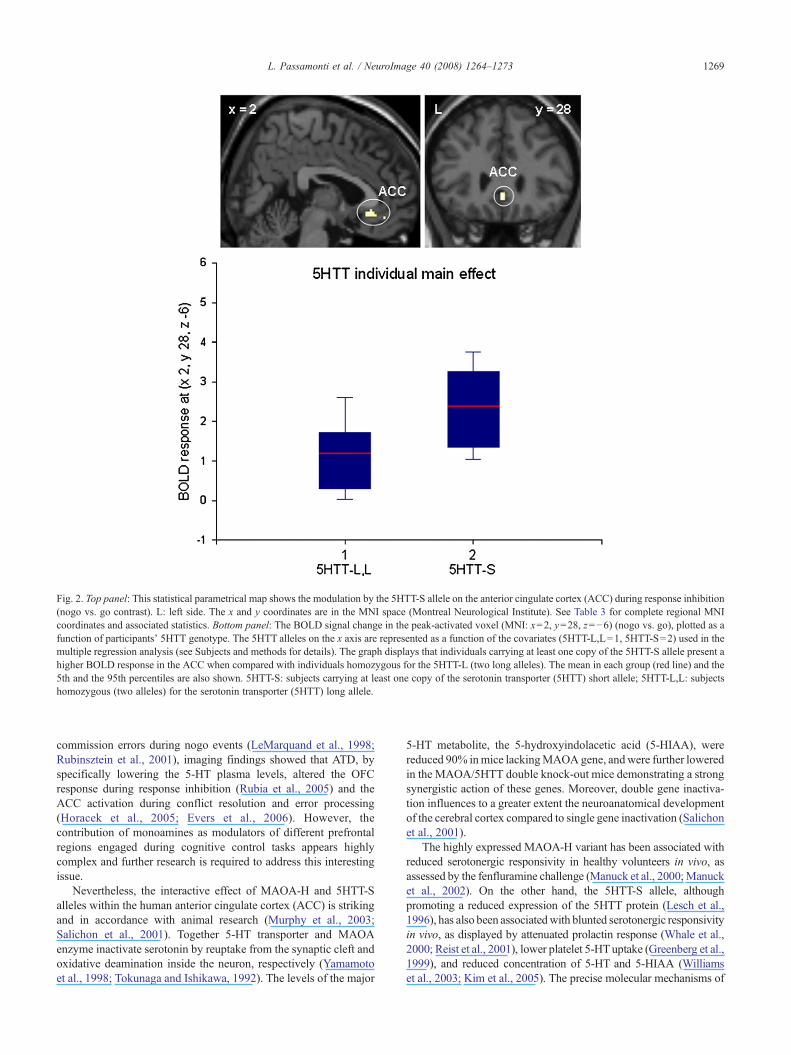
Fig. 2. Top panel: This statistical parametrical map shows the modulation by the 5HTT-S allele on the anterior cingulate cortex (ACC) during response inhibition(nogo vs. go contrast). L: left side. The x and y coordinates are in the MNI space (Montreal Neurological Institute). See Table 3 for complete regional MNIcoordinates and associated statistics. Bottom panel: The BOLD signal change in the peak-activated voxel (MNI: x=2, y=28, z=−6) (nogo vs. go), plotted as afunction of participants' 5HTT genotype. The 5HTT alleles on the x axis are represented as a function of the covariates (5HTT-L,L=1, 5HTT-S=2) used in themultiple regression analysis (see Subjects and methods for details). The graph displays that individuals carrying at least one copy of the 5HTT-S allele present ahigher BOLD response in the ACC when compared with individuals homozygous for the 5HTT-L (two long alleles). The mean in each group (red line) and the5th and the 95th percentiles are also shown. 5HTT-S: subjects carrying at least one copy of the serotonin transporter (5HTT) short allele; 5HTT-L,L: subjectshomozygous (two alleles) for the serotonin transporter (5HTT) long allele.
1269L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
commission errors during nogo events (LeMarquand et al., 1998;Rubinsztein et al., 2001), imaging findings showed that ATD, byspecifically lowering the 5-HT plasma levels, altered the OFCresponse during response inhibition (Rubia et al., 2005) and theACC activation during conflict resolution and error processing(Horacek et al., 2005; Evers et al., 2006). However, thecontribution of monoamines as modulators of different prefrontalregions engaged during cognitive control tasks appears highlycomplex and further research is required to address this interestingissue.
Nevertheless, the interactive effect of MAOA-H and 5HTT-Salleles within the human anterior cingulate cortex (ACC) is strikingand in accordance with animal research (Murphy et al., 2003;Salichon et al., 2001). Together 5-HT transporter and MAOAenzyme inactivate serotonin by reuptake from the synaptic cleft andoxidative deamination inside the neuron, respectively (Yamamotoet al., 1998; Tokunaga and Ishikawa, 1992). The levels of the major
5-HT metabolite, the 5-hydroxyindolacetic acid (5-HIAA), werereduced 90% inmice lackingMAOAgene, andwere further loweredin the MAOA/5HTT double knock-out mice demonstrating a strongsynergistic action of these genes. Moreover, double gene inactiva-tion influences to a greater extent the neuroanatomical developmentof the cerebral cortex compared to single gene inactivation (Salichonet al., 2001).
The highly expressed MAOA-H variant has been associated withreduced serotonergic responsivity in healthy volunteers in vivo, asassessed by the fenfluramine challenge (Manuck et al., 2000;Manucket al., 2002). On the other hand, the 5HTT-S allele, althoughpromoting a reduced expression of the 5HTT protein (Lesch et al.,1996), has also been associatedwith blunted serotonergic responsivityin vivo, as displayed by attenuated prolactin response (Whale et al.,2000; Reist et al., 2001), lower platelet 5-HTuptake (Greenberg et al.,1999), and reduced concentration of 5-HT and 5-HIAA (Williamset al., 2003; Kim et al., 2005). The precise molecular mechanisms of
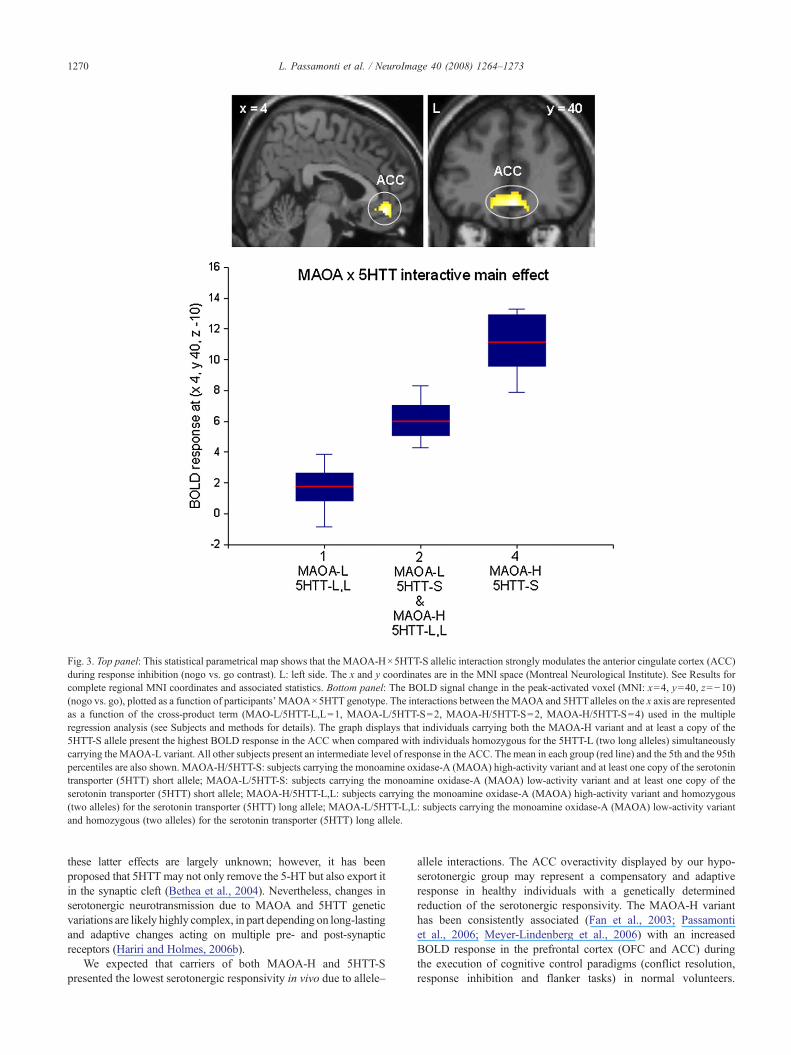
Fig. 3. Top panel: This statistical parametrical map shows that the MAOA-H×5HTT-S allelic interaction strongly modulates the anterior cingulate cortex (ACC)during response inhibition (nogo vs. go contrast). L: left side. The x and y coordinates are in the MNI space (Montreal Neurological Institute). See Results forcomplete regional MNI coordinates and associated statistics. Bottom panel: The BOLD signal change in the peak-activated voxel (MNI: x=4, y=40, z=−10)(nogo vs. go), plotted as a function of participants' MAOA×5HTT genotype. The interactions between the MAOA and 5HTTalleles on the x axis are representedas a function of the cross-product term (MAO-L/5HTT-L,L=1, MAOA-L/5HTT-S=2, MAOA-H/5HTT-S=2, MAOA-H/5HTT-S=4) used in the multipleregression analysis (see Subjects and methods for details). The graph displays that individuals carrying both the MAOA-H variant and at least a copy of the5HTT-S allele present the highest BOLD response in the ACC when compared with individuals homozygous for the 5HTT-L (two long alleles) simultaneouslycarrying the MAOA-L variant. All other subjects present an intermediate level of response in the ACC. The mean in each group (red line) and the 5th and the 95thpercentiles are also shown. MAOA-H/5HTT-S: subjects carrying the monoamine oxidase-A (MAOA) high-activity variant and at least one copy of the serotonintransporter (5HTT) short allele; MAOA-L/5HTT-S: subjects carrying the monoamine oxidase-A (MAOA) low-activity variant and at least one copy of theserotonin transporter (5HTT) short allele; MAOA-H/5HTT-L,L: subjects carrying the monoamine oxidase-A (MAOA) high-activity variant and homozygous(two alleles) for the serotonin transporter (5HTT) long allele; MAOA-L/5HTT-L,L: subjects carrying the monoamine oxidase-A (MAOA) low-activity variantand homozygous (two alleles) for the serotonin transporter (5HTT) long allele.
1270 L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
these latter effects are largely unknown; however, it has beenproposed that 5HTT may not only remove the 5-HT but also export itin the synaptic cleft (Bethea et al., 2004). Nevertheless, changes inserotonergic neurotransmission due to MAOA and 5HTT geneticvariations are likely highly complex, in part depending on long-lastingand adaptive changes acting on multiple pre- and post-synapticreceptors (Hariri and Holmes, 2006b).
We expected that carriers of both MAOA-H and 5HTT-Spresented the lowest serotonergic responsivity in vivo due to allele–
allele interactions. The ACC overactivity displayed by our hypo-serotonergic group may represent a compensatory and adaptiveresponse in healthy individuals with a genetically determinedreduction of the serotonergic responsivity. The MAOA-H varianthas been consistently associated (Fan et al., 2003; Passamontiet al., 2006; Meyer-Lindenberg et al., 2006) with an increasedBOLD response in the prefrontal cortex (OFC and ACC) duringthe execution of cognitive control paradigms (conflict resolution,response inhibition and flanker tasks) in normal volunteers.

1271L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
Moreover, previous electrophysiological findings (Fallgatter et al.,1999) demonstrated that subjects carrying at least one copy of the5HTT-S allele presented the electrical centroid associated with thenogo events significantly more anteriorly (i.e. in the prefrontalcortex) compared with individuals homozygous for the 5HTT-Lallele. Taken all together with our own results, these findings showboth an individual and an interactive effect of MAOA-H and5HTT-S in the human ACC during response inhibition.
The combinatory interplay between MAOA-H and 5HTT-S alsofits with the idea that 5-HT regulates specific local microcircuitsinside the prefrontal cortex (PFC). This region receives an exten-sive innervation from brainstem raphe nuclei and contains differentserotonergic receptors (i.e. 5-HT1A and 5-HT2A subtypes) (Barnesand Sharp, 1999). Both receptors are widely expressed in pyra-midal neurons, whereas 5HT2A receptors are mainly expressed inGABAergic interneurons. The activation of 5HT1A receptorshyperpolarizes in vitro pyramidal neurons by increasing membranepotassium conductance. On the other hand, 5-HT2A receptor acti-vation has been reported to evoke mixed responses, both excitatoryand inhibitory (Stein et al., 2007). While the stimulation of 5-HT1A
and 5-HT2A receptors has opposite effects on attentional per-formances (Winstanley et al., 2003), both receptors appear stronglyinvolved in impulsive behaviour and in the neural processes con-trolling for the suppression of premature responses (Carli et al.,2006). The higher BOLD response displayed by subjects withgenetically driven low serotonergic responsivity (i.e. MAOA-H/5HTT-S carriers) could reflect local adaptive mechanisms, actingvia 5-HT1A and 5-HT2A receptors within the PFC.
However, we did not directly measure the 5-HT inside the PFCand did not specifically investigate the actions of both 5-HT1A and5-HT2A receptors. Thus, our fMRI findings could be not neces-sarily related to the serotonergic neurotransmission. More complexand long-acting molecular mechanisms could be involved, as 5-HThas been highly implicated in development and differentiation ofneurons (Encinas et al., 2006; Malberg et al., 2000). Moreover, wetested for correlations between ACC response and trait impulsivity,but we only found a non-significant negative trend. This maydepend on the small sample used in the present study and it needsfurther investigations.
Nevertheless, the current findings contribute to validatingmodels studying the impact of allele–allele interactions on neuro-anatomical substrates of cognitive functions. Haplotype modelsstill receive little exploitation, but they could provide new insightsinto the neurobiology of complex brain functions, with clearimplications for mental health.
Acknowledgments
We thank Giuseppina Morganti and Andrea Fossati forcontributions to early drafts of the manuscript.
References
American Psychiatric Association, 1994. Diagnostic and Statistical Manual ofMental Disorders, 4th ed. American Psychiatric Press, Washington, DC.
Arnsten, A.F., Li, B.M., 2005. Neurobiology of executive functions:catecholamine influences on prefrontal cortical functions. Biol. Psychiatry57, 1377–1384.
Baca-Garcia, E., Vaquero, C., Diaz-Sastre, C., Garcia-Resa, E., Saiz-Ruiz,J., Fernandez-Piqueras, J., de Leon, J., 2004. Lack of associationbetween the serotonin transporter promoter gene polymorphism and
impulsivity or aggressive behavior among suicide attempters and healthyvolunteers. Psychiatry Res. 126, 99–106.
Barnes, N.M., Sharp, T., 1999. A review of central 5-HT receptors and theirfunction. Neuropharmacology 38, 1083–1152.
Bertolino, A., Arciero, G., Rubino, V., Latorre, V., De Candia, M., Mazzola,V., Blasi, G., Caforio, G., Hariri, A., Kolachana, B., Nardini, M.,Weinberger, D.R., Scarabino, T., 2005. Variation of human amygdalaresponse during threatening stimuli as a function of 5′HTTLPRgenotype and personality style. Biol. Psychiatry 57, 1517–1525.
Bethea, C.L., Streicher, J.M., Coleman, K., Pau, F.K., Moessner, R.,Cameron, J.L., 2004. Anxious behavior and fenfluramine-inducedprolactin secretion in young rhesus macaques with different alleles ofthe serotonin reuptake transporter polymorphism (5HTTLPR). Behav.Genet. 34, 295–307.
Carli, M., Baviera, M., Invernizzi, R.W., Balducci, C., 2006. Dissociablecontribution of 5-HT1A and 5-HT2A receptors in the medial prefrontalcortex to different aspects of executive control such as impulsivity andcompulsive perseveration in rats. Neuropsychopharmacology 31,757–767.
Carrel, L., Cottle, A.A., Goglin, K.C., Willard, H.F., 1999. A first-generationX-inactivation profile of the human X chromosome. Proc. Natl. Acad.Sci. U. S. A. 96, 14440–14444.
Caspi, A., McClay, J., Moffitt, T.E., Mill, J., Martin, J., Craig, I.W., Taylor,A., Poulton, R., 2002. Role of genotype in the cycle of violence inmaltreated children. Science 297, 851–854.
Caspi, A., Sugden, K., Moffitt, T.E., Taylor, A., Craig, I.W., Harrington, H.,McClay, J., Mill, J., Martin, J., Braithwaite, A., Poulton, R., 2003.Influence of life stress on depression: moderation by a polymorphism inthe 5-HTT gene. Science 301, 386–389.
Chamberlain, S.R., Muller, U., Blackwell, A.D., Clark, L., Robbins, T.W.,Sahakian, B.J., 2006. Neurochemical modulation of response inhibitionand probabilistic learning in humans. Science 311, 861–863.
Clarke, H.F., Dalley, J.W., Crofts, H.S., Robbins, T.W., Roberts, A.C., 2004.Cognitive inflexibility after prefrontal serotonin depletion. Science 304,878–880.
Congdon, E., Canli, T., 2005. The endophenotype of impulsivity: reachingconsilience through behavioral, genetic, and neuroimaging approaches.Behav. Cogn. Neurosci. Rev. 4, 262–281.
Courtet, P., Picot, M.C., Bellivier, F., Torres, S., Jollant, F., Michelon, C.,Castelnau, D., Astruc, B., Buresi, C., Malafosse, A., 2004. Serotonintransporter gene may be involved in short-term risk of subsequentsuicide attempts. Biol. Psychiatry 55, 46–51.
Davidson, R.J., Putnam, K.M., Larson, C.L., 2000. Dysfunction in theneural circuitry of emotion regulation—a possible prelude to violence.Science 289, 591–594.
Denney, R.M., Koch, H., Craig, I.W., 1999. Association between mono-amine oxidase A activity in human male skin fibroblasts and genotype ofthe MAOA promoter-associated variable number tandem repeat. Hum.Genet. 105, 542–551.
Encinas, J.M., Vaahtokari, A., Enikolopov, G., 2006. Fluoxetine targetsearly progenitor cells in the adult brain. Proc. Natl. Acad. Sci. U. S. A.103, 8233–8238.
Evers, E.A., van der Veen, F.M., Jolles, J., Deutz, N.E., Schmitt, J.A., 2006.Acute tryptophan depletion improves performance and modulates theBOLD response during a Stroop task in healthy females. Neuroimage32, 248–255.
Fallgatter, A.J., Jatzke, S., Bartsch, A.J., Hamelbeck, B., Lesch, K.P., 1999.Serotonin transporter promoter polymorphism influences topography ofinhibitory motor control. Int. J. Neuropsychopharmcol. 2, 115–120.
Fan, J., Fossella, J., Sommer, T., Wu, Y., Posner, M.I., 2003. Mapping thegenetic variation of executive attention onto brain activity. Proc. Natl.Acad. Sci. U. S. A. 100, 7406–7411.
Fossati, A., Di Ceglie, A., Acquarini, E., Barratt, E.S., 2001. Psychometricproperties of an Italian version of the Barratt Impulsiveness Scale–11(BIS-11) in nonclinical subjects. J. Clin. Psychol. 57, 815–828.
Fowler, J.S., MacGregor, R.R., Wolf, A.P., Arnett, C.D., Dewey, S.L.,Schlyer, D., Christman, D., Logan, J., Smith, M., Sachs, H., et al., 1987.

1272 L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
Mapping human brain monoamine oxidase A and B with 11C-labeledsuicide inactivators and PET. Science 235, 481–485.
Frankle, W.G., Lombardo, I., New, A.S., Goodman, M., Talbot, P.S., Huang,Y., Hwang, D.R., Slifstein, M., Curry, S., Abi-Dargham, A., Laruelle,M., Siever, L.J., 2005. Brain serotonin transporter distribution insubjects with impulsive aggressivity: a positron emission study with[11C]McN 5652. Am. J. Psychiatry 162, 915–923.
Friston, K.J., Holmes, A.P., Worsley, K.J., Poline, J.B., Frith, C.D.,Frackowiak, R.S.J., 1995. Statistical parametric maps in functionalimaging: a general approach. Hum. Brain Mapp. 2, 189–210.
Garavan, H., Ross, T.J., Kaufman, J., Stein, E.A., 2003. A midline disso-ciation between error-processing and response-conflict monitoring.Neuroimage 20, 1132–1139.
Gerra, G., Garofano, L., Santoro, G., Bosari, S., Pellegrini, C., Zaimovic, A.,Moi, G., Bussandri, M., Moi, A., Brambilla, F., Donnini, C., 2004.Association between low-activity serotonin transporter genotype andheroin dependence: behavioral and personality correlates. Am. J. Med.Genet. B Neuropsychiatr. Genet. 126, 37–42.
Greenberg, B.D., Tolliver, T.J., Huang, S.J., Li, Q., Bengel, D., Murphy,D.L., 1999. Genetic variation in the serotonin transporter promoterregion affects serotonin uptake in human blood platelets. Am. J. Med.Genet. 88, 83–87.
Hariri, A.R., Mattay, V.S., Tessitore, A., Kolachana, B., Fera, F., Goldman, D.,Egan, M.F., Weinberger, D.R., 2002. Serotonin transporter geneticvariation and the response of the human amygdala. Science 297, 400–403.
Hariri, A.R., Drabant, E.M., Weinberger, D.R., 2006a. Imaging genetics:perspectives from studies of genetically driven variation in serotoninfunction and corticolimbic affective processing. Biol. Psychiatry 59,888–897.
Hariri, A.R., Holmes, A., 2006b. Genetics of emotional regulation: the roleof the serotonin transporter in neural function. Trends Cogn. Sci. 10,182–191.
Heinz, A., Jones, D.W., Mazzanti, C., Goldman, D., Ragan, P., Hommer, D.,Linnoila, M., Weinberger, D.R., 2000. A relationship between serotonintransporter genotype and in vivo protein expression and alcoholneurotoxicity. Biol. Psychiatry 47, 643–649.
Holmes, A., Murphy, D.L., Crawley, J.N., 2002. Reduced aggression in micelacking the serotonin transporter. Psychopharmacology (Berl) 161,160–167.
Horacek, J., Zavesicka, L., Tintera, J., Dockery, C., Platilova, V., Kopecek, M.,Spaniel, F., Bubenikova, V., Hoschl, C., 2005. The effect of tryptophandepletion on brain activation measured by functional magnetic resonanceimaging during the Stroop test in healthy subjects. Physiol. Res. 54,235–244.
Kim, D.K., Tolliver, T.J., Huang, S.J., Martin, B.J., Andrews, A.M.,Wichems, C., Holmes, A., Lesch, K.P., Murphy, D.L., 2005. Alteredserotonin synthesis, turnover and dynamic regulation in multiple brainregions of mice lacking the serotonin transporter. Neuropharmacology49, 798–810.
LeMarquand, D.G., Pihl, R.O., Young, S.N., Tremblay, R.E., Seguin, J.R.,Palmour, R.M., Benkelfat, C., 1998. Tryptophan depletion, executivefunctions, and disinhibition in aggressive, adolescent males. Neuropsy-chopharmacology 19, 333–341.
Lesch, K.P., Bengel, D., Heils, A., Sabol, S.Z., Greenberg, B.D., Petri, S.,Benjamin, J., Muller, C.R., Hamer, D.H., Murphy, D.L., 1996. Asso-ciation of anxiety-related traits with a polymorphism in the serotonintransporter gene regulatory region. Science 274, 1527–1531.
Liddle, P.F., Kiehl, K.A., Smith, A.M., 2001. Event-related fMRI study ofresponse inhibition. Hum. Brain Mapp. 12, 100–109.
Little, K.Y., McLaughlin, D.P., Zhang, L., Livermore, C.S., Dalack, G.W.,McFinton, P.R., DelProposto, Z.S., Hill, E., Cassin, B.J., Watson, S.J.,Cook, E.H., 1998. Cocaine, ethanol, and genotype effects on humanmidbrain serotonin transporter binding sites and mRNA levels. Am. J.Psychiatry 155, 207–213.
Loeber, R., Keenan, K., 1994. Interaction between conduct disorder and itscomorbid conditions: Effects of age and gender. Clin. Psychol. 14,497–523.
Malberg, J.E., Eisch, A.J., Nestler, E.J., Duman, R.S., 2000. Chronicantidepressant treatment increases neurogenesis in adult rat hippocam-pus. J. Neurosci. 20, 9104–9110.
Manuck, S.B., Flory, J.D., Ferrell, R.E., Mann, J.J., Muldoon, M.F., 2000. Aregulatory polymorphism of the monoamine oxidase-A gene may beassociated with variability in aggression, impulsivity, and centralnervous system serotonergic responsivity. Psychiatry Res. 95, 9–23.
Manuck, S.B., Flory, J.D., Muldoon, M.F., Ferrell, R.E., 2002. Centralnervous system serotonergic responsivity and aggressive disposition inmen. Physiol. Behav. 77, 705–709.
Mathalon, D.H., Whitfield, S.L., Ford, J.M., 2003. Anatomy of an error:ERP and fMRI. Biol. Psychol. 64, 119–141.
Meyer-Lindenberg, A., Buckholtz, J.W., Kolachana, B., Hariri, A.R.,Pezawas, L., Blasi, G., Wabnitz, A., Honea, R., Verchinski, B., Callicott,J.H., Egan, M., Mattay, V., Weinberger, D.R., 2006. Neural mechanismsof genetic risk for impulsivity and violence in humans. Proc. Natl. Acad.Sci. U. S. A. 103, 6269–6274.
Mobini, S., Chiang, T.J., Al-Ruwaitea, A.S., Ho, M.Y., Bradshaw, C.M.,Szabadi, E., 2000. Effect of central 5-hydroxytryptamine depletion oninter-temporal choice: a quantitative analysis. Psychopharmacology(Berl) 149, 313–318.
Murphy, D.L., Uhl, G.R., Holmes, A., Ren-Patterson, R., Hall, F.S., Sora, I.,Detera-Wadleigh, S., Lesch, K.P., 2003. Experimental gene interactionstudies with SERT mutant mice as models for human polygenic andepistatic traits and disorders. Genes Brain Behav. 2, 350–364.
Noblett, K.L., Coccaro, E.F., 2005. Molecular genetics of personality. Curr.Psychiatry Rep. 7, 73–80.
Park, S.B., Coull, J.T., McShane, R.H., Young, A.H., Sahakian, B.J., Robbins,T.W., Cowen, P.J., 1994. Tryptophan depletion in normal volunteersproduces selective impairments in learning and memory. Neuropharma-cology 33, 575–588.
Passamonti, L., Fera, F.,Magariello, A., Cerasa, A., Gioia,M.C.,Muglia, M.,Nicoletti, G., Gallo, O., Provinciali, L., Quattrone, A., 2006. Monoamineoxidase-a genetic variations influence brain activity associated withinhibitory control: new insight into the neural correlates of impulsivity.Biol. Psychiatry 59, 334–340.
Reist, C., Mazzanti, C., Vu, R., Tran, D., Goldman, D., 2001. Sero-tonin transporter promoter polymorphism is associated with atten-uated prolactin response to fenfluramine. Am. J. Med. Genet. 105,363–368.
Robbins, T.W., 2005. Chemistry of the mind: neurochemical modulation ofprefrontal cortical function. J. Comp. Neurol. 493, 140–146.
Rubia, K., Smith, A.B., Brammer, M.J., Taylor, E., 2003. Right inferiorprefrontal cortex mediates response inhibition while mesial prefrontalcortex is responsible for error detection. Neuroimage 20, 351–358.
Rubia, K., Lee, F., Cleare, A.J., Tunstall, N., Fu, C.H., Brammer, M.,McGuire, P., 2005. Tryptophan depletion reduces right inferiorprefrontal activation during response inhibition in fast, event-relatedfMRI. Psychopharmacology (Berl) 179, 791–803.
Rubinsztein, J.S., Rogers, R.D., Riedel, W.J., Mehta, M.A., Robbins, T.W.,Sahakian, B.J., 2001. Acute dietary tryptophan depletion impairsmaintenance of “affective set” and delayed visual recognition in healthyvolunteers. Psychopharmacology (Berl) 154, 319–326.
Sabol, S.Z., Hu, S., Hamer, D., 1998. A functional polymorphism in themonoamine oxidase A gene promoter. Hum. Genet. 103, 273–279.
Salichon, N., Gaspar, P., Upton, A.L., Picaud, S., Hanoun, N., Hamon, M.,De Maeyer, E., Murphy, D.L., Mossner, R., Lesch, K.P., Hen, R., Seif, I.,2001. Excessive activation of serotonin (5-HT) 1B receptors disrupts theformation of sensory maps in monoamine oxidase A and 5-HTtransporter knock-out mice. J. Neurosci. 21, 884–896.
Smith, G.S., Lotrich, F.E., Malhotra, A.K., Lee, A.T., Ma, Y., Kramer, E.,Gregersen, P.K., Eidelberg, D., Pollock, B.G., 2004. Effects of serotonintransporter promoter polymorphisms on serotonin function. Neuropsy-chopharmacology 29, 2226–2234.
Stein,D.J., Hemmings, S.,Moolman-Smook,H.,Audenaert, K., 2007. 5-HT2A:its role in frontally mediated executive function and related psychopathol-ogy. CNS Spectr. 12, 512–516.

1273L. Passamonti et al. / NeuroImage 40 (2008) 1264–1273
Tokunaga, K., Ishikawa, K., 1992. Evidence for different sites ofmonoamine oxidase and catechol-O-methyltransferase in the striatum.Brain Res. 575, 164–166.
van Gaalen, M.M., van Koten, R., Schoffelmeer, A.N., Vanderschuren, L.J.,2006. Critical involvement of dopaminergic neurotransmission inimpulsive decision making. Biol. Psychiatry 60, 66–73.
Varnas, K., Halldin, C., Hall, H., 2004. Autoradiographic distribution ofserotonin transporters and receptor subtypes in human brain. Hum. BrainMapp. 22, 246–260.
Wechsler, D., 1981. Manual for the Wechsler Adult Intelligence Scale–Revised. [Italian version: WAIS-R Wechsler Adult Intelligence Scale–Revised. Manuale. Firenze: O.S. Organizzazioni Speciali], 1997.Psychological Corporation, New York.
Wellman, C.L., Izquierdo, A., Garrett, J.E., Martin, K.P., Carroll, J.,Millstein, R., Lesch, K.P., Murphy, D.L., Holmes, A., 2007. Impairedstress-coping and fear extinction and abnormal corticolimbic morphol-ogy in serotonin transporter knock-out mice. J. Neurosci. 27, 684–691.
Whale, R., Quested, D.J., Laver, D., Harrison, P.J., Cowen, P.J., 2000.Serotonin transporter (5-HTT) promoter genotype may influence the
prolactin response to clomipramine. Psychopharmacology (Berl) 150,120–122.
Williams, R.B., Marchuk, D.A., Gadde, K.M., Barefoot, J.C., Grichnik, K.,Helms, M.J., Kuhn, C.M., Lewis, J.G., Schanberg, S.M., Stafford-Smith,M., Suarez, E.C., Clary, G.L., Svenson, I.K., Siegler, I.C., 2003.Serotonin-related gene polymorphisms and central nervous systemserotonin function. Neuropsychopharmacology 28, 533–541.
Winstanley, C.A., Chudasama, Y., Dalley, J.W., Theobald, D.E., Glennon, J.C.,Robbins, T.W., 2003. Intra-prefrontal 8-OH-DPAT and M100907improve visuospatial attention and decrease impulsivity on the five-choice serial reaction time task in rats. Psychopharmacology (Berl) 167,304–314.
Worsley, K.J., Marrett, S., Neelin, P., Vandal, A.C., Friston, K.J., Evans,A.C., 1996. A unified statistical approach for determining signif-icant signals in images of cerebral activation. Hum. Brain Map. 4,58–73.
Yamamoto, H., Fujimiya, M., Shirai, Y., Nakashita, M., Oyasu, M., Saito,N., 1998. Immunohistochemical localization of serotonin transporter innormal and colchicine treated rat brain. Neurosci. Res. 32, 305–312.