effects of x-irradiation on nerve growth factor in the developing mouse brain
TRANSCRIPT

Toxicology Letters 90 (1997) 35-43
Effects of X-irradiation on nerve growth factor in the developing mouse brain
Toxicology Letters
Yvonne Dimberg”,*, Marcel0 Vazquez b, Stine SiiderstrGm”, Ted Ebendal”
“Department #of Radioecology, Swedish University of Agricultural Sciences, Box 7031, S-750 07 Uppsala, Sweden bEye Radiation and Environmental Research Laboratory, College of Surgeons and Physicians, Columbia University,
New York, USA “Department of Developmental Neuroscience, Uppsala University, Biomedical Center, Box 587, S-751 23 Uppsala, Sweden
Received 26 April 1996; revised 28 August 1996; accepted 2 September 1996
Abstract
The involvement of neurotrophins after radiation injury during brain development were studied in pregnant mice (C 57/Bl) exposed on gestation day 15 to X-ray doses of 0.02-2 Gy. Nerve growth factor protein (NGF) and different cholinergic markers were investigated on postnatal day 1 (Pl) and day 21 (P21); in situ hybridization with brain-derived neurotrophic factor (BDNF) and t&C (receptor serving to bind neurotrophin-3) probes was investi- gated on P21 in cortex, hippocampus, septum and cerebellum. The level of NGF protein was increased in irradiated forebrain on Pl in a dose-related manner. However, on P21 the NGF protein dropped down below the control levels in irradiated hippocampus and cerebellum. The response of acetylcholine esterase (AChE) activity in cerebellum at P21 was correlated with the changes in the amount of NGF. The intensity of cell labelling with trkC probe decreased after irradiation in the region of the hippocampus at P21, especially in dentate gyrus. The expression of BDNF mRNA was increased at P21 by low doses of irradiation (0.02-l Gy) but was decreased by a high dose (2 Gy) in the same area. Thus, the radiation induced an alteration of neurotrophins, and the changes varied depending on the dose or time after irradiation. Such alterations in the pattern of growth factor production may modulate the response of cells to radiation. Furthermore, NGF protein levels and the expression of BDNF and trkC mRNA were affected by radiation doses as low as 0.02 Gy, indicating that during development the neurotrophins and their receptors are very sensitive to radiation. Copyright 0 1997 Elsevier Science Ireland Ltd.
Keywords: Nerve gr’owth factor; Brain-derived neurotrophic factor; trkC; Radiation; In situ hybridization
* Corresponding author, Tel.: + 46 18 671280; fax: + 46 18 672886.
0378-4274/97/$17.00 Copyright 0 1997 Elsevier Science Ireland Ltd. All rights reserved
PII SO378-4274(96 103827-l

36 Y. Dimberg et al. / Toxicology Letters 90 (1997) 35-43
1. Introduction
The developing CNS is highly sensitive to low doses of certain chemical and physical agents such as ionizing radiation. Mechanisms at different levels have been discussed in many papers [for review see; [1,2]]. However, there is little informa- tion concerning the signaling transduction path- ways which may be involved in repair of radiation lesions in eukaryotic cells. Some recent discoveries that radiation induces the expression of several genes which can modulate the radiation response in mammalian cells point in this direction. For example, ionizing radiation has been demonstrated to induce or activate certain proteins that protect the cell against radiation stress [3,4]. Growth fac- tors are specific proteins that also have been demonstrated to be released after radiation expo- sure [5-71 and they may be involved in regulating radiation damage repair in cells. It has been pro- posed that radiation-induced basic fibroblast growth factor (bFGF) can stimulate potentially lethal damage repair pathway and this process can lead to recovery of cells from radiation lesions [8,9]. Furthermore, other growth factors, e.g., platelet-derived like growth factor and transform- ing growth factor, have been shown to be released from cells by radiation in a dose-related manner [lo-121. In cultured brain cells, radiation induced alterations in nerve growth factor (NGF) mRNA. The neurotrophic interactions mediated by changes induced in levels of neurotrophic factors may help to restore the balance disrupted by the radiation treatment [ 131.
In the developing central nervous system moder- ate and low doses of methyl mercury and radiation selectively affect certain areas, e.g., hippocampus, dentate gyrus, cerebellum and cortex [14- 161. It is still not clear how structural and behavioural changes induced by heavy metals and radiation are related to the NGF-dependent cholinergic system in the hippocampus-basal forebrain axis. Nerve growth factor plays a crucial role in the mainte- nance of cholinergic neurons in the basal forebrain during development and adulthood [17]. An alter- ation in the level of NGF could hypothetically lead to neurodegeneration or to a failure in the process of neural plasticity. It has been shown that beside
NGF, the hippocampus contains brain derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3), which influence the function of different types of nerve cells [18,19]. The local release of these growth factors can probably influence nerve cells in the hippocampus.
The aim of this study was to establish how low doses of y-radiation affect the development of the hippocampal-basal forebrain axis of the fetal mouse and neural plasticity in these areas and some involved neurotrophins (NGF, BDNF and NT-3 receptor). Using morphological and im- munochemical methods we report changes at the structural and biochemical levels.
2. Materials and methods
2.1. Animals
C57/Bl females were mated by being placed with males (two females to every male) overnight. The next morning they were examined to confirm the presence of vaginal plugs, and mated females were regarded as being in day 0 of pregnancy.
Pregnant mice were exposed to 0, 0.02, 0.1, 1 or 2 Gy of X-radiation on gestation day 15. The animals were housed individually under normal laboratory conditions at the animal facility of the Department of Pathology, Swedish University of Agricultural Sciences. A 12:12 h light-dark cycle was maintained, and food and drinking solutions were available for ad libitum consumption. The control groups were handled in a similar way. All the animals were weaned on P2 1. For the analysis animals of both sexes were randomly taken from different litter. On Pl, due to small volume, only forebrains were investigated. Cortex, hippocam- pus, septum and cerebellum were investigated on P21. The number of animal used was 8-26 (see also the figure text).
2.2. Irradiation
The irradiation was performed with a Muller MG-300 Roentgen machine operated at 260 kV and 11 mA and with an inherent filtration of 4 mm Al. The distance between the tube and the target was approximately 60 cm or 120 cm for the

Y. Dimberg et al. / Toxicology Letters 90 (1997) 35-43 31
low-dose experim,ent, resulting in a dose rate of 0.46 Gy/min or 0.1 Gy/min.
2.3. Enzyme assaJ)s
Acetylcholine esterase (AChE) was assayed ac- cording to the procedure of Ellman et al. [20]. Homogenate (50 ~1) and 10 mM dithio-bis(ni- trobenzoic acid) (15 ~1) were added to 1 ml of 0.1 M potassium phosphate buffer, pH 8.0. The reac- tion was started by adding 15 ~1 of 75 mM acetylthiocholine iodide and followed in a Shi- madzu UV-160 spectrophotometer at 25°C for 3-5 min (Ed12= 1.3600 M-‘.cn-‘).
Choline acetyltransferase (ChAT) was assayed according to the method of Wilson et al. [21], measuring the “C-labelled acetylcholine formed in a 50-~1 reaction mixture containing 0.2 mM [l-i4C] acetyl-CoA, (5.0 mCi/mmol), 20 mM choline chloride, 0.1 mM neostigmine methyl- sulphonate, 200 mM NaCl, 0.5% Triton X-100, 50 mM potassium phosphate (pH 6.8) EDTA and 20 ~1 of homogenate. The reaction mixtures were incubated for 20 min at 37°C and thereafter stopped by adding 0.5 ml ice-cold H,O. The mix- ture was passed through an ion exchange column (Dowex AG l-x8 [chloride form] packed in a Pasteur pipette, bed volume 0.5 ml); the tubes were washed with, two 0.5 ml portions of ice-cold H,O, and the washes were also passed through the column. Column effluents were collected in scintillation vials, whereafter 4 ml of Quickszint 1 Zinnser Analytic scintillation mixture was added, and radioactivity was measured in a Packard Tri- carb 1900A scintillation counter. The blank reac- tion for each sample was assayed with choline chloride omitted.
DNA was measured fluorimetrically in a Kon- tron SFM 25 spectrofluorometer, as described by Hinegardner [22], using the reaction of di- aminobenzoic acid with the deoxyribose residues. Protein content was determined by the method of Lowry et al. [23].
2.4. Measurement of NGF levels using enzyme immunoassay (EI.4)
The brains of pups sacrificed on Pl and P21
were dissected to obtain the cerebellum, hippocampus, septum and cortex in order to de- termine NGF levels. Samples were frozen and weighed prior to homogenizing them in 10 vol- umes of cold 0.02 M Tris-HCl (pH 7.4) and 0.5 M NaCl (TBS) containing 0.5% Tween 20, 2 mg/ml bovine serum albumin (BSA, USB), 10 mM EDTA, aprotinin (Sigma; 20 kallikerein units/ml), and 0.1 mM phenylmethyl sulfonyl fluoride (Sigma; PMSF). The samples were stored at - 20°C. The enzyme immunoassay for determina- tion of NGF has been described in detail elsewhere [24]. Briefly, the brain samples were centrifuged for 60 min at 1660 x g at 4°C. The supernatant was added to a black 96-microwell immunoplate precoated with mab-anti mouse p- NGF antibody 27/21 [25] and incubated overnight at 4°C. The wells were washed with TBS before incubation with the 27/21-P-galactosidase conju- gate. Purified mouse /I-NGF was added in known amounts to parallel wells and served as a stan- dard. The results were read in a microfluorometer after adding the substrate 4-methylumbelliferyl-B - galactoside to the wells.
2.5. In situ hybridization histochemistry
Entire brains were frozen on dry ice and kept at - 80°C until sectioned. Coronal sections (10 pm) of fresh-frozen brains from Pl and P21 mice (C57/Bl) were cut at - 20°C on a cryostat (Leitz, FRG). The sections were thawed on slides pre- treated with poly-L-lysine (50 pg/ml, Sigma, St. Louis, MO, USA), fixed in 4% paraformaldehyde at 4°C for 30 min and rinsed twice in PBS. Dehydration was carried out in a graded ethanol series and included a 5 min incubation in chloro- form. Slides were air-dried and stored at - 20°C until hybridized. Oligonucleotides were provided by Scandinavian Gene Synthesis, Koping, to de- tect the specific mRNA in the sections. The fol- lowing oligonucleotide probes were use for the present study: Rat BDNF oligoprobe TE35 5’- TTG GCC TTT TGA TAC CGG GAC TTT CTC CAG GAC TGT GAC CGT CCC ACC-3’, a 48-mer corresponding to nucleotides 559-606, amino acids 33-48 in the mature brain-derived
38 Y. Dimberg et al. 1 Toxicology Letters 90 (1997) 35-43
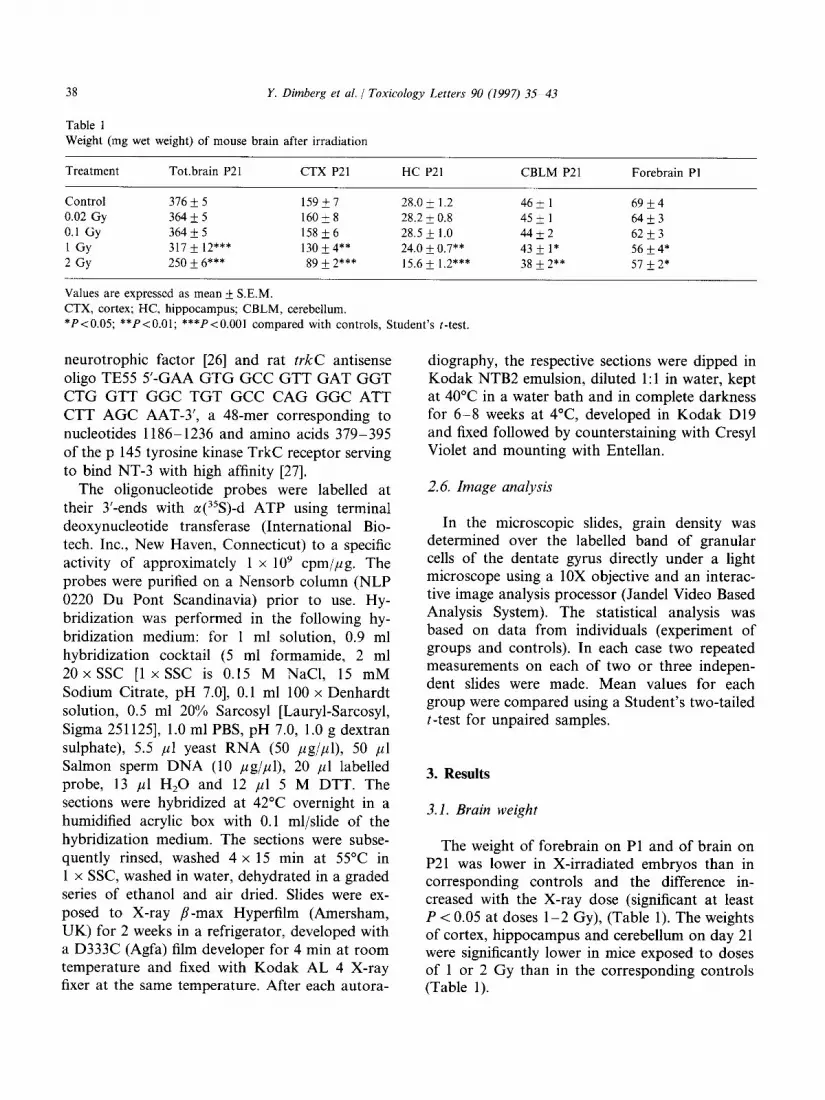
Table 1 Weight (mg wet weight) of mouse brain after irradiation
Treatment Tot.brain P21 CTX P21 HC P21 CBLM P21 Forebrain PI
Control 316 * 5 159*7 28.0 1.2 k 46 1 * 69 4 k
0.02 Gy 364*5 16Ok8 28.2 i 0.8 45 1 * 64 3 +
0.1 Gy 364 + 5 158&6 28.5 1.0 & 44 2 + 62 3 +
1 GY 317 k 12*** 130+4** 24.0 0.7** & 432 1* 56*4*
2 GY 250 f 6*** 89 * 2*** 15.6 k 1.2*** 38 2** * 57 2* *
Values are expressed as mean + S.E.M.
CTX, cortex; HC, hippocampus; CBLM, cerebellum.
*P<O.O5; **P<O.Ol; ***P<O.OOl compared with controls, Student’s t-test.
neurotrophic factor [26] and rat t&C antisense oligo TE55 5’-GAA GTG GCC GTT GAT GGT CTG GTT GGC TGT GCC CAG GGC ATT CTT AGC AAT-3’, a 48-mer corresponding to nucleotides 1186-1236 and amino acids 379-395 of the p 145 tyrosine kinase TrkC receptor serving to bind NT-3 with high affinity [27].
The oligonucleotide probes were labelled at their 3’-ends with a(35S)-d ATP using terminal deoxynucleotide transferase (International Bio- tech. Inc., New Haven, Connecticut) to a specific activity of approximately 1 x lo9 cpm/pg. The probes were purified on a Nensorb column (NLP 0220 Du Pont Scandinavia) prior to use. Hy- bridization was performed in the following hy- bridization medium: for 1 ml solution, 0.9 ml hybridization cocktail (5 ml formamide, 2 ml 20 x SSC [l x SSC is 0.15 M NaCl, 15 mM Sodium Citrate, pH 7.01, 0.1 ml 100 x Denhardt solution, 0.5 ml 20% Sarcosyl [Lauryl-Sarcosyl, Sigma 2511251, 1.0 ml PBS, pH 7.0, 1.0 g dextran sulphate), 5.5 ,ul yeast RNA (50 pggipl), 50 ~1 Salmon sperm DNA (10 pg/pl), 20 ~1 labelled probe, 13 ~1 H,O and 12 ~1 5 M DTT. The sections were hybridized at 42°C overnight in a humidified acrylic box with 0.1 ml/slide of the hybridization medium. The sections were subse- quently rinsed, washed 4 x 15 min at 55’C in 1 x SSC, washed in water, dehydrated in a graded series of ethanol and air dried. Slides were ex- posed to X-ray p-max Hyperfilm (Amersham, UK) for 2 weeks in a refrigerator, developed with a D333C (Agfa) film developer for 4 min at room temperature and fixed with Kodak AL 4 X-ray fixer at the same temperature. After each autora-
diography, the respective sections were dipped in Kodak NTB2 emulsion, diluted 1: 1 in water, kept at 40°C in a water bath and in complete darkness for 6-8 weeks at 4°C developed in Kodak D19 and fixed followed by counterstaining with Cresyl Violet and mounting with Entellan.
2.6. Image analysis
In the microscopic slides, grain density was determined over the labelled band of granular cells of the dentate gyrus directly under a light microscope using a 10X objective and an interac- tive image analysis processor (Jandel Video Based Analysis System). The statistical analysis was based on data from individuals (experiment of groups and controls). In each case two repeated measurements on each of two or three indepen- dent slides were made. Mean values for each group were compared using a Student’s two-tailed t-test for unpaired samples.
3. Results
3.1. Brain weight
The weight of forebrain on Pl and of brain on P21 was lower in X-irradiated embryos than in corresponding controls and the difference in- creased with the X-ray dose (significant at least P < 0.05 at doses l-2 Gy), (Table 1). The weights of cortex, hippocampus and cerebellum on day 21 were significantly lower in mice exposed to doses of 1 or 2 Gy than in the corresponding controls (Table 1).
Y. Dimberg et al. / Toxicology Letters 90 (1997) 35-43 39
Table 2
ChAT activity after irradiation (nmol/g wet weight x min)
Treatment CTXPZ 1 HC P21 SEP P21
Control 46.4 * 3.1 24.4 + 1.8 22.9 & 1.5
0.02 Gy 42.1 + 4.3 23.2 _t 1.2 21.6 k 0.7
0.1 Gy 44.3 & 3.0 24.3 + 1.4 19.8 f 0.9
1 GY 32.7 & 1.5** 22.8 + 0.4 29.8 + 2.8*
2 GY 35.9 * 0.8** 22.8 + 0.9 26.0 + 3.7
Values are expressed as mean f S.E.M.
CTX, cortex; HC, hippocampus; SEP, septum.
*P<O.O5; **PiO.Ol; ***P<O.OOl compared with controls,
Student’s t-test.
3.2. Marker enzymes
ChAT activity was decreased in cortex on P21 after irradiation at doses of l-2 Gy (P < 0.01). By contrast, in septum ChAT activity was in- creased by irradiation at 1 Gy (Table 2).
AChE activity in the forebrain of young mice on Pl was higher in those that had received l-2 Gy than in controls. Three weeks later mice that had received 1 or 2 Gy had a lower level of AChE activity in their cerebellum as compared with the controls (Table 3).
3.3. EZA
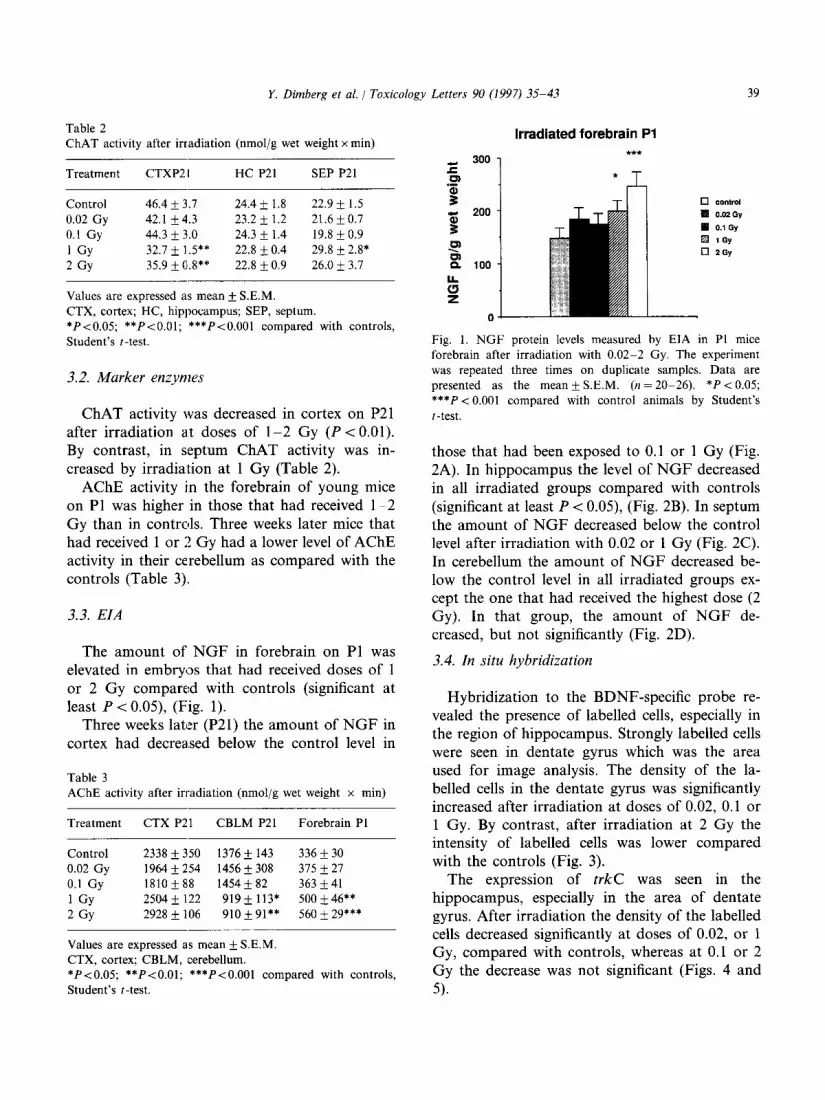
The amount of NGF in forebrain on Pl was elevated in embryos that had received doses of 1 or 2 Gy compared with controls (significant at least P < 0.05), (Fig. 1).
Three weeks later (P21) the amount of NGF in cortex had decreased below the control level in
Table 3
AChE activity after irradiation (nmol/g wet weight x min)
Treatment CTX P21 CBLM P21 Forebrain Pl
Control
0.02 Gy
0.1 Gy
1 GY
2 GY
2338 f 350 1376 + 143 336 k 30
1964 k 254 1456 k 308 375 &- 27 1810 k 88 1454 + 82 363 f 41
2504 & 122 919 * 113* 500 + 46**
2928 + 106 910+91** 560 k 29***
Values are expressed as mean & S.E.M.
CTX, cortex; CBLM, cerebellum.
*P<O.O5; **P<O.Ol; ***P<O.OOl compared with controls,
Student’s t-test.
Irradiated forebrain Pl
! Fig. 1. NGF protein levels measured by EIA in Pl mice
forebrain after irradiation with 0.02-2 Gy. The experiment
was repeated three times on duplicate samples. Data are
presented as the mean k S.E.M. (n = 20-26). *P < 0.05;
***P < 0.001 compared with control animals by Student’s
t-test.
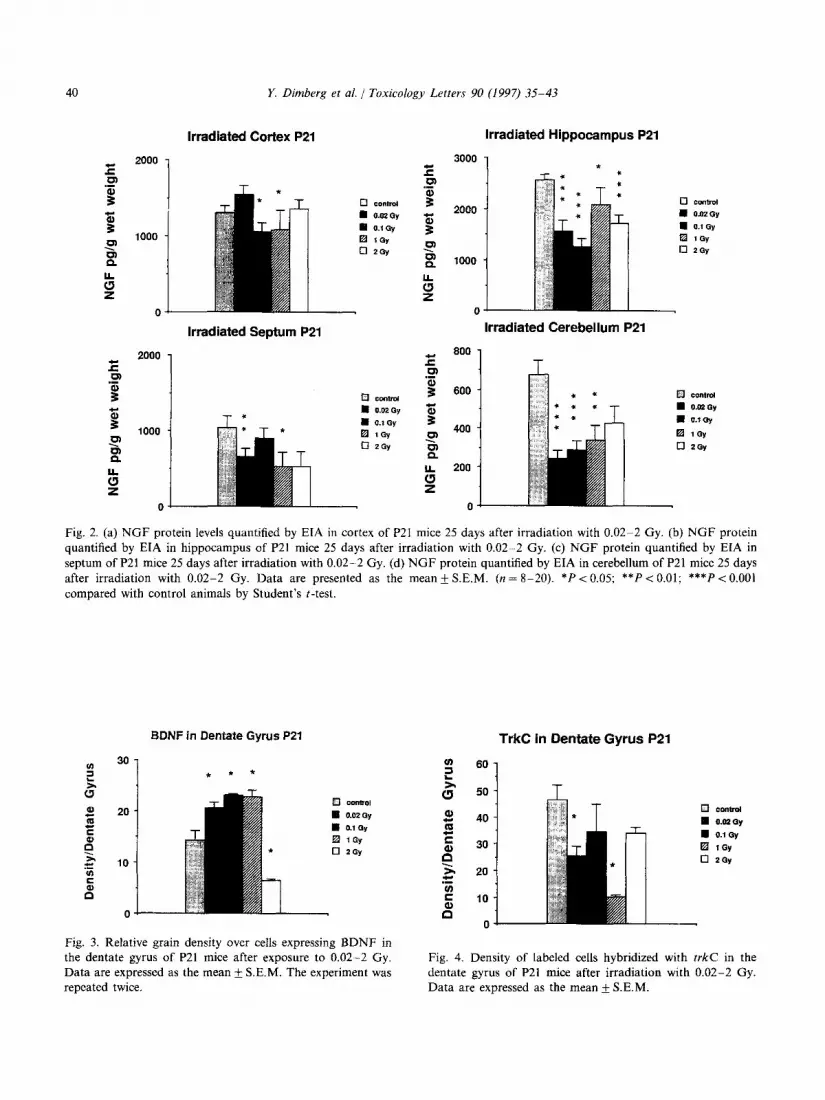
those that had been exposed to 0.1 or 1 Gy (Fig. 2A). In hippocampus the level of NGF decreased in all irradiated groups compared with controls (significant at least P < 0.05), (Fig. 2B). In septum the amount of NGF decreased below the control level after irradiation with 0.02 or 1 Gy (Fig. 2C). In cerebellum the amount of NGF decreased be- low the control level in all irradiated groups ex- cept the one that had received the highest dose (2 Gy). In that group, the amount of NGF de- creased, but not significantly (Fig. 2D).
3.4. In situ hybridization
Hybridization to the BDNF-specific probe re- vealed the presence of labelled cells, especially in the region of hippocampus. Strongly labelled cells were seen in dentate gyrus which was the area used for image analysis. The density of the la- belled cells in the dentate gyrus was significantly increased after irradiation at doses of 0.02, 0.1 or 1 Gy. By contrast, after irradiation at 2 Gy the intensity of labelled cells was lower compared with the controls (Fig. 3).
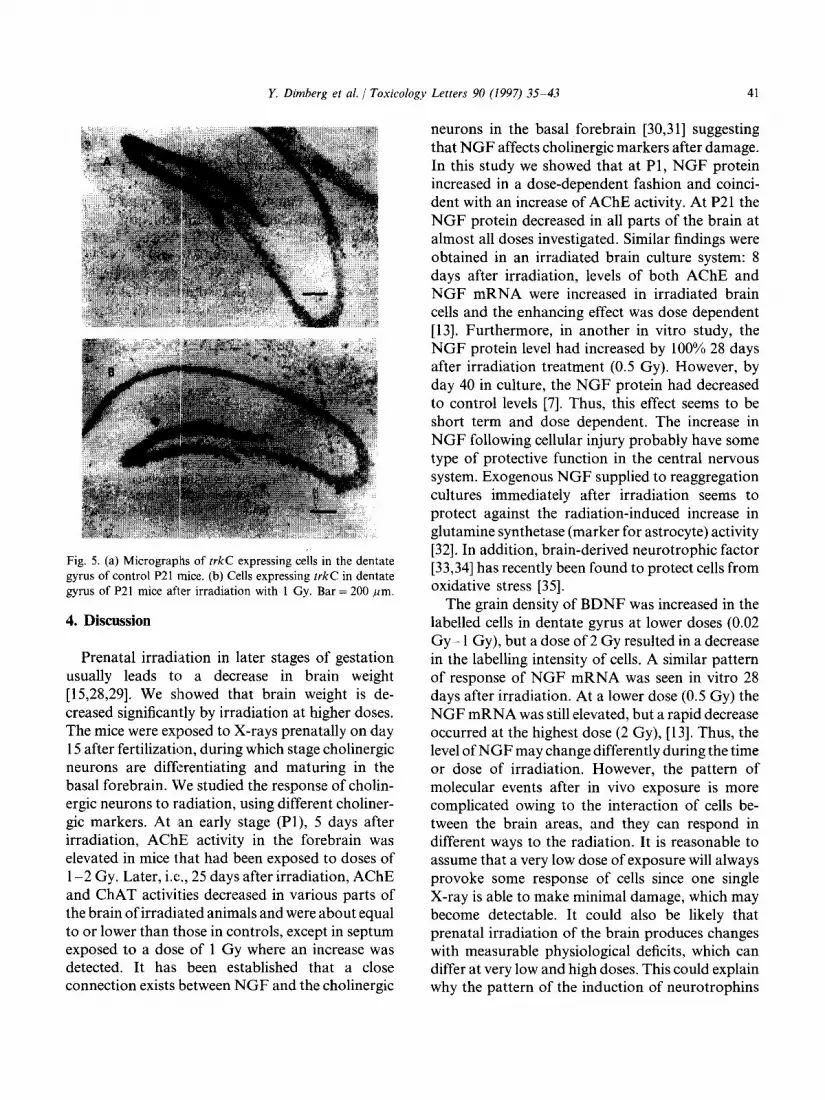
The expression of trkC was seen in the hippocampus, especially in the area of dentate gyrus. After irradiation the density of the labelled cells decreased significantly at doses of 0.02, or 1 Gy, compared with controls, whereas at 0.1 or 2 Gy the decrease was not significant (Figs. 4 and 5).
40 Y. Dimberg et al. J Toxicology Letters 90 (1997) 35-43
5 .zJ $ z
2000
B ul
2 1000
E 0
Irradiated Septum P21
J Irradiated Cerebellum P21
q cmtmr
E 800
.F
$ 800
5 B m 400
2 200
/ 0
Fig. 2. (a) NGF protein levels quantified by EIA in cortex of P21 mice 25 days after irradiation with 0.02-2 Gy. (b) NGF protein quantified by EIA in hippocampus of P21 mice 25 days after irradiation with 0.02-2 Gy. (c) NGF protein quantified by EIA in septum of P21 mice 25 days after irradiation with 0.02-2 Gy. (d) NGF protein quantified by EIA in cerebellum of P21 mice 25 days after irradiation with 0.02-2 Gy. Data are presented as the mean k S.E.M. (n = 8-20). *PC 0.05; **PC 0.01; ***P < 0.001 compared with control animals by Student’s t-test.
Irradiated Cortex P21 Irradiated Hippocampus P21
BDNF in Dentate Gyros P21
Fig. 3. Relative grain density over cells expressing BDNF in
TrkC in Dentate Gyrus P21
60 1
the dentate gyrus of P21 mice after exposure to 0.02-Z Gy. Fig. 4. Density of labeled cells hybridized with t&C in the Data are expressed as the mean k S.E.M. The experiment was dentate gyrus of P21 mice after irradiation with 0.02-2 Gy. repeated twice. Data are expressed as the mean + S.E.M.
Y. Dimberg et al. / Toxicology Letters 90 (1997) 35-43 41
Fig. 5. (a) Micrographs of trkC expressing cells in the dentate gyrus of control P21 mice. (b) Cells expressing trkC in dentate gyrus of P21 mice after irradiation with 1 Gy. Bar = 200 pm.
4. Discussion
Prenatal irradi(ation in later stages of gestation usually leads to a decrease in brain weight [15,28,29]. We showed that brain weight is de- creased significantly by irradiation at higher doses. The mice were exposed to X-rays prenatally on day 15 after fertilization, during which stage cholinergic neurons are differentiating and maturing in the basal forebrain. We studied the response of cholin- ergic neurons to radiation, using different choliner- gic markers. At .an early stage (Pl), 5 days after irradiation, AChE activity in the forebrain was elevated in mice that had been exposed to doses of l-2 Gy. Later, i.e., 25 days after irradiation, AChE and ChAT activities decreased in various parts of the brain of irradiated animals and were about equal to or lower than those in controls, except in septum exposed to a dose of 1 Gy where an increase was detected. It has been established that a close connection exists between NGF and the cholinergic
neurons in the basal forebrain [30,31] suggesting that NGF affects cholinergic markers after damage. In this study we showed that at Pl, NGF protein increased in a dose-dependent fashion and coinci- dent with an increase of AChE activity. At P21 the NGF protein decreased in all parts of the brain at almost all doses investigated. Similar findings were obtained in an irradiated brain culture system: 8 days after irradiation, levels of both AChE and NGF mRNA were increased in irradiated brain cells and the enhancing effect was dose dependent [13]. Furthermore, in another in vitro study, the NGF protein level had increased by 100% 28 days after irradiation treatment (0.5 Gy). However, by day 40 in culture, the NGF protein had decreased to control levels [7]. Thus, this effect seems to be short term and dose dependent. The increase in NGF following cellular injury probably have some type of protective function in the central nervous system. Exogenous NGF supplied to reaggregation cultures immediately after irradiation seems to protect against the radiation-induced increase in glutamine synthetase (marker for astrocyte) activity [32]. In addition, brain-derived neurotrophic factor [33,34] has recently been found to protect cells from oxidative stress [35].
The grain density of BDNF was increased in the labelled cells in dentate gyrus at lower doses (0.02 Gy- 1 Gy), but a dose of 2 Gy resulted in a decrease in the labelling intensity of cells. A similar pattern of response of NGF mRNA was seen in vitro 28 days after irradiation. At a lower dose (0.5 Gy) the NGF mRNA was still elevated, but a rapid decrease occurred at the highest dose (2 Gy), [13]. Thus, the level of NGF may change differently during the time or dose of irradiation. However, the pattern of molecular events after in vivo exposure is more complicated owing to the interaction of cells be- tween the brain areas, and they can respond in different ways to the radiation. It is reasonable to assume that a very low dose of exposure will always provoke some response of cells since one single X-ray is able to make minimal damage, which may become detectable. It could also be likely that prenatal irradiation of the brain produces changes with measurable physiological deficits, which can differ at very low and high doses. This could explain why the pattern of the induction of neurotrophins

42 Y. Dimberg e6 al. / Toxicology Letters 90 (1997) 35-43
was more complex in the present study, especially with regard to the dose-response relationship. In contrast to BDNF, a reduced intensity in labelled cells of @kc-specific probe was observed in den- tate gyrus. The mechanism which controls the expression of neurotrophin receptors during neu- ronal development is largely unknown but it can be assumed that a differential regulation of indi- vidual neurotrophins occurs in the brain. Some recent studies support this assumption. Rocamora et al. [36] found a very different pattern of expres- sion of neurotrophins after introhippocampal in- jection of excitotoxin quinolinic acid in the adult rat brain. A dramatic increase of NGF and BDNF mRNAs and a decrease of NT-3 mRNA were observed in dentate gyrus. In the hippocampal pyramidal layer only BDNF mRNA increased. In another study NGF and BDNF mRNAs appeared to be induced differential in response to impulse activity [37]. Furthermore, treatment of cultured sympathetic neurons from newborn rat superior cervical ganglia with retinoic acid suppressed the expression of trkA (receptor serving to bind NGF) mRNA and induced the expression of trkB (recep- tor serving to bind BDNF) mRNA [38]. These observations may be an explanation for mecha- nisms by which the cells signaling via autocrine or paracrine system to mediate specific cellular and tissue responses after damage. Previously, it has been shown that gene expression is altered by ionizing radiation [5,8,13], and it is believed that the radiation damage to DNA evokes specific cellular responses such as the induction of growth factors, enzymes or other cellular proteins [5,7,8,39]. Such growth factors may remain stimu- lated for days or weeks [39] but can subsequently be down regulated to normal [8,40] or below-nor- mal level [12,13].
In summary, X-irradiation of the prenatal brain induced alterations of NGF protein which coin- cided with changes in AChE activity. Further- more, the NGF protein level and the expression of BDNF and trkC mRNA were affected by doses as low as 0.02 Gy, indicating that the regulation of different neurotrophins and their receptors is very sensitive to radiation. The alternated production of the neurotrophins may modulate the response of cells to radiation.
Acknowledgements
Technical assistance with the X-ray procedure provided by Britt-Marie Svedenstal was greatly appreciated. Financial support was obtained from the Swedish Radiation Protection Institute and the Swedish Medical and Natural Science Research Councils (K94-03P-10159-03 1, B94-13X-09025- 05B, B-BU 04024-313). M.V. was supported by grants from the Swedish Institute and the Wenner- Gren Center Foundation for Scientific Research. These experiments were performed with the per- mission of the local Swedish Ethical Committee.
References
[l] Frankenberg-Schwager, M. (1989) Review of repair kinet- ics for DNA damage induced in eukaryotic cells in vitro by ionizing radiation. Radioth. Oncol. 14, 307-320.
[2] Thacker, .I. (1986) The use of recombinant DNA tech- niques to study radiation-induced damage, repair and genetic change in mammalian cells. Int. J. Rad. Biol. 50, t-30.
[3] Boothman, D.A., Bouvard, I. and Hughes, E.N. (1989) Identification and characterization of X-ray-induced proteins in human cells. Cancer Res. 49, 2871-2878.
[4] Singh, S.P. and Lavin, M.F. (1990) DNA-binding protein activated by y-radiation in human cells. Mol. Cell Biol. 10, 527995285.
[5] Hallahan, D.E., Spriggs, D.R., Beckett, M.A., Kufe, D.W. and Weichselbaum, R.R. (1989) Increased tumor necrosis factor a mRNA after cellular exposure to ionizing radia- tion. Proc. Natl. Acad. Sci. USA 86, 1010410107.
[6] Fuks, Z. and Weichselbaum, R.R. (1992) Radiation toler- ance and the new biology: growth factor involvement in radiation injury to the lung. Int. J. Radiat. Oncol. Biol. Phys. 24, 183-184.
[7] Dimberg, Y., Tottmar, O., Aspberg, A., Ebendal, T., Johansson, K-J. and Walinder, G. (1992) Effects of low- dose X-irradiation on mouse-brain aggregation cultures. Int. J. Radiat. Biol. 61, 355-363.
[8] Haimovitz-Friedman, A., Voldavsky, I., Witte, L. and Fuks, Z. (1991) Autocrine effects of fibroblast growth factor in the repair of radiation damage in endothelial cells. Cancer Res. 51, 2552-2558.
[9] Jung, M., Kern, F.G., Jorgensen, T.J., McLeskey, SW., Blair, O.C. and Dritschilo, A. (1994) Fibroblast growth factor-4 enhanced G, arrest and cell survival following ionizing radiation. Cancer Res. 54, 5194-5197.
[lo] Witte, L., Fuks, Z., Hamovitz-Friedman, A., Voldavsky, I., Goodman, D.W.S. and Eldor, A. (1989) Effects of irradiation on the release of growth factors from cultured bovine porcine, and human endothelial cells. Cancer Res. 49, 506665072.

Y. Dimberg et al. 1 Toxicology Letters 90 (1997) 35-43 43
1111
v-21
1131
1141
u51
[I61
v71
1181
[I91
PO1
WI
PI
1231
~241
[251
WI
Anscher, MS., Cracker, I.R. and Jirtle, R.L. (1990)
Transforming growth factor-/3 1 expression in irradiated
liver. Radiat. Res. 122, 77-85
Finkelstein, J.N., Johnston, C.J., Baggs, R. and Rubin, P.
(1994) Early alterations in extracellular matrix and trans-
forming growth factro /r gene expression in mouse lung
indicative of late radiation fibrosis. Int. J. Radiat. Oncol.
Biol. Phys. 28, 621-631.
Dimberg, Y. an,d Larkfors, L. (1994) Effects of irradia-
tion on cholinergic neurons and nerve growth factor
mRNA in mouse foetal brain aggregation cultures. Int. J.
Radiat. Biol. 66, 7933800.
Valcana, T., Liao, C. and Tmiras, P.S. (1974) Effects of
X-radiation on the subcellular distribution of cholinergic
enzymes in the developing rat cerebellum. Brain Res. 73,
105-120.
Brizzee, K.R., Cirdy, J.M., Kaack, M.B. and Beavers, T.
(1980) Effects of prenatal ionizing radiation on the visual
cortex and hippocampus of newborn squirrel monkeys. J.
Neuropathol. Exp. Neurol. 39, 5233540.
Larkfors, L., Osckarsson, A., Sundberg, J. and Ebendal,
T. (1991) Methylmercury induced alteration in the nerve
growth factor level in the developing brain. Develop.
Brain Res. 62, 287-291.
Ebendal, T. (1989) NGF in CNS: Experimental data and
clinical implications. Prog. Growth Factor Res. 1, 143-
159.
Hofer, M.M., Pagliusi, S.R., Hohn, A., Leibrock, J. and
Barde, Y-A. (1990) Regional distribution of brain-derived
neurotrophic factor mRNA in the adult mouse brain.
EMBO J. 9, 24519-2464.
Emfors, P., Wetmore, C., Olson, L. and Persson, H.
(1990) Identification of cells in rat brain and peripheral
tissues expressing mRNA for members of the nerve
growth factor family. Neuron 5, 51 l-526.
Ellman, G.T., Courtney, K.D., Andres, V. Jr. and Feath-
erstone, R.M. (1961) A new and rapid calorimetric deter-
mination of acetylcholinesterase activity. Biochem.
Pharmacol. 7, 813-95.
Wilson, S.H., Schrier, B.K., Farber, J.L., Thompson,
E.J., Rosenberg, R.N., Blume, A.J. and Nirenberg, M.W.
(1972) Markers for gene expression in cultured cells from
the nervous system. J. Biol. Chem. 247, 3159-3169.
Hinegardner, R.T. (1971) An improved fluorometric assay
for DNA. Anal. Biochem. 39, 197-201.
Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall,
R.J. (1951) Protein measurement with the folin phenol
reagent. J. Biol. Chem. 193, 265-275.
Siiderstrom, S., Hallbook, F., Ibanez, C.F., Persson, H.
and Ebendal, 1‘. (1990) Recombinant human p-nerve
growth factor (NGF): biological activity and properties in
an enzyme immunoassay. J. Neurosci. Res. 27, 665-677.
Korsching, S. and Thoenen, H. (1983) Nerve growth
factor in sympathetic ganglia and corresponding target
organs of the rat: correlation with density of sympathetic
innervation. Proc. Natl. Acad. Sci. USA 80, 3513-3516.
Maisonpierre, P.C., Belluscio, L., Friedman, B., Alder-
1271
1281
~291
1301
[311
~321
[331
[341
[351
1361
son, R.A., Wiegand, J., Furth, E., Lindsay, R.M. and
Yancopoulos, G.D. (1990) NT-3, BDNF, and NGF in
the developing rat nervous system: parallel as well as
reciprocal patterns of expression. Neuron 5, 501-509.
Merliot, J-P., Ernfors, P., Jaber, M. and Persson, H.
(1992) Molecular cloning of rat trkC and distribution of
cells expressing messenger RNAs for members of the trk family in the rat central nervous system. Neuroscience 51,
513-532.
Wanner, R.A. and Edwards, M.J. (1983) Comparison of
the effect of radiation of brain growth of guinea pigs. Br.
J. Radiol. 56, 33-39.
Konermann, G. (1987) Postimplantation defects in devel-
opment following ionizing irradiation. Adv. Radiat. Biol.
13, 91-165.
Hefti, F. and Weiner, W.J. (1986) Nerve Growth Factor
and Alzheimer’s disease. Ann. Neurol. 20, 2755281.
Hartikka, J. and Hefti, F. (1988) Comparison of nerve
growth factor’s effects on development of septum, stria-
turn and nucleus basalis cholinergic neurons in vitro. J.
Neurosci. Res. 21, 352-364.
Dimberg, Y., Aspberg, A. and Tottmar, 0. (1993) Effects
of nerve growth factor on X-irradiated reaggregation
cultures of rat brain cells. Int. J. Radiat. Biol. 64, 731-
737. Barde, Y-A., Edgar, D. and Thoenen, H. (1982) Purifica-
tion of a new neurotrophic factor from mammalian brain.
EMBO J. 1, 549-553.
Ebendal, T. (1992) Function and evolution in the NGF
family and its receptors. J. Neurosci. Res. 32, 461-470.
Spina, M.B., Squinto, S.P., Miller, J., Lindsay, R.M. and
Hyman, S. (1992) Brain-derived neurotrophic factor pro-
tects dopamine neurons against 6-hydroxydopamine and
N-methyl-4-phenylpyridinium ion toxicity: involvement of
the glutathione system. J. Neurochem. 59, 999106.
Rocamora, N., Massien, L., Boddeke, H.W.G.M., Pala-
cios, J.M. and Mengod, G. (1994) Differential regulation
of the expression of nerve growth factor, brain derived
neurotrophic factor and neurotrophin-3 mRNAs in adult
rat brain after intrahippocampal injection of quinolinic
acid. Mol. Brain Res. 26, 89998.
[37] Elliott, R.C., Inturrisi, C.E., Black, I.B. and Dreyfus,
CF. (1994) An improved method detects differential
NGF and BDNF gene expression in response to depolar-
ization in cultured hippocampal neurons. Mol. Brain Res.
26, 81-88.
[38] Kobayaski, M., Kurihara, K. and Matsuoka, I. (1994)
Retinoic acid induces BDNF responsivness of sympa-
thetic neurons by alteration of Trk neurotrophin receptor
expression. FEBS Letters 356, 60-65.
[39] Barcellos-Hoff, M.H. (1993) Radiation-induced trans-
forming growth factor p and subsequent extracellular
matrix reorganization in murine mammary gland. Cancer
Res. 53, 3880-3886. [40] Sherman, M.L., Datta, R., Hallahan, D.E., Weichsel-
baum, R.R. and Kufe, D.W. (1990) Ionizing radiation
regulates expression of the c-jun protooncogene. Proc.
Natl. Acad. Sci. USA 87, 5663-5666.