cortical-striatal integration of cocaine history and prefrontal dysfunction in animal modeling of...
TRANSCRIPT

CPDR
Bau
Mois
Rhvla
Ct
Ks
Dgua(lqgamu
da(icnh
F
A
R
0d
ortical-Striatal Integration of Cocaine History andrefrontal Dysfunction in Animal Modeling of Dualiagnosis
. Andrew Chambers, Alena M. Sentir, Susan K. Conroy, William A. Truitt, and Anantha Shekhar
ackground: High rates of substance disorders in schizophrenia and other mental illnesses may reflect biological vulnerabilities to theddiction process. Interactions between addictive drug effects and mental illness involving circuits that generate motivated behavior maynderpin this vulnerability.
ethods: We examined separate and combined effects of neonatal ventral hippocampal lesions (NVHLs)—a neurodevelopmental modelf schizophrenia (vs. SHAM-operated control animals)—and a behaviorally sensitizing cocaine history (15 mg/kg/day � 5 days vs. saline
njections) on acute cocaine-induced neural activation signaled by c-Fos expression. Stereological assessment of activation densitiespanned six ventral to dorsal cortical-striatal compartments.
esults: Cortically, NVHLs showed hypoactivation and decreased volume of the ventral medial prefrontal cortex. In contrast, cocaineistory was only expressed subcortically and as a hyperactivating effect in the dorsal striatum where significant NVHL-induced hyperacti-ation also emerged. Across all subjects and brain regions, only dorsal striatal activation was correlated with differences in sensitized
ocomotion. However, this activation was tightly correlated to a simple multiplicative function of ventral medial prefrontal hypoactivationnd cocaine history-related increases in striatal activation.
onclusions: These findings suggest drug history and developmental temporal limbic abnormalities associated with prefrontal dysfunc-
ion produce compounding effects within cortical-striatal circuits as mechanistic basis for dual diagnosis.ey Words: c-Fos, dual diagnosis, hippocampus, prefrontal cortex,tereology, striatum
rug addictions are mainstream in mentally ill populations,and “dual diagnosis” is associated with greater medical andpsychiatric illness and death (1). Growing evidence sug-
ests mental illnesses and addiction vulnerability co-emerge invol-ntarily from a unified etiology, definable on the neurocircuit levelnd contributed to by shared environmental and genetic factors2,3). Abnormalities of prefrontal cortex (PFC) and/or temporal-imbic regions (e.g., hippocampus) span mental disorders fre-uently co-occurring with addictions (4). These regions providelutamatergic projections to ventral striatal targets (e.g., nucleusccumbens [NAC]), where in conjunction with mesolimbic dopa-ine, they modulate information processing and neuroplasticitynderlying alterations in motivated behavior (5,6).
In animals, developmental temporal-limbic disruptions pro-uce complex neuropsychiatric syndromes including signs ofddiction vulnerability (4). Neonatal ventral hippocampal lesionNVHL) rats demonstrate multiple schizophrenia features, includ-ng positive-like neuroleptic-responsive symptoms of periadoles-ent onset, cognitive impairments, negative symptom traits, andeurobiological markers of PFC dysfunction (7). Neonatal ventralippocampal lesion rats also show accentuated long-term behav-
rom the Laboratory for Translational Neuroscience of Dual Diagnosis &Development (RAC, AMS, SKC), Department of Psychiatry; Departmentof Anatomy and Cell Biology (WAT); and Indiana Clinical and Transla-tional Sciences Institute & Department of Psychiatry (AS), Indiana Uni-versity School of Medicine, Indianapolis, Indiana.
ddress correspondence to R. Andrew Chambers, M.D., Laboratory forTranslational Neuroscience of Dual Diagnosis & Development, Depart-ment of Psychiatry, Indiana University School of Medicine, Institute ofPsychiatric Research, 791 Union Drive, Indianapolis, IN 46202; E-mail:[email protected].
eceived Mar 20, 2009; revised Sep 3, 2009; accepted Sep 4, 2009.
006-3223/10/$36.00oi:10.1016/j.biopsych.2009.09.006
ioral sensitization to cocaine (8), heightened baseline and co-caine history-induced impulsivity (9), and enhanced drug takingand drug seeking in cocaine self-administration (10).
Characterization of a neural integration of addictive drug historyand developmental temporal-limbic alterations underlying psychi-atric syndromes has been lacking. We investigated how a behav-iorally sensitizing cocaine history alters, 2 weeks later, cocaine-induced c-Fos early gene expression in NVHL versus SHAM-controlanimals. Applying stereology to measuring c-Fos activation as anonspecific marker of regional neural activation, we mapped sixcortical-striatal compartments: three cortical (ventral-medial pre-frontal, dorsal-medial prefrontal, secondary motor cortices [vmPFC,dmPFC, sMOC]) and three striatal (ventral-medial nucleus accum-bens, dorsal-lateral nucleus accumbens, dorsal-lateral caudate-pu-tamen [vmNAC, dlNAC, dlCAPU]). These regions were chosen askey components of parallel cortico-striato-thalamo-cortical loopsthought to interactively generate motivated behavior via a ventral-dorsal directionality of information processing (11–13). We soughtto 1) define separate and combined effects of NVHLs and cocainehistory on cortical-striatal activation; 2) determine which regionalactivations correspond to behavior; and 3) explore quantitativerelationships between regional activations as brought to light byNVHL and cocaine history effects.
Methods and Materials
Subjects/Surgeries/BehaviorPostnatal day 7 (PD-7), male Sprague-Dawley pups underwent
NVHL or SHAM surgery as described previously (10). Pups were raisedunder standard conditions through weaning (PD-21) then pair-housed(PD-35). On PD-70, rats were randomized to daily injections (intraperi-toneal) of saline (1 mL/kg) or cocaine hydrochloride (15 mg/kg insaline [1mL/kg]) for 5 days in homecages. Twoweeks after completinginjections (PD-89), a single cocaine injection (15 mg/kg) was deliveredto all rats after 1 hour of a 2-hour locomotor arena session. Allprocedures (Section 1 in Supplement 1) accorded with NIH Guide for
Care and Use of Laboratory Animals.BIOL PSYCHIATRY 2010;67:788–792© 2010 Society of Biological Psychiatry
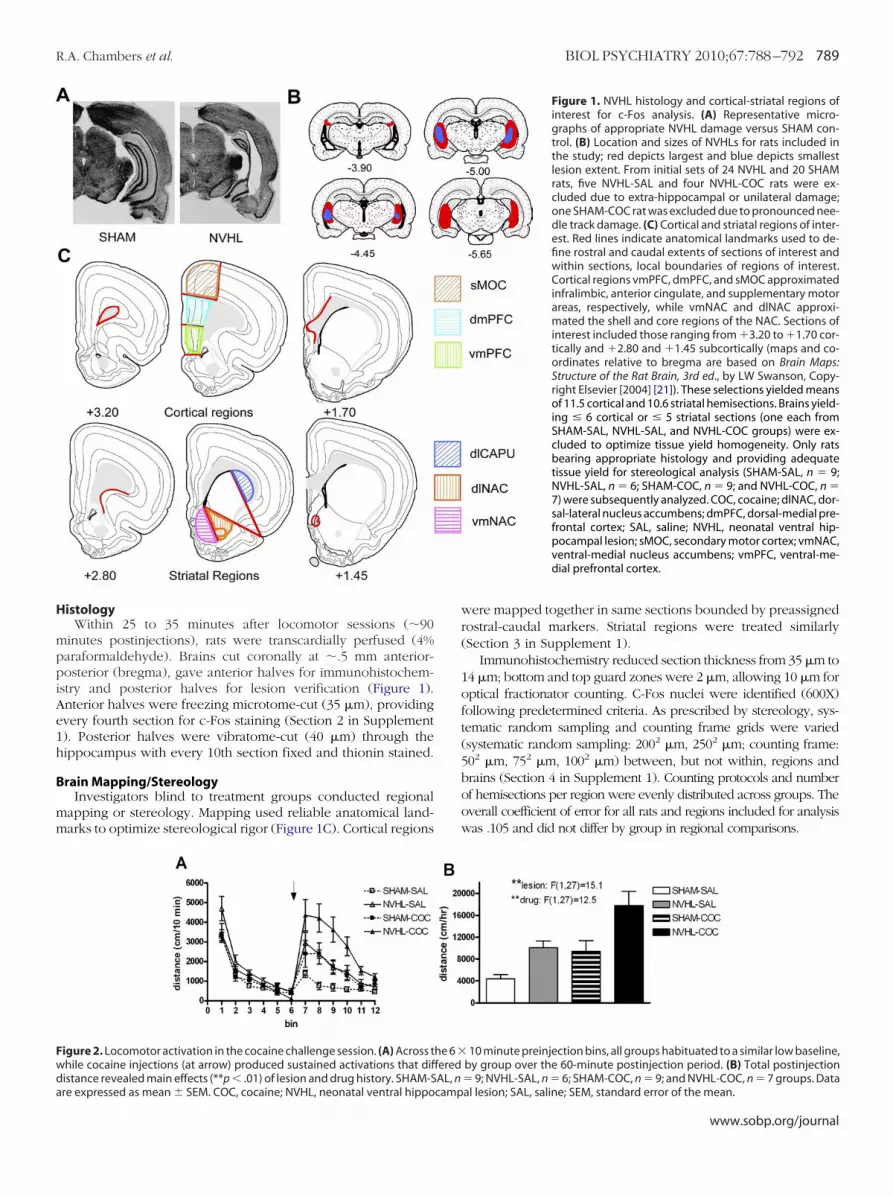
H
mppiAe1h
B
mm
Fwda
R.A. Chambers et al. BIOL PSYCHIATRY 2010;67:788–792 789
istologyWithin 25 to 35 minutes after locomotor sessions (�90
inutes postinjections), rats were transcardially perfused (4%araformaldehyde). Brains cut coronally at �.5 mm anterior-osterior (bregma), gave anterior halves for immunohistochem-
stry and posterior halves for lesion verification (Figure 1).nterior halves were freezing microtome-cut (35 �m), providingvery fourth section for c-Fos staining (Section 2 in Supplement). Posterior halves were vibratome-cut (40 �m) through theippocampus with every 10th section fixed and thionin stained.
rain Mapping/StereologyInvestigators blind to treatment groups conducted regional
apping or stereology. Mapping used reliable anatomical land-arks to optimize stereological rigor (Figure 1C). Cortical regions
igure 2. Locomotor activation in the cocaine challenge session. (A) Across thile cocaine injections (at arrow) produced sustained activations that difistance revealed main effects (**p � .01) of lesion and drug history. SHAM-S
re expressed as mean � SEM. COC, cocaine; NVHL, neonatal ventral hippocampwere mapped together in same sections bounded by preassignedrostral-caudal markers. Striatal regions were treated similarly(Section 3 in Supplement 1).
Immunohistochemistry reduced section thickness from 35 �m to14 �m; bottom and top guard zones were 2 �m, allowing 10 �m foroptical fractionator counting. C-Fos nuclei were identified (600X)following predetermined criteria. As prescribed by stereology, sys-tematic random sampling and counting frame grids were varied(systematic random sampling: 2002 �m, 2502 �m; counting frame:502 �m, 752 �m, 1002 �m) between, but not within, regions andbrains (Section 4 in Supplement 1). Counting protocols and numberof hemisections per region were evenly distributed across groups. Theoverall coefficient of error for all rats and regions included for analysiswas .105 and did not differ by group in regional comparisons.
Figure 1. NVHL histology and cortical-striatal regions ofinterest for c-Fos analysis. (A) Representative micro-graphs of appropriate NVHL damage versus SHAM con-trol. (B) Location and sizes of NVHLs for rats included inthe study; red depicts largest and blue depicts smallestlesion extent. From initial sets of 24 NVHL and 20 SHAMrats, five NVHL-SAL and four NVHL-COC rats were ex-cluded due to extra-hippocampal or unilateral damage;one SHAM-COC rat was excluded due to pronounced nee-dle track damage. (C) Cortical and striatal regions of inter-est. Red lines indicate anatomical landmarks used to de-fine rostral and caudal extents of sections of interest andwithin sections, local boundaries of regions of interest.Cortical regions vmPFC, dmPFC, and sMOC approximatedinfralimbic, anterior cingulate, and supplementary motorareas, respectively, while vmNAC and dlNAC approxi-mated the shell and core regions of the NAC. Sections ofinterest included those ranging from �3.20 to �1.70 cor-tically and �2.80 and �1.45 subcortically (maps and co-ordinates relative to bregma are based on Brain Maps:Structure of the Rat Brain, 3rd ed., by LW Swanson, Copy-right Elsevier [2004] [21]). These selections yielded meansof 11.5 cortical and 10.6 striatal hemisections. Brains yield-ing � 6 cortical or � 5 striatal sections (one each fromSHAM-SAL, NVHL-SAL, and NVHL-COC groups) were ex-cluded to optimize tissue yield homogeneity. Only ratsbearing appropriate histology and providing adequatetissue yield for stereological analysis (SHAM-SAL, n � 9;NVHL-SAL, n � 6; SHAM-COC, n � 9; and NVHL-COC, n �7) were subsequently analyzed. COC, cocaine; dlNAC, dor-sal-lateral nucleus accumbens; dmPFC, dorsal-medial pre-frontal cortex; SAL, saline; NVHL, neonatal ventral hip-pocampal lesion; sMOC, secondary motor cortex; vmNAC,ventral-medial nucleus accumbens; vmPFC, ventral-me-dial prefrontal cortex.
10 minute preinjection bins, all groups habituated to a similar low baseline,by group over the 60-minute postinjection period. (B) Total postinjection� 9; NVHL-SAL, n � 6; SHAM-COC, n � 9; and NVHL-COC, n � 7 groups. Data
he 6 �feredAL, n
al lesion; SAL, saline; SEM, standard error of the mean.
www.sobp.org/journal
R
B
m2it.F
S
(Cve
790 BIOL PSYCHIATRY 2010;67:788–792 R.A. Chambers et al.
w
esults
ehavioral SensitizationCocaine increased locomotion over habituated levels (10-
inute preinjection bin) that were similar among groups (FigureA). Neonatal ventral hippocampal lesions and cocaine historyncreased cocaine-induced locomotion independently and addi-ively [two-way analysis of variance: lesion: F (1,27) � 15.1, p �01; drug history: F (1,27) � 12.5, p � .01; lesion � drug history:(1,27) � .6, p � .5] (Figure 2B).
tereological Analysis of C-Fos ActivationDensities of c-Fos activation by region and treatment group
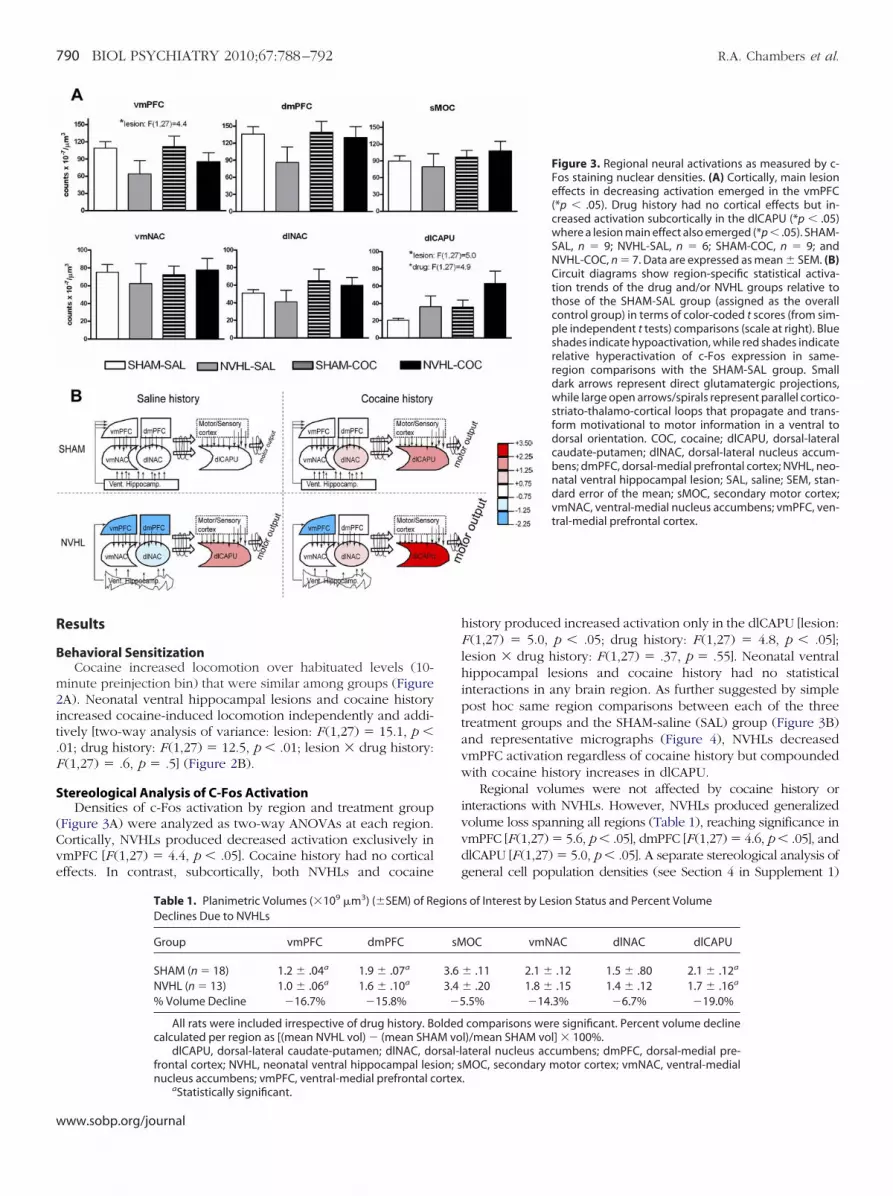
Figure 3A) were analyzed as two-way ANOVAs at each region.ortically, NVHLs produced decreased activation exclusively inmPFC [F (1,27) � 4.4, p � .05]. Cocaine history had no corticalffects. In contrast, subcortically, both NVHLs and cocaine
Table 1. Planimetric Volumes (�109 �m3) (�SEM) of ReDeclines Due to NVHLs
Group vmPFC dmPFC
SHAM (n � 18) 1.2 � .04a 1.9 � .07a
NVHL (n � 13) 1.0 � .06a 1.6 � .10a
% Volume Decline �16.7% �15.8%
All rats were included irrespective of drug history. Bcalculated per region as [(mean NVHL vol) � (mean SHA
dlCAPU, dorsal-lateral caudate-putamen; dlNAC, dofrontal cortex; NVHL, neonatal ventral hippocampal lesnucleus accumbens; vmPFC, ventral-medial prefrontal c
aStatistically significant.
ww.sobp.org/journal
history produced increased activation only in the dlCAPU [lesion:F (1,27) � 5.0, p � .05; drug history: F (1,27) � 4.8, p � .05];lesion � drug history: F (1,27) � .37, p � .55]. Neonatal ventralhippocampal lesions and cocaine history had no statisticalinteractions in any brain region. As further suggested by simplepost hoc same region comparisons between each of the threetreatment groups and the SHAM-saline (SAL) group (Figure 3B)and representative micrographs (Figure 4), NVHLs decreasedvmPFC activation regardless of cocaine history but compoundedwith cocaine history increases in dlCAPU.
Regional volumes were not affected by cocaine history orinteractions with NVHLs. However, NVHLs produced generalizedvolume loss spanning all regions (Table 1), reaching significance invmPFC [F(1,27) � 5.6, p � .05], dmPFC [F(1,27) � 4.6, p � .05], anddlCAPU [F(1,27) � 5.0, p � .05]. A separate stereological analysis ofgeneral cell population densities (see Section 4 in Supplement 1)
Figure 3. Regional neural activations as measured by c-Fos staining nuclear densities. (A) Cortically, main lesioneffects in decreasing activation emerged in the vmPFC(*p � .05). Drug history had no cortical effects but in-creased activation subcortically in the dlCAPU (*p � .05)where a lesion main effect also emerged (*p � .05). SHAM-SAL, n � 9; NVHL-SAL, n � 6; SHAM-COC, n � 9; andNVHL-COC, n � 7. Data are expressed as mean � SEM. (B)Circuit diagrams show region-specific statistical activa-tion trends of the drug and/or NVHL groups relative tothose of the SHAM-SAL group (assigned as the overallcontrol group) in terms of color-coded t scores (from sim-ple independent t tests) comparisons (scale at right). Blueshades indicate hypoactivation, while red shades indicaterelative hyperactivation of c-Fos expression in same-region comparisons with the SHAM-SAL group. Smalldark arrows represent direct glutamatergic projections,while large open arrows/spirals represent parallel cortico-striato-thalamo-cortical loops that propagate and trans-form motivational to motor information in a ventral todorsal orientation. COC, cocaine; dlCAPU, dorsal-lateralcaudate-putamen; dlNAC, dorsal-lateral nucleus accum-bens; dmPFC, dorsal-medial prefrontal cortex; NVHL, neo-natal ventral hippocampal lesion; SAL, saline; SEM, stan-dard error of the mean; sMOC, secondary motor cortex;vmNAC, ventral-medial nucleus accumbens; vmPFC, ven-tral-medial prefrontal cortex.
s of Interest by Lesion Status and Percent Volume
OC vmNAC dlNAC dlCAPU
� .11 2.1 � .12 1.5 � .80 2.1 � .12a
� .20 1.8 � .15 1.4 � .12 1.7 � .16a
.5% �14.3% �6.7% �19.0%
comparisons were significant. Percent volume declinel)/mean SHAM vol] � 100%.ateral nucleus accumbens; dmPFC, dorsal-medial pre-MOC, secondary motor cortex; vmNAC, ventral-medial.
gion
sM
3.63.4�5
oldedM vorsal-l
ion; sortex

e(tpSC
N
spawvc
ab(iFnp
dmdchbb.(d
FsRdSdtcp
R.A. Chambers et al. BIOL PSYCHIATRY 2010;67:788–792 791
xamined regions where significant c-Fos differences emergedvmPFC and dlCAPU). Neonatal ventral hippocampal lesionsended to increase cell-packing densities in both regions, mostrominently in the vmPFC (NVHL: 11.8 [� .7] � 10�5 cells/�m3;HAM: 10.7 [� .7] � 10�5 cells/�m3), although not significantly.ocaine history also had no effects on cell densities.
eural Activation and BehaviorAcross six separate Pearson correlations, each including all
ubjects, only dlCAPU activation was highly correlated withostinjection locomotion (Table 2). To explore whether dlCAPUctivation might represent a function of regional activationsithin PFC and NAC, we defined measures of vmPFC hypoacti-ation and drug history effects on striatal activation (d), whichould be quantified for each rat.
We defined (sMOC)/(vmPFC) as a measure of vmPFC hypo-ctivation, since from cortical regions, sMOC was least affectedy lesion (i.e., [sMOC] was the best cortical standard). CalculatingsMOC)/(vmPFC) for each rat provides a unitless measure thatncreases with greater vmPFC hypoactivation. As shown inigure 5A, (sMOC)/(vmPFC) was 1 for NVHLs and was sig-ificantly greater than for SHAMs [two-tailed t (29) � �3.6,�.01], which was � 1.With respect to cocaine history effects, activation levels
eclined dorsally and with regularity across striatal compart-ents, but cocaine history appeared to diminish the rate ofecline, leading to parametrically significant drug history in-reases in dlCAPU (Figure 5B). Quantifying this effect, in terms ofow drug history increases the ratio of dorsal striatal to accum-ens activation, we defined d � (dlCAPU)/(dlNAC) on a per ratasis, giving means of dc � .74 for cocaine history rats and ds �56 for saline history rats. As d algebraically rearranges todlCAPU) � d(dlNAC), we set a relationship predictive of
igure 4. Representative micrographs (100� magnification) of cortical andtriatal effects of lesion and drug history on c-Fos nuclear staining densities.egions of interest are bounded by green lines for vmPFC and blue lines forlCAPU. A paucity of c-Fos nuclei in the vmPFC of NVHL compared withHAM groups was evident for rats with either saline or cocaine histories. InlCAPU, NVHL-COC rats had the greatest c-Fos densities and SHAM-SAL rats
he least, with NVHL-SAL and SHAM-COC falling intermediately. COC, co-aine; dlCAPU, dorsal-lateral caudate-putamen; NVHL, neonatal ventral hip-ocampal lesion; SAL, saline; vmPFC, ventral-medial prefrontal cortex.
lCAPU activation: (dlCAPU)p � d(dlNAC). Incorporating the
vmPFC hypoactivation argument into this expression as a multi-plicative modifier of d(dlNAC) gives:
(dlCAPU)p � d(dlNAC)[(sMOC)/(vmPFC)]
where d ��.74 (cocaine history [dc])
.56 (saline history [ds])
Computing (dlCAPU)p for each rat and plotting it versus theiractual dlCAPU activations (Figure 5C) produces a linear regres-sion line (dlCAPU) � (.98 � .11) (dlCAPU)p � 1.4 that closelyapproximates an exact relationship expecting (dlCAPU) �1(dlCAPU)p � 0. This correlation was significant [F (1,30) � 86.0 �.001(R � .865)], explaining 75% of variation in the data. Althoughnone of vmPFC, sMOC, or dlNAC activations alone correlatedwith postinjection locomotion, their integration in (dlCAPU)pcorrelated with it significantly (Pearson [.57, p � .01]).
Discussion
This study reveals compounding effects of a mental illnessmodel and addictive drug history on cortical-striatal circuit functionand behavioral output. The vmPFC hypoactivation and volumeatrophy in NVHL rats generally agree with evidence for abnormal-ities in PFC neuronal function, excitatory and inhibitory neurotrans-mission, and deficits in neuroplasticity, and dendritic complexity inthe NVHL model, and with PFC atrophy and dysfunction in humanschizophrenia (7,14,15). Meanwhile, subcortical cocaine historyeffects that we observed likely reflect enduring alterations in striatalnetwork activation incurred by prior cocaine history (5), since allrats received the same cocaine dose before sacrifice.
Across all subjects and brain regions, dlCAPU activation wasexclusively and highly correlated with degree of behavioral sensi-tization consistent with Willuhn and Steiner (16), suggesting aprominent role for dorsal striatal activation in motoric effects ofpsychostimulants. Unlike the NAC, which receives cortical projec-tions predominantly from PFC (11), the caudate-putamen receivesinput from cortical motor neurons, firing in registry with them,corresponding to motor performance (17,18). Consistent with thesedata and evidence that ventral cortical-striatal circuits process mo-tivational information that influences dorsal cortical-striatal motorcontrol and learning (4,12,13), our data suggest the dlCAPU oper-ated as an output station where activations depend on represen-tational states within PFC and NAC. Specifically, we found
Table 2. Pearson Correlations Between Postinjection Locomotion andRegional c-Fos Activation Density
Brain Region R p
vmPFC �.247 .18dmPFC �.125 .50sMOC .195 .29vmNAC �.016 .93dlNAC .124 .51dlCAPUa .649 �.001
All rats were included (SHAM-SAL, n � 9; NVHL-SAL, n � 6; SHAM-COC,n � 9; and NVHL-COC, n � 7) for each analysis, irrespective of lesion or drughistory status.
COC, cocaine; dlCAPU, dorsal-lateral caudate-putamen; dlNAC, dorsal-lateral nucleus accumbens; dmPFC, dorsal-medial prefrontal cortex; SAL,saline; sMOC, secondary motor cortex; vmNAC, ventral-medial nucleus ac-cumbens; vmPFC, ventral-medial prefrontal cortex.
aStatistically significant.
quantitative evidence that vmPFC hypoactivation operates like a
www.sobp.org/journal

md
ancdird
FtrNhrt(msctfdcetga(otapdsc
792 BIOL PSYCHIATRY 2010;67:788–792 R.A. Chambers et al.
w
ultiplicative amplifier of drug history effects on the ratio oflCAPU to dlNAC activation, correlating with behavioral output.
Further studies using differential lesions and drug regimensre needed to better understand our findings and to defineeuronal subpopulations where c-Fos expression changes oc-urred. Nevertheless, our results provide initial animal modelingata suggesting that PFC dysfunction and/or atrophy, oftendentified in nonmentally ill addicted populations (3,19,20),epresents a key substrate mechanism in the generation of dual
igure 5. Quantitative relationships between degree of vmPFC hypoactiva-ion, cocaine history-related effects, and dlCAPU activation, inclusive of allats in the study (SHAM-SAL, n � 9; NVHL-SAL, n � 6; SHAM-COC, n � 9; andVHL-COC, n � 7). Data are expressed as mean � SEM. (A) Degree of vmPFCypoactivation calculated as the ratio of sMOC to vmPFC activation on a per
at basis was significantly increased in NVHLs compared with SHAMs (two-ailed t test, p � .05) (lesion groupings are irrespective of drug history).B) C-Fos activation densities of drug history groups across striatal compart-
ents (drug groupings are irrespective of lesion status) suggest a progres-ively increasing separation following a dorsal orientation reaching signifi-ance at the dlCAPU (drug history: p � .05). Calculating d � ratio of dlCAPUo dlNAC activation densities on a per rat basis provides means of dc � .74or cocaine history rats and ds � .56 for saline history rats. (C) Plot of actuallCAPU activation versus predicted CAPU activation [(CAPU)p] for each rat,alculated as the multiplicative product of their corresponding drug historyffect value (dc or ds), dlNAC activation, and degree of vmPFC hypoactiva-ion [(sMOC)/(vmPFC)], suggests differential levels of dlCAPU activation, asenerated by cocaine versus saline history, are further split into sublevels bymultiplicative operation of vmPFC hypoactivation. Actual (dlCAPU) and
dlCAPU)p were highly correlated (***p � .001) and the linear regression linef (dlCAPU) � .98(dlCAPU)p � 1.39 (solid line) closely approximates a line
hat demarks a perfect relationship between (CAPU)p and actual CAPUctivation (dotted line). Units are c-Fos counts � 10� 7/�m3. CAPU, caudate-utamen; COC, cocaine; dlCAPU, dorsal-lateral caudate-putamen; dlNAC,orsal-lateral nucleus accumbens; NVHL, neonatal ventral hippocampal le-ion; SAL, saline; SEM, standard error of the mean; sMOC, secondary motorortex; vmPFC, ventral-medial prefrontal cortex.
iagnoses. These findings impart novel clinical/translational sig-
ww.sobp.org/journal
nificance to basic research characterizing relationships betweenventral and dorsal cortical-striatal circuits in the generation andadaptability of motivated behavior.
This work was supported by the US National Institutes ofHealth, NIDA-K08 DA019850 (RAC). No commercial interestswere involved in the funding or conduct of this research.
The authors report no biomedical financial interests or po-tential conflicts of interest.
Supplementary material cited in this article is availableonline.
1. O’Brien CP, Charney DS, Lewis L, Cornish JW, Post RM, Woody GE, et al.(2004): Priority actions to improve the care of persons with co-occurringsubstance abuse and other mental disorders: A call to action. Biol Psy-chiatry 56:703– 818.
2. Chambers RA, Krystal JK, Self DW (2001): A neurobiological basis for sub-stance abuse comorbidity in schizophrenia. Biol Psychiatry 50:71–83.
3. Brewer JA, Potenza MN (2007): The neurobiology and genetics of im-pulse control disorders: Relationships to drug addictions. Biochem Phar-macol 75:63–75.
4. Chambers RA, Bickel WK, Potenza MN (2007): A scale-free systems the-ory of motivation and addiction. Neurosci Biobehav Rev 31:1017–1045.
5. Kauer JA, Malenka RC (2007): Synaptic plasticity and addiction. Nat RevNeurosci 8:844 – 858.
6. Grace A, Floresco SB, Goto Y, Lodge DJ (2007): Regulation of firing ofdopaminergic neurons and control of goal-directed behaviors. TrendsNeurosci 30:220 –227.
7. Tseng KY, Chambers RA, Lipska BK (2009): The neonatal ventral hip-pocampal lesion as a heuristic neurodevelopmental animal model ofschizophrenia. Behav Brain Res 204:295–305.
8. Chambers RA, Taylor JR (2004): Animal modeling dual diagnosis schizo-phrenia: Sensitization to cocaine in rats with neonatal ventral hip-pocampal lesions. Biol Psychiatry 56:308 –316.
9. Chambers RA, Jones RM, Brown S, Taylor JR (2005): Natural rewardrelated learning in rats with neonatal ventral hippocampal lesions andprior cocaine exposure. Psychopharmacology (Berl) 179:470 – 478.
10. Chambers RA, Self DW (2002): Motivational responses to natural and drugrewards in rats with neonatal ventral hippocampal lesions: An animal model ofdual diagnosis schizophrenia. Neuropsychopharmacology 27:889–905.
11. Brog JS, Salyapongse A, Deutch AY, Zahm DS (1993): The patterns ofafferent innervation of the core and shell in the “accumbens” part of theventral striatum: Immunohistochemical detection of retrogradely trans-ported fluoro-gold. J Comp Neurol 338:255–278.
12. Haber SN, Fudge JL, McFarland N (2000): Striatonigrostriatal pathwaysin primates form an ascending spiral from the shell to the dorsolateralstriatum. J Neurosci 20:2369 –2382.
13. Everitt BJ, Robbins TW (2005): Neural systems of reinforcement for drugaddiction: From actions to habits to compulsion. Nat Neurosci 8:1481–1489.
14. Lewis DA, Moghaddam B (2006): Cognitive dysfunction in schizophre-nia: Convergence of gamma-aminobutyric acid and glutamate alter-ations. Arch Neurol 63:1372–1376.
15. Gur RE, Cowell PE, Latshaw A, Turetsky BI, Grossman RI, Arnold SE, et al.(2000): Reduced dorsal and orbital prefrontal gray matter volumes inschizophrenia. Arch Gen Psychiatry 57:761–768.
16. Willuhn I, Steiner H (2006): Motor-skill learning-associated gene regulation inthe striatum: Effects of cocaine. Neuropsychopharmacology 31:2669–2682.
17. Carelli RM, West M (1991): Representation of the body by single neuronsin the dorsolateral striatum of the awake, unrestrained rat. J CompNeurol 309:231–249.
18. Saka E, Goodrich C, Harlan P, Madras BK, Graybiel AM (2004): Repetitivebehaviors in monkeys are linked to specific striatal activation patterns.J Neurosci 24:7557–7565.
19. Makris N, Gasic GP, Kennedy DN, Hodge SM, Kaiser JR, Lee JM, et al.(2008): Cortical thickness abnormalities in cocaine addiction-a reflec-tion of both drug use and a pre-existing disposition to drug abuse?Neuron 60:174 –188.
20. Redish AD, Jensen S, Johnson A (2008): A unified framework for addic-tion: Vulnerabilities in the decision process. Behav Brain Sci 31:415– 487.
21. Swanson LW (2004): Brain Maps: Structure of the Rat Brain, 3rd ed. New
York: Elsevier.
IVA
N
acdorHdmim
d7mapsns
pftforpss
7nttbmo
CD
rtrcipd
atJmot
0
N THIS ISSUE
olume 67, Number 8, April 15, 2010
brief summary of the articles appearing in this issue of Biological Psychiatry.icotine Effects: On Mood and In BrainPerkins et al. (pages 707–714) examined smokers randomly
ssigned to one of three smoking conditions (nicotine or deni-otinized cigarettes, or no smoking) while engaged in fourifferent negative mood induction procedures, one per session,r neutral mood. Smoking, regardless of nicotine, robustlyelieved negative affect after overnight smoking abstinence.owever, negative affect relief due to smoking was minimaluring various situational stressors and depended on the affecteasure used. These results suggest that smoking, and nicotine
n particular, alleviates negative affect in a context dependentanner.Cognitive alterations are a core symptom of nicotine with-
rawal, contributing to smoking relapse. Loughead et al. (pages15–721) provide new evidence that the smoking cessationedication varenicline increases working memory-related brain
ctivity during abstinence from smoking, with associated im-rovements in cognitive performance among highly dependentmokers. This finding provides further insight about the role oficotinic compounds on cognition and brain function in humanmokers.
To identify brain regions whose reactivity to smoking cuesredict relapse vulnerability, Janes et al. (pages 722–729) usedunctional magnetic resonance imaging (fMRI) in smokers beforehey attempted to quit smoking. Insula and anterior cingulateMRI reactivity to smoking cues predicted short-term cessationutcomes, suggesting that these regions modulate smokingelapse vulnerability. The findings also suggest that it may beossible to identify smokers likely to relapse before they quitmoking, which may guide treatment assignment and improvemoking cessation outcomes.
Using positron emission tomography, Biegon et al. (pages74–777) sought to evaluate whether there is a direct effect oficotine administration on brain aromatase. They demonstratedhat administration of nicotine results in blockade of aromatase,he enzyme responsible for estrogen synthesis, in the femaleaboon brain in vivo. This finding suggests a novel additionalechanism through which nicotine may exert some of its effectsn behavior and physiology.
ue Reactivity and Reward Processing in AddictiveisordersThere are large individual differences in the degree to which
eward-related cues act as incentive stimuli, spurring individualso action. Here, Saunders and Robinson (pages 730–736)eport that if individual rats find a food cue attractive, a cocaineue is also more effective in motivating drug-taking behavior andnstigating relapse. This suggests that it is possible to determinerior to drug experience which rats will have difficulty resistingrug cues, a trait that may confer vulnerability to addiction.
While the importance of dopamine signaling in the nucleusccumbens (NAc) to reward-seeking is known, several aspects ofhe timing and regulation of this signaling remain unknown.ones et al. (pages 737–744) employed electrochemistry toeasure NAc dopamine signaling in rats engaged in a cue-drivenperant task, in conjunction with pharmacological inactivation of
he basolateral amygdala. The findings demonstrate that basolat-006-3223/10/$36.00
eral amgydala activity modulates dopamine release within theNAc to augment reward-seeking evoked by reward-predictivestimuli.
Environmental cues associated with cocaine evoke sustainedseeking and this process, or cue reactivity, is a critical element ofcocaine addiction. Here Agatsuma et al. (pages 778–780) showthat mice deficient for striatal N-methyl-D-aspartic acid receptor(NMDAR) failed to develop an approach behavior toward cuesassociated with cocaine. This finding indicates that NMDAR levelserves as a determinant for susceptibility to this aspect of cocaineaddiction.
The pathological or problem gambler is a high risk-taker.Infrequent wins are accompanied by frequent losses, yet thegambler persists despite the negative consequences. Hewig et al.(pages 781–783) used measures of event-related brain potentials,recorded during a gambling task, to show that such persistence maybe attributed to a hypersensitive response to the reward associatedwith successful risk-taking. This high-risk behavior may be a sign ofa hyperactive dopaminergic response in gamblers.
Bühler et al. (pages 745–752) studied the mechanismsunderlying drug addiction by assessing the processing of ciga-rette and monetary reward in dependent and non-dependentsmokers using fMRI. Non-dependent smokers showed greateractivation of brain reward circuitry when presented with money-related cues than smoking-related cues. Nicotine-dependentsmokers showed equal activation of these brain circuits underboth conditions. This imbalance may represent a central mech-anism of drug addiction.
Pharmacogenetics of Motivation and BehaviorOff-label prescriptions of modafinil have increased, as re-
search on its putative pro-cognitive effects has increased. Themechanism(s) by which modafinil exerts these effects remainunclear. However, Young and Geyer (pages 784–787) havefound that modafinil increases motivation in mice, mimickingeffects of the dopamine transporter inhibitor GBR12909. Bothdrug effects were attenuated in mice having reduced dopamineD1 receptor expression. The pro-cognitive effects of modafinilmay be due to increased motivation to perform caused by aninhibition of the dopamine transporter and subsequent down-stream effects on D1 receptors.
The neuropeptide orexin has been well-studied for its role insleep as well as drug addiction, but its role in food intake is lesscertain. Here, Sharf et al. (pages 753–760) used genetic, phar-macological, and a unique viral-mediated adult knockdown oforexin in mice to demonstrate a role for orexin in learning andbeing motivated to work for food. Their findings suggest thatactivation of the orexin receptor is a necessary component offood-reinforced responding and/or motivation in normal mice.
Little is known about neurobiological risk factors for devel-oping impulse control disorders, including gambling addiction.Eisenegger et al. (pages 702–706) used a pharmacogeneticapproach in healthy subjects to assess how L-DOPA administra-tion interacts with a genetic risk factor for poor impulse controlto modulate gambling behavior. Results show that variation inthe dopamine receptor D4 gene modulates the impact of dopa-
minergic stimulation on gambling behavior.BIOL PSYCHIATRY 2010;67:697–698© 2010 Society of Biological Psychiatry

C
i7pbcctdc
R
pcp(poirR
698 BIOL PSYCHIATRY 2010;67:697–698
w
ortical-Striatal Circuits in Dual DiagnosisPersons with mental illness, especially schizophrenia, have
ncreased vulnerability to addictions. Chambers et al. (pages88–792) examined the effects of neonatal ventral hippocam-al lesions, an animal model of schizophrenia, and cocaineehavioral sensitization on neural activation levels signaled by-Fos expression. The authors found that the lesions andocaine exposure had a multiplicative effect upon the activa-ion of the dorsal striatum, a brain region associated with theevelopment and expression of habitual behavior and psy-hosis.
egion Specific Actions of RGS4Regulator of G protein signaling 4 (RGS4), linked to neuro-
sychiatric disorders, belongs to a family of proteins known toontrol signaling amplitude and duration via interactions with Grotein alpha subunits or other signaling molecules. Han et al.pages 761–769) used RGS4 knockout mice and mice overex-ressing RGS4 to examine the influence of RGS4 on responses topiates. Behavioral, electrophysiological, and biochemical stud-es reveal that, in the nucleus accumbens, RGS4 acts as a negativeegulator of morphine reward, while in the locus coeruleus,
GS4 opposes morphine physical dependence.ww.sobp.org/journal
Repetitive Transcranial Magnetic Stimulation forBulimic Treatment?
Craving is an important trigger for bingeing behavior, andcraving is associated with dysfunction of the dorsolateral prefrontalcortex. Van den Eynde et al. (pages 793–795) used real or shamrepetitive transcranial magnetic stimulation (rTMS) in people withbulimic eating disorders to evaluate its effect on craving. They foundthat real rTMS of the left frontal brain reduced food craving inresponse to being presented with high caloric foods and binge-eating over the following twenty-four hours, suggesting that rTMSmay be explored as a treatment for bulimic eating disorders.
FAST: New Gene Modulating SystemTanaka et al. (pages 770–773) have created an efficient
system for manipulating gene expression in vivo to rapidlyscreen animal models of disease: FAST (Flexible AcceleratedSTOP TetO-knockin). They obtained two distinct knock-in micefrom a single gene-targeting event, permitting generation ofmouse lines with 5 independent expression patterns. Thus, usingthe FAST system, multiple gain- and loss-of-function mutants canbe generated on a timescale not previously achievable. Thistechnique has widespread implications for investigation of neu-
ropsychiatric disease.