fenobam sulfate inhibits cocaine-taking and cocaine-seeking behavior in rats: implications for...
TRANSCRIPT

ORIGINAL INVESTIGATION
Fenobam sulfate inhibits cocaine-taking and cocaine-seekingbehavior in rats: implications for addiction treatmentin humans
Thomas M. Keck & Hong-Ju Yang & Guo-Hua Bi & Yong Huang &
Hai-Ying Zhang & Ratika Srivastava & Eliot L. Gardner &
Amy Hauck Newman & Zheng-Xiong Xi
Received: 23 December 2012 /Accepted: 30 March 2013# Springer-Verlag Berlin Heidelberg (outside the USA) 2013
AbstractRationale The metabotropic glutamate receptor subtype 5(mGluR5) has been reported to be critically involved in drugreward and addiction. Because themGluR5 negative allostericmodulators (NAMs) 2-methyl-6-(phenylethynyl)pyridine(MPEP) and 3-((2-methyl-1,3-thiazol-4-yl)ethynyl)pyridine(MTEP) significantly inhibit addictivelike behaviors of co-caine and other drugs of abuse in experimental animals, ithas been suggested that mGluR5 NAMs may have transla-tional potential for treatment of addiction in humans.However, neither MPEP nor MTEP have been evaluated inhumans due to their off-target actions and rapid metabolism.Objectives Herein, we evaluate a potential candidate fortranslational addiction research: a new sulfate salt formula-tion of fenobam, a selective mGluR5 NAM that has beeninvestigated in humans.Results In rats, fenobam sulfate had superior pharmacokineticscompared to the free base, with improved maximal plasmaconcentration (Cmax) and longer half life. Oral (p.o.)
administration of fenobam sulfate (30 or 60 mg/kg) inhibitedintravenous (i.v.) cocaine self-administration, cocaine-inducedreinstatement of drug-seeking behavior, and cocaine-associatedcue-induced cocaine-seeking behavior in rats. Fenobam sulfatealso inhibited p.o. sucrose self-administration and sucrose-induced reinstatement of sucrose-seeking behavior, but had noeffect on locomotion.Conclusions This study provides additional support for therole of mGluR5 signaling in cocaine addiction and suggeststhat fenobam sulfate may have translational potential inmedication development for the treatment of cocaine addic-tion in humans.
Keywords Cocaine . Fenobam . Self-administration .
Reinstatement . Cocaine-seeking behavior .
Incubation of cocaine craving . mGluR5
Introduction
Glutamate regulates neuronal activity via ionotropic andmetabotropic glutamate receptor (mGluR) subtypes. AmongmGluRs, members of the Group I family, mGluR1 andmGluR5, are coupled to intracellular phospholipase C via Gq
protein (Schoepp and Conn 1993). mGluR5 is widely expressedin many regions of the brain, including critical mesolimbicstructures such as the ventral tegmental area and the nucleusaccumbens (Romano et al. 1996; Shigemoto et al. 1993).mGluR5 is mostly located postsynaptically (Mitrano and Smith2007) and colocalized with adenosine A2A, dopamine D2, andN-methyl-D-aspartate (NMDA) receptors (Tebano et al. 2005).
mGluR5 became a major target of interest in the potentialtreatment of drug abuse in 2001 when it was reported that mice
T. M. Keck :A. H. Newman (*)Medicinal Chemistry Section, Molecular Targets and MedicationsDiscovery Branch, Baltimore, MD 21224, USAe-mail: [email protected]
H.-J. Yang :G.-H. Bi :H.-Y. Zhang : R. Srivastava :E. L. Gardner : Z.-X. Xi (*)Neuropsychopharmacology Section, Chemical Biology ResearchBranch, Intramural Research Program, National Institute on DrugAbuse, NIH, DHHS, Baltimore, MD 21224, USAe-mail: [email protected]
Y. HuangDepartment of Bioengineering and Therapeutic Sciences,University of California, San Francisco, CA 94143, USA
PsychopharmacologyDOI 10.1007/s00213-013-3106-9

lacking mGluR5 do not self-administer cocaine and that phar-macological blockade of mGluR5 by the negative allostericmodulator (NAM) 2-methyl-6-(phenylethynyl)pyridine(MPEP) inhibited cocaine self-administration (Chiamulera etal. 2001). Attenuation of mGluR5 signaling with MPEP or itsanalog 3-((2-methyl-1,3-thiazol-4-yl)ethynyl)pyridine (MTEP)has been found to inhibit multiple addictive-like behaviors inanimals, including cocaine self-administration (Kenny et al.2005; Lee et al. 2005; Martin-Fardon et al. 2009; Paterson andMarkou 2005; Tessari et al. 2004), cocaine-induced conditionedplace preference (CPP) (Herzig and Schmidt 2004; McGeehanand Olive 2003), cocaine-induced hyperactivity (McGeehan etal. 2004), and cocaine-, cue-, or stress-induced reinstatement ofdrug-seeking behavior (Backstrom and Hyytia 2006;Kumaresan et al. 2009; Lee et al. 2005; Martin-Fardon andWeiss 2012; Wang et al. 2013). These data strongly suggest thatmGluR5 plays an important role in cocaine abuse and thatmGluR5 NAMsmay have potential for the treatment of cocaineaddiction in humans (Heidbreder et al. 2003; Olive et al. 2005).
However, there are multiple reasons that MPEP and MTEPhave not advanced to clinical trials (Lindsley and Emmitte2009). MPEP blocks the norepinephrine transporter(Heidbreder et al. 2003), NMDA receptors (Movsesyan et al.2001; O'Leary et al. 2000), and monoamine oxidase A (Leaand Faden 2006), while activating mGluR4 (Mathiesen et al.2003). In addition, MTEP was reported to inhibit cytochromeP450 1A2 (Green et al. 2004), and both MPEP and MTEP arerapidlymetabolized after injection. Therefore, extensive effortshave been undertaken to develop novel potent, selective, met-abolically stable mGluR5 NAMs for use in humans (Emmitte2011; Nicoletti et al. 2011; Rocher et al. 2011).
Fenobam, a brain-penetrant nonbenzodiazepine anxiolyticadvanced to Phase II clinical trials in the early 1980s(Friedmann et al. 1980; Itil et al. 1978; Pecknold et al. 1982),has been identified as a selective mGluR5 NAM (Porter et al.2005). Fenobam was granted orphan drug status by the Foodand Drug Administration (FDA) in 2006 and has been recentlyinvestigated as an experimental clinical treatment for Fragile XSyndrome (Berry-Kravis et al. 2009) and L-DOPA-induceddyskinesias (Rylander et al. 2010). Fenobam was recentlyreported to inhibit brain stimulation reward (Cleva et al.2012) and attenuate methamphetamine-seeking behavior in rats(Watterson et al. 2013). Considering these results, as well as thesuccess of buprenorphine as an orphan drug approved fortreating morphine dependence (Jaffe and O'Keeffe 2003), andfenobam's superior mGluR5 selectivity compared to MPEP(Montana et al. 2009), fenobam is an intriguing clinical candi-date for the treatment of psychostimulant dependence.However, it is unclear whether fenobam inhibits cocaine-taking and cocaine-seeking behaviors as has been demonstrat-ed with MPEP and MTEP. In addition, clinical tests haveutilized the free base monohydrate formulation of fenobamand variability in response to fenobam has been correlated with
high variability in plasma levels of the drug after oral (p.o.)administration (Itil et al. 1978; Berry-Kravis et al. 2009).Therefore, development of new formulations of fenobam withimproved oral bioavailability was deemed important for use infuture clinical trials.
In this study, we first compared the pharmacokinetics andoral bioavailability of fenobam free base and fenobam sul-fate, a new salt formulation of fenobam, and then evaluatedthe pharmacological effects of fenobam sulfate on intrave-nous (i.v.) cocaine self-administration, cocaine-induced re-instatement of drug-seeking behavior and cue-inducedcocaine-seeking behavior in rats—three commonly usedanimal models to evaluate drug reward, craving, and relapse(O'Brien and Gardner 2005). We also observed the effects offenobam sulfate on p.o. sucrose self-administration andsucrose-induced reinstatement of sucrose-seeking behaviorand locomotor activity. Overall, our aim was to evaluatefenobam sulfate in rat models of cocaine addiction and,thus, determine the potential for development of fenobamsulfate as a pharmacological treatment for cocaine addiction.
Materials and methods
Drugs
Fenobam free base and fenobam sulfate were provided by theDivision of Pharmacotherapies and Medical Consequences ofDrug Abuse (DPMCDA), National Institute on Drug Abuse(NIDA). Cocaine HCl was provided by NIDA and dissolved inphysiological saline. Since fenobam free base has low solubil-ity in 40 % hydroxypropyl-β-cyclodextrin (BCD), it was firstdissolved in 10 % dimethylsulfoxide (DMSO) and brought tofinal concentration in 40 % BCD; fenobam sulfate wasdissolved in 40 % BCD. There are few reports that such alow concentration (10 %) of DMSO alters the pharmacokineticprofile of drugs. Therefore, it is commonly used in animalpharmacokinetic studies to increase drug solubility. In all be-havioral studies, fenobam sulfate was dissolved in 40 % BCDand water for p.o. administration.
Experiment 1: pharmacokinetic comparison of fenobam freebase and fenobam sulfate
Pharmacokinetic data were obtained under NIDA ContractN01DA-9-8883. Briefly, two groups of Sprague–Dawleyrats (Charles River Laboratories, Raleigh, NC, USA, n=3per sex, 6 per group, 12 animals in total) were used. I.v.catheterization of the right external jugular vein for bloodsampling was performed under sodium pentobarbital(60 mg/kg, i.p.) anesthesia, utilizing standard aseptic surgi-cal techniques as we reported previously. After recoveryfrom surgery, each animal received a single p.o. dose of
Psychopharmacology

30 mg/kg of fenobam free base or fenobam sulfate. The doseadministered of each test material, fenobam free base orfenobam sulfate, was based on the anhydrous free baseequivalents of fenobam. Blood samples were collected viajugular vein catheter prior to dosing and after dosing atdifferent time points (0.25, 0.5, 1, 2, 3, 4, 8, 12, and 24 hafter p.o. administration). Blood samples were centrifugedto obtain plasma and all samples were stored at −20 °C untilanalysis. Fenobam concentrations in plasma samples wereanalyzed by a validated liquid chromatography/tandemmass spectrometry (LC/MS) method with a quantizationlimit of 5 ng/ml. Rat plasma samples (50 μl) were analyzedfor fenobam with an LC/MS procedure in a Sciex API 3000system equipped with a Synergi Polar-RP column (4.6×75 mm, 4 μm particle size), a mobile phase system consistingof acetonitrile:water:formic acid (42.5:57.5:0.01) (v:v:v) with5 mM ammonium acetate and mass spectrometric detec-tion with positive ionization by atmospheric pressurechemical ionization (APCI) and mass scanning by mul-tiple reaction monitoring (MRM) analysis. The transitionions for fenobam were m/z 267>114. Sample prepara-tion consisted of addition of internal standard(bupropion), extraction of 50 μl of plasma with1.00 ml of methyl t-butyl ether prior to separation byLC/MS/MS. The assay was validated from 5.00 to1,000 ng/ml. The coefficients of variance and relativeerror for low, mid, and high concentrations of qualitycontrols were <15 %.
The PK parameters of peak plasma concentration (Cmax),half-life (T1/2), and area under the curve (AUC) were calculatedusing WinNonlin software (Professional Edition, 5.2.1 ver.,Pharsight Corp., CA, USA) in the noncompartmental mode.Cmax was defined as the highest level of plasma fenobamamong nine time points detected. AUC was calculated bysummation of plasma drug concentrations at all nine timepoints. Cmax and AUC in both fenobam free base and fenobamsulfate groups were compared using unpaired, two-tailed t tests.
Experiment 2: cocaine self-administration
Animals
Male Long–Evans rats (250–300 g; Charles RiverLaboratories, Raleigh, NC, USA) were used for all experi-ments. They were individually housed in a climate-controlled room on a reverse light–dark cycle (lights on at1900 h, lights off at 0700 h) with ad libitum access to foodand water. All experimental procedures were conducted inaccordance with the Guide for the Care and Use ofLaboratory Animals (US National Academy of Sciences)and were approved by the Animal Care and UseCommittee of the National Institute on Drug Abuse of theUS National Institutes of Health. The animal facility was
fully accredited by the Association for Assessment andAccreditation of Laboratory Animal Care International.
Surgery
I.v. catheterization of the right external jugular vein wasperformed under sodium pentobarbital (60 mg/kg, i.p.) anes-thesia, utilizing standard aseptic surgical techniques as wereported previously (Xi et al. 2006a, b). To help prevent clog-ging, the catheters were flushed daily with a gentamicin–heparin–saline solution (0.1 mg/ml gentamicin, 30 IU/ml hep-arin; ICN Biochemicals, Cleveland, OH, USA).
Apparatus
Self-administration chambers from Med Associates Inc.(Saint Albans, VT, USA) were the same as used previously(Xi et al. 2006a, b).
Self-administration training
After recovery from surgery, each rat was placed into a testchamber and allowed to lever press for i.v. cocaine(1 mg/kg/infusion) on a fixed ratio 1 (FR1) reinforcementschedule. Each cocaine infusion delivered a volume of0.08 ml/infusion over 4.65 s and was paired with the simul-taneous presentation of a stimulus light and tone (eachlasting 4.65 s). All self-administration sessions lasted 3 h.FR1 reinforcement was used for 6 days to establish stableself-administration. Subjects were then allowed to continuecocaine (0.5 mg/kg/infusion) self-administration under FR2reinforcement. To avoid cocaine overdose during the self-administration period, each animal was limited to a maxi-mum of 50 cocaine injections per 3-h session. After stableself-administration was established on the FR2 schedule(less than 10 % variation in total infusions over three con-secutive days), the effects of fenobam sulfate (0, 30, and60 mg/kg, p.o.) on cocaine self-administration were deter-mined in one group of rats (n=11). Fenobam sulfate or vehiclewas administered 30 min prior to self-administration testing.Each animal received three doses of fenobam sulfate over aweek of testing, with the order of testing for various dosescounterbalanced. After each drug test, animals continued dailycocaine self-administration until stable self-administrationwas re-established; typically, stable self-administration wasre-established within 1–2 days. Each animal received a totalof three injections corresponding to three drug doses in eachgroup.
Multiple-dose cocaine self-administration training
To determine whether the pharmacological action offenobam sulfate was also cocaine dose-dependent, we
Psychopharmacology

further studied the effects of fenobam sulfate on cocaine self-administration maintained by a full range of cocaine doses(0.03, 0.06, 0.125, 0.25, and 0.5 mg/kg/infusion) in a singlesession (Mantsch et al. 2007; Hiranita et al. 2009; Song et al.2012). The session consisted of five sequential 20-min com-ponents, each preceded by a 20-min time-out period for chang-ing cocaine dose. The infusion volumes and durations in eachcomponent were identical, except that cocaine concentrationsfor corresponding unit cocaine doses differed. There was a 30-min extinction period (0 mg/kg cocaine) before daily cocaineself-administration training. Training continued until: (1) aminimum of 5.0 mg/kg cocaine was self-administered withina session with less than 20 % variation in the total number ofcocaine injections compared with the previous session, (2) thedose of cocaine that maintained maximal response rates variedby no more than one-half log unit over two consecutive testsessions, and (3) maximal response rates were at least five-foldhigher than response rates maintained during extinction. Then,each rat randomly received either vehicle or one of two dosesof fenobam sulfate (30 and 60 mg/kg, p.o.) 30 min prior to thetest session. Animals then received an additional 3–4 days ofself-administration of cocaine alone until the baseline responserate was re-established prior to testing the next dose offenobam sulfate. The order of testing for the various doses ofdrug or vehicle was counterbalanced.
Experiment 3: sucrose self-administration
The procedures for sucrose self-administration were identicalto the procedures for cocaine self-administration, except forthe following: (1) no surgery was performed on the animals,(2) active lever presses led to delivery of 0.1 ml of 5 % sucrosesolution into a liquid food tray on the operant chamber wallalong with presentation of a stimulus light and tone, (3) 3-hFR1 training sessions were capped at 100 deliveries, and (4)test sessions were 90-min in length and capped at 100 deliv-eries. After stable sucrose self-administration was establishedon the FR1 schedule (less than 10 % variation in total deliv-eries over three consecutive days), the effects of fenobamsulfate (0, 30, and 60 mg/kg, p.o.) on sucrose self-administration were determined in one group of rats (n=7).Each animal received three doses of fenobam sulfate over aweek of testing, with the order of testing for various dosescounterbalanced. After each drug test, animals continued dailysucrose self-administration until stable self-administrationwas re-established; typically, stable self-administration wasre-established within 1–2 days.
Experiment 4: sucrose-triggered reinstatementof sucrose-seeking behavior
Two additional group of rats (n=11 total) were used toevaluate the effects of fenobam sulfate on sucrose-induced
reinstatement of sucrose-seeking behavior. After stable su-crose self-administration was achieved, animals underwentextinction training until sucrose-seeking behavior wasextinguished. To determine whether fenobam sulfatepretreatment inhibits sucrose-seeking behavior, extinguishedanimals were treated with fenobam sulfate or vehicle prior tothe triggering of reinstatement via five noncontingent sucrosedeliveries. The animals were divided into two dose groups.Group 1 (n=6) randomly received vehicle or 30 mg/kg offenobam sulfate and received the matched treatment 6 dayslater after two intervening extinction trials. Group 2 (n=5)randomly received vehicle or 60 mg/kg of fenobam sulfateand received the matched treatment 6 days later after twointervening extinction trials. Lever presses during the rein-statement tests were recorded but did not lead to either sucrosedelivery or presentation of the conditioned cue light and tone.
Experiment 5: cocaine-primed reinstatementof cocaine-seeking behavior
After the completion of Experiment 1, animals continuedcocaine self-administration for at least 3–5 days until stableself-administration was re-established. Then, the animalsunderwent extinction training, during which cocaine wasreplaced by saline and the light and sound cues that previ-ously accompanied cocaine infusions were turned off. Afterdrug-seeking behavior was extinguished, defined as ≤10active lever presses during each 3-h session for at least threeconsecutive days, the animals were divided into three dosegroups (between-subjects design) to evaluate the effects offenobam sulfate on cocaine-primed reinstatement of drug-seeking behavior (n=9–10 per group). On the reinstatementtest day, each group of animals received one dose (0, 30, or60 mg/kg, p.o.) fenobam sulfate alone or 30 min prior tococaine priming (10 mg/kg, i.p.). Then, the animals wereplaced into the operant chambers that were previouslypaired with cocaine self-administration. Reinstatement con-ditions were identical to those in the extinction sessions, i.e.,active lever presses were recorded without drug infusions oraccompanying cues for 3 h. Effects of fenobam sulfate ondrug-seeking or cocaine-induced reinstatement wereassessed by comparing the mean number of active leverpresses per test session.
Experiment 6: contextual cue-induced cocaine seeking
Two additional groups of rats were used in this experiment.One group of rats (n=32) was used to evaluate contextualcue-induced incubation of cocaine craving by comparingextinction responding after 2 or 21 days (n=16 rats in eachsubgroup) of withdrawal from cocaine self-administration.Another group of rats (n=24) was used to examine theeffects of fenobam sulfate (0, 30, and 60 mg/kg, p.o.,
Psychopharmacology

30 min prior to testing) on contextual cue-induced incuba-tion of cocaine craving (between-subjects design,n=6–8 each subgroup). On the test day, rats were placed intothe same self-administration chambers in which they weretrained, and cocaine cue-induced drug-seeking behavior (i.e.,extinction responding) was assessed under extinction condi-tions during which cocaine and cocaine-associated discrete cuelight and tone were unavailable, and lever pressing resulted inno consequences. Each extinction session lasted 3 h.
Experiment 7: effects of fenobam sulfate on locomotorbehavior
Finally, to determine whether the reduction in cocaine-taking and cocaine-seeking behavior was due to nonspecificlocomotor impairment or sedative effects, we observed theeffects of the same doses of fenobam sulfate on basal loco-motor behavior in a separate group of rats (n=8). Beforetesting, drug-naïve rats were habituated in a locomotordetection chamber (AccuScan, Columbus, OH, USA) for1 h each day on two consecutive days. On each testingday, 1-h basal levels of locomotor activity were recordedprior to vehicle or fenobam sulfate administration; drug-induced changes in locomotor behavior were recorded foran additional 3 h. Each animal was tested three times withdifferent drug doses. The order of testing for various dosesof fenobam sulfate was counterbalanced. The time intervalsbetween testing were 2–3 days.
Data analysis
All data are presented as means±SEM. One-way ANOVAwas used to determine the significance of the changes inreward-taking or reward-seeking behavior after drug admin-istration. Whenever a significant main effect was found,individual group comparisons were carried out usingpreplanned Bonferroni t tests. Paired two-tailed t tests wereused to compare the differences in fenobam sulfate effectson sucrose reinstatement. Unpaired two-tailed t tests wereused to compare the differences in Cmax and AUC betweenfenobam free base and fenobam sulfate treatment groups.
Results
Fenobam sulfate has superior absorption characteristicscompared to fenobam free base
Figure 1 illustrates the comparative pharmacokinetic prop-erties of fenobam free base and fenobam sulfate. Fenobamsulfate had a higher Cmax (Fig. 1a: p<0.05) and longeraverage T1/2 [3.47±2.57 (mean±SD) versus 0.4±0.15 h infemale rats (n=3); 1.27±1.28 versus 1.37±1.77 in male rats
(n=3)] than fenobam free base. Since the T1/2 data showslarge variability across animals, we calculated the AUC ofthe plasma fenobam concentrations in each animal andpooled data from male and female rats together to increasethe power for statistical analysis. We found that the AUCvalues in fenobam sulfate-treated rats were significantlyhigher than those in fenobam free base-treated rats(Fig. 1b: p<0.01). These data suggest that the sulfate for-mulation of fenobam has higher oral bioavailability thanfenobam free base.
Fenobam sulfate inhibits cocaine self-administration
Figure 2 illustrates the effects of fenobam sulfate on cocaineself-administration, demonstrating that p.o. administrationof fenobam sulfate (30–60 mg/kg) significantly inhibitedcocaine self-administration in a dose-dependent manner(Fig. 2a: F2,10=5.36, p<0.05). P.o. administration offenobam sulfate also significantly reduced active leverresponding in a dose-dependent manner (Fig. 2b: F2,10=8.05, p<0.01). Individual group comparisons demonstratedthat fenobam sulfate, at 60 mg/kg, significantly decreasedcocaine infusions (t=3.22, p<0.05) and active lever presses(t=4.01, p<0.01). These effects lasted for less than 24 h ascocaine self-administration on the following day returned toprevious stable levels. There were no significant differencesin inactive lever responding across treatments (Fig. 2b).
Figure 2c illustrates that fenobam sulfate, at the same doses(30–60 mg/kg, p.o.), significantly and dose-dependentlyinhibited cocaine self-administration maintained by lower dosesof cocaine (0.06, 0.125, and 0.25 mg/kg/infusion) and shiftedthe cocaine dose–response self-administration curve downward.Two-way ANOVA with two-factor repeated measures overcocaine and fenobam sulfate doses revealed a significant treat-ment (vehicle versus fenobam) main effect (F2,75=10.98,p<0.01), significant time (cocaine dose) main effect (F5,75=7.69, p<0.001) and a significant treatment×time interaction(F10,75=5.95, p<0.001). Individual group comparisons revealeda statistically significant reduction in cocaine self-administrationfollowing 30 mg/kg fenobam treatment at cocaine doses of 0.06(t=5.98, p<0.001) and 0.125 mg/kg/infusion cocaine (t=4.76,p<0.001) and following 60mg/kg fenobam treatment at cocainedoses of 0.06 (t=6.47, p<0.001), 0.125 (t=5.11, p<0.001), and0.25 mg/kg/infusion cocaine (t=2.90, p<0.05), when comparedto the vehicle control group.
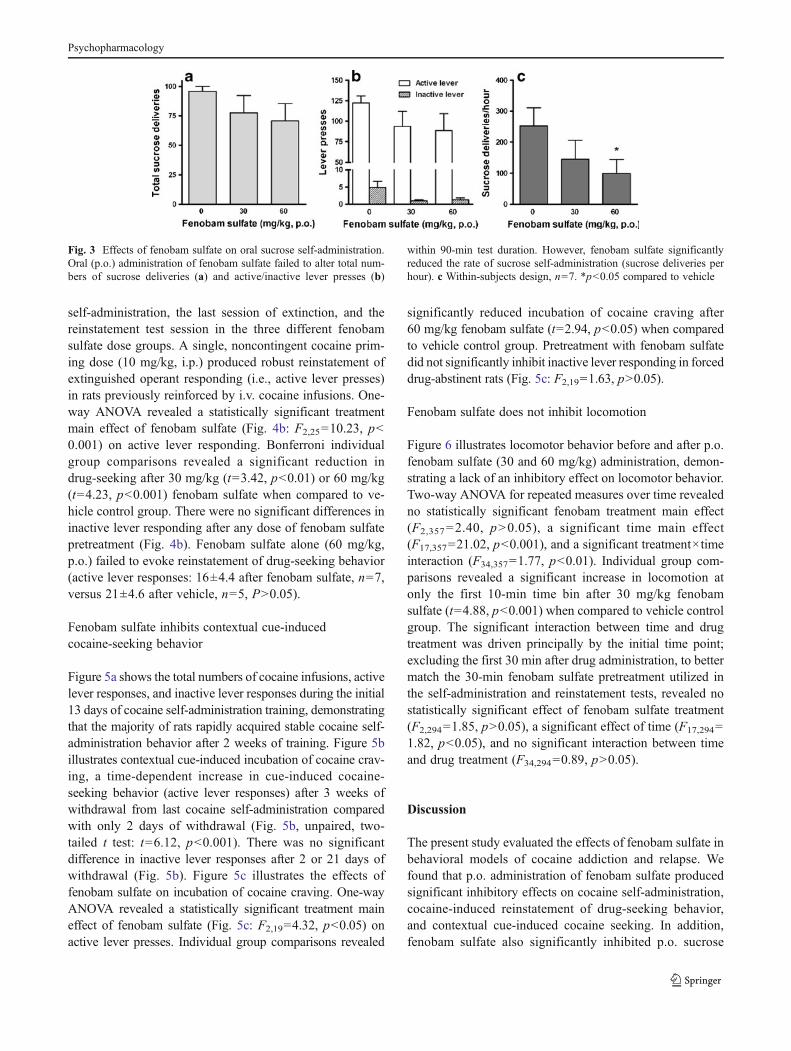
Fenobam sulfate inhibits oral sucrose-takingand sucrose-seeking behavior
Figure 3 illustrates the effects of fenobam sulfate on p.o.sucrose self-administration. Systemic administration offenobam sulfate failed to alter the total number of sucrosedeliveries (Fig. 3a: F2,6=1.14, p>0.05) or the total numbers of
Psychopharmacology

active lever presses (Fig. 3b: F2,6=1.46, p>0.05). Althoughone-way ANOVA revealed a statistically significant treatmentmain effect of fenobam on inactive lever responding (Fig. 3b:F2,6=4.34, p<0.05), post hoc individual group comparisonsrevealed no significant reduction in inactive lever respondingafter 30mg/kg (t=2.65, p>0.05) or 60 mg/kg (t=2.45, p>0.05)fenobam sulfate when compared to vehicle control group.
Since sucrose deliveries were limited to maximal 100deliveries during 90-min testing and the majority of animalscompleted the maximal 100 deliveries in less time, werenormalized the data to the rate of sucrose deliveries perhour. The normalized data show that systemic administra-tion of fenobam sulfate significantly and dose-dependentlyinhibited p.o. sucrose self-administration (Fig. 3c: F2,6=5.19, p<0.05). Individual group comparisons revealed a
significant reduction in the rate of sucrose delivery after60 mg/kg fenobam sulfate (t=3.14, p<0.05).
Figure 4a shows the effects of fenobam sulfate on rein-statement of sucrose-seeking behavior. Pretreatment with60 mg/kg (paired, two-tailed t test: t=3.22, p<0.05), but not30 mg/kg (t=0.55, p>0.05), fenobam sulfate significantlyinhibited sucrose-induced reinstatement of sucrose-seekingbehavior. There were no statistically significant variations inthe total inactive lever responses across treatments.
Fenobam sulfate inhibits cocaine-primed reinstatementof cocaine-seeking behavior
Figure 4b illustrates the total number of active and inactivelever presses observed during the last session of cocaine
Fig. 1 Blood fenobam concentrations following oral administration of30 mg/kg fenobam free base or fenobam sulfate. (a) Cmax, maximal(peak) plasma concentration, and (b) AUC, area under the curve of
plasma fenobam concentrations over time (all nine time points within24 h) after oral administration of 30 mg/kg fenobam free base orfenobam sulfate. Free base, n=6; sulfate, n=6. *p<0.05, **p<0.01
Fig. 2 Effects of fenobamsulfate on intravenous cocaineself-administration. Oral (p.o.)administration of fenobamsulfate significantly inhibitedcocaine self-administration (a)and active lever responses (b) inrats. Within-subjects design, n=11. *p<0.05, **p<0.01compared to vehicle. It alsodose-dependently shiftedcocaine dose–response self-administration curve downward(c; vehicle (filled square),30 mg/kg (unfilled triangle),60 mg/kg (unfilledcircle)).Within-subjects design,n=6; 30 mg/kg fenobam versusvehicle: †††p<0.001; 60 mg/kgfenobam versus vehicle: *p<0.05, ***p<0.001
Psychopharmacology
self-administration, the last session of extinction, and thereinstatement test session in the three different fenobamsulfate dose groups. A single, noncontingent cocaine prim-ing dose (10 mg/kg, i.p.) produced robust reinstatement ofextinguished operant responding (i.e., active lever presses)in rats previously reinforced by i.v. cocaine infusions. One-way ANOVA revealed a statistically significant treatmentmain effect of fenobam sulfate (Fig. 4b: F2,25=10.23, p<0.001) on active lever responding. Bonferroni individualgroup comparisons revealed a significant reduction indrug-seeking after 30 mg/kg (t=3.42, p<0.01) or 60 mg/kg(t=4.23, p<0.001) fenobam sulfate when compared to ve-hicle control group. There were no significant differences ininactive lever responding after any dose of fenobam sulfatepretreatment (Fig. 4b). Fenobam sulfate alone (60 mg/kg,p.o.) failed to evoke reinstatement of drug-seeking behavior(active lever responses: 16±4.4 after fenobam sulfate, n=7,versus 21±4.6 after vehicle, n=5, P>0.05).
Fenobam sulfate inhibits contextual cue-inducedcocaine-seeking behavior
Figure 5a shows the total numbers of cocaine infusions, activelever responses, and inactive lever responses during the initial13 days of cocaine self-administration training, demonstratingthat the majority of rats rapidly acquired stable cocaine self-administration behavior after 2 weeks of training. Figure 5billustrates contextual cue-induced incubation of cocaine crav-ing, a time-dependent increase in cue-induced cocaine-seeking behavior (active lever responses) after 3 weeks ofwithdrawal from last cocaine self-administration comparedwith only 2 days of withdrawal (Fig. 5b, unpaired, two-tailed t test: t=6.12, p<0.001). There was no significantdifference in inactive lever responses after 2 or 21 days ofwithdrawal (Fig. 5b). Figure 5c illustrates the effects offenobam sulfate on incubation of cocaine craving. One-wayANOVA revealed a statistically significant treatment maineffect of fenobam sulfate (Fig. 5c: F2,19=4.32, p<0.05) onactive lever presses. Individual group comparisons revealed
significantly reduced incubation of cocaine craving after60 mg/kg fenobam sulfate (t=2.94, p<0.05) when comparedto vehicle control group. Pretreatment with fenobam sulfatedid not significantly inhibit inactive lever responding in forceddrug-abstinent rats (Fig. 5c: F2,19=1.63, p>0.05).
Fenobam sulfate does not inhibit locomotion
Figure 6 illustrates locomotor behavior before and after p.o.fenobam sulfate (30 and 60 mg/kg) administration, demon-strating a lack of an inhibitory effect on locomotor behavior.Two-way ANOVA for repeated measures over time revealedno statistically significant fenobam treatment main effect(F2,357=2.40, p>0.05), a significant time main effect(F17,357=21.02, p<0.001), and a significant treatment×timeinteraction (F34,357=1.77, p<0.01). Individual group com-parisons revealed a significant increase in locomotion atonly the first 10-min time bin after 30 mg/kg fenobamsulfate (t=4.88, p<0.001) when compared to vehicle controlgroup. The significant interaction between time and drugtreatment was driven principally by the initial time point;excluding the first 30 min after drug administration, to bettermatch the 30-min fenobam sulfate pretreatment utilized inthe self-administration and reinstatement tests, revealed nostatistically significant effect of fenobam sulfate treatment(F2,294=1.85, p>0.05), a significant effect of time (F17,294=1.82, p<0.05), and no significant interaction between timeand drug treatment (F34,294=0.89, p>0.05).
Discussion
The present study evaluated the effects of fenobam sulfate inbehavioral models of cocaine addiction and relapse. Wefound that p.o. administration of fenobam sulfate producedsignificant inhibitory effects on cocaine self-administration,cocaine-induced reinstatement of drug-seeking behavior,and contextual cue-induced cocaine seeking. In addition,fenobam sulfate also significantly inhibited p.o. sucrose
Fig. 3 Effects of fenobam sulfate on oral sucrose self-administration.Oral (p.o.) administration of fenobam sulfate failed to alter total num-bers of sucrose deliveries (a) and active/inactive lever presses (b)
within 90-min test duration. However, fenobam sulfate significantlyreduced the rate of sucrose self-administration (sucrose deliveries perhour). c Within-subjects design, n=7. *p<0.05 compared to vehicle
Psychopharmacology

self-administration and sucrose-induced reinstatement. Thepresent findings, combined with the use of fenobam inhuman clinical studies, suggest that fenobam sulfate is apotential candidate for translational development in thetreatment of cocaine addiction.
The behavioral inhibition in the above experiments wasunlikely due to nonspecific sedation or locomotor impair-ment because fenobam sulfate did not inhibit open fieldlocomotor activity. Indeed, there was a significant increase
in locomotion within 10 min after p.o. fenobam sulfateadministration. However, we do not think that this increasein locomotion contributed to the effects on cocaine- orsucrose-driven self-administration and reinstatement behav-iors because the operant behavioral tests began 30 min afterfenobam sulfate administration; no fenobam effect on loco-motor activity was detectable after 30 min.
Drug addiction is characterized by compulsive drug-taking and drug-seeking behavior after abstinence (Gawin
Fig. 4 Effects of fenobam sulfate on sucrose-induced reinstatement ofreward-seeking behavior and cocaine-primed reinstatement of drug-seek-ing behavior. (a) Two groups of animals (vehicle and 30 mg/kg, n=6;vehicle and 60 mg/kg, n=5) were trained to self-administer sucrose andthen sucrose-taking/sucrose-seeking behavior was extinguished.Pretreatment with 60 mg/kg, but not 30 mg/kg, fenobam sulfate signifi-cantly inhibited reinstatement of extinguished sucrose-seeking behavior
following noncontingent sucrose delivery. (b) Three groups of animals(vehicle, n=10; 30 mg/kg fenobam sulfate, n=9; 60 mg/kg fenobamsulfate, n=9) were trained to self-administer cocaine and then cocaine-taking/cocaine-seeking behavior was extinguished. Pretreatment withfenobam sulfate (0, 30, or 60 mg/kg, p.o.) dose-dependently inhibitedcocaine-induced reinstatement of extinguished drug-seeking behavior inrats. *p<0.05, **p<0.01, ***p<0.001, compared to vehicle
Fig. 5 Effects of fenobam sulfate on contextual cue-induced incuba-tion of cocaine craving. (a) Total numbers of cocaine infusions(unfilled circle), active (filled square), and inactive lever (unfilledtriangle) responses during initial 13 days of cocaine self-administrationtraining. (b) Extinction responses after 2 or 21 days of withdrawal fromcocaine self-administration (n=16 rats in each subgroup), illustrating a
progressive increase in cue-induced cocaine-seeking (i.e., active leverpresses) over time of withdrawal. (c) Pretreatment with fenobam sul-fate (0, 30, or 60 mg/kg, p.o.) inhibited cue-induced cocaine-seekingbehavior. Fenobam sulfate test: vehicle, n=6; 30 mg/kg, n=8; 60 mg/kg, n=8. *p<0.05, ***p<0.001
Psychopharmacology

and Kleber 1986; Satel et al. 1991). I.v. drug self-administration is one of the most commonly used animalmodels for studying a drug's reinforcing effects (Panlilio andGoldberg 2007). The present finding that fenobam sulfatesignificantly inhibits cocaine self-administration is consis-tent with previous studies using the prototypic MPEP, inwhich it was found that MPEP significantly inhibits cocaineself-administration under fixed-ratio reinforcement in mice(Chiamulera et al. 2001) or under progressive-ratio rein-forcement in rats (Paterson and Markou 2005), cocaine-induced CPP (Herzig and Schmidt 2004; McGeehan andOlive 2003), cocaine-induced hyperlocomotion (Herzig andSchmidt 2004), and cocaine-induced locomotor sensitiza-tion (Dravolina et al. 2006). The neural mechanisms under-lying the antagonism of cocaine reward by mGluR5 NAMsare not fully understood. Several studies suggest that block-ade of mGluR5 decreases brain reward functioning as mea-sured by electrical intracranial self-stimulation (Cleva et al.2012; Kenny et al. 2005), suggesting that a diminishedrewarding response to cocaine may underlie the antagonismof cocaine self-administration after mGluR5 blockade.
Two animal models are commonly used to evaluate drug-or cue-induced relapse to drug-seeking behavior (Grimm et al.2001; O'Brien and Gardner 2005). One is drug priming-induced reinstatement (relapse) of drug-seeking behavior, inwhich animals are first trained for cocaine self-administrationand then drug-seeking behavior is extinguished in the absenceof cocaine and cocaine-paired cues (light and tone). Rats arethen tested for cocaine priming-induced reinstatement(relapse) of extinguished drug-seeking behavior (O'Brienand Gardner 2005). The second model is incubation of crav-ing, in which animals are forcibly withdrawn from cocaineself-administration without behavioral extinction of the
previously reinforced operant responding. After varying pe-riods of withdrawal time in their home cages, animals are re-exposed to the environmental context previously paired withdrug self-administration, i.e., the self-administration cham-bers. Subsequent lever responding (i.e., extinctionresponding) leads to no consequence (no drug, no conditionedcues) or to only conditioned cues (Lu et al. 2004). Strikingly,there is a progressive increase in drug seeking over duration ofwithdrawal. Since the time course of the increase in cocaineseeking is somewhat similar to that of the expression ofpsychostimulant sensitization after withdrawal and that ofdrug craving in humans during abstinence, it has been termed“cue-induced incubation of cocaine craving” (Grimm et al.2001; Lu et al. 2004).
Because fenobam is a selective mGluR5 NAM (Porter et al.2005) and has had some success in clinical trials for treatmentof anxiety and Fragile X Syndrome (Friedmann et al. 1980;Pecknold et al. 1982; Berry-Kravis et al. 2009), it is an intrigu-ing candidate for possible clinical development for the treat-ment of other neuropsychiatric disorders, includingpsychostimulant addiction (Carroll, 2008). However, previousclinical work utilizing fenobam free base monohydrate waslimited by variable bioavailability (Itil et al. 1978; Berry-Kravis et al. 2009). The low water solubility of fenobam freebasemay have contributed to its inconsistent absorption profile,whereas fenobam sulfate has improved water solubility thatmay contribute to its greater overall bioavailability in ratscompared to the free base as measured byCmax, T1/2, and AUC.
An important finding in the present study is that systemicadministration of fenobam sulfate significantly inhibited co-caine self-administration in a dose-dependent manner. This isconsistent with previous reports using MPEP or MTEP(Kenny et al. 2005; Lee et al. 2005; Martin-Fardon et al.2009; Paterson andMarkou 2005). To determine whether sucha reduction in drug-taking behavior was due to a reduction incocaine's rewarding strength, we observed the effects offenobam sulfate on cocaine self-administration dose–responsecurve based upon a well-accepted view that a leftward orupward shift of a dose–response curve is usually interpretedas an increase in pharmacological action and vice versa(Hiranita et al., 2009). We found that fenobam sulfate dose-dependently shifted the cocaine self-administration dose–re-sponse curve downward, suggesting a reduction in cocaine'srewarding efficacy after mGluR5 blockade. We note that awithin-session, not between-session, multiple-dose cocaineself-administration paradigm was used in the present studybecause it is less time-consuming and more resource (drugs,animals)-efficient with fewer concerns about tolerance orsensitization caused by repeated drug administration used inbetween-session experiments (Bentzley et al., 2013). Themajor weaknesses include: (1) the cocaine dose–responsecurve observed in a within-session design may not preciselyreveal the dose–response relationship because residual drug
Fig. 6 Effects of fenobam sulfate on locomotor activity. Compared tovehicle (filled square), fenobam sulfate, at 30 mg/kg (unfilled triangle),but not 60 mg/kg (unfilled circle), produced a transient increase inlocomotion within the initial 10 min, but not at any other time pointsfollowing oral drug administration. Within-subjects design, n=8.***p<0.001 compared to vehicle
Psychopharmacology

may still be present in the brain before the next cocaine dosetesting, which may overlay and interact with subsequent doseinjections and (2) if the time course of the pharmacologicalaction of a tested drug (for example, fenobam) is short than theduration (3 h) of multiple-dose self-administration testing, itmay produce an artifactual diminution of the effects of thetested drug on higher doses of cocaine tested later inthe paradigm. Given that very low doses (0.125–1.0 mg/kg/infusion) of cocaine were used in self-administration, suggesting that brain cocaine may be rapidlycleared after each testing and that the pharmacological actionof many testing drugs last at least 2–3 h, the within-sessionmultiple-dose of cocaine self-administration paradigm is rou-tinely used in this field (Mantsch et al.,2007; Hiranita et al.2009; Song et al. 2012).
In addition, we also found that fenobam sulfate signifi-cantly and dose-dependently inhibited cocaine priming-induced reinstatement of drug-seeking behavior. This isconsistent with previous reports that MPEP (Backstromand Hyytia 2007; Lee et al. 2005) or MTEP (Kumaresanet al. 2009; Martin-Fardon et al. 2009; Martin-Fardon andWeiss 2012; Wang et al. 2013) inhibit cocaine-, cue-, orstress-induced reinstatement of drug-seeking behavior. Inaddition, the present study also, for the first time, demon-strated that blockade of mGluR5s by fenobam sulfateinhibited contextual cue-induced cocaine seeking in rats,supporting an important role of mGluR5 in relapse todrug-seeking behavior. The neural mechanisms underlyingthese actions are not fully understood. Previous studiessuggest that cocaine priming, footshock stress, or re-exposure to cocaine-associated cues evokes glutamate re-lease within the ventral tegmental area (VTA) and nucleusaccumbens (NAc) (McFarland et al. 2003; Miguens et al.2008; Wang et al. 2005, 2007; Xi et al. 2006a; You et al.2007), which has been shown to play an important role inrelapse to drug-seeking behavior (Kalivas and Volkow 2011;Knackstedt and Kalivas 2009). Given that mGluR5s aremostly located on postsynaptic cells (Mitrano et al. 2008;Mitrano and Smith 2007) and colocalized with adenosineA2a, dopamine D2, and NMDA receptors (Tebano et al.2005), it is plausible to hypothesize that blockade of post-synaptic mGluR5 may attenuate cocaine- or cue-inducedincreases in synaptic glutamate transmission within theVTA and NAc, thereby inhibiting cocaine- or cue-inducedreinstatement of drug-seeking behavior. More studies arerequired to test this hypothesis and other possible mecha-nisms such as possible interactions between mGluR5 andNMDA or dopamine D2-like receptors.
In addition to inhibition of cocaine self-administration,fenobam sulfate also inhibited sucrose self-administrationand sucrose-induced reinstatement. This is consistent withprevious reports that MPEP or MTEP decrease food-takingbehavior in rats and non-human primates (Paterson and
Markou 2005; Platt et al. 2008) and with a recent report thatfenobam free base inhibits food- and sucrose-induced rein-statement of reward-seeking behavior (Watterson et al.2013). We note, however, that in the present study, fenobamsulfate appears to be more effective in inhibiting cocaine-taking and cocaine-seeking behavior than in inhibitingsucrose-taking and sucrose-seeking behavior (Figs. 2,3 and4). For example, although fenobam sulfate (30–60 mg/kg,p.o.) significantly lowered the rate of sucrose delivery perhour, it failed to alter the total numbers of sucrose deliveriesand active/inactive lever presses during the 90-min testingduration. In addition, fenobam sulfate, at 30 mg/kg, signif-icantly inhibited cocaine-induced, but not sucrose-induced,reinstatement. At 60 mg/kg, fenobam sulfate produced morethan 80 % reduction in cocaine-induced reinstatement, but~50 % reduction in sucrose-induced reinstatement. Thesedata suggest that mGluR5 does regulate general appetitivebehaviors as reported previously (Bradbury et al. 2005).However, within an appropriate therapeutic dose window,mGluR5 NAMs may selectively decrease cocaine-takingand cocaine-seeking behavior without significant effectson other appetitive behaviors.
Initially, research into the pharmacology of mGluR5s wasfocused on the identification and characterization of competi-tive glutamate analogs (Schoepp and Conn 1993), but successwas limited due to a high degree of homology betweenmGluRs at the orthosteric glutamate binding site located in alarge bilobed N-terminal domain. Subsequently, substantialefforts have been taken to develop noncompetitive mGluR5antagonists that target allosteric binding sites (Nicoletti et al.2011) and demonstrate mGluR subtype selectivity. SIB-1757was the first mGluR5-selective antagonist that inhibitedglutamate-induced receptor activation in a noncompetitivemanner (Varney et al. 1999). Subsequent lead optimization ofSIB-1757 led to the discovery of MPEP, a more potent, sys-temically active mGluR5 NAM (Gasparini et al. 1999). Sincethen, MPEP has served as an important research tool.However, off-target effects, notably NMDA receptor antago-nism (Movsesyan et al. 2001; O'Leary et al. 2000), precludedfurther clinical development (Lindsley and Emmitte 2009).MTEP is a second-generation mGluR5 NAM that is bothstructurally and pharmacologically similar to MPEP (Cosfordet al. 2003) but without substantial NMDA receptor activity.However, MTEP was not developed further due to potentinhibition of cytochrome P450 1A2 and a rapid metabolism(Smith et al. 2004). Therefore, several groups have exploredthe MPEP and MTEP pharmacophores in search of high-affinity mGluR5 ligands (Emmitte 2011; Rocher et al. 2011).
Fenobam is a selective mGluR5 NAM that has beenadministered to humans. A recent open-label clinical trialfor the treatment of Fragile X Syndrome reported no adverseeffects of fenobam, but some inconsistent measures of effi-cacy, possibly associated with large inter-individual
Psychopharmacology

variations in reported plasma levels of fenobam followingp.o. administration (Berry-Kravis et al. 2009). In the presentstudy, a new sulfate salt formulation of fenobam gave Cmax
and AUC values approximately two- to three-fold higherthan the free base. Orally active fenobam sulfate attenuatedcocaine self-administration, cocaine-induced reinstatementof drug-seeking behavior, and cue-induced incubation ofcocaine craving in rats and, thus, may be more effectivethan the free base in the clinic.
In summary, the present study reports that fenobam sul-fate is efficacious in several preclinical models of cocaineaddiction. Further, as fenobam has previously been ap-proved for clinical studies and, more recently, has beensuggested for the treatment of Fragile X Syndrome and L-DOPA-induced dyskinesias (Berry-Kravis et al. 2009;Rylander et al. 2010), there appears to be significant poten-tial for this new formulation for translational use as a treat-ment for cocaine abuse. These preclinical data providestrong support for designing human studies using fenobamsulfate as the first clinical candidate in this class to treatcocaine abuse and addiction.
Acknowledgments This research was supported by the IntramuralResearch Program of the National Institute on Drug Abuse, NationalInstitutes of Health, Department of Health and Human Services.T.M.K. was supported by an NIH Postdoctoral Intramural ResearchTraining Associate (IRTA) Fellowship. Pharmacokinetic data was pro-vided under NIDA Contract, N01DA-9-8883. We thank Drs. PhilSkolnick and Amrat Patel, Division of Pharmacotherapies and MedicalConsequences of Drug Abuse, National Institute on Drug Abuse, forproviding the fenobam free base and fenobam sulfate, helpful discus-sions, and critical reading of this manuscript.
Conflict of interest None.
References
Bäckström P, Hyytiä P (2006) Ionotropic and metabotropic glutamatereceptor antagonism attenuates cue-induced cocaine seeking.Neuropsychopharmacology 31:778–786
Backstrom P, Hyytia P (2007) Involvement of AMPA/kainate, NMDA,and mGlu5 receptors in the nucleus accumbens core in cue-induced reinstatement of cocaine seeking in rats. Psychopharma-cology 192:571–580
Bentzley BS, Fender KM, Aston-Jones G (2013) The behavioral eco-nomics of drug self-administration: a review and new analyticalapproach for within-session procedures. Psychopharmacology226:113–125
Berry-Kravis E, Hessl D, Coffey S, Hervey C, Schneider A,Yuhas J, Hutchison J, Snape M, Tranfaglia M, Nguyen DV,Hagerman R (2009) A pilot open label, single dose trial offenobam in adults with fragile X syndrome. J Med Genet46:266–271
Bradbury MJ, Campbell U, Giracello D, Chapman D, King C, TehraniL, Cosford ND, Anderson J, Varney MA, Strack AM (2005)
Metabotropic glutamate receptor mGlu5 is a mediator of appetiteand energy balance in rats and mice. J Pharmacol Exp Ther313:395–402
Carroll FI (2008) Antagonists at metabotropic glutamate receptorsubtype 5: structure activity relationships and therapeutic poten-tial for addiction. Ann N YAcad Sci 1141:221–232
Chiamulera C, Epping-Jordan MP, Zocchi A, Marcon C, Cottiny C,Tacconi S, Corsi M, Orzi F, Conquet F (2001) Reinforcing andlocomotor stimulant effects of cocaine are absent in mGluR5 nullmutant mice. Nat Neurosci 4:873–874
Cleva RM, Watterson LR, Johnson MA, Olive MF (2012) Differ-ential modulation of thresholds for intracranial self-stimulation by mGlu5 positive and negative allosteric modu-lators: implications for effects on drug self-administration.Front Pharmacol 2:93
Cosford ND, Tehrani L, Roppe J, Schweiger E, Smith ND, Anderson J,Bristow L, Brodkin J, Jiang X, McDonald I, Rao S, Washburn M,Varney MA (2003) 3-[(2-Methyl-1,3-thiazol-4-yl)ethynyl]-pyri-dine: a potent and highly selective metabotropic glutamatesubtype 5 receptor antagonist with anxiolytic activity. J MedChem 46:204–206
Dravolina OA, Danysz W, Bespalov AY (2006) Effects of group Imetabotropic glutamate receptor antagonists on the behavioralsensitization to motor effects of cocaine in rats. Psychopharma-cology 187:397–404
Emmitte KA (2011) Recent advances in the design and development ofnovel negative allosteric modulators of mGlu5. ACS ChemNeurosci 2:411–432
Friedmann CTH, Davis LJ, Ciccone PE, Rubin RT (1980) Phase IIdouble blind controlled study of a new anxiolytic, fenobam(McN-3377) vs. placebo. Curr Ther Res 27:144–151
Gasparini F, Lingenhöhl K, Stoehr N, Flor PJ, Heinrich M, Vranesic I,Biollaz M, Allgeier H, Heckendorn R, Urwyler S, Varney MA,Johnson EC, Hess SD, Rao SP, Sacaan AI, Santori EM, VelicelebiG, Kuhn R (1999) 2-Methyl-6-(phenylethynyl)-pyridine (MPEP),a potent, selective and systemically active mGlu5 receptor antag-onist. Neuropharmacology 38:1493–1503
Gawin FH, Kleber HD (1986) Abstinence symptomatology and psy-chiatric diagnosis in cocaine abusers. Clinical observations. ArchGen Psychiatry 43:107–113
Green MD, Jiang X, King CD (2004) Inhibition of human hepatic CYPisoforms by mGluR5 antagonists. Life Sci 75:947–953
Grimm JW, Hope BT, Wise RA, Shaham Y (2001) Neuroadaptation.Incubation of cocaine craving after withdrawal. Nature 412:141–142
Heidbreder CA, Bianchi M, Lacroix LP, Faedo S, Perdona E, RemelliR, Cavanni P, Crespi F (2003) Evidence that the metabotropicglutamate receptor 5 antagonist MPEP may act as an inhibitor ofthe norepinephrine transporter in vitro and in vivo. Synapse50:269–276
Herzig V, Schmidt WJ (2004) Effects of MPEP on locomotion, sensi-tization and conditioned reward induced by cocaine or morphine.Neuropharmacology 47:973–984
Hiranita T, Soto PL, Newman AH, Katz JL (2009) Assessment ofreinforcing effects of benztropine analogs and their effects oncocaine self-administration in rats: comparisons with monoamineuptake inhibitors. J Pharmacol Exp Ther 329:677–686
Itil T, Seaman B, HuqueM,Mukhopadhyay S, Blasucci D, NqK, CicconeP (1978) The clinical and quantitative EEG effects and plasma levelsof fenobam (McN-3377) in subjects with anxiety: an open rising dosetolerance and efficacy study. Curr Ther Res 24:708–724
Jaffe JH, O'Keeffe C (2003) From morphine clinics to buprenorphine:regulating opioid agonist treatment of addiction in the UnitedStates. Drug Alcohol Depend 70:S3–S11
Kalivas PW, Volkow ND (2011) New medications for drug addictionhiding in glutamatergic neuroplasticity. Mol Psychiatry 16:974–986
Psychopharmacology

Kenny PJ, Boutrel B, Gasparini F, Koob GF, Markou A (2005)Metabotropic glutamate 5 receptor blockade may attenuate co-caine self-administration by decreasing brain reward function inrats. Psychopharmacology 179:247–254
Knackstedt LA, Kalivas PW (2009) Glutamate and reinstatement. CurrOpin Pharmacol 9:59–64
Kumaresan V, Yuan M, Yee J, Famous KR, Anderson SM, Schmidt HD,Pierce RC (2009) Metabotropic glutamate receptor 5 (mGluR5)antagonists attenuate cocaine priming- and cue-induced reinstate-ment of cocaine seeking. Behav Brain Res 202:238–244
Lea PM, Faden AI (2006) Metabotropic glutamate receptor subtype 5antagonists MPEP and MTEP. CNS Drug Rev 12:149–166
Lee B, Platt DM, Rowlett JK, Adewale AS, Spealman RD (2005)Attenuation of behavioral effects of cocaine by the metabotropicglutamate receptor 5 antagonist 2-methyl-6-(phenylethynyl)-pyri-dine in squirrel monkeys: comparison with dizocilpine. JPharmacol Exp Ther 312:1232–1240
Lindsley CW, Emmitte KA (2009) Recent progress in the discoveryand development of negative allosteric modulators of mGluR5.Curr Opin Drug Discov Devel 12:446–457
Lu L, Grimm JW, Hope BT, Shaham Y (2004) Incubation of cocainecraving after withdrawal: a review of preclinical data. Neurophar-macology 47(Suppl 1):214–226
Mantsch JR, Li SJ, Risinger R, Awad S, Katz E, Baker DA, Yang Z(2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-induced reinstatement in rats. Psycho-pharmacology 192:581–591
Martin-Fardon R, Baptista MA, Dayas CV, Weiss F (2009) Dissociationof the effects of MTEP [3-[(2-methyl-1,3-thiazol-4-yl)ethynyl]piperidine] on conditioned reinstatement and reinforcement: com-parison between cocaine and a conventional reinforcer. J PharmacolExp Ther 329:1084–1090
Martin-Fardon R,Weiss F (2012) (−)-2-Oxa-4-aminobicylco[3.1.0]hexane-4,6-dicarboxylic acid (LY379268) and 3-[(2-methyl-1,3-thiazol-4-yl)ethynyl]piperidine (MTEP) similarly attenuate stress-induced rein-statement of cocaine seeking. Addict Biol 17:557–564
Mathiesen JM, Svendsen N, Bräuner-Osborne H, Thomsen C, RamirezMT (2003) Positive allosteric modulation of the humanmetabotropic glutamate receptor 4 (hmGluR4) by SIB-1893 andMPEP. Br J Pharmacol 138:1026–1030
McFarland K, Lapish CC, Kalivas PW (2003) Prefrontal glutamaterelease into the core of the nucleus accumbens mediates cocaine-induced reinstatement of drug-seeking behavior. J Neurosci23:3531–3537
McGeehan AJ, Janak PH, Olive MF (2004) Effect of the mGluR5antagonist 6-methyl-2-(phenylethynyl)pyridine (MPEP) on theacute locomotor stimulant properties of cocaine, D-amphetamine,and the dopamine reuptake inhibitor GBR12909 in mice. Psycho-pharmacology 174:266–273
McGeehan AJ, Olive MF (2003) The mGluR5 antagonist MPEP re-duces the conditioned rewarding effects of cocaine but not otherdrugs of abuse. Synapse 47:240–242
Miguéns M, Del Olmo N, Higuera-Matas A, Torres I, García-Lecumberri C, Ambrosio E (2008) Glutamate and aspartate levelsin the nucleus accumbens during cocaine self-administration andextinction: a time course microdialysis study. Psychopharmacol-ogy 196:303–313
Mitrano DA, Arnold C, Smith Y (2008) Subcellular and subsynapticlocalization of group I metabotropic glutamate receptors in thenucleus accumbens of cocaine-treated rats. Neuroscience154:653–666
Mitrano DA, Smith Y (2007) Comparative analysis of the subcellularand subsynaptic localization of mGluR1a and mGluR5metabotropic glutamate receptors in the shell and core of thenucleus accumbens in rat and monkey. J Comp Neurol500:788–806
Montana MC, Cavallone LF, Stubbert KK, Stefanescu AD, KharaschED, Gereau RW IV (2009) The metabotropic glutamate receptorsubtype 5 antagonist fenobam is analgesic and has improved invivo selectivity compared with the prototypical antagonist 2-methyl-6-(phenylethynyl)-pyridine. J Pharmacol Exp Ther330:834–843
Movsesyan VA, O'Leary DM, Fan L, Bao W, Mullins PG, KnoblachSM, Faden AI (2001) mGluR5 antagonists 2-methyl-6-(phenylethynyl)-pyridine and (E)-2-methyl-6-(2-phenylethenyl)-pyridine reduce traumatic neuronal injury in vitro and in vivo byantagonizing N-methyl-D-aspartate receptors. J Pharmacol ExpTher 296:41–47
Nicoletti F, Bockaert J, Collingridge GL, Conn PJ, Ferraguti F,Schoepp DD, Wroblewski JT, Pin JP (2011) Metabotropic gluta-mate receptors: from the workbench to the bedside. Neurophar-macology 60:1017–1041
O'Brien CP, Gardner EL (2005) Critical assessment of how to studyaddiction and its treatment: human and non-human animalmodels. Pharmacol Ther 108:18–58
O'Leary DM, Movsesyan V, Vicini S, Faden AI (2000) SelectivemGluR5 antagonists MPEP and SIB-1893 decrease NMDA orglutamate-mediated neuronal toxicity through actions that reflectNMDA receptor antagonism. Br J Pharmacol 131:1429–1437
Olive MF, McGeehan AJ, Kinder JR, McMahon T, Hodge CW, JanakPH, Messing RO (2005) The mGluR5 antagonist 6-methyl-2-(phenylethynyl)pyridine decreases ethanol consumption via aprotein kinase Cε-dependent mechanism. Mol Pharmacol67:349–355
Panlilio LV, Goldberg SR (2007) Self-administration of drugs in ani-mals and humans as a model and an investigative tool. Addiction102:1863–1870
Paterson NE, Markou A (2005) The metabotropic glutamate receptor 5antagonist MPEP decreased break points for nicotine, cocaine andfood in rats. Psychopharmacology 179:255–261
Pecknold JC, McClure DJ, Appeltauer L, Wrzesinski L, Allan T (1982)Treatment of anxiety using fenobam (a nonbenzodiazepine) in adouble-blind standard (diazepam) placebo-controlled study. J ClinPsychopharmacol 2:129–133
Platt DM, Rowlett JK, Spealman RD (2008) Attenuation of cocaineself-administration in squirrel monkeys following repeated ad-ministration of the mGluR5 antagonist MPEP: comparison withdizocilpine. Psychopharmacology 200:167–176
Porter RH, Jaeschke G, Spooren W, Ballard TM, Buttelmann B,Kolczewski S, Peters JU, Prinssen E, Wichmann J, Vieira E,Muhlemann A, Gatti S, Mutel V, Malherbe P (2005) Fenobam:a clinically validated nonbenzodiazepine anxiolytic is a potent,selective, and noncompetitive mGlu5 receptor antagonist withinverse agonist activity. J Pharmacol Exp Ther 315:711–721
Rocher JP, Bonnét B, Boléa C, Lütjens R, Le Poul E, Poli S, Epping-Jordan M, Bessis AS, Ludwig B, Mutel V (2011) mGluR5 neg-ative allosteric modulators overview: a medicinal chemistry ap-proach towards a series of novel therapeutic agents. Curr Top MedChem 11:680–695
Romano C, van den Pol AN, O'Malley KL (1996) Enhanced earlydevelopmental expression of the metabotropic glutamate receptormGluR5 in rat brain: protein, mRNA splice variants, and regionaldistribution. J Comp Neurol 367:403–412
Rylander D, Iderberg H, Li Q, Dekundy A, Zhang J, Li H, BaishenR, Danysz W, Bezard E, Cenci MA (2010) A mGluR5 antag-onist under clinical development improves L-DOPA-induceddyskinesia in parkinsonian rats and monkeys. Neurobiol Dis39:352–361
Satel SL, Southwick SM, Gawin FH (1991) Clinical features ofcocaine-induced paranoia. Am J Psychiatry 148:495–498
Schoepp DD, Conn PJ (1993) Metabotropic glutamate receptors inbrain function and pathology. Trends Pharmacol Sci 14:13–20
Psychopharmacology

Shigemoto R, Nomura S, Ohishi H, Sugihara H, Nakanishi S, Mizuno N(1993) Immunohistochemical localization of a metabotropic gluta-mate receptor, mGluR5, in the rat brain. Neurosci Lett 163:53–57
Smith ND, Poon SF, Huang D, Green M, King C, Tehrani L, Roppe JR,Chung J, Chapman DP, Cramer M, Cosford ND (2004) Discoveryof highly potent, selective, orally bioavailable, metabotropic glu-tamate subtype 5 (mGlu5) receptor antagonists devoid of cyto-chrome P450 1A2 inhibitory activity. Bioorg Med Chem Lett14:5481–5484
Song R, Zhang HY, Li X, Bi GH, Gardner EL, Xi ZX (2012) Increasedvulnerability to cocaine in mice lacking dopamine D3 receptors.Proc Natl Acad Sci USA 109:17675–17680
Tebano MT, Martire A, Rebola N, Pepponi R, Domenici MR, Gro MC,Schwarzschild MA, Chen JF, Cunha RA, Popoli P (2005) Aden-osine A2A receptors and metabotropic glutamate 5 receptors areco-localized and functionally interact in the hippocampus: a pos-sible key mechanism in the modulation of N-methyl-D-aspartateeffects. J Neurochem 95:1188–1200
Tessari M, Pilla M, Andreoli M, Hutcheson DM, Heidbreder CA(2004) Antagonism at metabotropic glutamate 5 receptors inhibitsnicotine- and cocaine-taking behaviours and prevents nicotine-triggered relapse to nicotine-seeking. Eur J Pharmacol 499:121–133
Varney MA, Cosford ND, Jachec C, Rao SP, Sacaan A, Lin FF,Bleicher L, Santori EM, Flor PJ, Allgeier H, Gasparini F, KuhnR, Hess SD, Velicelebi G, Johnson EC (1999) SIB-1757 and SIB-1893: selective, noncompetitive antagonists of metabotropic glu-tamate receptor type 5. J Pharmacol Exp Ther 290:170–181
Wang B, Shaham Y, Zitzman D, Azari S, Wise RA, You ZB (2005)Cocaine experience establishes control of midbrain glutamate anddopamine by corticotropin-releasing factor: a role in stress-induced relapse to drug seeking. J Neurosci 25:5389–5396
Wang B, You ZB, Rice KC, Wise RA (2007) Stress-induced relapse tococaine seeking: roles for the CRF2 receptor and CRF-bindingprotein in the ventral tegmental area of the rat. Psychopharmacol-ogy 193:283–294
Wang X, Moussawi K, Knackstedt L, Shen H, Kalivas PW (2013) Roleof mGluR5 neurotransmission in reinstated cocaine-seeking. Ad-dict Biol 18:40–49
Watterson LR, Kufahl PR, Nemirovsky NE, Sewalia K, Hood LE, OliveMF (2013) Attenuation of reinstatement of methamphetamine-,sucrose-, and food-seeking behavior in rats by fenobam, ametabotropic glutamate receptor 5 negative allosteric modulator.Psychopharmacology 225:151–159
Xi ZX, Gilbert JG, Peng XQ, Pak AC, Li X, Gardner EL (2006a)Cannabinoid CB1 receptor antagonist AM251 inhibits cocaine-primed relapse in rats: role of glutamate in the nucleusaccumbens. J Neurosci 26:8531–8536
Xi ZX, Newman AH, Gilbert JG, Pak AC, Peng XQ, Ashby CR Jr,Gitajn L, Gardner EL (2006b) The novel dopamine D3 receptorantagonist NGB 2904 inhibits cocaine's rewarding effects andcocaine-induced reinstatement of drug-seeking behavior in rats.Neuropsychopharmacology 31:1393–1405
You ZB, Wang B, Zitzman D, Azari S, Wise RA (2007) A role forconditioned ventral tegmental glutamate release in cocaine seek-ing. J Neurosci 27:10546–10555
Psychopharmacology