changes in oxidative stress parameters and neurodegeneration markers in the brain of the...
TRANSCRIPT

Changes in oxidative stress parameters and neurodegeneration markers
in the brain of the senescence-accelerated mice SAMP-8
Francesc X. Sureda a,*, Javier Gutierrez-Cuesta b, Marta Romeu a, Miquel Mulero a,
Anna Maria Canudas b, Antoni Camins b, Jordi Mallol a, Merce Pallas b
a Unitat de Farmacologia, Facultat de Medicina i Ciencies de la Salut, Universitat Rovira i Virgili, c./St. Llorenc 21, E-43201 Reus, Tarragona, Spainb Unitat de Farmacologia i Farmacognosia, Facultat de Farmacia, Universitat de Barcelona, Nucli Universitari de Pedralbes, E-08028 Barcelona, Spain
Received 28 November 2005; received in revised form 26 January 2006; accepted 31 January 2006
Available online 20 March 2006
Abstract
The senescence-accelerated strains of mice (SAMP) are well-characterized animal models of senescence. Senescence may be related to
enhanced production or defective control of reactive oxygen species, which lead to neuronal damage. Therefore, the activity of various oxidative-
stress related enzymes was determined in the cortex of 5 months-old senescence-accelerated mice prone-8 (SAMP-8) of both sexes and compared
with senescence-accelerated mice-resistant-1 (SAMR-1). Gluthatione reductase and peroxidase activities in SAMP-8 male mice were lower than
in male SAMR-1, and a decreased catalase activity was found in both male and female SAMP-8 mice, which correlates with the lower catalase
expression found byWestern blotting. Nissl staining showed marked loss of neuronal cells in the cerebral cortex of five months-old SAMP-8 mice.
SAMP-8 mice also had marked astrogliosis and microgliosis. We also found an increase in caspase-3 and calpain activity in the cortex. In addition,
we observed morphological changes in the immunostaining of tau protein in SAMP-8, indicative of a loss of their structural function. Altogether,
these results show that, at as early as 5 months of age, SAMP-8 mice have cytological and molecular alterations indicative of neurodegeneration in
the cerebral cortex and suggestive of altered control of the production of oxidative species and hyper-activation of calcium-dependent enzymes.
q 2006 Elsevier Inc. All rights reserved.
Keywords: Senescence; Oxidative stress; Catalase; Calpain; Tau; Neurodegeneration
1. Introduction
The study of the ageing process is an important area of
research. There is growing interest in establishing the
molecular targets that contribute to the neurobiology of ageing
and, ultimately, in developing drugs that could modulate the
activity of such targets and mitigate the multi-organic
dysfunction that is the common feature of ageing. To better
understand the ageing process, several animal models have
been developed. Among them, the senescence-accelerated
mice (SAM) model is widely used in experimental gerontol-
ogy. The SAM strain was generated in the mid-1970s by
Takeda and colleagues at the University of Kyoto, and is
characterized by a reduced life span and the presence of early
signs of senescence (lordokyphosis, loss of activity, neuro-
logical signs, among others). The senescence-accelerated
0531-5565/$ - see front matter q 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.exger.2006.01.015
* Corresponding author. Tel.: C34 977 75 93 57; fax: C34 977 75 93 78.
E-mail address: [email protected] (F.X. Sureda).
mice-prone (SAMP) model and its equivalent with normal
ageing, the senescence-accelerated mice-resistant (SAMR)
model, are both established animal models for studying ageing
and age-associated pathologies.
A number of studies using different strains of SAM mice
indicate that alterations in the control of oxidative stress are
responsible for this accelerated ageing (for a review, see
Hosokawa, 2002). For instance, parameters indicative of lipid
or protein oxidation are greater in SAMP-8 than in SAMR-1
(Kim et al., 2002), and it has been suggested that changes in the
activity of several key enzymes account for this enhanced
oxidative stress (Liu and Mori, 1993). Based on these
observations, it has been postulated that antioxidant com-
pounds would have beneficial effects as anti-ageing com-
pounds, and several studies have been carried out to ascertain
this. In fact, melatonin reduces oxidative damage in the brain of
SAMP-8 mice (Okatani et al., 2002). Alpha-lipoic acid or
N-acetylcisteine have also shown to reverse memory impair-
ment in aged SAMP-8 (Farr et al., 2003), and Boldyrev and
coworkers found an enhanced life span in SAMP-1 treated with
the antioxidant carnosine (Stvolinskii et al., 2003).
Experimental Gerontology 41 (2006) 360–367
www.elsevier.com/locate/expgero

F.X. Sureda et al. / Experimental Gerontology 41 (2006) 360–367 361
However, the search for a particular key step in the
oxidative stress defence machinery has been inconclusive.
Although several authors point to an alteration of a particular
enzyme that might be responsible for increased oxidative
stress, others found transient changes or no change at all.
Apart from this, no studies have been carried out to ascertain
gender differences in enzyme activity or expression in these
strains. Whatever the cause of increased oxidative stress, a
number of publications have described changes indicative of
neuronal cell death in the brain of these strains of mice.
Extensive literature describes the involvement of caspases
(cysteine proteases) in neuronal cell death. In addition, several
studies performed in post-mortem human brain tissues suggest
the participation of these enzymes in neurodegenerative
processes and in ageing (Maccioni et al., 2001). Another
class of cysteine proteases involved in neuronal cell death is
the calcium-activated protease, calpain, with up to 12
isoenzyme forms (Nixon, 2003). The two most widely
distributed isoforms, calpain I (m-calpain) and calpain II (m-
calpain), have nearly identical substrate specificities and, so
far, have been shown to differ mainly in their calcium
requirements for activation in vitro and their tissue distri-
butions. By making selective and limited proteolytic
cleavages, they modulate the activity of enzymes, including
key signalling molecules, and induce specific cytoskeletal
rearrangements, which accounts for their roles in cell motility,
signal transduction, vesicular trafficking and structural
stabilization. Enhanced calpain activation is involved in
several ageing phenomena and diseases of late life, including
cataract formation, erythrocyte senescence, type-2 Diabetes
mellitus, hypertension, arthritis, and neurodegenerative dis-
orders. Increased calpain activation could be part of the
neurodegenerative pathway process in several neuro-
degenerative disorders such as Parkinson’s, Alzheimer’s and
Huntington’s diseases (Nixon et al., 1994). Calpains directly
or indirectly activate cdk5 and ERK/MAP kinase (Nath et al.,
2000; Patrick et al., 1999), which phosphorylate tau and
potentially promote tau-related cytoskeletal disorganization,
as well as reducing neuronal survival by other mechanisms
(Delacourte et al., 2002; Zhu et al., 2004). The overactivation
of calpains, in particular, has figured prominently in
hypotheses on cellular ageing beginning with the observation
that levels of calpain activity in the brain correlate inversely
with lifespan in across several orders of mammals (Nixon,
2003). Consistent with observations of increased intracellular
or calcium influx in ageing tissues, calpain activities also rise
during ageing (Benuck et al., 1996) The basis for high calpain
activity in ageing is, most commonly, increased activation
without any change in overall expression (Averna et al., 2003;
Nixon, 2003).
The aim of this study was to establish differences in several
key enzymes in the regulation of oxidative stress between
SAMP-8 and SAMR-1 in 5 months-old mice. Moreover, we
have established new features that are indicative of neuronal
damage and that shed light on the underlying causes of
accelerated ageing in this animal model.
2. Materials and methods
2.1. Animals
Senescence-accelerated mice prone-8 (SAMP-8) and senes-
cence-accelerated mice resistant-1 (SAMR-1) were provided
by Harlan Interfauna Iberica (Barcelona, Spain). All mice were
housed in a room equipped with automatic light cycles (12 h
light/dark), and maintained at 22G2 8C and relative humidity
of 40–60%. Food (Panlab rat chow, Panlab, Barcelona) and tap
water were offered ad libitum throughout the study.
2.2. Sample processing
For enzyme activity assays, 5 months-old mice were
anesthetized with a mixture of xylazine and ketamine (i.p. 10
and 100 mg/kg, respectively) dissolved in 0.2 M phosphate
buffered saline. Immediately after death, the brain was
removed and cerebral cortices were dissected on an ice–cold
surface. After homogenization in 0.2 M cold sodium
phosphate buffer (1:4) at pH 6.25 in a Potter–Elvehjem
homogenizer fitted with a teflon pestle, the crude soluble
fraction was obtained (centrifugation at 1,05,000!g for
60 min) and the pellet was discarded. The fresh crude soluble
fraction was divided to determine the parameters described
below. An aliquot was reserved for the glutathione-S-
transferase (GST) activity assay, which was conducted the
same day as the sample processing. The remainder was
distributed in different aliquots and maintained at K80 8C
until the day of assay.
For Western blot, caspase-3 and calpain activity determi-
nations the cortices were quickly excised and weighted.
Afterwards, they were homogenized using a Polytron
(Kinematica) in 10 volumes of cold buffer (0.32 sucrose and
5 mM tris–HCl, pH 7.4). Homogenates were centrifuged twice
at 15,000!g for 30 min. Finally, the pellet was dissolved in
50 mM Tris–HCl buffer, pH 7.4, with a Potter–Elvejhem
homogenizer.
Protein was determined with Bradford method, with using
bovine serum albumin used as standard (Merck, Germany).
For immunohistochemistry studies, mice were anesthetized
with sodium pentobarbital (75 mg/kg, i.p.) and exsanguinated
with 25 ml cold 0.1 M phosphate-buffered saline, pH 7.4 (PBS)
followed by fixation with 25 ml 4% paraformaldehyde in 0.1 M
(PBS), pH 7.4. Brains were removed and postfixed overnight in
4% paraformaldehyde and then cryoprotected in 20% sucrose
in 0.1 M PBS. The fixed brains were coronally cut into 30-mmthick sections on a cryostate and stored in 4% paraformalde-
hyde plus sodium azide (0.2%) at 4 8C.
2.3. Enzyme activity assays
Glutathione-S-transferase (GST) activity was determined
immediately after centrifugation, using the method of Habig et
al. (1975) and with CDNB (1-chloro-2, 4-dinitrobenzene) as a
substrate. As CDNB is a non-specific substrate, total GST
activity was determined.
F.X. Sureda et al. / Experimental Gerontology 41 (2006) 360–367362
Glutathione reductase (GR) and glutathione peroxidase
(GPx) activities were determined by the Wheeler et al. (1990)
method in a COBAS–MIRA autoanalyzer (Roche Diagnostics)
and expressed in units/g of mg protein (one unit, UZ1 mmol of
NADPH transformed/min).
Catalase (CAT) activity was determined by the Cohen et al.
(1970) method, by monitoring the rate of disappearance of
hydrogen peroxide (15 mM) in a Lambda 2 PerkinElmer
spectrophotometer. Values are expressed in terms of units/mg
of protein (one unit, UZ1 mmol of hydrogen peroxide
transformed/min).
Caspase-3 activity was determined by using the colorimetric
substrate Ac-DEVD-p-nitroaniline, a synthetic substrate of
caspase-3. Briefly, the protein aliquots were incubated with
200 mM of Ac-DEVD-p-nitroaniline in assay buffer in 96-well
plates at 37 8C for 24 h. Absorbance of the cleaved product was
measured at 405 nm in a microplate reader (BioRad). Results
are expressed as arbitrary units of absorbance.
Calpain activity was measured by using a commercial kit
(MBL International). In brief, fluorimetric calpain substrate
Ac-LLY-AFC was used as follows: cortex samples were
collected in a lysis buffer and 0.5 mg/ml of protein was
incubated with 200 mM of fluorometric substrate in assay
buffer in 96-well plates at 37 8C for 24 h. Negative control was
through using the calpain inhibitor Z-LLY-FMK. The
fluorescence intensity of the liberated AFC was monitored by
using a microplate spectrofluorometer (PerkinElmer Victor 3)
at an excitation wavelength of 400 nm and an emission
wavelength of 505 nm. Calpain activity is expressed as
Relative Fluorescence Units (RFU) per milligram of protein.
2.4. Western-blot analysis
Aliquots of tissue homogenate, containing 30 mg (catalase
and cdk5) or 5 mg (spectrin) of protein per sample, were placed
in sample buffer (0.5 M Tris–HCl pH 6.8, 10% glycerol, 2%
(W/V) SDS, 5% (V/V) 2-b-mercaptoethanol, 0.05% bromo-
phenol blue) and denaturised by boiling at 95–100 8C for
5 min. Samples were separated by electrophoresis on 10%
acrylamide gels. Subsequently, proteins were transferred to
polyvinylidene fluoride (PVDF) sheets (Immobilone-P,
Millipore Corp.) by means of a transblot apparatus (BioRad).
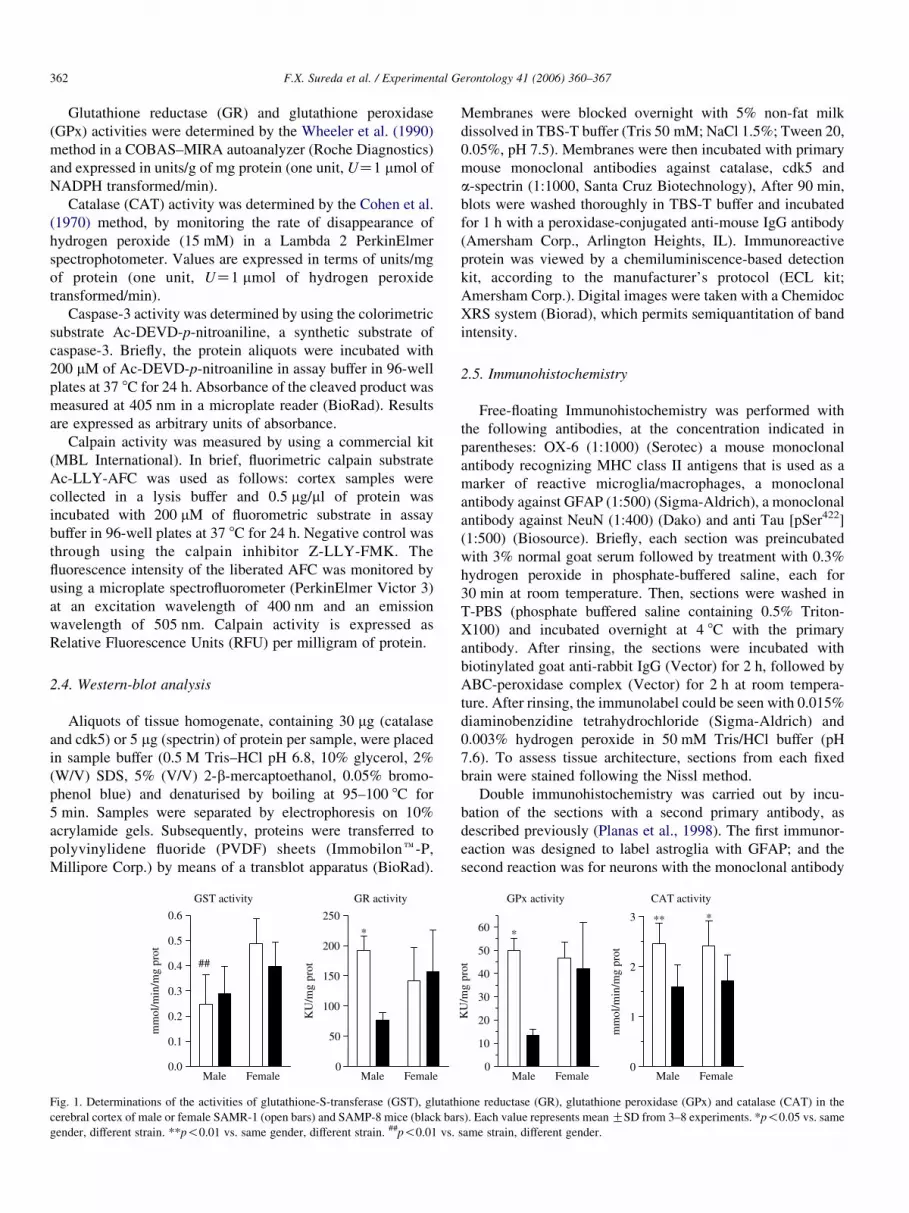
Fig. 1. Determinations of the activities of glutathione-S-transferase (GST), glutath
cerebral cortex of male or female SAMR-1 (open bars) and SAMP-8 mice (black bar
gender, different strain. **p!0.01 vs. same gender, different strain. ##p!0.01 vs.
Membranes were blocked overnight with 5% non-fat milk
dissolved in TBS-T buffer (Tris 50 mM; NaCl 1.5%; Tween 20,
0.05%, pH 7.5). Membranes were then incubated with primary
mouse monoclonal antibodies against catalase, cdk5 and
a-spectrin (1:1000, Santa Cruz Biotechnology), After 90 min,
blots were washed thoroughly in TBS-T buffer and incubated
for 1 h with a peroxidase-conjugated anti-mouse IgG antibody
(Amersham Corp., Arlington Heights, IL). Immunoreactive
protein was viewed by a chemiluminiscence-based detection
kit, according to the manufacturer’s protocol (ECL kit;
Amersham Corp.). Digital images were taken with a Chemidoc
XRS system (Biorad), which permits semiquantitation of band
intensity.
2.5. Immunohistochemistry
Free-floating Immunohistochemistry was performed with
the following antibodies, at the concentration indicated in
parentheses: OX-6 (1:1000) (Serotec) a mouse monoclonal
antibody recognizing MHC class II antigens that is used as a
marker of reactive microglia/macrophages, a monoclonal
antibody against GFAP (1:500) (Sigma-Aldrich), a monoclonal
antibody against NeuN (1:400) (Dako) and anti Tau [pSer422]
(1:500) (Biosource). Briefly, each section was preincubated
with 3% normal goat serum followed by treatment with 0.3%
hydrogen peroxide in phosphate-buffered saline, each for
30 min at room temperature. Then, sections were washed in
T-PBS (phosphate buffered saline containing 0.5% Triton-
X100) and incubated overnight at 4 8C with the primary
antibody. After rinsing, the sections were incubated with
biotinylated goat anti-rabbit IgG (Vector) for 2 h, followed by
ABC-peroxidase complex (Vector) for 2 h at room tempera-
ture. After rinsing, the immunolabel could be seen with 0.015%
diaminobenzidine tetrahydrochloride (Sigma-Aldrich) and
0.003% hydrogen peroxide in 50 mM Tris/HCl buffer (pH
7.6). To assess tissue architecture, sections from each fixed
brain were stained following the Nissl method.
Double immunohistochemistry was carried out by incu-
bation of the sections with a second primary antibody, as
described previously (Planas et al., 1998). The first immunor-
eaction was designed to label astroglia with GFAP; and the
second reaction was for neurons with the monoclonal antibody
ione reductase (GR), glutathione peroxidase (GPx) and catalase (CAT) in the
s). Each value represents meanGSD from 3–8 experiments. *p!0.05 vs. same
same strain, different gender.

F.X. Sureda et al. / Experimental Gerontology 41 (2006) 360–367 363
against NeuN. After extensive washing following the first
immunoreaction, sections were incubated with 3% normal
horse serum for 2 h to block nonspecific binding sites for NeuN
immunohistochemistry. Sections were incubated overnight at
4 8C with the corresponding primary antibody, followed by
treatment with biotinylated secondary antibody (1:200), and
the ABC complex. Finally, sections were washed in 0.01 M
sodium phosphate buffer, pH 6, preincubated in 0.01%
benzidine dihydrochloride and 0.025% sodium nitroferricya-
nide in 0.01 M sodium phosphate buffer for 10 min and then
developed in the same solution containing 0.005% of H2O2.
3. Results
3.1. Changes in activity and expression of oxidative-stress
related enzymes
GST activity does not differ between SAMP-8 mice and
SAMR-1 mice. However, we found a statistically significant
Fig. 2. Representative images of 3–4 experiments of Nissl staining, NeuN, GFAP, an
Arrows indicate representative positive cells in each experiment.
difference in GST activity between males and females in
SAMR-1 mice. Specifically, GST activity in male SAMR-1
mice was approximately half than that of females. However, no
other differences were found for this enzyme. Male SAMP-8
mice had lower GR (60% less) and GPx activity (74% less)
than SAMR-1, but this difference was not found in females.
CAT activity was the only enzyme that showed a consistently
reduced activity in both SAMP-8 male (35% decrease) and
female (29%) mice than in SAMR-1 (Fig.1).
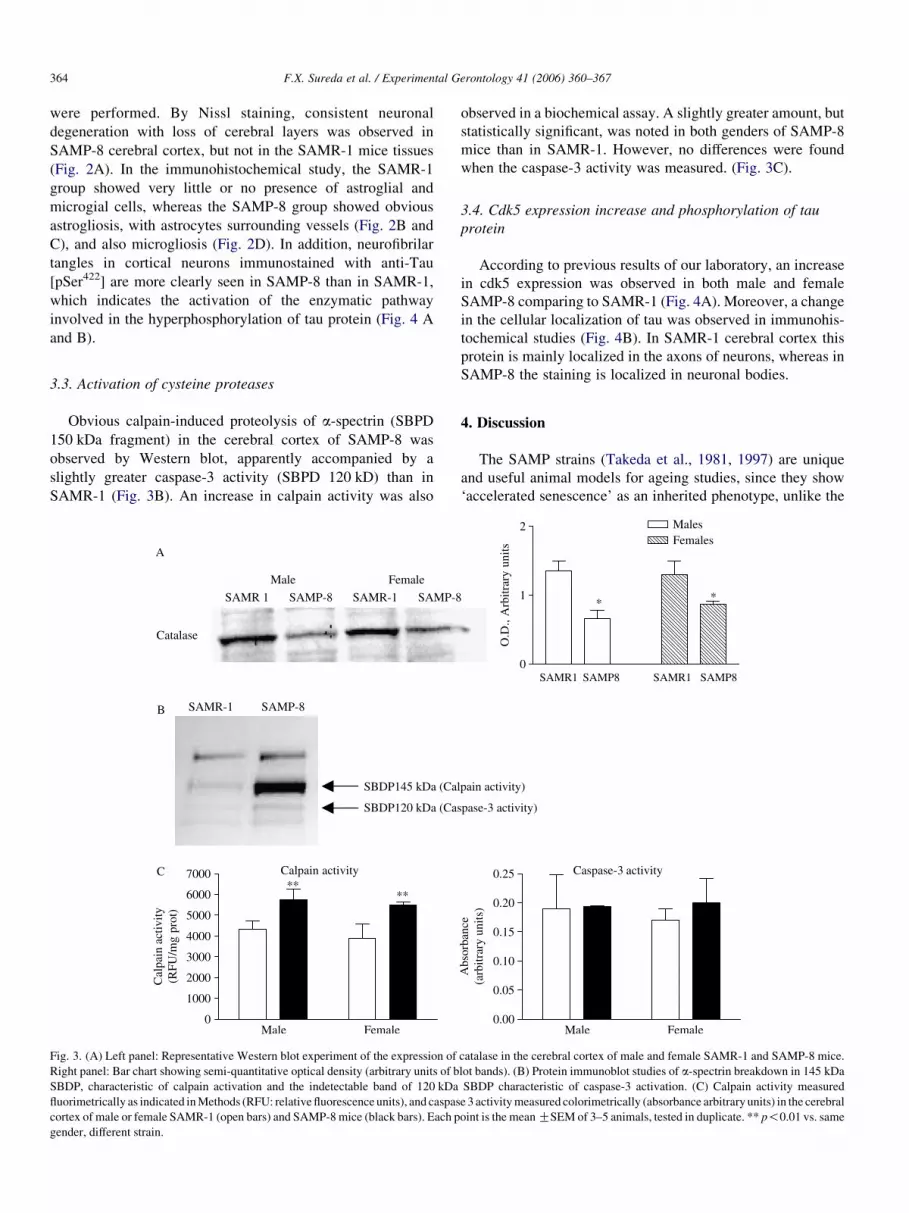
To ascertain whether so little activity was due to less enzyme
expression, aWestern blot analysis was performed. As shown in
Fig. 3A a significantly reduced CAT expression was found in
both male and female SAMP-8 mice than in SAMR-1 mice.
3.2. Immunohistochemistry and histological study with Nissl
staining
To assess the presence of neurodegenerative features, Nissl
staining and immunostaining with neuronal and glial markers
d OX6 immunohistochemistry in cerebral cortex of male SAMR-1 and SAMP-8.
F.X. Sureda et al. / Experimental Gerontology 41 (2006) 360–367364
were performed. By Nissl staining, consistent neuronal
degeneration with loss of cerebral layers was observed in
SAMP-8 cerebral cortex, but not in the SAMR-1 mice tissues
(Fig. 2A). In the immunohistochemical study, the SAMR-1
group showed very little or no presence of astroglial and
microgial cells, whereas the SAMP-8 group showed obvious
astrogliosis, with astrocytes surrounding vessels (Fig. 2B and
C), and also microgliosis (Fig. 2D). In addition, neurofibrilar
tangles in cortical neurons immunostained with anti-Tau
[pSer422] are more clearly seen in SAMP-8 than in SAMR-1,
which indicates the activation of the enzymatic pathway
involved in the hyperphosphorylation of tau protein (Fig. 4 A
and B).
3.3. Activation of cysteine proteases
Obvious calpain-induced proteolysis of a-spectrin (SBPD
150 kDa fragment) in the cerebral cortex of SAMP-8 was
observed by Western blot, apparently accompanied by a
slightly greater caspase-3 activity (SBPD 120 kD) than in
SAMR-1 (Fig. 3B). An increase in calpain activity was also
Fig. 3. (A) Left panel: Representative Western blot experiment of the expression of
Right panel: Bar chart showing semi-quantitative optical density (arbitrary units of b
SBDP, characteristic of calpain activation and the indetectable band of 120 kDa
fluorimetrically as indicated inMethods (RFU: relative fluorescence units), and caspas
cortex of male or female SAMR-1 (open bars) and SAMP-8mice (black bars). Each p
gender, different strain.
observed in a biochemical assay. A slightly greater amount, but
statistically significant, was noted in both genders of SAMP-8
mice than in SAMR-1. However, no differences were found
when the caspase-3 activity was measured. (Fig. 3C).
3.4. Cdk5 expression increase and phosphorylation of tau
protein
According to previous results of our laboratory, an increase
in cdk5 expression was observed in both male and female
SAMP-8 comparing to SAMR-1 (Fig. 4A). Moreover, a change
in the cellular localization of tau was observed in immunohis-
tochemical studies (Fig. 4B). In SAMR-1 cerebral cortex this
protein is mainly localized in the axons of neurons, whereas in
SAMP-8 the staining is localized in neuronal bodies.
4. Discussion
The SAMP strains (Takeda et al., 1981, 1997) are unique
and useful animal models for ageing studies, since they show
‘accelerated senescence’ as an inherited phenotype, unlike the
catalase in the cerebral cortex of male and female SAMR-1 and SAMP-8 mice.
lot bands). (B) Protein immunoblot studies of a-spectrin breakdown in 145 kDa
SBDP characteristic of caspase-3 activation. (C) Calpain activity measured
e 3 activitymeasured colorimetrically (absorbance arbitrary units) in the cerebral
oint is the meanGSEM of 3–5 animals, tested in duplicate. ** p!0.01 vs. same
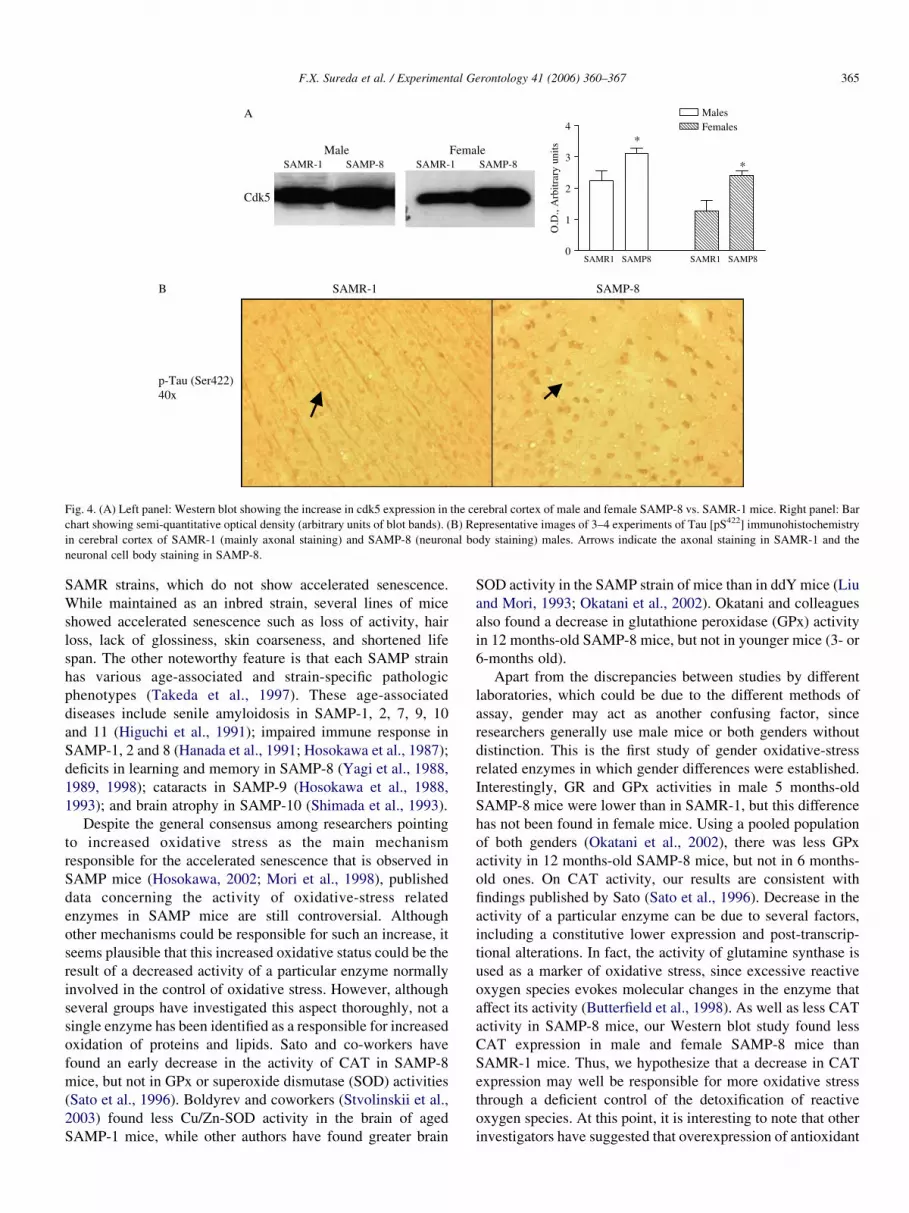
Fig. 4. (A) Left panel: Western blot showing the increase in cdk5 expression in the cerebral cortex of male and female SAMP-8 vs. SAMR-1 mice. Right panel: Bar
chart showing semi-quantitative optical density (arbitrary units of blot bands). (B) Representative images of 3–4 experiments of Tau [pS422] immunohistochemistry
in cerebral cortex of SAMR-1 (mainly axonal staining) and SAMP-8 (neuronal body staining) males. Arrows indicate the axonal staining in SAMR-1 and the
neuronal cell body staining in SAMP-8.
F.X. Sureda et al. / Experimental Gerontology 41 (2006) 360–367 365
SAMR strains, which do not show accelerated senescence.
While maintained as an inbred strain, several lines of mice
showed accelerated senescence such as loss of activity, hair
loss, lack of glossiness, skin coarseness, and shortened life
span. The other noteworthy feature is that each SAMP strain
has various age-associated and strain-specific pathologic
phenotypes (Takeda et al., 1997). These age-associated
diseases include senile amyloidosis in SAMP-1, 2, 7, 9, 10
and 11 (Higuchi et al., 1991); impaired immune response in
SAMP-1, 2 and 8 (Hanada et al., 1991; Hosokawa et al., 1987);
deficits in learning and memory in SAMP-8 (Yagi et al., 1988,
1989, 1998); cataracts in SAMP-9 (Hosokawa et al., 1988,
1993); and brain atrophy in SAMP-10 (Shimada et al., 1993).
Despite the general consensus among researchers pointing
to increased oxidative stress as the main mechanism
responsible for the accelerated senescence that is observed in
SAMP mice (Hosokawa, 2002; Mori et al., 1998), published
data concerning the activity of oxidative-stress related
enzymes in SAMP mice are still controversial. Although
other mechanisms could be responsible for such an increase, it
seems plausible that this increased oxidative status could be the
result of a decreased activity of a particular enzyme normally
involved in the control of oxidative stress. However, although
several groups have investigated this aspect thoroughly, not a
single enzyme has been identified as a responsible for increased
oxidation of proteins and lipids. Sato and co-workers have
found an early decrease in the activity of CAT in SAMP-8
mice, but not in GPx or superoxide dismutase (SOD) activities
(Sato et al., 1996). Boldyrev and coworkers (Stvolinskii et al.,
2003) found less Cu/Zn-SOD activity in the brain of aged
SAMP-1 mice, while other authors have found greater brain
SOD activity in the SAMP strain of mice than in ddY mice (Liu
and Mori, 1993; Okatani et al., 2002). Okatani and colleagues
also found a decrease in glutathione peroxidase (GPx) activity
in 12 months-old SAMP-8 mice, but not in younger mice (3- or
6-months old).
Apart from the discrepancies between studies by different
laboratories, which could be due to the different methods of
assay, gender may act as another confusing factor, since
researchers generally use male mice or both genders without
distinction. This is the first study of gender oxidative-stress
related enzymes in which gender differences were established.
Interestingly, GR and GPx activities in male 5 months-old
SAMP-8 mice were lower than in SAMR-1, but this difference
has not been found in female mice. Using a pooled population
of both genders (Okatani et al., 2002), there was less GPx
activity in 12 months-old SAMP-8 mice, but not in 6 months-
old ones. On CAT activity, our results are consistent with
findings published by Sato (Sato et al., 1996). Decrease in the
activity of a particular enzyme can be due to several factors,
including a constitutive lower expression and post-transcrip-
tional alterations. In fact, the activity of glutamine synthase is
used as a marker of oxidative stress, since excessive reactive
oxygen species evokes molecular changes in the enzyme that
affect its activity (Butterfield et al., 1998). As well as less CAT
activity in SAMP-8 mice, our Western blot study found less
CAT expression in male and female SAMP-8 mice than
SAMR-1 mice. Thus, we hypothesize that a decrease in CAT
expression may well be responsible for more oxidative stress
through a deficient control of the detoxification of reactive
oxygen species. At this point, it is interesting to note that other
investigators have suggested that overexpression of antioxidant

F.X. Sureda et al. / Experimental Gerontology 41 (2006) 360–367366
enzymes, such as Cu/Zn superoxide dismutase (Cu/Zn SOD)
might be responsible for the delayed ageing that is observed in
the dwarf mice (Hauck and Bartke, 2000). These researchers
also found greater hypothalamic CAT activity, but failed to
demonstrate greater CAT expression. However, other mech-
anisms cannot be ruled out: Poon et al. have suggested that the
b-amyloid peptide may increase oxidative stress in the brain of
SAMP-8 mice, since the intracerebroventricular administration
of antisense oligonucleotides of the APP gene reduces lipid
peroxidation and protein oxidation (Poon et al., 2004).
We found signs of gliosis and neurodegeneration in the brain
of SAMP-8, which was consistent with the oxidative stress
features. Immunohistochemistry techniques revealed important
neuroglial changes at as early as 5 months of age in the SAMP-8
strain of mice. There was a significant increase in reactive
astrocytes and microgliosis is demonstrated, according to other
previous reports on the SAM strain (Katoh-Semba and Kato,
1994). The loss of cellular bodies and cerebral cortex structures
are evident in the SAMP-8 at 5 months andmay be due in part to
oxidative stress and glial reactivity but other mechanisms
cannot be ruled out. Although recent reports indicate that
neuronal death in the SAMP-8 hippocampus is not mediated by
the intrinsic apoptotic pathway (Wu et al., 2005), the
participation of other proteases has not been studied till now.
Here we demonstrate the participation of other proteases,
calpains, in the neuronal death involved in senescence in
SAMP-8. Once activated, calpains and caspases degrade the
cytosketal protein a-spectrin (280 kD) into three main
fragments of 145 kDa (calpains), 120 kDa (caspase-3) and
150 kDa (calpains-caspases). Both the presence of a-spectrinbreakdown products (SBDP 150 kDa) and the increased activity
of this enzyme demonstrate a solid relationship between
neuronal death and ageing in this model of senescence.
Calpain is activated by increased intracellular calcium
concentrations (Nixon et al., 1994). Our results indicate that in
SAMP-8 there are certain biochemical changes, probably
through a deficient control of ROS production causing a rise in
the intracellular calcium concentration, thus increasing the
calpain activation in the cortex. Several mechanisms for
explaining the intracellular calcium concentration that would
lead to calpain activation have been put forward. For example,
it has been reported that increased cytosolic ROS causes the
calcium concentration in the mitochondrial matrix to increase,
which could interfere with the respiratory chain and in the
tricarboxylic acid cycle. This would promote the opening of the
mitochondrial permeability transition pore, thus releasing
proapoptotic factors into the cytosol (Wu et al., 2005).
Moreover, the neuronal calcium buffering capacity might
also be impaired due to an age-dependent decrease in calbindin
(Armbrecht et al., 1999).
Recent findings from our laboratory demonstrate that there
is a hyperphosphorylation of the microtubule-associated
protein tau in SAMP-8 at 5 months-old (Canudas et al.,
2005). Calpains are activated in neuronal processes before
abnormalities in the protein tau are detected. In a latter phase,
the activated protease associates prominently with abnormal
neuronal tau aggregates (neuropil threads and neurofibrillary
tangles), a hallmark of Alzheimer’s disease neuropathology
(Avila et al., 2004). It has been suggested that calpain breaks
cdk5/p35 down into cdk5/p25, which is followed by an
increase in cdk5/Gsk3ß activity, thus inducing post-transla-
tional modifications in tau (Baumann et al., 1993; Canudas
et al., 2005; Cho and Johnson, 2003) and neuronal cell death in
the cerebral cortex. The activation of cdk5/p25 activity was
demonstrated in 5 months-old SAMP-8 accompanied by tau
modifications (Canudas et al., 2005). Here we report an
alteration in an upstream step (increased calpain activity), that
would lead to cdk5 activation and, consequently, to tau protein
alterations. As it is illustrated by the immunohistochemical
studies against anti-Tau [pSer422], the typical staining of as
drop depositions can be seen, thus indicating a loss of the
original structure. Moreover, other age-related alterations in
several tissues have been attributed to increased calpain
activity, including cataract formation or characteristic
reductions in the major erythrocyte transmembrane and ion
transport protein in red cells (Hosokawa et al., 1984, 1988;
Nixon et al., 1994; Takeda et al., 1981; Takeda et al., 1991).
Ageing-induced cell death may be caused through altera-
tions of mitochondrial function, and in fact, our results
corroborate the hypothesis by which a defective expression
of CAT or other oxidative stress-related enzymes (namely
superoxide dismutase) in the cerebral cortex, may lead to
mitochondrial alterations an greater activity of proteases that
would enhance cdk5/Gsk3b thus induces an increase in tau
phosphorylation, neurofibrilar tangles formation and gliosis,
cell loss in cerebral cortex and the functional decline during
ageing in SAMP-8.
Acknowledgements
This study was made possible by a grant from the Spanish
‘Instituto de Salud Carlos III (ISCIII)’, through the G03/137
project, by grants SAF2005-05179-C02-01 from Ministerio de
Educacion y Ciencia, European Funds. We are grateful to
Vanessa Sanchez–Martos and Ana Isabel Dıez for their
excellent technical assistance. We thank Almirall–Prodesfarma
for Pharmacology Award 2005.
References
Armbrecht, H.J., Boltz, M.A., Kumar, V.B., Flood, J.F., Morley, J.E., 1999.
Effect of age on calcium-dependent proteins in hippocampus of senescence-
accelerated mice. Brain Res. 842, 287–293.
Averna, M., De Tullio, R., Capini, P., Salamino, F., Pontremoli, S., Melloni, E.,
2003. Changes in calpastatin localization and expression during calpain
activation: a new mechanism for the regulation of intracellular Ca(2C)-
dependent proteolysis. Cell. Mol. Life Sci. 60, 2669–2678.
Avila, J., Lucas, J.J., Perez, M., Hernandez, F., 2004. Role of tau protein in both
physiological and pathological conditions. Physiol. Rev. 84, 361–384.
Baumann,K.,Mandelkow,E.M.,Biernat, J., Piwnica-Worms,H.,Mandelkow,E.,
1993. Abnormal Alzheimer-like phosphorylation of tau-protein by cyclin-
dependent kinases cdk2 and cdk5. FEBS Lett. 336, 417–424.
Benuck, M., Banay-Schwartz, M., DeGuzman, T., Lajtha, A., 1996. Changes in
brain protease activity in aging. J. Neurochem. 67, 2019–2029.
Butterfield, D.A., Koppal, T., Howard, B., Subramaniam, R., Hall, N.,
Hensley, K., Yatin, S., Allen, K., Aksenov, M., Aksenova, M.,

F.X. Sureda et al. / Experimental Gerontology 41 (2006) 360–367 367
Carney, J., 1998. Structural and functional changes in proteins induced by
free radical-mediated oxidative stress and protective action of the
antioxidants N-tert-butyl-alpha-phenylnitrone and vitamin E. Ann. NY
Acad. Sci. 854, 448–462.
Canudas, A.M., Gutierrez-Cuesta, J., Rodriguez, M.I., Acuna-Castroviejo, D.,
Sureda, F.X., Camins, A., Pallas, M., 2005. Hyperphosphorylation of
microtubule-associated protein tau in senescence-accelerated mouse
(SAM). Mech. Ageing Dev. 126, 1300–1304.
Cho, J.H., Johnson, G.V., 2003. Glycogen synthase kinase 3beta phosphor-
ylates tau at both primed and unprimed sites. differential impact on
microtubule binding. J. Biol. Chem. 278, 187–193.
Cohen, G., Dembiec, D., Marcus, J., 1970. Measurement of catalase activity in
tissue extracts. Anal. Biochem. 34, 30–38.
Delacourte, A., Sergeant, N., Wattez, A., Maurage, C.A., Lebert, F., Pasquier, F.,
David, J.P., 2002. Tau aggregation in the hippocampal formation: an ageing or
a pathological process? Exp. Gerontol. 37, 1291–1296.
Farr, S.A., Poon, H.F., Dogrukol-Ak, D., Drake, J., Banks, W.A., Eyerman, E.,
Butterfield, D.A., Morley, J.E., 2003. The antioxidants alpha-lipoic acid
and N-acetylcysteine reverse memory impairment and brain oxidative
stress in aged SAMP8 mice. J. Neurochem. 84, 1173–1183.
Habig, W.H., Keen, J.H., Jakoby, W.B., 1975. Glutathione S-transferase in the
formation of cyanide from organic thiocyanates and as an organic nitrate
reductase. Biochem. Biophys. Res. Commun. 64, 501–506.
Hanada, K., Katoh, H., Hosokawa, T., Hosono, M., Takeda, T., 1991. Immune
responses in newly developed short-lived SAM mice. IV. chromosomal
location of a gene controlling defective helper T-cell activity. Immunology
74, 160–164.
Hauck, S.J.,Bartke,A., 2000.Effects of growthhormoneonhypothalamiccatalase
and Cu/Zn superoxide dismutase. Free Radic. Biol. Med. 28, 970–978.
Higuchi, K., Naiki, H., Kitagawa, K., Hosokawa, M., Takeda, T., 1991. Mouse
senile amyloidosis. ASSAM amyloidosis in mice presents universally as a
systemic age-associated amyloidosis. Virchows Arch. B. Cell Pathol. Incl.
Mol. Pathol. 60, 231–238.
Hosokawa, M., 2002. A higher oxidative status accelerates senescence and
aggravates age-dependent disorders in SAMP strains of mice. Mech.
Ageing Dev. 123, 1553–1561.
Hosokawa, M., Takeshita, S., Higuchi, K., Shimizu, K., Irino, M., Toda, K.,
Honma, A., Matsumura, A., Yasuhira, K., Takeda, T., 1984. Cataract and
other ophthalmic lesions in senescence accelerated mouse (SAM).
morphology and incidence of senescence associated ophthalmic changes
in mice. Exp. Eye Res. 38, 105–114.
Hosokawa, T., Hosono, M., Higuchi, K., Aoike, A., Kawai, K., Takeda, T.,
1987. Immune responses in newly developed short-lived SAMmice. I. age-
associated early decline in immune activities of cultured spleen cells.
Immunology 62, 419–423.
Hosokawa, M., Ashida, Y., Tsuboyama, T., Chen, W.H., Takeda, T., 1988.
Cataract in senescence accelerated mouse (SAM). 2. Development of a new
strain of mouse with late-appearing cataract. Exp. Eye Res. 47, 629–640.
Hosokawa, M., Ashida, Y., Matsushita, T., Takahashi, K., Takeda, T., 1993.
Persistent hyaloid vascular system in age-related cataract in a SAM strain of
mouse. Exp. Eye Res. 57, 427–434.
Katoh-Semba, R., Kato, K., 1994. Age-related changes in levels of the beta-
subunit of nerve growth factor in selected regions of the brain: comparison
between senescence-accelerated (SAM-P8) and senescence-resistant
(SAM-R1) mice. Neurosci. Res. 20, 251–256.
Kim, H.C., Bing, G., Jhoo, W.K., Kim, W.K., Shin, E.J., Park, E.S., Choi, Y.S.,
Lee, D.W., Shin, C.Y., Ryu, J.R., Ko, K.H., 2002. Oxidative damage causes
formation of lipofuscin-like substances in the hippocampus of the
senescence-accelerated mouse after kainate treatment. Behav. Brain Res.
131, 211–220.
Liu, J., Mori, A., 1993. Age-associated changes in superoxide dismutase
activity, thiobarbituric acid reactivity and reduced glutathione level in the
brain and liver in senescence accelerated mice (SAM): a comparison with
ddY mice. Mech. Ageing Dev. 71, 23–30.
Maccioni, R.B., Munoz, J.P., Barbeito, L., 2001. The molecular bases of
alzheimer’s disease and other neurodegenerative disorders. Arch. Med.
Res. 32, 367–381.
Mori, A., Utsumi, K., Liu, J., Hosokawa, M., 1998. Oxidative damage in the
senescence-accelerated mouse. Ann. NY Acad. Sci. 854, 239–250.
Nath, R., Davis, M., Probert, A.W., Kupina, N.C., Ren, X., Schielke, G.P.,
Wang, K.K., 2000. Processing of cdk5 activator p35 to its truncated form
(p25) by calpain in acutely injured neuronal cells. Biochem. Biophys. Res.
Commun. 274, 16–21.
Nixon, R.A., 2003. The calpains in aging and aging-related diseases. Ageing
Res. Rev. 2, 407–418.
Nixon, R.A., Saito, K.I., Grynspan, F., Griffin, W.R., Katayama, S., Honda, T.,
Mohan, P.S., Shea, T.B., Beermann, M., 1994. Calcium-activated neutral
proteinase (calpain) system in aging and Alzheimer’s disease. Ann. NY
Acad. Sci. 747, 77–91.
Okatani, Y., Wakatsuki, A., Reiter, R.J., Miyahara, Y., 2002. Melatonin
reduces oxidative damage of neural lipids and proteins in senescence-
accelerated mouse. Neurobiol. Aging 23, 639–644.
Patrick, G.N., Zukerberg, L., Nikolic, M., de la Monte, S., Dikkes, P.,
Tsai, L.H., 1999. Conversion of p35 to p25 deregulates Cdk5 activity and
promotes neurodegeneration. Nature 402, 615–622.
Planas, A.M., Justicia, C., Soriano, M.A., Ferrer, I., 1998. Epidermal growth
factor receptor in proliferating reactive glia following transient focal
ischemia in the rat brain. Glia 23, 120–129.
Poon, H.F., Joshi, G., Sultana, R., Farr, S.A., Banks, W.A., Morley, J.E.,
Calabrese, V., Butterfield, D.A., 2004. Antisense directed at the Abeta
region of APP decreases brain oxidative markers in aged senescence
accelerated mice. Brain Res. 1018, 86–96.
Sato, E., Kurokawa, T., Oda, N., Ishibashi, S., 1996. Early appearance of
abnormality of microperoxisomal enzymes in the cerebral cortex of
senescence-accelerated mouse. Mech. Ageing Dev. 92, 175–184.
Shimada, A., Ohta, A., Akiguchi, I., Takeda, T., 1993. Age-related
deterioration in conditional avoidance task in the SAM-P/10 mouse, an
animal model of spontaneous brain atrophy. Brain Res. 608, 266–272.
Stvolinskii, S.L., Fedorova, T.N., Yuneva, M.O., Boldyrev, A.A., 2003.
Protective effect of carnosine on Cu,Zn-superoxide dismutase during
impaired oxidative metabolism in the brain in vivo. Bull. Exp. Biol. Med.
135, 130–132.
Takeda, T., Hosokawa, M., Takeshita, S., Irino, M., Higuchi, K.,
Matsushita, T., Tomita, Y., Yasuhira, K., Hamamoto, H., Shimizu, K.,
Ishii, M., Yamamuro, T., 1981. A new murine model of accelerated
senescence. Mech. Ageing Dev. 17, 183–194.
Takeda, T., Hosokawa, M., Higuchi, K., 1991. Senescence-accelerated mouse
(SAM): a novel murine model of accelerated senescence. J. Am. Geriatr.
Soc. 39, 911–919.
Takeda, T., Hosokawa, M., Higuchi, K., 1997. Senescence-accelerated mouse
(SAM): a novel murine model of senescence. Exp. Gerontol. 32, 105–109.
Wheeler, C.R., Salzman, J.A., Elsayed,N.M., Omaye, S.T., Korte Jr., D.W., 1990.
Automated assays for superoxide dismutase, catalase, glutathione peroxidase,
and glutathione reductase activity. Anal. Biochem. 184, 193–199.
Wu, Y., Zhang, A.Q., Wai, M.S.M., Lai, H.W.L., Wu, S.X,. Yew, D.T.,
in press. Changes of apoptosis-related proteins in hippocampus of SAM
mouse in development and aging. Neurobiol. of Aging. Corrected proof.
doi:10.1016/j.neurobiolaging.2005.07.014.
Yagi, H., Katoh, S., Akiguchi, I., Takeda, T., 1988. Age-related deterioration of
ability of acquisition in memory and learning in senescence accelerated
mouse: SAM-P/8 as an animal model of disturbances in recent memory.
Brain Res. 474, 86–93.
Yagi, H., Irino, M., Matsushita, T., Katoh, S., Umezawa, M., Tsuboyama, T.,
Hosokawa, M., Akiguchi, I., Tokunaga, R., Takeda, T., 1989. Spontaneous
spongy degeneration of the brain stem in SAM-P/8 mice, a newly developed
memory-deficient strain. J. Neuropathol. Exp. Neurol. 48, 577–590.
Yagi, H., Akiguchi, I., Ohta, A., Yagi, N., Hosokawa, M., Takeda, T., 1998.
Spontaneous and artificial lesions of magnocellular reticular formation of
brainstem deteriorate avoidance learning in senescence-accelerated mouse
SAM. Brain Res. 791, 90–98.
Zhu, L.Q., Wang, S.H., Ling, Z.Q., Wang, D.L., Wang, J.Z., 2004. Effect of
inhibiting melatonin biosynthesis on spatial memory retention and tau
phosphorylation in rat. J. Pineal Res. 37, 71–77.