a dynamically regulated 14–3–3, slob, and slowpoke potassium channel complex in drosophila...
TRANSCRIPT

Neuron, Vol. 22, 809–818, April, 1999, Copyright 1999 by Cell Press
A Dynamically Regulated 14–3–3, Slob,and Slowpoke Potassium Channel Complexin Drosophila Presynaptic Nerve Terminals
Xiao et al., 1998), have dispelled any remaining notionthat channels might float freely in the membrane lipid.
Large conductance calcium-dependent potassium(KCa) channels are ubiquitous in neurons and other excit-able cells (Latorre et al., 1989) and play a critical role in
Yi Zhou,* W. Michael Schopperle,* Heather Murrey,*Angela Jaramillo,* Daniel Dagan,*§ Leslie C. Griffith,†and Irwin B. Levitan*‡
*Department of Biochemistry†Department of Biology
regulating neuronal firing patterns and neurotransmitterVolen Center for Complex Systemsrelease (e.g., Gho and Ganetzky, 1992; Robitaille et al.,Brandeis University1993). Because their activity is regulated by both theWaltham, Massachusetts 02454membrane voltage and intracellular calcium, they canbe thought of as a molecular locus for integrating electri-cal and biochemical signals. Hence, they are particularlySummaryattractive targets for modulation, and they are indeedknown to be modulated by a number of different proteinSlob is a novel protein that binds to the carboxy-termi-kinases (Chung et al., 1991; Reinhart et al., 1991; Whitenal domain of the Drosophila Slowpoke (dSlo) calcium-et al., 1991, 1993; Bielefeldt and Jackson, 1994; Reinhartdependent potassium (KCa) channel. A yeast two-and Levitan, 1995; Shipston and Armstrong, 1996; Narahybrid screen with Slob as bait identifies the z isoformet al., 1998; Tian et al., 1998). The first KCa channel toof 14–3–3 as a Slob-binding protein. Coimmunopre-be cloned was one encoded by the Slowpoke locus incipitation experiments from Drosophila heads andDrosophila (Atkinson et al., 1991; Adelman et al., 1992).transfected cells confirm that 14–3–3 interacts withThe amino acid sequence of this Drosophila SlowpokedSlo via Slob. All three proteins are colocalized presyn-(dSlo) channel reveals that it is considerably longer thanaptically at Drosophila neuromuscular junctions. Twoother voltage-dependent potassium channels, with anserine residues in Slob are required for 14–3–3 binding,extended carboxy-terminal domain that constitutes aboutand the binding is dynamically regulated in Drosophilatwo-thirds of the channel protein. The function of thisby calcium/calmodulin-dependent kinase II (CaMKII)large domain is not clear, although evidence from mam-phosphorylation. 14–3–3 coexpression dramatically al-malian Slowpoke homologs suggests that it may partici-ters dSlo channel properties when wild-type Slob ispate in calcium binding (Wei et al., 1994; Schreiber andpresent but not when a double serine mutant Slob thatSalkoff, 1997). Interestingly, the Drosophila and mam-is incapable of binding 14–3–3 is present. The resultsmalian Slowpokes have a large number of alternativeprovide evidence for a dSlo/Slob/14–3–3 regulatorysplice variants within this region of the protein (Adelmanprotein complex.et al., 1992; Tseng-Crank et al., 1994). Part or all of thecarboxy-terminal domain of dSlo have been used asIntroductionbaits in yeast two-hybrid screens, and two novel pro-teins that interact with and modulate dSlo have been
Modulation of membrane ion channels is of fundamentalidentified (Schopperle et al., 1998; Xia et al., 1998b). We
importance for the regulation of neuronal excitability, have named one of these proteins Slob, for Slowpoke-synaptic transmission, and neuronal plasticity. It is well- binding protein.established that both ligand-gated and voltage-depen- To investigate further the function of Slob, we havedent ion channels are important targets for protein used it as bait in another yeast two-hybrid screen. Wekinases, phosphatases, and other signaling proteins show here that this screen identifies the z isoform of(Swope et al., 1992; Catterall, 1993; Levitan, 1994). More 14–3–3 as a Slob-binding protein and that dSlo, Slob,recently, it has become evident that some ion channels and 14–3–3 interact in Drosophila tissues as well as inbind directly to signaling proteins that modulate channel transfected cells. The 14–3–3 proteins were first identi-properties (Swope and Huganir, 1994; Ehlers et al., 1996; fied some 30 years ago as a group of abundant, acidicFuhrer and Hall, 1996; Holmes et al., 1996; Colledge and brain proteins (reviewed by Aitken et al., 1992), but onlyFroehner, 1997; Yu et al., 1997; Schopperle et al., 1998; recently has their function begun to be investigated. ItXia et al., 1998a, 1998b). These findings, together with is becoming apparent that they may participate in cellthe emerging recognition that scaffolding proteins of cycle control, apoptosis, and in multiple signal transduc-various types can cause ion channels to localize and tion pathways by acting as scaffolds for and regulatorscluster at appropriate locations in the plasma membrane of signaling proteins such as protein kinases (Morrison,(Hall and Sanes, 1993; Wallace, 1995; Kim et al., 1995, 1994; Burbelo and Hall, 1995; Pawson and Scott, 1997).1998; Kornau et al., 1995; Muller et al., 1996; Apel et al., In Drosophila, 14–3–3 isoforms have been reported to1997; Brakeman et al., 1997; Dong et al., 1997; Ehlers regulate signaling mediated by the small GTPase Raset al., 1998; Lin et al., 1998; Niethammer et al., 1998; (Chang and Rubin, 1997) and are required for Raf kinase–
dependent photoreceptor development (Kockel et al.,1997). In addition, flies that contain a mutation in the‡ To whom correspondence should be addressed (e-mail: levitan@gene for the z isoform of 14–3–3 exhibit olfactory learningvolen.brandeis.edu).deficits (Skoulakis and Davis, 1996), as well as defects§ Permanent address: Bernard Katz Minerva Center for Cell Biophys-in synaptic transmission at the neuromuscular junctionics and Rappaport Faculty of Medicine, Technion, POB 9697, Haifa
31096, Israel. (Broadie et al., 1997). Our demonstration that 14–3–3

Neuron810
Figure 1. dSlo, Slob, and 14–3–3 Coimmunoprecipitate from Co-transfected Cells
tsA201 cells were transfected with 14–3–3 and Slob (lane 1), 14–3–3and dSlo (lane 2), or 14–3–3, Slob and dSlo (lane 3). 14–3–3 immuno-precipitates were probed on Western blots for Slob (A), dSlo (B), or14–3–3 (C).(A) Slob is detected in the 14–3–3 immunoprecipitate when it iscotransfected with 14–3–3, either in the absence (lane 1) or presence Figure 2. dSlo, Slob, and 14–3–3 Coimmunoprecipitate from Dro-(lane 3) of dSlo. sophila Heads(B) dSlo is detected in the 14–3–3 immunoprecipitate when it is 14–3–3 immunoprecipitates (left panels) from fly head or transfectedcotransfected with 14–3–3, but only in the presence of Slob (lane 3). cell lysates (right panels) were probed on Western blots for Slob(C) The amount of 14–3–3 expressed and immunoprecipitated is the (A), dSlo (B), or 14–3–3 (C).same under all transfection conditions. (A) A protein of molecular weight similar to that of recombinant Slob
(left panel, right lane) coimmunoprecipitates from fly heads with14–3–3 and is recognized by the Slob antibody (left panel, left lane).Bands of the same molecular weight are seen in the lysates (rightcan interact with and modulate dSlo via Slob raises thepanel). Other unidentified bands are also recognized by the Slobintriguing possibility that some of the actions of 14–3–3antibody.may result from its interaction with potassium channels.(B) A protein of molecular weight similar to that of recombinant dSlocoimmunoprecipitates from wild-type (WT) fly heads with 14–3–3
Results and is recognized by the dSlo antibody (left panel, left lane). Thisprotein is not present when the immunoprecipitate is prepared fromslo4 homozygous mutant flies that lack the dSlo channel (left panel,Two-Hybrid Screen with Slob as Baitright lane). A higher molecular weight species is recognized in theWe reported recently the identification and characteriza-lysates from both wild-type and mutant flies (right panel).tion of Slob, a novel protein that binds to dSlo (Schop-(C) Expression and immunoprecipitation of 14–3–3 are similar inperle et al., 1998). To determine whether Slob might wild-type and slo4 flies.
interact with other proteins and bring them into a com-plex with the channel, cDNA encoding the completeamino acid sequence of Slob was used as bait in a yeast
Slob are expressed together in the tsA201 mammaliantwo-hybrid screen (Fields and Song, 1989). Approxi-cell line (Margolskee et al., 1993) and 14–3–3 is immuno-mately 2,000,000 transformed yeast colonies wereprecipitated, Slob can be detected in the immunoprecip-screened, and 91 positive colonies were identified onitate (Figure 1A, lane 1). In contrast, when 14–3–3 isthe second day. Of the 15 colonies that were confirmedcoexpressed with dSlo, little dSlo coimmunoprecipi-as true positives by more stringent genetic testing, 2tates with 14–3–3 (Figure 1B, lane 2), indicating thatcontained cDNA inserts encoding the z isoform of Dro-these two proteins do not interact directly. However,sophila 14–3–3 (Swanson and Ganguly, 1992).when 14–3–3, dSlo, and Slob are all transfected to-gether, both Slob (Figure 1A, lane 3) and dSlo (FigureA Protein Complex Containing dSlo, Slob,1B, lane 3) are present in the 14–3–3 immunoprecipitate.and 14–3–3Note that the amount of 14–3–3 in the immunoprecipitateTo test further the suggestion from the yeast two-hybridis very similar under all the transfection conditions usedscreen that 14–3–3 binds to Slob, coimmunoprecipita-
tion experiments were carried out. When 14–3–3 and (Figure 1C). These results demonstrate clearly that 14–
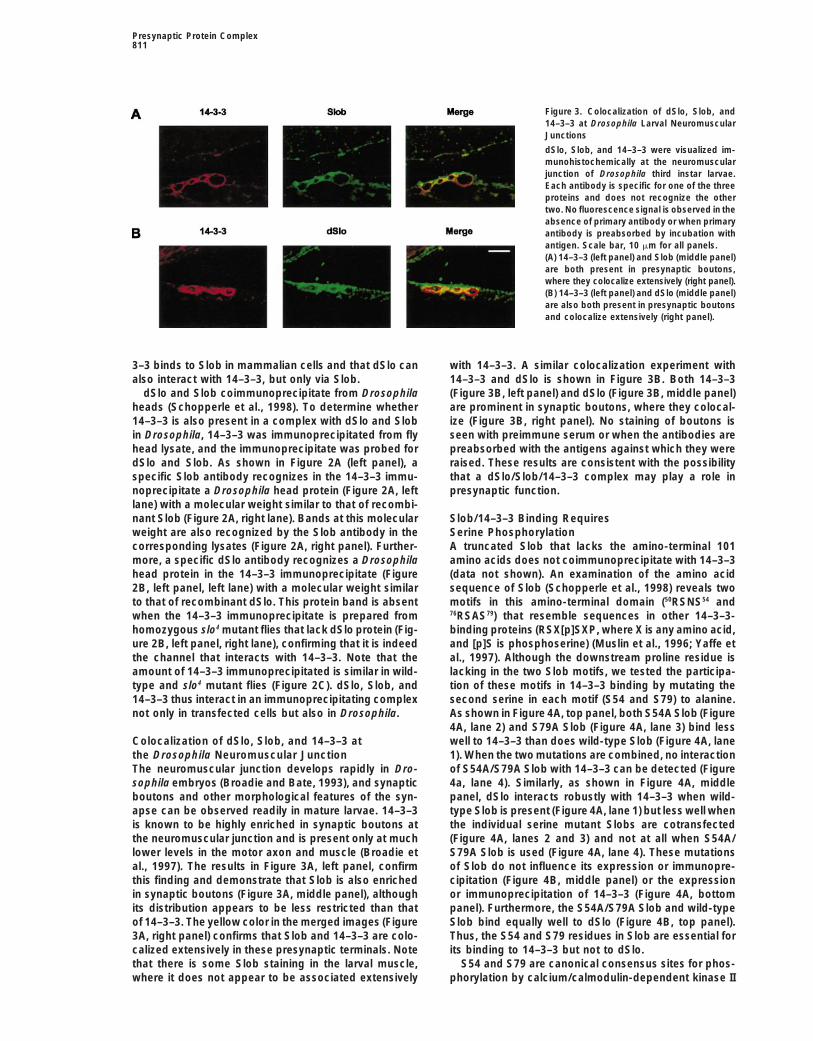
Presynaptic Protein Complex811
Figure 3. Colocalization of dSlo, Slob, and14–3–3 at Drosophila Larval NeuromuscularJunctions
dSlo, Slob, and 14–3–3 were visualized im-munohistochemically at the neuromuscularjunction of Drosophila third instar larvae.Each antibody is specific for one of the threeproteins and does not recognize the othertwo. No fluorescence signal is observed in theabsence of primary antibody or when primaryantibody is preabsorbed by incubation withantigen. Scale bar, 10 mm for all panels.(A) 14–3–3 (left panel) and Slob (middle panel)are both present in presynaptic boutons,where they colocalize extensively (right panel).(B) 14–3–3 (left panel) and dSlo (middle panel)are also both present in presynaptic boutonsand colocalize extensively (right panel).
3–3 binds to Slob in mammalian cells and that dSlo can with 14–3–3. A similar colocalization experiment with14–3–3 and dSlo is shown in Figure 3B. Both 14–3–3also interact with 14–3–3, but only via Slob.
dSlo and Slob coimmunoprecipitate from Drosophila (Figure 3B, left panel) and dSlo (Figure 3B, middle panel)are prominent in synaptic boutons, where they colocal-heads (Schopperle et al., 1998). To determine whether
14–3–3 is also present in a complex with dSlo and Slob ize (Figure 3B, right panel). No staining of boutons isseen with preimmune serum or when the antibodies arein Drosophila, 14–3–3 was immunoprecipitated from fly
head lysate, and the immunoprecipitate was probed for preabsorbed with the antigens against which they wereraised. These results are consistent with the possibilitydSlo and Slob. As shown in Figure 2A (left panel), a
specific Slob antibody recognizes in the 14–3–3 immu- that a dSlo/Slob/14–3–3 complex may play a role inpresynaptic function.noprecipitate a Drosophila head protein (Figure 2A, left
lane) with a molecular weight similar to that of recombi-nant Slob (Figure 2A, right lane). Bands at this molecular Slob/14–3–3 Binding Requires
Serine Phosphorylationweight are also recognized by the Slob antibody in thecorresponding lysates (Figure 2A, right panel). Further- A truncated Slob that lacks the amino-terminal 101
amino acids does not coimmunoprecipitate with 14–3–3more, a specific dSlo antibody recognizes a Drosophilahead protein in the 14–3–3 immunoprecipitate (Figure (data not shown). An examination of the amino acid
sequence of Slob (Schopperle et al., 1998) reveals two2B, left panel, left lane) with a molecular weight similarto that of recombinant dSlo. This protein band is absent motifs in this amino-terminal domain (50RSNS54 and
76RSAS79) that resemble sequences in other 14–3–3-when the 14–3–3 immunoprecipitate is prepared fromhomozygous slo4 mutant flies that lack dSlo protein (Fig- binding proteins (RSX[p]SXP, where X is any amino acid,
and [p]S is phosphoserine) (Muslin et al., 1996; Yaffe eture 2B, left panel, right lane), confirming that it is indeedthe channel that interacts with 14–3–3. Note that the al., 1997). Although the downstream proline residue is
lacking in the two Slob motifs, we tested the participa-amount of 14–3–3 immunoprecipitated is similar in wild-type and slo4 mutant flies (Figure 2C). dSlo, Slob, and tion of these motifs in 14–3–3 binding by mutating the
second serine in each motif (S54 and S79) to alanine.14–3–3 thus interact in an immunoprecipitating complexnot only in transfected cells but also in Drosophila. As shown in Figure 4A, top panel, both S54A Slob (Figure
4A, lane 2) and S79A Slob (Figure 4A, lane 3) bind lesswell to 14–3–3 than does wild-type Slob (Figure 4A, laneColocalization of dSlo, Slob, and 14–3–3 at
the Drosophila Neuromuscular Junction 1). When the two mutations are combined, no interactionof S54A/S79A Slob with 14–3–3 can be detected (FigureThe neuromuscular junction develops rapidly in Dro-
sophila embryos (Broadie and Bate, 1993), and synaptic 4a, lane 4). Similarly, as shown in Figure 4A, middlepanel, dSlo interacts robustly with 14–3–3 when wild-boutons and other morphological features of the syn-
apse can be observed readily in mature larvae. 14–3–3 type Slob is present (Figure 4A, lane 1) but less well whenthe individual serine mutant Slobs are cotransfectedis known to be highly enriched in synaptic boutons at
the neuromuscular junction and is present only at much (Figure 4A, lanes 2 and 3) and not at all when S54A/S79A Slob is used (Figure 4A, lane 4). These mutationslower levels in the motor axon and muscle (Broadie et
al., 1997). The results in Figure 3A, left panel, confirm of Slob do not influence its expression or immunopre-cipitation (Figure 4B, middle panel) or the expressionthis finding and demonstrate that Slob is also enriched
in synaptic boutons (Figure 3A, middle panel), although or immunoprecipitation of 14–3–3 (Figure 4A, bottompanel). Furthermore, the S54A/S79A Slob and wild-typeits distribution appears to be less restricted than that
of 14–3–3. The yellow color in the merged images (Figure Slob bind equally well to dSlo (Figure 4B, top panel).Thus, the S54 and S79 residues in Slob are essential for3A, right panel) confirms that Slob and 14–3–3 are colo-
calized extensively in these presynaptic terminals. Note its binding to 14–3–3 but not to dSlo.S54 and S79 are canonical consensus sites for phos-that there is some Slob staining in the larval muscle,
where it does not appear to be associated extensively phorylation by calcium/calmodulin-dependent kinase II
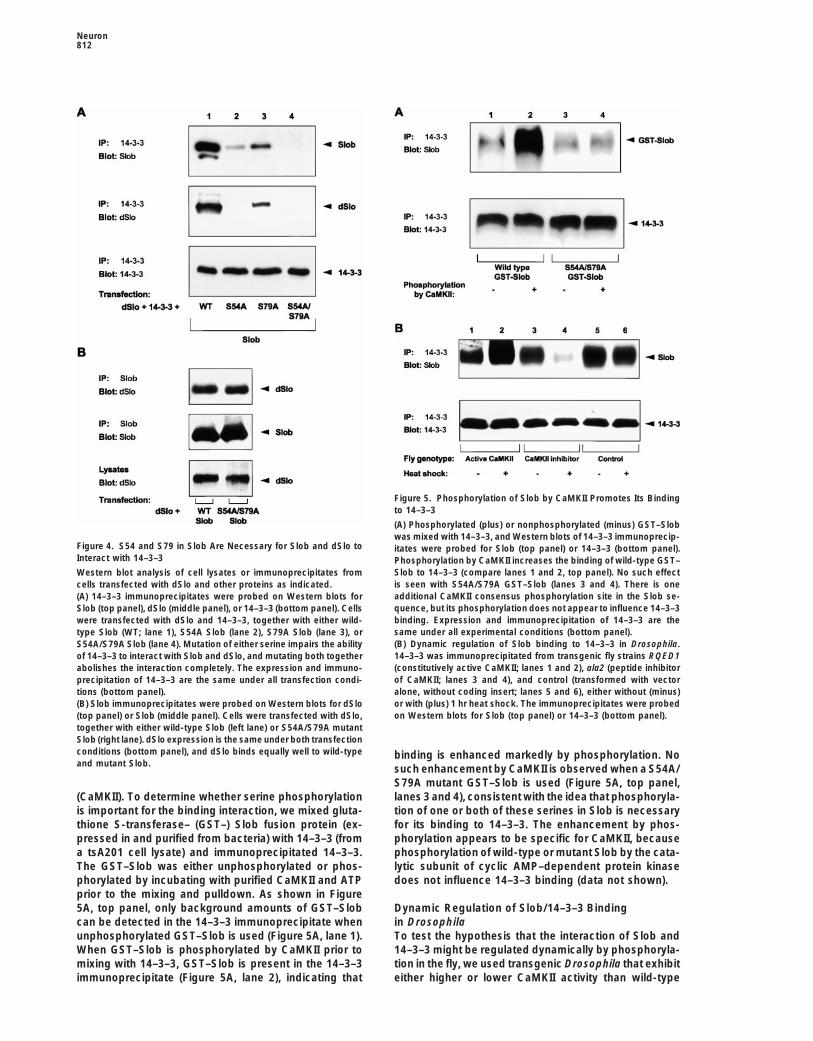
Neuron812
Figure 5. Phosphorylation of Slob by CaMKII Promotes Its Bindingto 14–3–3
(A) Phosphorylated (plus) or nonphosphorylated (minus) GST–Slobwas mixed with 14–3–3, and Western blots of 14–3–3 immunoprecip-
Figure 4. S54 and S79 in Slob Are Necessary for Slob and dSlo to itates were probed for Slob (top panel) or 14–3–3 (bottom panel).Interact with 14–3–3 Phosphorylation by CaMKII increases the binding of wild-type GST–
Slob to 14–3–3 (compare lanes 1 and 2, top panel). No such effectWestern blot analysis of cell lysates or immunoprecipitates fromis seen with S54A/S79A GST–Slob (lanes 3 and 4). There is onecells transfected with dSlo and other proteins as indicated.additional CaMKII consensus phosphorylation site in the Slob se-(A) 14–3–3 immunoprecipitates were probed on Western blots forquence, but its phosphorylation does not appear to influence 14–3–3Slob (top panel), dSlo (middle panel), or 14–3–3 (bottom panel). Cellsbinding. Expression and immunoprecipitation of 14–3–3 are thewere transfected with dSlo and 14–3–3, together with either wild-same under all experimental conditions (bottom panel).type Slob (WT; lane 1), S54A Slob (lane 2), S79A Slob (lane 3), or(B) Dynamic regulation of Slob binding to 14–3–3 in Drosophila.S54A/S79A Slob (lane 4). Mutation of either serine impairs the ability14–3–3 was immunoprecipitated from transgenic fly strains RQED1of 14–3–3 to interact with Slob and dSlo, and mutating both together(constitutively active CaMKII; lanes 1 and 2), ala2 (peptide inhibitorabolishes the interaction completely. The expression and immuno-of CaMKII; lanes 3 and 4), and control (transformed with vectorprecipitation of 14–3–3 are the same under all transfection condi-alone, without coding insert; lanes 5 and 6), either without (minus)tions (bottom panel).or with (plus) 1 hr heat shock. The immunoprecipitates were probed(B) Slob immunoprecipitates were probed on Western blots for dSloon Western blots for Slob (top panel) or 14–3–3 (bottom panel).(top panel) or Slob (middle panel). Cells were transfected with dSlo,
together with either wild-type Slob (left lane) or S54A/S79A mutantSlob (right lane). dSlo expression is the same under both transfectionconditions (bottom panel), and dSlo binds equally well to wild-type binding is enhanced markedly by phosphorylation. Noand mutant Slob. such enhancement by CaMKII is observed when a S54A/
S79A mutant GST–Slob is used (Figure 5A, top panel,lanes 3 and 4), consistent with the idea that phosphoryla-(CaMKII). To determine whether serine phosphorylation
is important for the binding interaction, we mixed gluta- tion of one or both of these serines in Slob is necessaryfor its binding to 14–3–3. The enhancement by phos-thione S-transferase– (GST–) Slob fusion protein (ex-
pressed in and purified from bacteria) with 14–3–3 (from phorylation appears to be specific for CaMKII, becausephosphorylation of wild-type or mutant Slob by the cata-a tsA201 cell lysate) and immunoprecipitated 14–3–3.
The GST–Slob was either unphosphorylated or phos- lytic subunit of cyclic AMP–dependent protein kinasedoes not influence 14–3–3 binding (data not shown).phorylated by incubating with purified CaMKII and ATP
prior to the mixing and pulldown. As shown in Figure5A, top panel, only background amounts of GST–Slob Dynamic Regulation of Slob/14–3–3 Binding
in Drosophilacan be detected in the 14–3–3 immunoprecipitate whenunphosphorylated GST–Slob is used (Figure 5A, lane 1). To test the hypothesis that the interaction of Slob and
14–3–3 might be regulated dynamically by phosphoryla-When GST–Slob is phosphorylated by CaMKII prior tomixing with 14–3–3, GST–Slob is present in the 14–3–3 tion in the fly, we used transgenic Drosophila that exhibit
either higher or lower CaMKII activity than wild-typeimmunoprecipitate (Figure 5A, lane 2), indicating that

Presynaptic Protein Complex813
Figure 6. Modulation of dSlo by 14–3–3, Acting via Slob
dSlo channel activity was evoked by 150 ms depolarizing voltage steps to 130 mV or 1130 mV in detached membrane patches, at a freeCa21 concentration of 30 mM. All traces were normalized to the Itail (max) (see Experimental Procedures), and hence, no current scale bar isshown.(A) Normalized currents evoked by depolarizations to 130 mV. Current amplitude is similar in patches from cells transfected with dSlo alone(i) or dSlo 1 14–3–3 (ii) and is higher in patches from cells transfected with dSlo 1 Slob (iii). Much less current is observed under theseconditions in patches from cells transfected with dSlo 1 14–3–3 1 wild-type Slob (iv), but not when S54A/S79A mutant Slob is used (v).(B) Left, traces iv and v from (A), aligned to show the effect of 14–3–3 mediated by wild-type but not mutant Slob at 130 mV. Right, normalizedcurrents evoked by depolarizations to 1130 mV in patches from cells transfected with dSlo, 14–3–3, and either wild-type (iv) or mutant (v)Slob.
flies. The RQED1 fly line (Broughton et al., 1996) ex- exhibiting both green and blue fluorescence. The chan-nel distribution is normal in triply transfected cells, andpresses a constitutively active rat CaMKII (Waldmann
et al., 1990) under the control of a heat shock promoter. identical electrophysiological results were obtained inother experiments (data not shown) using nontaggedIn contrast, the ala2 fly line expresses a peptide inhibitor
of CaMKII, also under heat shock control (Griffith et al., dSlo and 14–3–3 constructs. Current traces were nor-malized as described in the Experimental Procedures.1993). As shown in Figure 5B, top panel, expression of
constitutively active CaMKII increases the binding of dSlo current evoked by a depolarizing voltage step to130 mV (Figure 6A, trace i) is not affected by the coex-Slob and 14–3–3 (Figure 5B, lane 2) relative to the non-
heat-shocked control (Figure 5B, lane 1). In contrast, pression of 14–3–3 (Figure 6A, trace ii) and is somewhatlarger when the channel is coexpressed with Slob (Fig-expression of the peptide inhibitor of CaMKII decreases
the binding (Figure 5B, lane 4) compared with the non- ure 6A, trace iii). In contrast, much less dSlo current isevoked by the same depolarizing voltage step when allheat-shocked control (Figure 5B, lane 3). Heat shock
has no effect on the binding (Figure 5B, lanes 5 and 6) three proteins are expressed together (Figure 6A, traceiv), even though the GFP fluorescence confirms robustin another fly line transformed with vector alone (no
coding insert under heat shock control) (Griffith et al., expression and membrane targeting of dSlo in thesecells (data not shown). The mean relative peak conduc-1993). Note also that the expression and immunoprecipi-
tation of 14–3–3 are the same in all of the transgenic fly tance (GRel) evoked by voltage steps to 130 mV was0.64 6 0.05 (mean 6 SEM, n 5 6) in patches from cellslines, with or without heat shock (Figure 5B, bottom
panel). These results demonstrate clearly that the inter- expressing dSlo alone, 0.70 6 0.04 in patches from cellscoexpressing dSlo and 14–3–3 (n 5 9), 0.81 6 0.03 inaction of Slob and 14–3–3 is not static but can be influ-
enced rapidly by changes in CaMKII activity in the fly. patches from cells expressing dSlo and Slob (n 5 4),but only 0.29 6 0.09 in patches from cells expressingdSlo, Slob, and 14–3–3 (n 5 5). Thus, the dSlo current14–3–3 Modulates dSlo via Slob
dSlo channel activity was examined in detached mem- evoked by depolarization to 130 mV is inhibited about65% by 14–3–3. To determine whether 14–3–3 bindingbrane patches from transfected tsA201 cells to test the
possibility that the channel might be modulated by 14– to Slob is required for this effect, dSlo and 14–3–3 werecoexpressed with the S54A/S79A mutant Slob that can3–3. dSlo–green fluorescent protein (–GFP) and 14–3–3–
blue fluorescent protein (–BFP) constructs were used in bind dSlo but does not bind to 14–3–3 (see Figure 4).As shown in Figure 6A (trace v), dSlo current in patchesthe experiments shown here (see Experimental Proce-
dures), and patch recording was done only on cells from these cells (GRel, 0.82 6 0.03; n 5 6) is essentially
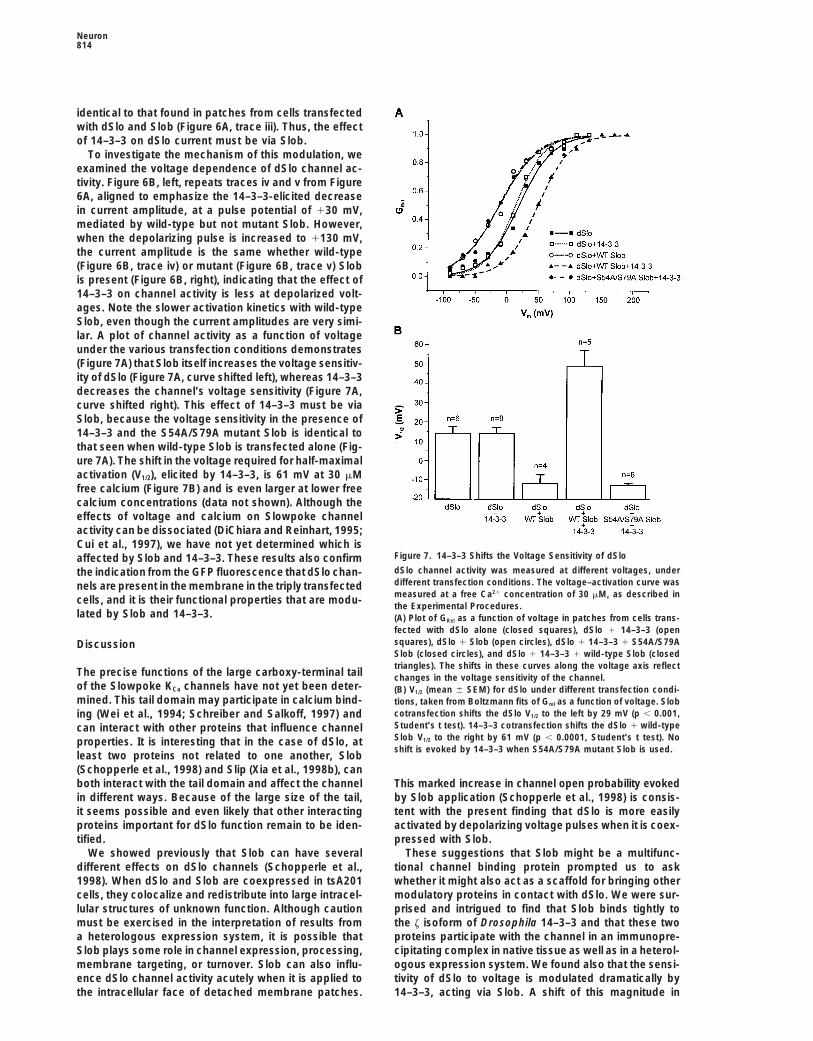
Neuron814
identical to that found in patches from cells transfectedwith dSlo and Slob (Figure 6A, trace iii). Thus, the effectof 14–3–3 on dSlo current must be via Slob.
To investigate the mechanism of this modulation, weexamined the voltage dependence of dSlo channel ac-tivity. Figure 6B, left, repeats traces iv and v from Figure6A, aligned to emphasize the 14–3–3-elicited decreasein current amplitude, at a pulse potential of 130 mV,mediated by wild-type but not mutant Slob. However,when the depolarizing pulse is increased to 1130 mV,the current amplitude is the same whether wild-type(Figure 6B, trace iv) or mutant (Figure 6B, trace v) Slobis present (Figure 6B, right), indicating that the effect of14–3–3 on channel activity is less at depolarized volt-ages. Note the slower activation kinetics with wild-typeSlob, even though the current amplitudes are very simi-lar. A plot of channel activity as a function of voltageunder the various transfection conditions demonstrates(Figure 7A) that Slob itself increases the voltage sensitiv-ity of dSlo (Figure 7A, curve shifted left), whereas 14–3–3decreases the channel’s voltage sensitivity (Figure 7A,curve shifted right). This effect of 14–3–3 must be viaSlob, because the voltage sensitivity in the presence of14–3–3 and the S54A/S79A mutant Slob is identical tothat seen when wild-type Slob is transfected alone (Fig-ure 7A). The shift in the voltage required for half-maximalactivation (V1/2), elicited by 14–3–3, is 61 mV at 30 mMfree calcium (Figure 7B) and is even larger at lower freecalcium concentrations (data not shown). Although theeffects of voltage and calcium on Slowpoke channelactivity can be dissociated (DiChiara and Reinhart, 1995;Cui et al., 1997), we have not yet determined which is
Figure 7. 14–3–3 Shifts the Voltage Sensitivity of dSloaffected by Slob and 14–3–3. These results also confirmdSlo channel activity was measured at different voltages, underthe indication from the GFP fluorescence that dSlo chan-different transfection conditions. The voltage–activation curve wasnels are present in the membrane in the triply transfectedmeasured at a free Ca21 concentration of 30 mM, as described incells, and it is their functional properties that are modu-the Experimental Procedures.
lated by Slob and 14–3–3. (A) Plot of GRel as a function of voltage in patches from cells trans-fected with dSlo alone (closed squares), dSlo 1 14–3–3 (opensquares), dSlo 1 Slob (open circles), dSlo 1 14–3–3 1 S54A/S79ADiscussionSlob (closed circles), and dSlo 1 14–3–3 1 wild-type Slob (closedtriangles). The shifts in these curves along the voltage axis reflect
The precise functions of the large carboxy-terminal tail changes in the voltage sensitivity of the channel.of the Slowpoke KCa channels have not yet been deter- (B) V1/2 (mean 6 SEM) for dSlo under different transfection condi-mined. This tail domain may participate in calcium bind- tions, taken from Boltzmann fits of Grel as a function of voltage. Slob
cotransfection shifts the dSlo V1/2 to the left by 29 mV (p , 0.001,ing (Wei et al., 1994; Schreiber and Salkoff, 1997) andStudent’s t test). 14–3–3 cotransfection shifts the dSlo 1 wild-typecan interact with other proteins that influence channelSlob V1/2 to the right by 61 mV (p , 0.0001, Student’s t test). Noproperties. It is interesting that in the case of dSlo, atshift is evoked by 14–3–3 when S54A/S79A mutant Slob is used.
least two proteins not related to one another, Slob(Schopperle et al., 1998) and Slip (Xia et al., 1998b), canboth interact with the tail domain and affect the channel This marked increase in channel open probability evoked
by Slob application (Schopperle et al., 1998) is consis-in different ways. Because of the large size of the tail,it seems possible and even likely that other interacting tent with the present finding that dSlo is more easily
activated by depolarizing voltage pulses when it is coex-proteins important for dSlo function remain to be iden-tified. pressed with Slob.
These suggestions that Slob might be a multifunc-We showed previously that Slob can have severaldifferent effects on dSlo channels (Schopperle et al., tional channel binding protein prompted us to ask
whether it might also act as a scaffold for bringing other1998). When dSlo and Slob are coexpressed in tsA201cells, they colocalize and redistribute into large intracel- modulatory proteins in contact with dSlo. We were sur-
prised and intrigued to find that Slob binds tightly tolular structures of unknown function. Although cautionmust be exercised in the interpretation of results from the z isoform of Drosophila 14–3–3 and that these two
proteins participate with the channel in an immunopre-a heterologous expression system, it is possible thatSlob plays some role in channel expression, processing, cipitating complex in native tissue as well as in a heterol-
ogous expression system. We found also that the sensi-membrane targeting, or turnover. Slob can also influ-ence dSlo channel activity acutely when it is applied to tivity of dSlo to voltage is modulated dramatically by
14–3–3, acting via Slob. A shift of this magnitude inthe intracellular face of detached membrane patches.

Presynaptic Protein Complex815
voltage sensitivity will profoundly influence dSlo channel transmission. We have confirmed this presynaptic local-activity in neurons. Voltage and calcium interact in ization of 14–3–3 and show in addition that it colocalizescomplex ways to activate Slowpoke channels (DiChiara with both dSlo and Slob in presynaptic boutons. Slow-and Reinhart, 1995; Cui et al., 1997), and we have not poke channels are also enriched in presynaptic endingsyet determined the precise biophysical mechanism by in rat brain (Knaus et al., 1996) and frog neuromuscularwhich 14–3–3 modulates dSlo voltage sensitivity. junction (Robitaille et al., 1993), where they influence
What are the molecular details of the profound down- transmitter release (Robitaille and Charlton, 1992). Sinceregulation of dSlo channel activity by 14–3–3, described dSlo current contributes to membrane repolarizationhere? Members of the family of KCa channels are subject and helps to limit transmitter release from Drosophilato modulation by a variety of molecular mechanisms, nerve terminals (Gho and Ganetzky, 1992), and 14–3–3ranging from protein phosphorylation (Levitan, 1994) to downregulates dSlo via Slob, it is plausible that thereoxidation/reduction reactions (DiChiara and Reinhart, is greater nerve terminal dSlo current in 14–3–3 mutant1997). It is conceivable that simply the binding of 14–3–3 flies and that this accounts for the decreased synapticto dSlo via Slob is sufficient to alter the gating of the transmission seen in these mutants (Broadie et al.,channel, as appears to be the case for b subunit interac- 1997). The possibility that a modulatory complex associ-tions with Slowpoke (McManus et al., 1995) and other ated with a neuronal ion channel may influence synapticpotassium channels (Rettig et al., 1994). Alternatively, transmission, and ultimately higher brain functions, is14–3–3 may act as another scaffolding component, to an attractive hypothesis for future investigation.bring one of the protein kinases that it is known to bind(Morrison, 1994; Pawson and Scott, 1997) into the prox- Experimental Proceduresimity of the channel. Indeed, because 14–3–3 dimerizes,
Yeast Two-Hybrid Screenit might bridge the interactions of several differentThe two-hybrid screen was carried out according to a standardsignaling proteins with the channel (Braselmann andprotocol as described previously (Schopperle et al., 1998). cDNAMcCormick, 1995; Tzivion et al., 1998). It will be interest-containing the entire coding sequence of Slob was subcloned intoing to determine whether the Raf protein kinase, onethe bait vector, pEG202, which contains the LexA DNA binding
of the kinases that binds 14–3–3 (Fu et al., 1994), can domain and a histidine selection marker. This plasmid was trans-phosphorylate and modulate dSlo, because Raf is a key formed into EGY48 yeast cells, followed by the transformation of aplayer in the mitogen-activated protein (MAP) kinase LacZ reporter plasmid, pSH18–34, which has a LexA binding site and
a uracil selection marker. The yeast were subsequently transformedpathway that conveys signals from the plasma mem-with a Drosophila head cDNA library subcloned into the activationbrane to the cell nucleus (Burbelo and Hall, 1995). Activa-domain vector, pJG4–5. Approximately 2,000,000 transformed yeasttion of this pathway can influence ion channel expres-colonies were screened for Slob-interacting clones. Positive clones
sion and activity (Huang and Rane, 1994; Hilborn et were selected on plates lacking leucine, tryptophan, histidine, andal., 1998), and potassium channel activity in turn can uracil and assayed for b-galactosidase activity. The library plasmidsmodulate tyrosine kinase signaling in cells (Holmes et of true positive clones were isolated from yeast cells, and the se-
quences of their cDNAs were determined by DNA sequencing.al., 1997). Thus, the present findings raise the intriguingpossibility that a potassium channel regulatory complex
DNA Constructs and Mutagenesisis involved in MAP kinase signaling and the regulationcDNA containing the entire coding sequence of the z isoform ofof many fundamental cell processes.Drosophila 14–3–3 (kindly provided by R. Ganguly, University ofOur finding that the interaction of Slob with 14–3–3Tennessee) was subcloned into the mammalian expression vector
requires Slob phosphorylation is consistent with studies pcDNA3 with a 1D4 epitope tag (ETSQVAPA) attached to its carboxylof other 14–3–3 binding proteins (e.g., Muslin et al., 1996; terminus. GST–14–3–3 was constructed by PCR amplification ofYaffe et al., 1997). It is especially intriguing that the 14–3–3 sequence and subcloning into pGEX4T-1 vector (Phar-
macia). A similar approach was used to make 14–3–3–BFP, whichbinding can be regulated in vivo by changes in the activ-contains both the 14–3–3 and BFP coding sequences in the pcDNA3ity of CaMKII; these results suggest that there may bevector. The dSlo–GFP construct was made by excising the GFPdynamic physiological regulation of dSlo channel activ-coding sequence from the pEGFP-N1 vector (Clontech Laboratories)ity by 14–3–3 that depends on the phosphorylation stateand subcloning it at the carboxyl terminus of the dSlo cDNA in the
of Slob. In view of the presynaptic colocalization of the pcDNA3 vector. Slob in pcDNA3, GST–Slob, and Slob–GFP werethree proteins described here, it is interesting that it constructed as reported previously (Schopperle et al., 1998). Site-is the CaMKII phosphorylation of Slob that regulates directed mutagenesis of the Slob cDNA was performed using the
QuickChange strategy (Stratagene), with appropriate primers to14–3–3 binding. CaMKII is also present at a high concen-separately introduce single serine-to-alanine mutations at S54 ortration presynaptically in Drosophila (Wang et al., 1994),S79. After verifying the mutations by DNA sequencing, the S79Aand thus the same calcium rise that evokes transmittermutation was introduced into the S54A mutant by another Quick-release might promote phosphorylation of Slob, bindingChange reaction to produce the double mutant S54A/S79A Slob.
of 14–3–3, and downregulation of dSlo.14–3–3z is required for photoreceptor development in Fly Strains and Heat Shock Induction
Drosophila (Kockel et al., 1997), while another isoform, All fly stocks were raised at 208–258C on standard Drosophila me-14–3–3e, also influences photoreceptor development by dium. The Drosophila stocks used in the 14–3–3/Slob/dSlo coimmu-regulating Ras-mediated signaling pathways (Chang noprecipitation experiments were Canton S (wild-type) and slo4 ho-
mozygous; the latter contains a null mutation in the dSlo gene andand Rubin, 1997). It is particularly interesting that fliesdoes not produce any channel protein (Atkinson et al., 1991). Thelacking 14–3–3z are severely impaired in an olfactorytransgenic RQED1 flies that express constitutively active CaMKIIlearning task (Skoulakis and Davis, 1996) and exhibit(Broughton et al., 1996) or the ala2 flies that express an inhibitory
defects in basal synaptic transmission as well as in syn- peptide for CaMKII (Griffith et al., 1993) were generated by P ele-aptic plasticity (Broadie et al., 1997). 14–3–3 is enriched ment–mediated germline transformation (Spradling, 1986). Controlin presynaptic boutons of the neuromuscular junction transgenic flies were transformed with the CasPer-SV40 P element
vector, with no coding insert (Griffith et al., 1993). In all cases, gene(Broadie et al., 1997), consistent with a role in synaptic

Neuron816
expression was under the control of the heat shock promoter, antibodies. Samples were washed and then incubated with FITC-conjugated donkey anti-rabbit IgG (Jackson Laboratories) for 2 hrhsp70. Induction of the inhibitory peptide or active CaMKII in these
transgenic strains was accomplished by placing the adult flies, at room temperature. After further washes, the preparations wereincubated overnight at 48C with a mouse anti-FITC monoclonal anti-raised at 208–258C, into a 378C temperature-controlled water bath
for 60 min. The heat-shocked flies were then maintained at 208–258C body (Sigma), followed by incubation at room temperature with anFITC-conjugated donkey anti-mouse IgG (Jackson Laboratories).for an additional 30 min, and heads were harvested by freezing in
liquid nitrogen. For double labeling experiments, anti-14–3–3 antibody was appliedsimultaneously with anti-FITC antibody, followed by simultaneousincubation with both Texas Red–conjugated donkey anti-rabbit andAntibodies, Coimmunoprecipitation, and WesternFITC-conjugated donkey anti-mouse IgG. No fluorescence was ob-Blot Analysisserved in controls that lacked one of the primary antibodies or whenRabbit polyclonal anti-dSlo and anti-Slob antibodies were generatedprimary antibody was preabsorbed with antigen.and characterized as described previously (Schopperle et al., 1998).
After staining, the preparations were dehydrated with an ethanolThe anti-14–3–3 antibody was generated by immunizing rabbits withseries, cleared in 70% glycerol, and mounted in Vectashield mount-a GST–14–3–3 fusion protein that includes the entire sequence ofing medium (Vector Laboratories). Optical sections of the neuromus-Drosophila 14–3–3z. A specific antibody against 14–3–3 was ob-cular junction were collected on a confocal microscope (MRC-600,tained by affinity purification using CNBr-sepharose bead–coupledBio-Rad) using equal gain and aperture settings for all images.GST–14–3–3. This antibody but not the preimmune serum specifi-
cally recognizes 14–3–3 in immunoprecipitation, Western blot, andimmunohistochemistry experiments. Anti-1D4 monoclonal antibody Electrophysiologythat specifically recognizes the 1D4-tagged 14–3–3 was provided dSlo channel activity was recorded in inside-out detached mem-by D. Oprian (Brandeis University). brane patches from tsA201 cells expressing dSlo–GFP, either alone
tsA201 cells (Margolskee et al., 1993) were maintained in Dulbec- or together with Slob and/or 14–3–3–BFP. The functional propertiesco’s modified Eagle’s medium supplemented with 10% fetal bovine of dSlo and dSlo–GFP are indistinguishable, and the same resultsserum. A calcium phosphate transfection protocol (Sambrook et al., were obtained in other experiments using nontagged dSlo and 14–1989) was used to introduce cDNA into the cells. Forty-eight hours 3–3 (data not shown). cDNAs were transfected by a calcium phos-after transfection, cells were lysed in lysis buffer containing 1% phate protocol, as above. Recordings were done 1–3 days afterCHAPS, 20 mM Tris-HCl (pH, 7.5), 10 mM EDTA, 120 mM NaCl, 50 transfection on an Axiovert 25 inverted fluorescence microscopemM KCl, 1 mM Na3VO4, 2 mM DTT and protease inhibitors (1 mM (Zeiss). Cotransfected cells expressing both GFP and BFP fluores-PMSF, 1 mg/ml each aprotonin, leupeptin, and pepstatin A [Sigma]). cence were identified with an FITC and 4’,6-diamidino-2-phenylin-Drosophila head lysates were made by isolating heads using a sizing dole filter set. Patch electrodes with resistances of 2–4 MV weresieve and homogenizing them in the lysis buffer. After centrifugation pulled from borosilicate glass, fire polished and silicone coated.to remove insoluble debris from the lysates, the supernatant was Both the electrode and bath solutions contained 150 mM KCl, 2 mMused for coimmunoprecipitation by incubation with anti-1D4 or anti- MgCl2, 10 mM HEPES (pH, 7.2), and 0.5 mM BAPTA. Free calcium14–3–3 antibodies for 2 hr at 48C. The complexes were precipitated concentrations in the bath solutions were established as describedby incubation with protein A– or protein A/G–agarose. To immuno- previously (Schopperle et al., 1998). Macroscopic currents wereprecipitate 14–3–3 from fly heads, the polyclonal anti-14–3–3 anti- recorded from the inside-out patches with an Axopatch 200A ampli-bodies were coupled covalently to protein A–agarose beads by a fier. Experimental control and data acquisition were done usingstandard protocol (Harlow and Lane, 1988), and the complexes were pClamp 6 software. dSlo currents were evoked by 150 ms depolariz-eluted with 100 mM glycine (pH, 2.5). No IgG was observed in the ing voltage pulses from a holding potential of 2100 mV, and theeluates when these covalently coupled antibodies were used. deactivating tail currents measured after the ends of the depolariz-
Proteins in the cell lysates or immunoprecipitates were separated ing pulses were used to analyze the data at a constant driving forceon polyacrylamide gels and transferred to nitrocellulose mem- (at 2100 mV). To generate a voltage–activation curve, the peakbranes. After blocking with 5% nonfat milk in TBST (0.1% Tween tail current following activation at a given test voltage (Itail [V]) was20 in Tris-buffered saline), the blots were probed with appropriate normalized to the maximum peak tail current in that patch (Itail [max],primary antibodies in blocking buffer at 48C overnight. The mem- evoked by depolarizations to 1130 mV or higher) (Liman et al., 1991).branes were then washed with TBST and incubated with horseradish The normalized tail currents (Itail [V]/Itail [max]) 5 Grel) were then fit to aperoxidase–conjugated sheep anti-mouse or donkey anti-rabbit IgG Boltzmann function.(Amersham) for 1 hr at room temperature. The proteins were visual-ized with an enhanced chemiluminescence detection system (Am-
Acknowledgmentsersham).
This work was supported by grants to I. B. L. and L. C. G. We areIn Vitro Phosphorylation of GST Fusion Proteinsgrateful to R. Ganguly for providing us with 14–3–3z cDNA, DanPhosphorylation of wild-type and mutant GST–Slobs by CaMKII wasOprian for anti-1D4 monoclonal antibody, and Haipeng Yang forcarried out in 50 ml reactions containing 2 mg of purified GST–Slobcontributing to the Slob mutagenesis experiments. We also thankfusion protein, 50 mM PIPES (pH, 7.0), 15 mM MgCl2, 1 mM CaCl2,Mats Holmqvist and Ping Jin for helpful discussions and Ping Jin,50 mg/ml calmodulin, 50 mM ATP, and 2 ml Drosophila CaMKII R3Chris Miller, and Mats Holmqvist for their critical comments on theisoform, purified as described previously (GuptaRoy et al., 1996).manuscript.The samples were incubated at 308C for 1 hr. The mixture was then
added to cell lysates from tsA201 cells that had been transfectedwith 14–3–3, and 14–3–3 immunoprecipitation and Western blot Received November 25, 1998; revised February 26, 1999.analysis were carried out.
ReferencesImmunocytochemistryDrosophila larvae were examined by immunohistochemical staining
Adelman, J.P., Shen, K.-Z., Kavanaugh, M.P., Warren, R.A., Wu,using a procedure described previously (Tejedor et al., 1997). ThirdY.-N., Lagrutta, A., Bond, C.T., and North, R.A. (1992). Calcium-instar larvae of wild-type Drosophila were dissected along the dorsalactivated potassium channels expressed from cloned complemen-midline in Ca21-free saline, pinned open, and fixed at 48C for 15 mintary DNAs. Neuron 9, 209–216.in fresh Bouin’s solution (saturated picric acid, 37% formaldehyde,Aitken, A., Collinge, D.B., van Heusden, B.P.H., Isobe, T., Roseboom,and glacial acetic acid; 15:5:1) and then for 15 min at room tempera-P.H., Rosenfeld, G., and Soll, J. (1992). 14–3–3 proteins: a highlyture in Bouin’s solution containing 0.2% Triton X-100. After repeatedconserved, widespread family of eukaryotic proteins. Trends Bio-washing with 0.1% Triton X-100 in phosphate-buffered saline, thechem. Sci. 17, 498–501.preparations were incubated overnight at 48C for 1 hr at room tem-
perature with affinity-purified rabbit polyclonal anti-Slob or anti-dSlo Apel, E.D., Glass, D.J., Moscoso, L.M., Yancopoulos, G.D., and

Presynaptic Protein Complex817
Sanes, J.R. (1997). Rapsyn is required for MuSK signaling and re- GuptaRoy, B., Beckingham, K., and Griffith, L.C. (1996). Functionaldiversity in alternatively spliced isoforms of Drosophila calcium/cruits synaptic components to a MuSK-containing scaffold. Neuroncalmodulin-dependent protein kinase II. J. Biol. Chem. 271, 19846–18, 623–635.19851.Atkinson, N.S., Robertson, G.A., and Ganetzky, B. (1991). A compo-Hall, Z.W., and Sanes, J.R. (1993). Synaptic structure and develop-nent of calcium-activated potassium channels encoded by the Dro-ment: the neuromuscular junction. Cell 72, 99–121.sophila slo locus. Science 253, 551–555.
Harlow, E., and Lane, D. (1988). Antibodies: A Laboratory ManualBielefeldt, K., and Jackson, M.B. (1994). Phosphorylation and de-(Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press).phosphorylation modulate a Ca21-activated K1 channel in rat peptid-
ergic nerve terminals. J. Physiol. 475, 241–254. Hilborn, M.D., Vaillancourt, R.R., and Rane, S.G. (1998). Growth fac-tor receptor tyrosine kinases acutely regulate neuronal sodiumBrakeman, P.R., Lanahan, A.A., O’Brien, R., Roche, K., Barnes, C.A.,channels through the Src signaling pathway. J. Neurosci. 18,Huganir, R.L., and Worley, P.F. (1997). Homer: a protein that selec-590–600.tively binds metabotropic glutamate receptors. Nature 386, 284–288.Holmes, T.C., Fadool, D.A., Ren, R., and Levitan, I.B. (1996). Associa-Braselmann, S., and McCormick, F. (1995). BCR and RAF form ation of Src tyrosine kinase with a human potassium channel medi-complex in vivo via 14–3–3 proteins. EMBO J. 14, 4839–4848.ated by SH3 domain. Science 274, 2089–2091.Broadie, K., Rushton, E., Skoulakis, E.M.C., and Davis, R.L. (1997).Holmes, T.C., Berman, K., Swartz, J.E., Dagan, D., and Levitan, I.B.Leonardo, a Drosophila 14–3–3 protein involved in learning, regu-(1997). Expression of voltage-gated potassium channels decreaseslates presynaptic function. Neuron 19, 391–402.cellular protein tyrosine phosphorylation. J. Neurosci. 17, 8964–Broadie, K.S., and Bate, M. (1993). Development of the embryonic8974.neuromuscular synapse of Drosophila melanogaster. J. Neurosci.Huang, Y., and Rane, S.G. (1994). Potassium channel induction by13, 144–166.the Ras/Raf signal transduction cascade. J. Biol. Chem. 269, 31183–Broughton, S.J., Kane, N.S., Yoder, M., Greenspan, R.J., and Robi-31189.chon, A. (1996). Transport of CaM kinase along processes elicitedKim, E., Niethammer, M., Rothschild, A., Jan, Y.N., and Sheng, M.by neuronal contact evokes an inhibition of arborization and out-(1995). Clustering of Shaker-type K1 channels by interaction withgrowth in D. melanogaster cultured neurons. J. Cell. Biochem. 62,a family of membrane-associated guanylate kinases. Nature 378,484–494.85–88.
Burbelo, P.D., and Hall, A. (1995). Hot numbers in signal transduc-Kim, J.H., Liao, D., Lau, L.-F., and Huganir, R.L. (1998). SynGAP: ation. Curr. Biol. 5, 95–96.synaptic RasGAP that associates with the PSD-95/SAP90 protein
Catterall, W.A. (1993). Structure and modulation of Na1 and Ca21
family. Neuron 20, 683–691.channels. Ann. NY Acad. Sci. 707, 1–19.
Knaus, H.-G., Schwarzer, C., Koch, R.O.A., Eberhart, A., Kaczorow-Chang, H.C., and Rubin, G.M. (1997). 14–3–3e positively regulates ski, G.J., Glossmann, H., Wunder, F., Pongs, O., Garcia, M.L., andRas-mediated signaling in Drosophila. Genes Dev. 11, 1132–1139. Sperk, G. (1996). Distribution of high-conductance Ca21-activatedChung, S.K., Reinhart, P.H., Martin, B.L., Brautigan, D., and Levitan, K1 channels in rat brain: targeting to axons and nerve terminals. J.I.B. (1991). Protein kinase activity closely associated with a reconsti- Neurosci. 16, 955–963.tuted calcium-activated potassium channel. Science 253, 560–562. Kockel, L., Vorbruggen, G., Jackle, H., Mlodzik, M., and Bohmann,Colledge, M., and Froehner, S.C. (1997). Tyrosine phosphorylation D. (1997). Requirement for Drosophila 14–3–3zeta in Raf-dependentof nicotinic acetylcholine receptor mediates Grb2 binding. J. Neu- photoreceptor development. Genes Dev. 11, 1140–1147.rosci. 17, 5038–5045. Kornau, H.-C., Schenker, L.T., Kennedy, M.B., and Seeburg, P.H.Cui, J., Cox, D.H., and Aldrich, R.W. (1997). Intrinsic voltage depen- (1995). Domain interaction between NMDA receptor subunits anddence and Ca21 regulation of mslo large conductance Ca-activated the postsynaptic density protein PSD-95. Science 269, 1737–1740.K1 channels. J. Gen. Physiol. 109, 647–673. Latorre, R., Oberhauser, A., Labarca, P., and Alvarez, O. (1989).DiChiara, T.J., and Reinhart, P.H. (1995). Distinct effects of Ca21 and Varieties of calcium-activated potassium channels. Annu. Rev.voltage on the activation and deactivation of cloned Ca21-activated Physiol. 51, 385–399.K1 channels. J. Physiol. 489, 403–418. Levitan, I.B. (1994). Modulation of ion channels by protein phosphor-DiChiara, T.J., and Reinhart, P.H. (1997). Redox modulation of hslo ylation and dephosphorylation. Annu. Rev. Physiol. 56, 193–212.Ca21-activated K1 channels. J. Neurosci. 17, 4942–4955. Liman, E.R., Hess, P., Weaver, F., and Koren, G. (1991). Voltage-Dong, H., O’Brien, R.J., Fung, E.T., Lanahan, A.A., Worley, P.F., sensing residues in the S4 region of a mammalian K1 channel. Natureand Huganir, R.L. (1997). GRIP: a synaptic PDZ domain–containing 353, 752–756.protein that interacts with AMPA receptors. Nature 386, 279–284. Lin, J.W., Wyszynski, M., Madvavan, R., Sealock, R., Kim, J.U., andEhlers, M.D., Zhang, S., Bernhardt, J.P., and Huganir, R.L. (1996). Sheng, M. (1998). Yotiao, a novel protein of neuromuscular junctionInactivation of NMDA receptors by direct interaction of calmodulin and brain that interacts with specific splice variants of NMDA recep-with the NR1 subunit. Cell 84, 745–755. tor subunit NR1. J. Neurosci. 18, 2017–2027.Ehlers, M.D., Fung, E.T., O’Brien, R.J., and Huganir, R.L. (1998). Margolskee, R.F., McHendry-Rinde, B., and Horn, R. (1993). PanningSplice variant–specific interaction of the NMDA receptor subunit transfected cells for electrophysiological studies. Biotechniques 15,NR1 with neuronal intermediate filaments. J. Neurosci. 18, 720–730. 906–911.Fields, S., and Song, O. (1989). A novel genetic system to detect McManus, O.B., Helms, L.M.H., Pallanck, L., Ganetzky, B., Swanson,protein–protein interactions. Nature 340, 245–246. R., and Leonard, R.J. (1995). Functional role of the b subunit of high
conductance calcium-activated potassium channels. Neuron 14,Fu, H., Xia, K., Pallas, D.C., Cui, C., Conroy, K., Narsimhan, R.P.,645–650.Mamon, H., Collier, R.J., and Roberts, T.M. (1994). Interaction of the
protein kinase raf-1 with 14–3–3 proteins. Science 266, 126–129. Morrison, D. (1994). 14–3–3: modulators of signaling proteins? Sci-ence 266, 56–57.Fuhrer, C., and Hall, Z.W. (1996). Functional interaction of Src family
kinases with the acetylcholine receptor in C2 myotubes. J. Biol. Muller, B.M., Kistner, U., Kindler, S., Chung, W.J., Kuhlendahl, S.,Chem. 271, 32474–32481. Fenster, S.D., Lau, L.-F., Veh, R.W., Huganir, R.L., Gundelfiner, E.D.,
and Garner, C.C. (1996). SAP102, a novel postsynaptic protein thatGho, M., and Ganetzky, B. (1992). Analysis of repolarization of pre-interacts with NMDA receptor complexes in vivo. Neuron 17,synaptic motor terminals in Drosophila larvae using potassium chan-255–265.nel–blocking drugs and mutations. J. Exp. Biol. 170, 93–111.Muslin, A.J., Tanner, J.W., Allen, P.M., and Shaw, A.S. (1996). Inter-Griffith, L.C., Verselis, L.M., Aitken, K.M., Kyriacou, C.P., Danho,action of 14–3–3 with signaling proteins is mediated by the recogni-W., and Greenspan, R.J. (1993). Inhibition of calcium/calmodulin-tion of phosphoserine. Cell 84, 889–897.dependent protein kinase in Drosophila disrupts behavioral plastic-
ity. Neuron 10, 501–509. Nara, M., Dhulipala, P.D.K., Wang, Y.-X., and Kotlikoff, M.I. (1998).

Neuron818
Reconstitution of b-adrenergic modulation of large conductance, Wallace, B.G. (1995). Regulation of the interaction of nicotinic acetyl-calcium-activated potassium (Maxi-K) channels in Xenopus oo- choline receptors with the cytoskeleton by agrin-activated proteincytes. J. Biol. Chem. 273, 14920–14924. tyrosine kinase. J. Cell Biol. 128, 1121–1129.
Niethammer, M., Valtschanoff, J.G., Kapoor, T.M., Allison, D.W., Wang, J., Renger, J., Griffith, L.C., Greenspan, R.J., and Wu, C.-F.Weinberg, R.J., Craig, A.M., and Sheng, M. (1998). CRIPT, a novel (1994). Concomitant alterations of physiological and developmentalpostsynaptic protein that binds to the third PDZ domain of PSD- plasticity at CaM kinase II–inhibited synapses in Drosophila. Neuron95/SAP90. Neuron 20, 693–707. 13, 1373–1384.Pawson, T., and Scott, J.D. (1997). Signaling through scaffold, an- Wei, A., Solaro, C., Lingle, C., and Salkoff, L. (1994). Calcium sensitiv-choring, and adaptor proteins. Science 278, 2075–2080. ity of BK-type KCa channels determined by a separable domain.
Neuron 13, 671–681.Reinhart, P.H., and Levitan, I.B. (1995). Kinase and phosphataseactivities intimately associated with a reconstituted calcium-depen- White, R.E., Schonbrunn, A., and Armstrong, D.L. (1991). Somato-dent potassium channel. J. Neurosci. 15, 4572–4579. statin stimulates Ca21-activated K1 channels through protein de-Reinhart, P.H., Chung, S.K., Martin, B.L., Brautigan, D.L., and Levi- phosphorylation. Nature 351, 570–573.tan, I.B. (1991). Modulation of calcium-activated potassium channels White, R.E., Lee, A.B., Shcherbatko, A.D., Lincoln, T.M., Schon-from rat brain by protein kinase A and phosphatase 2A. J. Neurosci. brunn, A., and Armstrong, D.L. (1993). Potassium channel stimula-11, 1627–1635. tion by natriuretic peptides through cGMP-dependent dephosphor-Rettig, J., Heinemann, S.H., Wunder, G., Lorra, C., Parcej, D.N., ylation. Nature 361, 263–266.Dolly, J.O., and Pongs, O. (1994). Inactivation properties of voltage- Xia, X.-M., Fakler, B., Rivard, A., Wayman, G., Johnson-Pais, T.,gated K1 channels altered by presence of b-subunit. Nature 369, Keen, J.E., Ishii, T., Hirschberg, B., Bond, C.T., Lutsenko, S., Maylie,289–294. J., and Adelman, J.P. (1998a). Mechanism of calcium gating in small-Robitaille, R., and Charlton, M.P. (1992). Presynaptic calcium signals conductance calcium-activated potassium channels. Nature 395,and transmitter release are modulated by calcium-activated potas- 503–507.sium channels. J. Neurosci. 12, 297–305.
Xia, X.-M., Hirschberg, B., Smolik, S., Forte, M., and Adelman, J.P.Robitaille, R., Garcia, M.L., Kaczorowski, G.J., and Charlton, M.P. (1998b). dSlo interacting protein 1, a novel protein that interacts(1993). Functional colocalization of calcium and calcium-gated po- with large-conductance calcium-activated potassium channels. J.tassium channels in control of transmitter release. Neuron 11, Neurosci. 18, 2360–2369.645–655.
Xiao, B., Tu, J.C., Petralia, R.S., Yuan, J.P., Doan, A., Breder, C.D.,Sambrook, J., Fritsch, E.F., and Maniatas, T. (1989). Molecular Clon- Ruggiero, A., Lanahan, A.A., Wenthold, R.J., and Worley, P.F. (1998).ing: A Laboratory Manual (Cold Spring Harbor, NY: Cold Spring Homer regulates the association of Group 1 metabotropic glutamateHarbor Laboratory Press). receptors with multivalent complexes of homer-related, synapticSchopperle, W.M., Holmqvist, M.H., Zhou, Y., Wang, J., Wang, Z., proteins. Neuron 21, 707–716.Griffith, L.C., Keselman, I., Kusinitz, F., Dagan, D., and Levitan, I.B.
Yaffe, M.B., Rittinger, K., Volinia, S., Caron, P.R., Aitken, A., Leffers,(1998). Slob, a novel protein that interacts with the Slowpoke cal-
H., Gamblin, S.J., Smerdon, S.J., and Cantley, L.C. (1997). The struc-cium-dependent potassium channel. Neuron 20, 565–573.tural basis for 14–3–3:phosphopeptide binding specificity. Cell 91,
Schreiber, M., and Salkoff, L. (1997). A novel calcium-sensing do- 961–971.main in the BK channel. Biophys. J. 73, 1355–1363.
Yu, X.-M., Askalan, R., Keil, G.J., and Salter, M.W. (1997). NMDAShipston, M.J., and Armstrong, D.L. (1996). Activation of protein channel regulation by channel-associated protein tyrosine kinasekinase C inhibits calcium-activated potassium channels in rat pitu- Src. Science 275, 674–678.itary tumour cells. J. Physiol. 493, 665–672.
Skoulakis, E.M.C., and Davis, R.L. (1996). Olfactory learning deficitsin mutants for leonardo, a Drosophila gene encoding a 14–3–3 pro-tein. Neuron 17, 931–944.
Spradling, A.C. (1986). P element–mediated transformation. In Dro-sophila: A Practical Approach, D.B. Roberts, ed. (Oxford: IRL Press),pp. 175–197.
Swanson, K.D., and Ganguly, R. (1992). Characterization of a Dro-sophila melanogaster gene similar to the mammalian genes encod-ing the tyrosine/tryptophan hydroxylase activator and protein kinaseC inhibitor proteins. Gene 113, 183–190.
Swope, S.L., and Huganir, R.L. (1994). Binding of the nicotinic acetyl-choline receptor to SH2 domains of Fyn and Fyk protein tyrosinekinases. J. Biol. Chem. 269, 29817–29824.
Swope, S.L., Moss, S.J., Blackstone, C.D., and Huganir, R.L. (1992).Phosphorylation of ligand-gated ion channels: a possible mode ofsynaptic plasticity. FASEB J. 6, 2514–2523.
Tejedor, F.J., Bokhari, A., Rogero, O., Gorczyca, M., Zhang, J., Kim,E., Sheng, M., and Budnik, V. (1997). Essential role for dlg in synapticclustering of Shaker K1 channels in vivo. J. Neurosci. 17, 152–159.
Tian, L., Knaus, H.-G., and Shipston, M.J. (1998). Glucocorticoidregulation of calcium-activated potassium channels mediated byserine/threonine protein phosphatase. J. Biol. Chem. 273, 13531–13536.
Tseng-Crank, J., Foster, C.D., Krause, J.D., Mertz, R., Godinot, N.,DiChiara, T.J., and Reinhart, P.H. (1994). Cloning, expression, anddistribution of functionally distinct Ca21-activated K1 channel iso-forms from human brain. Neuron 13, 1315–1330.
Tzivion, G., Luo, Z., and Avruch, J. (1998). A dimeric 14–3–3 proteinis an essential cofactor for Raf kinase activity. Nature 394, 88–92.
Waldmann, R., Hanson, P.I., and Schulman, H. (1990). Multifunc-tional Ca21/calmodulin-dependent protein kinase made Ca21 inde-pendent for functional studies. Biochemistry 29, 1679–1684.