septin 3 (g-septin) is a developmentally regulated phosphoprotein enriched in presynaptic nerve...
TRANSCRIPT

Septin 3 (G-septin) is a developmentally regulated phosphoproteinenriched in presynaptic nerve terminals
Jing Xue,*,1 Christopher W. Tsang,�,1 Wei-Ping Gai,� Chandra S. Malladi,* William S. Trimble,�John A. P. Rostas§ and Phillip J. Robinson*
*Cell Signalling Unit, Children’s Medical Research Institute, New South Wales, Australia
�Programmes in Brain and Behaviour and Cell Biology, Hospital for Sick Children, Toronto, Canada
�Department of Physiology and Centre for Neuroscience, Flinders University, Bedford Park, South Australia, Australia
§School of Biomedical Sciences and the Hunter Medical Research Institute, The University of Newcastle, New South Wales, Australia
Abstract
The septins are GTPase enzymes with multiple roles in
cytokinesis, cell polarity or exocytosis. The proteins from the
mammalian septin genes are called Sept1–10. Most are
expressed in multiple tissues, but the mRNA for Sept5
(CDCrel-1) and Sept3 (G-septin) appear to be primarily ex-
pressed in brain. Sept3 is phosphorylated by cGMP-depend-
ent protein kinase I (PKG-I) and the cGMP/PKG pathway is
involved in presynaptic plasticity. Therefore to determine
whether Sept3 specifically associates with neurones and
nerve terminals we investigated its distribution in rat brain and
neuronal cultures. Sept3 protein was detected only in brain by
immunoblot, but not in 12 other tissues examined. Levels
were high in all adult brain regions, and reduced in those
enriched in white matter. Expression was developmentally
regulated, being absent in the early embryo, low in late
embryonic rat brain and increasing after birth. Like dynamin I,
Sept3 was specifically enriched in synaptosomes compared
with whole brain, and was only found in a peripheral mem-
brane extract and not in the soluble or membrane extracts.
Sept3 was particularly abundant in mossy fibre nerve termi-
nals in the hippocampus. In primary cultured hippocampal
neurones Sept3 immunoreactivity was punctate in neurites
and predominantly localized to presynaptic terminals, strongly
colocalizing with synaptophysin and dynamin I. The specific
nerve terminal localization was confirmed by immunogold
electron microscopy. Together this shows that Sept3 is a
neurone-specific protein highly enriched in nerve terminals
which supports a secretory role in synaptic vesicle recycling.
Keywords: cGMP, G-septin, PKG, protein phosphorylation,
septin 3, synaptosomes.
J. Neurochem. (2004) 91, 579–590.
The septins are a multigene family in mammals comprising10 genes, producing a variety of protein products by furtheralternative splicing. The proteins are named Sept1–10 andare GTPase enzymes of around 40–70 kDa (Macara et al.2002; Surka et al. 2002). The GTPase domain comprises thebulk of the sequence and no other large domains are found,although a few short functional motifs are known (Sarasteet al. 1990; Zhang et al. 1999). The septins have multiplecellular functions, primarily playing an essential role incytokinesis (Kinoshita et al. 1997; Hsu et al. 1998; Xie et al.1999; Surka et al. 2002). In non-mitotic cells septins appearto assemble into filaments and/or associate with the plasmamembrane (Xie et al. 1999; Fung and Scheller 1999; Zhanget al. 1999). Groups of different septins also assemble asfilaments in vitro (Field et al. 1996; Hsu et al. 1998;Kinoshita 2003). The filaments appear to be strongly
associated with either actin (Kinoshita et al. 1997; Kinoshitaet al. 2002) or tubulin networks (Surka et al. 2002), or insome cases with both (Surka et al. 2002). The filamentscan self-assemble into higher-order structures to form
Received April 22, 2004; revised manuscript received June 25, 2004;accepted June 29, 2004.Address correspondence and reprint requests to Phillip J. Robinson,
Children’s Medical Research Institute, 214 Hawkesbury Road West-mead, NSW 2145 Australia. E-mail: [email protected] authors contributed equally to this work.Abbreviations used: BSA, bovine serum albumin; GL, granular layer;
MF, mossy fibre; ML, molecular layer; NGS, normal goat serum; PBS,phosphate-buffered saline; PKG-I, cGMP-dependent protein kinase I;PL, Purkinje cell layer; SDS, sodium dodecyl sulphate; WM, whitematter.
Journal of Neurochemistry, 2004, 91, 579–590 doi:10.1111/j.1471-4159.2004.02755.x
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590 579

cytoplasmic rings and coils, or form linear bundles in thepresence of actin filaments (Kinoshita et al. 2002).
A number of septins are found in postmitotic neurones andhave potential roles associated with synaptic transmission.Sept7 is present in postsynaptic density fractions (Walikoniset al. 2000), suggesting a role in the cell architecture. Onegroup [Sept6 (KIAA0128), Sept7 (cdc10), Sept2 (Nedd5)and Sept4 (H5)] associates with the exocyst complex, whichis involved in polarized vesicle secretion (Hsu et al. 1998;Vega and Hsu 2003). Sept5 (CDCrel-1) and Sept2 coimmu-noprecipitate with the SNARE protein syntaxin (Beites et al.1999; Dent et al. 2002). Targeted disruption of the mouseSept5 gene produces no major phenotype (Peng et al. 2002),but platelets from Sept5 –/– mice are sensitized to collagen-induced activation and serotonin secretion (Dent et al. 2002),again supporting a role in secretion. Sept2 interacts with theastrocyte glutamate transporter GLAST and regulates glu-tamate uptake in the brain (Kinoshita et al. 2004).
Septins are linked to several neurodegenerative disorders.Sept2, Sept4, Sept1 and Sept7 are found in neurofibrillarytangles (Kinoshita et al. 1998) and Sept7 expression is down-regulated in mouse models of Alzheimer’s disease (Liauwet al. 2002). Sept4, but not 5 other septins, is found in thea-synuclein-positive cytoplasmic inclusions of Parkinson’sdisease, dementia with Lewy bodies, and multiple systematrophy (Ihara et al. 2003). Sept5 interacts with Parkin, an E3ubiquitin-protein ligase implicated in autosomal recessivefamilial Parkinson’s disease, promoting Sept5 degradation(Zhang et al. 2000). Sept5 over-expression in the brain inducesselective dopamine neurodegeneration and inhibits dopaminesecretion (Dong et al. 2003). These studies are indicative ofimportant roles for septins in the mature brain.
We previously reported the cloning of Sept3, and that it isa specific substrate for cGMP-dependent protein kinase(PKG) in vitro (Xue et al. 2000). Its phosphorylation isregulated by cyclic nucleotides in nerve terminals in vivo.Sept3 and Sept5 are the only two septins to date that seem tobe primarily brain-specific (Yagi et al. 1998; Beites et al.1999; Xue et al. 2000). In mouse brain Sept5 is abundant ininhibitory presynaptic terminals and is associated withGABAergic vesicles (Kinoshita et al. 2000). However,nothing is known concerning the tissue distribution of Sept3protein and its subcellular localization. Hence our primaryaim was to determine whether Sept3 might be specificallyenriched in nerve terminals. We report a highly specificlocalization of Sept3 in nerve terminals, suggesting a role inthe dynamics of synaptic vesicle recycling.
Materials and methods
Protein purification and expression
Sept3 (G-septin) was purified from rat brain (Xue et al. 2000). TheGST-tagged Sept2 (Nedd5, mouse sequence) plasmid was from
Makoto Kinoshita (Kyoto, Japan) and the recombinant protein was
expressed in Escherichia coli and purified on glutathione Sepharose
column (Amersham Pharmacia, Piscataway, NJ, USA) following the
manufacturer’s instructions.
Antibodies
Sheep anti-Sept3 antibodies were raised against two different
synthetic peptides Sept3 a97)112 and Sept3 a244)259 as previously
described (Xue et al. 2000). The rabbit anti-Sept2 antibodies were
raised against two different synthetic peptides: Sept297)112 EKI-
PKTVEIKAIGHVI and Sept2244)259 DLEDKTENDKIRQESM.
The sequences for Sept3 and Sept2 peptides were not present in
any other septin or other protein in GenBank. The peptides were
synthesized and purified to greater than 90% purity by Auspep
(Australia) and were individually conjugated by coupling through
C-terminal cysteines added to the sequences during their synthesis
to diphtheria toxoid as a carrier and conjugates were then mixed
together to immunize the same rabbit (Institute of Medical and
Veterinary Science, Adelaide, Australia). Rabbit polyclonal anti-
serum to Sept3 was generated by immunizing a rabbit with a
synthetic peptide (MSELVPEPRPKPAVPC, Sheldon Biotechno-
logy, Montreal, Canada) conjugated to keyhole limpet haemocyanin
(Pierce, Rockford, IL, USA). This sequence (excluding the C-
terminal cysteine) corresponds to amino acids 1–15 of Sept3 which
is present in all known Sept3 isoforms but not in any other known
proteins in GenBank. The rabbit Sept3 antibody was then affinity-
purified from the antisera using the peptide coupled to a SulfoLink
column. NeuN and synaptophysin monoclonal antibodies were
purchased from Chemicon (Temecula, CA, USA). Cy3-conjugated
anti-GFAP antibodies and MAP2 were purchased from Sigma (St
Louis, MO, USA). The dynamin I antibody was as previously
described (Powell and Robinson 1995).
Immunoblotting
Various organs of the rat, or different brain regions were dissected
out at 4�C and homogenized in 20 mM Tris/HCl buffer, pH 7.4,
containing 1 mM EDTA, 1 mM EGTA, 7 lg/mL leupeptin, and
1 mM phenylmethylsulfonyl fluoride. Normalized protein levels
from each sample (60 lg) were denatured in a reducing sodium
dodecyl sulphate (SDS) sample buffer and resolved on 12%
polyacrylamide mini-gels. The proteins were transferred onto a
nitrocellulose transfer membrane (0.45 lm, Schleicher & Schuell,
Dassel Town, Germany) (Towbin et al. 1979). The membranes were
blocked in phosphate-buffered saline (PBS), pH 7.4 with 5%
skimmed milk overnight and washed in a Tris-buffered saline
(pH 7.4) containing 0.1% Tween 20 (TBST), then incubated with
the first antibody for 2 h and with the second antibody for 1 h in
TBST containing 0.5% polyvinylpyrrolidone-40, and washed with
TBST buffer. Sept3 was detected by incubation of the membranes
with anti-Sept3 antiserum from sheep (Xue et al. 2000). Immuno-
blotting was performed by chemiluminescent detection (Pierce
SuperSignal West Dura).
Preparation of hippocampal mossy fibre (MF) synaptosomes
Rat brain hippocampal MF synaptosomes were prepared as
described (Taupin et al. 1994) with minor modifications. Briefly,
P2 synaptosomes were prepared from rat hippocampus in 0.32 M
sucrose containing 5 mM Tris/HCl (pH 7.4) and 1 mM MgCl2. The
P2 pellet, retaining the MF synaptosomes, was washed once by
resuspension in the same buffer and re-centrifuged. The resuspended
580 J. Xue et al.
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590

P2 was submitted to fractionation on a three-step Percoll gradient
(consisting of 2.3 mL of 10, 15, and 23% Percoll in the same buffer
as above) (Dunkley et al. 1986). The synaptosomes between 10 and
15% Percoll were collected as MF3 synaptosomes and those
between 15 and 23% Percoll were collected as MF4 synaptosomes.
The MF synaptosomes are larger and denser and are mostly
concentrated in MF4 (Taupin et al. 1994). Both fractions were
pelleted and washed once with 350 mM NaCl to remove extracel-
lular peripheral membrane protein contaminants, then twice with
PBS. Synaptosomes were then lysed in 5 mM Tris/HCl pH 7.4
containing both protease inhibitor tablets (Roche, Indianapolis, IN,
USA) and phosphatase inhibitor tablets (Calbiochem, San Diego,
CA, USA). After centrifugation at 20 000 · g for 15 min to collect
cytosol, the particulate fractions were extracted with 250 mM NaCl
in the same buffer for 15 min and re-centrifuged to collect the
peripheral membrane extract.
Immunohistochemistry
Adult Sprague-Dawley rats were treated with an overdose of
Nembutol, perfused transcardially with 50 mL of 0.1 M phosphate
buffer saline at pH 7.4 (PBS), followed by 200 mL 4% formalde-
hyde and 2% picric acid in PBS. The brain was removed, stored in
30% sucrose in PBS at 4�C, and cut on a frozen microtome to 40-
lm sections. Sections were washed three times for 10 min each with
50% alcohol and three times with 0.1 M Tris-buffered saline at
pH 7.4 (TBS). The sections were incubated with 20% normal horse
serum in TBS for 1 h at room temperature to block non-specific
antibody binding sites, then incubated 4�C overnight in affinity
purified sheep anti-Sept3 IgG diluted 1 : 500 in TBS containing 1%
normal horse serum, 0.25% Triton-X 100 and 0.1% sodium azide.
Following three times 10-min washes in TBS sections were
incubated 2 h at room temperature in TBS with biotin-conjugated
donkey anti-sheep IgG (1 : 200, Jackson Laboratory). Following
three times 10-min washes in TBS sections were incubated for
60 min at room temperature with streptavidin-biotin-peroxidase
solution prepared using the ABC kit (Vector, Laboratory, Burlin-
game, CA, USA). Immunoreactivity was visualized using the
diaminobenzidine (DAB)–H2O2 reaction. Sections were dehydrated,
cover-slipped, and examined using a BH-2 Olympic light micro-
scope.
Cell culture and immunostaining
Rat embryonic hippocampal cell cultures were prepared from
hippocampi of 18-day-old fetal rat brains. Dissection and removal
of the hippocampus was described previously (Banker and Goslin
1991). Hippocampal cells were plated and maintained in culture
as described (Brewer et al. 1993). On the first day, triturated cells
were plated in neurobasal media (Life Technologies) supplemen-
ted with 2% B27 (Life Technologies), 3% FBS, 0.5 mM L-
glutamine (Life Technologies) and 25 lM glutamic acid (Sigma, St
Louis, MO, USA) on glass coverslips (Carolina Biological
Supply) coated with poly D-lysine (Sigma). Twenty-four hours
later and every 3 days afterwards, half the media was replaced
with fresh Neurobasal supplemented with B27 and glutamine only.
After 14 days in vitro, the cells were fixed with 4% paraformal-
dehyde in PBS for 20 min, and then blocked with 5% bovine
serum albumin (BSA) in PBS for 1 h at room temperature.
Antibodies to Sept3 (1 : 500), synaptophysin (1 : 200), NeuN
(1 : 200), and GFAP (1 : 1000) were incubated overnight at 4�Cin PBS containing 0.5% BSA. In studies on the colocalization of
Sept3 and dynamin, after fixation the cells were permeabilized for
15 min with 0.1% SDS prior to blocking. Fluorescent-labelled
secondary antibodies were incubated for 1 h at room temperature
in PBS containing 0.5% BSA. For multilabelling experiments,
primary and secondary antibodies were incubated in succession.
Images were acquired with a Zeiss LSM510 laser scanning
confocal microscope and documented with Adobe Photoshop and
Illustrator. For pure glial cultures, freshly dissected hippocampal
cells were maintained in Neurobasal media supplemented with a
G-5 supplement (Invitrogen) which supports the growth of
primary glial but not neuronal cells. Glia were maintained by
exchanging half the bath volume with fresh media every three
days. Glial cells were solubilized directly in SDS loading buffer
for analysis by western blotting.
Immunogold electron microscopy
Rat embryonic hippocampal neurone cultures were prepared as
above. Cultured 14-day hippocampal neurones were fixed with 4%
paraformaldehyde and 0.05% glutaraldehyde in PBS for 20 min.
The fixative was quenched with 0.1% glycine (Sigma) and 0.05%
sodium borohydride in PBS for 10 min and then the neurones were
blocked with 5.0% normal goat serum (NGS) and 5.0% BSA in PBS
for 30 min. Endogenous Sept3 was labelled with a rabbit anti-Sept3
antibody (1 : 500) for 1.5 h followed by a goat anti-rabbit ultra
small gold-conjugated secondary antibody (1 : 100; Aurion,
Wageningen, the Netherlands) for 4 h. All steps were carried out
at room temperature in PBS with 1% NGS and 1% BSA. After
labelling with antibodies, the neurones were fixed with 2%
glutaraldehyde in phosphate buffer (PB) overnight at 4�C and then
washed with PB. The ultra small gold particles were enhanced with
silver (R-Gent SE-EM kit, Aurion, AA Wageningen, Netherlands)
after chelating non-specific nucleating agents with an enhancement
conditioning solution (ECS; Aurion) according to the manufac-
turer’s instructions. The silver-enhanced gold particles were re-
coated with gold (gold-toning) by incubation with a 0.05% gold
chloride solution for 10 min at 4�C first, followed by 0.5% oxalic
acid solution for 2 min at room temperature, and then a 1.0%
sodium thiosulfate solution for 45 min at room temperature.
Neurones were then stained with 0.5% osmium tetroxide in PB
for 15 min and 1% uranyl acetate in water for 15 min. Once the
neurones were dehydrated with increasing concentrations of ethanol
they were embedded in Spurr resin (Canemco Inc., Montreal,
Canada). Sections were cut en face, stained with 2.0% uranyl acetate
for 15 min and 0.2% lead citrate for 5 min and then imaged with a
Philips CM100 Electron Microscope.
Results
Sept3 is a brain-specific protein
We and others previously detected Sept3 mRNA exclusivelyin brain by northern blot analysis of tissues from rat, mouseor human (Xiong et al. 1999; Xue et al. 2000; Methner et al.2001). To determine whether the expression of Sept3 proteinwas also brain-specific immunoblot analysis was performedon 13 specific rat tissue extracts: brain, intestine, testis,stomach, kidney, skeletal muscle, pancreas, lung, liver, heart,spleen, aorta, and adrenal gland. Cytosolic, peripheral
Sept3 (G-septin) localization in nerve terminals 581
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590
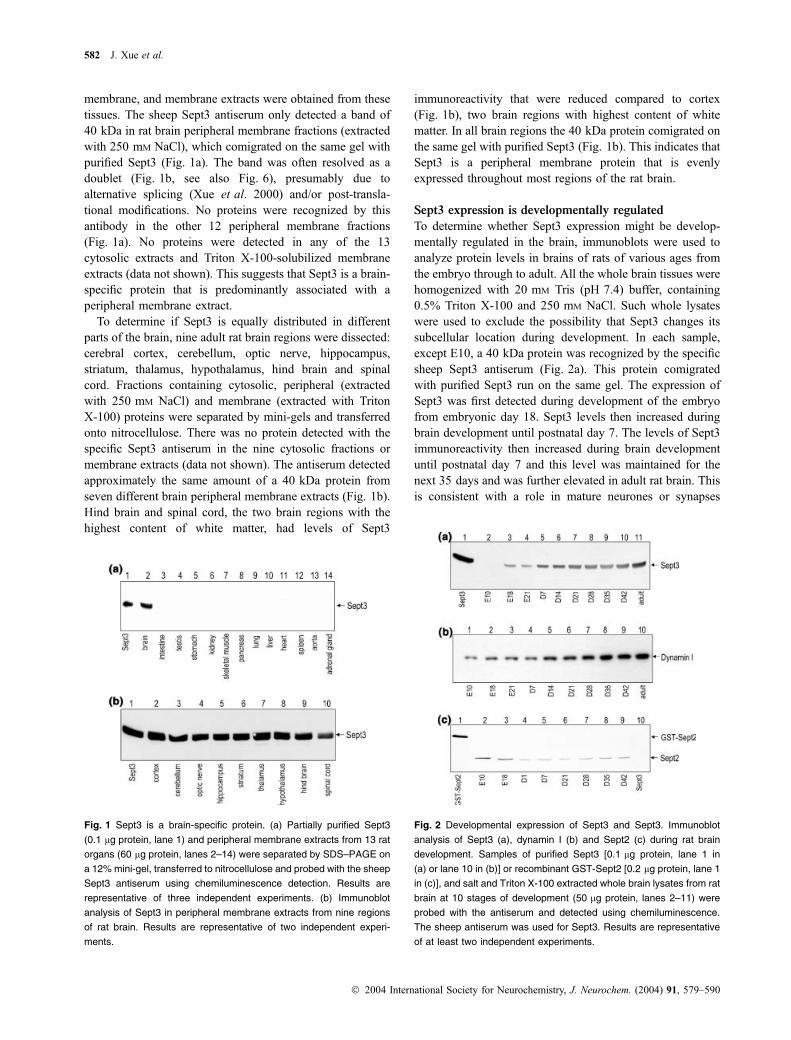
membrane, and membrane extracts were obtained from thesetissues. The sheep Sept3 antiserum only detected a band of40 kDa in rat brain peripheral membrane fractions (extractedwith 250 mM NaCl), which comigrated on the same gel withpurified Sept3 (Fig. 1a). The band was often resolved as adoublet (Fig. 1b, see also Fig. 6), presumably due toalternative splicing (Xue et al. 2000) and/or post-transla-tional modifications. No proteins were recognized by thisantibody in the other 12 peripheral membrane fractions(Fig. 1a). No proteins were detected in any of the 13cytosolic extracts and Triton X-100-solubilized membraneextracts (data not shown). This suggests that Sept3 is a brain-specific protein that is predominantly associated with aperipheral membrane extract.
To determine if Sept3 is equally distributed in differentparts of the brain, nine adult rat brain regions were dissected:cerebral cortex, cerebellum, optic nerve, hippocampus,striatum, thalamus, hypothalamus, hind brain and spinalcord. Fractions containing cytosolic, peripheral (extractedwith 250 mM NaCl) and membrane (extracted with TritonX-100) proteins were separated by mini-gels and transferredonto nitrocellulose. There was no protein detected with thespecific Sept3 antiserum in the nine cytosolic fractions ormembrane extracts (data not shown). The antiserum detectedapproximately the same amount of a 40 kDa protein fromseven different brain peripheral membrane extracts (Fig. 1b).Hind brain and spinal cord, the two brain regions with thehighest content of white matter, had levels of Sept3
immunoreactivity that were reduced compared to cortex(Fig. 1b), two brain regions with highest content of whitematter. In all brain regions the 40 kDa protein comigrated onthe same gel with purified Sept3 (Fig. 1b). This indicates thatSept3 is a peripheral membrane protein that is evenlyexpressed throughout most regions of the rat brain.
Sept3 expression is developmentally regulated
To determine whether Sept3 expression might be develop-mentally regulated in the brain, immunoblots were used toanalyze protein levels in brains of rats of various ages fromthe embryo through to adult. All the whole brain tissues werehomogenized with 20 mM Tris (pH 7.4) buffer, containing0.5% Triton X-100 and 250 mM NaCl. Such whole lysateswere used to exclude the possibility that Sept3 changes itssubcellular location during development. In each sample,except E10, a 40 kDa protein was recognized by the specificsheep Sept3 antiserum (Fig. 2a). This protein comigratedwith purified Sept3 run on the same gel. The expression ofSept3 was first detected during development of the embryofrom embryonic day 18. Sept3 levels then increased duringbrain development until postnatal day 7. The levels of Sept3immunoreactivity then increased during brain developmentuntil postnatal day 7 and this level was maintained for thenext 35 days and was further elevated in adult rat brain. Thisis consistent with a role in mature neurones or synapses
Fig. 1 Sept3 is a brain-specific protein. (a) Partially purified Sept3
(0.1 lg protein, lane 1) and peripheral membrane extracts from 13 rat
organs (60 lg protein, lanes 2–14) were separated by SDS–PAGE on
a 12% mini-gel, transferred to nitrocellulose and probed with the sheep
Sept3 antiserum using chemiluminescence detection. Results are
representative of three independent experiments. (b) Immunoblot
analysis of Sept3 in peripheral membrane extracts from nine regions
of rat brain. Results are representative of two independent experi-
ments.
Fig. 2 Developmental expression of Sept3 and Sept3. Immunoblot
analysis of Sept3 (a), dynamin I (b) and Sept2 (c) during rat brain
development. Samples of purified Sept3 [0.1 lg protein, lane 1 in
(a) or lane 10 in (b)] or recombinant GST-Sept2 [0.2 lg protein, lane 1
in (c)], and salt and Triton X-100 extracted whole brain lysates from rat
brain at 10 stages of development (50 lg protein, lanes 2–11) were
probed with the antiserum and detected using chemiluminescence.
The sheep antiserum was used for Sept3. Results are representative
of at least two independent experiments.
582 J. Xue et al.
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590
(Rostas et al. 1991). It also suggests that a single splicevariant of Sept3 predominates in the brain. For comparison,we investigated the expression of dynamin I, a presynapticGTPase required for endocytosis of small synaptic vesicles inthe nerve terminal and whose expression correlates withsynapse formation (Powell and Robinson 1995). The mem-brane was re-probed with anti-dynamin I antibodies. Dyn-amin I was detected in early E10 (Fig. 2b, lane 1) and itsexpression steadily increased with brain development(Fig. 2b, lanes 1–10).
Northern blot analysis and in situ hybridization previouslydemonstrated that the mRNA for the ubiquitously expressedSept2 (Nedd5) decreased during differentiation of adult brain(Kinoshita et al. 1997). As a second control, the expressionof Sept2 in rat brain was detected with specific anti-Sept2antibodies which did not cross-react with the purified Sept3from brain (Fig. 2c). Expression of Sept2 decreased duringdevelopment from the embryo to postnatal day 1, thenremained relatively stable throughout adulthood (Fig. 2c),consistent with previous findings (Kinoshita et al. 1997;Peng et al. 2002). The distinct expression pattern of septinsin brain suggests that they have different functions duringdevelopment.
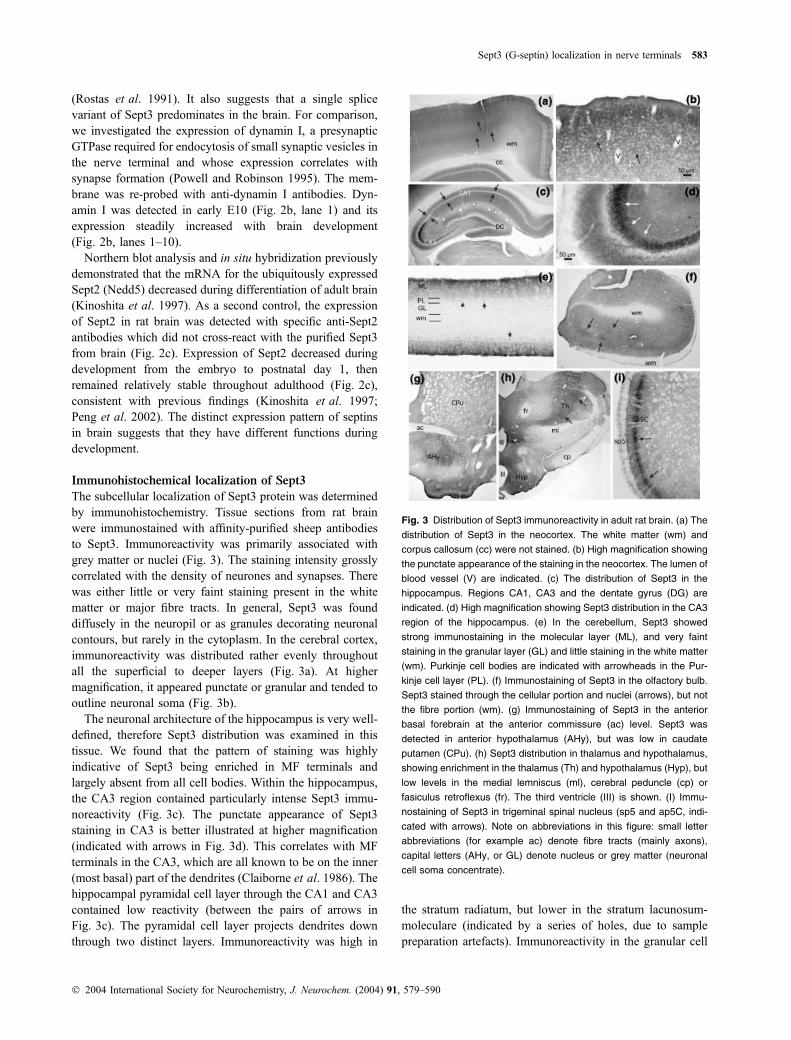
Immunohistochemical localization of Sept3
The subcellular localization of Sept3 protein was determinedby immunohistochemistry. Tissue sections from rat brainwere immunostained with affinity-purified sheep antibodiesto Sept3. Immunoreactivity was primarily associated withgrey matter or nuclei (Fig. 3). The staining intensity grosslycorrelated with the density of neurones and synapses. Therewas either little or very faint staining present in the whitematter or major fibre tracts. In general, Sept3 was founddiffusely in the neuropil or as granules decorating neuronalcontours, but rarely in the cytoplasm. In the cerebral cortex,immunoreactivity was distributed rather evenly throughoutall the superficial to deeper layers (Fig. 3a). At highermagnification, it appeared punctate or granular and tended tooutline neuronal soma (Fig. 3b).
The neuronal architecture of the hippocampus is very well-defined, therefore Sept3 distribution was examined in thistissue. We found that the pattern of staining was highlyindicative of Sept3 being enriched in MF terminals andlargely absent from all cell bodies. Within the hippocampus,the CA3 region contained particularly intense Sept3 immu-noreactivity (Fig. 3c). The punctate appearance of Sept3staining in CA3 is better illustrated at higher magnification(indicated with arrows in Fig. 3d). This correlates with MFterminals in the CA3, which are all known to be on the inner(most basal) part of the dendrites (Claiborne et al. 1986). Thehippocampal pyramidal cell layer through the CA1 and CA3contained low reactivity (between the pairs of arrows inFig. 3c). The pyramidal cell layer projects dendrites downthrough two distinct layers. Immunoreactivity was high in
the stratum radiatum, but lower in the stratum lacunosum-moleculare (indicated by a series of holes, due to samplepreparation artefacts). Immunoreactivity in the granular cell
Fig. 3 Distribution of Sept3 immunoreactivity in adult rat brain. (a) The
distribution of Sept3 in the neocortex. The white matter (wm) and
corpus callosum (cc) were not stained. (b) High magnification showing
the punctate appearance of the staining in the neocortex. The lumen of
blood vessel (V) are indicated. (c) The distribution of Sept3 in the
hippocampus. Regions CA1, CA3 and the dentate gyrus (DG) are
indicated. (d) High magnification showing Sept3 distribution in the CA3
region of the hippocampus. (e) In the cerebellum, Sept3 showed
strong immunostaining in the molecular layer (ML), and very faint
staining in the granular layer (GL) and little staining in the white matter
(wm). Purkinje cell bodies are indicated with arrowheads in the Pur-
kinje cell layer (PL). (f) Immunostaining of Sept3 in the olfactory bulb.
Sept3 stained through the cellular portion and nuclei (arrows), but not
the fibre portion (wm). (g) Immunostaining of Sept3 in the anterior
basal forebrain at the anterior commissure (ac) level. Sept3 was
detected in anterior hypothalamus (AHy), but was low in caudate
putamen (CPu). (h) Sept3 distribution in thalamus and hypothalamus,
showing enrichment in the thalamus (Th) and hypothalamus (Hyp), but
low levels in the medial lemniscus (ml), cerebral peduncle (cp) or
fasiculus retroflexus (fr). The third ventricle (III) is shown. (I) Immu-
nostaining of Sept3 in trigeminal spinal nucleus (sp5 and ap5C, indi-
cated with arrows). Note on abbreviations in this figure: small letter
abbreviations (for example ac) denote fibre tracts (mainly axons),
capital letters (AHy, or GL) denote nucleus or grey matter (neuronal
cell soma concentrate).
Sept3 (G-septin) localization in nerve terminals 583
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590

layer of the dentate gyrus (labelled as DG) was also very low,but there was strong staining in the underlying plexiformlayer (the hilus), which was also consistent with terminalsfrom the MF axons. The MF axons each give rise to aboutseven thinner collaterals which form synapses with thedendrites of neurones within the hilus. Each collateral formsabout 160 small varicosities (0.5–2.0 lm) and one giantvaricosity (3–5 lm) that resemble MF terminals in the CA3region (Scharfman et al. 1990).
The cerebellum was also selected for detailed analysisbecause it also has a well-defined neuronal architecture.There was a prominent immunostaining for Sept3 observedin the molecular layer (ML), and faint staining in the Purkinjecell layer (PL, arrow heads in Fig. 3e), but very little stainingin the granular layer (GL) and the white matter (WM) in thecerebellum (Fig. 3e). The granular layer is the site ofparticularly large clusters of up to 50 synapses calledglomeruli that stain well for dynamin (Powell and Robinson1995), but poorly for Sept3. In the olfactory bulb, Sept3 wasintensely stained through the cellular portion, but not in thefibre portion of the WM (wm in Fig. 3f). Prominent Sept3immunoreactivity was also present in anterior hypothalamus(AHy in Fig. 3g), thalamus and hypothalamus (Th and Hypin Fig. 3h). There was reduced staining in the medialleminiscus, cerebral peduncle and fasiculus retroflexus of thethalamic region (Fig. 3h), regions that are rich in axonbundles and therefore have reduced synapse density. In thebrainstem, moderate Sept3 immunostaining was present inmost nuclei. Intense staining was seen in sensory nucleiincluding the trigeminal spinal nucleus (arrows at Sp5C inFig. 3i). Together, these results strongly suggest that Sept3 iswidespread in neurones in the brain and spinal cord. Stainingintensity was generally relatively low in neuronal cell bodiesand staining intensity was much higher in regions with highsynapse density.
Sept3 is enriched in hippocampal MF nerve terminals
The strongest Sept3 immunohistochemical staining in brainwas the hippocampal CA3 region. This pattern was remark-ably similar to that previously reported for Timm’s stain,which specifically stains the zinc which is concentrated inglutamatergic synaptic vesicles of the MF terminals in thehippocampus (Claiborne et al. 1989; Henze et al. 2000).MFs are the axons of the dentate granule cells and theirterminals form multiple giant synapses with hippocampalCA3 pyramidal neurones and they play a critical role in thehippocampal neural network (Neuman et al. 1988; Henzeet al. 1997; Henze et al. 2000). To determine whether Sept3is specifically enriched in these terminals, MF synaptosomes(fractions MF3 and MF4) were isolated from hippocampi.The MF terminals are enriched in fraction MF4 (Taupin et al.1994). Cytosolic and peripheral membrane (250 mM NaCl-extracted) protein extracts were separated. Equal amount ofproteins from whole brain synaptosomes (BS), MF synapto-
somes (MF3 and MF4), whole hippocampus synaptosomes(HS), and from cerebral cortex synaptosomes (CS) wereapplied to nitrocellulose membrane with a slot blot. Thisapproach was used to mimic the immunohistochemistryapproach, and thus sample denaturation in SDS was avoided.Sheep antibodies to Sept3 were then used to probe themembrane and densitometry of the films provided a quan-titative analysis of the amount of Sept3 immunoreactivity inthe synaptosomes from each brain region. Sept3 was detectedin synaptosomes from each brain region, but surprisingly, itwas not elevated in the hippocampal or cortical synapto-somes relative to synaptosomes from whole brain. These tworegions were expected to contain a higher synapse densitythan whole brain. Importantly, Sept3 immunoreactivity wasenriched more than threefold in the MF terminals in MF4(Fig. 4). This supports the immunohistochemistry and indi-cates that the Sept3 is specifically enriched within MF nerveterminals.
Sept3 is enriched in forebrain brain nerve terminals
We previously demonstrated that Sept3 is phosphorylated byPKG in vitro and is a phosphoprotein in nerve terminals (Xueet al. 2000). To determine whether Sept3 is present insidenerve terminals and whether it might be enriched therein,immunoblots were used on soluble and peripheral membraneproteins from purified rat brain synaptosomes and comparedwith whole forebrain (Xue et al. 2000). In the case ofsynaptosomes, the soluble and peripheral membrane frac-tions primarily originate from within sealed synaptosomesand their proteins thus represent intraterminal proteins. Priorto synaptosomal lysis they were washed with 350 mM NaClto remove extra-terminal contaminating Sept3, and after lysis
Fig. 4 Specific enrichment of Sept3 in hippocampal MF nerve termi-
nals. Slot blot analysis of Sept3 in whole triton lysates of synapto-
somes isolated from different brain regions. Equal amounts of protein
(60 lg) from different synaptosomal extracts were spotted onto nitro-
cellulose with a slot blot device: whole brain synaptosomes (BS), MF
synaptosomes MF3 and MF4, synaptosomes from whole hippocam-
pus (HS), and from cerebral cortex (CS). The membrane probed with
sheep anti-Sept3 antibodies and the film was quantitated using den-
sitometry (n ¼ 3, error bars are SEM).
584 J. Xue et al.
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590
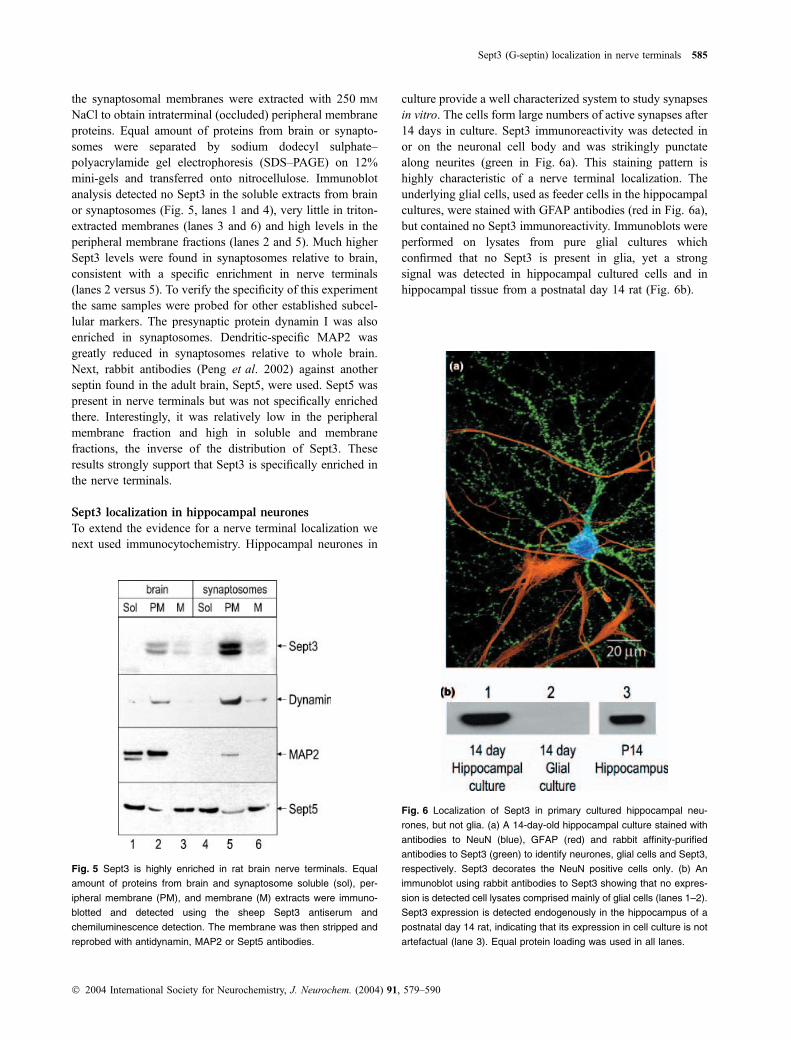
the synaptosomal membranes were extracted with 250 mM
NaCl to obtain intraterminal (occluded) peripheral membraneproteins. Equal amount of proteins from brain or synapto-somes were separated by sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS–PAGE) on 12%mini-gels and transferred onto nitrocellulose. Immunoblotanalysis detected no Sept3 in the soluble extracts from brainor synaptosomes (Fig. 5, lanes 1 and 4), very little in triton-extracted membranes (lanes 3 and 6) and high levels in theperipheral membrane fractions (lanes 2 and 5). Much higherSept3 levels were found in synaptosomes relative to brain,consistent with a specific enrichment in nerve terminals(lanes 2 versus 5). To verify the specificity of this experimentthe same samples were probed for other established subcel-lular markers. The presynaptic protein dynamin I was alsoenriched in synaptosomes. Dendritic-specific MAP2 wasgreatly reduced in synaptosomes relative to whole brain.Next, rabbit antibodies (Peng et al. 2002) against anotherseptin found in the adult brain, Sept5, were used. Sept5 waspresent in nerve terminals but was not specifically enrichedthere. Interestingly, it was relatively low in the peripheralmembrane fraction and high in soluble and membranefractions, the inverse of the distribution of Sept3. Theseresults strongly support that Sept3 is specifically enriched inthe nerve terminals.
Sept3 localization in hippocampal neurones
To extend the evidence for a nerve terminal localization wenext used immunocytochemistry. Hippocampal neurones in
culture provide a well characterized system to study synapsesin vitro. The cells form large numbers of active synapses after14 days in culture. Sept3 immunoreactivity was detected inor on the neuronal cell body and was strikingly punctatealong neurites (green in Fig. 6a). This staining pattern ishighly characteristic of a nerve terminal localization. Theunderlying glial cells, used as feeder cells in the hippocampalcultures, were stained with GFAP antibodies (red in Fig. 6a),but contained no Sept3 immunoreactivity. Immunoblots wereperformed on lysates from pure glial cultures whichconfirmed that no Sept3 is present in glia, yet a strongsignal was detected in hippocampal cultured cells and inhippocampal tissue from a postnatal day 14 rat (Fig. 6b).
Fig. 5 Sept3 is highly enriched in rat brain nerve terminals. Equal
amount of proteins from brain and synaptosome soluble (sol), per-
ipheral membrane (PM), and membrane (M) extracts were immuno-
blotted and detected using the sheep Sept3 antiserum and
chemiluminescence detection. The membrane was then stripped and
reprobed with antidynamin, MAP2 or Sept5 antibodies.
Fig. 6 Localization of Sept3 in primary cultured hippocampal neu-
rones, but not glia. (a) A 14-day-old hippocampal culture stained with
antibodies to NeuN (blue), GFAP (red) and rabbit affinity-purified
antibodies to Sept3 (green) to identify neurones, glial cells and Sept3,
respectively. Sept3 decorates the NeuN positive cells only. (b) An
immunoblot using rabbit antibodies to Sept3 showing that no expres-
sion is detected cell lysates comprised mainly of glial cells (lanes 1–2).
Sept3 expression is detected endogenously in the hippocampus of a
postnatal day 14 rat, indicating that its expression in cell culture is not
artefactual (lane 3). Equal protein loading was used in all lanes.
Sept3 (G-septin) localization in nerve terminals 585
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590

The punctate staining of neurites with Sept3 antibodieswas detected as early as 3 days in culture, but wasprogressively elevated at days 7 (data not shown) and 14(Fig. 7a,d,f). This correlates with synaptogenesis in thesecultures (Schikorski and Stevens 1997) and with thedevelopmental expression pattern in whole brain (Fig. 2).Synaptophysin is a synaptic vesicle protein which localizesto the surface of synaptic vesicles, as part of a complex that isrequired for the biogenesis of synaptic vesicles and it iswidely used as a marker that is enriched in nerve terminals(Thiele et al. 2000). When the cells were double-stained forSept3 and synaptophysin there was extensive overlap inpunctate regions (Fig. 7a–e). Dynamin I is also highlyenriched in nerve terminals (e.g. Fig. 5) and localizes toactive synaptic puncta in cultured neuronal cells (Tan et al.2003). Double-staining for Sept3 and dynamin I showedalmost complete overlap at numerous puncta along theneurites (Fig. 7f–h). In other studies using styryl dyes suchpuncta are demonstrated sites for exocytosis (Ahmari andSmith 2002; Tan et al. 2003).
At the level of light microscopy, it is difficult to determinewhether a protein is on the presynaptic or postsynaptic sideof the synapse. Therefore immunogold electron microscopywas used to provide further evidence that Sept3 was presentspecifically within presynaptic terminals of hippocampalneurones (Fig. 8). Concentrations of gold particles weredetected primarily within presynaptic terminals and onlyrarely within the postsynapse. Within the terminal, stainingwas primarily associated with the outside of synaptic vesiclesrather than their lumen, suggesting Sept3 associates with oradjacent to synaptic vesicles. However, tissue fixation,embedding and staining methods for immunogold electronmicroscopy do not exclude additional localizations inaccess-ible to the antibodies under these conditions, thus there areinherent difficulties in comparing immunocytochemistry andimmunogold electron microscopy due to such differences.
Discussion
In this report we have demonstrated that Sept3 is a brain-specific protein. Within the brain it is restricted to neuronesand within neurones there is a specific enrichment inpresynaptic nerve terminals. Its expression is developmen-tally regulated in a manner opposite to that of another septin,Sept2, such that expression correlates with synapse formationand maturation (Rostas et al. 1991). These results suggestthat Sept3 differs from most other septins in that it is unlikelyto play a role in cytokinesis in mitotic neuronal precursorcells. Rather, the results suggest it has a role in the functionof mature neurones, such as synapse maturation or synapticvesicle recycling.
Location in neurones
All mammalian septins examined to date are expressed in thebrain (Kinoshita et al. 2000). Sept2 is expressed in neuronalprecursor cells and is down-regulated in adult brain (Kumaret al. 1992). Septins 1–9 (except Sept3) coimmunoprecipi-
Fig. 7 Co-localization of Sept3 with synaptophysin or dynamin in
primary cultured hippocampal neurones. (a–e) A hippocampal neur-
one cultured for 14 days stained with rabbit antibodies against Sept3
(green) and synaptophysin (red). Sept3 shows a punctate distribution
in processes and a strong colocalization with synaptophysin. Sept3 is
expressed in punctate regions located extrasynaptically along the
edges of processes (arrows). The boxed area in (a,b) is zoomed in
(d,e) to show strong colocalization at synaptic puncta. (f–h) A hippo-
campal neurone cultured for 14 days stained with sheep antibodies
against Sept3 (green) and dynamin I (red). Sept3 staining shows
strong enrichment in synaptic puncta and is present at low levels
throughout the body of the neurite. Dynamin I staining is essentially
identical to Sept3 in the synaptic puncta.
Fig. 8 Presynaptic localization of Sept3 in cultured hippocampal
neurones. Two immunogold electron micrographs of 14-day cultured
hippocampal neurones. Immunolabelling with purified rabbit antibodies
to Sept3 (the black dots represent gold-conjugated antibodies) was
found in the presynaptic terminals (PRE, labelled with arrows), adja-
cent to synaptic vesicles.
586 J. Xue et al.
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590

tate in different groups from brain and form filaments (Hsuet al. 1998; Kinoshita et al. 2002). However, most septinsare also abundant in other tissues. In contrast, Sept5 wasthought to be largely a brain-specific protein with a specificfunction in the regulation of exocytosis (Beites et al. 1999;Kinoshita et al. 2000), however, it is also present inmegakaryocytes, particularly platelets (Yagi et al. 1998;Dent et al. 2002). Our results indicate that Sept3 is the onlyseptin to date specific to brain and is not found in platelets orcell lines derived from non-neuronal origin. Previous studiesshowed that Sept3 mRNA was detected only in brain (Xueet al. 2000). Immunoblot analysis confirmed that Sept3protein is only expressed in brain. Immunohistochemistryrevealed a wide-spread distribution in many brain regions.Highest levels occurred where neurones and nerve terminalsare concentrated, but not all neurones contained Sept3immunoreactivity. This correlates with the brain distributionof Sept5, but not Sept4 or Sept7 (Kinoshita et al. 2000).Cultured hippocampal neurones were also strongly immu-noreactive for Sept3. We found no evidence for Sept3expression in glia. This was also apparent from the wholebrain immunohistochemistry. For example, Sept3 levels werereduced in hind brain and spinal cord, regions containinghighest levels of white matter, but rich in glia. It was alsoconfirmed using primary cultured glial cells which lackeddetectable Sept3 by immunocytochemistry or by immuno-blot. We cannot exclude that some subpopulations of glialcells might express Sept3. Other septins have been localizedto neurones (Sept4–7), while Sept2, Sept4 and Sept7 areadditionally present in glia (Kinoshita et al. 2000; Ihara et al.2003; Kinoshita et al. 2004).
Location in nerve terminals
A variety of our data provides evidence that Sept3 isremarkably enriched in nerve terminals. First, areas of thebrain known to be highly enriched in nerve terminals wereparticularly immunoreactive for Sept3. In particular itsdistribution within the cortex, hippocampus, cerebellum,thalamus, hypothalamus and brain stem sensory nucleicorrelated with high synapse density in these regions (Powellet al. 2000). This conclusion was supported by culturedhippocampal neurones where Sept3 expression was greatlyincreased after 14 days in culture and was highly punctatealong cell processes. This is consistent with a specificdistribution in the synapses onto the hippocampal neuronesin culture (Ahmari and Smith 2002). Second, the specificenrichment of Sept3 in nerve terminals was revealed byimmunoblot of occluded protein from synaptosomes. Sept3was associated with a peripheral membrane extract, consis-tent with filament formation or superficial membrane attach-ment. Third, Sept3 localization determined by confocalmicroscopy revealed strong colocalization with the presy-naptic marker synaptophysin in brain (data not shown) and inprimary hippocampal neurones (Fig. 7). The results are
similar to those reported for Sept5 in the same cells, whichpartly colocalizes with another synaptic vesicle markerVAMP2 (Peng et al. 2002) and interacts with synaptophysinand syntaxin (Honer et al. 1993; Caltagarone et al. 1998;Beites et al. 1999). As synaptophysin immunoreactivity is animperfect marker for active synapses (Ahmari and Smith2002), we used dynamin I localization as an additional index.Sept3 showed specific enrichment in synaptic puncta andalmost 100% colocalization with dynamin I. Finally, anintraterminal location was confirmed by immunogold elec-tron microscopy, where Sept3 was found adjacent to synapticvesicles. Sept5 is also present in synaptosomes wherein itforms a complex with Sept2 and Sept7 (Peng et al. 2002). Astrong association of Sept3 with Sept5 seems unlikely as theyhad opposite subcellular distribution in synaptosomes. Sept5and 6 have also been specifically localized to subpopulationsof nerve terminals by immunogold electron microscopy(Kinoshita et al. 2000). Sept5 was localized mainly aroundthe presynaptic vesicles, but not along the presynapticmembrane, and appeared to be restricted to inhibitorypresynaptic terminals and associated with GABAergic ves-icles. Similarly, Sept7 was found in a distinct, overlappingset of nerve terminals as well as glia. Thus we conclude thatSept3 is a presynaptic protein that is enriched in nerveterminals or terminals en passant. However, it should benoted that Sept3 might not be present at high levels in allpopulations of synapses. For example, its expression waslower in the synaptic glomeruli of the granular layer of thecerebellum, as well as subregions of the hippocampusbetween the CA1 and dentate gyrus.
Developmental regulation
Sept3 expression in rat brain was first detectable at a latestage of embryonic development (embryonic day 18), but notin early embryos. Expression levels then increased untilpostnatal day 7, then only slightly increased for the next35 days, suggesting that Sept3 expression is developmentallyregulated. This expression correlates well with previousmRNA studies suggesting a role for Sept3 in neuronaldevelopment or differentiation (Xiong et al. 1999; Methneret al. 2001) and correlates with expression of Sept5 in thebrain (Peng et al. 2002). Similar expression profiles werefound for another brain-specific protein, dynamin I, which isa 100 kDa GTPase involved in the endocytic cycle ofsynaptic vesicle membranes (Faire et al. 1992; Robinsonet al. 1993; De Camilli et al. 1995; De Camilli and Takei1996). The dynamin 3.6 kb mRNA transcript is preferen-tially expressed in the rat brain after postnatal day 7, with amaximal peak of mRNA and protein expression betweenpostnatal day 15 and 23 in the cerebral cortex and maximal atpostnatal day 35 in cerebellum (Nakata et al. 1991; Faireet al. 1992). This supports the known role of dynamin I inendocytosis in mature nerve terminals (De Camilli and Takei1996).
Sept3 (G-septin) localization in nerve terminals 587
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590

Association with the peripheral membrane
Sept3 was shown to be associated with peripheral membraneextracts. All known mammalian septins are thought to bepredominantly associated with membranes, although theylack transmembrane domains (reviewed in Field and Kellogg1999; Trimble 1999). Sept4 attaches to plasma membranes ina nucleotide-dependent manner by specifically binding to thephospholipids PtdIns(4,5)P2 and PtdIns(3,4,5)P3, and thebinding site is at a poly basic motif in Sept4135)141 (Zhanget al. 1999). Most mammalian septins have similar motifs,except Sept6 and Sept8. These basic amino acids are alsopresent in Sept352)58. We have recently found that Sept3 is alipid-binding protein which specifically binds phosphatidy-linositol 3-phosphate [PtdIns(3)P] and PtdIns(4)P, and thatbinding is blocked by prior phosphorylation with PKG (JXand PJR unpublished). Since the phosphorylation site forSept3 by PKG-I is near the first GTP binding motif(Sept368)76, JX and PJR unpublished), phosphorylation ofSept3 by PKG may regulate its GTP binding and thus alterthe localization of Sept3 within nerve terminals. A similarresult was also obtained with dynamin whose associationwith lipid was blocked by phosphorylation by cyclin-dependent protein kinase 5 (Tan et al. 2003).
Enrichment in MF terminals
A number of septins are present in the hippocampal CA3region (Kinoshita et al. 2000). Our study shows that Sept3immunoreactivity has a striking enrichment there. We showedit to be most highly enriched in MF nerve terminals.Hippocampal MF synapses of the CA3 contain an abundanceof Zn2+ sequestered in their glutamatergic synaptic vesiclesthrough synaptic vesicle zinc transporters (Li et al. 2001). TheMF synapse of the CA3 region in hippocampus exhibits short-term facilitation and plasticity (Salin et al. 1996), LTP (Staubliet al. 1990; Langdon et al. 1995), and LTD (Yokoi et al.1996). The induction of MF LTP is dependent on cAMP-dependent protein kinase A (PKA) (Lopez-Garcia et al. 1996;Yeckel et al. 1999) and involves phosphorylation of RIM1al-pha (Lonart et al. 2003). Sept3 is a poor PKA substrate in vitroand in not likely to be involved. However, there are interestinglinks between Sept3 and the CA1. LTP in hippocampal CA1region is suppressed by PKG inhibitors (Son et al. 1998) andpresynaptic injection of PKG can facilitate the induction ofLTP in cultured hippocampal CA1 neurones (Arancio et al.2001). LTP in the stratum radiatum of CA1, where Sept3immunoreactivity was high, is known to have a stronginvolvement of the nitric oxide pathway (Son et al. 1996).Chemically induced LTD is also mediated by PKG in the CA1region (Santschi et al. 1999), where elevation of cGMP levelsalso reduces exocytosis (Stanton et al. 2001). We showedpreviously that Sept3 is phosphorylated in intact synaptosomesfrom rat brain and is an in vitro PKG substrate (Xue et al.2000). Sept3 increases in phosphorylation in response tostimulation with cGMP analogues (JX and PJR unpublished
data). These results support the possibility that the phosphory-lation of Sept3 in neurones may occur in vivo and may be offunctional significance. Together this forms a potentiallyinteresting link between Sept3, cGMP and plasticity in thehippocampus and future studies should evaluate if Sept3mightbe involved in some of these plasticity changes.
In summary, we define Sept3 as a new and majorcomponent of the presynaptic nerve terminal in the mam-malian brain. Our data demonstrate that Sept3 is greatlyenriched in nerve terminals. Sept3 has interesting propertiesof GTP-regulated filament assembly, lipid association and isregulated by nitric oxide-mediated protein phosphorylation.Like other nerve terminal proteins, such as dynamin andsynapsins, Sept3 may have functions involved in the synapticvesicle life cycle, however, this proposal must be directlyconfirmed with functional studies.
Acknowledgements
Supported by grants from the Australian National Health and
Medical Research Council (NHMRC to PJR). Peter Rowe is
thanked for critical reading of the manuscript. We would like to
thank Robert Temkin in the Electron Microscopy Facility at the
Hospital for Sick Children for his expertise.
References
Ahmari S. E. and Smith S. J. (2002) Knowing a nascent synapse whenyou see it. Neuron 34, 333–336.
Arancio O., Antonova I., Gambaryan S., Lohmann S. M., Wood J. S.,Lawrence D. S. and Hawkins R. D. (2001) Presynaptic role ofcGMP-dependent protein kinase during long-lasting potentiation.J. Neurosci. 21, 143–149.
Banker G. and Goslin K. (1991) Culturing Nerve Cells, pp. 251–278.The MIT Press, Cambridge, MA.
Beites C. L., Xie H., Bowser R. and Trimble W. S. (1999) The septinCDCrel-1 binds syntaxin and inhibits exocytosis. Nat. Neurosci. 2,434–439.
Brewer G. J., Torricelli J. R., Evege E. K. and Price P. J. (1993) Opti-mized survival of hippocampal neurons in B27-supplementedneurobasal, a new serum-free medium combination. J. Neurosci.Res. 35, 567–576.
Caltagarone J., Rhodes J., Honer W. G. and Bowser R. (1998) Local-ization of a novel septin protein, hCDCrel-1, in neurons of humanbrain. Neuroreport 9, 2907–2912.
Claiborne B. J., Amaral D. G. and Cowan W. M. (1986) A light andelectron microscopic analysis of the mossy fibers of the rat dentategyrus. J. Comp. Neurol. 246, 435–458.
Claiborne B. J., Rea M. A. and Terrian D. M. (1989) Detection of zinc inisolated nerve terminals using a modified Timm’s sulfide-silvermethod. J. Neurosci. Methods 30, 17–22.
De Camilli P. and Takei K. (1996) Molecular mechanisms in synapticvesicle endocytosis and recycling. Neuron 16, 481–486.
De Camilli P., Takei K. and McPherson P. S. (1995) The function ofdynamin in endocytosis. Curr. Opin. Neurobiol. 5, 559–565.
Dent J., Kato K., Peng X.-R., Martinez C., Cattaneo M., Poujol C.,Nurden P., Nurden A., Trimble W. S. and Ware J. (2002) A pro-totypic platelet septin and its participation in secretion. Proc. NatlAcad. Sci. USA 99, 3064–3069.
588 J. Xue et al.
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590

Dong Z., Ferger B., Paterna J. C., Vogel D., Furler S., Osinde M., FeldonJ. and Bueler H. (2003) Dopamine-dependent neurodegeneration inrats induced by viral vector-mediated overexpression of the parkintarget protein, CDCrel-1. Proc. Natl Acad. Sci. USA 100, 12 438–12 443.
Dunkley P. R., Jarvie P. E., Heath J. W., Kidd G. J. and Rostas J. A.(1986) A rapid method for isolation of synaptosomes on Percollgradients. Brain Res. 372, 115–129.
Faire K., Trent F., Tepper J. M. and Bonder E. M. (1992) Analysis ofdynamin isoforms in mammalian brain: dynamin-1 expression isspatially and temporally regulated during postnatal development.Proc. Natl Acad. Sci. USA 89, 8376–8380.
Field C. M. and Kellogg D. (1999) Septins: cytoskeletal polymers orsignalling GTPases? Trends Cell Biol. 9, 387–394.
Field C. M., al-Awar O., Rosenblatt J., Wong M. L., Alberts B. andMitchison T. J. (1996) A purified Drosophila septin complex formsfilaments and exhibits GTPase activity. J. Cell Biol. 133, 605–616.
Fung E. T. and Scheller R. H. (1999) Identification of a novel alternat-ively spliced septin. FEBS Lett. 451, 203–208.
Henze D. A., Card J. P., Barrionuevo G. and Ben Ari Y. (1997) Largeamplitude miniature excitatory postsynaptic currents in hippo-campal CA3 pyramidal neurons are of mossy fiber origin. J.Neurophysiol. 77, 1075–1086.
Henze D. A., Urban N. N. and Barrionuevo G. (2000) The multifarioushippocampal mossy fiber pathway: a review. Neurosci. 98, 407–427.
Honer W. G., Hu L. and Davies P. (1993) Human synaptic proteins witha heterogeneous distribution in cerebellum and visual cortex. BrainRes. 609, 9–20.
Hsu S. C., Hazuka C. D., Roth R., Foletti D. L., Heuser J. and SchellerR. H. (1998) Subunit composition, protein interactions, andstructures of the mammalian brain sec6/8 complex and septin fil-aments. Neuron 20, 1111–1122.
Ihara M., Tomimoto H., Kitayama H., Morioka Y., Akiguchi I., Shiba-saki H., Noda M. and Kinoshita M. (2003) Association of thecytoskeletal GTP-binding protein Sept4/H5 with cytoplasmicinclusions found in Parkinson’s disease and other synucleinopa-thies. J. Biol. Chem. 278, 24 095–24 102.
Kinoshita A., Kinoshita M., Akiyama H., Tomimoto H., Akiguchi I.,Kumar S., Noda M. and Kimura J. (1998) Identification of septinsin neurofibrillary tangles in Alzheimer’s disease. Am. J. Pathol.153, 1551–1560.
Kinoshita A., Noda M. and Kinoshita M. (2000) Differential localizationof septins in the mouse brain. J. Comp. Neurol. 428, 223–239.
Kinoshita M. (2003) Assembly of mammalian septins. J. Biochem. 134,491–496.
Kinoshita M., Kumar S., Mizoguchi A., Ide C., Kinoshita A., HaraguchiT., Hiraoka Y. and Noda M. (1997) Nedd5, a mammalian septin, isa novel cytoskeletal component interacting with actin-basedstructures. Genes Dev. 11, 1535–1547.
Kinoshita M., Field C. M., Coughlin M. L., Straight A. F. and MitchisonT. J. (2002) Self- and actin-templated assembly of mammalianseptins. Dev. Cell 3, 791–802.
Kinoshita N., Kimura K., Matsumoto N., Watanabe M., Fukaya M.and Ide C. (2004) Mammalian septin Sept2 modulates the activityof GLAST, a glutamate transporter in astrocytes. Genes Cells 9,1–14.
Kumar S., Tomooka Y. and Noda M. (1992) Identification of a set ofgenes with developmentally down-regulated expression in themouse brain. Biochem. Biophys. Res. Commun. 185, 1155–1161.
Langdon R. B., Johnson J. W. and Barrionuevo G. (1995) Posttetanicpotentiation and presynaptically induced long-term potentiation atthe mossy fiber synapse in rat hippocampus. J. Neurobiol. 26, 370–385.
Li Y., Hough C. J., Frederickson C. J. and Sarvey J. M. (2001) Inductionof mossy fiber?CA3 long-term potentiation requires translocationof synaptically released Zn2+. J. Neurosci. 21, 8015–8025.
Liauw J., Nguyen V., Huang J., George-Hyslop P. and Rozmahel R.(2002) Differential display analysis of presenilin 1-deficient mousebrains. Brain Res. Mol. Brain Res. 109, 56–62.
Lonart G., Schoch S., Kaeser P. S., Larkin C. J., Sudhof T. C. and LindenD. J. (2003) Phosphorylation of RIM1alpha by PKA triggerspresynaptic long-term potentiation at cerebellar parallel fiber syn-apses. Cell 115, 49–60.
Lopez-Garcia J. C., Arancio O., Kandel E. R. and Baranes D. (1996)A presynaptic locus for long-term potentiation of elementarysynaptic transmission at mossy fiber synapses in culture. Proc. NatlAcad. Sci. USA 93, 4712–4717.
Macara I. G., Baldarelli R., Field C. M. et al. (2002) Mammalian septinsnomenclature. Mol. Biol. Cell 13, 4111–4113.
Methner A., Leypoldt F., Joost P. and Lewerenz J. (2001) Human septin3 on chromosome 22q13.2 is upregulated by neuronal differenti-ation. Biochem. Biophys. Res. Commun. 283, 48–56.
Nakata T., Iwamoto A., Noda Y., Takemura R., Yoshikura H. andHirokawa N. (1991) Predominant and developmentally regulatedexpression of dynamin in neurons. Neuron 7, 461–469.
Neuman R. S., Ben Ari Y. and Cherubini E. (1988) Antagonism ofspontaneous and evoked bursts by 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX) in the CA3 region of the in vitro hippocampus.Brain Res. 474, 201–203.
Peng X.-R., Jia Z., Zhang Y., Ware J. and Trimble W. S. (2002) Theseptin CDCrel-1 is dispensable for normal development and neu-rotransmitter release. Mol. Cell Biol. 22, 378–387.
Powell K. A. and Robinson P. J. (1995) Dephosphin/dynamin is aneuronal phosphoprotein concentrated in nerve terminals: evidencefrom rat cerebellum. Neuroscience 64, 821–833.
Powell K. A., Valova V. A., Malladi C. S., Jensen O. N., Larsen M. R.and Robinson P. J. (2000) Phosphorylation of dynamin I on Ser-795 by protein kinase C blocks its association with phospholipids.J. Biol. Chem. 275, 11 610–11 617.
Robinson P. J., Sontag J.-M., Liu J. P., Fykse E. M., Slaughter C.,McMahon H. T. and Sudhof T. C. (1993) Dynamin GTPaseregulated by protein kinase C phosphorylation in nerve terminals.Nature 365, 163–166.
Rostas J. A., Kavanagh J. M., Dodd P. R., Heath J. W. and Powis D. A.(1991) Mechanisms of synaptic plasticity: changes in postsynapticdensities and glutamate receptors in chicken forebrain duringmaturation. Mol. Neurobiol. 5, 203–216.
Salin P. A., Scanziani M., Malenka R. C. and Nicoll R. A. (1996)Distinct short-term plasticity at two excitatory synapses in thehippocampus. Proc. Natl Acad. Sci. USA 93, 13 304–13 309.
Santschi L., Reyes-Harde M. and Stanton P. K. (1999) Chemicallyinduced, activity-independent LTD elicited by simultaneous activa-tion of PKG and inhibition of PKA. J. Neurophysiol. 82, 1577–1589.
Saraste M., Sibbald P. R. and Wittinghofer A. (1990) The P-loop – acommon motif in ATP- and GTP-binding proteins. Trends Bio-chem. Sci. 15, 430–434.
Scharfman H. E., Kunkel D. D. and Schwartzkroin P. A. (1990) Synapticconnections of dentate granule cells and hilar neurons: results ofpaired intracellular recordings and intracellular horseradish per-oxidase injections. Neuroscience 37, 693–707.
Schikorski T. and Stevens C. F. (1997) Quantitative ultrastructural ana-lysis of hippocampal excitatory synapses. J. Neurosci. 17, 5858–5867.
SonH., HawkinsR.D.,MartinK., KieblerM., Huang P. L., FishmanM.C.and Kandel E. R. (1996) Long-term potentiation is reduced in micethat are doubly mutant in endothelial and neuronal nitric oxidesynthase. Cell 87, 1015–1023.
Sept3 (G-septin) localization in nerve terminals 589
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590

Son H., Lu Y. F., Zhuo M., Arancio O., Kandel E. R. and Hawkins R. D.(1998) The specific role of cGMP in hippocampal LTP. Learn.Mem. 5, 231–245.
Stanton P. K., Heinemann U. and Muller W. (2001) FM1-43 imagingreveals cGMP-dependent long-term depression of presynaptictransmitter release. J. Neurosci. 21, RC167.
Staubli U., Larson J. and Lynch G. (1990) Mossy fiber potentiation andlong-term potentiation involve different expression mechanisms.Synapse 5, 333–335.
Surka M. C., Tsang C. W. and Trimble W. S. (2002) The mammalianseptin MSF localizes with microtubules and is required for com-pletion of cytokinesis. Mol. Biol. Cell 13, 3532–3545.
Tan T. C., Valova V. A., Malladi C. S. et al. (2003) Cdk5 is essential forsynaptic vesicle endocytosis. Nat. Cell Biol. 5, 701–710.
Taupin P., Zini S., Cesselin F., Ben Ari Y. and Roisin M. P. (1994)Subcellular fractionation on Percoll gradient of mossy fiber syna-ptosomes: morphological and biochemical characterization incontrol and degranulated rat hippocampus. J. Neurochem. 62,1586–1595.
Thiele C., Hannah M. J., Fahrenholz F. and Huttner W. B. (2000)Cholesterol binds to synaptophysin and is required for biogenesisof synaptic vesicles. Nat. Cell Biol. 2, 42–49.
Towbin H., Staehlin T. and Gordon J. (1979) Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: proce-dure and some applications. Proc. Natl Acad. Sci. USA 76, 4350–4354.
Trimble W. S. (1999) Septins: a highly conserved family of membrane-associated GTPases with functions in cell division and beyond.J. Membr. Biol. 169, 75–81.
Vega I. E. and Hsu S. C. (2003) The septin protein Nedd5 associates withboth the exocyst complex and microtubules and disruption of itsGTPase activity promotes aberrant neurite sprouting in PC12 cells.Neuroreport 14, 31–37.
Walikonis R. S., Jensen O. N., Mann M., Provance D. W. J., Mercer J. A.and Kennedy M. B. (2000) Identification of proteins in thepostsynaptic density fraction by mass spectrometry. J. Neurosci.20, 4069–4080.
Xie H., Surka M., Howard J. and Trimble W. S. (1999) Characterizationof the mammalian septin H5: distinct patterns of cytoskeletal andmembrane association from other septin proteins. Cell Motil.Cytoskeleton 43, 52–62.
Xiong J. W., Leahy A. and Stuhlmann H. (1999) Retroviral promoter-trap insertion into a novel mammalian septin gene expressed duringmouse neuronal development. Mech. Dev. 86, 183–191.
Xue J., Wang X., Malladi C. S., Kinoshita M., Milburn P. J., Lengyel I.,Rostas J. A. and Robinson P. J. (2000) Phosphorylation of a newbrain-specific septin, G-septin, by cyclic GMP-dependent proteinkinase. J. Biol. Chem. 275, 10 047–10 056.
Yagi M., Zieger B., Roth G. J. and Ware J. (1998) Structure andexpression of the human septin gene HCDCREL-1. Gene 212,229–236.
Yeckel M. F., Kapur A. and Johnston D. (1999) Multiple forms of LTP inhippocampal CA3 neurons use a common postsynaptic mechan-ism. Nat. Neurosci. 2, 625–633.
Yokoi M., Kobayashi K., Manabe T. et al. (1996) Impairment of hip-pocampal mossy fiber LTD in mice lacking mGluR2. Science 273,645–647.
Zhang J., Kong C., Xie H., McPherson P. S., Grinstein S. and Trimble W.S. (1999) Phosphatidylinositol polyphosphate binding to themammalian septin H5 is modulated by GTP. Curr. Biol. 9, 1458–1467.
Zhang Y., Gao J., Chung K. K., Huang H., Dawson V. L. and Dawson T.M. (2000) Parkin functions as an E2-dependent ubiquitin-proteinligase and promotes the degradation of the synaptic vesicle-asso-ciated protein, CDCrel-1. Proc. Natl Acad. Sci. USA 97, 13 354–13 359.
590 J. Xue et al.
� 2004 International Society for Neurochemistry, J. Neurochem. (2004) 91, 579–590