the presynaptic particle web
TRANSCRIPT

Neuron, Vol. 32, 63–77, October 11, 2001, Copyright 2001 by Cell Press
The Presynaptic Particle Web: Ultrastructure,Composition, Dissolution, and Reconstitution
havior of each synapse (Foster and Sherrington, 1897).Ultrastructural observations have since led to the cur-rent notion of a “characteristic” CNS synapse: pre- and
Greg R. Phillips,1 Jeffrey K. Huang,1 Yun Wang,5
Hidekazu Tanaka,6 Lawrence Shapiro,2
Wandong Zhang,1 Wei-Song Shan,1 Kirsten Arndt,1
postsynaptic membranes are rigorously parallel, in reg-Marcus Frank,1 Ronald E. Gordon,3
ister with each other, and separated by a cleft of uniformMary Ann Gawinowicz,4 Yingming Zhao,7
distance (�25 nm, Peters et al., 1991). In orthogonaland David R. Colman1,8
projection, electron dense “pegs” project into the syn-1The Corinne Goldsmith Dickinson Centeraptic cleft from thickenings of the pre-and postsynapticfor Multiple Sclerosismembranes (Ichimura and Hashimoto, 1988), sug-Department of Neurology and The Fishberggesting that the pre- and postsynaptic membrane link-Research Center for Neurobiologyage is mediated by molecules interacting within that2 Program in Structural Biologyspace. These features of the CNS synaptic junctionalDepartment of Physiology and Biophysicscomplex are indicative of a strongly adhesive structure,3 Department of Pathologyand this conclusion is confirmed by cell fractionationThe Mount Sinai School of Medicineexperiments. Once formed, the pre- and postsynapticOne Gustave L. Levy Placemembranes resist physical separation, such that evenNew York, New York 10029the strong shearing forces of nitrogen cavitation fail to4 Protein Chemistry Core Facilityseparate them.Howard Hughes Medical Institute
The CNS synapse should therefore be thought of as aColumbia Universityprimary adhesive device—an adherens junction—uponNew York, New York 10032whose scaffold has been superimposed, over the course5 Kratos Analytical Inc.of evolution, the molecular entities that mediate its phys-Chestnut Ridge, New York 10977iology (Fannon and Colman, 1996; Colman, 1997). Both6 Department of Pharmacologythe adherens junction and the CNS synapse share simi-Osaka University Medical Schoollar features, such as an intercellular cleft of approxi-Osaka 565mately the same distance, discrete concentrations ofJapanmaterial, termed “densities,” held in register just below7 Department of Biochemistrythe cytoplasmic faces of the apposed membranes, andUniversity of Texas Southwestern Medical Centerthe presence of calcium-dependent cell-cell adhesionDallas, Texas 75390molecules—classic cadherins—which in all epitheliamediate strong, selective cell-cell adhesion (Colman,1997; Shapiro and Colman, 1999). The CNS synapse,Summaryhowever, of course exhibits properties that reflect itscomplex neurophysiological functions. On the presyn-We report the purification of a presynaptic “particleaptic side, proteins are coassembled that mediate syn-web” consisting of �50 nm pyramidally shaped parti-aptic vesicle clustering, exocytotic fusion, and mem-cles interconnected by �100 nm spaced fibrils. Thisbrane retrieval (Chen and Scheller, 2001; Slepnev andis the “presynaptic grid” described in early EM studies.De Camilli, 2000), while the postsynaptic compartmentIt is completely soluble above pH 8, but reconstitutesof the junction harbors neurotransmitter receptors andafter dialysis against pH 6. Interestingly, reconstitutedtheir associated signaling components (Rao et al., 2000;particles orient and bind PSDs asymmetrically. MassCraig and Boudin, 2001).
spectrometry of purified web components reveals ma-When viewed by electron microscopy, under optimal
jor proteins involved in the exocytosis of synaptic vesi- preparation conditions, differences between the pre-cles and in membrane retrieval. Our data support the and postsynaptic densities are apparent. While the post-idea that the CNS synaptic junction is organized by synaptic membrane exhibits a relatively continuoustransmembrane adhesion molecules interlinked in the thickening below its membrane (the PSD, Peters et al.,synaptic cleft, connected via their intracytoplasmic 1991), on the presynaptic side a prominent intermittentdomains to the presynaptic web on one side and to the “gridwork” or “web” has been noted (Bloom and Agha-postsynaptic density on the other. The CNS synaptic janian, 1968). This web was described as consisting ofjunction may therefore be conceptualized as a compli- electron dense particles (�50 nm in diameter) arrangedcated macromolecular scaffold that isostatically brid- in a regular network just beneath the plasma membraneges two closely aligned plasma membranes. (Pfenninger et al., 1972), and it was speculated that
these presynaptic particles participate in the alignmentIntroduction and docking of synaptic vesicles at the plasma mem-
brane. More contemporary investigations have de-The concept of the synapse was originally framed in scribed a “presynaptic cytomatrix,” comprising fibrillarphysiological terms, but it was also realized that there structures that originate at the presynaptic plasmahad to be a stable physical structure mediating the be- membrane and extend deeply into the presynaptic com-
partment (Landis et al., 1988; Hirokawa et al., 1989;Garner et al., 2000)). The presynaptic cytomatrix also8 Correspondence: [email protected]

Neuron64
likely plays a role in tethering and guiding synaptic vesi- ative between CNS synaptic membranes, and it indicatesthat the activation factors controlling transynaptic adhe-cles toward the presynaptic membrane.sion are likely to be different on pre- and postsynapticThe PSD has been well characterized (Kennedy, 2000),sides. We first sought to identify conditions which mightand has generally been considered the primary orga-maintain pre- to postsynaptic attachment after selectivenizer for the synaptic junction, an entity essentially inde-membrane solubilization. Starting with synaptosomalpendent of the presynaptic components of the synapsemembranes, we tested several extraction protocols, in-in terms of structural connections. However, the realiza-cluding a variety of detergents, salts, and pH titrationstion that pre- to postsynaptic adhesion is critical forin order to find conditions under which we could isolatethe maintenance, stabilization, and physiology of theintact synaptic junctions from vesicular and other mem-synapse has forced a reevaluation of this view. At thebranous components while preserving the adhesive link-CNS synapse, pre- and postsynaptic membranes withage. Insoluble pellets derived from these experimentstheir associated bona fide adhesion molecules align awere examined by electron microscopy. We found thatdynamic transcellular structural “scaffold” whose ele-extraction with 1% TX-100 at pH 6 solubilizes almostments penetrate both apposed membrane bilayers andall plasma membrane, but retains in the insoluble pelletprotrude into the cytoplasms of the pre- and postsynap-paired pre- and postsynaptic structures, held in registertic compartments (Tanaka et al., 2000). As at the neuro-by the 20–30 nm filaments or “pegs” that protrude intomuscular junction (Hall and Sanes, 1993), the moleculesthe cleft and which are observed in native synapses (Figurenecessary for inducing and processing neurotransmitter1C; arrowheads in [F]).release are embedded within or linked to the scaffold,
In addition, when viewed en face, perforated disksthus ensuring their positions in relation to the synapticwere frequently observed (Figure 1G) indicating that thiscleft. A major unanswered question is: how are the mole-morphology, when it occurs, is real, remarkably stable,cules which mediate physiological activity integratedand resistant to detergent extraction. pH 6 isolated syn-with those that create the structural scaffold of the syn-aptic junctions also exhibited a meshwork in both theaptic junctional complex?pre- and postsynaptic densities that was absent in junc-We have now experimentally determined conditionstions treated at pH 8 (compare Figures 1F and 1G with(pH 6) under which synaptic junctional complexes—with1D and 1E). pH 6-treated synaptic junctions exhibitedintact pre- to postsynaptic connections—can be iso-few or no membrane attachments. These results reveallated in high yield and high purity. Treatments that par-the existence of a stable, plasma membrane-indepen-tially disrupt synaptic junction organization (1% Triton,dent proteinaceous scaffold which links and holds inpH 7–7.5) revealed �50 nm presynaptic particles con-register the pre- and postsynaptic densities.nected to the PSD via filaments that are likely to partici-
It is known that the classic cadherins participate inpate, presumably indirectly, in cell-cell adhesion. Pro-adhesive interactions across the synaptic cleft (Fannongressive elevation of the pH to 8 disrupts this interactionand Colman, 1996; Uchida et al., 1996; Benson and Ta-and solubilizes the presynaptic particles. Remarkably,naka, 1998; Shapiro and Colman, 1999). We investigatedsubsequent lowering of pH to 6 via dialysis reconstituteswhether cadherins were retained in isolated synapsesthe particle web and permits its asymmetric reattach-obtained after treatment at pH 6. After immunogold-label-ment to PSDs. Mass spectrometry of presynaptic webing of purified junctions with an N-cadherin antibody (Fig-fractions demonstrates that the web is composed ofure 2), we found label at or near the synaptic cleft, espe-a subset of proteins involved in vesicle recycling andcially at sites where there was an apparent tight pre- tomembrane fusion. Thus, like the PSD, the presynapticpostsynaptic apposition (arrowheads). This supports theweb should perhaps be recognized as an authentic “mo-idea that cadherin-based adhesive interactions are inte-lecular machine,” integral to the complex functions ofgral components of synaptic junctional complexes (Fan-the presynaptic compartment. Our data also reveal thenon and Colman, 1996; Uchida et al., 1996). It should be
existence of a macromolecular scaffold which spans thenoted that many junctions did not label with N-cadherin
synaptic cleft and holds the presynaptic web and PSDantiserum, as expected, since this cadherin is specific
in strict register. only to certain synaptic junctions (Fannon and Colman,1996; Benson and Tanaka, 1998; Huntley and Benson,
Results 1999), while different classic cadherins operate to medi-ate adhesion at other synapses in the CNS.
A Stable Linkage between Pre- and Postsynaptic In detergent-treated synaptosomal membranes incu-Specializations Is Maintained even after Membrane bated at increasing pH values from 6 to 8 in 0.5 unitSolubilization increments, a stepwise disruption of the presynapticIncompletely characterized adhesive linkages maintain specialization was observed. A relatively intact presyn-the apposition of pre- and postsynaptic compartments aptic specialization was observed at pH 6 (Figures 3Ain synaptosomal membrane preparations (Figure 1A) de- and 3E; arrowheads in [A]), while at pH 6.5, the presynap-spite the high shear forces employed during homogeni- tic specialization was partially disrupted, revealing parti-zation of brain tissue. However, the presynaptic special- cles that appeared to be attached preferentially to theization is disintegrated by detergent treatment of PSD (arrowheads in Figures 3B and 3F). Extraction at pHsynaptosomal membranes at pH 8, thus destroying the 7 more clearly revealed the array of 50 nm presynapticadhesive interactions and leaving only the PSD as an particles (Figures 3C and 3G, arrowheads). At higher pHinsoluble pellet (Figure 1B). This is an interesting obser- values (pH 7.5–8), the particles were completely solubi-vation in light of the demonstration that homophilic ad- lized, leaving only the PSD (Figure 3D, arrowheads).
Furthermore, and of great interest, partial disruption ofhesive interactions (e.g., the classic cadherins) are oper-

Reconstitution of the CNS Presynaptic Web65
Figure 1. The pH of TX-100 Solubilization ofSynaptosomes Affects the Preservation ofSynaptic Junctions in the Detergent-Insolu-ble Pellet
Intact synaptosomes, showing vesicular andmitochondrial components and intact fila-mentous crossbridges (A) were treated with1% TX-100, at pH 8 (B, D, and E) or pH 6(C, F, and G), and the insoluble phase waspelleted, fixed, and processed for thin-sec-tion electron microscopy. PSDs were ob-tained when membranes were solubilizedwith 1.0% TX-100 buffered to pH 8 (B, D, andE) while paired electron dense profiles wereobserved when membranes were solubilizedwith 1% TX-100 at pH 6 (C, F, and G). Highmagnification images of single detergent-extracted synapses are shown in panels (D)–(G). The synaptic material found in PSD prep-arations was elongated and retained the di-mensions of PSDs observed in intact tissue,but the fine substructure was noticeably ab-sent and appeared collapsed (D and E). Incontrast, synaptic junctions observed in pH6, TX-100 extracts were characterized by thepresence of both pre- and postsynaptic den-sities that retained a file filamentous networkof “loops” (F and G). In between the two den-sities, filamentous material appeared to con-nect the pre- and postsynaptic densities (ar-rowheads in [F]). Perforations (G) or diskswere visible when synapses were viewed enface. Bar � 500 nm in (A)–(C) and 100 nm in(D)–(G).
the synaptic junction by extraction at an intermediate treated, disrupted junctions (compare with Figure 3H,arrowheads). We therefore conclude that the particlespH (pH 7) revealed filaments in the synaptic cleft that
connected the presynaptic particles with the PSD (Fig- are prominent, bona fide features of the presynapticcompartment where they underlie presynaptic mem-ure 3H, arrowheads).
Treatment of cortical tissue with ethanolic phospho- branes, appositional to the PSD. Both specializationsare likely to be connected to the cytoplasmic domainstungstic acid (EPTA) revealed regularly spaced presyn-
aptic particles as well as an intracleft dense line (see of integral membrane proteins whose extracellular seg-ments are bound to each other in the cleft. Together,Figure 3I, and Pfenninger et al., 1972). Isolated junctions
(TX-100, pH 6) treated with EPTA revealed that the PSD this assemblage may be considered a cell-cell adhesivescaffold upon which is superimposed the functional ele-is in register with an array of presynaptic particles (Fig-
ures 3J and 3K) similar to what is observed in intact ments of the CNS synapse.tissue, although EPTA-treated, detergent-isolated junc-tions did not display the intracleft dense line (Figure Separation and Identification of Pre-
and Postsynaptic Components3, compare I with J–K). Interestingly, in EPTA-treatedjunctions, extraction revealed fine filaments connecting We showed above that treatment with TX-100 at pH
6 preserves the linkage of the presynaptic membranepresynaptic particles with the PSD (Figure 3L, arrow-head) remarkably similar to those observed in osmium- specialization, including that of the presynaptic particles

Neuron66
Figure 2. Immunogold Localization of N-Cadherin in pH 6 Detergent-Extracted Synaptic Junctions
The insoluble pellet derived from extraction of synaptosomes with 1% TX-100 at pH 6.0 was fixed with paraformaldehyde and processed forthin-section electron microscopy without osmication. Antibodies to N-cadherin labeled the cleft of isolated junctions when the junctions wereviewed from the side (arrowheads). N-cadherin antibody also labeled synapses sectioned en face (arrow). Bar � 500 nm.
to the PSD, while treatment with the same detergent at SNAP-25, or Munc-18 (data not shown). In contrast tothe differential TX-100 solubility of syntaxin, SNAP-25,pH 8 disrupts the pre- to postsynaptic scaffold, solubiliz-
ing the presynaptic specialization and the particle web and Munc-18 with respect to pH, the presynaptic cyto-matrix components rim and bassoon were primarily in-embedded in it, leaving only the PSD in the detergent-
insoluble pellet. Therefore, we might expect that molecules soluble at all pH values, although small amounts of bothproteins were solubilized at pH 8–9 (Figure 4, lanes 5–8).involved in synaptic vesicle dynamics at the presynaptic
membrane, if connected to the synaptic scaffold, would These results indicate that these proteins either remainassociated with PSDs via interactions with other mole-be retained in the detergent-insoluble pH 6 synaptic junc-
tional pellet, but might be soluble at pH 8. Most postsyn- cules and under conditions in which there is no visiblepresynaptic element, or, they become detached fromaptic proteins comprising the PSD would be predicted
to be insoluble between pH 6–8 in TX-100. We might PSDs by detergent extraction, but are insoluble underthe extraction conditions and are therefore pelleted (seealso expect that molecules loosely connected to the
synaptic junctional scaffold might be solubilized in TX- Discussion). In contrast, the synaptic vesicle proteinsynaptophysin was completely solubilized even at pH 6,100 at pH 6. To evaluate these possibilities, we examined
by immunoblot the soluble and insoluble fractions from indicating that the synaptic vesicle membrane is morelabile than the presynaptic membrane specializationintact, partially disrupted and completely disrupted syn-
aptic junctions obtained by detergent extraction at in- (Figure 4, lanes 1–4).There were incremental transfers of N-cadherin andcreasing pH values (Figure 4). The representative mark-
ers were synaptophysin (vesicle); syntaxin (presynaptic �-catenin into the soluble fraction when the pH wasincreased from 6.0 to 9.0 in the presence of 1% TX-membrane); Munc-18 and SNAP-25 (presynaptic mem-
brane-associated); rim and bassoon (presynaptic cyto- 100 (Figure 4, lanes 1–4) as both molecules decreasedproportionally from the insoluble fraction (Figure 4, lanesmatrix); the glutamate receptor subunit NMDAR1 (post-
synaptic); and the adhesion-related molecules N-cadherin, 5–8). At the highest pH values (9), �50% of N-cadherinand �-catenin remained in the pellet, revealing that these�-catenin, and the “extrasynaptic” neural cell adhesion
molecule (N-CAM). molecules are components of the residual PSD fraction,although it is clear that, as expected (Fannon and Col-Syntaxin, SNAP-25, and Munc-18 were present in the
insoluble junctional fraction at low pH, and very little man, 1996; Tanaka et al., 2000), N-cadherin is also foundon the presynaptic side. NMDAR1 was completely insol-remained in this fraction at pH values above 7.5 (Figure
4, lanes 5–8), consistent with the localization of these uble at both low and high pH (Figure 4, lanes 5–8), con-sistent with its tight association with the PSD. N-CAMproteins within the presynaptic membrane specializa-
tion, and possibly within the 50 nm presynaptic particles was completely solubilized at both low and high pH(Figure 4, lanes 1–4), as might be expected consideringthemselves. A significant proportion of the above pro-
teins was solubilized at pH 6, however (Figure 4, lane its “extrasynaptic” disposition (Tanaka et al., 2000).The EM data revealed that presynaptic web compo-1), revealing synaptic and nonsynaptic pools of these
molecules. Consistent with this, repeated extraction of nents are isolated together with PSDs by detergent solu-bilization at pH 6, and are separated from PSDs byisolated synaptic junctions with TX-100 (pH 6) did not
solubilize the junction-associated pools of syntaxin, detergent at pH 8. We reasoned, therefore, that a frac-

Reconstitution of the CNS Presynaptic Web67
tion highly enriched in presynaptic components should membranes (reviewed in Hilfiker et al., 1999), were alsobe obtained in a final supernatant by sequential deter- present in the pellet fraction.gent extraction of synaptosomes, first at pH 6, and then Four other proteins found in these fractions, ATPat pH 8 (Figure 5A). The removal of nonsynaptic proteins synthase, hexokinase I (brain form), fumarate hydra-and the enrichment of synaptic proteins by sequential tase, and a putative motor protein (accession numberextraction from pH 6–8 was assessed by SDS-PAGE and NM_006839), may be derived from mitochondria whichsilver staining (Figure 5B). Synaptosomal membranes are found in both pre- and postsynaptic compartments(Figure 5B, lane 1) were treated with 1% TX-100 at pH (see Figure 1A), although the possibility exists that they6 to yield an insoluble intact synaptic junctional pellet are exported to synaptic sites (Soltys and Gupta, 1999).fraction (Figure 5B, lane 2) and a soluble fraction (Figure Of note, it has been reported that hexokinase I displays5B, lane 3). The junctions were then treated with 1% TX- fast axonal transport that is inconsistent with a mito-100 at pH 8 and pelleted. A subset of proteins originally chondrial localization and so this molecule has beenfound in the pH 6 pellet fraction was solubilized and suggested to be directly associated with presynapticrecovered in the pH 8.0 supernatant (Figure 5B, lane 4), terminals as well (Garner et al., 1996).while in the pH 8.0 pellet, a largely different set of protein It should be noted that in both fractions, spectrin �bands was revealed (Figure 5B, lane 5). The major pro- and � subunits were prominently represented, indicatingteins enriched in the pH 8 supernatant (presynaptic frac- that spectrin is likely to play structural roles on bothtion) migrated with Mrs �180, 100, 70, 60, 55, and 45k. sides of the synaptic junction.The major proteins enriched in the pH 8.0 pellet (post-synaptic fraction) had Mrs of 190, 160, 110, 90, 80, 70, The Presynaptic Web Self-Assemblesand 50k. Other proteins present equally in both fractions Evidence suggests that certain axon terminals, whenmigrated at 270 and 250k. appropriately triggered, can rapidly assemble a func-
We compared the proteins of the pre- and postsynap- tional presynaptic membrane with associated vesicletic fractions by mass spectrometry. The results of this clusters, suggesting that the presynaptic compartmentanalysis are shown in Figure 5D. Consistent with the exhibits a remarkable tendency to rapidly self-assembleexpectation that the pH 8 supernatant should consist (Ahmari et al., 2000; Scheiffele et al., 2000; Zhai et al.,mostly of presynaptic protein components, we found a 2001). These data suggested to us that perhaps thestriking enrichment of proteins which have been impli- presynaptic particles, once solubilized (in TX-100 at pHcated in either synaptic vesicle exocytosis or recycling, 8), might possibly be induced to re-form upon return toincluding the clathrin heavy chain (MW � 180 kDa), two conditions that maintain their assembly (pH 6). Accord-alternatively spliced forms of dynamin (MW � 98 and ingly, after disruption of isolated junctions (pH 8), and97 kDa), the clathrin-uncoating ATPase (Schlossman et removal of PSDs by pelleting, the supernatant wasal., 1984) hsc70 (MW � 70), and the syntaxin binding slowly dialyzed against pH 6. Remarkably, after dialysis,protein (Hata et al., 1993) Munc-18 (MW � 64). In addi- 50 nm particles were re-formed in the insoluble phasetion, a peptide sequence matching a segment of an (Figure 6A). The reconstituted particles were interlinkedexpressed DNA sequence tag (EST) in the mouse EST
by fine filaments that appeared to be similar to thosedatabase (MW � 55; accession number AA000408) was
observed linking the presynaptic web in synapses inidentified. The translated EST nucleotide sequence was
situ (Pfenninger et al., 1972), and these also appearedfound to correspond to the carboxyl terminus of what
to preserve the native spacing between particles. Thismust be a novel member of the septin family of GTPasesmorphology was reconstituted only by slow dialysis; athat is associated with membranes belonging to therapid shift from pH 8 to pH 6 yielded amorphous insolu-presynaptic compartment. As a group, the septins haveble material (Figure 6B). From these results, we concludebeen implicated in the establishment of diffusion barri-that proteins comprising the presynaptic web, when iso-ers which delineate distinct membrane compartmentslated and then allowed to slowly coassociate, are ablebetween the mother and daughter cells in budding yeastto reassemble structures that correspond to the presyn-(Adam et al., 2000; Barral et al., 2000; Takizawa et al.,aptic particles in native synapses.2000). This novel septin family member displayed almost
We were curious whether these reconstituted presyn-complete identity to human KIAA0128 and mouse septinaptic particles might be capable of reattaching to PSDs6 (accession numbers D50918 and AB023622, respec-in vitro. We disrupted pH 6-isolated junctions by solubili-tively) across the length of the EST sequence, but dif-zation at pH 8 and then reconstituted the presynapticfered in sequence with known septins within the peptideparticles by dialysis (pH 6). We found that the 50 nmregion identified by mass spectrometry (not shown).particles were able to reattach to the PSD (Figure 6C).Tentatively, we have termed this protein septin 6A.This association appeared to be specific because theIn contrast to the prominent representation of proteinsparticles were always found to be asymmetrically ar-implicated in synaptic vesicle function in the superna-rayed along one side of the PSD. In contrast to thetant fraction, the pellet fraction contained many cy-specific reassociation of presynaptic particles with thetoskeletal or structural proteins, most of which havePSDs, when disrupted junctions were dialyzed againstbeen shown previously to be abundant within the PSDpH 6 buffer, no attachment of any material was observed(Kennedy, 1998; Kennedy et al., 1983; Walikonis et al.,when the dialysate was maintained at pH 8 (Figure 6D).2000). The PSD proteins identified in the pellet includedThe reconstituted particles shown in Figure 6E (rightspectrin, the myosin V heavy chain, neurofilament M,panel) closely resemble those found in native synapsesneurofilament L, and the CamKII � subunit (Figure 5C).from cortical tissue (Figure 6E, left and middle panels).The synapsins 1A/1B, which are actin binding proteins
involved in synaptic vesicle targeting to presynaptic In all cases, whether viewed in tissue from the side or

Neuron68
Figure 3. Presynaptic Particles and Their Tethers to the PSD Are Revealed upon Partial Disruption of Isolated Synaptic Junctions or byStaining Intact Junctions with EPTA
Presynaptic particles are obscured by other elements of the presynaptic specialization in pH 6 extracted synaptic junctions (A and E; arrowheadsin [A] show intact presynaptic specialization), but are progressively revealed after treatment of synaptosomes with TX-100 at pH 6.5 (B andF, arrowheads), and more so at pH 7 (C and G, arrowheads). Extraction at pH 8 completely removes all presynaptic elements leaving onlythe PSD in the insoluble pellet (D, arrowheads). Upon extraction at pH 7, sufficient material is removed from pre- and postsynaptic densities toreveal the presence of tethers linking the particles to the PSDs (arrowheads in [H]). Arrows in (F) and (G) indicate subsynaptic “bodies.” EPTAtreatment of intact isolated junctions (1% TX-100 at pH 6) reveals presynaptic particles remarkably similar to those seen by partial disruption atmildly alkaline pH (J–K). EPTA-stained particles in isolated junctions (J–K) were similar to those found in tissue (I). At high magnification, tetherslinking particles to PSDs were observed in EPTA-treated isolated junctions (arrowheads, L) that were identical to those observed in partially disruptedosmium-treated junctions (arrowheads, H). Bar � 500 nm in (A)–(D), 250 nm in (E)–(G) and (I)–(K), and 100 nm in (H) and (L).

Reconstitution of the CNS Presynaptic Web69
en face (Figure 6E, left and middle panels, respectively)or in the reconstituted particle fraction (Figure 6E, rightpanels), the particles were �50 nm in size (brackets inFigure 6E).
We next investigated which of the proteins originallyfound in the presynaptic extract (see Figure 4, lane 4)are reconstituted as particles upon dialysis. Superna-tants and pellets obtained from the reconstitution reac-tion in the absence of PSDs were electrophoresed andproteins were detected by silver staining (Figure 7A) orby immunoblotting (Figure 7C). Most of the proteinsoriginally found in the pH 8 supernatant (compare withFigure 5B) were found in the insoluble pellet upon dial-ysis at pH 6 (Figure 7A, lane 2) and were largely depletedfrom the supernatant, except for a single protein bandthat was still prominently represented in the supernatantafter dialysis (Figure 7A, lane 1, asterisk). Mass spec-trometry revealed this to be hsc70 (Figure 7A, lane 1,asterisk, see also Figure 5). Most of the proteins foundoriginally in the presynaptic extract were found to recon-stitute into the 50 nm particles, but not, however, septin6a, which appeared to have been degraded in pro-cessing. Of the presynaptic cytomatrix components, im-munoreactivity for both rim and bassoon was observedexclusively in the reconstituted pellet, although degra-dation of these two large protease-sensitive proteinswas also noted (Figure 7C).
To evaluate whether any of the adhesion moleculegroups known to be present at the synapse (Fannonand Colman, 1996; Song et al., 1999; Rao et al., 2000)might be involved in the attachment of the presynapticparticles to the PSD, we looked by immunoblot for thepresence of adhesion-related molecules in the presyn-aptic particle fraction (Figure 7C). We found that N-cadh-erin, �-catenin, and a �-protocadherin, as well as CASK(proposed to mediate the cytoskeletal interactions ofthe neurexins and neuroligins) were all present in theparticle fraction (Figure 7C, pellet). Multiple adhesivemechanisms probably play roles in the attachment ofthe particles to the PSD. This conclusion is supported
Figure 4. Differential Extraction of Synaptic Components by the fact that we observed some reattachment ofSynaptosomes were extracted with 1% TX-100 at the indicated pH, particles to PSDs in the presence of EDTA (not shown),and equal volumes of soluble and insoluble material were electro- which unequivocally inactivates cadherin-mediated in-phoresed, transferred to nitrocellulose, and probed with antibodies teractions.to the indicated proteins. Presynaptic markers syntaxin, Munc-18,
To verify the specificity of the reconstitution reaction,and SNAP-25 were present in the pellet at the lowest pH values butwe determined whether reconstituted presynaptic parti-were completely extracted at pH values above 7, suggesting thatcle components could be resolubilized and then subse-these molecules are part of the presynaptic specialization. Repeated
extraction of the pH 6 pellet with TX-100 at pH 6.0 never solubilized quently reconstituted (Figure 7A, lanes 3 and 4). All majorthe insoluble pool of syntaxin, Munc-18, or SNAP-25 (data not proteins found in the first reconstituted pellet (Figureshown). Presynaptic cytomatrix proteins rim and bassoon were in- 7A, lane 2) were also found in the second reconstitutedsoluble at all pH values between 6 and 9, indicating that these pellet (Figure 7A, lane 4, arrows and arrowheads). Someproteins remain associated with the PSD under conditions in which
additional minor contaminants were apparently re-there is an absence of a visible presynaptic element. Increases inmoved during the second reconstitution reaction re-the pH of extraction from 6.0 to 9.0, which gradually removed thesulting in a “sharpening” of bands in the second pellet.presynaptic specialization, caused gradual increases in the solubil-
ity of N-cadherin and �-catenin, consistent with the localization of This second purification of the particle fraction enabledthese proteins on both pre- and postsynaptic membranes. Vesicle the identification by mass spectrometry of additionalproteins such as synaptophysin and extrasynaptic proteins such as protein constituents (Figure 7A, lane 4, arrows). TheseN-CAM were completely extractable at all pH values between 6 and proteins included the N-ethyl-maleimide-sensitive fac-9 (soluble fractions, lanes 1–4), suggesting that the structures in
tor (NSF), and the vacuolar ATP synthase (v-ATPase)which these two proteins are found are not tightly linked to thesubunit A as well as the cytomatrix protein synapsin 1b.synaptic scaffold. The NMDA receptor subunit R1 was always foundBoth NSF and v-ATPase play well-characterized rolesin the insoluble fraction at both low and high pH consistent with its
tight linkage to the PSD. in mediating vesicle fusion and recycling, respectively(Zinsmaier and Bronk, 2001; Forgac, 2000). The synapsinfamily is associated with the presynaptic cytomatrix andparticipates in the localization of synaptic vesicles and

Neuron70
Figure 5. Segregation of Synaptic Proteins into Two Fractions Based on Their Differential Solubility
(A) Schematic representation of the sequential extraction. (B) Silver-stained SDS-PAGE of fractions derived from the sequential extraction ofsynaptosomes. Synaptosomal membranes (lane 1) were solubilized with 1% TX-100 at pH 6 yielding an insoluble junctional pellet (pH 6 pellet,lane 2) and soluble nonsynaptic supernatant (pH 6 supernatant, lane 3). The junctional pellet was re-extracted in 1% TX-100 at pH 8.0. Thesupernatant (pH 8 supernatant, lane 4) and pellet (pH 8 pellet, lane 5) fractions contained different sets of proteins that were originally foundin the intact junction pellet. Arrow indicates a nonsynaptic protein that was removed by solubilization at pH 6, arrowhead indicates a proteinenriched in the pH 8 supernatant, and the asterisk indicates a protein enriched in the pH 8 pellet. (C) Fifty micrograms of the pH 8 soluble(lane 1) or insoluble (lane 2) fractions was electrophoresed on SDS-PAGE and stained with colloidal Coomassie blue. The bands identified bymass spectrometry are indicated by numbered arrows and their identity is shown in (D).
is probably not a component of the presynaptic web structure had been observed, consisting of �50–80 nmparticles arrayed below the presynaptic membrane and(see Discussion). The components that we identified in
the present study as components of the presynaptic connected by fine fibrils spaced �100 nm apart. Wenow show that these components can be isolated andparticle web are listed in Table 1. From these studies,
it is clear that the presynaptic web forms a subset of reconstituted, and appear to be an important part of thecell-cell synaptic junctional scaffold. In our preparationsinterrelated proteins embedded in the presynaptic mem-
brane specialization and it is composed of synaptic vesi- derived from tissue fractionation, the particles, thespaces between particles, and the fibrillar network ex-cle exocytosis and recycling proteins.hibit the uniform dimensions observed in situ. The 50nm particles are generally separated by a distance ofDiscussionapproximately 50–100 nm by fibrils of approximately10 nm in width. Further support for the existence of theIn early EM studies of CNS tissue (Bloom and Aghajan-
ian, 1968; Pfenninger et al., 1972), a grid or web-like CNS presynaptic web comes from recent observations

Reconstitution of the CNS Presynaptic Web71
Figure 6. Solubilized Presynaptic Particles Independently Reconstitute Their Structure and Reattachments to PSDs
(A) Fifty nanometer presynaptic particles were formed by dialysis of the soluble presynaptic fraction against pH 6 buffer in the absence ofPSDs. Reconstituted particles were regular in size and were connected by fine filaments (arrowhead). (B) A precipitate with no resemblanceto presynaptic particles was formed when the presynaptic fraction was rapidly diluted 20-fold with 1% TX-100, 20 mM Tris pH 6.0, 1 mMCaCl2. (C) Reattachment of presynaptic particles to PSDs. The solubilized presynaptic fraction was separated from the PSD pellet. PSDs wereresuspended in an equal volume of the same buffer. One twentieth of the original volume of PSDs was added back to the presynaptic fractionand the suspension was dialyzed against 1% TX-100, 20 mM Tris, pH 6.0, 1 mM CaCl2 (C), or against 1% TX-100, 20 mM Tris, pH 8.0, 1mMCaCl2 (D). In the pellet obtained from the pH 6 dialyzate, numerous PSDs were found to have 50 nm particles attached by visible filamentsto only one side of the PSD (arrowheads, C). In contrast, the pellet from the pH 8 dialyzate contained mostly PSDs with no additionalattachments (D). (E) Presynaptic particles in tissues viewed either from the side (left) or en face (middle) exhibit the same dimensions asreconstituted particles (right). Brackets in (E) denote a distance of 50 �m. Bar � 500 nm in (A) and (B) and 350 nm in (C) and (D).
of a very similar structure within motor terminals at the that presynaptic particles can assemble and disassem-ble under defined conditions. These data have importantfrog neuromusclar junction (Harlow et al., 2001). We
conclude that the particle web at the presynaptic mem- implications for interpretation of work from other labora-tories that argues for a remarkable propensity for spon-brane is a real structure, intimately involved in synaptic
vesicle localization at the presynaptic membrane. taneous self-assembly of presynaptic components onsuitable substrates. It was first shown that presynapticOur studies reveal a sensitivity of presynaptic struc-
ture to relatively small changes in pH, and demonstrate structures are generated de novo in cultures when neurites
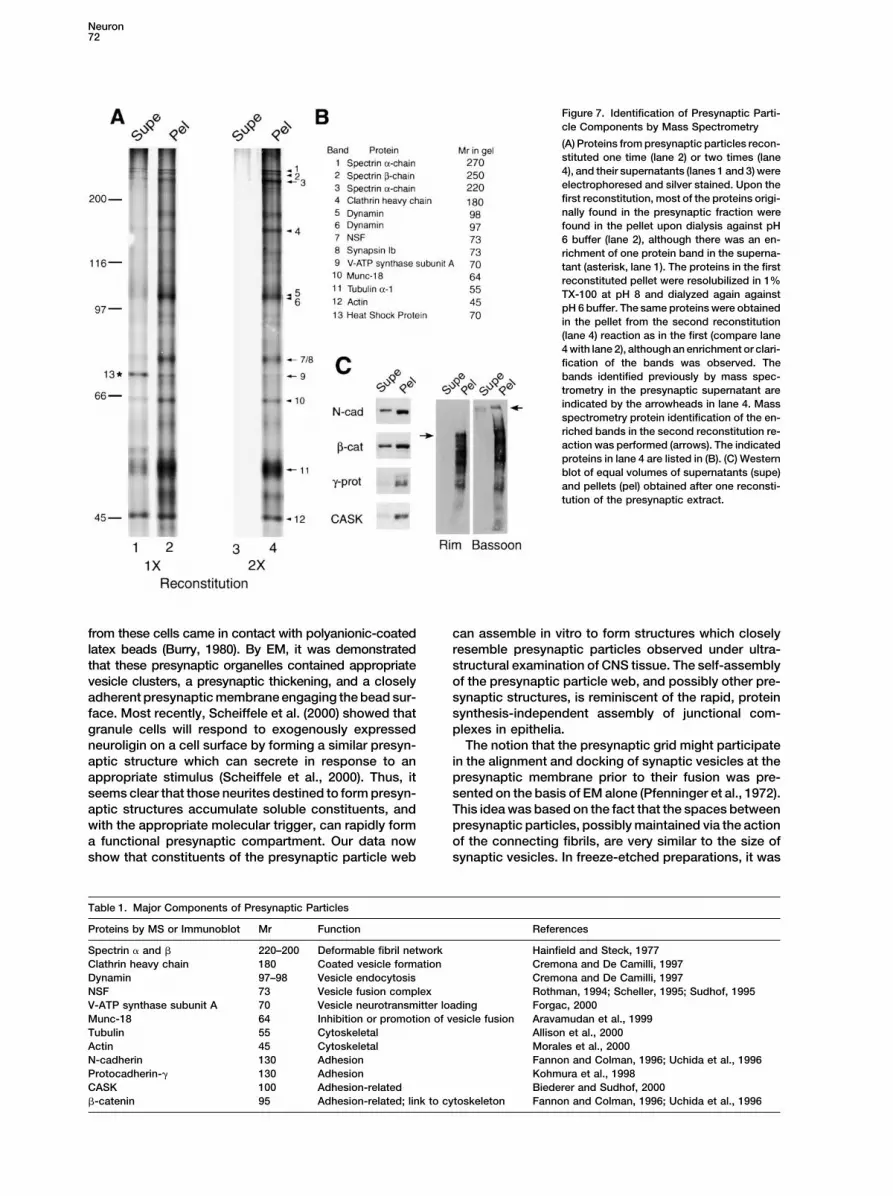
Neuron72
Figure 7. Identification of Presynaptic Parti-cle Components by Mass Spectrometry
(A) Proteins from presynaptic particles recon-stituted one time (lane 2) or two times (lane4), and their supernatants (lanes 1 and 3) wereelectrophoresed and silver stained. Upon thefirst reconstitution, most of the proteins origi-nally found in the presynaptic fraction werefound in the pellet upon dialysis against pH6 buffer (lane 2), although there was an en-richment of one protein band in the superna-tant (asterisk, lane 1). The proteins in the firstreconstituted pellet were resolubilized in 1%TX-100 at pH 8 and dialyzed again againstpH 6 buffer. The same proteins were obtainedin the pellet from the second reconstitution(lane 4) reaction as in the first (compare lane4 with lane 2), although an enrichment or clari-fication of the bands was observed. Thebands identified previously by mass spec-trometry in the presynaptic supernatant areindicated by the arrowheads in lane 4. Massspectrometry protein identification of the en-riched bands in the second reconstitution re-action was performed (arrows). The indicatedproteins in lane 4 are listed in (B). (C) Westernblot of equal volumes of supernatants (supe)and pellets (pel) obtained after one reconsti-tution of the presynaptic extract.
from these cells came in contact with polyanionic-coated can assemble in vitro to form structures which closelyresemble presynaptic particles observed under ultra-latex beads (Burry, 1980). By EM, it was demonstrated
that these presynaptic organelles contained appropriate structural examination of CNS tissue. The self-assemblyof the presynaptic particle web, and possibly other pre-vesicle clusters, a presynaptic thickening, and a closely
adherent presynaptic membrane engaging the bead sur- synaptic structures, is reminiscent of the rapid, proteinsynthesis-independent assembly of junctional com-face. Most recently, Scheiffele et al. (2000) showed that
granule cells will respond to exogenously expressed plexes in epithelia.The notion that the presynaptic grid might participateneuroligin on a cell surface by forming a similar presyn-
aptic structure which can secrete in response to an in the alignment and docking of synaptic vesicles at thepresynaptic membrane prior to their fusion was pre-appropriate stimulus (Scheiffele et al., 2000). Thus, it
seems clear that those neurites destined to form presyn- sented on the basis of EM alone (Pfenninger et al., 1972).This idea was based on the fact that the spaces betweenaptic structures accumulate soluble constituents, and
with the appropriate molecular trigger, can rapidly form presynaptic particles, possibly maintained via the actionof the connecting fibrils, are very similar to the size ofa functional presynaptic compartment. Our data now
show that constituents of the presynaptic particle web synaptic vesicles. In freeze-etched preparations, it was
Table 1. Major Components of Presynaptic Particles
Proteins by MS or Immunoblot Mr Function References
Spectrin � and � 220–200 Deformable fibril network Hainfield and Steck, 1977Clathrin heavy chain 180 Coated vesicle formation Cremona and De Camilli, 1997Dynamin 97–98 Vesicle endocytosis Cremona and De Camilli, 1997NSF 73 Vesicle fusion complex Rothman, 1994; Scheller, 1995; Sudhof, 1995V-ATP synthase subunit A 70 Vesicle neurotransmitter loading Forgac, 2000Munc-18 64 Inhibition or promotion of vesicle fusion Aravamudan et al., 1999Tubulin 55 Cytoskeletal Allison et al., 2000Actin 45 Cytoskeletal Morales et al., 2000N-cadherin 130 Adhesion Fannon and Colman, 1996; Uchida et al., 1996Protocadherin-� 130 Adhesion Kohmura et al., 1998CASK 100 Adhesion-related Biederer and Sudhof, 2000�-catenin 95 Adhesion-related; link to cytoskeleton Fannon and Colman, 1996; Uchida et al., 1996

Reconstitution of the CNS Presynaptic Web73
Figure 8. Model of the Presynaptic Web and Its Relationship with the Presynaptic Cytomatrix
(A) A model for the organization of the presynaptic specialization and its attachment to the PSD is shown. Fibrillar components originatingat the presynaptic membrane and projecting into the cytoplasm have been described in freeze-etch electron microscopy studies (Hirokawaet al., 1989; Landis et al., 1988). These fibrils may contain the presynaptic cytomatrix proteins (Garner et al., 2000). Shown superimposedupon the fibrils are the presynaptic particles which exhibit a visible attachment to the PSD (solid line, right panel). Under conditions in whichpresynaptic particles are stripped from the PSD (1% TX-100, pH 8) and hence solubilized, presynaptic cytomatrix components remain in theinsoluble pellet along with the PSD, suggesting the possibility of a distinct adhesive system linking the cytomatrix with the PSD (dotted linesin right panel). (B) Three-dimensional model of the synaptic scaffold. Presynaptic particles are linked by adhesion molecules across thesynaptic cleft to the PSD. The particles form an hexagonal array that may support the localization of synaptic vesicles to the membrane andtheir subsequent fusion. Presynaptic particles also contain components necessary for the retrieval of vesicle membrane proteins after theirfusion with the plasma membrane. The particles may serve to immobilize endocytosis components at high concentrations next to thepresynaptic membrane such that they might be readily available after vesicle fusion.
shown that within the spaces between particles, there tion. In agreement with this analogy, the spacing be-tween web elements at the presynaptic and erythrocyteare depressions in the presynaptic membrane which
appear to be sites of vesicle attachment (Pfenninger membranes appears to be consistent (50–100 nm). Thepresynaptic web may also exhibit analogies to anotheret al., 1972). Upon synaptic stimulation, the distance
between particles transiently increases, suggesting an spectrin-based structure—the terminal web of intestinalepithelial cells (Hull and Staehelin, 1979). This structureinsertion of material between the particles as a conse-
quence of fusion of the synaptic vesicle with the plasma contains a dense spectrin network that overlays a com-plex of intermediate filaments. This arrangement helpsmembrane (Triller and Korn, 1985). Thus the web ap-
pears to provide a framework to allow the docking and to keep actin bundles orthogonal to the membrane sur-face. Thus, the presynaptic web may participate in main-fusion of synaptic vesicles at the presynaptic mem-
brane. However, it is not known how the protein compo- taining presynaptic components in the proper orienta-tion orthogonal to the synaptic plane. In several othernents of the presynaptic web identified in the present
study actively participate in the synaptic vesicle cycle. respects, the presynaptic web identified here resemblesthe “active zone material” (AZM) found at the presynap-Our studies suggest that one important role of the pre-
synaptic web may be in the sequestration of vesicle tic plasma membrane of the neuromuscular junction.Using electron microscope tomography, Harlow, et al.recycling proteins such as clathrin and dynamin in cir-
cumscribed presynaptic microdomains. Given that (2001) have resolved the AZM into individual physicalcomponents—“ribs,” “beams,” and “pegs”—that formclathrin and dynamin exhibit the tendency to self-
assemble endocytotic structures around naked lipid a complex presynaptic framework whose intracellularposition and structure suggest that its componentsvesicles (Takei et al., 1998), a sequestering mechanism
may be needed to prevent the precocious self-assembly function in docking and vesicle fusion. It is postulatedthat components of the AZM are linked to transmem-of these molecules.
The regular array of particles of defined size, intercon- brane molecules embedded in the synaptic membranesimilar to the observed linkage for the presynaptic webnected by short fibrillar components, observed at the
presynaptic membrane is highly reminiscent of the spec- to PSDs in our study.trin-based web underlying the erythrocyte plasma mem-brane (Hainfeld and Steck, 1977). Indeed, the presence An Asymmetric Transcellular Synaptic Scaffold
The central nervous system synapse, while optimizedof spectrin as a prominent component in our recon-stituted presynaptic web fraction suggests the spacing over the course of evolution for polarized cell-cell com-
munication, nevertheless retains many of the propertiesbetween particles could be maintained by a flexiblespectrin-based filament network and this structural of the adherens junction from which it appears to be
derived (Colman, 1997; Shapiro and Colman, 1999). Theframework might explain the dynamic changes in spac-ing of the presynaptic particles upon synaptic stimula- adherens junction adhesively links two apposing parallel

Neuron74
plasma membranes and locks them into a transcellular The presynaptic cytomatrix most likely emanates intoscaffold, half of which is contributed by each participat- the cytoplasm from the presynaptic membrane as fibrilsing cell. The scaffold consists of transmembrane adhe- that extend well into the presynaptic compartment assion molecules that interlock in the intercellular cleft with has been observed in freeze-etched preparations (Hiro-cytoplasmic domains that protrude into the intracellular kawa et al., 1989; Landis et al., 1988). These fibrils mightmilieu. Anchored to these cytoplasmic domains in each tether or guide synaptic vesicles via the action of thecell are signaling molecules and cytoskeletal elements synapsins. While clearly distinct from the presynapticthat organize and cluster beneath the junctional mem- particles, the fact that the cytomatrix proteins rim andbrane plates (Aberle et al., 1996). In terms of adhesive bassoon are found in the reconstituted presynaptic par-forces, the adherens junction is symmetrical and is held ticle fraction (see Figure 7) suggests that these proteinstogether by homophilic cadherin-mediated interactions. may interact with proteins found in the particles. It is
On the other hand, while cadherin-based adhesive intriguing to speculate that the cytomatrix fibers, firmlyinteractions are undoubtedly important for the mainte- anchored to the presynaptic membrane, provide nucle-nance of the rigorously parallel synaptic cleft, their ap- ation sites for the components of the presynaptic parti-proximately equal partitioning and differential solubility cles (Figure 8). The presynaptic particle proteins wouldin both pre- and postsynaptic fractions suggests that then be available at high concentrations directly under-these molecules may have differential roles in organizing neath the presynaptic membrane. In conclusion, the re-or aligning synaptic elements on both the pre- and post- sults of this study shed light on the organization of thesynaptic side (Tanaka et al., 2000). On the postsynaptic synaptic junctional complex and expand the frameworkside, our study suggests that cadherin molecules are upon which to address the interrelationships of its pro-firmly anchored to the PSD via interactions of their cyto- tein components.plasmic domains either directly or indirectly with PSD
Experimental Proceduresstructural proteins. In contrast to the tight anchorage ofpostsynaptic cadherins, the presynaptic cadherins are
Antibodiesmore easily released from the junction by detergent, areThe N-cadherin antiserum has been described (Tanaka et al., 2000).more dynamic, and thus may subserve other roles. It isA polyclonal antibody to �-protocadherins, corresponding to the
interesting that we have noted that the classic cadherins conserved portion of the cytoplasmic domain (Wu and Maniatis,possess within their cytoplasmic domains two tyrosine- 1999), was raised in rabbits. Some antibodies (synaptophysin, syn-based consensus sorting sequences (YXXø; ø � large aptotagmin, Munc-18, SNAP-25, rim, and �-catenin) were from
Transduction Laboratories (Lexington, KY). NMDA receptor R1 andaliphatic) for the AP-1 adaptor proteins (Vincent et al.,N-CAM antibodies were obtained from Pharmingen (San Diego, CA).1997) involved in clathrin-mediated endocytosis. ThusBassoon antisera were obtained from Stressgen Biotechnologiesit is possible that presynaptic cadherins may have a role(Victoria, BC). All other antisera were from Sigma (St. Louis, MO).
in organizing or localizing the presynaptic endocytosiscomplex which may correspond to the presynaptic parti- Synaptosome Preparationcles (Figure 8B). Synaptosomes were prepared using a one-step synaptosome prep-
aration method based on the known isopycnic densities of variousThe machinery that guides synaptic vesicles to theircellular components (Cohen et al., 1977). Cortices from five adultappropriate location within the presynaptic compart-male rats (5–6 g total wet weight) were homogenized in 15 ml ofment prior to docking and fusion (the presynaptic cyto-solution (0.32 M sucrose, 0.1 mM CaCl2, 1 mM MgCl2, 0.1 mM PMSF)matrix; Garner et al., 2000) appears to be linked to theat 4�C using a Teflon-glass homogenizer. The homogenate was
synaptic junction by a different mechanism than that of brought to a final sucrose concentration of 1.25 M by the additionthe presynaptic particles (Figure 8A). Prior to docking, of 2 M sucrose (70 ml) and 30 ml 0.1 mM CaCl2 and divided into sixsynaptic vesicles are aligned at their position within the 25 � 89 mm ultracentrifuge tubes. The homogenate was overlaid
with 10 ml 1.0 M sucrose, 0.1 mM CaCl2, and with 5 ml homogeniza-presynaptic compartment by structural cytomatrix ele-tion solution and centrifuged (100,000 � g; 3 hr; 4�C). After centrifu-ments which involve proteins such as the synapsins,gation, a band representing synaptosomal membranes was col-bassoon, and piccolo (Cases-Langhoff et al., 1996; Hil-lected at the 1.25 M/1.0 M sucrose interface.fiker et al., 1999; tom Dieck et al., 1998). These molecules
appear to be very tightly linked to the PSD across the Solubility Analysis of Synapses and Synaptic Proteinssynaptic cleft, as evidenced by their presence in the Synaptosomes were diluted 1:10 with ice cold 0.1 mM CaCl2 andmost detergent-resistant fraction along with the PSD— divided into 100 �l aliquots. An equal volume of 2� solubilization
buffer was added (2% TX-100, 0.2 mM CaCl2, 40 mM Tris bufferedconditions where no presynaptic elements are detectedto various pH values; see Results); the samples were then mixedby electron microscopy (see Figures 3 and 4). Theseand incubated (20 min on ice). The samples were then centrifugedresults are consistent with the finding that some of theat 10,000 � g for 30 min at 4�C. Supernatants were collected andcytomatrix proteins are highly enriched in PSD fractionspellets resuspended in an equal volume (200 �l) of 0.1 mM CaCl2.(tom Dieck et al., 1998; Wang et al., 1999). Alternatively, An equal volume of 2� SDS-PAGE sample buffer was added and
the presence of these presynaptic vesicle targeting mol- the samples were loaded onto 7.5% SDS gels. Gels were transferredecules in the PSD fraction may simply reflect their rela- to nitrocellulose and the membranes were probed with antibodies.tive insolubility when compared to the rest of the pre-
Electron Microscopysynaptic machinery and not their specific linkage toFor EM, rats were anesthetized and perfused transcardially (4%the PSD.paraformaldehyde, 2% glutaraldehyde; Hank’s solution with 15 mMRegardless of their linkage mechanisms to the PSD,Hepes buffer, pH 7.0; 1 mM CaCl2). Brains were removed and post-
the differential solubility of the presynaptic cytomatrix fixed. Cortical sections were treated with 2% OsO4, 2 hr, and stainedand the presynaptic web elements suggests that these en bloc with uranyl acetate, dehydrated, and embedded in Eponare two distinctly organized, yet interrelated, substruc- (Embed 812, Electron Microscopy Sciences). Alternatively, cortical
sections were treated with 1% phosphotungstic acid in absolutetures within the presynaptic compartment (Figure 8A).

Reconstitution of the CNS Presynaptic Web75
ethanol containing a trace of water (2 drops of 95% ethanol per data-dependent mode where the machine measured the intensityof all peptide ions in the mass range of 400 to 1400 (mass to charge10 ml) for 2 hr at 50�C. The tissue was then rinsed in ethanol and
Epon-embedded. For isolated synaptic junctions, pellets were fixed ratio) and isolated the peptide peak with the highest intensity forcollision-induced dissociation. Thus, the masses of both the peptidewith 4% glutaraldehyde; phosphate buffer-pH 7.4. For standard EM,
pellets were treated with 2% osmium tetroxide in acetate buffer, and its daughter ions were detected. The accurately measuredmasses of the tryptic peptide and its fragments were used to searchstained en bloc with uranyl acetate, dehydrated through ethanols,
and embedded in Epon. EPTA treatment of isolated junctions was for protein candidates in the protein sequence database with theprogram “PepFrag” (Fenyo et al., 1998) (http://prowl1.rockefeller.done as above for rat cortex. Ultra-thin sections were cut and, for
osmicated specimens, stained with uranyl acetate-lead citrate and edu/prowl/pepfragch.html).viewed in an electron microscope at 8,300–50,000� magnification.
Immunogold labeling was performed essentially as described MALDI-MS Analysis(Phend, et al., 1995). Gel bands were prepared for digestion by washing twice with 200
�l 0.05 M Tris, pH 8.5/50% acetonitrile for 20 min with shaking. AfterSeparation of Pre- and Postsynaptic Proteins removing the washes, the gel pieces were dried for 30 min in aSynaptosomes were extracted sequentially with 1% TX-100 at pH Speed-Vac concentrator. Gels were digested by adding 0.05 �g6.0 followed by pH 8.0. Synaptosomes (4 ml, prepared as described modified trypsin in 13–15 �l of 0.025 M Tris, pH 8.5. The tubes wereabove) were diluted 1:10 with ice cold 0.1 mM CaCl2. The suspension placed in a heating block at 32�C and left overnight. Peptides werewas brought to a final concentration of 20 mM Tris, pH 6.0 and 1% extracted twice with 50 �l 50% acetonitrile/2% TFA and the com-TX-100. The membranes were extracted (30 min) and the insoluble bined extracts were dried and resuspended in matrix solution. Matrixmaterial was pelleted (40,000 � g, 30 min). The supernatant was solution was prepared by making a 10 mg/ml solution of 4-hydroxy-decanted and proteins precipitated with 10 volumes acetone at �-cyanocinnamic acid (Sigma; St. Louis, MO) in 50% acetonitrile/�20�C and recovered by centrifugation at 15,000 � g for 30 min. 0.1% TFA and adding two internal standards, angiotensin and bo-The pellet was resuspended in 10 ml 20 mM Tris, pH 6.0 and 1% TX- vine insulin, to the matrix solution. The dried digest was dissolved100; a small aliquot was taken for gel electrophoresis, reextracted, in 3 �l matrix/standard solution and 0.7 �l was spotted onto theprecipitated, and pelleted as above. The insoluble pellet was resus- sample plate. When the spot was completely dried, it was washedpended (10 ml of 20 mM Tris, pH 8.0, 1% TX-100), extracted, centri- twice with water to remove buffer salts. MALDI mass spectrometricfuged, and the supernatant reprecipitated as above. The insoluble analysis was performed on the digest using a PerSeptive Voyagerpellets as well as the recovered supernatants from all three extractions DE-RP mass spectrometer in the linear mode. Peptide masses werewere dissolved in 5% SDS and the protein concentrations were deter- used to search the protein database with the program ProFoundmined using the BCA protein assay (Pierce; Rockford, IL). (http://129.85.10.192/profound_bin/WebProFound.exe).
Reconstitution of Presynaptic Particles Acknowledgmentsfrom Solubilized FractionsThe presynaptic fraction was prepared as above and subsequently This work was supported by grants from the NIH (NS20147 anddialyzed against 1% TX-100, 20 mM Tris, pH 6.0, 0.1 mM CaCl2 for NS041687) and the NY State Spinal Cord Injury Research Programthree changes at 10 hr each. Control experiments included dialysis (C016883) to D.R.C. G.R.P. was supported by an NRSA (NS10836)for the same time but at pH 8.0. The dialysates were centrifuged at from the NIH and W.Z. by NS37731.15,000 � g and equal volumes of the supernatant and pellets wereelectrophoresed on SDS-PAGE and silver-stained, Coomassie
Received March 19, 2001; revised July 20, 2001.stained, or transferred to membranes for Western blotting. Pelletswere fixed and processed as above for EM. In some cases, purified
ReferencesPSDs were added to the reconstitution reaction. The PSD pelletwas resuspended in 1%TX-100, 20 mM Tris, pH 8, 0.1 mM CaCl2,
Aberle, H., Schwartz, H., and Kemler, R. (1996). Cadherin-cateninin a volume equal to that of the supernatant. One twentieth of thecomplex: protein interactions and their implications for cadherinoriginal volume of PSDs was added back to the presynaptic super-function. J. Cell. Biochem. 61, 514–523.natant prior to dialysis.
Adam, J.C., Pringle, J.R., and Peifer, M. (2000). Evidence for func-tional differentiation among Drosophila septins in cytokinesis andMS/MS Analysiscellularization. Mol. Biol. Cell 11, 3123–3135.Twenty to fifty micrograms of each protein sample (see above) was
loaded onto a 16 � 24 cm 7.5% SDS-PAGE gel and electrophoresed Ahmari, S.E., Buchanan, J., and Smith, S.J. (2000). Assembly ofat 50V overnight. Proteins were visualized by staining with silver or presynaptic active zones from cytoplasmic transport packets. Nat.with colloidal Coomassie blue (Novex; San Diego, CA). Gel slices Neurosci. 3, 445–451.containing the bands of interest were excised from the Coomassie-
Allison, D.W., Chervin, A.S., Gelfand, V.I., and Craig, A.M. (2000).stained gel, destained in 25 mM NH4HCO3 buffer (pH 8.0; metha-
Postsynaptic scaffolds of excitatory and inhibitory synapses in hip-nol:water, 1:1), cleaned in a fixation solution (water:acetonitrile:ace-
pocampal neurons: maintenance of core components independenttic acid, 45:45:10) for 20 hr, swollen with water for 2 hr, and then
of actin filaments and microtubules. J. Neurosci. 20, 4545–4554.equilibrated with digestion buffer (50 mM NH4HC03, pH 8.0) for 4
Aravamudan, B., Fergestad, T., Davis, W.S., Rodesch, C.K., andmin. Tryptic digestion was carried out at 37�C for 2 hr in 10 to 20Broadie, K. (1999). Drosophila UNC-13 is essential for synaptic trans-�l of digestion buffer including 0.2 �g of sequencing grade modifiedmission. Nat. Neurosci. 2, 965–971.trypsin (Boehringer Mannheim, Indianapolis, IN). The tryptic pep-
tides were extracted twice with 70% acetonitrile in water and con- Barral, Y., Mermall, V., Mooseker, M.S., and Snyder, M. (2000). Com-centrated in a speed vac concentrator (Savant; Holbrook, NY). The partmentalization of the cell cortex by septins is required for mainte-dried sample was dissolved in 6 �l of HPLC solution A (water:aceto- nance of cell polarity in yeast. Mol. Cell 5, 841–851.nitrile:acetic acid, 97.5:2:0.5) for mass analysis. Benson, D.L., and Tanaka, H. (1998). N-cadherin redistribution dur-
HPLC-MS/MS analysis was performed in a LCQ (Finnigan MAT) ing synaptogenesis in hippocampal neurons. J. Neurosci. 18, 6892–coupled online with a capillary HPLC system (Microm BioRe- 6904.sources). Two microliters of the peptide solution was loaded on
Biederer, T., and Sudhof, T.C. (2000). Mints as adaptors. Directcapillary HPLC connected with a C18 column (10 cm length, 75 mmbinding to neurexins and recruitment of munc18. J. Biol. Chem. 275,internal diameter). The peptides were sequentially eluted from the39803–39806.HPLC column with a gradient of 5%–90% of HPLC solution B (aceto-
nitrile:water:acetic acid, 2:97.5:0.5) at a flow rate of 0.7 ml/min and Bloom, F.E., and Aghajanian, G.K. (1968). Fine structural and cyto-chemical analysis of the staining of synaptic junctions with phospho-were sprayed directly from the tip of the capillary column to a LCQ
mass spectrometer for MS/MS analysis. LCQ was operated in a tungstic acid. J. Ultrastruct. Res. 22, 361–375.

Neuron76
Burry, R.W. (1980). Formation of apparent presynaptic elements in protein” is a subunit of a calmodulin-dependent protein kinase. Proc.Natl. Acad. Sci. USA 80, 7357–7361.response to poly-basic compounds. Brain Res. 184, 85–98.
Cases-Langhoff, C., Voss, B., Garner, A.M., Appeltauer, U., Takei, Kohmura, N., Senzaki, K., Hamada, S., Kai, N., Yasuda, R., Wata-K., Kindler, S., Veh, R.W., De Camilli, P., Gundelfinger, E.D., and nabe, M., Ishii, H., Yasuda, M., Mishina, M., and Yagi, T. (1998).Garner, C.C. (1996). Piccolo, a novel 420 kDa protein associated Diversity revealed by a novel family of cadherins expressed in neu-with the presynaptic cytomatrix. Eur. J. Cell Biol. 69, 214–223. rons at a synaptic complex. Neuron 20, 1137–1151.
Chen, Y.A., and Scheller, R.H. (2001). SNARE-mediated membrane Landis, D.M., Hall, A.K., Weinstein, L.A., and Reese, T.S. (1988). Thefusion. Nat. Rev. Mol. Cell Biol. 2, 98–106. organization of cytoplasm at the presynaptic active zone of a central
nervous system synapse. Neuron 1, 201–209.Cohen, R.S., Blomberg, F., Berzins, K., and Siekevitz, P. (1977).The structure of postsynaptic densities isolated from dog cerebral Morales, M., Colicos, M.A., and Goda, Y. (2000). Actin-dependentcortex. I. Overall morphology and protein composition. J. Cell Biol. regulation of neurotransmitter release at central synapses. Neuron74, 181–203. 27, 539–550.
Colman, D.R. (1997). Neurites, cadherins and synapses reconciled. Peters, A., Palay, S.L., and Webster, H.D.F. (1991). The fine structureMol. Cell. Neurosci. of the nervous system (New York: Oxford).
Craig, A.M., and Boudin, H. (2001). Molecular heterogeneity of cen- Pfenninger, K., Akert, K., Moor, H., and Sandri, C. (1972). The finetral synapses: afferent and target regulation. Nat. Neurosci. 4, structure of freeze-fractured presynaptic membranes. J Neurocytol569–578. 1, 129–149.Cremona, O., and De Camilli, P. (1997). Synaptic vesicle endocyto- Phend, K.D., Rustioni, A., and Weinberg, R.J. (1995). An osmium-sis. Curr. Opin. Neurobiol. 7, 323–330. free method of epon embedment that preserves both ultrastructure
and antigenicity for post-embedding immunocytochemistry. J. His-Fannon, A.M., and Colman, D.R. (1996). A model for central synaptictochem. Cytochem. 43, 283–292.junctional complex formation based on the differential adhesive
specificities of the cadherins. Neuron 17, 423–434. Rao, A., Harms, K.J., and Craig, A.M. (2000). Neuroligation: buildingsynapses around the neurexin-neuroligin link. Nat. Neurosci. 3,Fenyo, D., Qin, J., and Chait, B.T. (1998). Protein identification using747–749.mass spectrometric information. Electrophoresis 19, 998–1005.
Rothman, J.E. (1994). Mechanisms of intracellular protein transport.Forgac, M. (2000). Structure, mechanism and regulation of theNature 372, 55–63.clathrin-coated vesicle and yeast vacuolar H()-ATPases. J. Exp.
Biol. 203, 71–80. Scheiffele, P., Fan, J., Choih, J., Fetter, R., and Serafini, T. (2000).Neuroligin expressed in nonneuronal cells triggers presynaptic de-Foster, M., and Sherrington, C.S. (1897). A Text Book of Physiology,velopment in contacting axons. Cell 101, 657–669.Part III: The Central Nervous System, 7 Edition (London: Macmillan).
Scheller, R.H. (1995). Membrane trafficking in the presynaptic nerveGarner, J.A., Linse, K.D., and Polk, R.K. (1996). Type I brain hexoki-terminal. Neuron 14, 893–897.nase: axonal transport and membrane associations within central
nervous system presynaptic terminals. J. Neurochem. 67, 845–856. Schlossman, D.M., Schmid, S.L., Braell, W.A., and Rothman, J.E.Garner, C.C., Kindler, S., and Gundelfinger, E.D. (2000). Molecular (1984). An enzyme that removes clathrin coats: purification of andeterminants of presynaptic active zones. Curr. Opin. Neurobiol. 10, uncoating ATPase. J. Cell Biol. 99, 723–733.321–327. Shapiro, L., and Colman, D.R. (1999). The diversity of cadherins andHainfeld, J.F., and Steck, T.L. (1977). The sub-membrane reticulum implications for a synaptic adhesive code in the CNS. Neuron 23,of the human erythrocyte: a scanning electron microscope study. 427–430.J. Supramol. Struct. 6, 301–311. Slepnev, V.I., and De Camilli, P. (2000). Accessory factors in clathrin-Hall, Z.W., and Sanes, J.R. (1993). Synaptic structure and develop- dependent synaptic vesicle endocytosis. Nat. Rev. Neurosci. 1,ment: the neuromuscular junction. Cell Suppl. 72, 99–121. 161–172.
Harlow, M.L., Ress, D., Stoschek, A., Marshall, R.M., and McMahan, Soltys, B.J., and Gupta, R.S. (1999). Mitochondrial-matrix proteinsU.J. (2001). The architecture of active zone material at the frog’s at unexpected locations: are they exported? Trends Biochem. Sci.neuromuscular junction. Nature 409, 479–484. 24, 174–177.
Hata, Y., Slaughter, C.A., and Sudhof, T.C. (1993). Synaptic vesicle Song, J.Y., Ichtchenko, K., Sudhof, T.C., and Brose, N. (1999). Neu-fusion complex contains unc-18 homologue bound to syntaxin. Na- roligin 1 is a postsynaptic cell-adhesion molecule of excitatory syn-ture 366, 347–351. apses. Proc. Natl. Acad. Sci. USA 96, 1100–1105.Hilfiker, S., Pieribone, V.A., Czernik, A.J., Kao, H.T., Augustine, G.J., Sudhof, T.C. (1995). The synaptic vesicle cycle: a cascade of protein-and Greengard, P. (1999). Synapsins as regulators of neurotransmit- protein interactions. Nature 375, 645–653.ter release. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 354, 269–279. Takei, K., Haucke, V., Slepnev, V., Farsad, K., Salazar, M., Chen, H.,Hirokawa, N., Sobue, K., Kanda, K., Harada, A., and Yorifuji, H. and De Camilli, P. (1998). Generation of coated intermediates of(1989). The cytoskeletal architecture of the presynaptic terminal and clathrin-mediated endocytosis on protein-free liposomes. Cell 94,molecular structure of synapsin 1. J. Cell Biol. 108, 111–126. 131–141.Hull, B.E., and Staehelin, L.A. (1979). The terminal web. A reevalua- Takizawa, P.A., DeRisi, J.L., Wilhelm, J.E., and Vale, R.D. (2000).tion of its structure and function. J. Cell Biol. 81, 67–82. Plasma membrane compartmentalization in yeast by messenger
RNA transport and a septin diffusion barrier. Science 290, 341–344.Huntley, G.W., and Benson, D.L. (1999). Neural (N)-cadherin at devel-oping thalamocortical synapses provides an adhesion mechanism Tanaka, H., Shan, W., Phillips, G.R., Arndt, K., Bozdagi, O., Shapiro,for the formation of somatopically organized connections. J. Comp. L., Huntley, G.W., Benson, D.L., and Colman, D.R. (2000). MolecularNeurol. 407, 453–471. modification of N-cadherin in response to synaptic activity. Neuron
25, 93–107.Ichimura, T., and Hashimoto, P.H. (1988). Structural components inthe synaptic cleft captured by freeze- substitution and deep etching tom Dieck, S., Sanmarti-Vila, L., Langnaese, K., Richter, K., Kindler,of directly frozen cerebellar cortex. J. Neurocytol. 17, 3–12. S., Soyke, A., Wex, H., Smalla, K.H., Kampf, U., Franzer, J.T., et al.
(1998). Bassoon, a novel zinc-finger CAG/glutamine-repeat proteinKennedy, M.B. (1998). Signal transduction molecules at the gluta-selectively localized at the active zone of presynaptic nerve termi-matergic postsynaptic membrane. Brain Res. Brain Res. Rev 26,nals. J. Cell Biol. 142, 499–509.243–257.
Triller, A., and Korn, H. (1985). Activity-dependent deformations ofKennedy, M.B. (2000). Signal-processing machines at the postsyn-presynaptic grids at central synapses. J. Neurocytol. 14, 177–192.aptic density. Science 290, 750–754.
Kennedy, M.B., Bennett, M.K., and Erondu, N.E. (1983). Biochemical Uchida, N., Honjo, Y., Johnson, K.R., Wheelock, M.J., and Takeichi,M. (1996). The catenin/cadherin adhesion system is localized inand immunochemical evidence that the “major postsynaptic density

Reconstitution of the CNS Presynaptic Web77
synaptic junctions bordering transmitter release zones. J. Cell Biol.135, 767–779.
Vincent, V., Goffin, V., Rozakis-Adcock, M., Mornon, J.P., and Kelly,P.A. (1997). Identification of cytoplasmic motifs required for shortprolactin receptor internalization. J. Biol. Chem. 272, 7062–7068.
Walikonis, R.S., Jensen, O.N., Mann, M., Provance, D.W., Jr., Mercer,J.A., and Kennedy, M.B. (2000). Identification of proteins in the post-synaptic density fraction by mass spectrometry. J. Neurosci. 20,4069–4080.
Wang, X., Kibschull, M., Laue, M.M., Lichte, B., Petrasch-Parwez,E., and Kilimann, M.W. (1999). Aczonin, a 550-kD putative scaffoldingprotein of presynaptic active zones, shares homology regions withRim and Bassoon and binds profilin. J. Cell Biol. 147, 151–162.
Wu, Q., and Maniatis, T. (1999). A striking organization of a largefamily of human neural cadherin-like cell adhesion genes. Cell 97,779–790.
Zhai, R.G., Vardinon-Friedman, H., Cases-Langhoff, C., Becker, B.,Gundelfinger, E.D., Ziv, N.E., and Garner, C.C. (2001). Assemblingthe presynaptic active zone. A characterization of an active zoneprecursor vesicle. Neuron 29, 131–143.
Zinsmaier, K.E., and Bronk, P. (2001). Molecular chaperones andthe regulation of neurotransmitter exocytosis. Biochem. Pharmacol.62, 1–11.