6468.full.pdf - the journal of immunology
TRANSCRIPT

of January 13, 2022.This information is current as
T CellsδγUp-Regulation and Augment Cell Division in
αSelect Plant Tannins Induce IL-2R
Mark A. JutilaPatrick McCurley, Pati M. Glee, Aiyappa Palecanda andKerns, Miranda Radke, Jodi F. Hedges, Charles Petrie, Jeff Holderness, Larissa Jackiw, Emily Kimmel, Hannah
http://www.jimmunol.org/content/179/10/6468doi: 10.4049/jimmunol.179.10.6468
2007; 179:6468-6478; ;J Immunol
Referenceshttp://www.jimmunol.org/content/179/10/6468.full#ref-list-1
, 26 of which you can access for free at: cites 72 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 13, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

Select Plant Tannins Induce IL-2R� Up-Regulation andAugment Cell Division in �� T Cells1
Jeff Holderness,* Larissa Jackiw,* Emily Kimmel,* Hannah Kerns,* Miranda Radke,*Jodi F. Hedges,* Charles Petrie,† Patrick McCurley,† Pati M. Glee,† Aiyappa Palecanda,†
and Mark A. Jutila2*
�� T cells are innate immune cells that participate in host responses against many pathogens and cancers. Recently, phospho-antigen-based drugs, capable of expanding �� T cells in vivo, entered clinical trials with the goal of enhancing innate immunesystem functions. Potential shortcomings of these drugs include the induction of nonresponsiveness upon repeated use and theexpansion of only the V�2 subset of human �� T cells. V�1 T cells, the major tissue subset, are unaffected by phosphoantigenagonists. Using FACS-based assays, we screened primary bovine cells for novel �� T cell agonists with activities not encompassedby the current treatments in an effort to realize the full therapeutic potential of �� T cells. We identified �� T cell agonists derivedfrom the condensed tannin fractions of Uncaria tomentosa (Cat’s Claw) and Malus domestica (apple). Based on superior potency,the apple extract was selected for detailed analyses on human cells. The apple extract was a potent agonist for both human V�1and V�2 T cells and NK cells. Additionally, the extract greatly enhanced phosphoantigen-induced �� T cell expansion. Ouranalyses suggest that a tannin-based drug may complement the phosphoantigen-based drugs, thereby enhancing the therapeuticpotential of �� T cells. The Journal of Immunology, 2007, 179: 6468–6478.
T he �� T cell is one of the primary lymphocytes consideredto participate in innate immune responses. They are foundin virtually all portals of entry into the body where they
act as sentinels of the extracellular environment by respondingrapidly to tissue damage (1, 2) and pathogens through recognitionof conserved pathogen-associated molecular patterns and specificAgs (3–5). Human �� T cells can be categorized into two major,functionally distinct populations, V�1 and V�2 T cells, which arecharacterized by their respective TCR � chain usage. Expression ofsurface molecules, such as chemokine receptors (6), varies be-tween the �� T cell subsets, as does their physical locale within thebody. V�2 T cells are primarily found in the blood and lymphaticsystems, whereas V�1 T cells are typically found in tissues, suchas the gut mucosa and skin (7, 8). V�2 T cells are potent antimi-crobial and antitumor effector cells (9–11). V�1 cells have impor-tant immunoregulatory functions and, if properly stimulated, canbecome potent effector cells (12).
Observations that the majority of V�2 T cells proliferate in re-sponse to intermediates of the cholesterol synthesis pathway(phosphoantigens) led to clinical trials testing these compounds as
potential drugs for the treatment of some cancers and infections(13). Other known �� T cell agonists, such as bisphosphonates(14) and alkylamines (15), induce a similar response by increasingthe endogenous concentration of phosphoantigens. Notably, phos-phoantigen-based proliferation requires a costimulatory factor foroptimal V�2 T cell expansion, typically IL-2 or IL-15 (16–18).Though expansion of V�2 T cells in vivo is clearly demonstratedusing these new drugs, the benefit of these expanded cells on hostdefenses is not fully understood due to a minimal or nonexistentresponse in non-primate animal models (19). Additional caveatsfor the clinical use of phosphoantigen-based drugs include the re-quirement for large amounts of agonist (20), induction of rapid cellanergy to treatment (20), and activation of only the V�2 subset,which must then traffic to the tumor or infection to be effective (8,12, 20, 21).
We initiated a drug discovery effort to identify novel �� T cellagonists that overcome the shortcomings of phosphoantigen-baseddrugs. Traditional medicine uses extracts from plants and microbesto treat diseases, many of which have been shown to contain phar-maceutically relevant components. In fact, 47% of the small mol-ecule drugs developed over the last 25 years are either naturalproducts or derived from natural products (22). Phosphostim (In-nate Pharma), a �� T cell agonist in clinical trials, is an exampleof a drug designed from the structure of naturally occurring phos-phoantigens (23, 24). Due to the dominance of pharmaceuticallyrelevant compounds in nature, in an ongoing effort we initiated ascreen of �100,000 natural products, including common nutri-tional supplements, using a FACS-based assay, which measuresbovine �� T cell activation via up-regulation of the IL-2R � chain(IL-2R�). Bovine cells were used in the primary screens because,among other reasons, they do not robustly respond to phosphoan-tigens (25), thus avoiding the detection of these common �� T cellagonists. In this study we report nonphosphoantigen agonist activ-ity that was observed for both bovine and human �� T cells inspecific plant species including extracts of nonripe Malus domestica(apple) peel, Uncaria tomentosa (Cat’s Claw), Angelica sinensis
*Veterinary Molecular Biology, Montana State University, and †LigoCyte Pharma-ceuticals, Inc., Bozeman, MT 59718
Received for publication July 30, 2007. Accepted for publication August 30, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by funding in whole or in part from the National Instituteof Allergy and Infectious Diseases, National Institutes of Health, and by ContractHHSN266200400009/N01-AI40009 from the Department of Health and Human Ser-vices. This work is also supported by Contract 2000-04446 from the U.S. Department ofAgriculture Initiative Future Agriculture and Food Safety, and National Research Initia-tive. LigoCyte Pharmaceuticals, Inc., together with Montana State University, also holdsa National Institutes of Health contract that partially funded this work.2 Address correspondence and reprint requests to Dr. Mark A. Jutila, Veterinary Mo-lecular Biology, Molecular Biosciences Building, Montana State University, 960Technology Boulevard, Bozeman, MT 59718. E-mail address: [email protected]
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

(Dong Quai), and Funtumia elastica (Yamoa). We traced activity inthe apple and Cat’s Claw extracts to the tannin fraction, a class ofpolyphenols characterized by their capacity to bind proteins. Furtheranalysis identified the bioactive population of tannins as condensedtannins. Detailed dose analyses suggested that superior activity ex-isted in the apple extract. In studies on human cells, apple-derivedtannins demonstrated agonist activity for human V�1 and V�2 T cellsas well as NK cells. These studies suggest that unique tannin agonistsfor innate lymphocytes may account, at least in part, for the immu-nomodulatory properties of some plant extracts. They also suggestthat a tannin-based drug may complement the phosphoantigen-baseddrugs, thereby enhancing the therapeutic potential of �� T cells.
Materials and MethodsPreparation of bovine and human PBMC
Whole blood was collected from 1- to 3-mo-old bull Holstein calves intosodium heparin tubes (BD Biosciences) and from healthy human adultdonors with ACD solution B anti-coagulant tubes (BD Biosciences). Leu-kocytes were separated from whole blood using Histopaque 1077 (Sigma-Aldrich) for bovine cells as previously described (26) and for human cellsas per the manufacturer’s instructions. Additionally, bovine RBC were re-moved by hypotonic lysis. All experiments were performed in accordancewith National Institutes of Health guidelines and approved by the Institu-tional Animal Care and Use Committee and Institutional Review Board ofMontana State University (Bozeman, MT).
Preparation of plant extracts
Malus domestica (apple) non-ripe peel (APP3; Apple Poly), Uncaria to-mentosa (Cat’s Claw; Raintree Nutrition), Angelica sinensis (Dong Quai;Nature’s Way Products), and Funtumia elastica (Yamoa; NHC) were typicallysuspended in room temperature water (Cat’s Claw 10 ml/g, Dong Quai 5.0ml/g, apple peel 8.0 ml/g, and Yamoa 3.0 ml/g), although use of near boilingwater had no detectable impact on the preparations, and were agitated for 5min before centrifugation to remove insoluble particles. Extracts were sterilefiltered (0.2 �m) and stored at �80°C until use. Extracts were lyophilized todetermine their approximate dry weights: Cat’s Claw 17.6 mg/ml, APP 84.8mg/ml, Dong Quai 87.0 mg/ml, and Yamoa 32.1 mg/ml.
FACS-based IL-2R� and CFSE analysis of human andbovine PBMC
PBMC preparations were suspended in X-VIVO 15 medium (Cambrex) at1 � 106 cells/ml for bovine cells or 2 � 106 cells/ml for human cells. Forthe measurement of IL-2R� expression, cells were cultured for 48 h at37°C, 10% CO2 in the presence or absence of known or test agonists, andthe expression of IL-2R� measured by multicolor FACS (see below). Tomeasure cell proliferation, bovine or human PBMCs were labeled with 2.5�M CFSE for 5 min in HBSS, washed, and then cultured at 37°C, 10%CO2 in X-VIVO 15 medium in the presence or absence of known or testagonists for 5 days. Human CFSE cultures were typically treated withlow-dose human recombinant IL-15 (PeproTech) at the beginning of theculture period. For these cultures, all samples, including controls, received1.0 ng/ml IL-15. Cell division was quantified by determining the percent-age of cells that had divided at least once, based on a 50% or greaterreduction in CFSE intensity, using multicolor FACS. Positive control ago-nists were Con A (0.5 ng/ml; Sigma-Aldrich) with human recombinantIL-2 (1.0 ng/ml; Peprotech) or LPS (10 �g/ml, Escherichia coli O111:B4;Sigma-Aldrich), and negative controls included appropriate buffers dilutedin the assay medium.
Cells were analyzed by FACS using a FACSCalibur equipped with anHTS loader (BD Biosciences) or a FACSCanto (BD Biosciences). Bovinecells were stained with IL-2R� (LCTB2A; VMRD) and/or �� TCR (GD3.8(27)). Human cells were stained with IL-2R� (MEM-181; Serotec), ��TCR (11F2; BD Biosciences), CD19 (HIB19; BD Biosciences), CD94(HP-3B1; Beckman Coulter), CD56 (B159; BD Biosciences), CD3(UCHT1; Beckman Coulter), V�1 TCR (T58.2; Endogen), and/or V�2TCR (IMMU389; Beckman Coulter) Abs. Abs were directly labeled(FITC, PE, PE-Cy5, PE-Cy7, allophycocyanin, or allophycocyanin-Cy7)or indirectly labeled with goat anti-mouse PE, FITC, or allophycocyanin(Jackson ImmunoResearch Laboratories). Second stage reagents alone were
used to determine the level of background staining in the FACS analyses. Onlylive cells (as determined by FACS light scatter) were included in the analyses.
HPLC analysis of Cat’s Claw
An aqueous extract of Cat’s Claw (1.3 mg in 100 �l) was applied to areverse phase analytical C18 column (4.6 � 250 mm; Grace Vydac) equil-ibrated in 0.1% trifluoroacetic acid (Pierce). The extract was eluted with alinear gradient of 0% CH3CN/0.08% trifluoroacetic acid to 100% CH3CN/0.08% trifluoroacetic acid at a flow rate of 1 ml/min. UV absorbance wasmeasured at a wavelength of 254 nm. Fractions were collected at 1-minintervals, dried (SpeedVac; Savant Instruments), and suspended in PBS.
CD69 gene expression
Bovine CD69 gene expression was determined using the Quantigene assay(Panomics) following the manufacturer’s procedure. Briefly, bovinePBMCs (2 � 106 cells/ml) were cultured with the HPLC separated Cat’sClaw fractions (1/20 dilution) for 4 h in complete RPMI 1640 medium, aspreviously described (3) before lysis with supplied buffer. Lysates wereadded to prepared 96-well plates containing hybridization probes (providedby the manufacturer) for bovine CD69 mRNA. Presence of hybridizedCD69 mRNA was detected with luminescent DNA probes using an LB960Centro (Berthold Technologies). CD69 gene expression was represented bythe fold increase in CD69 signal compared with medium controls.
Separation of tannins from plant extracts
Aqueous extract of Cat’s Claw (10 ml) was treated with 5 ml of polyvi-nylpolypyrrolidone (PVPP; Sigma-Aldrich) slurry in 0.0625 M HCl atroom temperature overnight. The resulting slurry was filtered through a 0.2�m membrane to remove PVPP and PVPP-bound tannins. The eluent wasbrought to a pH of 7.2 with NaOH. The tannin-bound PVPP was washedwith water before treatment with 0.1 M NaOH for 60 min to remove boundtannins. The solution was again filtered to remove PVPP and brought to apH of 7.2 with hydrogen chloride. Removal of tannins from Yamoa, DongQuai, and APP was performed by incubating 1 ml of extract with 100 mgof PVPP for 15 min at 4°C. The unbound fractions were separated from thetannin-bound PVPP by centrifugation. The supernatant fluid was treatedwith an additional 100 mg of PVPP. This process was repeated a total offive times, reserving samples from each round of depletion.
Tannin fractions from APP, Cat’s Claw, and Dong Quai extracts weresubjected to a modified Folin-Ciocalteu assay (28) to measure total phe-nolics. Briefly, 2 ml of each extract was transferred into a bed volume of3 ml of PVPP and incubated for 1 h. The nontannin fraction was collectedby washing with 10 ml of water. Tannins were eluted with 10 ml of 1.0 MNaOH. Extracts and fractions thereof were serially diluted 1/1 to 1/4050 inwater before mixing 500 �l of diluted sample with 250 �l of 1 N Folin-Ciocalteu reagent (Sigma-Aldrich) and 1.25 ml of 20% sodium carbonate.The samples were mixed and incubated at room temperature for 40 minbefore absorbance was read at 725 nm using a DU800 spectrophotometer(Beckman Coulter). Tannic acid (Sigma-Aldrich) dilutions (40–200 �g/ml) were freshly prepared and used as a standard.
To separate condensed tannins by size, 1.5 ml of APP (�150 mg) wasadsorbed to LH-20 resin and washed with 30 ml of water. To elute, 30 mlof 25, 50, 75, and 95% MeOH and 70% acetone were used in successionand the fractions collected. The 95% MeOH and 70% acetone fractionswere collected in two 15-ml fractions. Activity was predominantly found inthe later fractions, therefore these fractions were used for analyses. Allfractions were lyophilized and resuspended in 750 �l of water.
TLR activation and Limulus assays
APP was tested on THP1-Blue CD14 cells (InvivoGen) for TLR agonistactivity, according to the manufacturer’s protocol. THP1-Blue CD14 cellsexpress TLRs 1, 2, 4–8, and 10, overexpress CD14, and are transfectedwith a reporter plasmid containing secreted embryonic alkaline phospha-tase (SEAP) under the control of both an NF-�B and AP-1 inducible pro-moter. TLR activation was determined by quantifying SEAP activity.Briefly, THP1-Blue CD14 cells at a concentration of 2 � 106 cells/ml werecultured in complete RPMI 1640 medium containing 10% FBS, glucose(4.5 mg/ml), zeocin (200 �g/ml), and blasticidin (10 �g/ml) (all fromInvivoGen) followed by PMA (50 ng/ml) treatment for 18 h. PMA wasused to differentiate the THP1 cells to induce expression of TLRs. Cellswere washed to remove residual PMA and the glucose, zeocin, and blas-ticidin treatment was discontinued. Cells were stimulated with LPS (aTLR4 agonist), as a positive control, and different concentrations of APPor Cat’s Claw extract in complete RPMI 1640 medium for 24 h at 37°C and10% CO2. Supernatant fluid was removed and added to Quanti Blue col-orimetric assay reagent for 24 h at 37°C and 10% CO2. After 24 h, samples
3 Abbreviations used in this paper: APP, apple polyphenol; PVPP, polyvinylpolypyr-rolidone; SEAP, secreted embryonic alkaline phosphatase; LAL, Limulus amebocytelysate.
6469The Journal of Immunology
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
were read at an OD of 655 nm by a VERSAmax tunable microplate reader(Molecular Devices). All samples were run in quadruplicate, from whichaverages and SDs were determined.
A kinetic turbidimetric Limulus amebocyte lysate (LAL; Associates ofCape Cod) assay was performed on APP as per the manufacturer’s instruc-tions. Briefly, APP was diluted 1/6 in LAL Reagent water and analyzed forendotoxin using LAL Reagent resuspended in Glucashield buffer. LALagglutination was measured using a ThermoMax Microplate Reader (Mo-lecular Devices). Samples were analyzed in triplicate.
Protease digestion of tannin-associated proteins
APP (42.4 mg) was treated with PVPP (50 mg) and either PBS, chymo-trypsin (20 U; Sigma-Aldrich), or trypsin (50,000U; Sigma-Aldrich) for 1 hat 37°C. PVPP was washed, and the tannins eluted with sodium hydroxide.
Alkaline phosphatase inactivation of phosphoantigens
APP extract was passed over a column of polyvinyl spheres containingimmobilized alkaline phosphatase (MoBiTec). The phosphoantigen HD-MAPP (1-hydroxy-2-methyl-2-buten-4-yl 4-diphosphate; Echelon) or iso-pentenyl pyrophosphate (Sigma-Aldrich) (both at 500 �g/ml) or APP (42.4mg/ml) were equilibrated with reaction buffer (final concentrations: 25 mMTris-HCl, 0.05 mM ZnCl2, 0.5 mM MgCl2 (pH 8.0)) and passed over thecolumn twice. Control samples for were equilibrated with reaction bufferbut not passed over the column.
Cell separation using magnetic beads and flow cytometry
PBMCs from bovine and human donors were sorted to create purified cellpopulations via magnetic bead separation (Miltenyi Biotec), as per themanufacturer’s instructions. Monocytes from bovine and human PBMCswere depleted with magnetic beads conjugated to CD14 mAb using an LDcolumn (Miltenyi Biotec). Purification of human �� T cells was achievedusing either positive or negative sorting techniques. Briefly, human PB-MCs were treated with mAbs either to the �� TCR or to a mixture of mAbsnot expressed on �� T cells (positive and negative sorting, respectively;Miltenyi Biotec). The labeled cells were then linked to magnetic beads andtransferred through LS columns (Miltenyi Biotec). The �� T cells werethen collected as the column bound or unbound fractions (positive andnegatively sorted �� T cells, respectively). Bovine �� T cells were sortedfrom PBMC by labeling with �� TCR mAb (GD3.8) and magnetic beadsconjugated with anti-mouse Abs (Miltenyi Biotec) as per the manufactur-er’s instructions or by using a FACSVantage SE (BD Biosciences) as pre-viously described (26). All sorted cells were cultured under the conditionsnoted for PBMC cultures, except sorted human �� T cell cultures, whichwere cultured between 1 � 105 and 5 � 105 cells/ml.
Purities of magnetically sorted cells were verified by FACS before cul-ture. CD14 depletion was confirmed as �0.5% monocyte contaminationusing CD14 mAbs (human, UCH-M1; Santa Cruz Biotechnology or bo-vine, CAM36A; VMRD). �� T cell purity was determined using �� TCRmAbs (human, 11F2 or bovine, GD3.8).
ResultsExtracts from common dietary plant supplements induceIL-2R� expression and cell division from �� T cells in PBMCpreparations
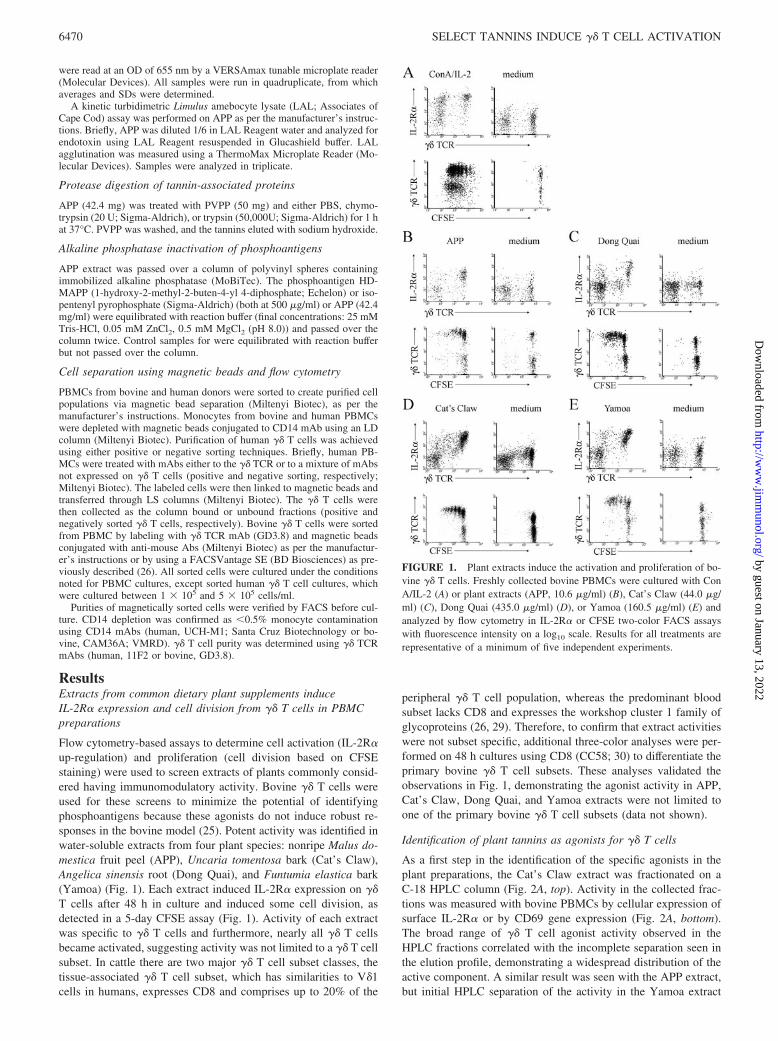
Flow cytometry-based assays to determine cell activation (IL-2R�up-regulation) and proliferation (cell division based on CFSEstaining) were used to screen extracts of plants commonly consid-ered having immunomodulatory activity. Bovine �� T cells wereused for these screens to minimize the potential of identifyingphosphoantigens because these agonists do not induce robust re-sponses in the bovine model (25). Potent activity was identified inwater-soluble extracts from four plant species: nonripe Malus do-mestica fruit peel (APP), Uncaria tomentosa bark (Cat’s Claw),Angelica sinensis root (Dong Quai), and Funtumia elastica bark(Yamoa) (Fig. 1). Each extract induced IL-2R� expression on ��T cells after 48 h in culture and induced some cell division, asdetected in a 5-day CFSE assay (Fig. 1). Activity of each extractwas specific to �� T cells and furthermore, nearly all �� T cellsbecame activated, suggesting activity was not limited to a �� T cellsubset. In cattle there are two major �� T cell subset classes, thetissue-associated �� T cell subset, which has similarities to V�1cells in humans, expresses CD8 and comprises up to 20% of the
peripheral �� T cell population, whereas the predominant bloodsubset lacks CD8 and expresses the workshop cluster 1 family ofglycoproteins (26, 29). Therefore, to confirm that extract activitieswere not subset specific, additional three-color analyses were per-formed on 48 h cultures using CD8 (CC58; 30) to differentiate theprimary bovine �� T cell subsets. These analyses validated theobservations in Fig. 1, demonstrating the agonist activity in APP,Cat’s Claw, Dong Quai, and Yamoa extracts were not limited toone of the primary bovine �� T cell subsets (data not shown).
Identification of plant tannins as agonists for �� T cells
As a first step in the identification of the specific agonists in theplant preparations, the Cat’s Claw extract was fractionated on aC-18 HPLC column (Fig. 2A, top). Activity in the collected frac-tions was measured with bovine PBMCs by cellular expression ofsurface IL-2R� or by CD69 gene expression (Fig. 2A, bottom).The broad range of �� T cell agonist activity observed in theHPLC fractions correlated with the incomplete separation seen inthe elution profile, demonstrating a widespread distribution of theactive component. A similar result was seen with the APP extract,but initial HPLC separation of the activity in the Yamoa extract
FIGURE 1. Plant extracts induce the activation and proliferation of bo-vine �� T cells. Freshly collected bovine PBMCs were cultured with ConA/IL-2 (A) or plant extracts (APP, 10.6 �g/ml) (B), Cat’s Claw (44.0 �g/ml) (C), Dong Quai (435.0 �g/ml) (D), or Yamoa (160.5 �g/ml) (E) andanalyzed by flow cytometry in IL-2R� or CFSE two-color FACS assayswith fluorescence intensity on a log10 scale. Results for all treatments arerepresentative of a minimum of five independent experiments.
6470 SELECT TANNINS INDUCE �� T CELL ACTIVATION
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
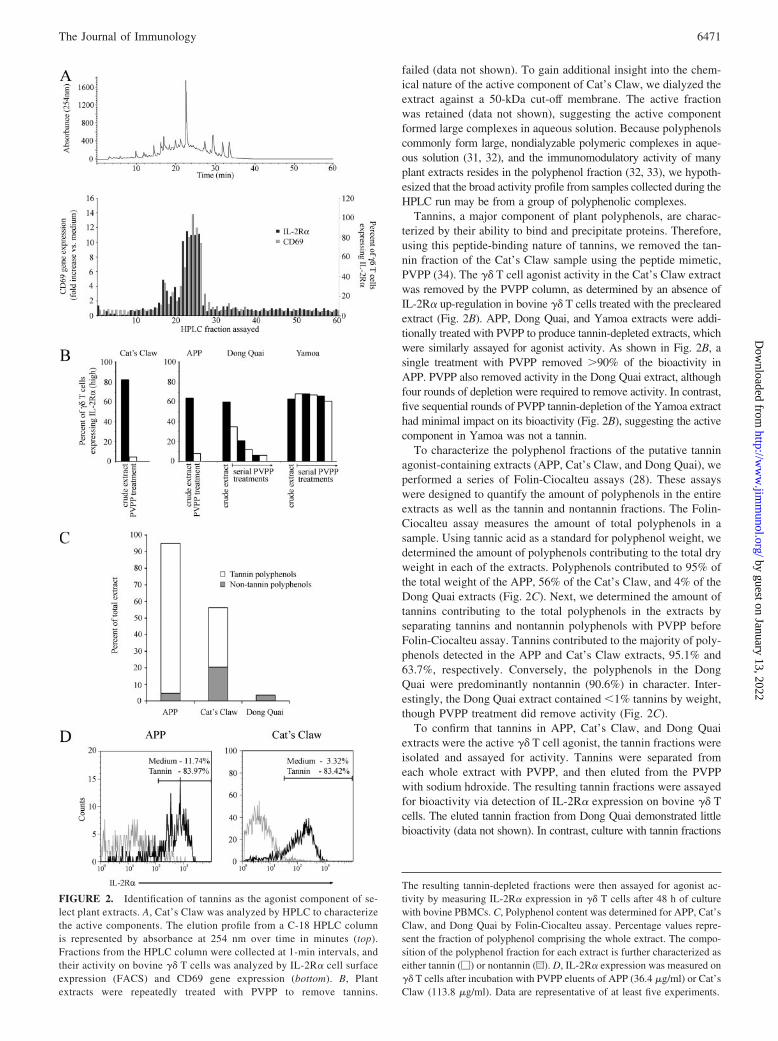
failed (data not shown). To gain additional insight into the chem-ical nature of the active component of Cat’s Claw, we dialyzed theextract against a 50-kDa cut-off membrane. The active fractionwas retained (data not shown), suggesting the active componentformed large complexes in aqueous solution. Because polyphenolscommonly form large, nondialyzable polymeric complexes in aque-ous solution (31, 32), and the immunomodulatory activity of manyplant extracts resides in the polyphenol fraction (32, 33), we hypoth-esized that the broad activity profile from samples collected during theHPLC run may be from a group of polyphenolic complexes.
Tannins, a major component of plant polyphenols, are charac-terized by their ability to bind and precipitate proteins. Therefore,using this peptide-binding nature of tannins, we removed the tan-nin fraction of the Cat’s Claw sample using the peptide mimetic,PVPP (34). The �� T cell agonist activity in the Cat’s Claw extractwas removed by the PVPP column, as determined by an absence ofIL-2R� up-regulation in bovine �� T cells treated with the preclearedextract (Fig. 2B). APP, Dong Quai, and Yamoa extracts were addi-tionally treated with PVPP to produce tannin-depleted extracts, whichwere similarly assayed for agonist activity. As shown in Fig. 2B, asingle treatment with PVPP removed �90% of the bioactivity inAPP. PVPP also removed activity in the Dong Quai extract, althoughfour rounds of depletion were required to remove activity. In contrast,five sequential rounds of PVPP tannin-depletion of the Yamoa extracthad minimal impact on its bioactivity (Fig. 2B), suggesting the activecomponent in Yamoa was not a tannin.
To characterize the polyphenol fractions of the putative tanninagonist-containing extracts (APP, Cat’s Claw, and Dong Quai), weperformed a series of Folin-Ciocalteu assays (28). These assayswere designed to quantify the amount of polyphenols in the entireextracts as well as the tannin and nontannin fractions. The Folin-Ciocalteu assay measures the amount of total polyphenols in asample. Using tannic acid as a standard for polyphenol weight, wedetermined the amount of polyphenols contributing to the total dryweight in each of the extracts. Polyphenols contributed to 95% ofthe total weight of the APP, 56% of the Cat’s Claw, and 4% of theDong Quai extracts (Fig. 2C). Next, we determined the amount oftannins contributing to the total polyphenols in the extracts byseparating tannins and nontannin polyphenols with PVPP beforeFolin-Ciocalteu assay. Tannins contributed to the majority of poly-phenols detected in the APP and Cat’s Claw extracts, 95.1% and63.7%, respectively. Conversely, the polyphenols in the DongQuai were predominantly nontannin (90.6%) in character. Inter-estingly, the Dong Quai extract contained �1% tannins by weight,though PVPP treatment did remove activity (Fig. 2C).
To confirm that tannins in APP, Cat’s Claw, and Dong Quaiextracts were the active �� T cell agonist, the tannin fractions wereisolated and assayed for activity. Tannins were separated fromeach whole extract with PVPP, and then eluted from the PVPPwith sodium hdroxide. The resulting tannin fractions were assayedfor bioactivity via detection of IL-2R� expression on bovine �� Tcells. The eluted tannin fraction from Dong Quai demonstrated littlebioactivity (data not shown). In contrast, culture with tannin fractions
FIGURE 2. Identification of tannins as the agonist component of se-lect plant extracts. A, Cat’s Claw was analyzed by HPLC to characterizethe active components. The elution profile from a C-18 HPLC columnis represented by absorbance at 254 nm over time in minutes (top).Fractions from the HPLC column were collected at 1-min intervals, andtheir activity on bovine �� T cells was analyzed by IL-2R� cell surfaceexpression (FACS) and CD69 gene expression (bottom). B, Plantextracts were repeatedly treated with PVPP to remove tannins.
The resulting tannin-depleted fractions were then assayed for agonist ac-tivity by measuring IL-2R� expression in �� T cells after 48 h of culturewith bovine PBMCs. C, Polyphenol content was determined for APP, Cat’sClaw, and Dong Quai by Folin-Ciocalteu assay. Percentage values repre-sent the fraction of polyphenol comprising the whole extract. The compo-sition of the polyphenol fraction for each extract is further characterized aseither tannin (�) or nontannin (u). D, IL-2R� expression was measured on�� T cells after incubation with PVPP eluents of APP (36.4 �g/ml) or Cat’sClaw (113.8 �g/ml). Data are representative of at least five experiments.
6471The Journal of Immunology
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
isolated from Cat’s Claw and APP up-regulated IL-2R� expressionon �� T cells (Fig. 2D), thereby verifying that the �� T cell agonist inthese two extracts was a tannin. Analysis of the PVPP-precipitatedtannin fraction via HPLC resulted in a broad elution profile similar tothat shown in Fig. 2A (data not shown), confirming the heterogeneousnature of the tannin fraction in these extracts.
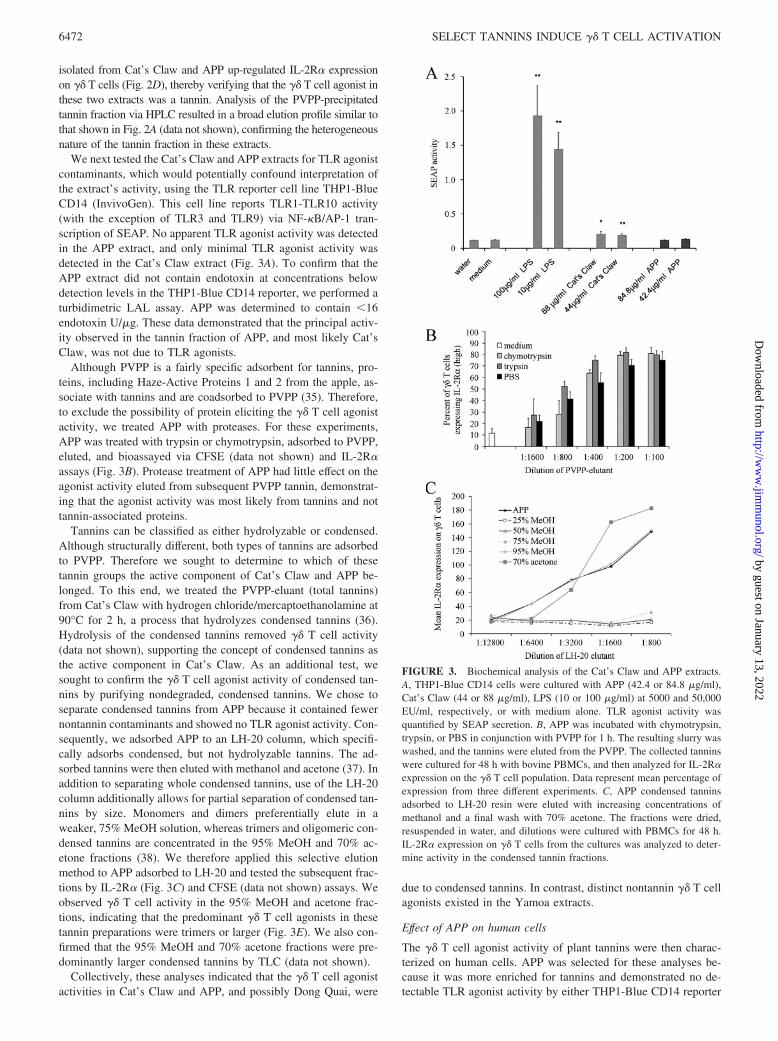
We next tested the Cat’s Claw and APP extracts for TLR agonistcontaminants, which would potentially confound interpretation ofthe extract’s activity, using the TLR reporter cell line THP1-BlueCD14 (InvivoGen). This cell line reports TLR1-TLR10 activity(with the exception of TLR3 and TLR9) via NF-�B/AP-1 tran-scription of SEAP. No apparent TLR agonist activity was detectedin the APP extract, and only minimal TLR agonist activity wasdetected in the Cat’s Claw extract (Fig. 3A). To confirm that theAPP extract did not contain endotoxin at concentrations belowdetection levels in the THP1-Blue CD14 reporter, we performed aturbidimetric LAL assay. APP was determined to contain �16endotoxin U/�g. These data demonstrated that the principal activ-ity observed in the tannin fraction of APP, and most likely Cat’sClaw, was not due to TLR agonists.
Although PVPP is a fairly specific adsorbent for tannins, pro-teins, including Haze-Active Proteins 1 and 2 from the apple, as-sociate with tannins and are coadsorbed to PVPP (35). Therefore,to exclude the possibility of protein eliciting the �� T cell agonistactivity, we treated APP with proteases. For these experiments,APP was treated with trypsin or chymotrypsin, adsorbed to PVPP,eluted, and bioassayed via CFSE (data not shown) and IL-2R�assays (Fig. 3B). Protease treatment of APP had little effect on theagonist activity eluted from subsequent PVPP tannin, demonstrat-ing that the agonist activity was most likely from tannins and nottannin-associated proteins.
Tannins can be classified as either hydrolyzable or condensed.Although structurally different, both types of tannins are adsorbedto PVPP. Therefore we sought to determine to which of thesetannin groups the active component of Cat’s Claw and APP be-longed. To this end, we treated the PVPP-eluant (total tannins)from Cat’s Claw with hydrogen chloride/mercaptoethanolamine at90°C for 2 h, a process that hydrolyzes condensed tannins (36).Hydrolysis of the condensed tannins removed �� T cell activity(data not shown), supporting the concept of condensed tannins asthe active component in Cat’s Claw. As an additional test, wesought to confirm the �� T cell agonist activity of condensed tan-nins by purifying nondegraded, condensed tannins. We chose toseparate condensed tannins from APP because it contained fewernontannin contaminants and showed no TLR agonist activity. Con-sequently, we adsorbed APP to an LH-20 column, which specifi-cally adsorbs condensed, but not hydrolyzable tannins. The ad-sorbed tannins were then eluted with methanol and acetone (37). Inaddition to separating whole condensed tannins, use of the LH-20column additionally allows for partial separation of condensed tan-nins by size. Monomers and dimers preferentially elute in aweaker, 75% MeOH solution, whereas trimers and oligomeric con-densed tannins are concentrated in the 95% MeOH and 70% ac-etone fractions (38). We therefore applied this selective elutionmethod to APP adsorbed to LH-20 and tested the subsequent frac-tions by IL-2R� (Fig. 3C) and CFSE (data not shown) assays. Weobserved �� T cell activity in the 95% MeOH and acetone frac-tions, indicating that the predominant �� T cell agonists in thesetannin preparations were trimers or larger (Fig. 3E). We also con-firmed that the 95% MeOH and 70% acetone fractions were pre-dominantly larger condensed tannins by TLC (data not shown).
Collectively, these analyses indicated that the �� T cell agonistactivities in Cat’s Claw and APP, and possibly Dong Quai, were
due to condensed tannins. In contrast, distinct nontannin �� T cellagonists existed in the Yamoa extracts.
Effect of APP on human cells
The �� T cell agonist activity of plant tannins were then charac-terized on human cells. APP was selected for these analyses be-cause it was more enriched for tannins and demonstrated no de-tectable TLR agonist activity by either THP1-Blue CD14 reporter
FIGURE 3. Biochemical analysis of the Cat’s Claw and APP extracts.A, THP1-Blue CD14 cells were cultured with APP (42.4 or 84.8 �g/ml),Cat’s Claw (44 or 88 �g/ml), LPS (10 or 100 �g/ml) at 5000 and 50,000EU/ml, respectively, or with medium alone. TLR agonist activity wasquantified by SEAP secretion. B, APP was incubated with chymotrypsin,trypsin, or PBS in conjunction with PVPP for 1 h. The resulting slurry waswashed, and the tannins were eluted from the PVPP. The collected tanninswere cultured for 48 h with bovine PBMCs, and then analyzed for IL-2R�expression on the �� T cell population. Data represent mean percentage ofexpression from three different experiments. C, APP condensed tanninsadsorbed to LH-20 resin were eluted with increasing concentrations ofmethanol and a final wash with 70% acetone. The fractions were dried,resuspended in water, and dilutions were cultured with PBMCs for 48 h.IL-2R� expression on �� T cells from the cultures was analyzed to deter-mine activity in the condensed tannin fractions.
6472 SELECT TANNINS INDUCE �� T CELL ACTIVATION
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
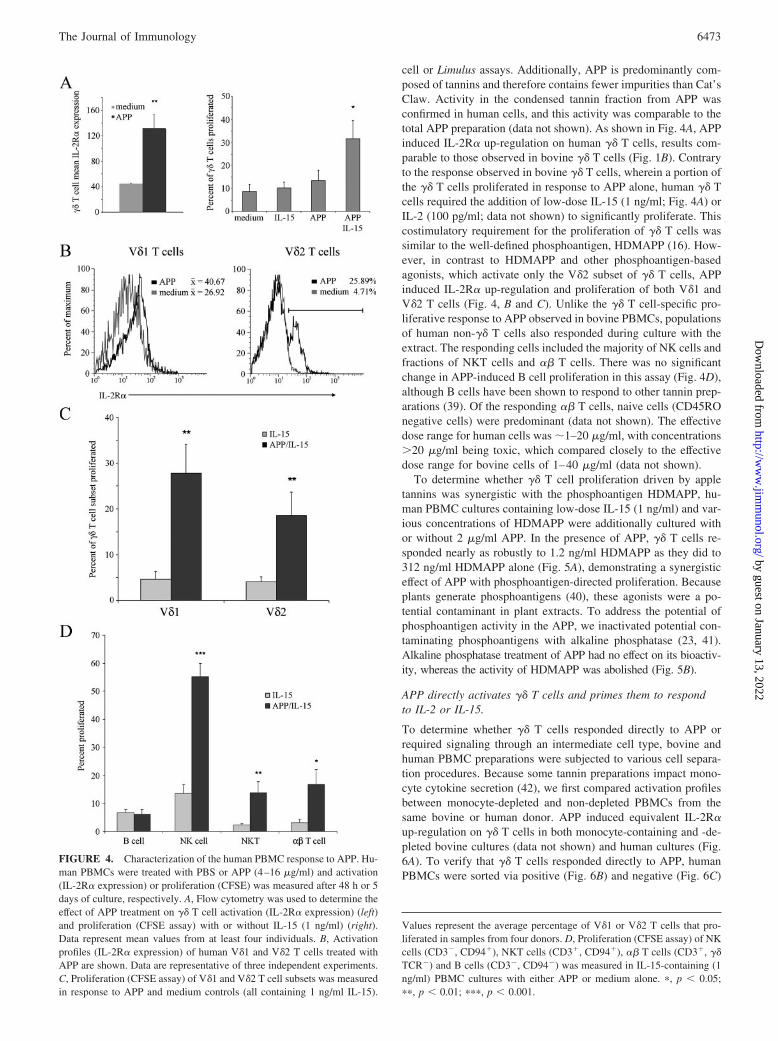
cell or Limulus assays. Additionally, APP is predominantly com-posed of tannins and therefore contains fewer impurities than Cat’sClaw. Activity in the condensed tannin fraction from APP wasconfirmed in human cells, and this activity was comparable to thetotal APP preparation (data not shown). As shown in Fig. 4A, APPinduced IL-2R� up-regulation on human �� T cells, results com-parable to those observed in bovine �� T cells (Fig. 1B). Contraryto the response observed in bovine �� T cells, wherein a portion ofthe �� T cells proliferated in response to APP alone, human �� Tcells required the addition of low-dose IL-15 (1 ng/ml; Fig. 4A) orIL-2 (100 pg/ml; data not shown) to significantly proliferate. Thiscostimulatory requirement for the proliferation of �� T cells wassimilar to the well-defined phosphoantigen, HDMAPP (16). How-ever, in contrast to HDMAPP and other phosphoantigen-basedagonists, which activate only the V�2 subset of �� T cells, APPinduced IL-2R� up-regulation and proliferation of both V�1 andV�2 T cells (Fig. 4, B and C). Unlike the �� T cell-specific pro-liferative response to APP observed in bovine PBMCs, populationsof human non-�� T cells also responded during culture with theextract. The responding cells included the majority of NK cells andfractions of NKT cells and �� T cells. There was no significantchange in APP-induced B cell proliferation in this assay (Fig. 4D),although B cells have been shown to respond to other tannin prep-arations (39). Of the responding �� T cells, naive cells (CD45ROnegative cells) were predominant (data not shown). The effectivedose range for human cells was �1–20 �g/ml, with concentrations�20 �g/ml being toxic, which compared closely to the effectivedose range for bovine cells of 1–40 �g/ml (data not shown).
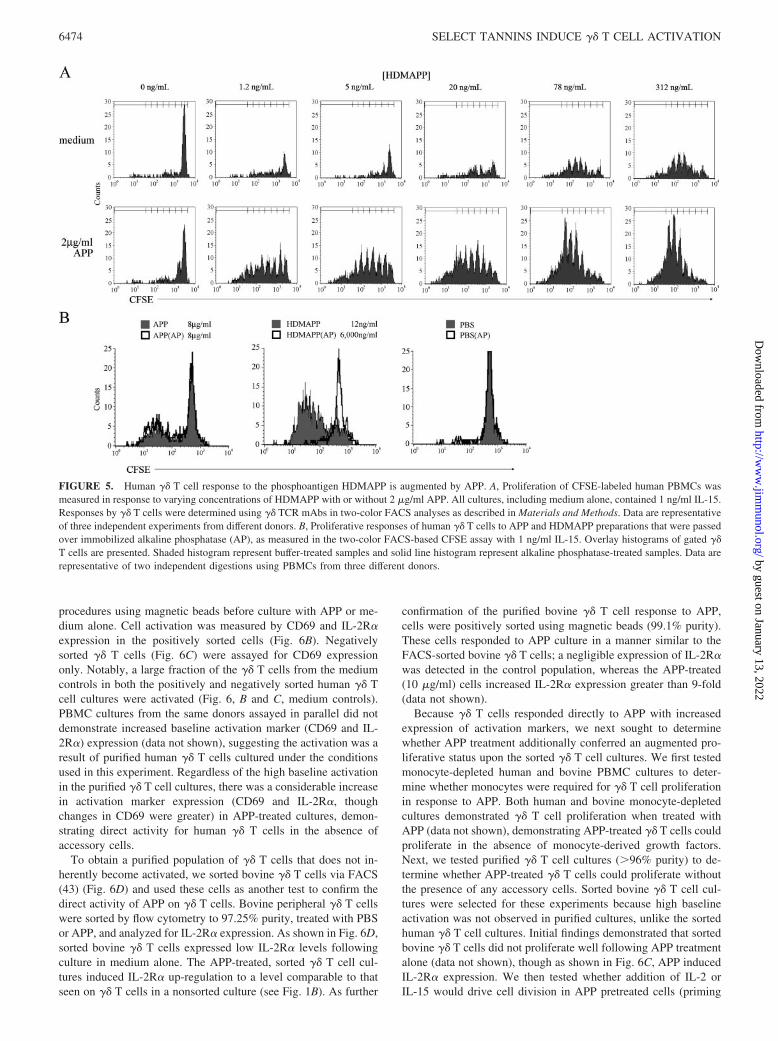
To determine whether �� T cell proliferation driven by appletannins was synergistic with the phosphoantigen HDMAPP, hu-man PBMC cultures containing low-dose IL-15 (1 ng/ml) and var-ious concentrations of HDMAPP were additionally cultured withor without 2 �g/ml APP. In the presence of APP, �� T cells re-sponded nearly as robustly to 1.2 ng/ml HDMAPP as they did to312 ng/ml HDMAPP alone (Fig. 5A), demonstrating a synergisticeffect of APP with phosphoantigen-directed proliferation. Becauseplants generate phosphoantigens (40), these agonists were a po-tential contaminant in plant extracts. To address the potential ofphosphoantigen activity in the APP, we inactivated potential con-taminating phosphoantigens with alkaline phosphatase (23, 41).Alkaline phosphatase treatment of APP had no effect on its bioactiv-ity, whereas the activity of HDMAPP was abolished (Fig. 5B).
APP directly activates �� T cells and primes them to respondto IL-2 or IL-15.
To determine whether �� T cells responded directly to APP orrequired signaling through an intermediate cell type, bovine andhuman PBMC preparations were subjected to various cell separa-tion procedures. Because some tannin preparations impact mono-cyte cytokine secretion (42), we first compared activation profilesbetween monocyte-depleted and non-depleted PBMCs from thesame bovine or human donor. APP induced equivalent IL-2R�up-regulation on �� T cells in both monocyte-containing and -de-pleted bovine cultures (data not shown) and human cultures (Fig.6A). To verify that �� T cells responded directly to APP, humanPBMCs were sorted via positive (Fig. 6B) and negative (Fig. 6C)
FIGURE 4. Characterization of the human PBMC response to APP. Hu-man PBMCs were treated with PBS or APP (4–16 �g/ml) and activation(IL-2R� expression) or proliferation (CFSE) was measured after 48 h or 5days of culture, respectively. A, Flow cytometry was used to determine theeffect of APP treatment on �� T cell activation (IL-2R� expression) (left)and proliferation (CFSE assay) with or without IL-15 (1 ng/ml) (right).Data represent mean values from at least four individuals. B, Activationprofiles (IL-2R� expression) of human V�1 and V�2 T cells treated withAPP are shown. Data are representative of three independent experiments.C, Proliferation (CFSE assay) of V�1 and V�2 T cell subsets was measuredin response to APP and medium controls (all containing 1 ng/ml IL-15).
Values represent the average percentage of V�1 or V�2 T cells that pro-liferated in samples from four donors. D, Proliferation (CFSE assay) of NKcells (CD3�, CD94�), NKT cells (CD3�, CD94�), �� T cells (CD3�, ��TCR�) and B cells (CD3�, CD94�) was measured in IL-15-containing (1ng/ml) PBMC cultures with either APP or medium alone. �, p � 0.05;��, p � 0.01; ���, p � 0.001.
6473The Journal of Immunology
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
procedures using magnetic beads before culture with APP or me-dium alone. Cell activation was measured by CD69 and IL-2R�expression in the positively sorted cells (Fig. 6B). Negativelysorted �� T cells (Fig. 6C) were assayed for CD69 expressiononly. Notably, a large fraction of the �� T cells from the mediumcontrols in both the positively and negatively sorted human �� Tcell cultures were activated (Fig. 6, B and C, medium controls).PBMC cultures from the same donors assayed in parallel did notdemonstrate increased baseline activation marker (CD69 and IL-2R�) expression (data not shown), suggesting the activation was aresult of purified human �� T cells cultured under the conditionsused in this experiment. Regardless of the high baseline activationin the purified �� T cell cultures, there was a considerable increasein activation marker expression (CD69 and IL-2R�, thoughchanges in CD69 were greater) in APP-treated cultures, demon-strating direct activity for human �� T cells in the absence ofaccessory cells.
To obtain a purified population of �� T cells that does not in-herently become activated, we sorted bovine �� T cells via FACS(43) (Fig. 6D) and used these cells as another test to confirm thedirect activity of APP on �� T cells. Bovine peripheral �� T cellswere sorted by flow cytometry to 97.25% purity, treated with PBSor APP, and analyzed for IL-2R� expression. As shown in Fig. 6D,sorted bovine �� T cells expressed low IL-2R� levels followingculture in medium alone. The APP-treated, sorted �� T cell cul-tures induced IL-2R� up-regulation to a level comparable to thatseen on �� T cells in a nonsorted culture (see Fig. 1B). As further
confirmation of the purified bovine �� T cell response to APP,cells were positively sorted using magnetic beads (99.1% purity).These cells responded to APP culture in a manner similar to theFACS-sorted bovine �� T cells; a negligible expression of IL-2R�was detected in the control population, whereas the APP-treated(10 �g/ml) cells increased IL-2R� expression greater than 9-fold(data not shown).
Because �� T cells responded directly to APP with increasedexpression of activation markers, we next sought to determinewhether APP treatment additionally conferred an augmented pro-liferative status upon the sorted �� T cell cultures. We first testedmonocyte-depleted human and bovine PBMC cultures to deter-mine whether monocytes were required for �� T cell proliferationin response to APP. Both human and bovine monocyte-depletedcultures demonstrated �� T cell proliferation when treated withAPP (data not shown), demonstrating APP-treated �� T cells couldproliferate in the absence of monocyte-derived growth factors.Next, we tested purified �� T cell cultures (�96% purity) to de-termine whether APP-treated �� T cells could proliferate withoutthe presence of any accessory cells. Sorted bovine �� T cell cul-tures were selected for these experiments because high baselineactivation was not observed in purified cultures, unlike the sortedhuman �� T cell cultures. Initial findings demonstrated that sortedbovine �� T cells did not proliferate well following APP treatmentalone (data not shown), though as shown in Fig. 6C, APP inducedIL-2R� expression. We then tested whether addition of IL-2 orIL-15 would drive cell division in APP pretreated cells (priming
FIGURE 5. Human �� T cell response to the phosphoantigen HDMAPP is augmented by APP. A, Proliferation of CFSE-labeled human PBMCs wasmeasured in response to varying concentrations of HDMAPP with or without 2 �g/ml APP. All cultures, including medium alone, contained 1 ng/ml IL-15.Responses by �� T cells were determined using �� TCR mAbs in two-color FACS analyses as described in Materials and Methods. Data are representativeof three independent experiments from different donors. B, Proliferative responses of human �� T cells to APP and HDMAPP preparations that were passedover immobilized alkaline phosphatase (AP), as measured in the two-color FACS-based CFSE assay with 1 ng/ml IL-15. Overlay histograms of gated ��T cells are presented. Shaded histogram represent buffer-treated samples and solid line histogram represent alkaline phosphatase-treated samples. Data arerepresentative of two independent digestions using PBMCs from three different donors.
6474 SELECT TANNINS INDUCE �� T CELL ACTIVATION
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

assay). To this end, we cultured FACS-sorted, CFSE-labeled bo-vine �� T cells for 48 h in culture medium containing either PBSor APP (priming event), washed, and then replaced the mediumwith either IL-2 (1 ng/ml) or IL-15 (10 ng/ml). After an additional3 days of culture with the cytokines, the amount of cell division inAPP-primed or PBS-treated cells was compared via FACS analy-sis. As shown in Fig. 7, APP-primed �� T cells from three differentanimals responded robustly (77–94% responding cells) to eitherIL-2 or IL-15, whereas the PBS-primed cells showed minimal re-sponse to either cytokine (18–32% responding cells). Thus, APPdirectly primed �� T cells to proliferate in response to IL-2 orIL-15 in the absence of accessory cells.
Discussion�� T cells are one of four lymphocyte populations thought to beimportant in innate immunity, with the other three being NK cells,NKT cells, and B1 B cells. Recently, clinical approaches weredeveloped to expand the number and/or function of human �� Tcells as a potential therapeutic modality for some cancers (44) andto increase innate resistance to infection (7, 10). The current phos-phoantigen “drugs” expand the major �� T cell subset found inblood (V�2), which display potent antitumor cell activity and im-mune function (13, 45, 46). However, these drugs have no effect onthe tissue-predominant human �� T cell subset (V�1) (47). Thus itis unclear whether phosphoantigen-based drugs will induce thedesired effect within tissues, such as the gut mucosa, the skin, andother portals of pathogen entry into the body (8, 12). In this studywe show that condensed tannins derived from the unripe peel ofthe apple fruit are agonists for both human V�1 and V�2 T cells,leading to increased expression of IL-2R� and cell proliferation.The condensed tannins were active on both human and bovine ��T cells but showed broader leukocyte subset specificity with hu-man cells. In particular, the human NK cell population was largelyresponsive to the condensed tannins. Of the two major human ��T cell subsets, a greater percentage of V�1 T cells responded to thecondensed tannins, suggesting that this tissue population might be
FIGURE 6. APP induces �� T cell activation in the absence of ac-cessory cells. Human and bovine �� T cells were sorted to varyingdegrees of cell purity and cultured with APP or medium alone for 48 hbefore FACS analysis of activation marker expression on �� T cells. A,Monocyte-depleted and nondepleted human PBMCs from four donorswere incubated with medium control or APP, and the effect on IL-2R�expression was determined by FACS. Data represent the concentrationof APP inducing maximal activation (4 –16 �g/ml). �, p � 0.05 or���, p � 0.001. B, Positively sorted �� T cells (91.14%, top, and83.63%, bottom) obtained from different donors were plated with eitherAPP (12 �g/ml) or medium alone and then were analyzed by FACS forCD69 and IL-2R� expression. C, Human �� T cells were collected bynegative sort (94.18%) and then cultured with 14 �g/ml APP or mediumalone before analysis for CD69 expression. D, IL-2R� expression wasmeasured on sorted bovine �� T cells (97.25%) after culture with me-dium control or APP (31.3 �g/ml).
FIGURE 7. APP-primed, purified �� T cell cultures proliferate in re-sponse to IL-2 or IL-15. Sorted bovine �� T cells were labeled with CFSEand cultured with APP (25 �g/ml) or PBS in culture medium. After 48 h,the medium was replaced with fresh medium containing IL-2 or IL-15, andthen cells were cultured for an additional 72–96 h before FACS analysis.All cultures were plated in triplicate. p � 0.001 for all experiments ascalculated from the ratio of proliferating to nonproliferating �� T cells inboth of the PBS- and APP-treated controls. Data displayed are pooled fromthe triplicates collected for each treatment and calf.
6475The Journal of Immunology
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

more responsive to a tannin-based drug preparation in vivo.Though APP was not particularly effective in driving V�2 T cellproliferation unaided, the extract greatly augmented (nearly 300-fold) the proliferative responses of these cells to phosphoantigensand therefore has the potential to be used in combination withphosphoantigens for clinical therapy.
Studies in animals show tissue �� T cells not only serve aseffector cells, but also participate in immunoregulation (7), tissuehomeostasis (2), and epithelial wound repair (48). Increasing thesefunctions would directly and indirectly enhance host innate anddownstream adaptive immune responses. In this study, we foundthat the tissue-associated �� T cells in both human (V�1 T cells)and bovine (CD8� �� T cells) respond to APP. Likewise, Akiyamaet al. (49) show that mouse mucosal �� T cells expand in responseto apple-condensed tannins in vivo, which we have confirmed within vitro assays (M.A. Jutila, unpublished results). Because theseresponses to tannins are conserved in various species, tannin rec-ognition may represent an ancient mechanism used by these cellsto sense and respond to their environment. One would expect planttannin-based agonists to be variably represented in the diet of eachof these animals, including humans. Ruminants are exposed to avery high load of plant derived tannins and this may account for,in part, the very large numbers of �� T cells seen in these animals.
The best-defined agonists that drive �� T cell expansion arephosphoantigens, but they are specific to human and non-humanprimate cells (19) making in vivo studies difficult. Limited studiesin non-human primates demonstrate proof-of-principle that �� Tcells expand in vivo (20), but efficacy studies have not been re-ported in these animals. To circumvent the difficulties inherent inusing non-human primates for in vivo studies, efficacy studies havebeen performed in SCID mice reconstituted with phosphoantigen-expanded human V�2 T cells. These studies demonstrate phos-phoantigen-expanded V�2 T cell effector function against tumors(45) and bacteria (9). However, challenge studies in immunocom-petent animals have not been performed. Until these studies areperformed, understanding the effectiveness of phosphoantigen-ex-panded �� T cells in a clinical setting will remain unresolved.
Because the �� T cell response to condensed tannins is conservedin a wide range of species, there are multiple models available to testthe in vivo efficacy of these condensed tannins in infectious disease,wound repair/healing, and cancers. Limited work already publishedon the extracts examined in this study support the likelihood for suc-cess in these experiments. For instance, Cat’s Claw is a traditionalSouth American medicine used to treat a variety of illnesses includingallergies, cancer, colitis, dysentery, and urinary tract infection (50).Interestingly, these illnesses are commonly associated with �� T cellregulatory activities (7, 51–54), suggesting the described immunos-timulatory properties of the Cat’s Claw plant could be partially due tothe activation of �� T cells. Preparations containing the apple-con-densed tannins we described are also recognized for similar immu-nostimulatory properties (49, 55–57). These observations on the im-munological effects of Cat’s Claw and apple tannins support theprospect for condensed tannins to be used as a therapy to increase ��T cell function in innate immunity and wound healing.
The results of this study may also provide another explanationfor the effect of tea consumption on human �� T cell activation.Kamath et al. (58) demonstrate an in vivo �� T cell priming eventcorresponding to tea intake. After 14 days of tea consumption, thestudy shows that �� T cells become more responsive to in vitrophosphoantigen-based stimulation. They attribute the effect of teato L-theonine, which is converted to alkylamines by the liver. Thealkylamines then purportedly drive the expansion/priming of thephosphoantigen-responsive �� T cells. However, tea is a richsource of immunomodulatory tannins, and we have found that hot
water extracts of black tea induce bovine �� T cell responses(M.A. Jutila, unpublished observations). The tannin fraction of teamay prime human �� T cells, in a manner akin to the APP responsedescribed in this experiment (Fig. 5), making them more respon-sive to in vitro phosphoantigen stimulation. Thus, tannins derivedfrom tea may account, at least in part, for the priming effects on ��T cells observed in vivo following tea consumption.
A major implication of these observations is that a tannin-baseddrug may enhance the effectiveness of current phosphoantigen-based drugs and other approaches to increase innate responses bylymphocytes. Enhancement of phosphoantigen responses may beof particular benefit to individuals afflicted with hyporesponsiveV�2 populations due to either repeat phosphoantigen treatment(20) or persistent infection (59–61). As shown in Fig. 5A, APPimproves the proliferation of HDMAPP-stimulated �� T cells.Therefore, in these settings, it may be possible to increase thenumber of expanding V�2 cells by coadministering a tannin prep-aration. The effects of a tannin agonist would likely extend to otherhuman lymphocytes as well. As shown in Fig. 5, NK cells wereactually more responsive to APP in the CFSE assay than �� Tcells. NK cells are important in a variety of innate functions, in-cluding roles as potent antiviral and antitumor cells as well asproducers of IFN-�. Currently, attempts to expand NK cells forclinical benefit focus on cytokine-based expansion and are there-fore troubled with toxicity due to non-target cell activation (62,63). Our in vitro data suggest the potential for tannin-derived ago-nists to aid, in a manner akin to phosphoantigen/IL-2 stimulation,the cytokine-based expansion of this innate cell population. A frac-tion of �� T cells also responded within our assays. Analysis ofCD45RO expression suggested that the majority of the responding�� T cells were, surprisingly, naive T cells. This suggests thatsome primary Ag-driven �� T cell responses might be enhancedby these tannins, but this requires further testing.
A tannin-based drug has the potential to not only be used ther-apeutically, but prophylactically as well. A concern with the pro-phylactic use of any immunomodulatory compound is the induc-tion of chronic inflammation. Current phosphoantigen-based drugslead to overt cytokine release in humans (64, 65) including theproinflammatory cytokines TNF-� and IFN-�, which would con-tribute to systemic inflammation in the patient. In a concurrentstudy, we found that the tannin preparation we tested up-regulatedCD11b on �� T cells, but importantly, did not alter transcript lev-els for TNF-�. However, GM-CSF and MIP1� gene transcriptionwas induced (43). Thus, activation occurs following tannin treat-ment, but the response appears to be far more subtle than thatobserved with phosphoantigens. Our data suggests that under aprophylactic treatment regimen, tannin agonists prime but do notovertly activate cells. Upon introduction of an insult, such as ex-posure to a mucosal pathogen, we postulate that “primed” �� Tcells would respond faster and more robustly to secondary agonistssuch as pathogen-associated molecular patterns and cytokinessecreted from other innate cells, such as macrophages.
The chemical nature of the apple tannins responsible for theagonist activity for �� T cell is not defined. Most plants producetannins, but their chemical characteristics vary considerably be-tween different species. Though we are only now beginning tocatalog these results, it is clear that not all plant tannins are �� Tcell agonists. In screens of common dietary supplements, potentagonist activity was detected in extracts of the plants described inthis study, but the majority of plants tested contained minimal tono �� T cell agonist activity. In our ongoing drug discovery efforts,we are characterizing eight plant extracts with potent non-tannin�� T cell agonist activity (A. Palecanda, and M. Jutila, unpub-lished observations). Yamoa is an example of one such plant;
6476 SELECT TANNINS INDUCE �� T CELL ACTIVATION
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

although it produces complex tannins (data not shown), its activityfor �� T cells resides in its non-tannin fraction (Fig. 3A).
Condensed tannins range from simple monomers to highly com-plex oligomers (33). The �� T cell agonist described in this ex-periment elutes with the larger condensed tannin population (Fig.3E). Large condensed tannins are prohibitive to synthesize, butmay be amendable to bulk purification procedures. Techniques ofbulk tannin purification previously developed by the food industrymay be adaptable to purify the tannin complex involved in theseresponses (66, 67). Using these tannin isolation procedures forpurification and characterization of the minimal tannin complexfrom APP is a primary focus in further evaluating the potential foruse of these compounds as therapeutic and/or prophylactic agents.
Questions remain regarding the specific mechanisms of tanninactivity on leukocytes. Originally defined as low-affinity and non-specific protein binding complexes with antioxidant activity, tan-nins are increasingly portrayed as having high-affinity counter re-ceptors (68, 69). An example of an immunologically relevanttannin-protein specific interaction is found in tannic acid. Tannicacid is able to block the chemokine CXCL12 from binding to itsreceptor, CXCR4. The action of tannic acid is specific to CXCL12/CXCR4 given that other chemokines/chemoattractants are not ef-fected by this tannin preparation (70). Furthermore, apple tanninsblock Fc�R1/IgE binding (55) and prevent epidermal growth fac-tor signaling, presumably through physically blocking the receptor(57). Similar polyphenols from tea bind CD11b on monocytes,which we recently found for APP as well (43). Characterizing apotential receptor mediated response triggered by APP in �� Tcells was well beyond the intent of this initial study. However, thedata presented in this study are consistent with APP acting throughone, or perhaps a restricted number of, receptors on the T cell andnot through a nonspecific mechanism, such as antioxidant activity.Assuming the active agonist represents a fraction of the total crudematerial, activity is likely within nanogram per milliliter levels.This level of activity is consistent with signaling through a specificreceptor. Studies describing the specific binding of tannic acid toCXCR4 show this interaction competitively inhibits CXCL12binding at concentrations (IC50, 360 ng/ml) (70) similar to thosewe predict for the active component of APP. The restrictive patternof gene regulation induced by APP in �� T cells, selective up-regulation of IL-2R�, CD69, MIP1�, and GM-CSF shown in thisstudy and by Graff and Jutila (43), is also consistent with a recep-tor-mediated event. We are currently investigating the receptorsthat may be involved in these responses.
The effective dose range of crude APP required to induce IL-2R� on �� T cells was quite limited (1–40 �g/ml for bovine cellsand 1–20 �g/ml for human cells), but this was due to toxicity(induction of apoptosis) of the preparations at higher concentra-tions under the assay conditions tested. This level of toxicity isconsistent with other tannin preparations (70). These in vitro datasuggest a relatively narrow therapeutic and nontoxic range for tan-nins. However, in vivo studies in rats and mice demonstrate high-dose oral administration without toxic effects. Apple tannins pro-vided to mice ad libitium in their drinking water at 1.0% w/v leadto expansion of �� T cells in the gut mucosa, but this dose does notinduce obvious toxicity (49). Furthermore, when APP is given orallyto rats at concentrations up to 2000 mg/kg, all animals survive, gainbody weight, and overt inflammation of the gut mucosa or any otherorgan is not observed over a 14-day period (71). The reason such highdoses of APP administered orally are not toxic may be because the gutappears to regulate the uptake of orally administered tannins and pre-vents toxic levels from making it to the bloodstream. Specifically,plasma uptake of tannins in rats plateaus at 10.2 �g/ml (1000 mg/kgoral dose) and does not increase when rats are fed 2000 mg/kg of a
tannin preparation (72). This suggests oral administration may natu-rally prevent overdose, and regulate tannin concentrations in theplasma at optimal mitogenic concentrations.
In summary, although phosphoantigens are potent stimulators ofhuman �� T cells, they possess limitations that may prevent theirreliable expansion of �� T cell populations for potential treatmentof a variety of diseases. Therefore, the search for additional �� Tcell agonists capable of improving phosphoantigen stimulation ofV�2 T cells as well as agonists capable of expanding the V�1subset is warranted. Our results indicate tannins derived fromplants tissues, such as Cat’s Claw bark and nonripe apple peel,represent distinct and complementary agonists to phosphoantigens.Therefore, tannin-based therapeutics can potentially address someshortcomings of phosphoantigen-based drugs by functioning asstand-alone drugs or drugs used in combination with other immu-nomodulatory compounds.
AcknowledgmentsWe thank Matt Calverley for preparing the THP1-Blue CD14 TLR assays,Evelyn Benson for performing the Limulus assays, and Kathryn Crist, KirkLubick, and Jill Graff for critical review and several helpful suggestions.
DisclosuresM. A. Jutila holds shares in LigoCyte Pharmaceuticals, Inc., which togetherwith Montana State University, holds a National Institutes of Health con-tract that partially funded this work. All other Montana State Universityauthors have no financial conflict of interest.
References1. Chen, Y., K. Chou, E. Fuchs, W. L. Havran, and R. Boismenu. 2002. Protection
of the intestinal mucosa by intraepithelial �� T cells. Proc. Natl. Acad. Sci. USA99: 14338–14343.
2. Sharp, L. L., J. M. Jameson, G. Cauvi, and W. L. Havran. 2005. Dendritic epi-dermal T cells regulate skin homeostasis through local production of insulin-likegrowth factor 1. Nat. Immunol. 6: 73–79.
3. Hedges, J. F., K. J. Lubick, and M. A. Jutila. 2005. �� T cells respond directlyto pathogen-associated molecular patterns. J. Immunol. 174: 6045–6053.
4. Deetz, C. O., A. M. Hebbeler, N. A. Propp, C. Cairo, I. Tikhonov, andC. D. Pauza. 2006. Gamma interferon secretion by human V�2V�2 T cells afterstimulation with antibody against the T-cell receptor plus the Toll-Like receptor2 agonist Pam3Cys. Infect. Immun. 74: 4505–4511.
5. Lubick, K., and M. A. Jutila. 2006. LTA recognition by bovine �� T cells in-volves CD36. J. Leukoc. Biol. 79: 1268–1270.
6. Kabelitz, D., and D. Wesch. 2003. Features and functions of �� T lymphocytes:focus on chemokines and their receptors. Crit. Rev. Immunol. 23: 339–370.
7. Girardi, M. 2006. Immunosurveillance and immunoregulation by �� T cells.J. Invest. Dermatol. 126: 25–31.
8. Deusch, K., F. Luling, K. Reich, M. Classen, H. Wagner, and K. Pfeffer. 1991.A major fraction of human intraepithelial lymphocytes simultaneously expressesthe �/� T cell receptor, the CD8 accessory molecule and preferentially uses theV�1 gene segment. Eur. J. Immunol. 21: 1053–1059.
9. Wang, L., A. Kamath, H. Das, L. Li, and J. F. Bukowski. 2001. Antibacterialeffect of human V�2V�2 T cells in vivo. J. Clin. Invest. 108: 1349–1357.
10. Poccia, F., C. Agrati, F. Martini, G. Mejia, M. Wallace, and M. Malkovsky. 2005.V�9V�2 T cell-mediated non-cytolytic antiviral mechanisms and their potentialfor cell-based therapy. Immunol. Lett. 100: 14–20.
11. Lamb, L. S., Jr., and R. D. Lopez. 2005. �� T cells: a new frontier for immu-notherapy? Biol. Blood Marrow Transplant 11: 161–168.
12. Ebert, L. M., S. Meuter, and B. Moser. 2006. Homing and function of human skin�� T cells and NK cells: relevance for tumor surveillance. J. Immunol. 176:4331–4336.
13. Viey, E., G. Fromont, B. Escudier, Y. Morel, S. Da Rocha, S. Chouaib, andA. Caignard. 2005. Phosphostim-activated �� T cells kill autologous metastaticrenal cell carcinoma. J. Immunol. 174: 1338–1347.
14. Dunford, J. E., K. Thompson, F. P. Coxon, S. P. Luckman, F. M. Hahn,C. D. Poulter, F. H. Ebetino, and M. J. Rogers. 2001. Structure-activity relation-ships for inhibition of farnesyl diphosphate synthase in vitro and inhibition ofbone resorption in vivo by nitrogen-containing bisphosphonates. J. Pharmacol.Exp. Ther. 296: 235–242.
15. Thompson, K., J. Rojas-Navea, and M. J. Rogers. 2006. Alkylamines causeV�9V�2 T-cell activation and proliferation by inhibiting the mevalonate path-way. Blood 107: 651–654.
16. Amslinger, S., S. Hecht, F. Rohdich, W. Eisenreich, P. Adam, A. Bacher, andS. Bauer. 2007. Stimulation of V�9/V�2 T-lymphocyte proliferation by the iso-prenoid precursor, (E)-1-hydroxy-2-methyl-but-2-enyl 4-diphosphate. Immuno-biology 212: 47–55.
17. Sato, K., S. Kimura, H. Segawa, A. Yokota, S. Matsumoto, J. Kuroda,M. Nogawa, T. Yuasa, Y. Kiyono, H. Wada, and T. Maekawa. 2005. Cytotoxic
6477The Journal of Immunology
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

effects of �� T cells expanded ex vivo by a third generation bisphosphonate forcancer immunotherapy. Int. J. Cancer 116: 94–99.
18. Garcia, V. E., D. Jullien, M. Song, K. Uyemura, K. Shuai, C. T. Morita, andR. L. Modlin. 1998. IL-15 enhances the response of human �� T cells to nonpeptide[correction of nonpetide] microbial antigens. J. Immunol. 160: 4322–4329.
19. Wang, H., H. K. Lee, J. F. Bukowski, H. Li, R. A. Mariuzza, Z. W. Chen,K. H. Nam, and C. T. Morita. 2003. Conservation of nonpeptide antigen recog-nition by rhesus monkey V�2V�2 T cells. J. Immunol. 170: 3696–3706.
20. Sicard, H., S. Ingoure, B. Luciani, C. Serraz, J. J. Fournie, M. Bonneville,J. Tiollier, and F. Romagne. 2005. In vivo immunomanipulation of V�9V�2 Tcells with a synthetic phosphoantigen in a preclinical nonhuman primate model.J. Immunol. 175: 5471–5480.
21. Kunzmann, V., E. Bauer, and M. Wilhelm. 1999. �/� T-cell stimulation by pam-idronate. N. Engl. J. Med. 340: 737–738.
22. Newman, D. J., and G. M. Cragg. 2007. Natural products as sources of new drugsover the last 25 years. J. Nat. Prod. 70: 461–477.
23. Tanaka, Y., S. Sano, E. Nieves, G. De Libero, D. Rosa, R. L. Modlin,M. B. Brenner, B. R. Bloom, and C. T. Morita. 1994. Nonpeptide ligands forhuman �� T cells. Proc. Natl. Acad. Sci. USA 91: 8175–8179.
24. Poupot, M., and J. J. Fournie. 2004. Non-peptide antigens activating human V�9/V�2 T lymphocytes. Immunol. Lett. 95: 129–138.
25. Vesosky, B., O. C. Turner, J. Turner, and I. M. Orme. 2004. Gamma interferonproduction by bovine �� T cells following stimulation with mycobacterial my-colylarabinogalactan peptidoglycan. Infect. Immun. 72: 4612–4618.
26. Hedges, J. F., D. Cockrell, L. Jackiw, N. Meissner, and M. A. Jutila. 2003.Differential mRNA expression in circulating �� T lymphocyte subsets definesunique tissue-specific functions. J. Leukoc. Biol. 73: 306–314.
27. Wilson, E., B. Walcheck, W. C. Davis, and M. A. Jutila. 1998. Preferential tissuelocalization of bovine �� T cell subsets defined by anti-T cell receptor for antigenantibodies. Immunol. Lett. 64: 39–44.
28. Singleton, V. L., and J. A. Rossi, Jr. 1965. Colorimetry of total phenolics withphosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticulture 16:144–158.
29. MacHugh, N. D., J. K. Mburu, M. J. Carol, C. R. Wyatt, J. A. Orden, andW. C. Davis. 1997. Identification of two distinct subsets of bovine �� T cells withunique cell surface phenotype and tissue distribution. Immunology 92: 340–345.
30. MacHugh, N. D., A. Bensaid, C. J. Howard, W. C. Davis, and W. I. Morrison.1991. Analysis of the reactivity of anti-bovine CD8 monoclonal antibodies withcloned T cell lines and mouse L-cells transfected with bovine CD8. Vet. Immunol.Immunopathol. 27: 169–172.
31. Rohrbach, M. S., T. J. Kreofsky, and B. G. Chan. 1990. Structural determinantsof the platelet agonist activity of cotton bract condensed tannin. Environ. Res. 52:199–209.
32. Chan, W. S., S. Chowdhry, T. Chang, and R. R. Kew. 1999. Initial character-ization of the complement activating compounds in extracts of smokeless to-bacco. Immunobiology 201: 64–73.
33. Okuda, T. 2005. Systematics and health effects of chemically distinct tannins inmedicinal plants. Phytochemistry 66: 2012–2031.
34. Toth, G. B., and H. Pavia. 2001. Removal of dissolved brown algal phlorotanninsusing insoluble polyvinylpolypyrrolidone (PVPP). J. Chem. Ecol. 27: 1899–1910.
35. Wu, L. C., and K. J. Siebert. 2002. Characterization of haze-active proteins inapple juice. J. Agric. Food Chem. 50: 3828–3834.
36. Selga, A., and J. L. Torres. 2005. Efficient preparation of catechin thio conjugatesby one step extraction/depolymerization of pine (Pinus pinaster) bark procyani-dins. J. Agric. Food Chem. 53: 7760–7765.
37. Strumeyer, D. H., and M. J. Malin. 1975. Condensed tannins in grain sorghum:isolation, fractionation, and characterization. J. Agric. Food Chem. 23: 909–914.
38. Taylor, A. W., E. Barofsky, J. A. Kennedy, and M. L. Deinzer. 2003. Hop (Humuluslupulus L.) proanthocyanidins characterized by mass spectrometry, acid catalysis, andgel permeation chromatography. J. Agric. Food Chem. 51: 4101–4110.
39. Kenny, T. P., C. L. Keen, H. H. Schmitz, and M. E. Gershwin. 2007. Immuneeffects of cocoa procyanidin oligomers on peripheral blood mononuclear cells.Exp. Biol. Med. 232: 293–300.
40. Eisenreich, W., A. Bacher, D. Arigoni, and F. Rohdich. 2004. Biosynthesis ofisoprenoids via the non-mevalonate pathway. Cell Mol. Life Sci. 61: 1401–1426.
41. Fischer, S., A. Scheffler, and D. Kabelitz. 1996. Activation of human �� T-cellsby heat-treated mistletoe plant extracts. Immunol. Lett. 52: 69–72.
42. Kolodziej, H., and A. F. Kiderlen. 2005. Antileishmanial activity and immunemodulatory effects of tannins and related compounds on Leishmania parasitisedRAW 264.7 cells. Phytochemistry 66: 2056–2071.
43. Graff, J. C., and M. A. Jutila. 2007. Differential regulation of CD11b on �� T cellsand monocytes in response to unripe apple polyphenols. J. Leukoc. Biol. 82:603–607.
44. Kabelitz, D., D. Wesch, and W. He. 2007. Perspectives of �� T cells in tumorimmunology. Cancer Res. 67: 5–8.
45. Kabelitz, D., D. Wesch, E. Pitters, and M. Zoller. 2004. Characterization of tumorreactivity of human V�9V�2 �� T cells in vitro and in SCID mice in vivo.J. Immunol. 173: 6767–6776.
46. Poccia, F., L. Battistini, B. Cipriani, G. Mancino, F. Martini, M. L. Gougeon, andV. Colizzi. 1999. Phosphoantigen-reactive V�9V�2 T lymphocytes suppress invitro human immunodeficiency virus type 1 replication by cell-released antiviralfactors including CC chemokines. J. Infect. Dis. 180: 858–861.
47. Tanaka, Y., C. T. Morita, Y. Tanaka, E. Nieves, M. B. Brenner, and B. R. Bloom.1995. Natural and synthetic non-peptide antigens recognized by human �� Tcells. Nature 375: 155–158.
48. Jameson, J. M., G. Cauvi, L. L. Sharp, D. A. Witherden, and W. L. Havran. 2005.�� T cell-induced hyaluronan production by epithelial cells regulates inflamma-tion. J. Exp. Med. 201: 1269–1279.
49. Akiyama, H., Y. Sato, T. Watanabe, M. H. Nagaoka, Y. Yoshioka, T. Shoji,T. Kanda, K. Yamada, M. Totsuka, R. Teshima, J. Sawada, et al. 2005. Dietaryunripe apple polyphenol inhibits the development of food allergies in murinemodels. FEBS Lett. 579: 4485–4491.
50. Heitzman, M. E., C. C. Neto, E. Winiarz, A. J. Vaisberg, and G. B. Hammond.2005. Ethnobotany, phytochemistry and pharmacology of Uncaria (Rubiaceae).Phytochemistry 66: 5–29.
51. Tsuchiya, T., S. Fukuda, H. Hamada, A. Nakamura, Y. Kohama, H. Ishikawa,K. Tsujikawa, and H. Yamamoto. 2003. Role of �� T cells in the inflammatoryresponse of experimental colitis mice. J. Immunol. 171: 5507–5513.
52. Jonasson, R., A. Johannisson, M. Jacobson, C. Fellstrom, and M. Jensen-Waern.2004. Differences in lymphocyte subpopulations and cell counts before and afterexperimentally induced swine dysentery. J. Med. Microbiol. 53: 267–272.
53. Cairo, C., E. Arabito, F. Landi, A. Casati, E. Brunetti, G. Mancino, and E. Galli.2005. Analysis of circulating �� T cells in children affected by IgE-associated andnon-IgE-associated allergic atopic eczema/dermatitis syndrome. Clin. Exp. Im-munol. 141: 116–121.
54. Jones-Carson, J., E. Balish, and D. T. Uehling. 1999. Susceptibility of immuno-deficient gene-knockout mice to urinary tract infection. J. Urol. 161: 338–341.
55. Tokura, T., N. Nakano, T. Ito, H. Matsuda, Y. Nagasako-Akazome, T. Kanda,M. Ikeda, K. Okumura, H. Ogawa, and C. Nishiyama. 2005. Inhibitory effect ofpolyphenol-enriched apple extracts on mast cell degranulation in vitro targetingthe binding between IgE and Fc�RI. Biosci. Biotechnol. Biochem. 69:1974–1977.
56. Enomoto, T., Y. Nagasako-Akazome, T. Kanda, M. Ikeda, and Y. Dake. 2006.Clinical effects of apple polyphenols on persistent allergic rhinitis: a randomizeddouble-blind placebo-controlled parallel arm study. J. Investig. Allergol. Clin.Immunol. 16: 283–289.
57. Kern, M., Z. Tjaden, Y. Ngiewih, N. Puppel, F. Will, H. Dietrich, G. Pahlke, andD. Marko. 2005. Inhibitors of the epidermal growth factor receptor in apple juiceextract. Mol. Nutr. Food Res. 49: 317–328.
58. Kamath, A. B., L. Wang, H. Das, L. Li, V. N. Reinhold, and J. F. Bukowski.2003. Antigens in tea-beverage prime human V�2V�2 T cells in vitro and in vivofor memory and nonmemory antibacterial cytokine responses. Proc. Natl. Acad.Sci. USA 100: 6009–6014.
59. Poccia, F., S. Boullier, H. Lecoeur, M. Cochet, Y. Poquet, V. Colizzi,J. J. Fournie, and M. L. Gougeon. 1996. Peripheral V�9/V�2 T cell deletion andanergy to nonpeptidic mycobacterial antigens in asymptomatic HIV-1-infectedpersons. J. Immunol. 157: 449–461.
60. Wilhelm, M., V. Kunzmann, S. Eckstein, P. Reimer, F. Weissinger, T. Ruediger,and H. P. Tony. 2003. �� T cells for immune therapy of patients with lymphoidmalignancies. Blood 102: 200–206.
61. Li, B., M. D. Rossman, T. Imir, A. F. Oner-Eyuboglu, C. W. Lee, R. Biancaniello,and S. R. Carding. 1996. Disease-specific changes in �� T cell repertoire and functionin patients with pulmonary tuberculosis. J. Immunol. 157: 4222–4229.
62. Duvic, M., M. L. Sherman, G. S. Wood, T. M. Kuzel, E. Olsen, F. Foss,R. J. Laliberte, J. L. Ryan, K. Zonno, and A. H. Rook. 2006. A phase II open-label study of recombinant human interleukin-12 in patients with stage IA, IB, orIIA mycosis fungoides. J. Am. Acad. Dermatol. 55: 807–813.
63. Wang, G., M. Tschoi, R. Spolski, Y. Lou, K. Ozaki, C. Feng, G. Kim,W. J. Leonard, and P. Hwu. 2003. In vivo antitumor activity of interleukin 21mediated by natural killer cells. Cancer Res. 63: 9016–9022.
64. Conti, L., R. Casetti, M. Cardone, B. Varano, A. Martino, F. Belardelli, F. Poccia,and S. Gessani. 2005. Reciprocal activating interaction between dendritic cellsand pamidronate-stimulated �� T cells: role of CD86 and inflammatory cyto-kines. J. Immunol. 174: 252–260.
65. Casetti, R., G. Perretta, A. Taglioni, M. Mattei, V. Colizzi, F. Dieli, G. D’Offizi,M. Malkovsky, and F. Poccia. 2005. Drug-induced expansion and differentiationof V�9V�2 T cells in vivo: the role of exogenous IL-2. J. Immunol. 175:1593–1598.
66. Laborde, B., V. Moine-Ledoux, T. Richard, C. Saucier, D. Dubourdieu, andJ. P. Monti. 2006. PVPP-polyphenol complexes: a molecular approach. J. Agric.Food Chem. 54: 4383–4389.
67. Saucier, C., M. Jourdes, Y. Glories, and S. Quideau. 2006. Extraction, detection,and quantification of flavano-ellagitannins and ethylvescalagin in a Bordeaux redwine aged in oak barrels. J. Agric. Food Chem. 54: 7349–7354.
68. Hagerman, A. E., and L. G. Butler. 1981. The specificity of proanthocyanidin-protein interactions. J. Biol. Chem. 256: 4494–4497.
69. Frazier, R. A., A. Papadopoulou, I. Mueller-Harvey, D. Kissoon, and R. J. Green.2003. Probing protein-tannin interactions by isothermal titration microcalorim-etry. J. Agric. Food Chem. 51: 5189–5195.
70. Chen, X., J. A. Beutler, T. G. McCloud, A. Loehfelm, L. Yang, H. F. Dong,O. Y. Chertov, R. Salcedo, J. J. Oppenheim, and O. M. Howard. 2003. Tannicacid is an inhibitor of CXCL12 (SDF-1�)/CXCR4 with antiangiogenic activity.Clin. Cancer Res. 9: 3115–3123.
71. Shoji, T., Y. Akazome, T. Kanda, and M. Ikeda. 2004. The toxicology and safetyof apple polyphenol extract. Food Chem. Toxicol. 42: 959–967.
72. Shoji, T., S. Masumoto, N. Moriichi, H. Akiyama, T. Kanda, Y. Ohtake, andY. Goda. 2006. Apple procyanidin oligomers absorption in rats after oral admin-istration: analysis of procyanidins in plasma using the porter method and high-performance liquid chromatography/tandem mass spectrometry. J. Agric. FoodChem. 54: 884–892.
6478 SELECT TANNINS INDUCE �� T CELL ACTIVATION
by guest on January 13, 2022http://w
ww
.jimm
unol.org/D
ownloaded from