carbohydrate oxidase from a red alga, · carbohydrate oxidase from a red alga, iridophycus...
TRANSCRIPT

CARBOHYDRATE OXIDASE FROM A RED ALGA, IRIDOPHYCUS FLACCIDUM”
BY R. C. BEANt AND W. Z. HASSID
(From the Department of Plant Biochemistry, College of Agriculture, University of California, Berkeley, California)
(Received for publication, May 27, 1955)
Enzymes capable of oxidizing D-glucose to n-gluconic acid are known to be present in bacteria (1, 2)) fungi (3)) and animal tissues (4, 5). How- ever, no such enzyme has hit,herto been reported in a photosynthetic organism. It was observed in the course of a study of the carbohydrate metabolism of the marine red alga, Iridophycus JEaccidum (6), that cell-free extracts of this plant had the ability to oxidize D-glucose to n-gluconic acid. Further investigation revealed that this preparation will also oxidize D-galactose to n-galactonic acid, and several reducing disaccharides, namely maltose, lactose, and cellobiose, to their corresponding aldobionic acids. It therefore appears that this oxidase differs from other D-glucose oxidases in that it can utilize a variety of sugar substrates.
The present communication is concerned with the preparation of active oxidase fractions from this red alga, the purification, and study of the properties of the enzyme.
EXPERIMENTAL
Assay Methods-A standard Warburg manometric method (7) was employed for assay of the enzyme. An aliquot of 0.01 to 1.0 ml. of the enzyme solution, together with 1.0 ml. of a 0.5 M maleate buffer, pH 5.2, was placed in the main chamber of the Warburg vessel and diluted with water to a volume of 2 ml. A 0.5 ml. aliquot of 0.5 M solution of D-
glucose (saturation of the enzyme occurs at approximately 0.02 M glucose) or other carbohydrate used as substrate was placed in the side arm. Since no carbon dioxide was evolved in the course of the reaction, it was not necessary to use alkali in the center well of the main chamber. Materials which were tested as inhibitors or activators were mixed with the enzyme in the main chamber or added from a second side arm, depending on the conditions of the experiment. Hydrogen peroxide was generated during t,he reaction. In the absence of added catalase the peroxide was not
* This work was supported in part by a research contract with the United States Atomic Energy Commission.
t Present address, Department of Plant Biochemistry, Citrus Experiment Sta- tion, University of California, Riverside, California.
425
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

426 CARBOHYDRATE OXIDASE FROM I. FLACCIDUM
decomposed. Determinations were made without the addition of catalase in order to take advantage of the greater rate of oxygen uptake in the absence of this enzyme. An enzyme unit of oxidase was the amount of enzyme required to cause the uptake of 1 pmole of oxygen per hour at 30”. This value of oxygen uptake also corresponded to the oxidation of 1 pmole of n-glucose to n-gluconic acid per hour.
In addition to the manometric assay, a photometric determination for the enzyme, with 2,6-dichlorophenol-indophenol, was occasionally em- ployed. This assay proved to be useful to rapid checks of oxidase activity during the process of fractionation. However, since the photometric assay could not be carried out at pH 5, which is the optimum of the reac- tion, it was not generally used.
Analysis for the gluconolactone was made by the method of Hestrin (8). The enzymatic reaction was carried out for varying periods in Warburg flasks in arsenate buffer (the maleate buffer interfered with the lactone determination) at pH 5. The reaction was then stopped by the addition of the hydroxylamine in alkaline solution and the hydroxamic acid forma- tion carried out according to Hestrin’s procedure.
Extraction Procedure and Fractionation of Enzyme Preparations-Fresh I. Jlaccidum (6) plants were rinsed with distilled water, the excess water was wiped off, and the material placed in a cold room at - 12’ for 2 to 4 hours. The partially frozen thalli were ground in the cold with a hand meat grinder having 0.25 inch apertures, 1 kilo of the coarsely ground material was taken up in 1500 ml. of distilled water, and the macerated tissue allowed to stand overnight at 04”. After infiltration of the ground material by squeezing through four layers of cheese-cloth, 1050 ml. of solution were obtained. Upon extraction of the solid residue with 1 liter of water and filtration through a cheese-cloth as before, another liter of solution was obtained. The extracts, having almost identical oxidase activity, were combined, and the extracted residue, which still contained a small amount of activity, was discarded. Before the barium-methanol fractionation, the extract was dialyzed overnight against tap water.
Barium-Methanol Fractionation-60 gm. of BaCha 2HsO were dissolved in 350 ml. of the dialyzed red alga extract and the solution was frozen overnight. The frozen extract was allowed to thaw, the insoluble material was removed by centrifugation (10,000 X g for 10 minutes), the solution cooled in ice to lo, and 35 ml. of methanol at -10” were added over a period of 15 minutes. The precipitate of low activity was removed by centrifuging for 10 minutes at 09. The supernatant solution was treated with another 115 ml. of cold methanol, the temperature being kept below 2”. After standing for half an hour with occasional stirring, the second precipitate was removed by centrifugation.
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from
R. C. BEAN AND TV. Z. HASSID 427
The third supernatant solution was further treated with a slow addition of 200 ml. of methanol so that the temperature did not rise above 2”. After centrifuging for 10 minutes in the cold, the precipitate (Fraction 1, Table I) was suspended in 10 ml. of water, dialyzed against tap water for 2 hours, and then overnight at 1” in distilled water. This fraction (11 ml.) had an activity of 332 units per ml.
After precipitation of Fraction 1, the supernatant solution was devoid of enzymatic activity. The first two methanol precipitates could be resolved into an inactive component and a fraction with a specific activity approximating that of Fraction 1 by subjecting them to a repetition of the above procedure.
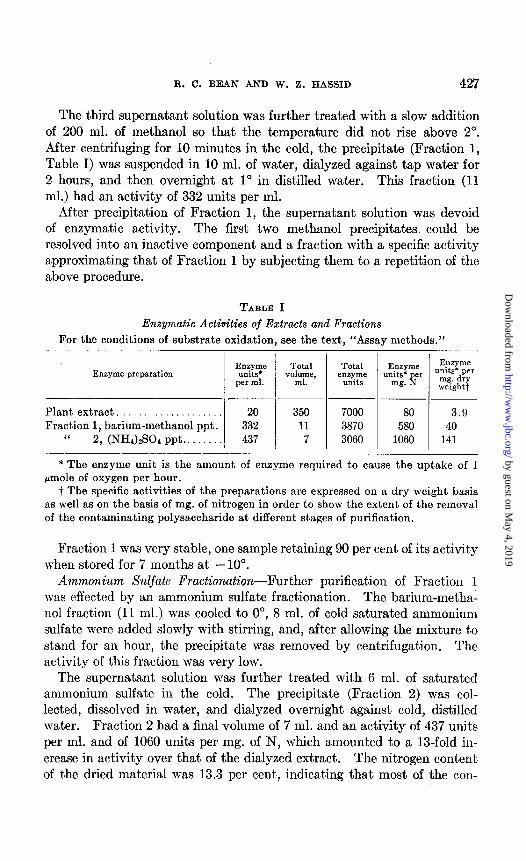
TABLE I
Enzymatic Activities of Extracts and Fractions For the conditions of substrate oxidation, see the text, “Assay methods.”
___~ Plant extract. . 20 350 7000 80 3.9 Fraction 1, barium-methanol ppt. 332 11 3870 580 40
I‘ 2, (NH&S04 ppt.. 437 7 3060 1060 141
* The enzyme unit is the amount of enzyme required to cause the uptake of 1 pmole of oxygen per hour.
t The specific activities of the preparations are expressed on a dry weight basis as well as on the basis of mg. of nitrogen in order to show the extent of the removal of the contaminating polysaccharide at different stages of purification.
Fraction 1 was very stable, one sample retaining 90 per cent of its activity when stored for 7 months at -10”.
Ammonium Sulfale Fractionation-Further purification of Fraction 1 was effected by an ammonium sulfate fractionation. The barium-metha- nol fraction (11 ml.) was cooled to 0”, 8 ml. of cold saturated ammonium sulfate were added slowly with stirring, and, after allowing the mixture to stand for an hour, the precipitate was removed by centrifugation. The activity of this fraction was very low.
The supernatant solution was further treated with 6 ml. of saturated ammonium sulfate in the cold. The precipitate (Fraction 2) was col- lected, dissolved in water, and dialyzed overnight against cold, distilled water. Fraction 2 had a final volume of 7 ml. and an activity of 437 units per ml. and of 1060 units per mg. of N, which amounted to a 13-fold in- crease in activity over that of the dialyzed extract. The nitrogen content of the dried material was 13.3 per cent, indicating that most of the con-
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

428 CARBOHYDRATE OXIDASE FROM I. FLACCIDUM
taminating polysaccharide had been removed at this stage of purification. Expressed on dry weight basis, the final “purification” amounted to a 36-fold increase in activity. Fraction 2 was not as stable as the less purified fractions. Most of its activity was lost on further attempts of purification. On remaining frozen at -20” for a month, it lost 50 per cent of its activity.
Oxidation of Sugars and Identification of Aldonic Acids-The n-gluconic and n-galactonic acids derived from the oxidation of n-glucose and D-
galactose, respectively, by the oxidase were identified by a combination of radioactive tracer and paper chromatographic techniques. In addition, the oxidation product of n-glucose was isolated as the barium gluconate salt and as crystalline n-gluconolactone, and the compounds were identified by their physical and chemical properties.
C14-labeled glucose, galactose, and maltose were incubated with the enzyme and the products were identified chromatographically by com- paring with synthetic samples of gluconic, galactonic, and maltobionic acids, procedures previously described being used (6). The results showed that the corresponding aldonic acid was formed from each sugar in all three cases. The chromatographic patterns of the reaction products were found to be identical with those of n-gluconic, n-galactonic, and malto- bionic acids, which had been prepared by bromine oxidation of the radio- active n-glucose, n-galactose, and n-maltose, respectively (9). Hydrolysis of the enzymatically formed maltobionic acid produced glucose and gluconic acids.
The formation of gluconic acid in the oxidation of the C14-labeled glucose was further confirmed by degradation of the chromatographically isolated oxidation product to arabinose by treatment with hydrogen peroxide in the presence of a ferric acetate catalyst (10).
In another experiment 250 mg. of glucose were incubated with the en- zyme. Barium gluconate was isolated from the reaction mixture and the product recrystallized from methanol and water to give 95.1 mg. of the pure salt. This enzymatically prepared crystalline form had a melting point of 147-148”, and a specific rotation, [CX] D $7.9” (c, 1 in water). These constants are in good agreement with those found for authentic gluconate which had been crystallized in the same manner.
Gluconolactone was prepared from pooled barium gluconate fractions by a modification of the method of Isbell et al. (11). A yield of 39.8 mg. of the lactone was obtained. Polarization of this lactone showed an initial [a] D +65.9” after 8 minutes, which gradually changed to a constant value of 7.6” after 6 hours. These values agreed closely with those of n-gluconolactone obtained by other workers (12).
The primary product of the reaction was actually the lactone of the
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from
R. C. BEAN AND W. Z. HASSID 429
aldonic acid rather than the free acid identified above. That this was the case was shown in two ways. When the enzyme was allowed to act on glucose in the absence of buffer in an oxygen atmosphere, there was a slow change in pH at the beginning of the reaction. After replacing the oxygen with nitrogen to stop the reaction, the gradual drop in pH con- tinued to occur, indicating that an acidic group was still being liberated even in the absence of oxidation. It was also possible to show formation of hydroxamic acid from the la&one, when the oxidation was carried out for 15 minutes and then alkaline hydroxylamine was added to stop the
200
100
0 IO 20 ; TIME IN MINUTES
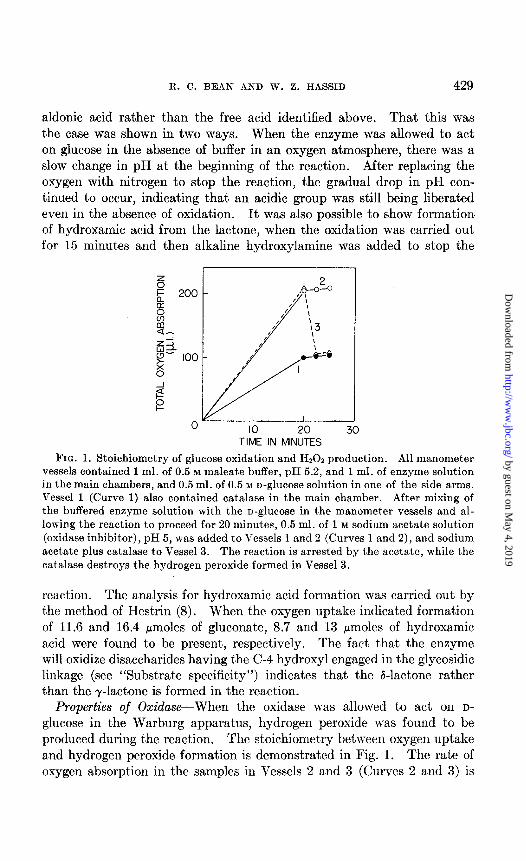
FIG. 1. Stoichiometry of glucose oxidation and Hz02 production. All manometer vessels contained 1 ml. of 0.5 M maleate buffer, pH 5.2, and 1 ml. of enzyme solution in the main chambers, and 0.5 ml. of 0.5 M D-glucose solution in one of the side arms. Vessel 1 (Curve 1) also contained catalase in the main chamber. After mixing of the buffered enzyme solution with the o-glucose in the manometer vessels and al- lowing the reaction to proceed for 20 minutes, 0.5 ml. of 1 M sodium acetate solution (oxidase inhibitor), pH 5, was added to Vessels 1 and 2 (Curves 1 and 2), and sodium acetate plus catalase to Vessel 3. The reaction is arrested by the acetate, while the
catalase destroys the hydrogen peroxide formed in Vessel 3.
react,ion. The analysis for hydroxamic acid format,ion was carried out by t’he method of Hestrin (8). When the oxygen uptake indicated formation of 11.6 and 16.4 pmoles of gluconate, 8.7 and 13 pmoles of hydroxamic acid were found to be present, respectively. The fact that the enzyme will oxidize disaccharides having the C-4 hydroxyl engaged in the glycosidic linkage (see “Substrate specificity”) indicates that the d-lactone rather than the y-lactone is formed in the reaction.
Properties of Oxidase-When the oxidase was allowed to act on D-
glucose in the Warburg apparatus, hydrogen peroxide was found to be produced during the reaction. The stoichiometry between oxygen uptake and hydrogen peroxide formation is demonstrated in Fig. 1. The rate of oxygen absorption in the samples in Vessels 2 and 3 (Curves 2 and 3) is
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

430 CARBOHYDRATE OXIDASE FROM I. FLACCIDUM
twice as great as that in Vessel 1 to which catalase had been added at the beginning of the experiment. When catalase was added after 20 minutes to Vessel 3 in the presence of an oxidase inhibitor (sodium acetate), oxygen was rapidly evolved equivalent to half that consumed in the absence of catalase. This indicates that the samples are completely free of catalase and that 1 mole of hydrogen peroxide is formed per mole of oxygen con- sumed.
The enzyme can utilize 2,6-dichlorophenol-indophenol as a hydrogen acceptor; in the presence of oxygen, both the dye and oxygen are reduced. Oxygen appears to be reduced more readily than the dye. Other com- pounds, such as methylene blue, tetrazolium violet, or ferricyanide in ranges from 1W5 to 1e3 M which usually react with flavoproteins, are not reduced by this enzyme under anaerobic conditions. The oxidase does not require the addition of any coenzymes and retains its full activity after long periods of dialysis. Attempts to split the protein from the prosthetic group by treatment with acid and then to reconstitute the enzyme were not successful. When the enzyme was treated with acid in the presence of ammonium sulfate at pH 3, there was no loss of activity in the precipitated protein. At pH 2 the enzymatic activity was com- pletely lost and could not be restored by the addition of flavin adenine dinucleotide or with concentrates of a supernatant solution from boiled enzyme preparations. Dialysis against dilute acid (0.01 M) also destroyed the activity irreversibly.
Absorption spectra given by the partially purified enzyme and by the split-products after acid treatment of the enzyme did not show the charac- teristic flavin absorption maxima peaks at 450 and 380 rnp. Strong absorption maxima at 265 and 330 rnp were given by the split-products. However, there was no correlation between the material responsible for these absorption peaks and the enzymatic activity in the fractionations; the same absorption peaks were found to be even more pronounced in some inactive fractions.
Substrate Xpecifiity-The following carbohydrates were tested as possi- ble substrates: n-glucose, n-galactose, maltose, lactose, cellobiose, L-
glucose, n-fructose, n-mannose, melibiose, raffinose, n-fucose, n-arabinose, n-xylose, n-galacturonic acid, n-glucuronolactone, n-glucose-6-phosphate, dulcitol, glycerol, glycerol phosphate, cY-methyl-n-glucoside, and 2,3- methyl-n-glucose. Of these compounds only the first five listed were found to be attacked by the oxidase preparations (see Table II).
‘All the organic compounds tested as inhibitors (Table IV) were also examined to determine whether they caused oxygen absorption or carbon dioxide evolution in the presence of the enzyme under normal conditions of enzymatic oxidation. None of these substances were found to be active in this respect.
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

R. C. BEAN AND W. Z. HASSID 431
Table II represents the rate of oxidation of five carbohydrates found to be attacked by the oxidase preparation. That the same enzyme is prob- ably responsible for the oxidation of both n-glucose and n-galactose (and possibly for the three disaccharides) is shown by the fact that the ratio of the rates of oxidation of D-glucose and D-galactose is constant at all stages of the enzyme purification (Table III). An experiment in which saturat- ing amounts of both n-glucose and n-galactose were added to the incuba-
TABLE II
Rates of Oxidatiolz of Various Carbohydrate Substrates by I. Jlaccidum Oxidase
For conditions of substrate oxidation, see the text, “Assay methods.” 0.1 M concentrations of sugars were used.
Substrate Micromoles 02 per min. per ml. enzyme
n-Glucose........................................ 4.3 D-Galactose...................................... 4.3 Lactose.......................................... 2.1 Maltose.......................................... .._. 2.0 Cellobiose........................ 1.3
TABLE III
Comparison of Rates of Oxidation of D-Glucose and n-Galactose at Different Stages of Enzyme Fractionation
0.1 M concentrations of n-glucose and n-galactose were used, and the oxidations were carried out as described under “Assay methods.” The values are given in micromoles of 02 per hour per ml. of enzyme.
Enzyme preparation D-GlUCOSe D-Galactose Activity ratio, D- galactose to D-glucose
Extract.......................... 20 18 0.90 Methanol ppt.. . 40 34 0.85 Fraction 1, barium-methanol. 332 300 0.90 Fraction 2, ammonium sulfate.. 437 390 0.89
tion mixture showed that no increase in the rate of oxygen uptake occurred over that found when n-glucose alone was present.
The Michaelis constant for n-glucose was found to be 0.0025 M and that for D-galactose 0.005 M. These constants are of the same order of magni- tude as Km for n-glucose wit.h notatin, which is given by Keilin and Har- tree (3) as 0.0042 M.
The disaccharides appear to be oxidized directly without preliminary hydrolysis. This is shown by the formation of maltobionic acid from maltose and the fact that, when the reactions are allowed to proceed to completion, the total oxygen consumption with the disaccharides is equal to that obtained with equimolar amounts of the monosaccharides.
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

432 CARBOHYDRATE OXIDASE FROM I. FLACCIDUM
0 IO 20 TIME (MINUTES)
FIG. 2. Comparison of enzymatic action on LY and p isomers of glucose. 0, a-n-glucose; l , @-n-glucose. 0.75 ml. of arsenate buffer, 0.25 ml. of enzyme solution, and 2 mg. of dry, crystalline 01. or &n-glucose were added from the side arm at zero time to give 0.011 M concentration. The p-n-glucose contained 85 per cent of the fi isomer.
TABLE IV
Inhibition of Sugar Dehydrogenase from I. fiaccidum
Mercuric chloride .......... Lead acetate ............... Silver nitrate .............. p-Chloromercuribenzoate. .. Acetate,sodium ........... Propionate, sodium ........ Pyruvate, potassium ....... Benzoate, sodium .......... n-Mannoset ...............
............
............ ............ ............
. . . . . . . . . . . .
. . . . . . . . . .
..__...._...
I- Per cent inhibition’s1
10-5 M
51:
50 2-5
- 10-a M
100 100 100
250 21 19
10-1 Y
63 60 20
100
lo-’ M
95 90
40
Assay conditions, except in the case of n-mannose and n-xylose: 1 ml. of maleate buffer, 0.5 ml. of enzyme solution in the main chamber of the manometer vessel; 0.5 ml. of 0.5 M n-glucose in Side Arm 1; 0.5 ml. of inhibitor test solution in Side Arm 2. Rates of oxygen uptake were checked with n-glucose alone in each experiment, and then the inhibitor was added from Side Arm 2 to determine the inhibited rate.
* Other compounds were tested which caused no inhibitory effects: 10e2 M con- centrations of copper sulfate, aluminum sulfate, zinc chloride, magnesium chloride, calcium chloride, sodium formate, sodium butyrate, sodium lactate, sodium glyco- late, sodium glyoxylate, sodium citrate, sodium succinate, sodium Versenate, hydroxylamine hydrochloride; 10-r M barium chloride, and n-xylose and acetalde- hyde.
t The glucose in the mannose and xylose experiments was added as 0.01 M solu- tion to give a final concentration of 0.002 M instead of the 0.1 M final concentration used in the rest of the experiments.
$ Mercuric chloride at lo+ M gave 60 per cent inhibition. 5 Actual concentration of p-chloromercuribenzoate was 2 X lo-+ M.
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from
R. C. BEAN AND TV. 2. HASSID 433
When the CY and p isomers of n-glucose were tested with the enzyme at low concentrations, it was found that the /3-n-glucose was oxidized at a slightly higher rate than the (Y isomer (Fig. 2).
Inhibitor Action-The compounds tested for inhibition of the enzymatic activity are listed in Table IV. Although Hg++, Ag+, Pb++, and p-chloro- mercuribenzoate do not inhibit the enzyme, it is doubtful that the slight inhibitory effects at low concentrations are connected with any specific
‘4 FIG. 3
I I 1 I I I 0 4 6 8
PH
FIG. 4
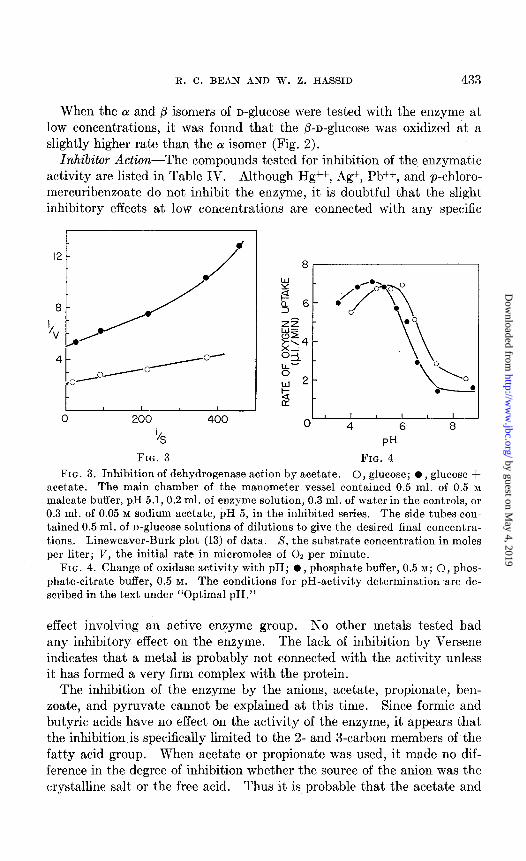
FIG. 3. Inhibition of dehydrogenase action by acetate. 0, glucose; l , glucose + acetate. The main chamber of the manometer vessel contained 0.5 ml. of 0.5 M
maleate buffer, pH 5.1, 0.2 ml. of enzyme solution, 0.3 ml. of water in the controls, or 0.3 ml. of 0.05 M sodium acetate, pH 5, in the inhibited series. The side tubes con- tained 0.5 ml. of n-glucose solutions of dilutions to give the desired final concentra- tions. Lineweaver-Burk plot (13) of data. S, the substrate concentration in moles per liter; V, the initial rate in micromoles of 02 per minute.
FIG. 4. Change of oxidase activity with pH; l , phosphate buffer, 0.5 M; 0, phos- phate-citrate buffer, 0.5 M. The conditions for pH-activity determination are de- scribed in the text under “Optimal PH.”
effect involving an active enzyme group. No other metals tested had any inhibitory effect on the enzyme. The lack of inhibition by Versene indicates that a metal is probably not connected with the activity unless it has formed a very firm complex with the protein.
The inhibition of the enzyme by the anions, acetate, propionate, ben- zoate, and pyruvate cannot be explained at this time. Since formic and butyric acids have no effect on the activity of the enzyme, it appears that the inhibition is specifically limited to the 2- and 3-carbon members of the fatty acid group. When acetate or propionate was used, it made no dif- ference in the degree of inhibition whether the source of the anion was the crystalline salt or the free acid. Thus it is probable that the acetate and
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from
434 CARBOHYDRATE OXIDASE FROM I. FLACCIDUM
propionate themselves, rather than some impurity, are responsible for the inhibition. As shown in Fig. 3, the effect of acetate is non-competitive with D-ghCOSe.
n-Mannose, but not n-xylose, acts as a weak competitive inhibitor of the reaction with n-glucose. The effect of n-mannose is perceptible only at fairly high ratios of n-mannose to n-glucose.
Optimal pH-The pH optimum for the oxidase was determined by using phosphate-citrate buffer as well as phosphate buffer alone, adjusted to the desired pH. 1 ml. of the enzyme solution was added to 1 ml. of the
61 I
0 IO 30 50 100 200 500 1000 2000
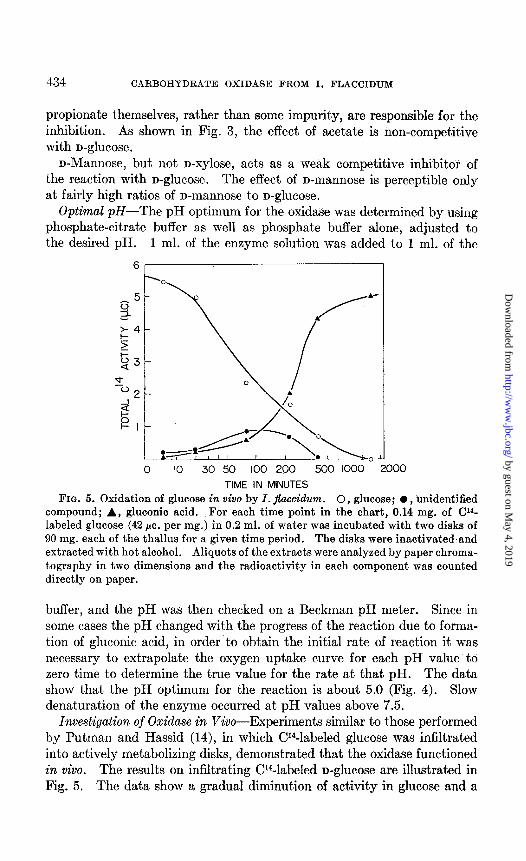
TIME IN MINUTES FIG. 5. Oxidation of glucose in vivo by I. jfaccidum. 0, glucose; 0, unidentified
compound; A, gluconic acid. For each time point in the chart, 0.14 mg. of CP- labeled glucose (42 pc. per mg.) in 0.2 ml. of water was incubated with two disks of 90 mg. each of the thallus for a given time period. The disks were inactivated and extracted with hot alcohol. Aliquots of the extracts were analyzed by paper chroma- tography in two dimensions and the radioactivity in each component was counted directly on paper.
buffer, and the pH was then checked on a Beckman pH meter. Since in some cases the pH changed with the progress of the reaction due to forma- tion of gluconic acid, in order’to obtain the initial rate of reaction it was necessary to extrapolate the oxygen uptake curve for each pH value to zero time to determine the true value for the rate at that pH. The data show that the pH optimum for the reaction is about 5.0 (Fig. 4). Slow denaturation of the enzyme occurred at pH values above 7.5.
Investigation of Oxidase in Viva-Experiments similar to those performed by Putman and Hassid (14), in which W-labeled glucose was infiltrated into actively metabolizing disks, demonstrated that the oxidase functioned in vivo. The results on infiltrating CY4-labeled n-glucose are illustrated in Fig. 5. The data show a gradual diminution of activity in glucose and a
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

R. C. BEAN AND W. 2. HASSID 435
simultaneous appearance of activity in gluconic acid and in a transient, unidentified compound. At the end of the reaction most of the activity was found in gluconic acid.
The results with infiltrated CP-labeled n-galactose were similar to those obtained with radioactive n-glucose. The aldonic acid and an unidentified compound were simultaneously produced. At the end of the incubation period the activity was found to reside almost quantitatively in the galac- tonic acid and its lactones.
Gluconic acid was also identified as one of the CY4-labeled products when the plant was allowed to photosynthesize in the presence of CY402 (6).
DISCUSSION
Two systems which oxidize n-glucose to D-&lCOniC acid have been thoroughly studied. The first, n-glucose oxidase (notatin), which exists in Penicillium notatum and possesses flavin adenine dinucleotide as its prosthetic group (3), is quite specific for n-glucose; no other sugar is oxidized by this enzyme at a rate greater than 1 per cent of that of D-
glucose. Sodium nitrite, fl-hydroxyquinoline, and semicarbazide are the only substances found to have an appreciable inhibitory effect on notatin. The second, the mammalian n-glucose dehydrogenase system (4), depends upon the pyridine nucleotides as its coenzymes. This enzyme is also highly specific for n-glucose, although it attacks n-xylose to some extent (5). It is inhibited by a low concentration of heavy metals. A third system, which depends upon a modified cytochrome transport path (l), is found in Pseudomonas Jluorescens. This system is specific for n-glucose and is strongly inhibited by cyanide.
Since the generation of hydrogen peroxide is frequently associated with flavoprotein oxidation, it is probable that this enzyme contains a flavin coenzyme. However, spectrographic analysis of the enzyme preparation did not show the presence of a flavin group. The inability to demonstrate this group may be attributed to the low purity of the enzyme preparation. Comparing the rate of oxidation of n-glucose by this impure oxidase prep- aration with that by pure notatin on dry weight basis, it is found that the former is about 25 times less active than the latter. In its behavior to- ward heavy metals, the I;ridophycus enzyme appears to be similar to the mammalian dehydrogenase. However, in its substrate specificity it dif- fers radically from all the other known oxidases. The fact that the Ir- idophycus oxidase is capable of oxidizing n-glucose and n-galactose but not n-mannose indicates that the enzyme is specific with regard to the C-2 configuration and is non-specific with regard to that of C-4 in the hexose chain. Since this enzyme is also capable of oxidizing the disac- charides, lactose, maltose, and cellobiose, it can be concluded that the
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

436 CARBOHYDRATE OXIDASE FROM I. FLACCIDUM
hydroxyl of C-4 may be substituted by a n-glucose or n-galactose unit without greatly affecting the activity of the enzyme. A 6-carbon sugar is apparently required, since the enzyme shows no activity toward D-xylose or L-arabinose, which are the pentose analogues of n-glucose and n-galac- tose, respectively. Inasmuch as modification of the hydroxyl group at C-6 of n-glucose by substitution, as in melibiose, or by oxidation of C-6 of D-galactose, as in galacturonic acid, prevents the enzymatic reaction from taking place, the primary C-6 hydroxyl appears to be indispensabie.
The function of direct oxidation of sugars in living cells is not clear. Although there is evidence that the gluconate produced in the red alga is slowly utilized, its rate of accumulation appears to exceed greatly t*hat of utilization.
SUMMARY
A cell-free enzyme preparation capable of oxidizing D-glucose to D-
gluconic acid, D-galactose to n-galactonic acid, and the disaccharides, maltose, lactose, and cellobiose, to their corresponding aldobionic acids has been obtained from the red alga Iridophycus jloxcidum. In these reactions oxygen is consumed and Hz02 is formed. The optimal pH for the oxidation is approximately 5.0.
The enzyme appears to be specific with regard to the C-2 configuration of the hexose chain. It remains active when the C-4 configuration is changed or when the hydroxyl on this carbon is substituted by a n-glucose unit.
The enzyme from the algal extract was partially purified by fractiona- tion with methanol in t,he presence of barium followed by ammonium sulfate precipitation.
BIBLIOGRAPHY
1. Wood, W. A., and Schwerdt, R. F., J. Biol. C&m., 201,501 (1953). 2. Ikada, Y., J. Agr. Chem. Sot. Japan, 24, 51 (1950). 3. Keilin, D., and Hartree, E. F., Biochem. J., 42, 221 (1948). 4. Eichel, B., and Wainio, W. W., J. Biol. Chem., 176, 155 (1948). 5. Strecker, H. J., and Korkes, S., J. Biol. Chem., 196, 769 (1952). 6. Bean, R. C., and Hassid, W. Z., J. Biol. Chem., 212, 411 (1955). 7. Umbreit, W. W., Burris, R. H., and Stauffer, J. F., Manometric techniques and
related methods for the study of tissue metabolism, Minneapolis (1945). 8. Hestrin, S., J. Biol. Chem., 160, 249 (1949). 9. Hudson, C. S., and Isbell, H. S., J. Am. Chem. Sot., 51,2225 (1929).
10. Hackett, R. C., and Hudson, C. S., J. Am. Chem. Sot., 66,1632 (1934). 11. Isbell, H. S., Karabinos, J. V., Frush, H. L., Holt, N. B., Schwebel, A., and
Galkowski, T. T., J. Res., Nat. Bur. Standards, 48, 163 (1952). 12. Brackenbury, J. M., and Upson, F. W., J. Am. Chem. Sot., 66, 2512 (1933). 13. Lineweaver, H., and Burk, D., J. Am. Chem. Sot., 56, 658 (1934). 14. Putman, E. W., and Hassid, W. Z., J. BioZ. Chem., 207,885 (1954).
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

R. C. Bean and W. Z. HassidFLACCIDUM
RED ALGA, IRIDOPHYCUS CARBOHYDRATE OXIDASE FROM A
1956, 218:425-436.J. Biol. Chem.
http://www.jbc.org/content/218/1/425.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/218/1/425.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from