universidade federal do rio grande do norte centro … · por permanganometria.....37 figura 9:...
TRANSCRIPT

1
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
AVALIAÇÃO DE PARÂMETROS DE ESTRESSE OXIDATIVO EM PLANTAS
DE CANA-DE-AÇÚCAR TRATADAS COM PERÓXIDO DE HIDROGÊNIO
KELLYA FRANCISCA MENDONÇA BARRETO
NATAL – RN
2013

2
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
AVALIAÇÃO DE PARÂMETROS DE ESTRESSE OXIDATIVO EM PLANTAS
DE CANA-DE-AÇÚCAR TRATADAS COM PERÓXIDO DE HIDROGÊNIO
KELLYA FRANCISCA MENDONÇA BARRETO
Dissertação apresentada ao programa
de Pós-Graduação em Ciências
Biológicas, do Centro de Biociências da
Universidade Federal do Rio Grande do
Norte, como parte dos requisitos para
obtenção do título de Mestre em
Ciências Biológicas.
Orientadora: Profª. Drª. Katia Castanho Scortecci Co-orientadora: Profª Drª. Adriana Ferreira Uchôa
NATAL – RN
2013

Barreto, Kellya Francisca Mendonça. Avaliação de parâmetros de estresse oxidativo em plantas de cana-de-açúcar tratadas com peróxico de hidrogênio / Kellya FranciscaMendonça Barreto. - Natal, 2013. 73f: il.
Orientadora: Profa. Dra. Katia Castanho Scortecci. Coorientadora: Profa. Dra. Adriana Ferreira Uchôa. Dissertação (Mestrado) - Universidade Federal do Rio Grande doNorte. Centro de Biociências. Programa de Pós-Graduação emCiências Biológicas.
1. Enzimas antioxidantes - Dissertação. 2. Sequenciamento depeptídeos - Dissertação. 3. Sinalização - Dissertação. I. Scortecci,Kátia Castanho. II. Uchôa, Adriana Ferreira. III. Universidade Federaldo Rio Grande do Norte. IV. Título.
RN/UF/BSE01 CDU 57
Catalogação da Publicação na FonteUniversidade Federal do Rio Grande do Norte - UFRN


3
Dedico esta obra ao meu marido Ezio Barreto Por sua infinita compreensão, sabedoria e apoio.
Sei que nos momentos mais difíceis, pude contar com você me encorajando e me fortalecendo.
E ao meu filho Kezio Barreto, que mesmo na sua mais tenra juventude, Por muitas vezes pude encontrar nos seus gestos e palavras
o mais profundo amor.

4
AGRADECIMENTOS
Inicialmente, agradeço a Deus por tudo em minha vida, mesmo sabendo que muitos negam a sua inexistência. Quero registrar aqui nesta obra, que ao estudar a biologia, contemplo a criação de Deus molecularmente e tento compreender a sua assinatura genômica em todos os seres viventes. Aos meus pais, Geraldo e Francisca por ter me ensinado a ser uma pessoa resiliente em meio às adversidades da vida; Aos meus irmãos, José Francisco e Klebya Suely e aos meus sobrinho(a)s por me acompanharem nesta jornada dia após dia, torcendo e me incentivando; Aos meus amigos da igreja que estiveram comigo e com a minha família em oração, muito obrigada; Aos colegas do grupo de plantas, que juntos, brotamos como uma semente, nos tornando uma árvore ou uma família. Árvore esta, que julgo ser a dos saberes multidisciplinar, por ter ramos em várias áreas, bem como, pesquisadores fortes, humildes e dedicados ao seu trabalho; À minha orientadora, Professora Katia Scortecci, pelos ensinamentos, amizade, simpatia e boa disposição e também, por confiar e acreditar em minha capacidade de superar os obstáculos que por vezes surgiram nesta trajetória; À minha co-orientadora Adriana Uchôa, por ter dividido os seus conhecimentos durante os experimentos que contribuiu muito para a minha formação acadêmica; Aos colegas do LBMG, que quando necessitei de ajuda, vocês sempre me atenderam com carinho, em especial, agradeço a Erika Castro, Renata Kaline e Helaine Cristiane pela contribuição científica que muitas vezes sequestrou o tempo de vocês. Ajuda preciosa que abrilhantou este trabalho ; Aos técnicos da UFRN, Artejose e Socorro, que sem a dedicação e empenho de vocês, não conseguiria obter estes resultados tão satisfatório;
Aos professores, Raquel Cordeiro Theodoro e Eduardo Luiz Voigt pela contribuição muito valorosa durante o exame de qualificação; Aos professores, Gilberto Sachetto Martins e João Paulo Matos Santos Lima por aceitar o convite para participar da banca de defesa. Á todos os funcionários que compõem o Depto de Biologia Celular e Genética –
UFRN;
CNPq REUNI FIOCRUZ CNPEM

5
RESUMO A cana-de-açúcar é uma das principais monoculturas no Brasil devido à importância dos seus produtos: etanol e açúcar. A produtividade pode variar entre as diferentes regiões do Brasil como sudeste e nordeste, por adaptações destas gramíneas as diferentes condições edafo-climáticas. As condições do solo e da temperatura da região Nordeste podem promover o estresse oxidativo, hídrico e consequentemente o florescimento precoce o que acarreta perdas consideráveis na produção. Neste trabalho, procurou-se averiguar as prováveis alterações morfofisiológicas, bioquímicas e moleculares resultantes da resposta das plantas ao estresse oxidativo. Deste modo, plantas de cana-de-açúcar foram cultivadas em condições controladas e quando atingiram dois meses foram submetidas a uma condição de hidropônia por 24 horas para sua aclimatação e em seguida ao estresse oxidativo realizado por 8 horas utilizando as concentrações de peróxido de hidrogênio: 0 mM; 10 mM; 20 mM e 30 mM. Após este período, foram observadas as seguintes alterações morfológicas para as folhas: o fechamento foliar parcial ou total de acordo com a concentração de peróxido de hidrogênio utilizada. As análises histomorfológicas permitiram verificar para ambos os tecidos (foliar e radicular) que houve alterações estruturais relacionadas ao aumento do volume das células propiciando, assim, a quebra das paredes das membranas dos vasos condutores xilema e floema entre outras modificações anatômicas em diferentes regiões, assim como a lignificação em algumas regiões. Também foi analisada a atividade enzimática das enzimas Superóxido dismutase - SOD; Catalase - CAT e Ascorbato peroxidase - APX. Foi observado um aumento na atividade da enzima superóxido dismutase nas raízes. Nessa abordagem bioquímica, foi também analisada a peroxidação de lipídeos que mostrou que ocorreram danos nas membranas das folhas e raízes principalmente para a concentração de 30 mM. Entretanto, para a enzima catalase foi observada uma baixa atividade em folhas e esta atividade não foi detectada nas raízes. Contudo, os resultados histomorfologicos juntamente com os resultados bioquímicos fortalecem que o tratamento com peróxido de hidrogênio pode ter alterado a homeostase redox e as vias de sinalização promovendo assim as alterações morfológicas (aumento de células, lignificação) e bioquímicas (EROs e antioxidantes). Outra ferramenta utilizada nesse trabalho foi a proteômica, onde foi utilizado o sequenciamento tanto por meio de géis bidimensionais como também pelo sequenciamento direto utilizando o EZI Q–TOF. Os resultados obtidos permitiram com essa abordagem identificar mais de 3.000 proteínas com o score variando de 100-5000 íons. Algumas das proteínas identificadas foram: light Harvesting; oxygen-evolving; Thioredoxin; Ftsh-like protein Pftf precusor; Luminal-binding protein; 2 cys peroxiredoxin e Lipoxygenase. Estas proteínas estão envolvidas no metabolismo em resposta ao estresse oxidativo, proteínas do aparelho fotossintético, além de proteínas hipotéticas e/ou desconhecidas (30% do total). Desta forma, os dados obtidos nos permitem propor uma hipótese, onde às proteínas identificadas estariam envolvidas em processos fisiológicos, na remodelação de tecidos, dano/degradação e síntese de anti-oxidante/desintoxicante, correlacionada aos dados de resposta de defesa enzimática, acúmulo de H2O2 nos tecidos ativando a peroxidação lipídica nas paredes das membranas das células. Desta forma ativando várias vias de cascata de sinalização. Palavras-chave: Anatomia; sequenciamento de peptídeos; sinalização, enzimas antioxidantes; Saccharum spp.

6
ABSTRACT
The genus Saccharum belongs to Poaceae family. Sugarcane has become important monocultures in Brazil due to their products: ethanol and sugar. The production may change between different regions from Brazil. This difference is related to soil, climatic conditions and temperature that promotes oxidative stress that may induce an early flowering. The aim of this work was to identify the effects of oxidative stress. In order to analyse this, sugarcane plants were submitted to oxidative stress using hydrogen peroxide. After this treatment, the oxidative stress were analyzed Then, the plant responses were analyzed under different approaches, using morphophysiological, biochemical and molecular tools. Thus, sugarcane plants were grown under controlled conditions and until two months they were subjected first to a hydroponics condition for 24 hours in order to acclimation. After this period, these plants were submitted to oxidative stresse using 0 mM, 10 mM, 20 mM and 30 mM hydrogen peroxide during 8 hours. The histomorphometric analysis allowed us to verify that both root and leaf tissues had a structural changes as it was observed by the increased in cell volume, lignin accumulation in cell walls. Besides, this observation suggested that there was a change in redox balance. Also, it was analyzed the activity of the SOD, CAT and APX enzymes. It was observed an increase in the SOD activity in roots and it was also observed a lipid peroxidation in leaves and roots. Then, in order to identify proteins that were differently expressed in this conditions it was used the proteomic tool either by bidimensional gel or by direct sequencing using the Q-TOF EZI. The results obtained with this approach identified more than 3.000 proteins with the score ranging from 100-5000 ions. Some of the proteins identified were: light Harvesting; oxygen-evolving; Thioredoxin; Ftsh-like protein Pftf precusor; Luminal-binding protein; 2 cys peroxiredoxin e Lipoxygenase. All these proteins are involved in oxidative stress response, photsynthetic pathways, and some were classified hypothetical proteins and/or unknown (30% of total). Thus, our data allows us to propose that this treatment induced an oxidative stress and the plant in response changed its physiological process, it made changes in tissue, changed the redox response in order to survival to this new condition. Keywords: Morphophysiological; peptides sequencing, signal pathways, antioxidant enzymes; Saccharum spp.

7
LISTA DE ABREVIATURAS
ABA – Ácido abscísico
APX – Ascorbato peroxidase
AT – Azul de toluidina
CAT - Catalase
CIB – Conselho de Informações sobre Biotecnologia
DAB - Diaminobenzidina
FAA – Formaldeído, álcool e ácido acético
FAO – Organização das Nações Unidas para Alimentação e Agricultura
IBGE – Instituto Brasileiro de Geografia e Estatística
MEV – Microscopia Eletrônica de Varredura
N2 – Gás nitrogênio
NBT - Nitro-blue tetrazolium
NPK – Nitrogênio, Fósforo e Potássio
PAR – Luminosidade disponível para a fotossíntese
PMSF – Phenylmethylsulfonyl fluoride
EROs – Espécies Reativas de Oxigênio
SOD – Superóxido dismutase

8
LISTA DE FIGURAS
Figura 1: Esquema do mecanismo de manutenção do equilíbrio das EROs.................20 Figura 2: Vista área da Usina Estivas, Goianinha/RN....................................................23 Figura 3: Desenho esquemático representando o desenvolvimento das plantas de cana-de-açúcar utilizadas para o tratamento oxidativo.............................................25 Figura 4: Esquema das amostras dos tecidos para diferentes tratamentos..................26 Figura 5: Tratamento de hidropônia em cana-de-açucar...............................................27 Figura 6: Determinação do conteúdo de peróxido de hidrogênio na solução................29 Figura 7: Parafinização dos colmos................................................................................35 Figura 8: Gráfico de determinação de H2O2 por permanganometria..............................37 Figura 9: Alterações morfológicas das folhas de cana-de-açúcar tratadas com peróxido de hidrogênio em diferentes concentrações (0, 10, 20 e 30 mM)............38 Figura 10: Conteúdo de peróxido de hidrogênio em meristema de plantas de cana-de-açúcar submetidas ao tratamento com concentrações crescentes de peróxido de hidrogênio (10, 20 e 30 mM) e controle (sem peróxido).................................................39 Figura 11: Gráficos de peroxidação lipídica (TBARS)....................................................41 Figura 12: Secção transversal da folha de cana-de-açúcar nas condições normais...........................................................................................................................43 Figura 13: Secção transversal da raiz de cana-de-açúcar nas condições normais...........................................................................................................................43 Figura 14: Análise das folhas de cana-de-açúcar tratadas com o corante Safranina........................................................................................................................44 Figura 15: Análise das raízes de cana-de-açúcar pelo corante safranina.........................................................................................................................45 Figura 16: Análise das folhas de cana-de-açúcar tratadas com o corante Eosina de Metileno..........................................................................................................................46 Figura 17: Análise das raízes de cana-de-açúcar pelo corante Eosina de Metileno..........................................................................................................................47 Figura 18: Análise das folhas de cana-de-açúcar com a coloração Diaminobenzidina (DAB)..............................................................................................................................48 Figura 19: Análise das Raízes de cana-de-açúcar pela a coloração Diaminobenzidina (DAB) e cloreto de zinco iodado (CZI)............................................................................49

9 Figura 20: Análise das folhas de cana-de-açúcar pelo corante cloreto de zinco iodado.............................................................................................................................50 Figura 21: Análise em MEV das folhas...........................................................................52 Figura 22: Análise em MEV das folhas...........................................................................52 Figura 23: Análise em MEV das raízes..........................................................................53 Figura 24: Gráficos da atividade enzimática de ascorbato peroxidase (APX)................55 Figura 25: Gráficos da atividade enzimática de catalase (CAT) ....................................56 Figura 26: Gráficos da ação do superoxido dismutase (SOD).......................................58

10
LISTA DE TABELAS
Tabela 1: Relação de corantes..........................................................................31

11
SUMÁRIO
1. INTRODUÇÃO .......................................................................................................... 13
1.1. Cana-de-açúcar .................................................................................................. 13
1.2. Morfologia e mecanismos de concentração de CO2, ciclo C3 e C4 .................... 15
1.3. Fisiologia e Espécies reativas de oxigênio (EROs) e estresse. .......................... 17
1.4. Contribuições do estudo: estresse oxidativo em cana-de-açúcar ....................... 21
2. OBJETIVO ................................................................................................................ 22
2.1. Objetivo Geral ..................................................................................................... 22
2.2. Objetivos Específicos .......................................................................................... 22
3. METODOLOGIA ...................................................................................................... 23
3.1. Local da coleta .................................................................................................... 23
3.2. Protocolo de cultivo ............................................................................................. 23
3.2.1. Tratamento de hidroponia e peróxido de higrogênio ........................................ 26
3.2.2. Determinação do H2O2 por permanganometria ................................................ 28
3.2.3. Determinação do conteúdo de H2O2 nos tecidos. ............................................ 29
3.3. Análise histológica .............................................................................................. 29
3.3.1. Solução fixadora .............................................................................................. 29
3.3.2. Técnicas de emblocagem ................................................................................ 30
3.3.3. Microscopia Eletrônica de Varredura ............................................................... 31
3.4. Ensaios enzimáticos ........................................................................................... 31
3.4.1. Extração de proteínas solúveis totais .............................................................. 32
3.4.2. Determinação da atividade de ascorbato peroxidase (APX) ............................ 32
3.4.3. Determinação da atividade superóxido dismutase (SOD) ................................ 32
3.4.4. Determinação da atividade de catalase (CAT) ................................................. 33
3.4.5. Estimação da peroxidação de lipídios pelo (TBARS) ....................................... 33
3.4.6. Análises estatísticas ......................................................................................... 34
4. RESULTADOS .......................................................................................................... 35
4.1. Tratamento com peróxido de hidrogênio ............................................................. 35
4.1.2. Determinação do conteúdo de peróxido de hidrogênio nos tecidos ................. 38
4.1.3. Espécies reativas ao ácido 2 – Tiobarbitúrico (TBARS) ................................... 40
4.2. Análises histológicas de folha e raiz .................................................................. 42
4.2.1. Análise morfológica por microscopia óptica ..................................................... 42
4.2.2. Análise dos tecidos de folha e raízes por microscopia eletrônica de varredura.51
4.3. Análises enzimáticas........................................................................................... 54
4.3.1. Ascorbato peroxidase (APX) ............................................................................ 54
4.3.2. Catalase (CAT) ................................................................................................ 56
4.3.3. Superóxido dismutase (SOD) .......................................................................... 57
5. DISCUSSÃO ............................................................................................................. 59
6. CONCLUSÃO ........................................................................................................... 63

12 8. REFERÊNCIAS ......................................................................................................... 64

13
1. INTRODUÇÃO
1.1. Cana-de-açúcar
A cana-de-açúcar, pertencente à família Poaceae, foi à primeira espécie vegetal
introduzida no Brasil proveniente da Ilha da Madeira em 1502. A espécie Saccharum
officinarum L. foi caracterizada pelo seu alto teor de açúcar, porte elevado, colmo
grosso e pouco teor de fibras. Atualmente, ela é cultivada comercialmente em mais de
70 países, sendo os maiores produtores, o Brasil e a Índia. Assim, a cana-de-açúcar
vem sendo utilizada como matéria prima para a produção de açúcar e álcool (LIMA,
1984 apud MACHADO, 2009). Tornando-se, uma das mais importantes espécies de
vegetais cultivadas no mundo e em especial nas regiões tropicais e subtropicais,
alcançando uma área superior a 20 milhões de hectares em 101 países (FAO, 2010). O
Brasil é considerado o maior produtor mundial de etanol e açúcar, chegando a produzir
por ano 14 bilhões de litros de álcool e 24 milhões de toneladas de açúcar
(GOLDENBERG, 2007).
Com a crise do petróleo, surgiu no mercado à importância do desenvolvimento de
novas fontes de energia renováveis (SCORTECCI, 2012). Sendo assim, a cana-de-
açúcar veio a ser uma cultura de grande importância geradora de biocombustível
renovável de primeira geração, sendo utilizada assim para a produção de etanol e
biomassa, como fonte alternativa de energia (WANG et al., 2010). Entretanto, estudos
para compreender seu metabolismo fisiológico, a resistência a condições de estresse
biótica e abiótica, a resposta de defesa e vias de sinalização têm sido o foco dos
pesquisadores nos últimos anos para o melhoramento genético da cana-de-açúcar
(SCORTECCI, 2012).
O cultivo desta gramínea tornou-se bastante complexo, uma vez que as
variedades cultivadas atualmente correspondem a híbridos obtidos do cruzamento
entre Saccaharum officinarum (alto teor de açúcar) e Saccharum spontaneum (alta
resistência a doenças) (BERDING & ROACH, 1987). A complexidade do genoma
(elevado nível de ploidia) devido aos diferentes cruzamentos realizados durante o
melhoramento genético clássico tem causado uma morosidade no progresso
dosestudos genéticos da cana-de-açúcar (SREENIVASAN et al., 1987). Os atuais
híbridos de cana-de-açúcar apresentam o valor de 2n= 100–130 (n= número de
cromossomos haplóides) cromossomos derivado de uma combinação de S. officinarum
(2n= 80 cromossomos) e S. spontaneum (2n=40-128 cromossomos) (BUTTERFIELD et
al., 2001; D'HONT & GLASZMANN, 2001). Considerando o genoma monoplóide, a

14 cana-de-açúcar apresenta um tamanho do genoma em torno de 750 a 930 milhões de
pares de bases - Mbps, este valor pode ser comparável à do sorgo (~ 760 Mbp) e cerca
de duas vezes a do arroz (~ 430 Mbp) (D'HONT & GLASZMANN, 2001).
Atualmente, o estado de São Paulo vem sendo responsável por 87% de sua
produção nacional. Em 2011, este estado produziu 21 milhões de toneladas de açúcar
e 11 milhões de metros cúbicos de etanol, representados por 58% e 51% do total
produzido no Brasil (IBGE, 2012). Essa liderança na produtividade é devido a alguns
fatores peculiares dessa região. Um desses fatores é o tipo de solo com uma
composição argilosa rica em minerais e nutrientes que favoreceu o desenvolvimento
dessas gramíneas (SETZER, 1991). Em 2001 e 2011, a produção paulista de açúcar
cresceu 121% e a produção de etanol 82% incentivada pelo mercado estadual de
biocombustíveis (IBGE, 2012). Segundo CONAB (2011), a previsão de produção de
açúcar na safra 2012/13 é de 37,66 milhões de toneladas, deste total, 70,03%
produzido na Região Sudeste, 10,21% na Região Centro-Oeste, 8,83% na Região Sul
e 09,87% na Região Nordeste (CONAB, 2013). Estes valores fazem do Brasil o maior
produtor mundial de açúcar, respondendo unicamente por 45% de todo o produto
comercializado no mundo (SCORTECCI, 2012). Além disso, o Brasil é o segundo
maior em produção de etanol e divide o Ranking de maior produtor mundial com
Estados Unidos da América. Os dois países são responsáveis por 70% de todo etanol
produzido no planeta. Vale a resaltar que, o produto norte-americano é exclusivamente
derivado do milho e voltado para o comércio interno (CIB, 2009).
Com base nos dados acima, a região Nordeste, que foi um tradicional produtor,
contribui atualmente somente com 09,87% da produção nacional devido ao clima, solo
e o tempo prolongado de seca (CONAB, 2013). Estes valores são decorrentes do
Estado do Rio Grande do Norte situada na região Nordeste apresentarem um solo do
tipo arenoso, o que aumenta as condições de seca porque os grãos são muito porosos
e permeáveis, facilitando assim a passagem da água e sais minerais, impossibilitando
a permanência destes na superfície. Sendo este considerado um solo pobre em
nutrientes, consequentemente, desfavorece a produtividade de espécies vegetais não
resistentes a estas condições (CUNHA, 2010). Assim, conhecer a morfologia vegetal e
como a alteração climática afeta o seu desenvolvimento e produtividade é de extrema
importância econômica considerando que a cana-de-açúcar é de grande relevância
socioeconômica para o Brasil, em especial a região Nordeste.

15 1.2. Morfologia e mecanismos de concentração de CO2, ciclo C3 e C4
Um dos grandes passos evolutivos que possibilitou a existência de vida na terra
foi o surgimento de organismos que contém a capacidade da realização
fotossintetizante (PETER et al., 2011). Por décadas os organismos fotossintéticos tem
despertado a curiosidade dos pesquisadores que buscam a compreensão e o
conhecimento destes seres vivos quanto a sua forma estrutural e metabólica, por ser
muito importante para a manutenção vital da terra (GREER, 2012 apud PURY, 1997).
Tal adaptação estrutural e metabólica promoveu o surgimento de várias plantas
ambientadas ao seu nicho ecológico como as do tipo C3 e as plantas C4. A descoberta
da fixação de CO2 em 3-fosfoglicerato, como sendo, um intermediário na fotossíntese
foi determinante para a classificação das plantas nas quais este composto de três
carbonos é o primeiro intermediário. Essas plantas foram, então, chamadas de plantas
C3. Dentre as plantas C3 podemos encontrar algumas com valor comercial agronômico
como as espécies de trigo (MARCINSKA et al., 2013); arroz (XU et al., 2013) e soja
(XU et al., 2013). As plantas C4 desenvolveram mecanismos diferentes de fixação e
armazenamento do CO2, o que permitiu reduzir a taxa de fotorrespiração, melhorando
assim a adaptação destas espécies a condições de alta temperatura (GOWIK &
WESTHOFF, 2011). Entre alguns exemplos de plantas C4 podemos citar algumas
espécies de importância agronômica como milho, cana-de-açúcar, sorgo, Miscanthus e
Panicum virgatum. Estas duas últimas estão sendo pesquisadas como potenciais
modelos para a conservação do solo, produção de etanol e butanol na América do
Norte, bem como futuros projetos de fitorremediação e produção de energia elétrica
(LUDWIG, 2013).
Embora as plantas da família Poaceae, em destaque a cana-de-açúcar que é do
tipo C4 compreendem menos de 4% do total das espécies de plantas terrestres, estas
também são importantes no equilíbrio das três esferas da biologia: ecológico, agrícola e
atmosfera, por este último às plantas C4 são capazes de capturar um maior número de
CO2 atmosférico, contribuindo para a purificação do ar (EHLERINGER et al, 1997).
As plantas C3 e C4 além de apresentarem mecanismos diferentes para a fixação
do CO2 na fotosíntese bem como, diferenças estruturais. Dentre estas, podemos citar
as folhas das plantas C3 não apresentam uma disposição concêntrica nas células
parenquimáticas e nas células da bainha do feixe. Outra diferença estrutural nas folhas
das plantas C4 corresponde à presença de dois tipos distintos de células contendo
cloroplastos: o mesófilo e a bainha do feixe vascular, as quais são conectadas por uma

16 extensa rede de plasmodesmas. As células da bainha do feixe apresentam uma
anatomia diferenciada de forma que os tecidos estão dispostos em círculo em relação
aos tecidos vasculares do xilema e floema. Este padrão estrutural é conhecido como
anatomia de Kranz (MUHAIDAT, et al., 2007).
Bioquimicamente, as plantas C3 apresentam apenas o ciclo de Calvin, onde o
primeiro intermediário estável é o 3-fosfoglicerato, com três carbonos. Uma observação
importante é que nas plantas C3 a fotossíntese é realizada em um compartimento
celular denominado célula do mesófilo. Para isto, recebe ajuda da proteína rubisco,
porém, esta molécula não tem especificidade absoluta pelo CO2 como substrato, sendo
assim, o oxigênio molecular O2 também compete com o CO2 no sítio ativo e cerca de
uma vez a cada três ou quatro giros, a proteína rubisco catalisa a condensação do O2
com a ribulose-1,5-bifosfato para formar 3-fosfoglicerato e 2-fosfoglicolato, este último
parece ser um produto metabólico sem aplicação celular. Como resultado desta reação
com o O2, não ocorre fixação de carbono e tal reação parece ser desvantajosa para a
célula (KIRCHHOFF et al., 2013).
Contrastando com isso, as plantas denominadas C4 apresentam o processo de
fotossíntese dividida em dois compartimentos celulares, seguida de quatro fases, na
qual a primeira ocorre à fixação de CO2 com o substrato fosfoenolpiruvato-carboxilase
(HCO3) pela enzima carboxilase do fosfoenolpiruvato (PEP-Carboxilase) no citosol das
células do mesófilo, formando oxaloacetato. Este ácido orgânico é convertido então
para malato. Estes ácidos de quatro carbonos são os primeiros intermediários estáveis
da fotossíntese das plantas C4 (KIRCHHOFF et al., 2013). Na segunda fase, os ácidos
de quatro carbonos são translocados das células do mesófilo para as células da bainha
do feixe vascular, via plasmodesmas. Na terceira etapa, estes ácidos de quatro átomos
de carbono são descarboxilados nas células da bainha, liberando assim a molécula de
CO2 e produzindo a molécula de piruvato. A molécula de CO2 é então fixada pela
enzima ribulose 1,5-bifosfato carboxilase/oxigenase (Rubisco), que é encontrada
somente nas células da bainha nas plantas C4. Por fim, através do transporte do
piruvato de três carbonos para o mesófilo ocorre a regeneração do fosfoenolpiruvato
(PEP) com gastos de moléculas de ATP (GOWIK & WESTHOFF, 2011).
Além disso, a enzima Rubisco é encontrada apenas nas células da bainha
vascular, portanto as plantas C4 gastam menos nitrogênio do que as plantas C3
(VOZNESENSKAYA et al., 2001). A enzima PEP carboxilase tem elevada afinidade
pelo substrato HCO3, o que a permite atuar em baixas concentrações do substrato,
além do que a enzima fosfoenolpiruvato carboxilase utiliza como substrato o HCO3 que

17 não compete com a molécula de O2, e a fotorrespiração é suprida na região do
mesófilo (GOWIK & WESTHOFF, 2011). Como desvantagem, o mecanismo de
regeneração da enzima PEP carboxilase consome dois ATP, com isso as plantas C4
gastam 5 ATP para cada carbono fixado, enquanto as plantas C3 gastam apenas 3
ATP por CO2 fixado. Apesar deste maior consumo de ATP, o mecanismo de
fotossíntese C4 é considerado bastante eficiente nas condições de clima tropical
porque praticamente reduz a fotorrespiração e apresentam taxas de fotossíntese
líquida bem superior às das espécies C3 (DAL’MOLIN et al., 2010).
Com base neste estudo dos mecanismos de fixação de carbono, as plantas C4
apresentam vantagens para o cultivo nas regiões do Nordeste por serem capaz em se
adaptar em ambientes quentes e secos (SAGE, 2003). Desta forma, estas plantas
mostram-se mais eficientes na utilização da água que as plantas C3. Esta eficiência
está relacionada com o mecanismo de fixação de CO2, mesmo quando estes mantêm
seus estômatos fechados. Assim, a perda de água por transpiração é bem reduzida
(LARA & ANDREO, 2011).
1.3. Fisiologia e Espécies reativas de oxigênio (EROs) e estresse.
O crescimento da cana-de-açúcar é um processo fisiológico e complexo,
formado por vários estágios de desenvolvimento, e cada estágio tem a sua própria
necessidade ambiental e fisiológica. Fatores externos relacionados com o crescimento
são: altas ou baixas temperaturas, radiação solar, alta exposição ou escassez de água,
umidade e fertilidade do solo, entre outros (ARALDI et al., 2010; GRAY, 2012 ). Além
desses fatores, o solo arenoso e a alta salinidade uma consequencia da irrigação
descuidada ou sem acompanhamento podem desencadeiar respostas de sinalização
envolvendo diversas vias, que podem aumentar o acúmulo de espécies reativas de
oxigênio (EROs) (VOIGT et al., 2012).
Assim, todos estes tipos de agentes estressores podem elevar o aumento das
espécies reativas de oxigênio (EROs) que são produzidos naturalmente no processo
metabólico da célula em resposta ao estresse. Estas espécies também podem ser
geradas durante a respiração, fotorespiração e fotossíntese (SAGE, 2003). As espécies
reativas de oxigênio em: (1O2) oxigênio singlete, (O2*-) radical ânion superóxido, (OH)
radical hidroxila e peróxido de hidrogênio (H2O2) (GILL & TUTELA, 2010). A molécula
de H2O2 nas células vegetais, normalmente desempenha um duplo papel como um
subproduto tóxico do metabolismo celular e como uma molécula reguladora na
percepção de estresse e transdução de sinal (MACEDO et al., 2009). Assim, para as

18 células é importante manter este equilíbrio entre produção e degradação das EROs.
Considerando que, em plantas a manutenção deste balanço é importante, desta forma,
existem vários mecanismos com enzimas antioxidantes para reduzir estes produtos
(MYOUNG, 2010 apud WAN & LIU, 2008). As principais enzimas que atuam na
redução das espécies reativas de oxigênio são: a enzima ascorbato peroxidase (APX),
catalase (CAT), superóxido dismutase (SOD), glutationa peroxidase (GPx) e
peroxirredoxina (Prxs). Além disso, as células vegetais possuem moléculas como o
ácido ascórbico e glutationa que funcionam como substâncias antioxidantes
eliminando, em condições normais, a molécula de H2O2 reduzindo-a em água e
oxigênio livre (H2O2 = H2O + O2) (Figura 1) (PATADE & SUPRASANNA, 2010).
Entretanto, este equilíbrio entre a produção e a eliminação do H2O2 pode ser
perturbado por vários tipos de estresses bióticos e abióticos, influenciando assim a um
aumento rápido e transitório dos níveis de H2O2 intercelulares. Quando o nível da
molécula de H2O2 atinge um limiar, muitas vezes os tecidos e células sofrem danos
oxidativos que podem vir a desencadear, um distúrbio nos processos fisiológicos e nas
vias metabólicas, denominadas estas condições como estresse oxidativo (Figura 1)
(GILL & TUTEJA, 2010).
Diversos estudos têm observado que em plantas expostas a condições de
estresse como seca; frio; salinidade; a radiação UV; metais pesados; ferimentos;
hormônios; incluindo o ácido abscísico ou ácido jasmônico e patógenos utilizam a
molécula de H2O2 como sinalizador e assim ativam a expressão de alguns genes
responsivos que permitem que a planta possa ser tolerante a estas condições (Figura
1) (WAN & LIU, 2008).
Assim, as EROs são produtos do próprio metabolismo da célula vegetal sendo
produzidas, por exemplo, nos compartimentos celulares como: cloroplasto,
peroxissomos e mitocôndria. Nos cloroplastos, os fotossistemas I e II (PSI e PSII)
promovem à síntese de 1O2 e O2 durante o processo de fotossíntese. Na mitocôndria, o
complexo I, ubiquinona, o complexo III e a cadeia transportadora de elétrons são os
principais locais para a geração de EROs. Com isto, estes exemplos mostram que
normalmente estas espécies são produzidas na célula. Em contrapartida, as células
apresentam uma maquinaria produtora de moléculas antioxidantes capaz de reduzir os
efeitos das EROs. Porém, estas na presença de íons metálicos como o ferro (Fe), por
exemplo, ativam uma sequência de reações que levam a formação de um hidróxido
através da reação do Ciclo de Haber-Weiss que produz um hidroxila (OH-) (QURESHIA
et al., 2007). Existem, outras moléculas, que trabalham paralelo ao mecanismo

19 enzimático de defesa, também colaboram para a degradação das espécies reativas
como a: glutationa redutase (GR); monodehidroascorbato redutase (MDHAR);
dehidroascorbato redutase (DHAR); guaiacol peroxidase (GOPX) e glutationa
transferase (GST), além de moléculas antioxidantes que não apresentam atividade
enzimática como: ácido ascórbico (AA) e o sistema de defesa de compostos fenólicos,
alcalóides, aminoácidos não proteicos e alfa tocoferol (ASH); glutationa (GSH) em que
todos servem para reparar algum desequilíbrio na cascata de oxidação, assim
protegendo as células dos danos oxidativos pela eliminação das EROs (Figura 1) (GILL
& TUTELA, 2010).
Deste modo, para compreender a complexidade do metabolismo celular vegetal
visando correlacionar respostas de sinalização e síntese de EROs que geram a
produção de antioxidantes é importante a realização de estudos utilizando as
ferramentas genômica e proteômica. Estas técnicas permitem a identificação de genes
responsivos e proteínas diferencialmente expressas a estas condições de estresse
oxidativo (GIRAUD et al., 2012).

20
Figura 1: Esquema do mecanismo de manutenção do equilíbrio das EROs. A planta sofre o estresse abiótico que nas suas células, as organelas peroxissomo, cloroplasto e mitocôndria vão obter respostas enzimáticas e não-enzimáticas. Com o estresse ocorre o aumento de EROs, na primeira linha de resposta, as enzimas SODs removem o O2
.- catalisando a dismutação de um O2 reduzindo a H2O2 e outro oxida para O2 evitando a formação de OH pelo ciclo de Heber. CATs
são enzimas tetraméricas contendo um potencial de desmutar H2O2 em H2O e O2. APX está envolvida na eliminação de H2O2 em H2O e ciclos de α-tocoferol e glutationa - ASH-GSH como doador de elétrons. Glutationa redutase - GR e uma flavoproteína oxidorredutase catalisa a redução da GSH, uma molécula envolvida na regulação metabólica e processos antioxidantes, a GR e também catalisa NADPH, uma reação dependente de sulfeto, vinculado ao substrato de duas moléculas de glutationa – GSSG. Portanto, a importância para a manutenção desse sistema é vital para a planta. A seta em negrito indica um desequilíbrio de EROs, causando na planta danos moleculares que resultam na morte celular (ADAPTADA E MODIFICADA DE GILL & TUTELA, 2010).

21
1.4. Contribuições do estudo: estresse oxidativo em cana-de-açúcar
O crescimento populacional e a queda da fertilidade dos solos utilizados
por séculos de sucessivos plantios causaram entre outros problemas, a
redução nos alimentos e rápidas mudanças nos padrões de consumo. Sendo
assim, para alimentar as futuras gerações as taxas de produção terão que ser
aumentadas em média 50% até 2050. Para isso, o século XXI conta com os
avanços tecnológicos que estão alicerçados por auspicioso conhecimento
científico na área da biologia molecular, marcadores e melhoramento genético
contribuindo assim para a melhoria do rendimento do alimento (grãos, folhas,
etc) por hectare (MURCHIE et al., 2008). Com esta problemática, a
biotecnologia vem tendo cada vez mais importância para unir todos estes
fatores para que no final ocorra o aumento da produtividade vegetal. E para
isto, diferentes ferramentas têm sido utilizadas como a genômica, proteômica e
metabolômica (GIRAUD et al., 2012). Considerando que, um dos grandes
problemas da região Nordeste é o estresse hídrico, salino e a presença do solo
arenoso e que todos estes estresses abióticos podem promover o aumento das
EROs, este trabalho teve o intuito de analisar o efeito do tratamento de plantas
de cana-de-açúcar com peróxido de hidrogênio.
Atualmente, vários trabalhos mostram que quando as plantas são
submetidas a estresse hídrico, salino, alta intensidade de luz, entre outros
culminam em desbalanço/desequilíbrio do estado redox , o qual desencadeia
um processo metabólico de respostas nas células vegetais que até o momento
não é completamente compreendido (GAD et al., 2010).
Com base em todos os dados mencionados acima, neste trabalho foi
abordada à questão do estresse oxidativo em plantas de cana-de-açúcar
utilizando o peróxido de hidrogênio como um agente estressor com intuito de
identificar genes e proteínas diferencialmente expressas nesta condição. Os
dados obtidos poderão auxiliar na compreensão dos mecanismos de
sinalização e defesa voltada para a cana-de-açúcar, aplicando este
conhecimento para a identificação de marcadores que possam auxiliar no
melhoramento genético como na obtenção de plantas tolerantes a este fator.

22
2. OBJETIVO
2.1. Objetivo Geral
Prospectar genes e proteínas em resposta antioxidante ao estresse
oxidativo por meio do tratamento com peróxido de hidrogênio em plantas de
cana-de-açúcar.
2.2. Objetivos Específicos
Padronizar o protocolo de cultivo em laboratório para cana-de-açúcar;
Estabelecer um protocolo para tratamento das variedades de cana-de-
açúcar com H2O2;
Realizar análises anatômicas do material tratado e controle;
Analisar o padrão da atividade das proteínas SOD; CAT e APX por meio
de ensaios enzimáticos;
Padronizar o protocolo para extração de proteínas totais em tecidos de
raiz e folha;
Analisar o padrão de expressão diferencial proteica por eletroforese
bidimensional;
Identificar os spots isolados e sequenciar os peptídeos;

23
3. METODOLOGIA
3.1. Local da coleta
As plantas de cana-de-açúcar foram coletadas na Usina Estivas, situada
em Goianinha, no estado do Rio Grande do Norte (figura 2) (06°16′08″S e
35°12′32″W). A coleta e seleção dos exemplares foram iniciadas em janeiro de
2011-2013.
Ao todo foram realizadas mais de nove coletas nos campos da usina
Estivas. E com as plantas germinadas foram realizadas mais de cinco testes
com peróxido de hidrogênio sob condições de hidropônia e para as análises
envolvendo a extração proteica, atividade enzimática, anatomia e histologia.
Todas essas análises foram realizadas em triplicatas.
Figura 2: Vista área da Usina Estivas, Goianinha/RN. Fonte: (www.estiva.com.br).
3.2. Protocolo de cultivo
As etapas seguintes ocorreram na sala de cultivo de plantas no
laboratório de Biologia Molecular e Genômica (LBMG), no Centro de
Biociências da Universidade Federal do Rio Grande do Norte (UFRN). Para se
obter plantas de cana-de-açúcar viáveis, foi importante inicialmente padronizar
um protocolo de cultivo de plantas em condições de laboratório. Os colmos

24
trazidos para o laboratório foram cortados em colmos menores, estes contendo
de dois a três entrenós. Depois estes foram lavados com água corrente e
sabão neutro. Em seguida, estes tiveram as suas extremidades preenchidas
com parafina.
Depois disso, os colmos foram plantados em bandejas com a capacidade
para 4,5 L; medindo 6 cm de profundidade; 33 cm de comprimento por 28 cm
de largura, em cada bandeja foram plantados seis colmos, totalizando 50
colmos cultivados em substrato vermiculita expandida fina (partículas com
diâmetro entre 0,15 e 8,0 mm; 2 L de volume por bandeja) (BRASIL, 1992).
Sistematicamente, a cada 20 dias foram adicionados ao substrato, fertilizante
com água pura e NPK–nutriente contendo em sua composição Nitrogênio;
Fósforo e Potássio na proporção de 10:10:10 – (Heringer) e crescidas sob
condições de luz fluorescente branca e complementada na faixa do vermelho
com lâmpadas Grolux (PAR 250 ~μmol.m-2.s-1). A irrigação ocorreu em dias
intercalados e todas as plantas estavam sob a mesma condição de
temperatura (25-27°C), umidade relativa do ar (62%-65%) e fotoperíodo (12h
claro e 12h escuro). A figura 3 mostra o esquema de desenvolvimento das
plantas a partir do plantio até dois meses de idade.

25
Figura 3: Desenho esquemático representando o desenvolvimento das plantas de cana-de-açúcar utilizadas para o tratamento oxidativo. Em (1) é representado um colmo contendo duas gemas, (2) é representado o colmo em estado de pré-germinação, (3) Colmo apresentando perfilho primário e raízes do colmo e do perfilho, (4) colmo contendo planta com germinação de emergência ou brotamento, (5) colmo com planta no estado de formação foliar e (6) planta de cana-de-açúcar propriamente dita em estágio de formação completa com 60 dias de desenvolvimento. (Fonte: acervo pessoal).

26
3.2.1. Tratamento de hidroponia e peróxido de higrogênio
Plantas de cana-de-açúcar com 60 dias de desenvolvimento foram
selecionadas aleatoriamente e submetidas ao tratamento hidropônico e
posterior tratamento de estresse oxidativo (Figura 5). Para isso, as plantas
foram lavadas com água corrente para retirar o excesso do substrato
vermiculita e subsequentemente, foram selecionadas 25 plantas por
tratamento, estas foram fixadas com arame e organizadas nas bandejas
contendo água pura na presença de um areador (HONG RI HR-028) e
submetido por um período de 24 horas ao tratamento hidropônico para
aclimatação. Posteriormente, essa água pura foi descartada e adicionou-se
água pura mais as seguintes concentrações de peróxido de hidrogênio: 0 mM,
10 mM, 20 mM e 30 mM em cada bandeja contendo os colmos parafinizados.
Este tratamento foi realizado durante 8 horas a uma temperatura de 25-27 °C.
Decorrido este período, as plantas foram retiradas, isoladas e congeladas em
nitrogênio líquido e armazenadas no freezer -80 °C (Figura 4).
Figura 4: Esquema das amostras dos tecidos para diferentes tratamentos. Representação esquemática das amostras utilizadas para diferentes etapas: cultivo, tratamento hidropônico e estresse oxidativo para obter as análises das atividades enzimáticas, extração proteica, sequenciamento de proteínas e RNA mensageiro de forma a verificar os efeitos metabólicos no tratamento oxidativo (Fonte: Acervo pessoal).

27
Figura 5: Tratamento de hidropônica em cana-de-açúcar. (1) Bandeja contento água pura e os colmos, (2) suporte fixador de colmos, (3) oxigenador e (4) plantas com 60 dias de desenvolvimento.

28
3.2.2. Determinação do H2O2 por permanganometria
A determinação de peróxido de hidrogênio seguiu os seguintes passos:
em um Erlenmeyer de 500 mL contendo 100 mL da solução ácido sulfúrico
(H2SO4) foi adicionado com auxílio de uma bureta de 50 mL com o gotejamento
da solução de permaganato sob agitação manual até o ácido atingir a cor
violeta. A mudança da cor da solução incolor para violeta indica a saturação
que é nomeado de ponto final. Assim determinou-se o volume da saturação do
peróxido de hidrogênio para todos os tratamentos em questão. Para isso, a
cada uma hora foram retirados 50 mL da solução (H2O+H2O2) das bandejas
sob hidroponia. No total, foram retiradas nove amostras para a medição da
absorção do peróxido pela planta. O mesmo processo foi repetido em bandejas
conténdo apenas a solução (H2O mais H2O2) com suas respectivas
molaridades sem a presença de plantas, onde nove amostras foram analisadas
a fim de obter um parâmetro comparativo, totalizando dezoito amostras.
Os 50 mL das amostra foram misturadas a 100 mL de uma solução ácida
mantendo-se na cor incolor. Lentamente, cerca de 10 minutos, foi adicionado
gota a gota da solução de permanganato até atingir o ponto final, representado
pela cor violeta. Assim, titulando em mL na bureta, a absorção de peróxido de
hidrogênio da amostra.
Para o preparo das soluções como os cálculos para o permanganato;
oxalato de sódio (Na2C2O4) e ácido sulfúrico para a obtenção desta titulação,
foram seguidas conforme SOLVAY Peroxidos do Brasil Ltda.
(www.peroxidos.com.br) (Figura 6).

29
Figura 6: Determinação do conteúdo de peróxido de hidrogênio na solução. A titulação foi obtida como mostra a figura (A) em 100 mL de meio ácido mais 50 mL (água pura mais peróxido) retirados dos tratados, a amostra foi mantida em gotejamento de solução padronizada de permanganato de potássio, sob agitação manual; (B) mostra a continuidade do gotejamento na solução, revelando o momento da saturação evidenciada pela cor violeta, esta mantida por mais de 30 segundos. Assim determinou-se o volume da saturação do peróxido de hidrogênio para cada tratado (Fonte: www.peroxidos.com.br).
3.2.3. Determinação do conteúdo de H2O2 nos tecidos.
O conteúdo de H2O2 nos tecidos de folha, raiz e meristema foi mensurado
a partir dos extratos obtidos com 1% TCA (p/v) (item 3.1), por meio da reação
específica do H2O2 com o iodeto de potássio (KI) (VELIKOVA; YORDANOV;
EDREVA, 2000; ALEXIEVA et al., 2001; JALEEL et al., 2008; SILVA, 2010;
TORRES et al., 2013). Em tubos de ensaio, foi aplicado 100 µL de extrato,
diluídos com mais 400 µL de TCA 1% e, em seguida, adicionou-se 0,5 mL de
tampão fosfato de potássio 10 mM, pH 7,0, e 1 mL de solução de KI 1 M. Os
tubos contendo os ensaios foram colocados no escuro e após 30 mim, foram
lidos a 390 ηm contra um branco de referência composto de 1% TCA. O
conteúdo foi calculado a partir de uma curva padrão da solução reagente de
KI, realizada com soluções de diferentes concentrações de H2O2, preparadas
em 1% TCA e os resultados obtidos, expressos em µmol H2O2 . g-1 MF.
3.3. Análise histológica 3.3.1. Solução fixadora
Para as análises anatômicas foram utilizados a parte média dos tecidos
de folha e raiz de cana-de-açúcar que foram fixados em solução de FAA

30
(formaldeído, álcool, etílico 70% (V/V) e ácido acético - 1:18:1 respectivamente)
e foram mantidos por 48 horas em temperatura ambiente. Decorrido este
tempo, a solução de FAA foi substituída por etanol 70% até o momento da sua
utilização (KRAUS & ARDUIN, 1997).
3.3.2. Técnicas de emblocagem
Após a fixação no FAA, os tecidos vegetais foram desidratados em
concentrações crescentes de álcool etílico (70 – 100%). Depois o material foi
submetido ao clareamento com a solução xilol por 1 hora para garantir a
penetração em todo o material, seguindo com a sequência de xilol mais álcool
absoluto (1:1) e posteriormente xilol mais parafina (1:1).
Os tecidos vegetais, após o processo de desidratação e clareamento
estavam prontos para a emblocagem. Com o auxílio de suportes metálicos foi
possível à confecção dos blocos de parafina. Posteriormente, os suportes já
contendo a parafina histológica na forma líquida foram adicionados
rapidamente às amostras, sempre na mesma posição, os tecidos, de raízes e
folhas em triplicatas com a ajuda de uma pinça. Os blocos de parafina medindo
1,5 cm2 foram acondicionados em temperatura ambiente.
Para a preparação das lâminas foi necessário congelar a -20 ºC os
blocos contendo as amostras por 12 horas antes, a fim de reduzir os danos ao
tecido na hora do corte. Com o auxílio do micrótomo (modelo 820 da América
Optical Company -New York, EUA) foram feitos os cortes histológicos em 10
µm de espessura. Os cortes de parafina obtidos foram então mergulhados num
banho-maria a 35-45°C (modelo BH10R marca patologista) contendo gelatina
incolor comercial. Com auxílio das lâminas foi realizada a pesca dos cortes
histológicos. Em seguida, estas foram colocadas verticalmente no suporte à
temperatura ambiente. Após o período de 24 horas de secagem foi iniciada a
etapa de desparafinização mergulhando assim as lâminas por 5 minutos, três
vezes no xilol e uma vez em séries diferentes de álcool 100%, 80%, 70% e por
fim, na água pura para reidratação (Adaptado de KRAUS & ARDUIN, 1997).
Em seguida, iniciou-se a sequência de coloração com os corantes temporários
e permanentes conforme a Tabela 1.

31
Tabela 1: Relação de corantes. Estes corantes foram utilizados nas amostras analisadas por microscopia óptica com as estruturas evidenciadas por estes.
CORANTE INDICADOR
Diaminobenzidina (DAB) Peróxido de hidrogênio
Cloreto de zinco iodado (CZI)
Azul-acinzentado (celulose); Amarela (lignina) Negro-azulado (amido)
Safranina (S) Vermelho forte (lignina)
Azul de toluidina (AT) Estruturas ácidas (ácidos nucleicos) e não acidas (lignina)
Eosina de metileno (EM) Estruturas abundantes (citoplasma)
As lâminas histológicas foram analisadas com o auxílio do microscópio
óptico (Olympus X41) em campo claro, com aumento de 10x para as raízes e
40x para as folhas de cana-de-açúcar. As imagens foram capturadas pela
câmera Nikon DXM 1200 acoplada ao microscópio e o software Nikon ACTL,
equipamentos disponibilizado pelo Laboratório de Neuroanatomia, no centro de
Biociências, UFRN.
3.3.3. Microscopia Eletrônica de Varredura
Os tecidos de cana-de-açúcar (folha e raiz) foram analisados em
Microscopia Eletrônica de Varredura (MEV) no Núcleo de Estudos em Petróleo
e Gás Natural (NEPGN-UFRN). Seguindo o protocolo de fixação já descrito
anteriormente para a preparação da microscopia óptica, as amostras de folha e
raiz mantidas no álcool a 70% foram secadas a temperatura ambiente.
Posteriormente, estas amostras foram transferidas para um suporte de porta
amostras contendo stubs (pequenas estruturas metálicas), onde as amostras
foram montadas com auxílio de pinças, pequenos cortes de fita adesiva de
carbono condutora de elétrons foram colados nos stubs e acima da fita, as
amostras foram fixadas com cola prata (Agar). Em seguida, as amostras foram
conduzidas para a câmera de vácuo de metalização, onde elas foram
revestidas com ouro em pó.
3.4. Ensaios enzimáticos
Os tecidos de raiz e folha foram utilizadas para análises das atividades
das enzimas: ascorbato peroxidase (APX), superóxido desmutase (SOD) e
catalase (CAT). Estes ensaios foram realizados em triplicatas biológicas.

32
3.4.1. Extração de proteínas solúveis totais
Para a extração de proteínas enzimáticas foram utilizados o tampão
fosfato de potássio 100 mM pH 7,0 contendo EDTA 1 mM e ácido ascórbico 0,5
mM. Utilizou-se 250 mg de amostra de raízes e folhas armazenadas
previamente em freezer -80 °C. Estas amostras foram trituradas com o
nitrogênio líquido e transferidas para tubos de centrífuga de 2 mL, onde foi
adicionado 1,2 mL do tampão fosfato de potássio e mantidas a 0 ºC. Em
seguida, estas amostras foram centrifugadas a 13.000 xg, 4°C durante 20
minutos (MIKRO 200R – Hettich Zentrifugen). Em seguida, o sobrenadante foi
coletado e transferido para um novo tubo. As proteínas solúveis totais foram
dosadas pelo método de Bradford (1976), contra uma curva padrão de BSA.
3.4.2. Determinação da atividade de ascorbato peroxidase (APX)
O protocolo utilizado foi baseado em Nakano & Asada (1981). Para a
análise da atividade enzimática de ascorbato peroxidase APX utilizou-se 100
µL do extrato de cada amostra, a este foi adicionado 2,7 mL do tampão de
ensaio de APX (tampão fosfato de potássio 50 mM pH 6,0 contendo ácido L-
ascórbico 0,5 mM) e 200 µL peróxido de hidrogênio (30 mM). As leituras foram
realizadas em espectrofotômetro (GE) utilizando a absorbância de 290 nm.
Para o cálculo da atividade enzimática foi utilizado o coeficiente de extinção
molar do ascorbato de 2,8 mM-1cm-1 e os resultados foram expressos em nmol
de ascorbato/g de proteína por minuto.
3.4.3. Determinação da atividade superóxido dismutase (SOD)
O protocolo utilizado para a determinação da atividade de superóxido
dismutase foi modificado a partir de Peixoto et al. (1999). Este ensaio foi
realizado em microplacas de 96 poços tipo Elisa. Inicialmente para o ensaio
enzimático foi utilizado o tampão fosfato de potássio contendo: fosfato de
potássio 50 mM, pH 7,8; EDTA 0,1 mM; L-Metionina 13 mM. Para a leitura nas
microplacas foi seguida uma ordem para os reagentes: primeiro foi adicionado
o tampão fosfato de potássio 50 mM, seguido do NBT 750 µM (Nitro-blue

33
tetrazolium - Sigma), a amostra (extrato bruto) e riboflavina 1 mM. Também
foram realizados controles brancos (tampão). Esta microplaca depois de
completamente montada com todas as amostras e reagentes foi exposta a luz
branca por 2 minutos e em seguida foi realizada a sua leitura em 560 nm com o
leitor de Elisa (espectrofotômetro Epoch da Biotek). Uma unidade de enzima
correspondeu à quantidade de enzima capaz de inibir em 50% a fotorredução
do NBT e os resultados foram expressos em Unidade de Atividade Enzimática
por mg de proteína do extrato.
3.4.4. Determinação da atividade de catalase (CAT)
Esta análise foi realizada de acordo com Havir & McHale, (1987). Para
este ensaio de CAT foi utilizado o tampão fosfato (tampão de potássio 50 mM,
pH 7,0 a 30 °C) contendo peróxido de hidrogênio a 20 mM. As leituras foram
realizadas em espectrofotômetro no comprimento de onda de 240 nm. Antes da
leitura das amostras, foi zerado o equipamento com o tampão de atividade CAT
sem a adição do peróxido de hidrogênio. Em seguida, foi realizado a leitura do
branco com o tampão de atividade de CAT contendo peróxido de hidrogênio 20
mM. Posteriormente, foi realizada a leitura do branco com o tampão de
atividade de CAT contendo peróxido de hidrogênio e tampão de extração das
enzimas. Depois de realizado a leitura dos controles, as amostras foram lidas
utilizando 50 µL de cada amostra (extrato bruto) em tampão do ensaio
contendo peróxido de hidrogênio a 20 mM. O decaimento da absorbância foi
acompanhado durante 5 min, em intervalos de 30 s no espectrofotômetro a 240
nm. Os resultados foram expressos em µmol de H2O2 por grama de proteína
por minuto.
3.4.5. Estimação da peroxidação de lipídios pelo (TBARS)
A análise de substâncias reagentes ao TBA foi modificada a partir de
Vyncke (1970). Este teste foi realizado em tubos de ensaio com tampas e
adicionado o reagente TCA 20% - TBA 0,5% e 500 µL da amostra. Depois
foram mantidos em banho-maria a 100°C por 1 hora. Ao término dos 60
segundos, foi necessário parar a reação. Para isto, os tubos foram colocados
em banho de gelo e, em seguida, centrifugados a 3.500 xg a temperatura
ambiente. As amostras foram lidas no espectrofotômetro (GE) utilizando a

34
absorbância de 532 nm a 660 nm utilizando o branco de referência que conteve
2000 µL de TCA 20% - TBA 0,5% e 500 µL de TCA 1%. Esta análise foi
determinada em triplicatas de duas amostras de folhas e raízes de cana-de-
açúcar. Os resultados foram expressos em mg malonaldeído (MDA-TBA) por g
de tecido.
3.4.6. Análises estatísticas
Para as análises estatísticas foram realizadas aplicando a análise de
variância (ANOVA) e as diferenças localizadas pelo Teste de Tukey, com
p<0,05.

35
4. RESULTADOS 4.1. Tratamento com peróxido de hidrogênio
Inicialmente, os colmos foram colocados para germinar em areia
previamente lavada com água destilada. Entretanto, a frequência de
germinação obtida foi muito baixa, menor do que 10% e foi observado
problemas de contaminação por fungos e insetos. Posteriormente, foi
estabelecido o protocolo de cultivo para as plantas de cana-de-açúcar, que
atingiu a maior taxa de germinação com sucesso, no qual foi utilizada a
selagem dos colmos com parafina e o plantio utilizando o substrato de
vermiculita expandida (Figura 7).
Figura 7: Parafinização dos colmos. Em (A) é apresentado o colmo contendo dois entrenós com duas gemas (setas) e o surgimento de raízes adventícias. Em (B) parafinização (seta) e (C), é apresentado em detalhe a parafinização das extremidades dos cortes, (D) é apresentado um colmo após cinco dias do plantio, em germinação e (E) mostra as plantas com quinze dias após o plantio. Local: Sala de crescimento de plantas do Laboratório de Biologia Molecular e Genômica/UFRN (Fonte: Acervo pessoal).
Desta forma, plantas com 60 dias de desenvolvimento foram separadas
e utilizadas para o tratamento hidropônico. Nas primeiras 24 horas, as plantas
foram mantidas em hidroponia sem o acréscimo do peróxido de hidrogênio.
Este período foi determinado para a aclimatação dos vegetais a esta nova
condição, durante este tempo, não foram observadas alterações estruturais.
Passadas as 24 horas, deu-se início ao tratamento com peróxido de
hidrogênio nas seguintes concentrações (10 mM, 20 mM e 30 mM).

36
Para a titulação do peróxido de hidrogênio foi utilizada a técnica de
permanganometria, que é um método titulométrico de oxido-redução, que usa o
reagente permanganato de potássio, um forte oxidante, como titulante em meio
ácido, para facilitar a oxidação da substância em análise. Com isso, usou-se o
peróxido de hidrogênio para determinar a concentração da solução através da
volumetria de óxido-redução com permanganato de potássio. O teor de
peróxido de hidrogênio foi medido a cada hora do tratamento a fim de verificar
a sua absorção pelas plantas ou a ocorrência de uma possível degradação do
peróxido de hidrogênio. Assim, esse ensaio foi realizado para as concentrações
estabelecidas no tratamento das plantas (0 mM - controle, 10mM, 20 mM e 30
mM) de peróxido de hidrogênio, nas mesmas condições de hidroponia à qual
os vegetais foram submetidos, inclusive com a presença do aerador.
Na figura 8, a linha constante refere-se à titulação do conteúdo de
peróxido nas bandejas que foram mantidas nas condições de tratamento, mas
sem a presença das plantas. As outras linhas correspondem ao tratamento
realizado com as plantas de cana-de-açúcar. Comparando-se os dados obtidos
na titulação com os obtidos nas bandejas contendo as plantas submetidas ao
tratamento com H2O2, foi possível observar que a concentração de H2O2 foi
reduzida até zero, na oitava hora de tratamento. Estes resultados comprovam
que as plantas absorveram o H2O2 contido na solução de hidroponia.

37
Figura 8. Gráfico de determinação de H2O2 por permanganometria. Titulação de H2O2 em recipientes contendo apenas peróxido de hidrogênio (sem plantas) e recipientes contendo plantas submetidas às diferentes concentrações utilizadas nos tratamentos.
Durante o tratamento com peróxido de hidrogênio observaram-se
alterações foliares, a exemplo, o fechamento gradativo das folhas de maneira
proporcional à concentração de peróxido aplicada, indicando a perda do turgor:
folhas tratadas com 10 mM de H2O2 apresentaram fechamento das
extremidades no ápice foliar (Figura 9); já folhas tratadas com 20 mM de H2O2
apresentaram fechamento de ambas as extremidades, enquanto que folhas
tratadas com 30 mM de H2O2 tiveram um fechamento total, além de abscisão
foliar (Figura 9). As folhas pertencentes ao grupo controle (sem peróxido) não
apresentaram as alterações mencionadas acima (Figura 9).

38
Figura 9: Alterações morfológicas permanentes das folhas de cana-de-açúcar tratadas com peróxido de hidrogênio em diferentes concentrações (0, 10, 20 e 30 mM). À esquerda folha durante aclimatação hidropônica da planta durante 24 horas (A). À direita estão representadas as folhas após o tratamento com H2O2: (B) controle, (C) 10 mM de peróxido; (D) tratado com 20 mM de peróxido e (E) tratado com 30 mM de peróxido.
4.1.2. Determinação do conteúdo de peróxido de hidrogênio nos tecidos
A medição do conteúdo de peróxido de hidrogênio nos tecidos foi
realizada de modo a verificar se houve a absorção a partir das raízes até a
parte aérea. Na figura 10, o gráfico do meristema apical mostra que a
exposição à concentração de 10 mM de H2O2 levou a um aumento do conteúdo
de peróxido de hidrogênio uma vez e meia quando comparado ao controle. Já
para a concentração de 20 mM, o conteúdo de peróxido de hidrogênio mostra-
se semelhante quando comparado ao controle. Na concentração de 30 mM, o
conteúdo de peróxido de hidrogênio verificado está um pouco maior quando
comparado ao controle. Nos tecidos foliares, pode-se observar um pequeno
aumento do conteúdo de peróxido de hidrogênio para os tratamentos de 10
mM, 20 mM e 30 mM quando comparado ao controle. Observa-se, contudo,
que esse aumento não foi crescente conforme a concentração de peróxido de
hidrogênio utilizada nos tratamentos. Nas raízes, foi verificada uma
concentração maior de peróxido de hidrogênio para os tratamentos 10 mM e 30
mM quando comparado ao controle.

39
Figura 10: Conteúdo de peróxido de hidrogênio em meristema, folhas e raízes de plantas de cana-de-açúcar. Submetidas ao tratamento com concentrações crescentes de peróxido de hidrogênio (10, 20 e 30 mM) e controle (sem peróxido).
a,b,c,d Letras distintas significam
diferença significativa entre os diferentes tratamentos.Teste ANOVA-Tukey (p≤0,05).

40
4.1.3. Espécies reativas ao ácido 2 – Tiobarbitúrico (TBARS)
Como nosso objetivo foi compreender o mecanismo que envolve o
estresse oxidativo e como este afeta as plantas, no item acima foram descritos
a análise do conteúdo de H2O2 nos tecidos de folhas, raízes e meristema
associados à manutenção do balanço de EROs, e neste item foi avaliado a
peroxidação de lipídios que pode ser outro alvo das EROs.
A figura 11 apresenta os resultados obtidos com esta análise para três
tecidos diferentes: meristema, folha e raízes. Nos três tecidos foram
observados resultados semelhantes. Para os tratamentos de 10 mM e 30 mM
observamos que os valores obtidos para a peroxidação de lipídeos nas
membranas foi maior quando comparado com o controle, este aumento foi até
4 vezes quando comparado o controle com o tratamento na concentração de
10 mM. Já para a concentração de 30 mM foi de 0.5 a até 5 vezes aumentada.
No tratamento com 20 mM de peróxido de hidrogênio observamos que não
houve uma variação significativa em relação ao controle. Em todos os tecidos
observou-se maior conteúdo de H2O2 e um menor conteúdo de TBARS.

41
Figura 11: Gráficos de peroxidação lipídica (TBRAS). Nos tecidos de meristema; folha e raiz de plantas de cana-de-açúcar submetidas a estresse oxidativo, grupo controle; 10 mM; 20 mM e 30 mM de peróxido de hidrogênio.
a,b,c,d Letras distintas significam diferença significativa entre
os diferentes tratamentos.Teste ANOVA-Tukey (p≤0,05).

42
4.2. Análises histológicas de folha e raiz
4.2.1. Análise morfológica por microscopia óptica
A observação por microscopia óptica da estrutura celular dos tecidos de
folhas e raízes de plantas de cana-de-açúcar foi importante para quais seriam
as alterações morfológicas observadas durante o tratamento com peróxido de
hidrogênio. Algumas variações estruturais foram observadas nas folhas
referente ao xiliema quanto ao número, e esta variação foi considerada como
natural entre os vegetais.
A figura 12 é uma representação esquemática da estrutura anatômica de
uma folha em condições normais, isto é, sem o tratamento com o peróxido de
hidrogênio. Nessa figura observa-se, considerando a orientação da parte
interna para a externa, as estruturas do xilema; floema; células da anatomia de
kranz; células buliformes e, por fim, na parte superior (adaxial), observa-se o
colênquima; epiderme adaxial e tricoma; na parte inferior (abaxial) também se
observa o colênquima abaxial; epiderme abaxial e células guardas.
Da mesma maneira, na figura 13 é apresentado o esquema da estrutura
anatômica de uma raiz da planta de cana-de-açúcar em condições normais,
onde são observadas as estruturas: medula delimitada pelo cilindro vascular,
metaxilema e córtex na fase interna, na fase externa observa-se endoderme e
aerênquima.

43
Figura 12: Secção transversal da folha de cana-de-açúcar nas condições normais.
Desenho à mão livre sob técnica grafite. Observa-se as estruturas: Ead - epiderme adaxial;
Eab - epiderme abaxial; Cad-colênquima adaxial; Cab-colênquima abaxial; X xilema; Fl-floema;
CB-células buliformes e CK – Coroa de Kranz ou anatomia de Kranz. Imagem do acervo
pessoal.
Figura 13: Secção transversal da raiz de cana-de-açúcar nas condições normais. Desenho à mão livre sob técnica grafite. Observa-se as estruturas: Medula, Córtex; Metaxilema; Endoderme, Aerênquima e Epiderme. Imagem do acervo pessoal.

44
Na figura 14 A é apresentado à imagem de uma secção transversal com
o corante safranina de uma folha controle (sem adição de peróxido de
hidrogênio). Nesta figura é observada uma coloração avermelhada uniforme, o
mesmo não foi observado para os diferentes tratamentos com o peróxido de
hidrogenio. Quando esta imagem é comparada com tratamento com peróxido
na concentração de 10 mM foi observado um coloração uniforme com exceção
nas regiões correspondentes ao colênquima adaxial e abaxial (Figura 14 B).
Esta forte coloração é indicativo de acúmulo de lignina nessa região. No
tratamento com peróxido na concentração de 20 mM (Figura 14 C), observa-se
nas células que envolvem o floema um tom vermelho forte. Novamente esta
coloração sugere o acúmulo de lignina nessa região. Na figura 14 D, onde o
material foi tratado com a concentração de 30 mM foi observado que as células
do colênquima adaxial e abaxial, coroa de Kranz, xilema e floema
apresentaram essa forte coloração vermelha indicando a provável presença de
lignina nessa região.
Figura 14: Análise das folhas de cana-de-açúcar tratadas com o corante Safranina. Secção transversal em plantas submetidas ao estresse oxidativo. (A) controle; (B tratamento 10 mM); (C) tratamento 20 mM e (D) tratamento 30 mM. Aumento 40x.
A figura 15 mostra as raízes em secção transversal submetidas ao
estresse oxidativo pelo tratamento com peróxido de hidrogênio coradas com
safranina. Na figura 15 A é apresentado a imagem de uma raiz de uma planta

45
controle, onde pode ser observada uma coloração uniforme. No tratamento
com peróxido de hidrogênio na concentração de 10 mM é observado o
aumento das células (em volume) do aerênquima com rompimento celular e o
córtex lignificado com a coloração em vermelho escuro (Figura 15 B). Na raiz
tratada com 20 mM pode-se observar que a região do metaxilema apresentou
uma desorganização devido ao aumento do volume seguido de rompimento
nas paredes da célula e, além disso, a epiderme apresentou-se fortemente
corada em vermelho indicando a lignificação dessas paredes (Figura 15 C). O
tratamento com 30 mM de peróxido de hidrogênio permitiu observar a presença
de uma região de aerênquima desorganizada (Figura 15 D).
Figura 15: Análise das raízes de cana-de-açúcar pelo corante safranina. Secção transversal em plantas submetidas ao estresse oxidativo. (A) controle; (B) tratamento 10mM; (C) tratamento 20mM; (D) tratamento 30mM,. Aumento 10x.
Outro corante que foi utilizado e nesse estudo foi o corante Eosina de
Metileno (EM), que é um corante ácido que cora estruturas básicas da célula.
Estas estruturas são abundantes no citoplasma e são chamadas de acidófilas
(GARTNER & HIATT, 1999). Então, este corante foi utilizado com o propósito
de corar estruturas no citoplasma. A eosina se liga nas paredes dos
cloroplastos indicando na célula a sua real localização. Na figura 16 A, foi
observado a presença de cloroplastos nas células que compõem a coroa de
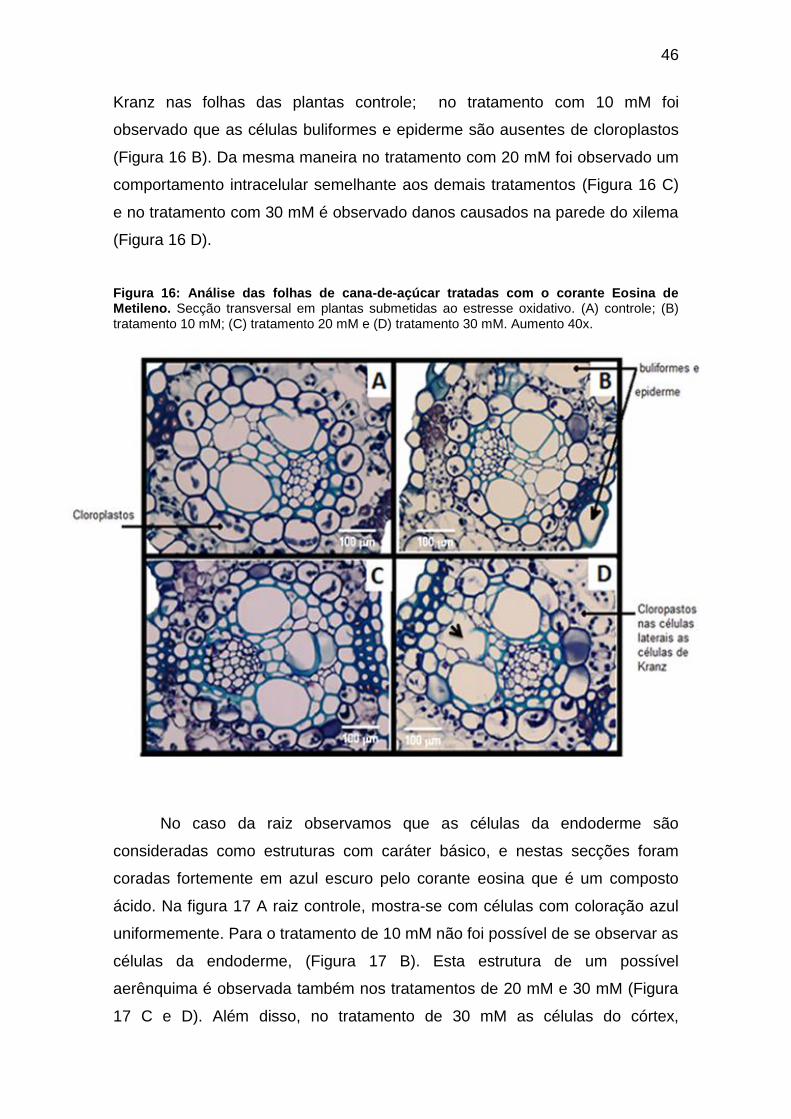
46
Kranz nas folhas das plantas controle; no tratamento com 10 mM foi
observado que as células buliformes e epiderme são ausentes de cloroplastos
(Figura 16 B). Da mesma maneira no tratamento com 20 mM foi observado um
comportamento intracelular semelhante aos demais tratamentos (Figura 16 C)
e no tratamento com 30 mM é observado danos causados na parede do xilema
(Figura 16 D).
Figura 16: Análise das folhas de cana-de-açúcar tratadas com o corante Eosina de Metileno. Secção transversal em plantas submetidas ao estresse oxidativo. (A) controle; (B) tratamento 10 mM; (C) tratamento 20 mM e (D) tratamento 30 mM. Aumento 40x.
No caso da raiz observamos que as células da endoderme são
consideradas como estruturas com caráter básico, e nestas secções foram
coradas fortemente em azul escuro pelo corante eosina que é um composto
ácido. Na figura 17 A raiz controle, mostra-se com células com coloração azul
uniformemente. Para o tratamento de 10 mM não foi possível de se observar as
células da endoderme, (Figura 17 B). Esta estrutura de um possível
aerênquima é observada também nos tratamentos de 20 mM e 30 mM (Figura
17 C e D). Além disso, no tratamento de 30 mM as células do córtex,

47
endoderme e epiderme mostraram-se fortemente lignificadas, com o tamanho
modificado e desorganizadas quando comparado ao controle.
Figura 17: Análise das raízes de cana-de-açúcar pelo corante Eosina de Metileno. Secção transversal em plantas submetidas ao estresse oxidativo. (A) controle; (B) tratamento 10 mM; (C) tratamento 20 mM e (D) tratamento 30 mM. Aumento 10x.
O corante Diaminobenzidina (DAB) possui a sua composição incolor e
quando este se liga a molécula de H2O2 o corante oxida mudando assim a sua
coloração para marrom ou caramelo. Na figura 18 A que representa a folha
controle (tecido sem a presença de peróxido) foi observada poucos
cloroplastos visíveis, a seta indica a visualização de um cloroplasto. Na folha
tratada com 10 mM de peróxido de hidrogênio fica evidente a presença de
muitos cloroplastos (Figura 18 B), sendo estes pouco corados por DAB, a seta
indica a localização do cloroplasto com partes de sua membrana corada em
marrom. Na figura 18 C, folha tratada com 20 mM de peróxido, a seta do centro
da imagem revela a membrana celular corada em marrom e a seta da direita
mostra os poucos cloroplastos fracamente corados em marrom próximo à
membrana celular. Na figura 18 D com a folha tratada com 30 mM de peróxido
de hidrogênio observa-se o aumento considerável do tamanho das células
buliformes e anatomia de Kranz, e a seta da esquerda mostra os cloroplastos
maiores e com maior intensidade da cor marrom para estas organelas. A seta

48
central evidencia a maior intensidade em marrom também para a membrana
celular.
Figura 18: Análise das folhas de cana-de-açúcar com a coloração Diaminobenzidina (DAB). Secção transversal em plantas submetidas ao estresse oxidativo. (A) controle; (B) tratamento 10 mM; (C) tratamento 20 mM; (D) tratamento 30 mM. Aumento 40x. (Acervo pessoal).
Nas raízes de plantas não submetidas ao estresse oxidativo,
observamos uma leve coloração com DAB na região da endoderme (Figura 19
A). Entretanto, nos tratamentos com 10 mM (Figura 19 B1) e 30 mM (Figura 19
D3) observamos uma forte coloração na região da endoderme e córtex. Ainda
na figura 19 (B4 C5 e D6) observamos o resultado obtido com outro corante de
lâminas temporárias que foi utilizado - o Cloreto de Zinco Iodado (CZI). Este
corante permite visualizar em colaração azul-acinzentado ou rosa o
componente de celulose, e com a coloração amarela para a lignina e com a cor
negra-azulada para o amido (JENSEN, 1962). Para as raízes no tratamento 10
mM de peróxido de hidrogênio (Figura 19 B4) a seta indica a presença de
lignina, e pela cor amarela na células da epiderme e nas células do córtex
evidencia a total degradação das células da endoderme. No tratamento 20 mM

49
de peróxido de hidrogênio a seta abaixo observa-se a lignificação das paredes
das células da endoderme pela presença da cor amarela intensa (Figura 19
C5), enquanto que a seta acima mostra a presença de celulose na região
medular pela cor rosa e no tratamento com 30 mM de peróxido de hidrogênio a
seta indica em amarelo intenso para a presença de lignina nas células da
endoderme (Figura 19 D6).
Figura 19: Análise das Raízes de cana-de-açúcar pela a coloração Diaminobenzidina (DAB). Secção transversal em plantas submetidas ao estresse oxidativo. (A1) controle. (B2) tratamento 10 mM de peróxido; (C3) tratamento 20 mM de peróxido; (D) tratamento 30 mM de peróxido. Corante cloreto de zinco iodado. (B4) tratamento 10 mM; (C5) tratamento 20 mM e (D6) tratamento 30 mM. Aumento 10x.
Na secção transversal da folha observamos uma região negro-azulada
na região do colênquima adaxial e abaxial correspondendo provavelmente a
reservas de amido (setas) na planta controle (Figura 20 A). Para o tratamento
com 10 mM e 20 mM de peróxido de hidrogênio observamos apenas a
presença dos cloroplastos no interior das células (setas) (Figura 20 B e C).
Entretanto, quando comparamos com o tratamento com 30 mM de peróxido de
hidrogênio observou-se uma coloração amarela intensa nas membranas dos

50
cloroplastos e como indicado pela seta acima e a seta mostra uma lignificação
na parede da membrana celular do xilema também corando as demais células
(Figura 20 D).
Figura 20: Análise das folhas de cana-de-açúcar pelo corante cloreto de zinco iodado. Secção transversal em plantas submetidas ao estresse oxidativo. . (A) controle; (B) tratamento 10 mM de peróxido; (C) tratamento 20 mM de peróxido; (D) tratamento 30 mM de peróxido. Aumento 40x.

51
4.2.2. Análise dos tecidos de folha e raízes por microscopia eletrônica de varredura.
A microscopia eletrônica de varredura (MEV) permitiu uma visualização
analítica do tecido após os tratamentos do estresse oxidativo na estrutura foliar
e na raíz.
A figura 21 mostra os resultados obtidos com o tecido de folhas que é
apresentado a imagem da estrutura foliar controle com o xilema, floema e
epiderme (Figura 21 A). Na amostra tratada com 10 mM observamos que
houve modificações estruturais como um aumento do xilema e alteração na
região do colênquima (Figura 21 B). Assim, foi observada a formação de mais
dois xilemas, assim como modificações na epiderme adaxial e abaxial que se
apresentam a sua estrutura celular aumentada, na coroa de Kranz, que
também, sofreu uma desorganização devido o aumento das células
circunvizinhas ou da própria célula da coroa de Kranz. Para a folha tratada com
20 mM observou-se um aumento celular no floema e colênquima adaxial e
abaxial. Para a região da epiderme, o que chama a atenção é a abertura
aumentada para todas estas células, a coroa de Kranz está com suas células
aumentadas e com o xilema traqueal bem aberto, dando a impressão de maior
rigidez (Figura 21 C). Novamente, no tratamento com 30 mM de peróxido de
hidrogênio observa-se uma desorganização foliar em relação ao material
controle (Figura 21 D), e também foi observado um aumento exacerbado do
tamanho das células. Na figura 22 A e A1 visualiza-se em um ângulo
amplificado o que ocorreu na parede entre os anéis do xilema traqueal após o
tratamento com 30 mM de peróxido de hidrogênio mostrando sua degradação.

52
Figura 21: Análise em MEV das folhas. Foto micrografia eletrônica de varredura com aumento de 500x de seção transversal de folhas de plantas de cana-de-açúcar submetidas ao estresse oxidativo na concentração de (A) 0 mM; (B) 10 mM; (C) 20 mM e (D) 30 mM de peróxido fixadas em suporte metálico específico nas posições vertical.
Figura 22: Análise em MEV das folhas. Foto micrografia eletrônica de varredura com aumento de 500x de seção transversal de folhas de plantas de cana-de-açúcar submetidas ao estresse oxidativo na concentração de (A) 30 mM de tratamento com peróxido fixadas em suporte específico nas posições vertical (B) Folha 30 mM aumentada 2000x.
Quando foi analisado o tecido da raiz após este tratamento foi observado
também algumas alterações nas estruturas de organização radicular como foi
evidenciado anteriormente pela microscopia óptica. Na figura 23 A e A1
observamos as raízes controle em dois diferentes aumentos, que nos auxiliou a

53
definir um padrão para este tecido em questão. A análise com o MEV permitiu
observar que para o tratamento com 10 mM de peróxido de hidrogênio houve
uma desorganização nas células da endoderme e um aumento nas células do
aerênquima (Figura 23 B e B1). No tratamento com 20 mM de peróxido de
hidrogênio foi observado novamente essa grande desorganização nas células
da endoderme e agora um aumento na circunferência do metaxilema, medula e
córtex (Figura 23 C e C1). Para as raízes tratadas com 30 mM de peróxido de
hidrogênio observamos que as células do córtex estão maiores quando
comparada com o controle, além da presença do aerênquima (Figura 23 D e
D1).
Figura 23: Análise em MEV das raízes. Foto micrografia eletrônica de varredura com aumento de 300x de seção transversal de raízes de plantas de cana-de-açúcar submetidas ao estresse oxidativo fixadas em suporte específico nas posições vertical. Raiz controle (A) e (A1) aumento 50x; tratamento 10 mM de peróxido (B) e (B1) aumento 60x; tratamento 20 mM de peróxido, (C) e (C1) aumento 60x e tratamento 30 mM de peróxido, (D) e (D1) aumento 60x.

54
4.3. Análises enzimáticas
Como mencionado acima, as plantas apresentam um conjunto de
enzimas que contribuem para a homeostase redox. Considerando isso, neste
trabalho foram analisadas três destas enzimas: ascorbato peroxidase, catalase
e superóxido dismutase com o intuito a compreender como o tratamento com o
peróxido de hidrogênio pode ter alterado esse balanço entre moléculas
antioxidantes e EROs.
4.3.1. Ascorbato peroxidase (APX)
Ao longo do monitoramento da taxa de oxidação do ascorbato a 290 ηm,
pode-se analisar a atividade da ascorbato peroxidase em folhas, revelando que
não houve diferença significativa entre as amostras submetidas ao estresse
oxidativo entre os tratamentos controle e tratados (Figura 24).
Para o sistema meristemático, a atividade enzima ascorbato peroxidase
aumentou na concentração 10 mM. Para os tratamentos com 20 mM e 30 mM
houve um pequeno aumento quando comparado com o controle, mas este não
foi considerado estatisticamente significativo. Já para o sistema radicular, foi
encontrado um aumento na atividade da APX apenas no tratamento de 20 mM
quando comparado com o controle. Para o tecido foliar foi verificado que os
tratamentos quando comparado ao controle não foi considerado
estatisticamente significativo.

55
Figura 24: Gráficos da atividade enzimática de ascorbato peroxidase (APX). Nos tecidos de meristema; folha e raiz de plantas de cana-de-açúcar submetidas a estresse oxidativo, grupo controle; 10 mM; 20 mM e 30 mM de peróxido de hidrogênio. As barras representam o desvio-padrão da média de três repetições.
a,b,c,d Letras distintas significam diferença significativa entre os diferentes tratamentos.Teste ANOVA-Tukey (p≤0,05).

56
4.3.2. Catalase (CAT)
A catalase é uma proteína chave para a quebra do peróxido de
hidrogênio nas células. Para o tecido meristemático e folha tratados com
peróxido de hidrogênio foi verificada a redução da atividade desta enzima
quando comparado ao controle. Em raiz, não foi detectada a atividade de
catalase tanto no controle quanto nos tratados (Figura 25).
Figura 25: Gráficos da atividade enzimática de catalase (CAT). Nos tecidos de meristema; folha e raiz de plantas de cana-de-açúcar submetidas a estresse oxidativo, grupo controle; 10 mM; 20 mM e 30 mM de peróxido de hidrogênio.
a,b,c,d Letras distintas significam diferença
significativa entre os diferentes tratamentos.Teste ANOVA-Tukey (p≤0,05).

57
4.3.3. Superóxido dismutase (SOD)
A determinação da atividade enzimática da enzima superóxido
dismutase foi por meio da sua capacidade em inibir em 50% a fotorredução do
azul de nitrotetrazólio (NBT). Na figura 26 observa-se no gráfico para o tecido
do meristema que o aumento na atividade desta enzima não foi significativo
quando comparado o tratamento com 10 mM com o tratamento controle.
Entretanto, quando foram analisados os tratamentos com 20 mM e 30 mM foi
verificado que a enzima SOD aumentou sua atividade em relação ao controle.
No gráfico que expressa os dados para os tecidos foliares verifica-se
que os valores são praticamente os mesmos para o controle e o tratamento
com 30 mM. Nos tratamentos com 10 mM e 20 mM foi observado uma redução
na atividade desta enzima quando comparada com o tratamento controle.
Ainda na figura 26 o gráfico que representa os tecidos radiculares a
atividade da enzima superoxido dismutase parece ser igual para todos os
tratamentos quando comparado ao controle.
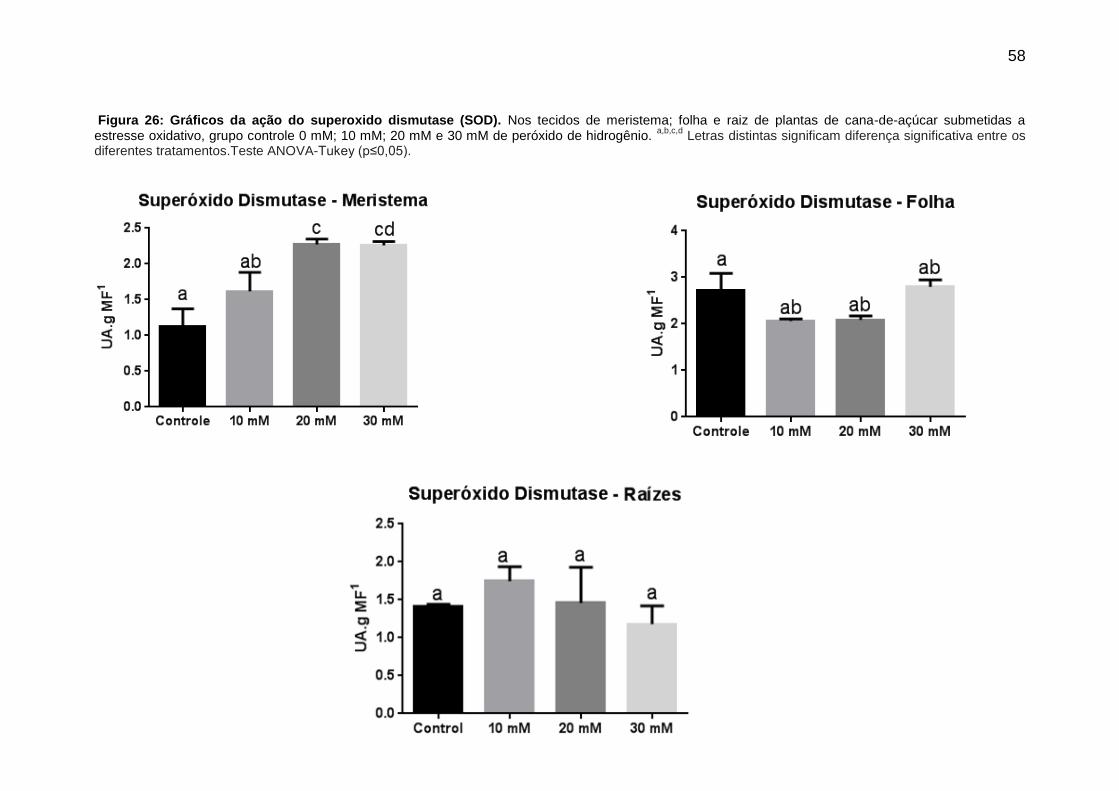
58
Figura 26: Gráficos da ação do superoxido dismutase (SOD). Nos tecidos de meristema; folha e raiz de plantas de cana-de-açúcar submetidas a estresse oxidativo, grupo controle 0 mM; 10 mM; 20 mM e 30 mM de peróxido de hidrogênio.
a,b,c,d Letras distintas significam diferença significativa entre os
diferentes tratamentos.Teste ANOVA-Tukey (p≤0,05).

59
5. DISCUSSÃO
Durante muito tempo, houve uma forte associação da presença de
espécies reativas de oxigênio (EROs) com disfunções fisiológicas que levavam
à morte celular (SCANDALIOS, 2005). Contudo, essa visão se tornou
ultrapassada, haja vista que essas moléculas são potentes sinalizadoras e
podem estar envolvidas tanto no controle de crescimento e desenvolvimento
das plantas, bem como em respostas de aclimatação e na resposta aos
estresses bióticos e/ou abióticos (LOIACONO et al., 2012; MILLER, 2010 ).
Apesar disso, os mecanismos que levam ao controle da homeostase redox
sistemas vegetais estão longe de ser elucidados (FOYER & SHIGEOKA, 2010).
A maioria das pesquisas envolvendo a avaliação do estado redox em
plantas sob condição de estresse tem utilizado muito o modelo genético
Arabidopsis thaliana que é uma planta herbácea, dicotiledônea, com
fotosíntese C3.
Desta forma, os resultados gerados neste trabalho contribuem para o
entendimento de como as plantas podem responder ao estresse oxidativo
utilizando diretamente o peróxido de hidrogênio nas plantas de cana-de-açúcar,
ao qual, corresponde a uma planta de mecanismo fotosintético C4. Esta planta
pertence à família Poaceae, é uma monocotiledônea e de grande valor sócio-
econômico-ambiental, em especial para a região Nordeste do Brasil.
Esse trabalho, portanto, apresenta resultados acerca da resposta de
plantas de cana-de-açúcar ao tratamento com peróxido de hidrogênio,
utilizando diversas abordagens: anatômicas, enzimáticas e proteômica. Além
disso, nesse trabalho foi verificado que as plantas absorveram o peróxido de
hidrogênio utilizado. Este foi verificado com os dados de titulação e a
determinação de conteúdo de peróxido de hidrogênio nos tecidos analisados.
Os resultados de histoquímica com microscopia óptica permitiram
observar o acúmulo de lignina nas paredes celulares das folhas e raízes
tratadas com H2O2. A lignina está presente na célula sempre em maior
abundância na resposta de defesa, aumentando a resistência na presença de
agentes estressores entre outros fatores (LACAYO et al., 2013; ZHONG et al.,
2000). Observou-se, ainda nas análises histológicas, um acúmulo de peróxido
de hidrogênio em folhas e raízes. Notou-se que a concentração de H2O2 foi
maior para o tratamento com 30 mM, este possivelmente aumentou a síntese
de lignina em resposta ao estresse. Outros estudos o mesmo pode ser

60
observado em plantas de trigo sob estresse seca e salinidade (LI et al., 2012) e
em soja (SANTOS et al., 2008). Também foi descrito em estudos com
tomateiro (Solanum lycopersicum) que a enzima peroxidase provavelmente
estaria envolvida no processo de síntese da lignina nas raízes (QUIRONGA et
al., 2000). DENESS et al. (2011) e RECORET et al. (2011) trabalhando com
Arabidopsis e Eucalyptus globulus, respectivamente, correlacionaram que o
acúmulo da algumas enzimas na parede celular poderia ser associado com a a
síntese de lignina, e consequentemente aumentando a espessura das paredes
de membrana nas células como uma defesa. Além disso, a degradação de
paredes e a biossíntese de lignina por meio da interferência com o
metabolismo redox tem sido mais uma evidência, fortalecendo assim a
correlação lignina-redox (VERA-SIRELA et al., 2010). Deste modo, o aumento
da lignina para os tecidos de folhas e raízes em cana-de-açúcar, bem como o
aumento do volume das células pode ser associado provavelmente ao estresse
provocado pelo tratamento com peróxido de hidrogênio (FAN et al., 2006).
Assim, os resultados da microscopia óptica e de varredura permitiram
observar que provavelmente houve um aumento do tamanho em todas as
células que compõe os tecidos de folhas e raízes nos diferentes tratamentos
(10 mM, 20 mM, 30 mM), onde pode ser verificados danos na membrana do
xilema traqueal. Estas células tem papel central na fisiologia da planta de
transportar e distribuir os fotoassimilados (KITIN et al., 2010; MILINHOS &
MIGUEL, 2013). Na folha tratada com 30 mM de H2O2 observamos em
destaque a degradação da parede do xilema causado pela possível ação do
H2O2. A morte celular de elementos traqueais é um exemplo bem conhecido da
morte celular programada. Este processo envolve o colapso do vacúolo
seguido de hidrólise do DNA nuclear e os conteúdos citoplasmáticos, como
resultado da ativação ou a libertação de enzimas hidrolíticas liberadas a partir
do vacúolo. Mesmo que o processo de morte celular do xilema esteja bem
descrito, pouco se sabe sobre o mecanismo molecular que o regula (VERA-
SIRELA et al., 2010).
A determinação do conteúdo de peróxido de hidrogênio nos tecidos foi
de extrema importância para este trabalho, assim confirmando a condição de
um estresse oxidativo. Por meio da análise dos dados da peroxidação lipídica
nos tecidos do meristema; folha e raiz foram observadas um aumento da
peroxidação lipídica principalmente nas concentrações de 10 mM e 30 mM, em

61
20 mM não foi observado a peroxidação lipídica quando comparado ao
controle.
Para a anatomia, os dados da microscopia óptica e do MEV, permitiram
observar também o aumento do tamanho das células que culminaram numa
desorganização estrutural das folhas, em especial nas células da coroa de
Kranz, xilema, floema e células buliformes, não sendo diferente para as raízes
que também tiveram as suas estruturas epidérmicas e endodérmicas com
formação de aerênquimas foram gravemente rompidas pelo aumento do
tamanho celular. Pode-se perceber que para a concentração 10 e 20 mM não
ocorreu muitos danos, já para 30 mM houve danos e degradação na região do
xilema. Em outros estudos na planta macieira (WANG et al., 2012) e arroz (LEE
et al., 2013) foram observados também com alterações semelhantes como a
lignificação e modificações morfológicas dos tecidos similar como observamos
para as plantas de cana-de-açúcar. Sendo assim, esta análise foi importante
por que inferiu que as plantas de cana-de-açúcar absorveram o H2O2 a que
foram expostas, e este provavelmente chegou no tecido foliar. Para as análises
enzimáticas foi observado que durante o tratamento com peróxido de
hidrogênio houve um aumento da EROs e uma maior atividade ou uma
redução de algumas enzimas que compõem o sistema antioxidante em plantas
como: superóxido dismutase (SOD), catalase (CAT) e ascorbato peroxidase
(APX). Os dados obtidos com as atividades enzimáticas indicaram um aumento
da SOD no meristema nas concentrações de 20 mM e 30mM; nas folhas na
concentração 30 mM e nas raízes na concentração 10 mM. As enzimas SODs
formam a primeira linha de defesa contra as espécies reativas de oxigênio,
catalizando assim a dismutação O2- em O2 e em seguida em H2O2. Sendo
assim, as enzimas SODs estão presentes nas células de acordo com o seu
metal cofator em três grupos: FeSOD presente nos cloroplastos; MnSOD
presente nas mitocôndrias e peroxissomas e as Cu/Zn-SOD presentes nos
cloroplastos, peroxissomos no citosol e possivelmente no espaço extracelular
(MOLINA-RUEDA et al., 2013) quando ocorre o acúmulo de H2O2 identifica-se
a lignificação nas paredes das membranas (VERA-SIRELA, et al., 2010). Como
foi visto nos resultados já citados pela análise da microscopia óptica e da
microscopia eletrônica em cana-de-açúcar.
A enzima ascosbato peroxidase (APX) apresentou aumento na atividade
para a concentração 10 mM no meristema e um aumento na concentração de

62
20 mM para a raíz quando comparado ao controle, sendo que, a APX para
folhas se mantiveram no mesmo nível de atividade para os diferentes tratados
quando comparado ao controle. A diferença entre a expressão das atividades
enzimáticas antioxidante nas folhas e raízes corroboram com os dados já
descritos em outros trabalhos: Nas plantas de arroz sob estresse salino,
observaram divergências nas atividades de APX e CAT (LEE et al., 2001); já na
planta Pinhão-manso sob condições de estresse de calor e seca, estas
apresentaram diferentes padrões para a atividade enzimatica de SOD; CAT e
APX (SILVA et al., 2010). Estudos com feijão caupi sob diferentes níveis de
salinidade, também foi observado diferentes atividades para as enzimas CAT e
SOD (MAIA et al., 2012) e em milho foi observado nos tecidos de folha a
disparidade entre as enzimas SOD; CAT e APX (JIANG & ZHANG, 2002).
Com base nos dados, uma hipótese para explicar por que a atividade de
APX não se alterou nas folhas de cana-de-açúcar durante o estresse oxidativo
seria devido a uma resposta de recuperação associado a outras vias anti-
oxidantes. Já para o decréscimo na atividade da CAT nas folhas e raízes de
cana-de-açúcar, pode ter comtribuído para o acúmulo de H2O2, aumentando a
peroxidação de lipídeos e danos nas paredes das membranas das células
associado a outras enzimas de membrana sob este estresse (SILVA et al.,
2010).

63
6. CONCLUSÃO
Com este trabalho, foi possível concluir que o tratamento com peróxido
de hidrogênio em diferentes concentrações (10 mM, 20 mM e 30 mM)
influenciou nas respostas integradas (através de interações morfológicas e
bioquímicas) de plantas de cana-de-açúcar submetidas a esse estímulo.
Alterações nos perfis de atividade das enzimas do sistema antioxidante de
plantas, associado à expressão diferencial de proteínas e modificações
estruturais de raízes e partes aéreas da cana-de-açúcar, permitiram a obtenção
de dados que podem contribuir para a criação de um modelo de homeostase
redox para plantas C4. Os dados dessa pesquisa possibilitaram compreender
um pouco mais acerca dos mecanismos de defesa de plantas de cana-de-
açúcar quando submetidas ao tratamento com peróxido de hidrogênio. Haja
vista a existência de uma rede integrada de resposta a estresse em sistemas
vegetais, o entendimento de como esses organismos se comportam diante
desse estímulo pode auxliar no melhoramento de plantas de grande valor
econômico como a cana-de-açúcar.

64
8. REFERÊNCIAS
ALLISON E.MCDONALD. Alternative oxidase: What information con protein sequence comparisons give us?.2009.Physiologia plantarum., 328-341. ALLEVA KARINA ; OSVALDO CHARA E GABRIELA AMODEO. 2012.Aquaporins: Another piece in the osmotic puzzle. FEBS Letters., 2991–2999. ANDREA BRA¨UTIGAM, KAISA KAJALA, JULIA WULLENWEBER, MANUEL SOMMER, DAVID GAGNEUL, KATRIN L. WEBER, KEVIN M. CARR, UDO GOWIK, JANINA MA, MARTIN J. LERCHER, PETER WESTHOFF, JULIAN M. HIBBERD, AND ANDREAS P.M. WEBER.2011.An mRNA Blueprint for C4 Photosynthesis Derived from Comparative Transcriptomics of Closely Related C3 and C4 Species. Plant Physiology., 142–156. ANA MILHINHOS CELIA M. MIGUEL.2013.Hormone interactions in xylem development: a matter of signals. Plant Cell., 867-83 AMALRAJ, R. S. et al. 2010. Sugarcane proteomics: Establishment of a protein extraction method for 2-DE in stalk tissues and initiation of sugarcane proteome reference map. Electrophoresis .,1959–1974. ANDRADE A; CELEDÓN P.A.F; LABATE C.A. O proteoma da madeira. Biotecnologia ciência & desenvolvimento. 2006. Disponível em: http://www.biotecnologia.com.br/revista/bio36/proteoma_36.pdf> Acesso em: 13 out. 2010. ARI PEKKA MA¨HO¨ NEN, MARTIN BONKE,LEILA AUPPINEN,MARJUKKA RIIKONEN,PHILIP N. BENFEY, AND YKA¨ HELARIUTTA.2000. A novel two-componenthybrid molecule regulatesvascular morphogenesis of the Arabidopsis root. GENES & DEVELOPMENT .,14:2938–2943. ARALDII ROSILAINE, FERDINANDO MARCOS LIMA SILVAI, ELIZABETH ORIKA ONOII,JOÃO DOMINGUES RODRIGUES.2010. Flowering in sugarcane. Ciência Rural., 694-702. BRECHENMARCHER, L. et al.2009. Establishment of a protein referente Map for soybean root hair cells. Plant Sciences., 670-682. BHATTACHARYA,MARIAN SABIA, MD. M. RAHMAN, YAN LI,ROCHELLE WEI, ASISH CHAUDHURI E HOLLY VAN. 2013. Genetic ablation of 12/15-lipoxygenase but not 5-lipoxygenase protects against denervation-induced muscle atrophy. ELSEVIER. BERDING, N. & ROACH, B.T. 1987. Germplasm collection,maintenance, and use. In: Sugarcane Improvement Through Breeding (Heinz D.J., ed.), Elsevier, Amsterdam, pp. 143-210. BRUTNELL, THOMAS P. 2012.Engineering C4 photosynthetic regulatory networks Sarit Weissmann and Thomas P Brutnell. Current Opinion in Biotechnology ., 23:298–304

65
BARTHA, R.1986 Biotechnology of petroleum pollutant biodegradation. Microbiol Ecol.,155.1986. BUTTERFIELD, M.K., D’HONT, A. AND BERDING, N. 2001. The sugarcane genome: A synthesis of current understanding, and lessons for breeding and biotechnology. Proc. S. Afr. Sug., 75: 1-5. BERTOMEU, JESU S MUN˜ OZ-, MARI´A ANGELES BERMU´ DEZ, JUAN SEGURA AND ROC ROS.2011.Arabidopsis plants deficient in plastidial glyceraldehyde-3-phosphate dehydrogenase show alterations in abscisic acid (ABA) signal transduction: interaction between ABA and primary metabolism. Journal of Experimental Botany., 1229–1239. BERTOMEU, JESÚS MUÑOZ-, BORJA CASCALES-MIÑANA, MANUEL ALAIZ, JUAN SEGURA AND ROC ROS.2010.A critical role of plastidial glycolytic glyceraldehyde-3-phosphate dehydrogenase in the control of plant metabolism and development. Plant Signaling & Behavior., 5:1, 67-69 CUI SUXIA, FANG HUANG, JIEWANG, XIAO MA, YONGSHENG CHENG AND JINYUAN LIU. 2005. A proteomic analysis of cold stress responses in rice seedlings. Proteomics., 3162–3172. CHRISTOPHE LALOI, DOMINIKA P. & KLAUS Apel. 2006. A genetic approach towards elucidating the biological activity of different restive oxygen species in Arabidopsis thaliana. Journal of Experimental Botany.,1719-1724. CUNHA, R.A.P. Atualmente do setor sucroenergético no Nordeste. Jornal Folha de Pernambuco.Disponível em:<http://www.canaldoprodutor.com.br>. Acesso: 24.09.2010. CONAB. A Companhia Nacional de Abastecimento.Disponível em:< http://www.conab.gov.br>. Acesso: 23.05.2013 CIB. Conselho de Informações sobre Biotecnologia. Disponível em:< http://cib.org.br>. Acesso: 14.11.2009. CARL W. GILBERT AND DENNIS E. BUETOW.1981.Gel Electrophoresis of Chloroplast Polypeptides: Comparison of One-Dimensional and Two-Dimensional Gel Analyses of Chloroplast Polypeptides From Euglena gracilis. Plant Physiol.,67, 623-628. CHEESEMAN, JOHN M..2013. Hydrogen Peroxide and Plant Stress: A Challenging Relationship. Plant Stress., 4-15. DAL’MOLIN, CRISTIANA GOMES DE OLIVEIRA, LAKE-EE QUEK, ROBIN WILLIAM PALFREYMAN,STEVENS MICHAEL BRUMBLEY, AND LARS KELD NIELSEN. 2010. C4GEM, a Genome-Scale Metabolic Model to Study C4. Plant Physiology., 1871–1885

66
D’HONT, A. AND GLASZMANN, J.C. 2001. Sugarcane genome analysis with molecular markers, a first decade of research. Proc. Int. Soc. Sugarcane. Technol. 24: 556-559. DOMONT G. B. et al. 2004. Fundamentos da análise proteômica. Laboratório de química de proteínas. Rio de Janeiro. DAVIDARIZA, et al. 2008. Changes in the protein profile of Quercus ilex leaves in response to drought stress and recovery. Journal of plant physiology p. 233-245. DAL’MOLIN, CRISTIANA GOMES DE OLIVEIRA, LAKE-EE QUEK, ROBIN WILLIAM PALFREYMAN,STEVENS MICHAEL BRUMBLEY, AND LARS KELD NIELSEN.2010. C4GEM, a Genome-Scale Metabolic Model to Study C4 Plant Metabolism. Plant Physiology., 1871–1885. DAVID L. NELSON, MICHAEL M. COX. Principios de bioquímica de LEHNINGER. 2011. ERIKA J. EDWARDS, COLIN P. OSBORNE, CAROLINE A. E. STRÖMBERG, STEPHEN A. SMITH. 2010.The Origins of C4 Grasslands:Integrating. SCIENCE.,328 EHLERINGER, JAMES R. Á THURE E. CERLING BRENT R. HELLIKER. 1997.C4 photosynthesis, atmospheric CO2. Oecologia., 112:285±299. FOYER, CHRISTINE H. AND SHIGERU SHIGEOKA. 2011. Understanding Oxidative Stress and Antioxidant Functions to Enhance Photosynthesis. Plant Physiology., 93–100. FU, JIANMING, IVANAMOMˇCILOVI´C, AND P. V. VARA PRASAD.2012. Roles of Protein Synthesis Elongation Factor EF-Tu in HeatTolerance in Plants. Journal of Botany., 835836 FRANCISCO VERA-SIRERA, EUGENIO G. MINGUET, SUNIL KUMAR SINGH, KARIN LJUNG,HANNELE TUOMINEN, MIGUEL A. BLÁZQUEZ, JUAN CARBONELL.2010. Role of polyamines in plant vascular development. Plant Physiology.,534-539. FAO. Organização das Nações Unidas para Alimentação e Agricultura. Disponivel em:<https://www.fao.org.br> . Acesso 18:26 01/04/13 GRIMALDI FILHO, G. Manual de técnica histologica. Rio de Janeiro: CME/IOC/Fiocruz, 1981. GOWIK & WESTHOFF. 2011.The Path from C3 to C4 Photosynthesis. Plant Physiology., 56–63. GOWIK, UDO AND PETER WESTHOFF. 2011.The Path from C3 to C4 Photosynthesis.Plant Physiology., 56–63.

67
GILL, SARVAJEET SINGH, NARENDRA TUTEJA. 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem., 909-30. GUEDES, H.V; et al. Quantificação de proteínas totais de bactérias Diazotróficas crescidas em meio de cultivo semi-sólido. EMBRAPA. 2007. GILBERTO BARBANTE KERBAUY. Fisiolofia Vegetal. 2004. GREER DH,WEEDON MM.2012.Modelling photosynthetic resposes to temperature of grapevine (Vitis vinifera cv. Semillon) leaves on vines grown in a hot climate. Plant, Cell and Environtnent., 20, 537-557 GOWIK, UDO AND PETER WESTHOFF.2011.The Path from C3 to C4 Photosynthesis. Plant Physiology., 56-63 GOLDEMBERG , JOSE, PATREICIA GARDABASSI.2010. The potencial for first geration ethanol production from sugarcane. Biofuels, Bioprod Bioref., 4 17-24 GERALD E. EDWARDS, VINCENT R. FRANCESCHI, AND ELENA V. VOZNESENSKAYA. 2004.Single-cell c4 photosynthesis versus the dual-cell (kranz) paradigm. Plant Biol., 55:173–96 GIRAUD, ESTELLE, OLIVIER VAN AKEN, VINDYA UGGALLA & JAMES WHELAN. 2012.REDOX regulation of mitochondrial function in plants.Plant, Cell and Environment.,35, 271–280 HURKMAN J. WILLIAM, TANAKA K. CHARLENE.1986. Solubilization of plant membrane proteins for analysis by two-dimensional gel electrophoresis. Plant physiol., 802-806. HOBE, STEPHAN, HOLGER FEY, HANS ROGL, HARALD PAULSEN. 2003. Protein structure and foldin: Determination of relative chlorophyll binding affinities in the major Ligth-havesting choropyll a/b complex. J Bio., 278:59. HELMUT, KIRCHHOFF, RICHARD M. SHARPE, MIROSLAVA HERBSTOVA, ROBERT YARBROUGH, AND GERALD E. EDWARDS.2013. Differential Mobility of Pigment-Protein Complexes in Granal and Agranal Thylakoid Membranes of C3 and C4 Plants. American Society of Plant Biologists.,497–507. IBGE – INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATISTICA, Produção Agrícola municipal 2006. Disponivel em: <http://www.ibge.gov.br/> Acesso em: 22 set. 2012 JIANG, MINGYI AND JIANHUA ZHANG.2002.Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J Exp Bot., 2401.

68
JELLOULI, N. et al. 2007.Proteomic analysis of Tunisian grapevine cultivar Razegui under salt stress. Journal of plant physiology., 471-481. JOANNE CHORY. 2010. Light signal transduction: an infinite spectrum of possibilities. The Plant Journal., 982-991 JUNQUEIRA L. C. U.; JUNQUEIRA, L. M. M. S. Tecnicas basicas de citologia e histologia. 1983. J PARKER, MENGMENG ZHU, AND SIXUE CHEN. 2012. Profiling Thiol Redox Proteome Using Isotope Tagging Mass Spectrometry. J Vis Exp., 61: 3766. JOHN M. CHEESEMAN.2007. Hydrogen Peroxide and Plant Stress: A Challenging Relationship. Plant Stress., 4-15 KRAUS, JANE ELIZABETH E MARCOS ARDUIN. Manual básico de métodos em morfologia vegetal .1997. KOLLER, YVES BALMER, ANTONIUS, GREGORIO DEL VAL, WANDA MANIERI, PETER CHURMANN, AND BOB B. BUCHANAN.2003. Proteomics gives insight into the regulatory function of chloroplast thioredoxins.PNAS.,100. KALDENHOFF RALF , LEI KAI, NORBERT UEHLEIN. 2013. Aquaporins and membrane diffusion of CO2 in living organisms. Biochimica et Biophysica Acta (BBA) - General Subjects., 64287. KOLLÁROVÁ R, OBLOŽINSKÝ M, KOVÁČIKOVÁ V.2013. Physiological aspects of lipoxygenase in plant signaling systems part I. Octadecanoid pathway. Ceska Slov Farm.,59-64. LEE, DONG-GI, et al. 2008. Chilling stress-induced proteomic changes in rice roots. Elsevier, p. 1-11. LEE MIN HEE, SEUNG GON WI,HYOUNGWOO BAE, JI EUN KIM, JAE-YOUNG CHO,SUNGBEOM LEE,JIN-HONG KIM,BYUNG YEOUP CHUNG.2013. Divergences in morphological changes and antioxidant responses in salt-tolerant and salt-sensitive rice seedlings after salt stress.Plant Physiology and Biochemistry., 325–335. LEE, DONG HEE,YOUNG SANG KIM, CHIN BUM LEE. 2001. The inductive responses of the antioxidant enzymes by salt stress in the rice (Oryza sativa L.) .Journal of Plant Physiology., 737–745 LI XIAONING , YINGLI YANG, JIN ZHANG, LINYUN JIA, QIAOXIA LI,TENGGUO ZHANG, KEXIONG QIAO, SHENGCHUN MA.2012. Zinc induced phytotoxicity mechanism involved in root growth of Triticum aestivum L.Ecotoxicology and Environmental Safety., 198–203. LOIACONO, F. VANESSA, AND MARIO C. DE TULLIO. 2012. Why We Should Stop Inferring Simple Correlations Between Antioxidants and Plant Stress Resistance: Towards the Antioxidomic Era. A Journal of Integrative Biology.,10.1089.

69
LANGDALE JA. 2011.C4 Cycles: Past, Present, and Future Research on C4 Photosynthesis. The Plant Cell., 23: 3879–3892. LOUISE E. ANDERSON AND IVAN PACOLD.1972.Chloroplast and Cytoplasmic Enzymes: IV. pea leaf fructose 1,6-diphosphate aldolases1'2 Plant Physiol., 49 393-397. MYOUNG DUCK KIM. et al. 2010.Enhanced tolerance to methyl viologen-induced oxidative stress and high temperature in transgenic potato plants overexpressing the CuZnSOD, APX and NDPK2 genes. Physiologia plantarum., 153-62. MACEDO S. ELISETE, et al.2009. Physiologic responses and gene diversity indicate olive alternative oxidase as a otential source for markers involved in efficient adventitious root induction. Physiologia plantarum., 532-552. MACHADO A. LAERTE; HABIB MOHAMED. Perspectivas e impactos da cultura de cana-de-açúcar no Brasil. Infobibos. 2009. MARTA S. LOPES, JOSE LUIS ARAUS, PHILIPPUS D. R. VAN HEERDEN AND CHRISTINE H. FOYER. 2011. Enhancing drought tolerance in C4 crops. Journal of Experimental Botany., 10.1093. MARÍA VALERIA LARA AND CARLOS SANTIAGO ANDREO. 2011.C4 Plants Adaptation to High Levels of CO2 andto Drought Environments. Abiotic Stress in Plants - Mechanisms and Adaptations., 428 MICHAEL W. GRAY, GERTRAUD BURGER, B. FRANZ LANG.1999.Mitochondrial Evolution. Science.,10.1126. MOORE. PAUL H. 2005.Integration of sucrose accumulation processes across hierarchical scales: towards developing an understanding of the gene-to-crop continuum. Plant Physiologist., 119-135 MURCHIE, E. H., M. PINTO AND P. HORTON. 2008. Agriculture and the new challenges for photosynthesis research.Journal compilation © New Phytologist., 10.1111. MILLER, GAD, NOBUHIRO SUZUKI1, SULTAN CIFTCI-YILMAZ1 & RON MITTLER. 2010. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant, Cell and Environment., 33, 453–467. MURCHIE, E. H., M. PINTO2 AND P. HORTON.2008. Agriculture and the new challenges for photosynthesis research. New Phytologist., 181: 532–552 MOLINA-RUEDA, JUAN JESU´ S CHUNG JUI TSAI, EDWARD G. KIRBY.2013. The Populus Superoxide Dismutase Gene Family and Its Responses to Drought Stress in Transgenic Poplar Overexpressing a Pine Cytosolic Glutamine Synthetase(GS1a).PLoS One.,10.1371.

70
MAIA, JOSEMIR MOURA, SERGIO LUIZ FERREIRA-SILVA, EDUARDO LUIZ VOIGT EDUARDO, CRISTIANE ELIZABETH COSTA DE MACEDO,LUIZ FERREIRA AGUIAR PONTE E JOAQUIM ALBENISIO GOMES SILVEIRA.2012. Atividade de enzimas antioxidantes e inibição do crescimento radicular de feijão caupi sob diferentes níveis de salinidade. Acta Botanica brasileira., 26. NEREA UBIERNA, WEI SUN, DAVID M. KRAMER & ASAPH B. COUSINS. 2013.The efficiency of C4 photosynthesis under low light conditions in Zea mays, Miscanthus x giganteus and Flaveria bidentis..Plant, Cell and Environment ., 36, 365–381. OLIVIER VAN AKEN, ESTELLE GIRAUD, RACHEL CLIFTON AND JAMES WHELAN. 2009.Alternative oxidase: a target and regulator of stress responses. Physiologia plantarum., 354-361. OLIVIER V. A; et al. 2009.Alternative oxidase: a target and regulator of stress responses. Physiologia Plantarum., 354-361. PATADE V. Y.; RAI A.N.; SUPRASANNA P.2010. Expression analysis of sugarcane shaggy-like kinase (SuSK) gene identified through cDNA subtractive hybridization in sugarcane (Saccharum officinarum L.). Protoplasma., 612-21.
PETER HR; RAY FE; SUSAN EE. Biologia Vegetal. 2011. PURY, D. G. G. DE & G. D. EARQUHAR.1997.Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models. Plant, Cell and Environtnent., 20, 537-557. PETER H. RAVEN; RAY F. EVENT, SUSAN E. EICHHORN. BIOLOGIA VEGETAL, 2011. PANT, B.D.; BUHTZ, A.; KEHR, J.; SCHEIBLE, W.R. 2008. MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis. The Plant Journal, 53: 731–738. PUERTO-GALÁN L, PÉREZ-RUIZ JM, FERRÁNDEZ J, CANO B, NARANJO B, NÁJERA VA, GONZÁLEZ M, LINDAHL AM, CEJUDO FJ.2013.Overoxidation of chloroplast 2-Cys peroxiredoxins: balancing toxic and signaling activities of hydrogen peroxide. Front Plant Sci.,4:310. PFAFFL, M. W. 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research, 29(9):45 QURESHI, M. I; S. qadir; LELLO zolla. 2007. Proteomics-based dissection of stress-responsive pathways in plants. Journal of plant physiology., 1239-1260. QUIROGA MÓNICA, CONSUELO GUERRERO, MIGUEL A. BOTELLA,ARACELI BARCELÓ, IRAIDA AMAYA, MAR A I. MEDINA, FRANCISCO J. ALONSO, SILVIA MILRAD DE FORCHETTI, HORACIO TIGIER & VICTORIANO VALPUESTA.2000. A Tomato Peroxidase Involved in the Synthesis of Lignin and Suberin. Plant Physiology.,1119-1128.

71
ROWAN F. SAGE.2003.The evolution of C4 photosynthesis. New Phytologist., 341–370. RAMAKERS, C.; RUIJTER, J.M.; DEPREZ, R.H.; MOORMAN, A.F. 2003. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR). Elsevier., 62-6. REDINALDO, S. S; BATISTA S. 2009.Extração e analise eletroforética em gel de poliacrilamida (SDS-PAGE) de proteínas totais de folha e raiz de Piper tuberculatum. Acta amazonica., 255-260. RAMOS O.H.P. & H.S. SELISTRE-DE-ARAUJO. 2001. Identification of metalloprotease gene families in sugarcane. Genetics and Molecular Biology., 285-290. RAIK WAGNER, HARALD AIGNER, CHRISTIANE FUNK.2012. FtsH proteases located in the plant chloroplast. Physiologia plantarum,203-14. SETZER J.As caracteristicas dos principais tipos de solo do estado de São Paulo. Boletim técnico do instituto agronômico do estado de São Paulo – BRAGANTIA. 1991. SCANDALIOS, J.G. 2005. Oxidative stress: molecular perception and transdution of signals triggering antioxidant gene defenses. Braz J Med Biol Res., 996-997. SREENIVASAN, T.V., AHLOOWALIA, B.S. AND HEINZ, D.J. 1987. Cytogenetics. In: Sugarcane Improvement Through Breeding. Elsevier., 211-253 SILVA, P. P. 2006. Divergência genética em genótipos de cana-de-açúcar (Saccharum sp.) através de características morfoagronômicos e por marcadores moleculares. Dissertação (mestrado) – Universidade de Alagoas, Centro de Ciências Agrárias. Rio Largo. SAVAJEET SINGH GILL, NARENDRA TUTEJA. 2010.Reactive oxygenspecies and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Molecular Biology., 110 067. SFAR, SONIA, RAOUDHA BOUSSOFFARA, MOHAMED TAHAR SFAR AND ABDELHAMID KERKENI.2013. Antioxidant enzymes activities in obese Tunisian Children. Nutrition Journal.,10.1186. SILVIERO, FÁBIO. BIOLOGIA CELULAR: bases moleculares e metodologia de pesquisa/organizador, 2013. SILVA, EVANDRO NASCIMENTO et al.2010. Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in Jatropha curcas plants. Plant Physiology., 1157-64.

72
SHEN GUOXIN, ZACH ADAM, HONG ZHANG.2007.The E3 ligase AtCHIP ubiquitylates FtsH1, a component of the chloroplast FtsH protease, and affects protein degradation in chloroplasts. The Plant Journal.,10.1111. SCORTECCI C., KATIA ; CRESTE, SILVANA ; CALSA, TERCILIO ; A., MAURO ; A. LANDELL, MARCOS G. ; FIGUEIRA, ANTONIO ; A., VAGNER. 2012. Challenges, Opportunities and Recent Advances in Sugarcane Breeding. In: Ibrokhim Y. Abdurakhmonov. (Org.). Plant Breeding. 1ed.Rijeka, Croatia: InTech., v., p. 267-296. SANTOS W. D. DOS,M. L. L. FERRARESE,C. V. NAKAMURA,K. S. M. MOURÃO,C. A. MANGOLIN,O. FERRARESE-FILHO.2008.Soybean (Glycine max) Root Lignification Induced by Ferulic Acid. J Chem Ecol.,1230–1241. TAKASHI H. KASUO S. 2010. Research on plant abiotic stress responses in the post-genome era: past, present and future. The Plant Journal., 1041-1052. TAKAHASHI , HIROKO, MASAKAZU IWAI, YUICHIRO TAKAHASHI, AND JUN INAGAWA.2006. Identification of the mobile light-harvesting complex II polypeptides for state transitions in Chlamydomonas reinhardtii. PNAS., 477–482. TAYLOR, NICOLAS L., YEW-FOON TAN, RICHARD P. JACOBY, A. HARVEY MILLAR.2009. Abiotic environmental stress induced changes in the Arabidopsis thaliana chloroplast, mitochondria and peroxisome proteomes.Journal of proteomics., 367-378. TRIVEDI DIPESH KUMAR, HARSHESH BHATT, RAVI KANT PAL, RENU TUTEJA, BHARTI GARG1, ATUL KUMAR JOHRI5,NEEL SAROVAR BHAVESH & NARENDRA TUTEJA.2013. Structure of RNA-interacting Cyclophilin A-like protein from Piriformospora indica that provides salinity-stress tolerance in plants. Scientific reports., 3 : 3001. User Guide For Reversed-Phase ZipTip C18, P10 – Millipore, Pipette Tips for SamplePreparation),(modificado). http://www.millipore.com/ziptip VAN DAMME, E.J.M., PEUMANS, W.J., BARRE, A., ROUGÉ, P. 1998. Plant lectins: a composite of several distinct families of structurally and evolutionary related proteins with diverse biological roles. Crit. Rev. Plant Sci. 17, 575–692. VANDENBORRE, GIANNI, GUY SMAGGHE B, ELS J.M. VAN DAMME.2011. Plant lectins as defense proteins against phytophagous insects. Plant defense. VELIKOVA, V.; YORDANOV, I.; EDREVA, 2000. A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants Protective role of exogenous polyamines. Plant Science, v. 151, p. 59 - 66, VOIGT EDUARDO LUIZ,JOSEMIR MOURA MAIA, SERGIO LUIZ FERREIRA-SILVA,CRISTIANE ELIZABETH COSTA DE MACEDO,LUIZ FERREIRA AGUIAR PONTE E JOAQUIM ALBENISIO GOMES SILVEIRA.2012.Atividade

73
de enzimas antioxidantes e inibição do crescimento radicular de feijão caupi sob diferentes níveis de salinidade. Acta Botanica Brasilica.,342-349. WANG, CHUANLI, LONGYUN GUO, YIXUE LI, ZHUO WANG. 2012.Systematic Comparison of C3 and C4 Plants Based on Metabolic Network Analysis. Systems Biology.,10.1186. WANNES DERMAUWA, NICKY WYBOUWA, STEPHANE ROMBAUTSB,C, BJÖRN MENTEND, JOHN VONTASE, MIODRAG GRBI_CF,G, RICHARD M. CLARKH, RENÉ FEYEREISENJ, AND THOMAS VAN LEEUWENA. 2012. A link between host plant adaptation and pesticide resistance in the polyphagous spider miteTetranychus urticae.PNAS.,113-122. WANG J. et al. 2010.Microllinearity between autopolyploid sugarcane and diploid sorghum genomes. BMC Genomics., 1471-2164. WANG SHUNCAI , DONG LIANG, CHAO LI, YONGLU HAO,FENGWANG MA, HUAIRUI SHU.2012. Influence of drought stress on the cellular ultrastructure and antioxidant system in leaves of drought-tolerant and drought-sensitive apple rootstocks. Plant Physiology and Biochemistry., 81–89. XIN-GUANG ZHU , LANLAN SHAN, YU WANG AND WILLIAM PAUL QUICK. 2010. C4 Rice – an Ideal Arena for Systems Biology Research. Journal of Integrative Plant Biology ., 8: 762–770. XU H, XU W, XI H, MA W, HE Z, MA M.2013.The ER luminal binding protein (BiP) alleviates Cd(2+)-induced programmed cell death through endoplasmic reticulum stress-cell death signaling pathway in tobacco cells. J Plant Physiol.,1434-41. YOSHIOKA, YASUSI YAMAMOTO.2011.Quality control of Photosystem II: Where and how does the degradation of the D1 protein by FtsH proteases start under light stress? – Facts and hypotheses.Journal of Photochemistry and Photobiology B: Biology., 229–235. YOSHIDA MITSUNORI, SHINSUKE OHNUKI,YOKO YASHIRODA, YOSHIKAZU OHYA.2013. Profilin is required for Ca2+ homeostasis and Ca2+-modulated bud formation in yeast. Original paper- springer., 288:317–328. ZAIA, D. A.M. Determinação de proteínas totais via espectrofotometria: vantagens e desvantagens dos métodos existentes. Química Nova. 1998. ZÖRB C, HERBST R, FORREITER C, SCHUBERT S.2009.Short-term effects of salt exposure on the maize chloroplast protein pattern. Proteomics., 4209-20.