effects of hydrodynamic regime on photosynthesis in the green alga
TRANSCRIPT

University of South FloridaScholar Commons
Graduate Theses and Dissertations Graduate School
2004
Effects of hydrodynamic regime on photosynthesisin the green alga CaulerpaMark D. DriscollUniversity of South Florida
Follow this and additional works at: http://scholarcommons.usf.edu/etd
Part of the American Studies Commons
This Thesis is brought to you for free and open access by the Graduate School at Scholar Commons. It has been accepted for inclusion in GraduateTheses and Dissertations by an authorized administrator of Scholar Commons. For more information, please contact [email protected].
Scholar Commons CitationDriscoll, Mark D., "Effects of hydrodynamic regime on photosynthesis in the green alga Caulerpa" (2004). Graduate Theses andDissertations.http://scholarcommons.usf.edu/etd/1018

Effects of hydrodynamic regime on photosynthesis in the green alga Caulerpa.
by
Mark D. Driscoll
A thesis submitted in partial fulfillment
of the requirements for the degree of Master of Science
Department of Biology College of Arts and Science University of South Florida
Major Professor: Florence I.M. Thomas, Ph.D.
Kevin S. Beach, Ph.D. Clinton J. Dawes, Ph.D.
Date of Approval: March 19, 2004
Keywords: PAM fluorescence, hydrodynamics, primary production, boundary layers,
photosystem II kinematics.
© Copyright 2004, Mark D. Driscoll

Dedication
This thesis is dedicated to my wife, Jodie Corsetti Driscoll, for her patience,
encouragement and support.

Acknowledgements
I would like to thank Dr. Flo Thomas for her friendship, support and guidance
over the past few years, Dr. Kevin Beach for introducing me to the world of marine
algae, and for encouraging me to pursue a graduate degree, and Dr. Clinton Dawes for
thoughtful review of my thesis. This research was made possible by a National Science
Foundation PECASE award (OCE-9996361) to Florence I. M. Thomas and a SIGMA XI
grants in aid of research award to Mark D. Driscoll. I would also like to thank Chris
Cornelisen and Sean Kinane for field assistance, Jodie Corsetti Driscoll and Dan Huber
for laboratory assistance and Dan Huber, Justin Bowels, Dr. Bruce Cowell, and Dr. Gary
Huxel for assistance with statistical analysis. I would also like to thank my parents, John
and Karen Driscoll, for their un-wavering support throughout my educational career.

i
Table of Contents
List of Tables ii List of Figures iii Abstract v Introduction 1 Limits of Algal Productivty 2 How Hydrodynamics affects algal productivity 6 Objectives 12 Background information on Caulerpa spp. 13 Accoustic Doppler Velocimenter 16 The PAM Fluorescence measuring technique 17 Materials and Methods 22 Caulerpa 24 Measurement of PSII kinematics 25 Field Experiments 28 Time in Flow 30 Results 33 Does the magnetic sample holder affect values of Pmax? 33 Laboratory measurements of PSII kinematics in C. mexicana after a short eposure to hydrodynamic regime. 33 In situ field measurements of PSII kinematics on C. sertularioides. 34 Laboratory measurements of PSII kinematics in C. racemosa after long term exposure to hydrodynamic regime. 34 Discussion 36 References 67

ii
List of Tables
Table 1. Values of Pmax, a and Ik measured on C. sertularioides in situ. 62 Table 2. Values of Pmax, a, Ik, qP and NPQ measured on C. racemosa in laboratory flumes over 7 hours. 63 Table 3. Values of NPQ at each measurement hour. 64 Table 4. Values of Pmax, a, Ik, qP and NPQ measured on C. racemosa Over 5 hours in a laboratory flume. 65 Table 5. Values of NPQ after 3 hours of immersion. 66

iii
List of Figures
Figure 1. Diagram of anti-herbivory metabolite caulerpenyne 42 Figure 2. The three main energy pathways for photosystem II 43 Figure 3. Skematic diagram of the pulse amplitude modulated fluorescence measuring principle in both light and dark adapted samples. 44 Figure 4. Representative curve that would be analyzed using the Platt inhibition model. 45 Figure 5. Representative curve that would be analyzed using the hyperbolic tangent model. 46 Figure 6. Drawing of Caulerpa sertularioides. 47 Figure 7. Drawing of Caulerpa racemosa. 48 Figure 8. Drawing of Caulerpa mexicana. 49 Figure 9. Collection site for C. racemosa. 50 Figure 10. 110 Litre racetrack flume. 51 Figure 11. Maps of study sites in the Florida Keys. 52 Figure 12. Orientation of the acoustic doppler velocimeter during field measurements. 53 Figure 13. Ratio of Pmax in fast flow to Pmax in slow flow at each measuring interval. 54 Figure 14. Results of dark acclimation trial using magnetic leaf clips under full Illumination and a dark room. 55 Figure 15. Values of Pmax for C. mexicana run at high and low velocities in a laboratory flume. 56 Figure 16. In situ measures of Pmax in C. sertularioides. 57

iv
Figure 17. In situ measurements of saturation irradiance in C. sertularioides. 58 Figure 18. Laboratory measurements of NPQ in C. racemosa. 59 Figure 19. Laboratory measurements of Pmax in C. racemosa. 60 Figure 20. Laboratory measurements of NPQ in C. racemosa. 61

v
Effects of hydrodynamics on photosynthesis in the green alga Caulerpa.
Mark D. Driscoll
ABSTRACT
The delivery of nutrients to the surface of marine algae can be controlled by the
local hydrodynamic regime: in higher flow velocities, the Diffusive Boundary Layer
(DBL) at the uptake surface is thinner, which can increase the flux of dissolved chemicals
to the algal surface. If the primary productivity of an alga is controlled by the availability
of a dissolved chemical, increased water flow should result in greater primary
productivity due to increased chemical flux. To test the hypothesis that increased water
flow will increase Photosystem II kinematics (PSII) in the green alga Caulerpa we used a
Diving Pam Fluorometer to measure the maximum relative electron transport rate
(Pmax), Saturation Irradiance (Ik), Non-photochemical quenching (NPQ), the light
limited slope of photosynthesis vs. irradiance curve (α) and photo-chemical quenching
(qP) and compared these measured values among treatments of varying flow speeds in a
portable laboratory flume. We also measured the influence of water flow on values of
Pmax, Ik, α , qP and NPQ in the field. Results showed that in C. racemosa collected from
Tampa bay, and tested in a laboratory flume, values of Pmax and Ik were positively
correlated to increase water flow, possibly indicating mass-transfer limitation. C.
mexicana, collected from the Florida Keys, showed a decrease in values of Pmax, and Ik

vi
with increasing water velocity in flume experiments, indicating that the increased flow
was resulting in physiological stress. This result was supported with field measurements
for C. sertularioides, which showed a negative correlation between Pmax and flow
velocity and Ik and flow velocity.

1
Introduction
Marine macro-algal communities represent a highly productive and often
overlooked component of marine systems (Hader 1999, Hurd 2000). As primary
producers, providers of food and shelter to a wide variety of marine life and as sediment
stabilizers (Williams 1990), macroalgae play an integral role in the health and
development of many coastal marine ecosystems (Wheeler and Björnsäter 1992, Gacia et
al. 1996, Campbell 2001). Rates of algal productivity have been shown to be some of the
highest on the planet (Littler 1980, Hurd 2000) and in near shore marine communities,
macroalgae play an important role in overall production (Wing and Patterson 1993,
Hader et al. 2001). While the vast majority of macroalgae are a healthy component of
marine ecosystems, some are dangerous invaders, destroying native eco-systems and
remaining unresponsive to eradication efforts (Hill et al. 1998, Argyrou et al. 1999,
Aussem and Hill 1999, Ceccherelli et al. 2000, Uchimura et al. 2000, Stimson et al. 2001,
Harris and Tyrell 2001).
Members of the genus Caulerpa (Bryiopsidacea) are important components of
tropical marine ecosystems worldwide (Collado-Vides and Robledo 1999, Collado-Vides
2002). With approximately 70 species within the genus, Caulerpa is often one of the
dominant macro-algal species within tropical shallow water habitats (Gacia et al. 1996).
Recently coastal marine systems in California and the Mediterranean Sea have been

2
invaded by a rogue strain of C. taxifolia (Vahl) C. Agardh, which has overgrown native
seagrass beds and detrimentally affected the native marine ecosystems (Ceccherelli and
Cinelli 1999, Cecherelli et al. 2000, Meinesz et al. 2001, Ceccherelli and Sechi 2002,
Ceccherelli and Campo 2002, Boudouresque et al. 2002). It is clear that macroalgae are
important components of healthy marine systems, as well as devastating components of
some disturbed systems. Due to the substantial impact that macro-algae can have on
coastal marine systems it is important that we understand the factors that control algal
production.
Limits of Algal Productivity:
Marine macrophyte physiology shows an incredible amount of diversity (Littler
1980, Littler and Arnold 1982, Saffo 1987, Koehl and Alberte 1988, Jensen et al 1985,
Hader et al 1996, Beach et al 1997, Stevens and Hurd 1997, Larned 1998, Williams and
Carpenter 1998, Hurd 2000, McCook et al 2000, Stimson and Larned 2000, Hillis 2001,).
This diversity allows algal communities to flourish in a vast range of environmental
conditions. Researchers have shown that macroalgae are able to change their overall
morphology (Koehl and Alberte 1988), pigment quality and quantity (Beach and Smith
1996a) and photosystem structure (Fork and Satoh 1986, Beach and Smith 1996b) in
response to varying environmental conditions. However, even with the highly plastic
nature of these organisms, light and nutrients can severely limit photo-production

3
(Hatcher and Larkum 1983, Duke et al. 1989, Wheeler and Björnsäter 1992, Fong et al.
2001, Harrison and Hurd 2001).
The availability of saturating irradiance is a controlling factor for photosynthesis
in marine macroalgae (Engleman 1883, Beach and Smith 1997a, Beach and Smith 1997b
Beach et al. 1997, Gomez et al. 1998, Cloern 1999, Yentsch et al. 2002). The availability
of light also contributes to the vertical zonation of marine algae (Engleman 1883). While
it is well known that not enough light can limit the primary production of algae, it must
also be considered that an overabundance of light can be detrimental given certain
circumstances (Jensen et al 1985, Beach et al 1995, Beach and Smith 1996, Hader et al
1996, Beach and Smith 1997a, Beach and Smith 1997b, Beach et al 1997, Hader et al.
1999, Hader et al. 2000, Gorbunov et al. 2000). Species that are low light acclimated can
see a drastic drop in photosynthetic activity when brought into drastically higher light
regimes (Hader et al. 1996). It is believed that this decrease is caused by damage to the
D1 protein complex within photosystem II of the electron transport chain (Greene et al.
1994, Hader 1996, Hader 1999). If damage to the D1 protein occurs, the protein must be
replaced before normal functions resume. Light inhibition can often be overcome if the
algae are acclimated in small steps to the desired level of irradiance (Hader et al 1996).
Researchers have shown that given enough time to acclimate to a new light regime,
macroalgae are able to adjust pigment levels and pigment ratios, allowing the organism to
more efficiently utilize the available light regime (Riechert and Dawes 1986).
When light levels are saturating, nutrient availability becomes the major limiting
factor to macroalgal production (Chapman 1974, Duke et al. 1989, Björnsäter and

4
Wheeler 1990, Wheeler and Björnsäter 1992, Greene et al. 1994, Martin et al. 1994,
Jones et al. 1996, Pedersen and Borum 1996, Landry et al. 1997, Ceccherelli and Cinelli
1999, Schwartz et al. 2000). This is especially evident in tropical environments where
coastal waters are known to be nutrient deplete (Hatcher and Larkum 1983, Carpenter et
al. 1991, Stimson and Larned 2000, Fong et al. 2001). The major limiting nutrients in the
marine environment are nitrogen (N) (Ryther and Dunstan 1971, Kinsley 1974,
Björnsäter and Wheeler 1990, Wheeler and Björnsäter 1992, Duke et al. 1989), phosphate
(P) (Kinsey and Domm 1974, Björnsäter and Wheeler 1990, Wheeler and Björnsäter
1992, Atkinson and Smith 1983), carbon (C) (Björnsäter and Wheeler 1990, Wheeler and
Björnsäter 1992) which is most often in the form of bi-carbonate in marine systems, and
micronutrients such as iron (Fe) (Greene et al. 1994, Martin et al. 1994).
Many studies have experimentally examined the effect of nutrient limitation in
algae; however, due to the metabolic requirements of individual species, nutrient
limitation varies by species and study site. For example increased productivity was seen
when samples of the coral reef algae Chlorodesmis fastigiata were artificially fertilized
with nitrate and phosphate (Kinsey and Domnm 1974). Similarly, chlorophyll content
and overall biomass was shown to increase after N and N+P additions to the water
column (Menendez et al. 2002). Greene et al. (1994) proposed that in areas where C, N
and P are not limiting, Fe is the main limiting nutrient to phytoplankton growth. By
seeding the open ocean with Fe Green et al. (1994) showed that plankton growth and
productivity was shown to drastically increase.

5
Nutrient enrichment is not always beneficial to marine vegetative assemblages.
Taylor et al. (1995) showed that while nutrients may positively affect the growth of one
component of an ecosystem, the same nutrient pulse could be detrimental to other
components. Their research showed that the biomass of Nannochloropsis spp. increased
as a result of N fertilization as well as N+P fertilization, the nutrient pulse resulted in a
loss of below ground biomass for the associated seagrass community.
Certain species of algae are known to display mechanisms that allow them to
obtain nutrients from sources other than the water column. The thallus of Dictyospheria
cavernosa grows in such a manner as to isolate a parcel of water between it and the
benthos (Larned and Stimpson 1996). This “chamber” provides two sources of nutrients
other than what is available from the water column. The first source is metabolic waste
products from invertebrates that colonize the inside of the chamber, and the second
source is the efflux of nutrients from the sediment that is trapped under the algal thallus.
The nutrient concentrations inside these chambers were found to be ten times higher than
the surrounding water column (Larned and Stimpson 1996). This mechanism was
hypothesized to be the factor that allows D. cavernosa to thrive in waters where the
ambient water column nutrients are not high enough for algal growth (Larned and
Stimpson 1996). Transport of nutrients from the sediment to the vegetative tissue of
algae has also been seen . Uptake of labeled 15N by the rhizoids of the alga Caulerpa
cupressoides was shown by Williams (1984), demonstrating that in some species of
Caulerpa sediment nutrients can play a large role in the productivity of the algae.

6
In terms of nutrient availability, organisms can be considered to be either
physically limited, biologically limited or in between the two. In a physically limited
system, the organism is not experiencing a high enough nutrient flux to saturate its
physiological processes (e.g. uptake potential, enzyme loads). Biologically limited
systems receive more than enough nutrient flux to saturate their processes; however,
these organisms do not have the capability to process all of the available nutrients (e.g.
not enough uptake sites, insufficient enzyme stores).
How hydrodynamics affects algal productivity:
Many researchers have proposed that the hydrodynamic regime experienced by
an organism can have a large effect on its metabolic processes (Odum 1956, Wheeler
1980, Riber and Wetzel 1987, Fonseca and Kenworthy 1987, Koehl and Alberte 1988,
Patterson et al. 1991, Koch 1993, Williams and Carpenter 1998, Kuffner 2001). Studies
have shown that relative irradiance levels (Dromgoole 1987, Koehl and Alberte 1988,
Patterson 1992, Wing and Patterson 1993, Wing et al. 1993) and nutrient delivery (Koehl
and Alberte 1988, Bilger and Atkinson 1992) can be drastically changed depending on
the ambient hydrodynamic regime of a given area. Changes in the local hydrodynamic
regime can affect an organism in two major ways. The three dimensional orientation of
the organism can be changed, affecting the relative light levels and the structural reaction
to flow (Koehl and Alberte 1988). Secondly, the nutrient delivery to the organism’s

7
surface can be affected by disruptions to the boundary layers surrounding the organism
(Gerard 1982, Thomas and Cornelisen 2003)
The flow of water past a submerged organism results in drag forces acting on that
particular organism, which can have a major impact on its physiology. Organisms
located in high-energy areas generally experience more drag than those found in sheltered
areas. Temperate kelps are able to protect themselves from this increased drag by
changing the morphology of their blades so as to reduce form drag (Koehl and Alberte
1988). In high velocity areas the blades of Nereocystis leutkiana take on a smooth and
narrow form that allows the blades to clump together into a streamlined cone, reducing
drag. In low velocity areas the same blades take on a more ruffled, wider form (Koehl
and Alberte 1988). This clumping in high velocity reduces the cumulative amount of
light that reaches the entire plant due to self-shading. In low velocity areas the blades do
not clump together, resulting in a higher surface area exposed to irradiance.
The clumping of blades into a more streamlined shape is not the only condition
related to hydrodynamics regimes that can be detrimental to plants. Dromgoole (1987),
Wing and Patterson (1993) and Wing et al. (1993) showed that hydrodynamic regimes
can drastically alter the light climate reaching a benthic community. Light flecks are
periods of intense light caused by canopy movement or surface waves. Surface waves in
particular can cause a short term, intense focusing of light, up to five times the ambient
levels onto the benthos (Sagert and Schubert 2000, Schubert et al. 2001). It is possible
that under certain conditions, these periods of intense light are strong enough to cause

8
permanent damage to the photosystems of aquatic primary producers, resulting in photo-
inhibition.
The local hydrodynamic regime can affect the nutrient delivery to an algal thallus
by increasing advection of nutrients over the surface and by decreasing the thickness of
the diffusive boundary layer (DBL (a gradient in concentration where chemical transport
is controlled only by molecular diffusion, not momentum.)). Hydrodynamics controls
the thickness of the DBL and therefore greatly affects nutrient delivery (Kohel and
Alberte 1988, Bilger and Atkinson 1992). If nutrient delivery is influenced by
hydrodynamics, then metabolic processes dependent on that delivery are then linked to
hydrodynamics as well.
The momentum boundary layer (MBL) is usually a completely turbulent region,
where the rates of chemical transport are dominated by inertial forces. The turbulence
in this layer is a function of water velocity and the properties of the benthic roughness
elements. Due to the presence of turbulence in the water column, the rates of chemical
transport in this layer are much higher than would be expected from molecular diffusion
alone (Kays and Crawford 1993). Eddies vertically mix dissolved chemicals through this
layer to the surface of the viscous sub layer, transporting them through the water column
faster than molecular convection or molecular diffusion.
The viscous sub layer (VSL) is a thin region immediately next to the surface of a
solid. In this layer, the vertical transport of momentum due to turbulence is no longer
effective, often resulting in a region of laminar flow. Chemical transport in this region is
governed by viscous transport, or molecular conduction between the layers (Kays and

9
Crawford 1993). The concentration of solutes within the viscous sub layer is controlled
by the delivery of chemicals from the momentum boundary layer. In areas of low
turbulence, the transport of these solutes may be slower, resulting in a lower
concentration within the viscous sub layer.
Below the viscous sub layer is the diffusive boundary layer (DBL). In this
region, the transport of solutes to and from a solid is completely reliant on molecular
diffusion. If an organism uses a chemical faster than it can be delivered through this
layer, it will become physically limited. Similarly, if an organism excretes a metabolic bi-
product into this layer faster than it can diffuse out, then an elevated concentration of that
bi-product will occur in the water next to the tissue surface. For plants, boundary layers
can cause a major problem. All photosysnthesis results in the production of oxygen,
which can compete with carbon for binding sites on the enzyme Ribulose 1-5
Bisphosphate Carboxylase Oxygenase (RUBISCO). When this happens, primary
productivity may be reduced due to the oxygen build up in the DBL.
Hydrodynamic regimes may greatly affect the nutrient limited metabolic
processes such as photosynthesis under some circumstances. As the boundary layers
surrounding a submerged solid are broken up by turbulent energy, or made thinner by
increased velocity, molecular diffusion to that solid is increased. In this manner, even in
low nutrient waters, the relative nutrient flux experienced by an organism can be quite
high if the area is swept by high velocity water flow. Conversely, if there is little
turbulent energy or velocity in the water column, an organism in a nutrient rich
environment may experience nutrient limitation due to the thickness of the diffusive

10
boundary layer. Two branches of research have developed models of how water flow
affects aquatic organisms. The first branch focuses on the effects of water flow on
photosynthesis and gross morphology, and the second branch has demonstrated the
dependence of nutrient uptake on water velocity.
Wheeler (1980) demonstrated that increased water velocity influenced the
photosynthetic rates of Macrocystis pyrifera during laboratory experiments using 3.0 cm
tissue discs. Oxygen evolution was shown to increase from a level of 0.3µmol O2 cm-2 in
stagnant water to a level of 1.3 µmol O2 cm-2 in water moving at the equivalent of 7.0 cm
s-1. Similar results were reported by Gerard (1982), who determined that small tissue
samples of M. pyrifera is mass transfer limited in flows less than 3.0 to 4.0 cm s-1 . These
studies were followed by Koehl and Alberte (1988) who demonstrated increased
photosynthetic activity in tissue discs of Nereocystis leutkiana as a result of increased
flow regimes in laboratory experiments. Koehl and Alberte (1988) also demonstrated
that the morphology of N. leutkiana is highly variable depending upon environmental
conditions. In high flows, blades were found to be smooth and without marginal spines,
allowing the blades to clump together and reduce form drag, where as in low flow, blades
were found to have large marginal spines and ruggose morphology, increasing turbulence
around the blade. This increased turbulence acts to disrupt the viscous sub-layer and
increase the nutrient flux to the edge of the diffusive boundary layer. In several studies
that utilized whole plants, instead of dissected tissue disks and blades, rates of primary
production ([µgO2 {µg Chl a}-1 h-1]) and acetylene reduction, an indicator of nitrogen
fixation, in tropical algal turfs have been positively correlated to water velocity (0-22cm

11
s-1) (Carpenter et al. 1991, Williams and Carpenter 1998). The effect of water velocity on
photo-production in marine algae is not uniform, Koch (1993) demonstrated that
increasing friction velocities (u* (a measure of the intensity of the turbulence in a
system)) from 0.0 cm s -1 to 0.3 cm s -1 significantly increased the photosynthetic rate of
Ulva lactuca. However, increasing µ* beyond 0.3 cm s -1 had no effect on the algae, and it
was postulated that under less than saturating irradiance increased water flow could be
detrimental to photosynthesis. Koch postulated that this decreased performance in higher
flows may have been due to the algae being accustomed to the flow regime in the area
where it was collected. By changing this regime, the plants may have become stressed,
reducing their efficiency (Koch 1993).
In addition to measuring primary productivity in response to varying
hydrodynamic regimes, several studies have investigated the affects of water flow on
nutrient uptake rates. Increased N and P uptake rates as a function of velocity have been
widely reported in the literature (Atkinson and Bilger 1992, Bilger and Atkinson 1992,
Bilger and Atkinson 1995, Thomas and Atkinson 1997, Thomas et al. 2000, Thomas and
Cornelisen 2003). Atkinson and Bilger have mainly worked on organisms in flume
systems. These studies noted dependence of the first order rate constant for the uptake of
important nutrients by coral reef assemblages on hydrodynamic regime. Along the same
lines, Thomas and Cornelisen have focused on field based experiments using a large field
flume capable of encompassing 3.7 m2 of the benthos. These experiments have
demonstrated that nutrient uptake of an entire community in situ is directly correlated to
water velocity. Additionally, these studies showed that the uptake rates of individuals

12
within the community are correlated to water flow and to whole community uptake
(Cornelisen and Thomas 2002).
Objectives:
To clarify the role of hydrodynamics in photo-production new technology that
allows for measurements of hydrodynamic regimes and direct measurements of
photosystem II kinematics will be used in situ. By measuring PSII kinematics in
laboratory experiments, I intend to show that a Diving PAM Fluorometer is capable of
measuring changes in PSII kinematics due to varying hydrodynamic regimes. I also plan
on utilizing field measurements to directly measure PSII kinematics without removing
the plants from their natural environmental conditions, which has been suggested as a
stressor to aquatic vegetation (Koch 1993). These studies will focus on the genus
Caulerpa due to its extensive presence in near-shore tropical and sub-tropical marine
systems. The detailed goals of this thesis are:
1. To determine if hydrodynamic regime affects photosystem II kinematics
in situ and in a controlled laboratory flume.
2. To determine if the effects of hydrodynamic regime on photosystem II
kinematics can be measured by integrating data collected with an Acoustic
Doppler Velocimeter and a Diving Pam Flourometer.

13
3. To determine the magnitude of the dependence of photosystem II
kinematics on hydrodynamic regime within the genus Caulerpa.
Background information on Caulerpa spp.
Research has shown that members of the genus Caulerpa are important
components of tropical marine ecosystems around the world (Collado-Vides and Robledo
1999, Collado-Vides 2002). The morphology of the genus Caulerpa is highly diverse,
and within species the morphology is amazingly plastic (Gacia et al. 1996, Benzie et al.
2000, Collado-Vides 2002), changing drastically in accordance to environmental
conditions. As early as 1906 water motion and ambient light levels were being
considered as important factors affecting the morphology of Caulerpa (Svedelius 1906).
Barilotti (1970) showed that the morphology of C. prolifera (FØrsskål) Lamouroux was
determined by available light levels; Peterson (1972) then showed a similar dependence
on ambient light levels in C. racemosa.
The genus Caulerpa has drawn special attention in the Mediterranean Sea, where
two species, Caulerpa taxifolia and C. racemosa (Forsskål) J. Agardh, have become
dominant invaders (Ceccherelli and Cinelli 1999, Cecherelli et al. 2000, Ceccherelli and
Piazzi 2001, Meinesz et al. 2001, Ceccherelli and Sechi 2002, Ceccherelli et al. 2002,
Ceccherelli and Campo 2002, Verlaque et al. 2002). Since its introduction off the coast
of Monaco in 1984, C. taxifolia has been shown to out-compete the seagrasses Posidonia
oceanica (Ceccherelli and Cinelli 1999, Cecherelli et al. 2000) and Cymodocea nodosa

14
(Ceccerelli and Sechi 2002). Caulerpa racemosa, which is an immigrant species from
the red sea, has shown a similar pattern as C. taxifolia in the Southern Mediterranean Sea,
out-competing C. nodosa, Zostera noltii (Ceccherelli et al. 2002) and Posidonia oceanica
(Ceccherelli et al. 2000). The negative effect of these algae has not been limited to the
overgrowth of submerged aquatic vegetation. Arigoni et al (2002) showed that native
labrid fishes underwent a drastic color change in C. taxifolia bed as compared to normal
color morphs in P. oceanica beds. Decreases in invertebrate abundance (Bellan-Santini
et al. 1996), species richness, and fish biomass (Francour et al. 1995, Relini et al. 2000)
were also seen within C. taxifolia beds as compared to natural seagrass assemblages. The
high levels of secondary metabolites present in Caulerpa spp may explain this decline in
species diversity. The acetylenic sesquiterpene metabolite caulerpenyne (Figure 1), which
is the main anti-herbivory defense in Caulerpa spp. has been shown to compromise a
substantial (0.2-13.0%) component of the frond weight depending on the season (Dumay
et al. 2002). The high levels of secondary metabolites in Caulerpa is not surprising,
given that the genus is mainly found in tropical waters which are known for high rates of
herbivory (Pennings and Paul 1992, Cronin and Hay 1996).
Data collected on Caulerpa spp. has demonstrated in many systems its ability to
colonize a wide range of substrate types, ranging from muddy unconsolidated sediments
to rocky intertidal areas (Meinesz et al. 1995, Chisholm and Jaubert 1997, Ceccherelli
and Cinelli 1998, Arigoni et al. 2002). Caulerpa has been reported at depths of 50
meters, well below the photic zone of most green algae (Arigoni et al. 2002). This
flexibility allows Caulerpa a vast range of options when settlement of fragments or

15
zygotes occurs, giving Caulerpa an advantage over species that are more selective in
their substratum choices. Ecologically, Caulerpa has shown that it is an important
component in the succession of tropical seagrass beds (Williams 1990). By consolidating
loose sediments, Caulerpa makes it possible for climax species, such as seagrasses to
colonize an area.
The growth strategy of Caulerpa is based on an extremely fast rate of vegetative
expansion (Chisholm and Jaubert 1997, Ceccherelli and Piazzi 2001). This rapid growth
allows the alga to quickly extend new tissue into a large area immediately surrounding
the initial settlement plot. Caulerpa is known to grow at night, as is seen by the white
growing tips visible in the early morning (Dawes and Barilotti 1969). As light levels
increase, cytoplasm replaces the “meristemplasm” that occupies the white areas of the
cell and Chloroplasts re-position themselves in this newly available area (Dawes and
Barilotti 1969). Like many members of the Bryopsidales, Caulerpa has the ability to
reproduce both sexually via holocarpy (Zuljevic and Antolic 2000) and asexually through
vegetative fragmentation (Smith and Walters 1999, Ceccherelli and Piazzi 2001, Walters
et al. 2002). Vegetative fragmentation is an extremely effective mechanism of
reproduction in Caulerpa, fragments as small as 2.0 mm have been shown to successfully
settle, attach and grown into full size plants (Goddard and Dawes 1983, Smith and
Walters 1999, Ceccherelli and Piazzi 2001). The effectiveness of this growth strategy is
seen in C. taxifolia, which, in the Mediterranean has spread solely through vegetative
fragmentation, as only male gametes have been observed (Zuljevic and Antolic 2000).
The same is true for C. racemosa in the southern Mediterranean; no gamete production

16
has been seen in the alga, meaning that while sexual reproduction of C. racemosa is
important in tropical waters, vegetative growth and fragmentation are solely responsible
for the alga’s success in the Mediterranean Sea (Ceccherelli and Piazzi 2001).
Acoustic Doppler Velocimeter
Early studies utilized methods such as hot film anemometers, plaster of paris
dissolution, and “life saver” dissolution to measure bulk flow velocities (Wheeler 1980,
Koehl and Alberte 1988) Plaster of paris and life saver dissolution give a general idea of
water velocity by measuring the amount of mass lost over a given time. Identical
samples can be calibrated at a known water velocity and compared to samples tested
(Wheeler 1980, Koehl and Alberte 1988). Hot film anemometers measure the advection
of heat from the probe tip and correlate it with velocity (Koch 1993). These techniques,
while informative, do not measure the turbulent energy in a system, which affects the
DBL. Acoustic Doppler Velocimeters measure water flow in three vectors
simultaneously; “x” which is parallel to the bulk flow, “y” which is perpendicular to the
bulk flow, and “z” which is the vertical perpendicular to the bulk flow. The amount of
variation between these measures allows researchers to collect data on the amount of
energy, or turbulence in the system.

17
The PAM Fluorescence Measuring Technique
Traditional methods of measuring primary production have relied on gas
exchange or increased biomass as the primary indicating factor. While looking at an
increase of biomass or gas exchange over time can give a gross estimate of primary
productivity, these estimates may not give accurate insights into the efficiency of algal
photosystems. The advances seen in chlorophyll fluorescence technology in the past
decade have drastically changed the ability of eco-physiologists to discern photosystem II
(PSII) kinematics in the laboratory and in situ.
Pulse amplitude modulated fluorescence technology is based on exposing
photosynthetic tissue to a 655nm light source that is rapidly switched on and off at a rate
of 20 kHz. A photodiode then detects only the fluorescence at wavelengths greater than
700 nm to determine the fluorescent output of the sample (Walz manual 1998, Maxwell
and Johnson 2000). The ability to collect a non-invasive, real time fluorescence reading
from samples in situ has expanded the scope of the questions that can be addressed
concerning plant physiology.
The basics of chlorophyll fluorescence are relatively uncomplicated. Energy that
enters the photosynthetic apparatus of a given plant can follow three different pathways
(Figure 2). These pathways control the photochemistry of the system or the actual
conversion of light energy to glucose, and the dissipation of excess energy before it can
cause irreparable damage to the photosystem. Energy entering the first such pathway is
used to drive photosynthesis (production of ATP and NADPH) and is known as

18
photochemical quenching (qP). Energy that enters the second such pathway is dissipated
as heat; this pathway is linked to photo-protection, the dissipation of excess energy before
it can cause permanent damage to PSII (NPQ). The third option is for the energy to be
re-emitted from the PSII antennae complex at a higher wavelength (fluorescence).
Fluorescence is thought to be responsible for the dissipation of approximately
5% of excess the energy present in a given PSII antennae complex. The three energy
pathways of PSII are competing circuits, if the amount of energy entering one increases,
it will in turn decrease the amount entering the other two (Maxwell and Johnson 2000,
Krouse and Weis 1991). By looking at the relative amounts of energy in each pathway, it
is possible to look at the overall efficiency of PSII.
The performance of PSII can be determined from data collected from the changes
in fluorescent levels between times of no, low and high light (Figure 3). PAM
fluorescence can be used with the plants either in a dark acclimated or light acclimated
state. In dark acclimated conditions, the vegetative tissue is kept in complete darkness
for a varying length of time; this darkness ensures that all of the photo-receptors are in a
non-oxidized or open state. A small measuring light (0.15µ mol photon per m2) is
applied to the sample to determine the background fluorescence level (Fo). A pulse of
light with intensity high enough to oxidize (close) all available photo-receptors is then
applied to obtain the maximum fluorescent yield (Fm) for the sample. The difference
between the values of Fm, and Fo is known as the variable fluorescence or Fv. After the
saturating light pulse, a photosynthesis driving light is applied to the sample for a
prescribed length of time. A second saturating pulse is then switched on and the samples

19
maximum fluorescence in the light (Fm’) is measured. Immediately before the second
saturation pulse is applied a measure of the samples residual fluorescence (Ft) is taken.
The comparison of these values allows one to investigate the efficiency of the
photosystem.
The effective quantum yield of energy conversion in a given sample is determined
from the following equation Yield = (Fm’-Ft)/Fm’ (Maxwell and Johnson 2000, Waltz
Manual 1998). This value can be used to interpret how much energy is being consumed
in photochemistry. The potential quantum yield of PSII (ΦPSII) can be determined by
dividing Fv/Fm = (Fm-Fo)/Fm = PSII/qP. This measure can be used to determine the
possible yield for a sample if all of its reaction centers are “open” (Maxwell and Johnson
2000). Photochemical quenching (qP) is an indication of how many reaction centers are
open, or able to accept solar energy. Calculated as (Fm’-Ft)/ (Fm’-Fo’), qP allows one to
look at the actual processes affecting yield (Maxwell and Johnson 2000). Non-
photochemical quenching or NPQ, measures the amount in heat dissipation in dark
acclimated samples.
Fm cannot be measured without the presence of non-photochemical quenching
(Maxwell and Johnson 2000). Due to this fact if an accurate measure of Fm is desired,
samples must be dark acclimated prior to the initial reading, and a relative measure taken
from this dark adapted state. Dark acclimation ensures that all available photo-centers
are in the “open” state, meaning that they are capable of accepting photo-chemical
energy. qN can also affect the level of Fo in some situations, which in turn leads to a
reduction in the yield of Fo`. When qN reaches a point above 0.4, Fo can be quenched,

20
reducing the yield of Fo`. This quenching of Fo` can be addressed only by the use of a far
red light pulse to enhance Qa-reoxidation. This light source however, is not available in
the diving PAM. The diving PAM does however give a second measure of non-
photochemical quenching (NPQ) that is not as sensitive to Fo` quenching. Where qN is
affected by this quenching at 0.4, NPQ is affected by quenching at 4.0 (Maxwell and
Johnson 2000). Due to this increased tolerance, the value of NPQ has become the
measure of choice for heat dissipation analysis.
Field studies using PAM fluorometery have demonstrated the usefulness of this
technology in measuring the parameters surrounding marine primary productivity. Much
of the present work has focused on the effects of increased irradiance on PSII (Hader et
al. 1996. Hader et al. 1999). This work has indicated that the diving PAM fluorometry
system is able to measure in situ changes in quantum yield and qN in PSII as well
recovery time due to photo-inhibition. A correlation between increased photokinetics and
irradiance levels was also shown in seagrasses (Ralph et al. 1998), sponges (Beer and Ilan
1998a) and corals (Beer et al. 1998b). This work has set the base for continued work
using PAM technology to look at PSII kinematics as a function of various other
environmental fluctuations (e.g. water flow and salinity).
The development of chlorophyll fluorescence technology along with advances in
acoustic doppler velocimeters has made research questions that were not measurable a
short time ago valid research avenues. The ability to take detailed in situ measurements
of hydrodynamic regime and tie them in with direct measures of photosystem kinematics
may allow eco-physiologists, physical biologists, and oceanographers to produce more

21
accurate models of aquatic productivity. This thesis investigates the hypothesis that
increased water flow will increase PSII kinematics in Caulerpa spp. This hypothesis is
based on the fact that in higher water flow; the boundary layer is reduced, allowing for a
higher rate of nutrient transport to the algal thallus, or a higher rate of O2 removal from
the surface. This study is the first to use a direct measure photosystem activity to look at
the effects of flowing water on aquatic vegetation.

22
Materials and Methods
To determine the effects of water flow on the photoproduction of the genus
Caulerpa. I preformed three sets of experiments. These experiments were conducted
with specific goals in mind. The first set of experiments was conducted to determine
whether photosystem efficiency was dependent on changes in water velocity in a
laboratory flume. The second set of experiments was conducted to determine the effects
of water flow on photosynthesis in a natural setting, and the third set of experiments was
conducted in order to determine the effects of water flow on photosynthesis over varying
lengths of time.
Measurements on several species of macroalgae within the genus Caulerpa
from the Florida Keys and Tampa Bay were used to characterize the effects of
hydrodynamic regime on photosystem II (PSII) kinematics. Laboratory measurements
were collected in order to determine if photosystem kinematics are a function of
hydrodynamic regime and field measurements were collected in order to elucidate the
role of hydrodynamics in algal productivity in situ. All photosynthetic parameters
analyzed in this study were measured using rapid light curves produced by a Diving PAM
Fluorometer. PSII kinematics measurements were taken approximately half way up the
frond from the rhizome, and in no case was new tissue (complete lack of pigment) used
for measurement of photosynthetic activity. Unlike past studies on productivity in
flowing waters, the development of the Diving PAM Fluorometer allows for non-

23
invasive, direct measurements of PSII kinematics. In all experiments, values of
maximum electron transport rate (Pmax), the initial slope of the photosynthetic curve (α)
and saturation irradiance (Ik) were analyzed. In several experiments photochemical
quenching (qP) and non-photochemical quenching (NPQ) were also analyzed.
In all of the experiments conducted, rapid light curves were collected and analyzed
with two different non-linear models using the sigma plot software suite. The Platt
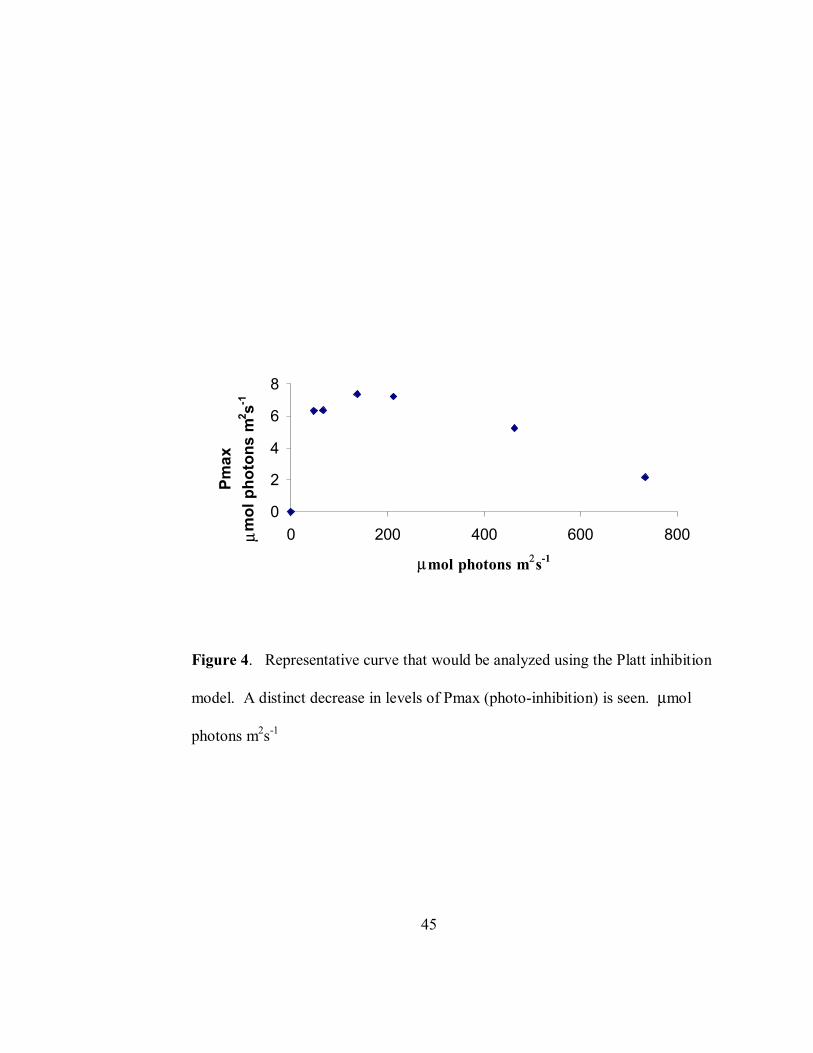
inhibition model, (f=px(1-exp((-axx)/p))x(exp(-bxx/p)) (Henely 1993) was used when light
curves showed a marked inhibition of photosystem structure (Figure 4) and the
hyperbolic tangent model (f=Pmax*tanh((alphaxx)/Pmax)+Rd) (Henely 1993) was used
when there was no inhibition in the light curve (Figure 5). Only curves where the model
fit with a r2 value of 0.90 or better were used in the data analysis.
The recent advances in technology used to measure hydrodynamic regimes have also
drastically changed the scope of the questions that can be raised concerning the
interactions of biological processes and water flow. In all flume experiments, the water
velocity was measured by taking a velocity profile (measures of velocity were collected
at 1.0 cm intervals with an ADV) through the water column and calculating a depth
average velocity. Field measurements of velocity were bulk flow measures taken at the
approximate midpoint of the water column. Care was taken when near a surface, to
ensure that the ADV signal was not being intercepted by the substrate (Finelli 1999).

24
Caulerpa
Caulerpa racemosa, C. sertularioides and C. mexicana were chosen for this study
due to their abundance in Florida’s coastal eco-systems. The unifying characteristics
Caulerpa spp. is a coenocytic thallus, a rhizomatous mass used to anchor the algae to the
substrate and trabeculae supporting the exterior cell walls. Caulerpa spp. often grow in
an intertwined matt of rhizomes, acting to stabilize unconsolidated sediments and
allowing the settlement of climax species such as seagrasses which require solid
sediments.
Caulerpa sertularioides (Figure 6) is characterized by an erect featherlike frond
up to 20.0 cm in length and 1.0-2.0 cm in width. The branchlets or pinnules are
oppositely branched and approximately 3.0-11.0 cm long and pointed at the tip. The
rhizome is up to 2.5 mm in diameter and can grow up to 2.0 m in length. C.
sertularioides is most often found in shallow waters on sandy substrates. C.
sertularioides is found from the shallow subtidal down to a depth of 10.0m. The range of
C. sertularioides is from Florida in the North throughout the Southern Caribbean (Littler
and Littler 2000).
Caulerpa racemosa (Figure 7) is characterized by a creeping or erect frond that is
1.0-5.0 cm tall. The pinnules, of C. racemosa are highly diversified and may include
rounded, mushroom capped, or oval shaped apices. The stolon of C. racemosa is thick
(2.0-3.0 mm) and creeping with a large number of branched rhizoids. C. racemosa is
often found attached to rocks in areas of considerable surf as well as sandy sediments that

25
experience little to no water flow, such as in thick seagrass beds. C. racemosa is found
from the intertidal zone to a depth of 50.0 m. The range of C. racemosa is from Florida
in the North throughout the Southern Caribbean (Littler and Littler 2000).
Caulerpa mexicana (Figure 8) is characterized by an erect frond that can range
from 2.0 cm in height in high-energy areas to 25.0 cm in height in low-energy
environments. Pinnules are opposite of each other and resemble the fronds of a fern. The
stolon of C. mexicana can be relatively thin ranging from 0.6 mm to 1.5 mm in diameter.
The rhizoids are thin and numerous, allowing the algae to anchor in very soft sediment.
The depth range of C. mexicana is reported as the shallow subtidal through a depth of
15.0 m. The species range of C. mexicana is the same as the other two species
mentioned, with a northern limit of Florida throughout the Southern Caribbean (Littler
and Littler 2000).
Measurement of PSII kinematics
Before the start of each experiment, samples were dark acclimated in order to
normalize the light history of individual plants. This dark acclimation involved securing
the sample inside the magnetic leaf clip of the Diving PAM Fluorometer for a period of
ten minutes. The acclimation period ensured that all of the reaction centers within PSII
were oxidized, or open for the capture of photochemical energy. The sample clips
required by the Diving PAM Fluorometer to dark acclimate samples present an
immediate problem when studying the effects of hydrodynamic regime on photosystem

26
function. Samples must be held in the clips for at least 11.5 minutes (10 minutes for dark
acclimation and at least 1.5 minutes for a rapid light curve) during which time the
boundary layer around the thallus is increased due to the added size of the leaf clip. This
increased boundary layer may act to slow the acquisition of nutrients by the algae. To
ensure that this increased boundary layer around the plants due to the presence of the
magnetic leaf clip was not a source of nutrient (e.g. C, N, P) limitation a small flume
experiment was conducted to determine if there was a difference in Pmax when samples
were acclimated using the clips vs. when they were acclimated in a dark room. Samples
of Caulerpa racemosa were collected from the South East jetty of the Sunshine Skyway
Bridge at the mouth of Tampa Bay (Figure 9). All samples were transported in aerated
seawater back to the lab within one hour, cleaned of all macro-epiphytes and kept for 24
hours in aerated seawater. The samples were then placed into a 110L racetrack flume
filled with artificial seawater with a salinity of 35ppt (Figure 10). Samples were held in
place using zip ties attached to a false floor in the flume and allowed to acclimate to the
hydrodynamic regime for 30 minutes before measurements were taken. A light bank of
10,000 K Coralife© full spectrum grow lights suspended above the flume provided
illumination. Samples were dark acclimated using either the magnetic leaf clip (Waltz,
Germany) under full artificial illumination or with the room in complete darkness. Water
flow in the flume was controlled using a small trolling motor and kept at approximately
15 cm s-1. After dark acclimation, rapid light curves using light levels of 0, 8, 12, 22, 30,
48, 66, 104, 155 µmol quanta m-2 s-1 were conducted. To determine if the magnetic leaf

27
clip was negatively affecting Pmax between the two treatments, an ANOVA (SYSTAT)
was run after assessing the normality of the data (SYSTAT).
An initial, small scale experiment was conducted in order to determine whether or
not a diving pam fluorometer could be used to measure short-term changes in
photosystem kinematics due to changes in water velocity. A series of small flume
experiments were conducted at the Keys Marine Lab (KML), on Long Key Florida
(Figure 11). Samples of Caulerpa mexicana were collected from the Thalassia
testudinum bed behind KML over five days of December of 2002. Samples were
transferred within five minutes to a re-circulating seawater table and stored for 24 hours
under natural light conditions before experiments were conducted. Samples were
cleaned of all macro-epiphytes and placed into a 110L racetrack flume filled with un-
filtered seawater (Figure 10), where the water velocity was controlled using a small
trolling motor. High velocity trials were run at 25.0-30.0 cm s-1, and low velocity trials
were run at 5.0-10.0cm s-1. Water velocity profiles were measured using a Sontek
Acoustic Doppler Velocimeter (ADV) at a sample rate of (25 hz) and a depth average
velocity was calculated. The samples were held in place using zip ties attached to a false
floor in the flume. The plants were allowed to acclimate to the flow conditions for one
hour before measurements were taken. A bank of overhead fluorescent lights placed
approximately 30.0 cm above the flume provided irradiance. Samples of C. mexicana
were placed in a magnetic leaf clip (Waltz, Germany) so that no pinnules were
overlapping and dark acclimated for 10 minutes. Rapid light curves using a Diving PAM
Fluorometer were taken using light levels of 0, 48, 66, 99, 138, 212, 311, 463, 734 µmol

28
quanta m-2 s-1. Light curves were analyzed for maximum electron transport rate (Pmax),
saturation irradiance (Ik) and the initial ascending slope of the curve (α) by fitting the data
to either the hyperbolic tangent model or the Platt inhibition model (Henely 1993). Data
were tested for normality using SYSTAT 10 statistical packages and values of Ik, were
normalized to the square root of the data (SIGMA STAT). An ANOVA (SYSTAT) was
used to assess the affect of hydrodynamic regime on Pmax, Ik and α.
Field Experiments
Unlike laboratory experiments that use constant uni-directional flow, organisms
living in natural conditions experience constant changes in the magnitude and direction of
water flow due to tidal changes and wind driven waves (Hearn et al. 2001, Wing and
Patterson 1993). To assess the role of hydrodynamics in algal photo-physiology in situ a
series of field measurements were taken on samples of Caulerpa sertularioides growing
on a carbonate shoal approximately 2.0 km North-West of the Long Key Channel Bridge.
Rapid light curves were measured on samples of Caulerpa sertularioides growing on Old
Sweat Bank, on the Florida Bay side of Long Key, Florida (Figure 11) between June 6,
2002 and June 9, 2002. Much of the water from Florida Bay, including water originating
from the Everglades empties into the Atlantic Ocean through the Long Key Channel
(Wang 1998). Water velocities on the shoal range from near 0.0 cm s-1 at slack tides to
greater than 1.0 m s-1 during flood tides. Old Sweat Bank is characterized as a carbonate
shoal composed mainly of Halimeda spp. segments and crushed coral. Vegetative

29
growth on the shoal is comprised of Thallassia testudinum and Syringonia filiforme,
extensive Halimeda spp. and Penicillus spp. beds, and intermittent patches of Caulerpa
sertularioides. Caulerpa patches were approximately 1.0 m in diameter and were located
along the midline of the shoal. Large patches of Porites porites are scattered over the
shoal along with various species of sponges. The bank is surrounded on all sides by
dense T. testudinum beds, and cut into three sections by two tidal channels (Figure 11).
Water depth over the shoal ranges from approximately 1.0 m at flood tide to <10cm at
neap tide. All light curves were run between 7:30 am and 8:00 pm to ensure that a
variety of water flow velocities (>1.0 ms-1 <0.05 cms-1) were encountered at all times of
photosynthetic activity for the algae.
Samples of Caulerpa sertularioides were placed into the magnetic leaf clip in
such a manner that no pinnules overlapped and dark acclimated in situ for ten minutes
(Waltz, Germany). Rapid light curves were then run on all samples using a diving pam
fluorometer at light levels of 0, 48, 66, 99, 138, 212, 311, 463, 734 µmol quanta m-2 s-1.
Light curves were analyzed for maximum electron transport rate (Pmax), saturation
irradiance (Ik) and the initial ascending slope of the curve (α) by fitting the data to either
the hyperbolic tangent model or the Platt inhibition model (Henely 1993). Ambient light
levels were measured using the external light sensor on the Diving PAM Fluorometer.
Light levels at the algal thallus ranged from 40-1167 µmol quanta m-2 s-1. The 2π
quantum sensor was mounted onto the top of fluorometer, which was the approximate
height of the algae off the substrate. Simultaneous acoustic doppler velocimeter (ADV,
Nortek) readings were taken adjacent to the algae. To measure bulk flow over the shoal

30
at mid water depth, the ADV sensor head was mounted perpendicular to the substrate
when water height above the bottom was >15cm (Figure 12). When water height was
<15cm the sensor was mounted parallel to the substrate (Figure 12). ADV samples were
taken at 25hertz with burst intervals of 5 minutes. Using the internal clocks features of
the fluorometer and the ADV, individual light curves were matched by time to the
corresponding bulk flow measurement over the shoal. Water velocities over the shoal
were separated into three discreet categories, <20cm s-1, 20-30cm s-1, and >30cm s-1. To
determine if photosystem kinematics were dependent on hydrodynamic regime a two way
ANOVA (SYSTAT) was used to test for differences in Pmax, Ik, and α, with time of day
as a covariate. Data was tested for normality (SYSTAT) and subsequently normalized
(SIGMA STAT) to the square root of the data. All outliers were removed according to
Grubbs outliers test (Rolph and Sokal 1981) before analysis was preformed.
Time in flow
In an attempt to isolate the effects of hydrodynamics on the photosystem
kinematics, and to determine the effects of “time in flow” on the PSII kinematics of C.
racemosa. a series of small flume experiments was conducted in a controlled laboratory
setting. In this manner the amount of light, nutrient levels and salinity can be controlled
with hydrodynamic regime as the only fluctuating component.
Samples of Caulerpa racemosa were collected from the South East jetty of the
Sunshine Skyway Bridge at the mouth of Tampa Bay (Figure 9) between July 2, 2002

31
and August 5, 2002. Samples were collected just after sunrise and were returned to the
lab within one hour. Samples were cleaned of macro-epiphytes and kept in aerated
seawater for 24 hrs before experiments were conducted. Only samples that had not been
damaged in the collection process were used for experiments. Three individual plants
were then placed into each of two identical 110L racetrack flume (Figure 10) filled with
artificial seawater (35 ppt). The sea water was fertilized with 0.0161g l-1 PO4, 0.031 g l-1
NH4, and 0.005 g l-1 Nitrate. These nutrient levels correspond to a ten year average
during the summer months at the Sunshine Skyway Bridge
(http://www.floridamarine.org, 2003). Water velocity within the flumes was controlled
by a small trolling motor. Velocity profiles within the flume were taken using an ADV
(SONTEK) and a depth average velocity was calculated for use as a bulk flow
measurement. Illumination was provided by an overhead bank of 10,000 K Coralife©
grow lights suspended over the flumes. Samples were allowed to acclimate to the
hydrodynamic regime for a period of one hour before Rapid Light Curves were collected
using light levels of 0, 8, 12, 22, 30, 48, 66, 104, 155 µmol quanta m-2 s-1. Rapid light
curves were taken every hour for 7 hours. Light curves were analyzed for maximum
electron transport rate (Pmax), saturation irradiance (Ik) and the initial ascending slope of
the curve (α) by fitting the data to either the hyperbolic tangent model or the Platt
inhibition model (Henely 1993). Photochemical quenching (qP) and non-photochemical
quenching (NPQ) values were obtained for each sample by selecting the set of data
within each curve nearest the saturation irradiance for the sample. To assess the role of
hydrodynamic regime on the photosystem kinematics of C. racemosa a two way repeated

32
measures ANOVA (SIGMA STAT) was preformed with water flow as one factor and the
amount of time in the flow as the second factor. The measurements from each of the
three plants in each flume were averaged to form one set of data per trial. The data was
then assessed for normality (SYSTAT) and transformed (SIGMA STAT) to the square
root of the data for Pmax, Ik, and NPQ. Data was transformed to the ln of the data for α,
and qP. All outliers were removed using Grubbs outliers test (Rohlf and Sokal 1981).
The ratio of Pmax in high flow to Pmax in low flow was plotted for each of the
seven time periods (FIGURE 13). A distinct drop in the ratios was seen after two hours
of immersion. The data for measurements three through seven was then assessed for
normality (SYSTAT) and transformed (SIGMA STAT) to the square root of the data for
Pmax, Ik, and NPQ. Data was transformed to the ln of the data for α, and qP. All outliers
were removed using Grubbs outliers test (Rohlf and Sokal 1981). A two way repeated
measures ANOVA was then used to assess the role of hydrodynamics after two hours of
immersion.

33
Results
Does the magnetic sample holder affect values of Pmax?
The differences in values of Pmax between trials using a magnetic leaf clip to
dark acclimate a sample and trials using a dark room to dark acclimate samples were not
significantly different (P=0.790, F=0.073, Figure 14). Samples acclimate with clips had a
mean Pmax of 8.6 (SE = 1.4, n=9) and samples acclimated in a darkened room had a
mean Pmax of 8.9 (SE=3.1, n=9).
Laboratory measurements of PSII kinematics on C. mexicana after a short exposure
to hydrodynamic regime.
The effect of water velocity on values of Pmax from C. mexicana was significant
(P<0.05, F=6.811, Figure 15). Samples run at high (25-30cm s-1) and low (5-10 cm s-1)
velocities had mean values of Pmax of 26.5 (SE=7.0, n=4) and 52.3 (SE=7.0, n=4)
respectively. Values of α from C. mexicana in the two different hydrodynamic regimes
were not significantly different. The effect of water velocity on saturation irradiance was
very close to being significant, (P=0.055, F=5.66). The mean of Ik in high flow was 16.2
(SE=1.7 n=4) and the mean of Ik in low flow was 22.0 (SE=1.7, n=4).

34
In situ field measurements of PSII kinematics on C. sertularioides
Water velocity had a significant affect on the values of Pmax and Ik in Caulerpa
sertularioides measured in situ (P<0.001, F=9.387, Figure 16, Table 1 and P<0.05,
F=3.754, Figure 17, Table 1 respectively). Water flow did not affect values of α
(P=0.570, F=5.67) Table 1). There was no significant interaction of time on the values of
Pmax, α or Ik (Table 1).
Laboratory measurements of PSII kinematics in C. racemosa after long term
exposure to hydrodynamic regime.
Values of Pmax (P=0.067, F=3.865), α (P=0.100, F=3.044,), Ik (P=0.893,
F=0.0188), qP (P=0.198, F=1.800) and NPQ (P=0.490, F=0.497) for C. racemosa run in
laboratory flumes at different water flow velocities over a period of seven hours were not
significantly different (Table 2). There was no significant difference in Values of Pmax
(P=0.343, F=1.143), α (P=0.104, F=1.818), Ik (P= 0.674, F=0.669) and qP (P=.135,
F=1.677) due to the time spent in the flow (Table 2). There was a significant effect of
time in flow for values of NPQ (P<.005, F=4.825, Figure 18, Table 2, Table 3). The
interaction of time spent in water flow and water velocity for Pmax (P=0.266, F=1.297),
α (P=0.409, F=1.032), Ik (P=0.078, F=1.972), qP (P=0.164, F=1.570) and NPQ (P=0.286,
F=1.259) was not significant (Table 2).
The ratio of Pmax in fast flow to Pmax in slow flow was calculated to determine
if this value changed over time (Figure 13). The ratio was found to be relatively constant
for the first two hours, then the ratio dropped, indicating that there may be a delay in

35
response to changes in water flow. If the data is analyzed for the time after two hours
there is a significant difference between Pmax in fast flow and Pmax in slow flow. A
second repeated measures ANOVA was run for measurements three through seven.
Water velocity significantly affected values of Pmax values at times three through seven
(P<0.05, F=4.816, Figure 19, Table 4). The mean value of Pmax in low flow velocity
was 3.5 (SE=0.201, n=45), the mean value of Pmax in high flow was 4.7 (SE=0.278,
n=45). Water velocity did not affect the values of α, Ik, qP and NPQ (P=0.098, F=3.096,
P=0.971, F=0.00138, P=0.098, F=3.096 and P=0.321, F=1.039 respectively Table 4). The
amount of time spent in the hydrodynamic regime did not affect the values of Pmax, α,
Ik, and qP(P=0.264, F=1.342, P=0.167, F=1.65, P=0.665, F=0.618 and P=0.167, F=1.675
respectively, Table 3). The amount of time in the hydrodynamic regime did affect values
of NPQ significantly (P=.001, F=4.197, Figure 20, Table 4, Table 5). The interaction of
water velocity and time spent in flow did not affect the values of Pmax, α, Ik , qP and
NPQ (P=0.273, F=1.318, P=0.458, F=0.920, P=0.139, F=1.810, P=0.403, F=1.023 and
P=0.374, F=0.826 respectively, Table 3).

36
Discussion The objectives of this thesis were to determine if hydrodynamic regime affects
photosystem II kinematics in three species of the green algae Caulerpa, and to determine
the extent of the dependence of PSII kinematics on hydrodynamic regime. Experiments
also showed that the effect of hydrodynamics on members of the genus Caulerpa may be
species, or location specific.
The results of these experiments indicate that photosystem II kinematics of
Caulerpa sertularioides, C. mexicana, and C. racemosa are influenced by hydrodynamic
regime in lab experiments and in situ. Lab experiments on C. racemosa (Figure 19)
collected from Tampa Bay, support the findings of Wheeler (1980), Gerard (1982), Koehl
and Alberte (1988), Carpenter et al. (1991) and Williams and Carpenter (1998) that
increased water flow positively correlates with Pmax (figure b) and Ik (figure c) in marine
algae. These results support our hypothesis that increased water flow would positively
influence PSII kinematics. A possible explanation is that in Tampa Bay, C. racemosa is
physically limited. As water velocity was artificially increased more nutrients were
delivered to the plant surface, allowing for an increase in the electron transport rate.
Field measurements of C. sertularioides from the Florida Keys and lab
measurements of C. mexicana did not support our hypothesis. Results showed a drastic
decrease in Pmax as a function of water velocity (Figure 13, Figure 15). These findings
are supported by several studies on the effects of water flow on marine and freshwater

37
algae. Gerard and Mann (1979) noted that the growth of Laminaria longicrursis in
exposed sites was significantly slower than the growth of L. longicrursis in protected
sites. More recently, Borchardt (1994) demonstrated that in freshwater plants which are N
and P limited, flows greater than 30 cm s-1 were physiologically costly. Several
mechanisms may be responsible for these results, including nutrient limitation due to the
high water velocity, light flecking, and self shading.
Nutrient limitation is often thought to occur only during periods of low flow,
when nutrient delivery is impeded by the thickness of the diffusive boundary layer.
However, nutrient limitation can occur during periods of relatively high water flow if
there are competing mechanisms within an organism that are triggered by the increase in
water velocity. Gerard and Mann (1979) concluded that in high flowing waters N was
more limiting than in slow flows in L. longicrursis because of specialized structures the
algae needed to produce and maintain in order to survive in high velocity environments.
If the plants need N to build or maintain these structures, then less is available for the
cells photosynthetic pathways. Borchardt (1994) proposed that the actual nutrient
requirements of nutrient limited Spirogyra fluviatilis changed with changes in
hydrodynamic regime; the plants needed more of a limiting nutrient when flow was
increased. This conclusion supports the conclusions of Gerard and Mann (1979), that
while nutrient delivery is increased in higher flows, the actual cellular or organismal
quota is also increased. Carbon limitation may actually increase with flowing waters
depending on the form of C that a plant is capable of using (Borchardt 1994). If a plant
utilizes an extra cellular bi-carbonate converting mechanism to convert bi-carbonate to

38
CO2, increased water flow may actually increase C limitation. These conversion
mechanisms often rely on the algae actively pumping a high gradient of protons (H+) into
the diffusive boundary layer next to the thallus surface. These H+ are needed to convert
bi-carbonate to CO2. If water velocity increased and these H+ were transported away
from the thallus, then higher levels of N and P may be needed to increase H+ transport
across the cell membranes.
The light field experienced by any photosynthetic organism can drastically change
its rate of photosynthesis. Sun flecks are intense burst of irradiance that result from the
movement of canopy structures and surface waves that can account for significant
differences in photosynthetic capacity of marine plants (Dromgoole 1987, Wing and
Patterson 1993, Wing et al. 1993). Due to their focusing effect, surface waves result in
high-frequency light fluctuations (millisecond range ), while the movement of canopy
structures results in slower frequency and longer duration fluctuations (seconds to
minutes ) (Green and Gerard 1990, Wing and Patterson 1993, Sagert and Schubet 2000).
These fluctuations in light levels and intensity are known to be both beneficial and costly
to algae. Under high frequencies of fluctuating light, the photosynthetic rates of many
species have been shown to increase (Dromgoole 1987, Dromgoole 1988, Green and
Gerard 1990). Conversely, other studies have demonstrated a negative correlation
between light flecks and photosynthetic rates (Wellnitz and Rinne 1999). For a short
period of time (milliseconds) focusing of irradiance by surface waves can increase the
amount of light reaching the benthos by as much as five times the surface irradiance
(Sagert and Schubert 2000, Schubert et al. 2001). During periods of high water flow on

39
Old Sweat Bank, small surface waves ran over the shoal with a very high frequency
(Driscoll personal observations). The light regime resulting from this intense focusing of
light may cause photo-inhibition in C. sertularioides due to temporarily damage the
antennae complexes of PSII.
Caulerpa, like several other genera of the siphonous green algae have the ability
to translocate their chloroplasts in times of stress and growth (Dawes and Barilotti 1969,
Drew and Abel 1990, Chisholm and Jaubert 1997). This chloroplast movement was
witnessed in C. sertularioides and C. racemosa during periods of high water flow. This
chloroplast movement may have induced self-shading (of plastids) effects which could
have resulted in an underestimation of PSII kinematics in high flow conditions.
In high velocity flows marine algae have been shown to “clump” together or bend
to reduce the effects of form drag (Koehl and Alberte 1988). This clumping, while
reducing drag can also decrease the photosynthetic activity of a plant by limiting the
amount of light reaching the individual photosynthetic surfaces of the plant. The frond
axis of C. sertularioides in the Florida Keys remained upright at all water velocities,
however, observations showed the pinnules bending with the bulk flow so that they were
almost parallel to the main velocity vector. This bending of the pinnules, while not
quantified, may have caused self-shading during times of high water flow, decreasing
values of Pmax.
Comparison between sites in the Florida Keys and Tampa Bay are made difficult
by the differences in light regime and ambient nutrient concentration. The collection site
in Tampa Bay is often extremely turbid due to terrestrial run-off and storm activity,

40
limiting the amount of light penetrating the water column. Nutrient levels are also high
due to extensive anthropogenic inputs. Conversely, Old Sweat Bank has very little
turbidity in the water column, allowing high levels of irradiance to penetrate the water.
Like most tropical systems the nutrient levels of carbonate shoals in the keys are deplete
(Lapointe and Clark 1992). These factors make comparisons between locations
equivocal, continued experimentation at both sites will be needed to draw conclusions on
a wider scale than species level.
These experiments are the first to use a direct measure of PSII kinematics to look
at the role of hydrodynamics in photo-production. Previous studies have relied on
oxygen evolution as the measure of photosynthetic activity (Raven et. al. 1979, Carpenter
et. al. 1991, Gacia et al. 1996). The measure of oxygen evolution as a function of water
velocity creates a situation that may drastically affect the results of experiments. Just as
Nitrogen, Phosphate, and Carbon are mass transfer limited to photosynthetic surface,
oxygen may be mass transfer limited from the same surface. This limitation would mean
that in low flows a gradient of oxygen would form within the boundary layer with higher
levels near the tissue surface. By increasing water velocity this oxygen would be carried
into the water column as the boundary layer thickness is decreased, erroneously
increasing the measured rates of oxygen evolution. By measuring chlorophyll
fluorescence this artifact is removed since fluorescence originates in the PSII antennae
complex and the measure itself is not affected by associated boundary layers.
This research is the first set of data that looks at direct measures of photo-
production as a function of hydrodynamic regime. Our findings support the hypothesis

41
that hydrodynamic regimes can influence physiological processes in marine algae. The
method of this influence and the degree of the dependence of PSII on water flow
conditions are still unclear, however, the results of these experiments and observations
suggest that the use of PAM Fluorescence could be applied to elucidate the answers to
these questions. Future directions for this project may involve altering the concentrations
of nutrients in a variety of hydrodynamic regimes in an attempt to isolate the limiting
factor, if any, in the photosynthetic pathways of Caulerpa spp. In situ measurements on
several more species of Caulerpa would also be beneficial in determining the role of
hydrodynamics in photosystem function.

42
Figure 1. Diagram of the anti-herbivory metabolite caulerpenyne. This compound is on
of the main factors allowing for the success of Caulerpa spp. In certain cases
caulerpenyne may compose up to 13% of the wet frond weight of C. taxifolia, making the
algae in-edible to any known herbivore.

43
Figure 2. The three main energy pathways for photosystem II. (1) Energy is
shunted off to the electron transport chain, (2) energy is dissipated as heat before it
damages the photosystem and (3) energy is re-emitted from photosystem II at a higher
wavelength (Fluorescence).

44
Figure 3. Skematic diagram of the pulse amplitude modulated fluorescence
measuring principle in both light and dark adapted samples. (Walz 2001).
45
0
2
4
6
8
0 200 400 600 800
µmol photons m2s-1
Pmax
µ
mol
pho
tons
m2 s-1
Figure 4. Representative curve that would be analyzed using the Platt inhibition
model. A distinct decrease in levels of Pmax (photo-inhibition) is seen. µmol
photons m2s-1

46
05
10152025
0 200 400 600 800
µmol photons m2s-1
Pmax
µ
mol
pho
tons
m2 s-1
Figure 5. Representative curve that would be analyzed using the hyperbolic
tangent model. No photo-inhibition is seen in the curve.

47
Figure 6. Drawing of Caulerpa sertularioides from Caribbean Reef Plants (Littler and Littler 2000). The pinnules are clearly much finer than those found in C. mexicana and C. racemosa.
48
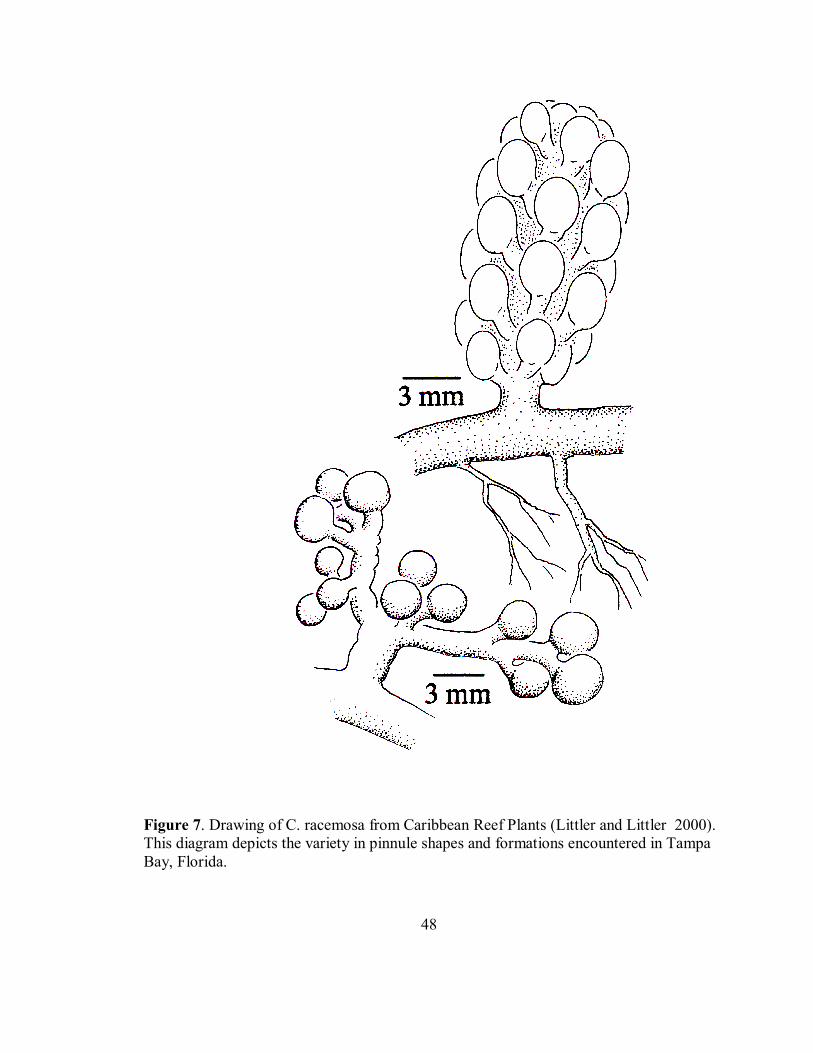
Figure 7. Drawing of C. racemosa from Caribbean Reef Plants (Littler and Littler 2000). This diagram depicts the variety in pinnule shapes and formations encountered in Tampa Bay, Florida.

49
Figure 8. Drawing of Caulerpa mexicana from Caribbean reef plants (Littler and Litter 2000). The pinnules of C. mexicana grown in a very flat broad shape, which is very similar to that of C. taxifolia.

50
Figure 9. Collection site for C. racemosa at the Sunshine Skyway Bridge south causeway. Specimens were collected from a depth of approximately 0.5m and returned to the lab within one hour of collection.

51
Figure 10. 110L racetrack flume. Water velocity within the flume is created by a small trolling motor and augmented using a diode control. Turbulence within the flume is dampened using three sets of flow straighteners.
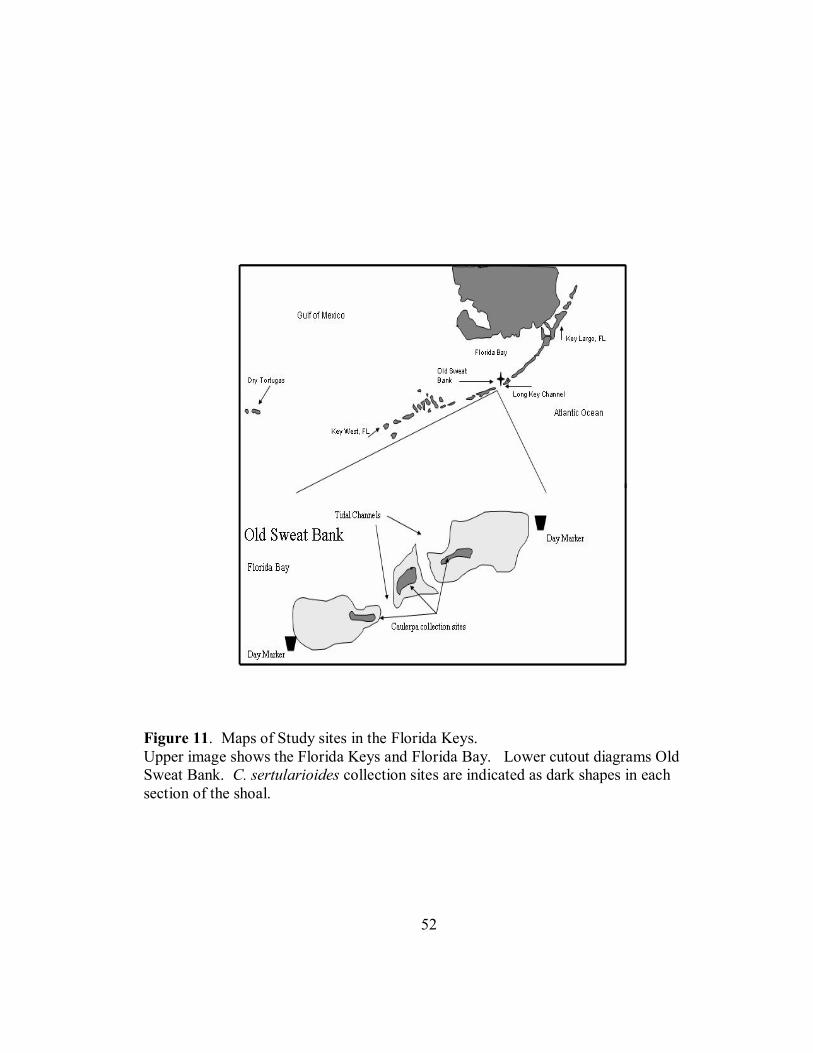
52
Figure 11. Maps of Study sites in the Florida Keys. Upper image shows the Florida Keys and Florida Bay. Lower cutout diagrams Old Sweat Bank. C. sertularioides collection sites are indicated as dark shapes in each section of the shoal.

53
Figure 12. Orientation of the acoustic doppler velocimeters during field measurements. The ADV was attached to a large ring stand device with cable ties and hose clamps, and height was adjusted using several ring stand clamps. The sensor head was placed perpendicular to the substrate when water depth was greater than 15 cm and it was oriented parallel to the substrate when the water depth was less than 15 cm.
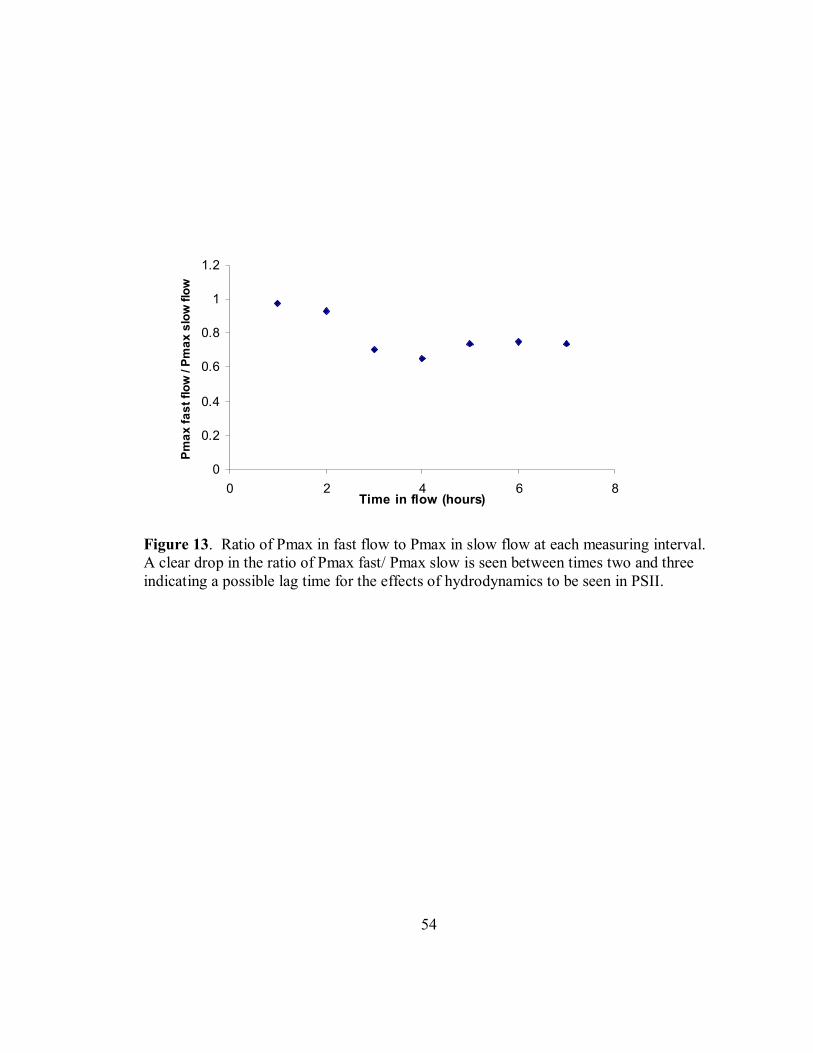
54
0
0.2
0.4
0.6
0.8
1
1.2
0 2 4 6 8Time in flow (hours)
Pm
ax fa
st fl
ow /
Pmax
slo
w fl
ow
Figure 13. Ratio of Pmax in fast flow to Pmax in slow flow at each measuring interval. A clear drop in the ratio of Pmax fast/ Pmax slow is seen between times two and three indicating a possible lag time for the effects of hydrodynamics to be seen in PSII.

55
0
5
10
15
Clip Dark room
Pmax
(µm
ol e
lect
ron
m2
s-1)
Figure 14. Results of dark acclimation trial using magnetic leaf clips under full illumination and a dark room. There was no significant difference (P value = 0.790) between trials utilizing the magnetic leaf clips and a dark room to dark acclimate tissue samples. These results indicate that the samples used were not nutrient limited by the presence of the magnetic leaf clip.

56
0 10
20 30
40 50
60 70
5-10 cm s-120-25 cm s-1
Pmax
(µm
ol e
lect
ron
m2
s-1)
Figure 15. Values of Pmax for C. mexicana run at high and low velocities in a laboratory flume. Water velocity was shown to significantly affect values of Pmax (P value = 0.04). High flow was 20 – 25 cm s -1 and low flow was 5-10 cm s-1 . Values of Pmax were significantly higher in slow hydrodynamic regimes.

57
0 2 4 6 8
10 12 14 16
<20 cm s-1 20-30 cm s-1 >30 cm s-1
Pm
ax
(µm
ol e
lect
ron
m2 s-1
)
Figure 16. In situ measures of Pmax in C. sertularioides. PSII kinematics parameters were measured without removing the plant from its natural environment. Water velocity was shown to significantly affect values of Pmax measured in the field using a Diving PAM Fluorometer (P value <0.001). Low flow was less than 20 cm s -1 , medium flow was 20 – 30 cm s -1 and high flow was above 30 cm s -1 . The highest values of Ik were seen when velocity was less than 20 cm s-1. The lowest values of Pmax were measured when water velocity was greater than 30 cm s-1.
A
B
C

58
0 10 20 30 40 50 60 70
<20 cm s -1 20-30 cm s -1 >30 cm s -1
I k (µ
mol
qua
nta
m-1
s-1)
Figure 17. In situ measurements of Saturation irradiance in C. sertularioides. Saturation irradiances measured on C. sertularioides measured in situ were significantly different as a function of water velocity (P value = 0.018). Low flow was less than 20 cm s -1 , medium flow was 20 – 30 cm s -1 and high flow was above 30 cm s -1 . The highest values of Ik were seen when velocity was less than 20 cm s-1.
A
B
C

59
0 0.020.040.060.080.1
0.120.14
1 2 3 4 5 6 7 Time in flow
NPQ
Figure 18. Laboratory measurements of NPQ in C. racemosa. Samples were collected from Tampa Bay, and PSII kinematics parameters were measured in a laboratory flume every hour for seven consecutive hours. Values of NPQ, measured from hours three through seven were shown to be significantly different as a function the amount of time spent in the water flow (P value = 0.002).
AA A
AA
BB

60
3.4 3.6
3.8 4
4.2 4.4
4.6 4.8
>15 cm s -1<15 cm s -1
Pm
ax
(µm
ol e
lect
ron
m2
s-1)
Figure 19. Laboratory measurements of Pmax in C. racemosa. C. racemosa was collected from the Sunshine Skyway jettie, and exposed to different water flow regimes in a small laboratory flume. After three hours in a hydrodynamic regime, values of Pmax from C. racemosa were shown to be significantly different as a function of water velocity (P value = 0.022). High flow was considered velocities above 15 cm s-1 and low flow was considered velocities lower than 15 cm s-1. Higher values of Pmax were measured in velocities greater than 15 cm s-1.

61
Figure 20. Laboratory measurements of NPQ in C. racemosa. Samples were collected from Tampa Bay, and PSII kinematics parameters were measured in a laboratory flume. Values of NPQ, measured from hours three through seven were shown to be significantly different as a function the amount of time spent in the water flow (P value = 0.002). Hours labeled “A” are significantly different than hours labeled "B".
0 0.02 0.04 0.06
0.08 0.1
0.12 0.14
3 4 5 6 7 Measurement hours
NPQ
A A
BB
B

62
Flow Speed
Low Medium High
P value
Mean
Mean Mean Velocity time
Pmax (µmol electron m2s-1)
12.8
(0.794)
9.4 (0.759)
7.2 (1.127)
<0.001 0.47
α (µmol electron m2s-1)
0.224 (0.017)
0.217 (0.019)
0.257 (0.031)
0.57 0.26
Ik (µmol quanta m2s-1)
56.8
(5.632)
46.4 (5.245)
30.3 (7.534)
<0.05 0.64
Table 1. Values of Pmax, α, and Ik measured on C. sertularioides in situ. Measurements were taken on plants that had not been removed from their natural habitats, and that were exposed to natural water flow and irradiance conditions. Values of Pmax, measured in situ were found to be significantly different as a function of water velocity (P value <0.001). Values of α and Ik were not found to be significantly different in different water flows when measured in situ.
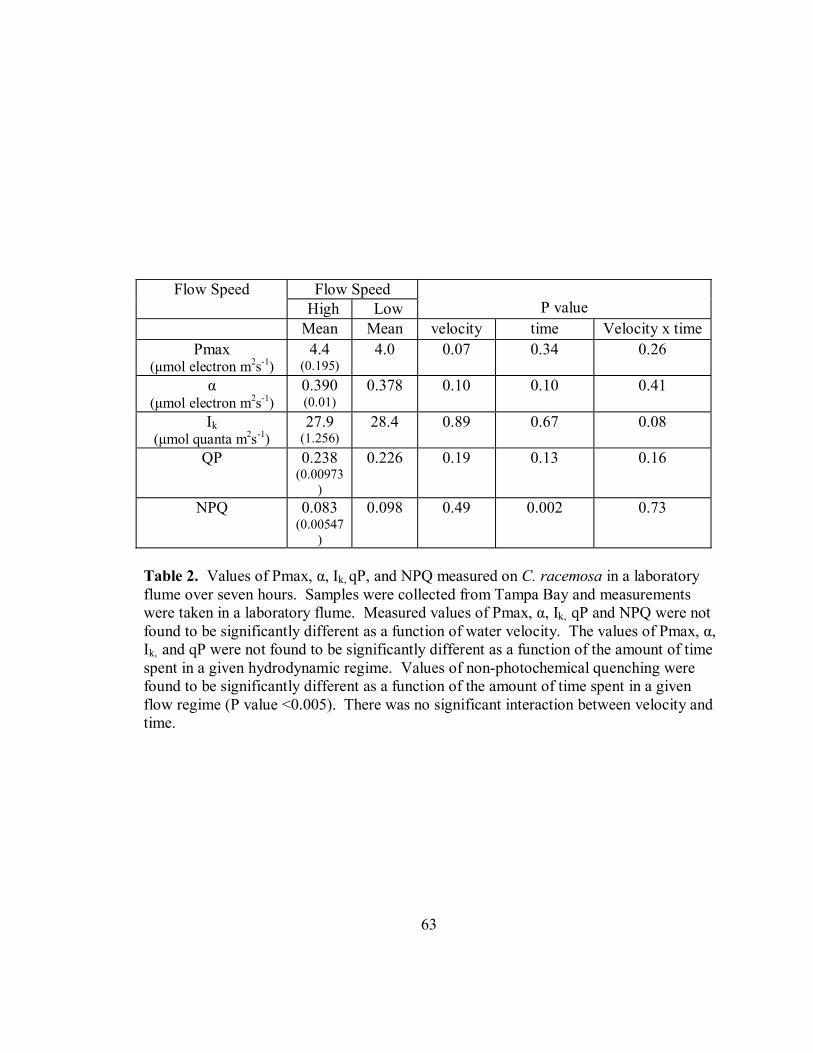
63
Flow Speed Flow Speed High Low
P value
Mean Mean velocity time Velocity x time Pmax
(µmol electron m2s-1) 4.4
(0.195) 4.0 0.07 0.34 0.26
α (µmol electron m2s-1)
0.390 (0.01)
0.378 0.10 0.10 0.41
Ik (µmol quanta m2s-1)
27.9 (1.256)
28.4 0.89 0.67 0.08
QP 0.238 (0.00973
)
0.226 0.19 0.13 0.16
NPQ 0.083 (0.00547
)
0.098 0.49 0.002 0.73
Table 2. Values of Pmax, α, Ik, qP, and NPQ measured on C. racemosa in a laboratory flume over seven hours. Samples were collected from Tampa Bay and measurements were taken in a laboratory flume. Measured values of Pmax, α, Ik, qP and NPQ were not found to be significantly different as a function of water velocity. The values of Pmax, α, Ik, and qP were not found to be significantly different as a function of the amount of time spent in a given hydrodynamic regime. Values of non-photochemical quenching were found to be significantly different as a function of the amount of time spent in a given flow regime (P value <0.005). There was no significant interaction between velocity and time.

64
Time NPQ Sample Size
Mean SE N 1 0.075 0.012 17 2 0.089 0.011 18 3 0.069 0.007 14 4 0.101 0.008 18 5 0.103 0.007 16 6 0.115 0.014 17 7 0.084 0.013 13
Table 3. Values of NPQ at each measurement hour. Measurements were taken on samples in a laboratory flume. NPQ values are significantly different as a function of time immersed in flow. Values of NPQ at measurement intervals of 3 and seven hours were significantly different than at the other five measurement times.

65
Flow Speed Flow Speed High Low
Pvalue
Mean Mean Velocity time Velocity x timePmax
(µmol electron m2s-1) 4.7
(0.278) 3.5
(0.201) <0.05 0.26 0.27
α (µmol electron m2s-1)
0.168 (0.00755)
0.141 (0.00904)
0.98 0.17 0.36
Ik (µmol quanta m2s-1)
27.8 (1.150)
27.7 (1.426)
0.97 0.67 0.14
qP 1.967 (0.0595)
1.772 (0.0617)
0.09 0.17 1.12
NPQ 0.096 (0.00716)
0.090 (0.00586)
0.32 <0.002 0.83
Table 4. Values of Pmax, α, Ik, qP, and NPQ measured on C. racemosa over five hours in a laboratory flume. Samples were collected from Tampa Bay, and measurements were taken while the samples were immersed in a given water flow regime in a laboratory flume. Measurements were taken once every hour for seven hours. Measured values of Pmax for times three through seven were found to be significantly different as a function of water velocity (P value <0.05). There was a higher electron transport rate in samples experiencing a faster hydrodynamic regime. Values of α, Ik, qP and NPQ at times three through seven in fast and slow hydrodynamic regimes were shown to not be significantly different. Measured values of NPQ were shown to be significantly different as a function of the amount of time spent in a given hydrodynamic regime (P value = 0.002). Values of Pmax, α, Ik and qP were not significantly different in at different measuring periods. There was no interaction of time and velocity.
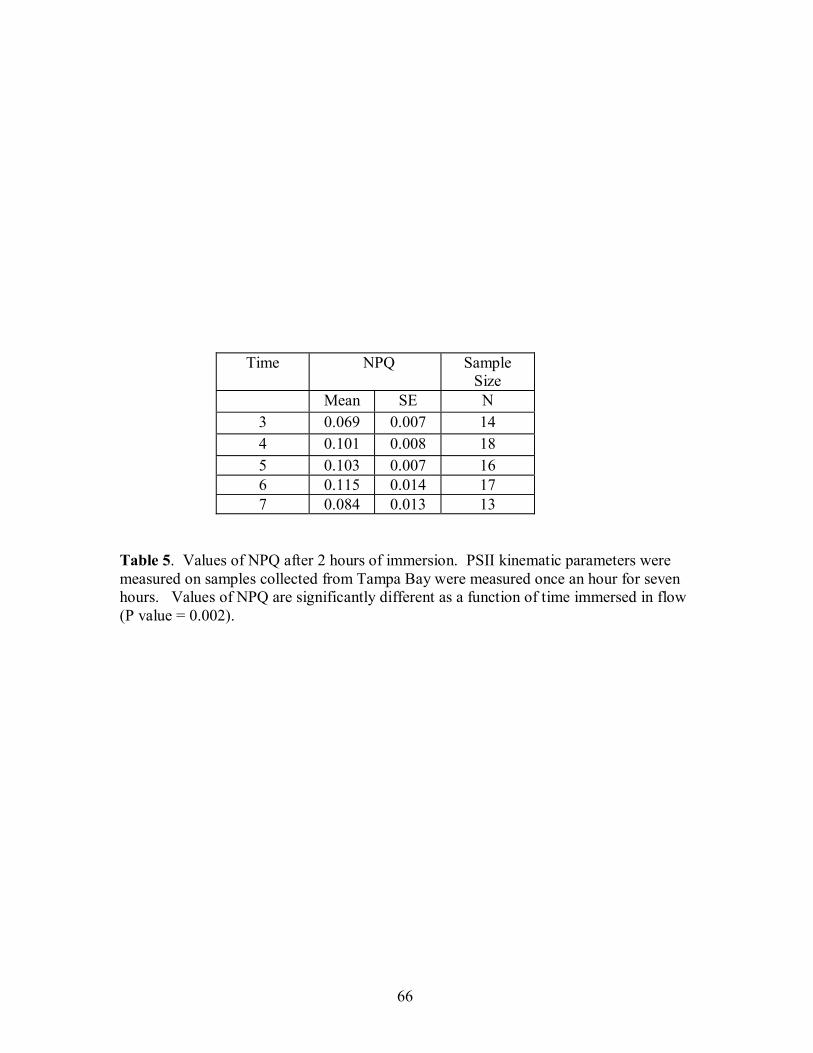
66
Time NPQ Sample Size
Mean SE N 3 0.069 0.007 14 4 0.101 0.008 18 5 0.103 0.007 16 6 0.115 0.014 17 7 0.084 0.013 13
Table 5. Values of NPQ after 2 hours of immersion. PSII kinematic parameters were measured on samples collected from Tampa Bay were measured once an hour for seven hours. Values of NPQ are significantly different as a function of time immersed in flow (P value = 0.002).

67
References Atkinson, M.J., R.W. Bilger., 1992. Effects of water velocity on phosphate uptake in coral reef-flat communities. Limnol. Oceanogr. 37, 273-279. Atkinson, M.J., S.V. Smith., 1983. C:N:P ratios of benthic marine plants. Limnol. Oceanogr. 28, 568-674. Argyrou, M., A. Demetropoulos, M. Hadjichristophorou., 1999. The impact of Caulerpa racemosa on the macrobenthic communities in the coastal waters of Cyprus. Proceedings of the workshop on invasive Caulerpa in the Mediterranean. Herkalion, Crete, Greece. 18-20 March, 1998. UNEP publ., Athens, Greece. 139-158. Arigoni, S., P. Francour, M. Harmelin-Vivien, L. Zaninetti., 2002. Adaptive colouration of Mediterranean labrid fishes to the new habitat provided by the introduced tropical alga Caulerpa taxifolia. J. Fish Biol. 60, 1486-1497. Aussem, A., D. Hill., 1999. Wedding connectionist and algorithmic modelling towards forecasting Caulerpa taxifolia development in the North-Western Mediterranean Sea. Ecol. Mod. 120, 225-236. Barilotti, D.C., 1970. Non-genetic morphological variation in Caulerpa prolifera (Forsskal) Lamouroux. Thesis. University of South Florida. Beach, K.S., C.M. Smith, T.M. Michael, H.W. Shin., 1995. Photosynthesis in reproductive unicells of Ulva fasciata and Enteromorpha flexuosa: Implication for ecological success. Mar. Ecol. Prog. Ser. 125, 229-237. Beach, K.S., and C.M. Smith, 1996a. Ecophysiology of tropical rhodophytes. I. Microscale acclimation in pigmentation. J. Phycol. 32, 701-710. Beach, K.S., C.M. Smith., 1996b. Ecophysiology of tropical Rhodophytes. II. Microscale acclimation in photosynthesis. J. Phycol. 32, 710-718. Beach, K.S., H.B. Borgeas, N.J. Nishimura, C.M. Smith., 1997. In vivo absorbance spectra and the ecophysiology of reef macroalgae. Coral Reefs. 16, 21-28. Benzie, J.A.H., E. Ballment, J.R.M. Chisholm, J.M. Jaubert., 2000. Genetic variation in the green alga Caulerpa taxifolia. Aquat. Bot. 66, 131-139.

68
Beer, S., M. Ilan., 1998a In situ measurements of photosynthetic irradiance responses of two Red Sea sponges growing under dim light conditions. Mar. Biol. 131, 613-617. Beer, S., M. Ilan, A. Eshel, A. Weil, I. Brickner., 1998b. Use of pulse amplitude modulated (PAM) fluorometry for in situ measurements of photosynthesis in two Red Sea flaviid corals. Mar. Biol. 131, 607-612. Bellan-Santini, D., P.M. Arnaud, G. Bellan, M. Verlaque., 1996. The influence of the introduced tropical alga Caulerpa taxifolia, on the biodiversity of the Mediterranean marine biota. J. Mar. Biol. Assoc. U.K. 76, 235-237. Bilger,R.W., M.J. Atkinson., 1992. Anamalous mass transfer of phosphate on coral reef flats. Limnol. Oceanogr. 37, 261-272. Bilger, R.W., M.J. Atkinson., 1995. Effects of nutrient loading on mass-transfer rates to a coral-reef community. Limnol Oceanogr. 40, 279-289. Bjornsater, B.R., P.A. Wheeler., 1990. Effect of nitrogen and phosphorus supply on growth and tissue composition of Ulva fenestrata and Enteromorpha intestinalis (Ulvales: Chlorophyta). J. Phycol. 26, 603-611. Borchardt, M.A. ,1994. Effects of flowing water on nitrogen- and phosphorus-limited photosynthesis and optimum N:P ratios by Spirogyra fluviatilis. J. Phycol. 30, 418-430. Carpenter, R.C., J.M. Hackney, W.H. Adey., 1991. Measurements of primary productivity and nitrogenase activity of coral reef algae in a chamber incorporating oscillatory flow. Limnol. Oceanogr. 36, 40-49. Ceccherelli, G., Campo., 2002. Different effects of Caulerpa racemosa on two co-occuring seagrasses in the Mediterranean. Bot. Mar. 45, 71-76. Ceccherelli, G., F. Cinelli., 1997. Short-term effects of nutrient enrichment of the sediment and interactions between the seagrass Cymodocea nodosa and the introduced green alga Caulerpa taxifolia in a Mediterranean bay. J. Exp. Mar. Biol. Ecol. 217, 165-177. Ceccherelli, G. F. Cinelli., 1999. A pilot study of nutrient enriched sediments in a Cymodocea nodosa bed invaded by the introduced alga Caulerpa taxifolia. Bot. Mar. 42, 409-417. Ceccherelli, G., L. Piazzi., 2001. Dispersal of Caulerpa racemosa fragments in the Mediterranean: lack of detachment time effect on Establishment. Bot. Mar. 44, 209-213.

69
Ceccherelli, G., L. Piazzi, F. Cinelli., 2000. Response of the non-indigenous Caulerpa racemosa (Forsskäl) J. Agardh to the native seagrass Posidonia oceanica (L.) Delile: effect of density of shoots and orientation of edges of meadows. J. Exp. Mar. Biol. Ecol. 243, 227-240. Ceccherelli, G., L. Piazzi, F. Cinelli., 2000. Response of the non-indigenous Caulerpa racemosa (Forsskål) J. Agardh to the native seagrass Posidonia oceanica (L.) Delile: effect of density of shoots and orientation of edges of meadows. J. Exp. Mar. Biol. Ecol. 243, 227-240. Ceccherelli, G., N. Sechi., 2002. Nutrient availability in the sediment and the reciprocal effects between the native seagrass Cymodocea nodosa and the introduced rhizophytic alga Caulerpa taxifolia. Hydrobiologica. 474, 57-66. Chapman, A.R.O., 1974. The ecology of macroscopic marine algae. An. Rev.Ecol. Syst. 5, 65-80. Chisholm, R.M., F. E. Fernex, D. Mathieu, J.M. Jaubert., 1997. Watewater discharge, seagrass decline and algal proliferation on the Côte d’Azur. Mar. Poll. Bull. 34, 78-84. Cloern, J.E., 1999. The relative importance of light and nutrient limitation of phytoplankton growth: a simple index of coastal ecosystem sensitivity to nutrient enrichment. Aquat. Ecol. vol. 33. pp. 3-16. Collado-Vides, L., D.Robledo., 1999. Morphology and photosynthesis of Caulerpa (Chlorophyta) in relation to growth form. J. Phycol. 35, 325-330. Collado-Vides, L., 2002. Morpholical plasticity of Caulerpa prolifera (Caulerpales-Chlorophyta) in relation to growth form in a coral reef lagoon. Bot. Mar. 45, 123-129. Collado-Vides, L., D. Robledo., 1999. Morphology and photosynthesis of Caulerpa (Chlorophyta) in relation to growth form. J. Phycol. 35, 325-330. Cornelisen, C.D., F.I.M. Thomas. 2002. Ammonium uptake by seagrass epiphytes: Isolation of the effects of water velocity using an isotope label. Limnol. Oceanogr. 47, 1223-1229. Cronin, G., M.E. Hay., 1996. Effects of light and nutrient availability on the growth, secondary chemistry, and resistance to herbivory of two brown seaweeds. Oikos. 77, 93-106. Dawes, C.J., D.C. Barilotti., 1969. Cytoplasmic organization and rhythmic streaming in growing blades of Caulerpa prolifera. Amer. J. Bot. 57, 8-16. Drew, E.A., K.M. Abel., 1990. Studies on Halimeda. III. A daily cycle of chloroplast migration within segments. Bot. Mar. 33, 31-45.

70
Dromgoole, F.I., 1987. Light fluctuations and photosynthesis in marine algae. 1. Adjustments of the rate in constant and fluctuating light regimes. Funct. Ecol. 1, 377-386. Duke, C.S., W. Litaker, J. Ramus., 1989. Effects of temperature, nitrogen supply and tissue nitrogen on ammonium uptake rates of the chlorophyte seaweeds Ulva curvata and Codium decorticatum. J. Phycol. 25, 113-120. Dumay, O., G. Pergent, C. Pergent-Martini, P Amade., 2002. Variations in caulerpenyne contents in Caulerpa taxifolia anc Caulerpa racemosa. J. Chem. Ecol. 28, 343-352. Englemann, T.W., 1883. Bacterium photometricum. Ein. Betrag zur vergleichenden Physiologie des Licht-und Farbensinnes. Arch. Physiol. 30, 95-124. Finelli, C.M., D.D. Hart, D.M. Fonseca., 1999. Evaluating the spatial resolution of an acoustic Doppler velocimeter and the consequences for measuring near-bed flows. Limnol. Oceanogr. 44, 1793-1801. Florida marine research institute. 2003. http://www.floridamarine.org Fong, P., K. Kamer, K.E. Boyer, K.A. Boyle., 2001. Nutrient content of macroalgae with differing morphologies may indicate sources of nutrients for tropical marine systems. Mar. Ecol. Prog. Ser. 220, 137-152. Fong, P., J. B. Zedler, R.M. Donohoe., 1993. Nitrogen vs. phosphorus limitation of algal biomass in shallow coastal lagoons. Limnol. Oceanogr. 38, 906-923. Fonseca, M.S., W.J. Kenworthy., 1987. Effects of current on photosynthesis and distribution of seagrasses. Aquat. Bot. 27, 59-78. Fork, D.C., K. Satoh., 1986. The control by state transitions of the distribution of excitation energy in photosynthesis. Annu. Rev. Plant Physiol. 37, 335-351. Francour, P., M. Harmelin-Viven, J.G. Harmelin, J. Duclerc., 1995. Impact of Caulerpa taxifolia colonization on the littoral ichtyofauna of North-Western Mediterranean: preliminary results. Hydrobiologica. 300-301, 345-353. Gacia, E., M.M. Littler, D.S. Littler., 1996. The relationships between morphology and photosynthetic parameters within the polymorphic genus Caulerpa. J. Exp. Mar. Biol. Ecol. 204, 209-224. Gerard, V.A., 1982. In situ water motion and nutrient uptake by the giant kelp Macrocystis pyrifera. Mar. Biol. 69, 51-54. Gerard, V.A., K.H. Mann., 1979. Growth and production of Laminaria longicruris populations exposed to different intensities of water movement. J. Phycol. 15,33-41.

71
Goddard, R.H., C.J. Dawes., 1983. An ultrastructural and histochemical study of the wound response in the coenocitic green alga Caulerpa ashmaedii (Caulerpales). Protoplasma. 114, 163-172. Gomez, I., E. Perez-Rodriguez, B. VInegla, F.L. Figueroa, U.I. Karsten., 1998. Effects of solar radiation on photosynthesis, UV-absorbing compounds and enzyme activities of the green alga Dasycladus vermicularis from southern Spain. J. Photochem. Photobiol. 47, 46-57. Gorbunov, M.Y., P.G. Falkowski, Z. Kolber., 2000. Measurement of photosynthetic parameters in benthic organisms in situ using a SCUBA-based fast repetition rate fluorometer. Limnol. Oceanogr. 45, 242-245. Greene, B.D., J.R. Dixon, J.M. Mueller, M.J. Whiting, O.W. Thornton Jr., 1994. Physiological limitation of phytoplankton photosynthesis in the eastern equatorial Pacific determined from variability in the quantum yield of fluorescence. Limnol. Oceanogr. 28, 165-172. Hader, D.P., H. Hermann, J. Schäfer, R. Santas., 1996. Photoinhibition in the Mediterranean green alga Halimeda tuna Ellis et Sol measured in situ. Photochem. Photobiol. 64, 66-73. Hader, D.P., M. Lebert, C. Jimenez, S. Salles, J. Aquilera, A, Flores-Moya, J. Mercado, B Vinegla, F.L. Figueroa., 1999. Pulse amplitude modulated fluorescence in the green macrophytes, Codium adherens, Enteromorpha muscoides, Ulva gigantean and Ulva rigida, from the Atlantic coast of Southern Spain. Environ. Exp. Bot. 41,247-255. Hader, D.P, M. Porst, M. Lebert., 2000. On site photosynthetic performance of Atlantic green algae. J. Photochem. Photobiol. B. 57, 159-168. Harris, L.G., M.C. Tyrell., 2001. Changing community states in the Gulf of Maine: synergism between invaders, overfishing and climate change. Biol. Invasions. 3, 9-21. Harrison, P.J., C.L. Hurd., 2001. Nutrient physiology of seaweeds: application of concepts to aquaculture. Cahir Biologie Marine. 42, 71-82. Hatcher, B.G., W.D. Larkum., 1983. An experimental analysis of factors controlling the standing crop of the epilithic algal community on a coral reef. J. Exp. Mar. Biol. Ecol. 69, 61-84. Hearn, C.J., M.J. Atkinson, J.L. Falter., 2001. A physical derivation of nutrient-uptake rates in coral reefs: effects of roughness and waves. Coral Reefs. 20, 347-356. Lapoint, B.E., M.W. Clark., 1992. Nutrient inputs from the watershed and coastal eutrophication in the Florida Keys. Estuaries. 15, 465-476.

72
Henely,W.J., 1993. Measurement and interpretation of photosynthetic irradiance-response curves in algae in the context of photo-inhibition and diel changes. J. Phycol. 29, 729-739. Hill, D., P. Coquillard, J. de Vaugelas, A. Meinesz., 1998. An algorithmic model for invasive species: Application to Caulerpa taxifolia (Vahl) C. Agardh development in the North-Weswtern Mediterranean Sea. Ecol. Mod. 109, 251-265. Hillis, L.W., 2001. The calcareous reef alga Halimeda (Chlorophyta, Bryopsidales): a cretaceous genus that diversified in the cenozoic. Palaeogeology Palaeoclimat. Palaeoecol. 166, 89-100. Hurd, C.L., 2000. Water motion, marine macroalgal physiology and production. J. Phycol. 36, 453-472. Jensen, P.R., R.A. Gibson, M.M. Littler, D.S. Littler., 1985. Photosynthesis and calcification in four deep-water Halimeda species (Chlorophycea, Caulerpales). Deep Sea Res. 32, 451-464. Jones, A.B., W.C. Dennison, G.R. Stewart., 1996. Macroalgal responses to nitrogen source and availability: amino acid metabolic profiling as a bioindicator using Gracilaria edulis (Rhodophyta). J. Phycol. 32, 757-766. Kays, W.M., M.E. Crawford., 1993. Convective heat and mass-transfer. Mcgraw-Hill. Kinsey, D.W., A. Domm., 1974. Effects of fertilization on a coral reef environment: primary production studies. Proceedings of the second international symposium on coral reefs (Brisbane, 1973). Great barrier reef committee, Brisbane, Australia. 1, 49-66. Koch, E.W., 1993. The effect of water flow on photosynthetic processes of the alga Ulva lactuca. Hydrobiologica. 260/261, 457-462. Koehl, M.A.R., R.S. Alberte., 1988. Flow, flapping and photosynthesis of Nereocystis luetkeana; a functional comparison of undulate and flat blade morphologies. Mar. Biol. 99, 435-444. Krouse, G.H., E. Weis., 1991. Chlorophyll fluroescence and photosynthesis: The basics. An. Rev. Plant Physiol. Plant Mol. Biol. 42, 313-349. Kuffner, I.B. 2001., Effects of ultraviolet (UV) radiation on larval settlement of the reef coral Pocillopra damicornis. Mar. Ecol. Prog. Ser. 217, 251-261.

73
Landry, M.R., R.T. Barber, R.R. Bidigare, F. Chai, K.H. Coale, H.G. Dam, M.R. Lewis, S.T. Lindley, J.J. McCarthy, M.R. Roman, D.K. Stoecker, P.G. Verity, J.R. White., 1997. Iron and grazing constraints on primary production in the central equatorial Pacific: an EqPac synthesis. Limnol. Oceanogr. 42, 405-418. Larkum, A.W.D., 1983. The primary productivity of plant communities on coral reefs. In Perspectives on coral reefs. 1983, pp. 221-230, CONTRIB. AUST. INST. MAR. SCI., no. 200 Larned. S., J. Stimson., 1996. Nitrogen-limited growth in the coral reef chlorophyte Dictyosphaeria cavernosa and the effect of exposure to sediment-derived nitrogen on growth. Mar. Ecol. Prog. Ser. 145, 95-108. Larned, S.T., 1998. Nitrogen- versus phosphorus-limited growth and sources of nutrients for coral reef macroalgae. Mar. Biol. 132, 409-421. Littler and Littler., 2000. Caribbean Reef Plants. Offshore graphics. Washington D.C. Littler, M.M., 1980. Morphological form and photosynthetic performances of marine macroalgae: tests of a functional-form hypothesis. Bot. Mar. 22, 161-165. Littler, M.M., K.E. Arnold. 1982. Primary productivity of marine macroalgal functional-form groups from Southwestern North America. J. Phycol. 18, 307-311. Martin, J.H., K.H. Coale, K.S. Johnson, S.E. Fitzwater, R.M. Gordon, S.J. Tanner, C.N. Hunter, V.A. Elrod, J.L. Nowicki, T.L. Coley, R.T. Barber, S. Lindley, A.J. Watson, K. Van Scoy, C.S. Law., 1994. Testing the iron hypothesis in ecosystems of the Equatorial Pacific Ocean. Nature. 371, 123-129. Maxwell, K. G.N. Johnson., 2000. Chlorophyll fluorescence – a practical guide. J. Exp. Bot. 51, 659-668. McCook, L.J., J. Jompa, G. Diaz-Pulido., 2000. Competition between corals and algae on coral reefs: a review of evidence and mechanisms. Springer Verlag. http://link.springer.ny.com/link/service/journ…/s000380000129chl00.htm Meinesz, A., L. Benichou, J. Blachier, T. Komatsu, R. Lemee, H. Molenaar, X. Mari., 1995. Variations in the structure, morphology and biomass of Caulerpa taxifolia in the Mediterranean Sea. Bot. Mar. 38, 599-508. Meinesz, A., T. Belsher, T. Thibaut, B. Antolic, K.,Ben Mustapha, C.F. Boudouresque, D. Chiaverni, F. Cinelli, J.M. Cottalorda, A. Djellouli, A. El Abed, C. Orestano, A.M. Grau, L. Ivesa, A. Janklin, A. Langar, E. Massuti-Pascual, A. Peirano, L. Tunesi, J. de Vaugelas, N. Zavodnik, A. Zuljevic., 2001. The introduced green alga Caulerpa taxifolia continues to spread in the Mediterranean. Biol. Invasions. 3, 201-210.

74
Menendez,M., J. Herrera, F. Comin., 2002. Effect of nitrogen and phosphorus supply on growth, chlorophyll content and tissue composition of the macroalga Chaetomorpha linum (O.F. Muell.) Kuetz in a Mediterranean coastal lagoon. Sci. Mar. (Barcelona.). 66, 355-364. Odum, H.T., 1956. Primary production in flowing water. Limnol. Oceanogr. 1, 102-117. Patterson, M.R., 1992. A chemical engineering view of cnidarian sybioses. Am. Zool. 32, 566-582. Patterson, M.R., K.P. Sebens, R.R. Olson., 1991. In situ measurements of flow effects on primary production and dark respiration in reef corals. Limnol. Oceanogr. 36, 936-948. Pedersen, M.F., J. Borum., 1996. Nutrient control of algal growth in estuarine waters. Nutirent limitation and the importance of nitrogen requirements and nitrogen storage among phytoplankton and species of macroalgae. Mar. Ecol. Prog. Ser. 142, 261-272. Pennings, S.C., V.J. Paul., 1992. Effect of plant toughness, calcification and chemistry on herbivory by Dolabella auricularia. Ecol. 73, 1606-1619. Peterson, R.D., 1972. Effects of light intensity on the morphology and productivity of Caulerpa racemosa (Försskal) J. Agardh. 8(1/2), 63-86. Ralph, P.J., R. Gademann, W.C. Dennison., 1998. In situ seagrass photosynthesis measured using a submersible, pulse-amplitude modulated fluorometer. Mar. Biol. 132, 367-373. Raven, J.A., F.A. Smith, S.M. Glidewell., 1979. Photosynthetic capacities and biological strategies of giant celled and small celled macro-algae. New Phytol.. 83, 299-309. Relini, G., M. Relini, G. Torchia., 2000. Fish population changes following the invasion of the allochthonous alga Caulerpa taxifolia in the Ligurian Sea (N.W. Mediterranean). ICES-CM. 2000/U:17.(http://www.ices.dk/asc/2000/abstracts/lists.u.htm). Riber, H.H., R.G. Wetzel., 1987. Boundary-layer and internal diffusion effects on phosphorus fluxes in lake periphyton. Limnol. Oceanogr. 32, 1181-1194. Riechert, R., C.J. Dawes., 1986. Acclimation of the green alga Caulerpa racemosa var. uvifera to light. Bot. Mar. 29, 533-537. Rolph, F.J., R.R. Sokal., 1981. Statistical Tables 2nd edition. W.H. Freeman Company. New York. Ryther, J.H., W.M. Dunstan., 1971. Nitrogen phosphorus and eutrophication in the coastal marine environment. Science. 171, 419-426.

75
Saffo, M.B., 1987. New Light on Seaweeds. Bioscience. 37, 654-664. Sagert,S., H. Schubert., 2000. Acclimation of Palmaria palmata (Rhodophyta) to light intensity: comparison between artificial and natural light field. J. Phycol. 36, 1119-1128. Schubert, H., S. Sagert, R.M. Forester., 2001. Evaluation of the different levels of variability in the underwater light field of a shallow estuary. Helgol. Mar. Res. 55, 12-22. Schwartz,A.M., M. Björk, T. Buluda, M. Mtolera, S. Beer., 2000. Photosynthetic utilization of carbon and light by two tropical seagrass species measured in situ. Mar. Biol. 137, 755-761. Smith, C.M., L.J. Walters., 1999. Fragmentation as a strategy for Caulerpa species: fates of fragments and implications for management of an invasive weed. Mar. Ecol. 20, 307-319. Stevens, C.L., C.L. Hurd., 1997. Boundary-layers around bladed aquatic macrophytes. Hydrobiologica. 346, 119-128. Stimson, J., S.T. Larned., 2000. Nitrogen efflux from the sediments of a subtropical bay and the potential contribution to macroalgal nutrient requirements. J. Exp. Mar. Biol. Ecol. 252, 159-180. Stimson, J., S.T. Larned, E. Conklin., 2001. Effects of herbivory, nutrient levels, and introduced algae on the distribution and abundance of the invasive macroalga Dictyoshaeria cavernosa in Kaneohe Bay, Hawaii. Coral Reefs. 19, 343-357. Svedelius, N., 1906. Reports on the marine algae of Ceylon. No. I. Ecological and systematic studies of the Ceylon species of Caulerpa. Ceylon. Mar. Biol. 4, 81-144. Taylor, D., S. Nixon, S. Granger, B. Buckley., 1995. Nutrient limitation and the eutrophication of coastal lagoons. Mar. Ecol. Prog. Ser. 127, 235-244. Thomas, F.I.M., M.J. Atkinson., 1997. Ammonium uptake by coral reefs: Effects of water velocity and surface roughness on mass transfer. Limnol. Oceanogr. 42, 81-88. Thomas, F.I.M., C.D. Cornelisen, J.M. Zande., 2000. Effects of water velocity and canopy morphology on ammonium uptake by seagrass communities. Ecology. 81, 2704-2713. Thomas, F.I.M., C.D. Cornelisen. 2003. Ammonium uptake by seagrass communities: effects of oscillatory versus unidirectional flow. Mar. Ecol. Prog. Ser. 247, 51-57.

76
Uchimura, M., A. Rival, A. Nato, R. Sandeaux, J. Sandeaux, J.C. Baccou., 2000. Potential use of Cu2+,K+ and Na+ for the destruction of Caulerpa taxifolia: differential effects on photosynthetic parameters. J. App. Phycol. 12, 15-23. Verlaque, M., C.F. Boudouresque, A. Meinesz, V. Gravez., 2002. The Caulerpa racemosa complex (Caulerpales, Ulvophyceae) in the Mediterranean Sea. Bot. Mar. 43, 49-68. Walters, L.J., C.M. Smith, J.A. Coyer, C.L. Hunter, K.S. Beach, P.S. Vroom., 2002. Asexual propagation in the coral reef macroalga Halimeda (Chlorophyta, Bryopsidales): Production, dispersal and attachment of small fragments. J. Exp. Mar. Biol. Ecol. 278, 47-65. Walz. Underwater Fluorometer Diving PAM. Handbook of Operation. 1st edition. July 1998. Effeltrich, Germany. pp. 101. Wang, J.D., 1998. Subtidal flow patterns in Western Florida Bay. Estuar. Coast. Shelf. Sci. 46, 901-915 Wellnitz, T., R. Rinne., 1999. Photosynthesis response of stream periphyton to fluctuating light regimes. J. Phycol. 35, 129-130. Wheeler, W.N., 1980. Effect of boundary layer transport on the fixation of carbon by the giant kelp Macrocystis pyrifera. Mar. Biol. 56, 103-110. Wheeler, P.A., B. R. Bjornsater., 1992. Seasonal fluctuations in tissue nitrogen, phosphorus, and N:P for five macroalgal species common to the pacific northwest coast. J. Phycol. 28, 1-6. Williams, S.L., R.C. Carpenter., 1998. Effects of unidirectional and oscillatory water flow on nitrogen fixation (acetylene reduction) in coral reef algal turfs, Kaneohe Bay, Hawaii. J. Exp. Mar. Biol. Ecol. 226, 293-316. Williams, S.L., 1984. Uptake of sediment ammonium and translocation in a marine green macroalgae Caulerpa cupressoides. Limnol. Oceanogr. 29, 374-379. Williams, S.L., R. C. Carpenter., 1998. Effects of unidirectional and oscillatory water flow on nitrogen fixation (acetylene reduction) in coral reef algal turfs, Kaneohe Bay, Hawaii. J. Exp. Mar. Biol. Ecol. 226, 293-316. Williams, S.L., 1990. Experimental studies of Carribean seagrass bed development. Ecol. Monogr. 60, 449-469. Wing, S.R., M.R. Patterson., 1993. Effects of wave-induced light flecks in the intertidal zone on photosynthesis in the macroalgae Postelsia palmaeformis and Hedophyllum sessile (Phaeophycea). Mar. Biol. 116, 519-525.

77
Wing, S.R., J.J. Leichter, M.W. Denny., 1993. A dynamic model for wave-induced light fluctuations in a kelp forest. Limnol. Oceanogr. 38, 396-407. Yentsch, C.S., C.M. Yentsch, J.J. Cullen, B. Lapointe, D.A. Phinney, S.W. Yentsch., 2002. Sunlight and water transparency: cornerstones in coral reasearch. J. Exp. Mar. Biol. Ecol. 268, 171-183. Zuljevic, A., B. Antolic 2000., Synchronous release of male gametes of Caulerpa taxifolia (Caulerpales, Chlorophyta) in the Mediterranean Sea. Phycologia. 39, 157-159.