varied carnivore effects and the prevalence of intertidal algal turfs
TRANSCRIPT

.I. E,YI~. MUV. Viol. Kcol., 166 (1993) 65-91 0 1993 Elscvier Science Publishers BV. All rights reserved 0022498 1~93/$06.00
JEMBE 01894
Varied carnivore effects and the prevalence of intertidal algal turfs
Carlos Robles and Jack Robb
(Received 6 January 1992; revision received 11 August 1992: accepted 3 I August 1992)
Abstract: A red algal turf covers mid-shore levels of much of the California Channel Islands. Our expcri- menta at Santa Catalina Island show that carnivores maintain the turf by consumingjuvenile mussels ,W~~tihr.c
spp. and associated invertebrates. Exclusion of spiny lobsters Punulirza inrerruptuy (Randall) from compar-
atively wave-cxposcd sites caused the complete and persistent replacement of the turf by the mussel asscm- blagc. On sites protected from wave action, the lobsters were joined by carnivorous fishes, i.c., Hrdichorw
.\emi.sinctu.c (Ayrcs). Ox~$dis crrhfbrnica (Gunther) and Srmico.r.~~phu.v pulcher (Ayrcs), and whelks, i.e..
Ccrcrro.rr~ma nr~tftrlli (Conrad) and MuxwrNiu ~emmrr (Sowcrby). Compensatory foraging shifts by the whelks prevented significant increases of mussels in lobster and fish cxclosures. Exclusion of all predators at the protected site caused a moderate but statistically significant increase. Thus. experiments at different sites indicated either “kcystonc” or “diffuse” effects of predators.
Experimental clumps of large Mj~tilir.~ ccrlifkwionus (Conrad) persisted longer after rc-exposure to preda- tion in the post-experimental periods than did clumps of smaller M. c&vrkurus or M. rdu/i.v L. Transplants of matched cohorts indicated that the size differences of mussels in the exclosure experiments wcrc caused
by diffcrcnccs in shell growth rates correlated with wave exposure. We speculate that changing growth rates may inHuencc the outcome of size-limited predation, accounting for some of the variation in the rclattvc abundance of turf and mussels over the wave exposure gradient.
Key words: Algal turf; Corallim~; Growth; Haemurop~r.~; Indirect effect; Intertidal zone; Kcystonc predator:
Labrid; ~Mrril~rs; Panulirus: Sire-limited predation: Whelk
After > 3 decades of field experiments (reviews in Sih et al., 1985; Menge & Far-
rell, 1989) it is clear that predation often molds the structure of benthic communities.
Yet, for many benthic communities, the effects of specific predators remain obscure.
Below, we examine two contrasting depictions of predator effects, the “keystone” and
“diffuse” predation hypotheses, in a rocky intertidal community.
We recognize a keystone predator by a consequence of its removal: marked shifts
in relative abundance at lower trophic levels, including the replacement of dominant
species (Paine, 1966, 1969, 1974, 1980). For example, urchin barrens apparently rc-
Correspondence address: C. Robles, Department of Biology. California State University. Los Angclcs, CA 90032, USA.
Contribution 143 to the publications of the University of Southern California Catalina Marine Science Ccntcr.

66 C. ROBLES AND J. ROBB
placed kelp beds following otter removals from shallow subtidal communities (c.g.,
Estes & Palmisano, 1974; Dayton, 1975; Simenstad et al., 1978; Duggins, 1980).
Mussel beds replaced perennial algae following sea star removals from rocky shores
(e.g., Paine, 1966, 1974). Both the original dominant species and their replacements
may harbor diverse “associated species” (e.g., Suchanek, 1979; Tsuchiya & Nishihira,
1986; Dean & Connell, 1987). The patterns of species interactions differ between the
pre- and postremoval assemblages because: (1) some species exist in only one assem-
blage; and (2) species shared by the assemblages may play different roles within each.
Thus, a transformation of community structure attends the change of dominants, even
though the alternative assemblages may share some species.
The concerted effects of several predator species, which we term “diffuse” predation,
is revealed by a different experimental design and outcome. The field experiments arc
compound removals, in which the different members of the predator fauna are removed
both separately and in combination (e.g., Peterson, 1979; Menge & Lubchenco, 198 1;
Lubchenco et al., 1984; Menge et al., 1986). Predator effects may show no interaction
between different treatments or they may interact. In the former, the probability of the
prey falling to one predator species does not depend on the presence or absence of a
second. With interaction, the probability of the prey falling to one predator species does
depend on whether others are removed (see Wilbur & Fauth, 1990, for the statistical
treatment of interaction among predator effects). Menge and Lubchenco (1981) present
an apparent case of interaction wherein significant increases in prey required the re-
moval of all or nearly all predator species. One explanation for such effects is the
compensatory foraging shifts hypothesis: after removal of one, the remaining predators
respond numerically (sensu Helling, 1959) and drive the prey’s densities towards
premanipulation levels. An increase in the proportion of that prey species in the diets
of the remaining predators also might be observed. However, no data exist confirm-
ing aggregation or diet shifts of unmanipulated predators.
Whether interactive or not, diffuse predation differs from keystone predation because
the removal of any one predator produces comparatively small changes in the abun-
dance of dominant prey and associated species (e.g., Menge & Lubchenco, 1981).
WC investigated predation on the M. cnlifornhnus assemblage in a community set-
ting very different from the original account of keystone predation (Paine. 1966, 1974).
The alternative to the mussel bed assemblage is a red algal turf that forms an unbro-
ken sward across much of the mid-shore levels of the California Channel Islands. The
turf was thought to owe its prevalence to tolerance of physical stresses and weak bi-
otic interactions, i.e., moderate browsing by fishes (Littler & Littler, 1980; Hay, 1981;
Stewart, 1982). Indirect effects of predators (sensu Kerfoot, 1987) had not been stud-
ied.
The predators included spiny lobster Punulirus interruptus (Randall) labrid fishes, i.e.,
Hnlichoeres semisinctus (Ayrcs), O.xyjulis calcjbrnica (Gunther), and Semicossyphus pul-
cher (Ayres), and whelks, i.e., Cerutostoma nuttalli (Conrad), and Maxwelliu gemnul
(Sowerby). Depending on differences in the abundance of predators among sites,

CARNIVORE EFFECTS 67
lobster exclusion alone or compound removal experiments were done. For the latter,
we collected data on compensatory foraging shifts and diffuse effects.
Earlier, C. Robles (1987; Robles et al., 1990) demonstrated that the lobster’s rapid
movement and short prey handling times denied mussels an upper intertidal spatial
refuge, such as that described for the sea star-mussel interaction (Paine, 1966, 1974).
Lobster exclusion increased the relative abundances of Mytilus spp. in the understory
of high intertidal beds of fucoid algae (Robles, 1987). Among the results we now re-
port is the complete and persistent replacement of a major vegetal formation, the algal
turf, in lobster exclosures below the zone of the fucoid algae.
METHODS
NATURAL HISTORY
We studied rocky shores near the Catalina Marine Science Center, Santa Catalina
Island (33’ 25’ N, 118’ 30’ W). Mean monthly surface temperatures varied from 14 ‘C
in January to 20.5 “C in September. The tidal excursion ranged from -0.7 to + 2.2 m
mean lower low water (MLLW).
In our initial Catalina observations and prior studies of temperate zone littoral
mussel predators (e.g., Menge, 1976, 1983; Peterson, 1979; Robles et al., 1989) rela-
tive abundances varied markedly across gradients of wave exposure. Therefore, we
chose three sites to span the range of wave exposures over which the algal turf occurred.
The protected site was a horizontal bench 0.5 m above MLLW on the southern side
of Big Fisherman’s Cove (Fig. 1). This site was protected from wave action by the walls
of the cove, a submerged reef, and a rocky islet (Bird Rock) ~0.75 km to the wind-
ward. The semiprotected and exposed sites were near the leeward and windward ends
of the islet. The semiprotected site was a horizontal bench 0.7 m above MLLW rc-
ceiving swells that refracted around the island. The exposed site was a similar platform
0.9 m above MLLW that received the full force of prevailing northwesterly swells.
The turf covers mid-shore levels (= 0.1 to + 1.0 m MLLW) of the islands from
sheltered covers to all but the most wave-exposed promontories (Emerson & Zelder.
1978; Murray & Littler, 1978; Murray et al., 1980; Thorn & Widdowson, 1980; Stewart,
1982). Only 1-3 cm thick, the turf consists of the branching thalli of the anchor spe-
cies, principally Corullina ojficinalis L., entwined with epiphytes, predominantly Gigrrr-
tinu canaliculrrtn (Harvey), Laurencia pacifica (Kylan), and Gelidium coulteri (Harvey).
The relative abundance of species in the turf varies seasonally, the epiphytes increas-
ing in summer (Murray & Littler, 1978; Stewart, 1982).
Invertebrates were present underneath or in gaps in the turf. These included barna-
cles, Tetruclitcr squamosa var. rubescens (Darwin); jewel box oysters Pseudochnma e.x--
og)wz (Conrad); and sessile tubiculous gastropods, Aletes squumigerus (Carpenter).
Three species of mussels occurred in the turf: Mytilus californianus, M. edulis and
Brcrchidontes czdumsianus (Dunker). [MacDonald & Koehn (1988) provide electro-

6X C. ROBLES AND J. ROBB
Exposed Site
Semi Protected Site
ISTHMUS REEF
Fig. 1. Location of study sites. Contour lines for 2 and 5 m below MLLkV.
phorctic data suggesting that the southern California form ofM. edulis is in fact ~4, ,$-
Ioprowk’ulis (Lamarck).] The M_~jtilus spp. recruit in densities sometimes exceeding
10000 m-‘, but few survive to reproductive sizes (2-3 cm long; Suchanek, 198.5; C.
Robles, pers. obs.). ~rae~~d~nte.~ is a small mussel (adults t3 cm long at Catalina).
Though numerous, it habitually nestles in crevices and does not form mussel beds
(Robles, 1987).
The predators included both resident predators, which remain within the intertidal
zone at all times, and transient predators, whose presence in the intertidal zone depends
on tidal, daylight, or migratory cycles. The resident predators were whelks, Cerf~rff.~~#r~~ff

CARNIVORE EFFECTS 69
~~utt~lli and ~a,~~el~ia gemmu. The transient predators were diurnal fishes in the La-
bridae, Haiichoeres sernisimtus, O.x_@dis culjbrniea, and Semicus.yvphus p&her; a bird,
the black oystercatcher, Haematopus bathmani Audubon, and P. interruptus. The vulnerability of mussels to different predators depends on their species and size.
Whelks, crabs, and lobsters prefer M. edulis to M. cal$mkwus @Larger, 1967. 1972;
C. Robles, pet-s. obs.). In laboratory feeding trials (Robles et al., 1990; C. Rob&
unpubl. data), the largest M. edulis occurring on Catalina (7 cm) were readily eaten by
all lobsters, even small subadults; whereas M. cal~fkziunus > 6 cm were invutnerable
to lobsters < 9.5 cm carapace length, z 80”; of those foraging intertidally. Gape limits
of the labrid fishes restrict their feeding to mussels <4 cm. H. b~~h~~~~zj, the black
oystercatcher. eat both ~~~j/M.~ spp, and can kill M. ~~l~i~~~?~~~~?u~ > 9 cm (Hartwick,
1976; this study).
OBSERVATIONAL METHODS
The apparent differences in wave action were confirmed by divers, who estimated
bottom velocities during high tide surveys of lobster abundance (see below). They timed
the oscillatory drift of neutrally buoyant spheres (perforated ping pong balls) or flot-
sam over grids painted on the sLlbstratun1. A grid was located on the protected site,
the s~miprotected site, and z 30 IX to the lee of the exposed site. The latter placement
and our un~~illingness to sample high wave action (8 m was the largest horizontal
displacement endured in the dive surveys) provided an underestimate of mean bottom
velocity of the exposed site. The velocities lo-40 cm above the substratum were esti-
mated on 21 nights in spring-summer 1986. The divers also noted horizontal displace-
ment (no period was estimated) in the middle of the cove and islet transects in all
seasons 1982-87.
Predator nbur~dunce and diet
Densities of whelks at the protected site were estimated by counting their numbers
in 50 9#0-cm’ square quadrats running c~~ntiguously along the mid-level of the turf
zone. Similar censuses were not done on the islet because years of close scrutiny of
the turf had failed to reveal a single whelk.
Divers using either snorkel or scuba estimated the densities of the labrid fishes, i.e.
S. pulr’her, H. .semisinctus, and 0. calijk-r&a, by swimming along permanently marked
20-m belt transects on the midline of the turf zone. Surveys were made on tides cov-
ering the turf by at least 1 m and falling between sunrise and sunset; the latter restriction
w’as imposed because the fishes are diurnal (Hobson et al., 198 1). The transects at the
protected site spanned the experimental area. On the islet, the 60 m of transects fell
midway between the semiprotected and the exposed site, with either end of the transect
line 30-40 m away from the two experimental areas. Lobster densities were estimated

70 C. ROBLES AND J. ROBB
by swimming transects during nocturnal high tides (within 1 h of predicted tides
> + 1.3 m MLLW, at least 1 h after sunset and 1 h before sunrise). Details of whelk,
fish, and lobster surveys appeared in Robles (1987).
Avian mussel predators (Marsh, 1986) were never observed at the protected site.
However, a pair of black oystercatchers used the islet as a nest site from April to August
each year. We estimated the foraging effort devoted to the turf zone by making repeated
spot checks of the birds’ location and activity. At all times of day and phases of the
tides, we motored along the shore of the islet in a small skiff. Once the birds were
sighted, their behavior and position relative to the turf zone were recorded. We used
this spotting method because the birds changed position and activity when people were
on the islet. z 100 surveys were made from 1985 to 1990. Here, we report data taken
when the turf zone was accessible to the birds (water level < + 0.6 m MLLW).
The apparent diets of whelks, fishes, and lobsters at the protected site were reported
previously (Robles, 1987). For comparison, we made additional collections of lobsters
from the turf zone of the islet. We also directly observed oystercatcher kills during some
of the spotting surveys, collecting the shells to measure sizes and confirm initial iden-
tifications.
EXPERIMENTAL METHODS
Experiments at the protected site
The predator abundance and diet surveys indicated that whelks, fishes, and lobsters
killed mussels at the protected site. Here, we excluded transient predators (lobsters and
fishes), either separately, or in combination with the whelks, in two successive exper-
iments.
The first experiment tested whether the transient predators limited the survival of
mussel recruits over a broad stretch of turf while predation by whelks was greatly re-
duced (Table I). Fishes and lobsters were excluded from plots with large mesh cages.
Hardware cloth with square openings of 2.5 cm (89”; open area) was bent into dome-
shaped cages ~20 cm high and 35 cm long, coated with a saltwater-resistant epoxy,
and nailed to the sandstone substratum. To control for possible artifacts caused by the
wire mesh, each cage exclosure was matched with a similarly prepared open-ended
cage, which admitted lobsters and fishes (Robles, 1987) and an open plot, with no mesh
(Fig. 2). The locations of treatments and control plots of three replicates were assigned
randomly along a 30-m strip of turf. Since the whelks could pass through the large
mesh, we removed them from the site by hand at monthly or shorter intervals. Initial
removals yielded hundreds of whelks, but after 10 months, near the end of the exper-
iment, 1 h of searching the site (E 100 m2) yielded only four.
The second experiment at the protected site tested whether the whelks could limit
the survival of mussel recruits in the absence of the transient predators, and if so,
whether this regulation could be explained by compensatory foraging shifts, i.e., the

CARNIVORE EFFECTS 71
TABLE I
Summary of experimental design
Experiment Date (month/year) Manipulation Predators cxcludcd
First protected
Second protected
12182 to 1 l/83
04186 to 12i86
Manual removals of cc helks
from bench Open plots Large mesh arch
Large mesh cage
Open plots Large mesh arch
Large mesh cage Small mesh arch Small mesh cage
Semiprotected 04186 to 02/87 Open plots None Large mesh arch None Large mesh cage Lobsters
Exposed OS/S5 to 03:‘86 Open plots Large mesh arch Large mesh cage
None None Lobsters
Whelks
Whelks Whelks, lobsters and fishes
None None Lobsters and fishes Lobsters and fishes
Lobsters, fishes and whelks
Ftp. 2. A replicate in the large mesh exclosure experiments. From left to right: arch control, open plot control. and cage treatment.

I? C. ROBLES AND J. ROBB
whelks aggregating in the transient predator exclosures. To do this, we used large and
small mesh cages to exclude transient predators either separately or in combination
with the whelks. Five different treatments were randomly assigned to turf plots and
replicated six times. The treatments were: (I) a small mesh cage, excluding both whelks
and the transient predators: (2) a small mesh arch, excluding only the transient prcd-
ators; (3) a large mesh cage, again excluding only the transient predators; (4) a large
mesh arch. accessible to all predators; and (5) an open plot, also accessible to ail
predators (Table I).
The small mesh exclosures were A-frame cages, 20 cm high, 25 cm wide at the base,
fashioned from hardware cloth of 1.2-cm” openings, and supported by a 0.5cm steel
rod running up the center of either end and along the peak. The ends of the rod were
set in concrete in l-cm diameter holes drilled in the rock. The stnall mesh arches were
similar in every way, except that a strip of the wire cloth was removed from the base
of the end panels to provide 2 cm clearance for the whelks. The small mesh cages were
not perfect cxclosures; occasionally a few whelks, small enough to fit through the mesh,
cntcrcd and consumed mussels. They were removed at regular intervals with long
forceps. Thus, the small mesh cages were most similar to the large mesh treatment in
the whelk manual removal experiment, testing the effect of transient predators when
whelk predation was reduced, but not eliminated.
The experiment used the entire turf zone of the protected site. The six replicates wcrc
arranged in two groups, three replicates along the shoreline of the first (whelk manual
removal) experiment, and three along the shore not subject to removals.
To test for compensatory foraging shifts, biweekly records of the experimental whelk
densities were analyzed using a multivariate general linear model of repeated measures
ANOVA (software: SYSTRT, Evanston, Illinois). The model contained two main,
between-subject, effects: treatment group (cage assignment) and location (previous
whelk manual removal or nonremoval shorelines). The variation among census dates
was partitioned as the within-subject effect, i.e., the repeated measure. Post hoc con-
trasts were used to determine which treatment groups differed significantly on specific
survey dates. If the whelks did respond numerically to increases in mussel abundances
resulting from the removal of lobsters and fishes, then their densities would increase
significantly in the large cage and small arch plots, but not in the large mesh arch plots
or open plots. Significant increases in the per cent cover of mussels would occur only
in the small cage treatment (excluding all predators). The comparisons between large
arch and open plots, and between small arch and large cage, tested for artifacts of wire
mesh, because the effects on predators were equivalent, but the cover of wire differed
within each pair.
E.qwriments at semiprotected and exposed sites
The predator abundance and diet surveys showed that lobsters were the primary
mussel predator on the isfet sites (see Results). Therefore, large mesh cages were

CARNIVORE EFFECTS 73
sufficient to stop predation. Three replicates were arranged on the turf of the semipro-
tccted site. The exclosures remained in place from 4 April 1986 until 28 February 1987.
when winter storms removed one of the exclosures (Table I).
From 1983-84, turf completely covered the most wave-exposed site. In May 198.5,
this site showed a heavy set of Myrilus spp.. Mussels covered 67”~” of the turf, reach-
ing maximum lengths of 3 cm. We installed three replicates of each treatment of the
large mesh exclosure experiment on 23 June 1985 and left them in place for 4 wk. On
22 July 1985, we detached the cages, recorded the results, and then reattached the cages
(Table I).
All experiments were concluded by removing the cages and visually estimating the
per cent covers of algae and invertebrates. We verified the accuracy of our visual per
cent cover estimates by comparing them with a conventional technique employing point
sampling of color transparencies (e.g., Cubit, 1984). Estimates of mean per cent cov-
ers differed between the two methods by < 5 “/, . For the first protected site and the
exposed site experiments we also scraped up and preserved for microscopic examina-
tion the cover of a 230-cm’ quadrat from the center of each experimental plot. These
samples provided densities and shell lengths for the mussels and herbivorous mollusks.
The scraping destroyed less than half of the substratum under the cages, allowing the
intact portion to be observed after the conclusion of an experiment. Notes were kept
on the cover and sizes of experimental mussels in the post experimental period. i.e..
after cages were removed, re-exposing the plots to predators.
The variances of per cent cover estimates were found to be “0” for some experimental
groups, and thus, heteroscedasticity and the differences among the means could not
be tested using F ratios. Kruskal-Wallis tests were applied to mean mussel covers of
all the groups in an experiment, and when these proved significant, Mann-Whitney C’
tests were used to compare covers on open plots with each of the other experimental
groups (Sokal & Rohlf, 1981). For the second protected site experiment (Table I),
which required four of the latter tests, a procedure-wise error rate of 5’:” was main-
tained using Bonferroni’s procedure (Sokal & Rohlf, 1981).
With one exception, original or square root-transformed densities were homosce-
dastic. The differences in means were tested with ANOVA, and statistically similar
means were grouped using Tukey’s HSD tests (Sokal & Rohlf, 1981). The exception
was sample variances of mussels > 1 cm, which on control plots often were at or near
zero, requiring Kruskal-Wallis tests.
RESULTS
WATERMOTION
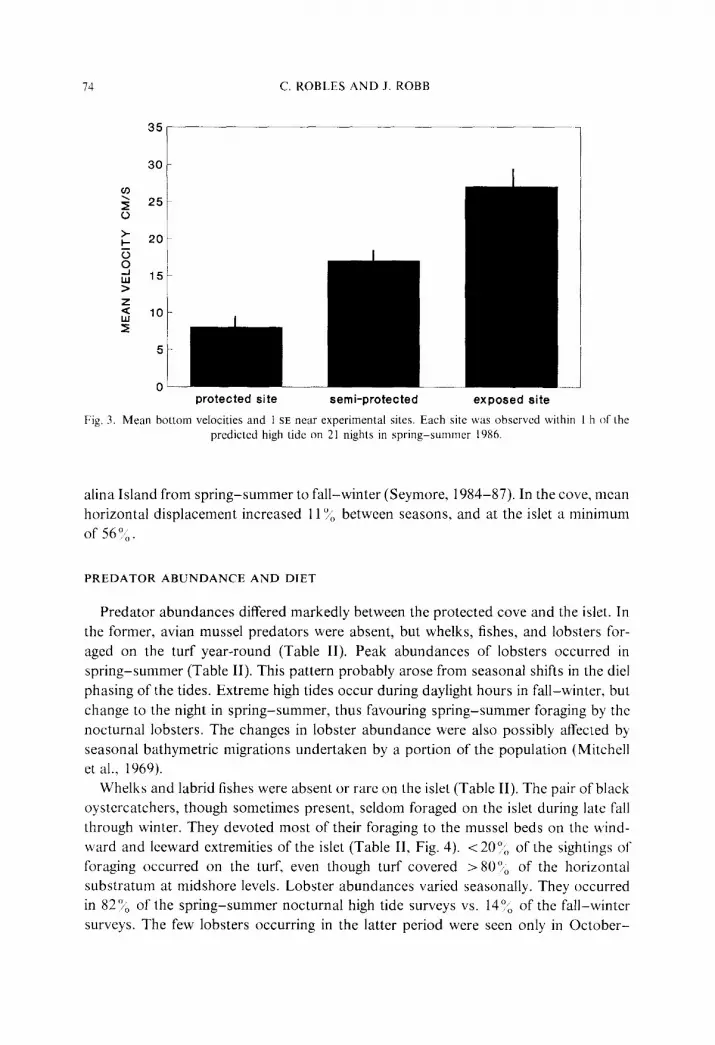
The mean velocity estimates differed more than three-fold between protected and
exposed sites (Fig. 3). Significant swell heights increased in the vicinity of Santa Cat-
C. ROBLES AND J. ROBB
35 I
30 -
0 protected site semi-protected exposed site
Fig. 3. Mean bottom velocities and 1 SE near experimental sites. Each site was observed withm I h of the predicted high tide on 21 nights in spring-summer 1986.
alina Island from spring-summer to fall-winter (Seymore, 1984-87). In the cove, mean
horizontal displacement increased 11 Y/l> between seasons. and at the islet a minimum
of 56”,.
PREDATOR ABUNDANCE AND DIET
Predator abundances differed markedly between the protected cove and the islet. In
the former, avian mussel predators were absent, but whelks, fishes, and lobsters for-
aged on the turf year-round (Table II). Peak abundances of lobsters occurred in
spring-summer (Table II). This pattern probably arose from seasonal shifts in the die1
phasing of the tides. Extreme high tides occur during daylight hours in fall-winter, but
change to the night in spring-summer, thus favouring spring-summer foraging by the
nocturnal lobsters. The changes in lobster abundance were also possibly affected by
seasonal bathymetric migrations undertaken by a portion of the population (Mitchell
et al., 1969).
Whelks and labrid fishes were absent or rare on the islet (Table II). The pair of black
oystercatchers, though sometimes present, seldom foraged on the islet during late fall
through winter. They devoted most of their foraging to the mussel beds on the wind-
ward and leeward extremities of the islet (Table II, Fig. 4). ~20”~ of the sightings of
foraging occurred on the turf, even though turf covered >80”, of the horizontal
substratum at midshore levels. Lobster abundances varied seasonally. They occurred
in 82’i,, of the spring-summer nocturnal high tide surveys vs. 141; of the fall-winter
surveys. The few lobsters occurring in the latter period were seen only in October-

CARNIVORE EFFECTS 75
TABLE II
Densities of mussel predators in the turf zone of the cove (protected) and the islet (semi-exposed and ex-
posed). Mean number f 1 SD,m ~’ are tabulated by location and season (s/s, spring-summer; f/w, fall-
winter). Number of surveys (N) and number of samples.survey. ’ (n) between parentheses.
Cerurostoma
cove b,‘S 2.1 f 4.7 (2!50) 2.3 i 4.1 (2/50) 0.046 2 0.330 (25.6) $‘\v 4.4 * 7. I (2!‘50) 1.3 k 2.5 (2.‘50) 0.028 + 0.360 (I 5 6)
lslct S/S 0.040 + 0.044 ( 19.6) 1.; w 0.001 t 0.003 (14’6)
cow $5 0.010+0.021 (17:3) 0.047 f 0.061 (7:3) 0.025 k 0.045 (7.3) S’\h 0.010 0.020 2 (23,/3) 0.053 f 0.077 (7/3) 0.000 t 0.000 (7:3)
Islet GS 0.000 2 0.000 (5;3) 0.002 + 0.005 (3:‘3) 0.006 &0.012(3.3) r;vv 0.001 0.003 + (20,‘3) 0.008 i 0.014 (7:3) 0.000 k 0.000 (7 3)
COW Islet \‘b s,‘s 4.8 x 10 J+9.8x 10 1 (23:‘l) I“\\ s’u 1.3 x 10 ‘i4.l x IO 1 (22,‘l)
70 i
b 30 -
5 20-
2 a lo
:
Mussel beds Surf Grass Turf
m Spring/summer m Fall/winter
Fig. 4. Frequency of black oystercatcher foraging on different intertidal substrates of the islet. Percent of total sightings of foraging birds are graphed by season and substratum type.

16 C. ROBLES .AND J. ROBB
November. Nevertheless, lobsters were by far the most abundant large predator on the
islet. reaching mean densities on the turf transects of 0.17.m-’ on single evenings in
spring-summer. Taking into consideration the frequency of feeding individuals within
total densities (100 vs. 7856, respectively, for lobsters and the black oystercatchers),
foraging lobsters were at least 100-fold more abundant than foraging birds on the turf
in spring-summer.
,411 predators ate A4~~ilus spp., and all but the whelks ate the associated herbivores:
limpets, chitons, and grapsid crabs (Robles, 1987; this study; see U’inget, 1968, for
discussion of lobster diets). Mussel shell fragments in the gut were found more frc-
qucntly and in greater volume in the lobsters than the labrid fishes from the protected
site (Roblcs, 1987). Of 25 lobsters collected on six occasions from the turf of the islet,
100°O contained Mytilus spp., 489, Bmchidontes. Afvtilus spp. also constituted by far
the largest proportion of recognizable fragments in the cardiac stomachs, though shells
of herbivorous gastropods were also found.
Of 57 kills observed during 18 black oystercatcher spotting surveys, 35”,, were
M. cal~j~wr~imus, 9’1, M. rdulis, 34”,, limpets, and 22O, coiled gastropods (A.Ptrcreu sp.,
Norrisiu sp., and Tegukz spp.). The birds killed M. cnlifbrnimus l-9 cm (mean length
5.2 cm). Small (< 3 cm) M. cul$minnus constituted < 5 “<, of the total mussel kills.
24.
Mean No. Mytilus per 100 cm2
Fig. 5. Proportion of mussels in the apparent diet of whelks. Dietary records were selected for periods within I month of one of four dates of mussel density surveys. The later were chosen to show the extreme tem- poral variation in MJ~U.T spp. abundance. Graphs are arranged by increasing density of musscls. Solid bar, .M. rdulis; fine hatch, M. cul$mianu.v; diag. hatch, Brcrchidonres sp.; clear, the barnacle, Trtruclircr sp.; and
coarse hatch, other prey.

CARNIVORE EFFECTS 71
To determine whether the whelks’ diet changed with the abundance of Mytilus spp.,
as expected under the compensatory foraging shifts hypothesis, we compared the
proportions of Myrilus spp. in the apparent diets at times of different mussel abun-
dances in the turf. The abundances of small mussels had been estimated by micro-
scopically examining samples of turf (details in Robles, 1987). When Mytilus spp.
abundances were high, they comprised the largest proportion in the diet; when low,
barnacles were the greatest proportion (Fig. 5).
EXPERIMENTS AT THE PROTECTED SITE
In the first experiment at the protected site (whelk manual removal, Table I), mean
total densities of M. edulis were significantly greater for the exclosures than the arch
controls (Table III). Total densities for the open controls were intermediate and did
not differ significantly from either the arch or exclosure means. Bruchidontes showed
similar, but nonsignificant, trends (Table III). The nonsignificant tendency for total
densities to be lower in the arch than open controls suggests that the wire mesh de-
pressed the numbers of small mussels, possibly by interfering with settlement. If so, then
the test for effects of predation would be conservative, because mussel recruitment
should be similarly depressed in the exclosures. Whelk aggregation under the mesh to
avoid desiccation stress would not have been a factor because after the initial removals
whelks were only found at the periphery of the site. Total densities of M. cal~fortzimus
increased slightly, though significantly, in exclosures relative to both the arch and open
controls.
Densities of mussels > 1 cm were uniformly low in both open and arch plots, and
significantly higher in the exclosures (Table III). The differences were significant for
TABLE III
Mean numbers f 1 SD of mussels.230-cm’ sample- ’ in the first protected site experiment (whelk manual removals) are tabulated with respect to experimental conditions and shell length. P values of ANOVA fo- low 4 values. The df values in the ANOVAs were 2 and 6, respectively. for effects of treatment and error. Tukey’s HSD tests (Sokal& Rohlf, 1981) were used to distinguish significant from nonsignificant differcncea
(underlined). Comparisons with SD values of 0 were tested with Kruskal-Wallis tests, and Y values were not grouped. The df values for these tests were 2.
Species Open Arch Cage P value
M. rduh
Total density Density > I cm
31.00~4.58 17.00 t 10.82 83.33 k 40.77* 0.028 0.00 & 0.00 0.00 + 0.00 33.33 i 6.69 0.022
.M. cul~fimiarzus
Total density Density > 1 cm
Bruchidontes
Total density Dcnsitj > 1 cm
0.33 + 0.58 0.33 & 0.58 4.67 + 1.53 0.003 0.00 i_ 0.00 0.00 & 0.00 2.00 f 2.00 0.105
256.00 f 46.03 178.67 + 82.72 447.672 157.91 0.05 1 5.33 & 1.53 8.00 2 1.73 52.00 5 3.61 i 0.000
* P value for Tukey’s HSD comparison of 7 values of open and caged plots = 0.1 I.

78 C. ROBLES AND J. ROBB
M. edulis and Bruchidontes, but not for M. californianus. Mussels begin to protrude
above the surface of the turf as they grow to 2 1 cm, presumably exposing themselves
to greater predation.
The Mjltilus spp. in exclosures overgrew the turf, causing moderate, but statistically
significant increases in their per cent cover (Fig. 6A). The arch and open controls
maintained unbroken covers of turf. These results demonstrate that the lobsters and
fishes were capable of extirpating mussel recruitment over a broad area (100 m’) even
while whelk predation was greatly reduced.
MYTILUS
FLESHY CORALLINE
MY-rlLUS
FLESHY CORALLINE
WfrlLUS
FLESHY CORALLINE
FLESHY CORALLINE
OPEN 0 LARGE CAGE 0 SMALL CAGE 0
LARGE ARCH A SMALL ARCH a
Fig. 6. Triangular coordinate plots of the relative proportions of the cover of coralline algal matrix. flesh! cpiphytes, and the M~~filus assemblage. Together these categories comprised > 90”,, of the mean total coc- crs of all experimental plots, with the exception of the second protected site experiment. as explained in Results. Positlon of the symbols within the triangles represents the mean relative proportions of the three types of cover. Thus, symbols falling on the base line of the triangle have no mussel cover. but differing
proportions of epiphytes to coralline algal matrix. Symbols at the center of the ficld represent approximate11 cqual mean amounts of all three covers. Symbols near the apex of the triangle represent covers approach- ing IOO”,, M,ri/us assemblage. Different symbol shapes represent different treatment groups. as indicated. Shading distinguishes means of experimental groups that differed significantly from those of the open con- trol plots in the Mann-Whitney U tests. Diagrams A-D are results for first protected site, second protected
site, semiprotected site, and exposed site experiments, respectively.

CARNIVORE EFFECTS 79
0 50 100 150 200
Days
Fig. 7. Plot of the mean abundances of whelks in different treatment plots of the second protected site CK-
periment. T, mean of data pooled from small arch and large cage trcatmcnts; C, mean of data pooled from large arch and open controls. Asterisks denote dates that the mean of treatments (T) differs signiticantl! from
the mean of controls (C; x= 0.05). 93”, of the whelks observed were C. rnmalli.
In the second experiment at the protected site (small and large mesh cages; Table
I), densities of whelks in the absence of transient predators (large mesh cage or small
mesh arch) were higher than in the control plots (large mesh arch or open plots, Fig. 7).
Repeated measures ANOVA yielded significant treatment effects (Table IV). In post
hoc contrasts of adjacent curves for single treatments in Fig. 7, only the large mesh
cage,‘large mesh arch comparison differed significantly. Since the differences among the
curves are transitive, transient predator exclosure curves (small mesh arch and large
mesh cage) do not differ significantly from one another, nor do the control curves (large
mesh arch and open). Pooled whelk densities for transient predator removals (means
of small mesh arch and large cage, the T curve in Fig. 7) and controls (means of large
TABLE IV
Repeated measures ANOVA of whelk densities in the second protected site experiment (small and large mesh cages). The upper table presents results for the effect of treatments, location (area previously used for the whelk removals or a nonremoval arca) and the interaction of treatment and location. The lower table prc-
sents results for the effect of time (repeated measures) and its interaction with the previous variables.
Source Sum of squares df Mean Square F ratio 1’ value
Trcatmcnt 52.50 3 17.50 4.47 Location 60.00 I 60.00 15.32 Interaction 12.23 3 4.08 1.04 Error 62.61 16 3.92
Time 21.25 9 3.03 1.70 Time x Treatment 45.92 21 1.70 0.95 Time x Location 12.75 9 1.42 0.80 Time x Treatment x Location 56.02 27 2.08 1.16 Error 256.67 144 1.78
0.0 I8
0.001 0.4OI
0.091
0.535
0.622 0.279

80 C. ROBLES AND J. ROBB
TABLEV
Mean per cent covers _t 1 SD of bare rock, undcrstorey invertebrates exposed with the decline of turf oovcr, and the turf in the second protected site experiment. P values for ANOVA of the differences in means among
treatment groups are also tabulated.
Treatment Bare rock Invertebrates TurS
Small cage Small arch
tar&x cage Large arch Open
P \altlc
19.50j 14.24 30.50+21.74 37.50 i_ 12.97 33.67 + 16.53 16.17 + 17.36 ah.33 _i 27.44
19.17i_5.81 11.50~21.69 52.SO + 12.07 20.00 + 8.65 16.17+22.19 63.33 + 77. IJ 30.50 & 13.25 IX.83 & 16.53 SO.67 * 10.75
0.140 0.437 0.313
mesh arch and open plots, the C curve in Fig. 7) differed signi~cantIy on five occasions.
Thus, whether grouping by the cage designs (single treatment curves) or by their effect
on transient predators (T and C curves) the results confirm that the whelks aggregated
in the transient predator exclosures.
The repeated measures ANOVA also revealed a significant location effect (Table IV).
Whelk densities were lower in the area used in the previous experiment for the whelk
manual removal, which would be expected if the whelk population had not fully re-
covered. Variation associated with sampling date followed a nonsignificant trend (Table
IV, P = 0.09).
Unlike the other experiments, in which the rock surfaces were never uncovered, the
thickness and cover of the turf assemblage declined in all plots, exposing either bare
rock or invertebrates, principally ~r~c~i~o~te~, barnacles (Tetr~clit~), and jewel box
oysters (Pseudoch~ma; Table V). Cage effects are not implicated because open and wire
covered plots were similarly affected (ANOVA; Table V). The turf is naturally thinner
at this site, and gaps in the turf appeared in other years.
The mussel assemblage increased according to the number of predators removed.
The percent cover of mussels in the large arch and open controls remained nil; small
arches and large cages, the transient predator removals, showed a nonsignificant in-
crease; and the small cage treatment, removing all but a few of the smaller whelks,
showed a statistically significant increase (Fig. 6B). Predation by whelks surely ac-
counted for most of the loss of mussels in plots protected from the transient preda-
tors. 93”; of a total of 69 empty &#.rtilus spp. shells retrieved from cage covered plots
bore whelk drill holes.
EXPERIMENTS AT THE SEMIPROTECTED AND EXPOSED SITES
Lobster exclosures at the semiprotected site (large mesh cages; Table I) produced
marked increases of the Mytilus assemblage. Mussels dominated the treatment plots,
while the turf remained unbroken on the controls (Fig. 6C). The cover of mussels in

CARNIVORE EFFECTS 81
f’lg. 8. ~‘h(~~~)~r~ph of an cxclosurc plot at the conclusion of lhc exposed site expwimenl. The crclt~trc cage has been I-cmovcd, but its outline on the turf is clearI) delineated by the cover of muss&. The rule IS
I5 cm long.
individual treatment plots was 45-93 OO. hf. calfjimiarzus comprised = 20”; of the total
cover of mussels. During nocturnal high tides, divers watched some lobsters foraging
on open and arch-covered control plots, while other lobsters pushed their foretegs
through the mesh of the cages, trying unsuccessfully to grasp the M~jtilus spp. protruding
from the turf.
Still greater increases in the Mq’tilus spp. assemblage were produced by lobster
exclosures on the exposed site (large mesh, Table I). At the start of the experiment, a
heavy set of juvenile mussels attached to the turf over the entire exposed site. During
the following 4 wk we observed the margin of the cover of mussels recede from its
seaward extremity shoreward across the bench until the turf was again the only sig-
nificant cover outside of the exclosures. inspection of the copious shell litter at the
receding margin revealed broken shells that bore sculptured fractures produced
by the lobsters (Robles et al., 1990; black oystercatchers make little or no mark
opening the shell). Mussels were cleared from control plots, but in the exclosurcs they
continued to grow rapidly, completely displacing the turf in two of the three ex-
closures (Fig. 8). The mean percent cover of mussels on exclosure plots exceeded
909, (Fig. 6D). Densities of ~~tjlu.~ spp. differed significantly between treattnents and
controfs (Table VI).
x2 C. ROBLES AND J. ROBB
TABLE VI
Mean numbers f 1 SD of mussels and herbivorous mollusks.230-cm’ sample- ’ in the experiment at the exposed site tabulated with respect to experimental conditions and shell length. Statistical analysis as in
Table III.
Species value Open Arch Cage P
M. edulis Total density 9.61 k 3.22 2.00 f 1.73
Density > 1 cm 1.33 2 1.58 1.67? 1.16
,M. culifbrnimus Total density Densit) > I cm
0.33 * 0.58 6.67 i 8.15 72.00 2 4.58 0.00 * 0.00 4.67 k 4.73 68.00* 5.57
Bmchidoniev Total density Density > 1 cm
Chitons > 0.5 cm Limpets > 0.5 cm Herbivorous mollusks
Total > 0.5 cm
16.00 f 10.54 6.22 i 3.06 1.00 + 1.73 0.67 i 0.58
3.33 +_ 2.08 0.33 +_ 0.58 3.00 k 4.36 3.00 + 1.00
6.33 k 5.51 3.33 k 1.16
8O.OOi30.12 i 0.000 7l.OO_t30.51 i 0.000
44.00 k 6.08 2.00* 1.73
7.33 +_ 2.3 1 14.33 + 8.74
21.675X.51
i 0.000 0.024
0.002 0.540
0.0 IO
0.079
0.018
MUSSEL SIZE AND PERSISTENCE
We observed a correspondence between prey size and persistence in the postexper-
imental periods. At the protected site, M. californianus comprised < 5 Oh of the total
mussel cover at the conclusion of the experiments. The mean length of the three largest
individuals of this species in each of the treatment plots was 1.3 & 0.7 cm, the longest
2.5 cm. All mussels disappeared in the first few weeks of the postexperimental periods
at this site.
Storms removed one cage from the semiprotected site in February 1987, just prior
to the spring resumption of lobster predation. z 15 y0 of the cover was large (4-5 cm)
M. calijxnianus, which persisted through spring and summer while a 701, cover of
M. edulis was lost. All experimental clumps and portions of the turf disappeared dur-
ing a stormy period late fall of 1987.
In the experiment on the exposed site, some M. californianus attained 4-5 cm lengths
as early as the July 1985 sampling (Fig. 9). By winter, when cages were removed, the
largest M. calijornianus had reached > 5 cm length (Fig. 9) and the cover of this spe-
cies was 40 YO. The experimentally induced clumps persisted until at least the winter
of 1991-92. increasing only slightly in area, but comprising > SOu/, M. ca/$wnimus.
DISCUSSION
DIFFUSE VS. KEYSTONE EFFECTS
Experimental results from the protected site support the hypothesis of diffuse pre-
dation through compensatory foraging shifts. Lobster and fish removals produced

CARNIVORE EFFECTS x3
5 -
o- - 0.0 0.0 1.0 1.6 20 1.6 ruI t.S 4.0 48 6.0 6.6 6.0
LENQTH CM
Fig. 9. Length-frequency distributions of M. cul~jhiunus in turf samples made on three different dates of the exposed site cxpcriment: the beginning, (23/06!85) conclusion (22/07iS5) and following winter (7;03,‘86).
Latter two samples are from the treatment plots.
significant changes only in combination with whelk removals. The whelks did aggrc-
gate in the transient predator exclosures. Mussel shell collections confirm that the
whelks caused most of the mortality in transient predator exclosures, and the chang-
ing proportions of Mjtihs spp. in the apparent whelk diets (Fig. 5) suggest that the
whelks might play a compensatory role in nature.
Predation on the wave-exposed site fits the keystone hypothesis: exclusion of lobsters
caused the persistent replacement of the turf by mussels. Considering the abundance

x4 C. ROBLES AND J. ROBB
and dietary information, contributory effects of the other predators are unlikely. Al-
though a single pair ofblack oystercatchers was sometimes present, the birds consumed
few M. cul(fornianus < 3 cm (Hart-wick, 1976; this study), the size range removed in the
experimental period. Furthermore, the experimental clumps of M. cnl$wnicrrm sur-
vived re-exposure to the lobsters and birds at lengths (> 5 cm) that were resistant to
the lobsters, but within the range killed by black oystercatchers in the natural mussel
beds.
The prior examples of keystone and the diffuse predation were separated from each
other by geographic region and species composition. The Catalina results suggest that
keystone and diffuse predation can operate in the same community, and in this case.
they may represent extremes of a continuum of the increasing effect of one prcdatot
relative to the others. We did not investigate the factors favoring the emergence of a
keystone predator. Although differences in the relative abundances of predators among
the sites coincide with the gradient of wave exposure, the cause of the differences rc-
mains unknown.
However, we caution that compound removal experiments have technical limita-
tions, and consequently, the diffuse effects at the protected site may be, at least in part.
an artifact. Out of practical necessity, compound removals couple the type of preda-
tor targeted with the scale of the manipulation (e.g., Menge & Lubchenco, 198 1). The
first experiment on the protected site (Table I) demonstrated that the transient pred-
ators were capable of limiting muss& over much of the available habitat (Z 100 m2
whelk removal bench). But in the second experiment (Table I), the whelks limited the
mussels only in the small areas of the transient predator exclosures ( < lo,, of the turf
zone) in which they congregated from the entire shore. The trend towards increased
mussel covers in large cage and small arch plots (Fig. 6B) shows that even with ag-
gregation, mussel removal by the whelks was incomplete. Therefore, the whelks might
not control the mussel assemblage over the entire turf zone, if large scale removals of
lobsters and fishes were feasible. Problems with experimental scale may exaggerate the
effects of compensatory foraging shifts.
Simultaneous removal of several predators does not measure the proportion of the
prey population falling to a specific predator. Apparent diets provide an indication, but
these may be biased by handling times (see Fairweather & Underwood, 1983. for
discussion). Consumption of mussels by large decapods and fishes is several orders of
magnitude faster than by gastropod predators (e.g., Chao, 1973; Brett, 1979; Edwards
et al., 1982; Menge, 1983). Thus, an arthropod, vertebrate, or other large consumer
within a diverse predator fauna may be the principal control of a dominant prey, yet
it remains unrecognized as a keystone under the limitations of our techniques.
KEYSTONE EFFECTS OF AN ARTHROPOD
Whether by natural constraints or the limitations of our assessments, strong effects
of single predator species are reported less frequently for: (1) terrestrial than aquatic

CARNIVORE EFFECTS x5
environments; (2) carnivores than herbivores; and (3) arthropods and mammals than
other taxa (Sih et al., 1985; but see Risch & Carroll, 1982; Lock, 1972; for marked
effects of insects and hippopotamus in terrestrial communities). Concurring with the
hypothesis that strong effects are rare at higher trophic levels (Menge & Sutherland,
1976) Sih et al. (1985) attribute the paucity of arthropod examples, especially in the
marine environment, to their characteristically high trophic status.
Heretofore, putative examples of marine arthropod keystones were supported pri-
marily by observations (e.g., Mann & Breen, 1972; Breen & Mann, 1976: see Elner &
Vadas. 1990, for discussion). Previous experimental studies of predation by marine
arthropods propose diffuse effects (eg., Menge & Lubchenco, 1981; Menge ct al.. 1986;
Hines ct al., 1990). Peterson (1979) describes a case in which observations suggested
a keystone effect of swimming crabs, but whelks and other predators prevented the
survival of mussels in crab exclosures. To our knowledge, the Catalina study provides
the first experimental demonstration of keystone effects of a marine arthropod.
CHANGING COMMUNITY INTERACTIONS
Removing predators cause more than the replacement of dominants. We illustrate
potential changes in community interactions with herbivorous mollusks. These are
among the most abundant of the > 200 invertebrate spp. found in mussel clumps
(Suchanek. 1979; Tsuchiya & Nishihira, 1986). In replicated turf samples (Robles,
1987; pers. obs.), combined densities of juvenile limpets and chitons ranged from 60
to 1050.11~’ However, large adults were very rare. The mollusks were found in the
lobster guts (Robles, 1987; this study) and the abundances of individuals >0.5 cm
increased significantly in lobster exclosures (Table VI), suggesting that they too are
limited by predation. Although the juvenile herbivorous mollusks do not clear the turf
from the rock, the adult populations associated with the mussel beds may prevent the
overgrowth of mature M. ccd~fivnirrnus by algae and other spatial competitors
(Suchanek, 1979; Paine & Suchanek, 1983). This overgrowth might otherwise lovvcr
mussel survivorship and reproduction (Witman & Suchanek, 1984; Dittman & Rob-
les, 1991). The results suggest a change in community interactions, even though the
assemblages share many species. [See Suchanek (1979) Tsuchiya & Nishihira (I 986),
and Dean & Connell (1987) for associated fauna of turf and mussel beds. Witman
(1985) describes community relationships of species associated with subtidal tnusscl
beds.]
SIZE RELATIONS AND PERSISTENCE OF EXPERIMENTAL CHANGES
The abilities of M. calijimGums to hold space and to resist predators accrue to large
body size (Paine, 1966, 1974, 1976; Dayton, 197 1; Harger, 1972; Roblcs et al., 1990).
The persistence of downward extensions of mussel zones following sea star removals
apparently depends on the mussels’ size range relative to that of the returning stars

86 C. ROBLES AND J. ROBB
(Paine et al., 1985). Farrell (1988) proposes a similar relationship for the persistence
of barnacles following a limpet removal. Therefore, size-limited predation probably
caused the longer persistence of relatively large M. californianus in the postexperimental
periods.
Differences in mussel growth rates among the sites apparently determined much of
the size differences of experimental mussels. Records of mussels lengths in the exclo-
sure plots indicated that M. califovniun~s grew out of the turf roughly twice as fast at
the exposed site than at the protected site. We tested for possible site-specific differ-
cnces in mussel growth rates by transplanting matched cohorts of small (2-4 cm)
M. calijbrniutzus simultaneously to the same shore level of each site and recording the
shell growth increment. The groups were covered with 1.2-cm’ opening wire mesh to
protect them from predators. Successive runs of the experiment were begun fall and
winter of 1990-91. The mussels did grow significantly faster at the wave-exposed sites
(Fig. 10).
Perennial beds of Mytilus spp. cover many wave-beaten shores of North America and
Europe. Wave stress in these areas favors higher mussel abundances by hampering
predator foraging (e.g., Menge, 1978a,b). M. californianus beds replace the turf on the
most wave-washed ends of the islet, and predator foraging ceased at these locations
in the season of highest wave action. Our transplant experiments, studies of natural
mussel populations in different wave exposures (Robles & Sherwood-Stephens; C.
Robles, unpubl. data) and indirect estimates (turnover of biomass; Leigh et al., 1987)
show a positive relationship between wave exposure and growth rate. We speculate that
81
protected site semi-protected exposed site
m Fall Run k?%@ Winter Run
Fig. 10. Mean increment and 1 SE of shell growth in two transplants of M. culijbrniunur to the three sites. The “fall” transplant ran from 15 September 1989 to 23 February 1990; the “winter” transplant from 4 March 1990 to 8 June 1990. For the latter, mussels were not transplanted to the semiprotected site. P values of
ANOVA of growth increments were 0.04 and 0.01. respectively, for fall and winter runs.

CARNIVORE EFFECTS x7
shifts in the relative abundance of turf and mussels over the wave exposure gradient
reflect changing relative levels of production and predation. Prey gaining resistance to
predators with size i.e., M. califovniunus, should display greater persistence on wave-
washed shores.
PHYSICAL STRESS, INDIRECT EFFECTS, AND PREVALENCE OF THE TURF
Prior explanations for the prevalence of the turf emphasized its tolerance to physi-
cal and biotic stresses (Littler & Littler, 1980; Hay, 1981). The mat’s ability to trap
water and sediments imparts resistance to desiccation (Hay, 1981). Desiccation stress
appears to be greatest in fall, when prolonged low tides coincide with strong sun and
dry winds (Seapy & Littler, 1982). If sun, storm waves, or other disturbances damage
the cover, vegetative propagation from intact thalli and sprouting from crusts quickly
close the gaps (Sousa et al.. 1981).
The turf resists some biotic challenges. The entrapped sediments may deprive some
spatial competitors of firm substrata for attachment (Seapy & Littler, 1982; Stewart,
1982, 1989). Seapy & Littler (1982) propose that fall desiccation frees space for the turf
by removing adult sedentary invertebrates. Rapid closure of gaps by vegetative prop-
agation may preclude other macroalgal species that colonize only by spores (Sousa
ct al., 198 1). Finally, the turfs tough tissues, low profile, and ability to increase branch-
ing when terminal portions are clipped evidently allows it to withstand moderate
browsing by fishes (Littler & Littler, 1980; Hay, 1981).
Although the mussels may be less tolerant of physical stress than the turf, the
replacement of turf by mussels in exclosures cannot be attributed to cage artifacts, i.e.,
reduced desiccation stress (Dayton & Oliver, 1980). Seapy & Littler (1982) cite algal
bleaching as evidence for desiccation stress. We compared per cent of total algal cover
bleached among open, arch, and cage plots during an episode of marked bleaching (fall
of 1982). Mean percentages of bleaching were 32.3, 28.7, and 26.7”, respectively, for
open, arch, and cage plots (ANOVA: F = 0.408; df = 2,6; P = 0.682). In all experiments,
50-loo’:, of the differences in mussel abundances between treatments and controls
developed in spring-summer, when physical stress are thought to be minimal, but
intertidal predators were most abundant. The hypothetical effects of desiccation would
have been preempted by predation. At the wave-exposed site (Table I), the mussels
survived the prolonged daytime emergence of the fall-winter after cages were removed.
In the second experiment on the protected site (Table I), we can not discount the
possibility that some of the treatment effect was an artifact, because there lvas no strict
small mesh control, i.e., a small mesh arch that excluded no predators. However, mean
mussel covers for small mesh arches and large mesh cages appeared more similar to
one another than to mean covers of either the large mesh arches or small mesh cages,
an outcome that would be expected if predation, rather than differing mesh sizes,
caused the differences among treatment groups (Fig. 6B).
While imparting resistance to some biotic and physical stresses, the turfs morphol-

xs ~.ROBLESA~DJ.ROBB
ogy may render it vulnerable to the mussels. Filamentous algae attract the settling stages
of the mussels (e.g., Seed, 1969a,b, 1976; Paine, 1974; Petraitis, 1979; Peterson, 1984).
and we found much higher densities of mussel plantigrades (recently settled individ-
uals < 1 cm) in the turfs than on bare rock (C. Robles, pers. obs.). On the islet, the
rapid growth of plantigrades quickly overtopped the low profile of the turf.
Resistance to desiccation, invasion of sporelings, and moderate browsing may well
explain the success of the turf relative to other algal assemblages, however. the prev-
alence of the turf also depends on the indirect effects of carnivores.
R. Desharnais, D. Levitan, R. Paine, P. Petraitis, IS. Pfister, S. Prescott, T. Raw-
lings, R. Sherwood-Stephens, and W. Sousa provided valuable criticism of preliminary
drafts. M. Alvarado, E. Carriazo, D. Dittman, C. Gramlich, G. Jung, K. Petit, H.
Ramos, D. Sweetnam, M. Vcla, and F. Villeda assisted the field work. The work was
supported by NSF Grants OCE8200474, R118505428, and RII8804679.
REFERENCES
Brcen. P.;\. & K.H. Mann. l976. Changing Jobster ahundan~~ and the destru~ti~ll of kelp beds h) SW-
urchins. .tft/v. Bioi., Vol. 34. pp. 137-142.
Brett, J. R.. 1979. SOW morphological adaptations of pile perch (~~ff~~1~/7i~~~.\ WCM) feeding on ~nusscls
(.bfuih &li~). CW. J. Zd. Vol. 57, pp. 658-663.
Chao. L. N.. 1973. Digestive system and feeding habits of the cunner. T~u~o~~~l~hnrs crd.~p<~w.s. a stomachless
fish. ES/I. Roll., Vol. 71. pp. 565-586.
Cubit. J. D., 1984. Herbivory and seasonal abundance of algae on a high intertidal rocky shore. Eu&+~,. Vol.
65. pp. 1904-1917.
Dayton, P. K., 1971. Competition, disturbance. and community organization: the provismn and suhscquent
utilization of space in a rocky intertidal community. Ed. Monqy., Vol. 41. pp. 351-389.
Dayton. P. K., 1975. Experimental studies of canopy interactions in a sea otter dominated kelp communit\;
at Amchitka Island. Alaska. Fi.&. &r/t.. Vol. 73. pp. 230-237.
Dayton. P. K. & J. S. Oliver. 1980. Xn evaluation of experimental analysis of p~~pulation and cor~lr~~unit~
patterns in benthic marine environt~~cnts. In. ~~u~~~ henrhic ~l,n~~~i~~s edited by K. R. Tcnore & B. P. Couil.
tinivcrsit? of South Carolina Press. Columbia. South Carolina, pp. 93-120.
Dean. R. L. & J. H. Conneli, 1987. Marine invcrtebratcs in algal succession. 1. Variations in abundance and
diversity with succession. .I. E-y. MN. Bird. Ed.. Vol. 109. pp. 195-215.
Dittman. D. E. & C. Robles, 1991. Effect of algal epiphytes on the mussel ~Mu~/uY c,rr/if;wGnw. Ew/o,q~,. Vol.
72. pp. 286-796.
Duggins, D.O., 1980. Kelp beds and sea otters: an experimental approach. Ec+y, Vol. 61, pp. 447-453.
EdwTards, C. D., D. 0. Conover & F. Sutter, 111, 1982. Mobile predators and the structure of marine intertidal
communities. Ecok~~>~, Vol. 63, pp. 1175-l 180.
Elncr, R.W. & R.L.. Vadas. 1990. lnferencc in Ecolog!: the sea urchin phenomenon in the northwestern
Atlantic. .4rit. ,l’trr.. Vol. 136, pp. 10%!2S.
Emerson. S. I<. & J. B. Zeldcr, 1978. Recolonization of intertidal algae: an experimental study. !%far. S&l..
Vol. 44. pp. 3 I s-324.

CARNIVORE EFFECTS 89
Estcs. J.A. & J. F. Palmisano, 1974. Sea otters: their role in structuring near shore communities. .Scic~nc,r.
Vol. 185, pp. 1059-1060.
Fairweather, P. G. & A. J. Underwood, 1983. The apparent die1 of predators and biases due to different
handling times of their prey. &co&in (Berlin), Vol. 56, pp. 169- 179.
Farrell, T.M., 1988. Community stability: effects of limpet removal and reintroduction in a rocky in(ertidal
community. Oecolngiu (Berlitz), Vol. 75, pp. 190-I 97.
Hargcr. J. R. E., 1967. Population studies on MJ~/&Y communities. Ph.D. dissertation, Univcrait) of Cali-
fornia at Santa Barbara, Santa Barbara, California. [University Microfilms. No. 69-17191.
l-larger. J. R. E., 1972. Competitive coexistence: maintenance of interacting associations of the sea mus~cla
.&f~~ilus edulis and Mytilus culiforniutms. Veliger. Vol. 14, pp. 387-410
Hartuick, E. B., 1976. Foraging stratcgq of the black oystcrcatchcr Huemutopuc hachrnmi Audubon. Ct/tl.
J. ZOO/.. Vol. 54, pp. 142-155.
Ha), M. E., 1981. The functional morphology of turf-forming seaweeds: persistence in stressful marine
habitats. E&o~L; Vol. 62, pp. 739-750.
Hines. A. H., A. M. Haddon & L. A. Weichcrt, 1990. Guild structure and the foraging impact of blue crabs
and epibenthic fish in a subestuary of Chesapeake Bay. Mrrr. kcol. Prog. Ser.. Vol. 67. pp. 105-126.
Hobson. E. S.. CV. M. McFarland & J. R. Chess. 1981. Crepuscular and nocturnal activities of California
ncarshorc fishes. with consideration of their scotopic visual pigments and the photic environment. F/\/T.
Bull.. Vol. 19. pp. l-29.
Helling. C. S.. 1959. The components of predation as revealed by a stud! of small mammal predation of
the European pine saw fly. Ccm. Entonwl., Vol. 91, pp. 293-320.
Kcrfoot. W. C.. 1987. Cascading effects and indirect pathways. In, Predutim: direct und indirecr irnpcrcr.\ itr
crquur;~~ umnnmities, edited by W. C. Kcrfoot & A. Sih, University Press of New England. Hanover. Nc\\
Hampshire. pp. 57-70.
Leigh. E.G., R.T. Paine, J. F. Quinn &T. H. Suchanck. 1987. Wave energy and intertidal productivit>. Proc
:Vutl. ncad. SC;., Vol. 84, pp. 1314-1318.
Littler, M.M. Br D.S. Littler, 1980. The evolution of algal thallus form and survival stratcgics in
bcnthic marine macroalgae: field and laboratory tests of a functional form model. Am. ,licrt.. Vol. 116. pp. 25-44.
Lock. J. M., 1971. The effects of Hippopotamus grazing on grasslands. J. f&l., Vol. 60, pp. 445-368.
1,ubchcnco J., B. 4. Menge, S. D. Garrity, P.J. Lubchenco, L. R. Ashkenaa, S. D. Gaines, R. Emlet, J. Lucas
& S. Strauss. 1984. Structure, persistence. and the role of consumers in a tropical rocky intertidal com-
munity (Tobaguilla Island, Bay of Panama). J. E.vp. Mm. Biol. Ecol.. Vol. 78, pp. 23-73.
MacDonald, J. H. & R. K. Koehn, 1988. The mussels M~likrs ~u/lopro~inc~irr/~.iF and ,M. trmtuhrs on the Pa-
cific Coast of North America. Mar. Biol., Vol. 99, pp. 1 I l-l 18.
Mann, K. H. & P.A. Brcen, 1972. The relation between lobster abundance. sea urchins and kelp beds. ./.
C‘frrl. Fish. RCF. Board, Vol. 29, pp. 603-609.
hlarbh, C. P., 19X6. Rocky intertidal community organization: the influence of avian predators on mussel
recruitment. fi&~~?: Vol. 67. pp. 771-786.
Mcngc. B.A.. 1976. Organization of the New England rocky intertidal community: role of predation.
competition and environmental heterogeneity. Ecol. Monoqr., Vol. 46, pp. 355-393
Mcngc. B.A.. 1978a. Predation intensity in a rocky intertidal community: relation between predator forag-
ing activity and environmental harshness. Oecologia (Berlin), Vol. 34. pp. I-16.
hlcnge, B. .A.. 1978b. Predation intensity in a rocky intertidal community: Effect of an algal canopy. \\avc
action and desxcation on predator feeding rates. Orcokgio (Berlin). Vol. 34, pp. 17-35.
Men&c. B.A.. 19X3. Components of predation intensity in the low zone of the New England rock? intcr-
tidal region. O~~~~/ogia (Berlin), Vol. 58. pp. 141- 155.
Mengc. B.A. & T.M. Farrell, 1989. Community Slructurc and interaction webs in shallow marme hard
bottom communities: tests of an environmental stress model. In, Adratxzr irz eco/qicu/ resrarch. edited
bq M. Begon er al.. Academic Press, San Diego. California. pp. 189-262.

90 C. ROBLES AND J. ROBB
Menge, B.A. & J. Lubchenco, 1981. Community organization in temperate and tropical rocky intertidal
habitats: prey refuges in relation to consumer pressure gradients. Ecol. Monogr., Vol. 51, pp. 423-450.
Menge, B.A.. J. Lubchenco, L. R. Ashkenas & F. Ramsey, 1986. Experimental separation of ctfects of
consumers on scssile prey in the low zone of a rocky short in the Bay of Panama: direct and indirect
consequences of food web complexity. J. E.qj. Mar. Viol. Ecol.. Vol. 100, pp. 225-269.
Mengc. B.A. & J.P. Sutherland, 1976. Spccics diversity gradients: synthesis of the roles of predation,
competition and temporal hcterogcneity. Am. ,Nat.. Vol. 110. pp. 351-369.
Mitch& C. T., C. H. Turner & A. R. Strachan, 1969. Observations on the biology and behavior of the
California spiny lobster. Punulinr.~ i~~t~rrrq~~.~ (Randall). Cu/$ /-‘is/i Gunzr, Vol. 55, pp. 12 I - I3 1.
Murray, S. N. & M. M. Littler, 1978. Patterns of algal succession in a pcrturbated marine intertidal com-
munity. J. fhyol., Vol. 14, pp. 506-512.
Murrq. S. N., M. M. Littler & 1. A. .Abbot, 1980. Biogeography of the California marine algae with emphasis
on the Southern California Islands. In, The Calrfinzic/ I.dands: Pmceeding.~ of (1 Mulridi.tri/,lir2r~l, .~~~mp-
.ciwr, cditcd by J. Power, Santa Barbara Muscum of Natural History, Santa Barbara, California. pp. 325%
339.
Paint, R.T.. 1966. Food web complexity and species diversity. Am. Nur., Vol. 100. pp. 65-76.
Paine. R.T.. 1969. The Pi.susrer-72&u interaction: prey patches. predator food preference and intertidal
community structure. Ecolqy, Vol. 50, pp. 950-961.
Pamc. R.T., 1974. Intertidal community structure: experimental studies on the relationships hctwccn a
dominant competitor and its principal predator. Oe&ogicr /Bet&j, Vol. 15, pp. 93- 120.
Pamc. R.T.. 1976. Sire-limited predation: an observational and experimental approach with the .~~~~ihr.~-
POcrtrer interaction. Ec&g,r, Vol. 57, pp. 858-873.
Paine. R. T.. 1980. Food wjcbs: linkage, interaction strength. and community infrastructure. J. A/Cm. Ed.
Vol. 49. pp. 667-685.
Paine R.T.. J.C. Castilla & J. Cancino. 1985. Perturbation and recovery patterns of starfish-dominated
intertidal assemblages in Chile. New Zealand and Washington State. An?. &or., Vol. 125. pp. 679-691.
Paint. R.T. & T. H. Suchanek, 1983. Convergence of ecological procescs between independently evolved
competitive dominants: a tunicatc-mussel comparison. Ec&g~~. Vol. 37. pp. 821-83 1.
Pctcrson. C. II.. 1979. The importance of predation and competition m organizing the mtcrtidal cpifaunal
communities of Barncgat Inlet, New Jersey. Oecolo~ia (Berlirrj, Vol. 39. pp. l-24.
Pctcrson. J. H.. 1984. Establishment of mussel beds: attachment behavior and distribution of rcccntly sct-
tied mussels (Mrtilzrs cul~fiwniunu.r). VeliRer. Vol. 27. pp. 7- 13.
Pctrams, P. S., 1979. Distributional patterns of juvcnilc h1~~ilirs eduli,~ and ,U,ti/u.r crrlifomicrnu.~. C’cliXer. Vol.
2 I, pp. 288-292.
Risch, S.J. & C. R. Carroll, 1982. Effects of a keystone predacious ant, Sdenyxi.c geminutcr. on arthropods
in 3 tropical agroecosystem. EuJ&~*. Vol. 63. pp. 1979-1983.
Roblcs. C.. 1987. Predator foraging characteristics and prey population structure on a sheltered short.
EC &g~~, Vol. 68. pp. 1502- I5 11. Roblcs, C., I). A. Swectnam & D. Dittman, 1989. Dicl variation of intertidal foraging by C~rzcr, /~~oducrrr.t
L. in British Columbia. J. krt. Nist., Vol. 23. pp. 1041-1049.
Roblcs. C., D.A. Swectnam & J. Eminikc. 1990. Lobster predation on mussels: shore-level dill’crcnccs in
prey vulnerability and predator prefcrcnce. Eco/ogj,, Vol. 71. pp. 1564-1577.
Scapy. R. R. & M. M. Littler. 1982. Population and species diversity fluctuations in a rocky intertidal
communit) rclativc to severe aerial exposure and sedtment burial. MU. Bio[., Vol. 71. pp. X7-96.
Seed. R.. 19693. The ecology of il;/?,tilus edu1i.s I.. (Lamellibranchiata) on exposed rocky shores. I. Brccd-
mg and scttlemcnt. Oecologiu /Berlin). Vol. 3. pp. 277-316.
Seed. R.. 1969b. The ecology of jvJ,rilrrr CL/U/~.F L. (Lamellibranchiata) on exposed rocky shores. II. Grovvth
and mortality. Oecohgia (Berlin). Vol. 3. pp. 3 17-350.
Seed, R., 1976. Chapter 2. Ecology. In, Mat& musse(.c: rheir ecdog~ crrld/,h,‘.violo~~,, edited by B. L. Baync.
Cambridge llniversity Press. Cambridge, IJK, pp. 17-60.

CARNIVORE EFFECTS 91
Scymorc. R. J.. 1984-1987. Monthly Summar) Reports. Coastal Data Information Program, Ncarahore
Research Group, Institute of Marine Resources. Scripps Institution of Oceanography. La Jolla, Califor-
nia.
Sih. A.. P. Crowley, M. McPeek, J. Petranka & K. Strohmeicr, 1985. Predation, competition and prq
communities: a review of field expcrimenta. Anrnr. Rev. Ecol. Sj,.~t.. Vol. 16, pp. 269-3 I I. Simcnatad, C. A.. J. A. Estes & K. W. Kcnyon. 1978. Alcuts. sea otters. and alternate stable communities.
S~.ieIIL.C. Vol. 200, pp. 403-4 I I. Sokal. R. R. & F. J. Rohlf, 198 I Bionw/q~. W. H. Freeman & Co.. San Francisco, California. second cdi-
non. 859 pp.
Sousa. W. P., S.C. Schroetcr & S. D. Gainca. 1981. Latitudinal variation in intertidal algal communlt~
structure: the influence of grazing and vegetative propagation. Orcwk~~iu /Bw/rnl. Vol. 48. pp. 297-307.
Stewart. J. G.. 1982. Anchor species and cpiphqtes in intertidal algal turf. Puce Sci., Vol. 36. pp. 45-59.
Stewart. J.G.. 1989. Establishment, pcrsistcnce and dominance of Cwc&w (Rhodophyta) in algal turf. ./.
PIIwJI., Vol. 25. pp. 436-446.
Suchanek, T. H.. 1979. The Mrrikrs c~~/ifiwnicmus communit): studies on the composition, structure. orgatr-
/ation, and dynamics of a mussel bed. Ph.D. dissertation. Department of Zoologq, IJni\,crsit) of Wash-
ington, Seattlc. Washington.
Suchanck. T. H., 1985. Mussels and their role in structuring rocky short communities. In. The rc~o/r~~q ~fror~l, I’
~w.vt.c. edited by P.G. Moore & R. Seed. Hoddcr and Stoughton Prcas. London. LJK. pp. 7ll-Oh.
Thorn. R. M. &T. B. Widdowson, 1978. A gradient in bcnthic intertidal algal assemblages along the southern
California coast. J. P/zIu~., Vol. 16. pp. 102-10X.
Tsuchiyr, M. & M. Nishihira, 1986. Islands of Mrrilas m/u/is as :I habitat for small intertidal animals: cf.
feet of ,24~~ri/u\ age structure on the species composition of the nssociatcd fauna and communit) organ-
ration. .%ftrr. Eu>/. Prog. Ser.. Vol. 3 I. pp. 17 I- 17X.
VVilbur. H. M. & J. P. Fauth, 1990. Experimental aquatic food webs: interactions between two predators and
t\+o prq, ,lrn lirr.. Vol. 135. pp. 176-204.
W’ingct. R.R.. 1968. Trophic rclatronship and metabolic cncrgy budget of the California spin! lobstct-.
Pmdiru.~ imrmpu.~ (Randall). M.Sc. thesis, San Diego State Univcrsitj. San Diego, Cahfornia.
\X’itman. J. D.. 1985. Refuges. biological disturbance. and rocky subtidal community structure in NIX EII-
gland. Ewl. jAl~~rwgr., Vol. 55, pp. 447-468.
Vv’itman. J. D. & T. H. Suchanek, 1984. Mussels in how: drag and dislodgement by cpiaoana. ‘Lltr~. I-.<,~J/.
I’nq. Ser., Vol. 16. pp. 259-268.